Introduction
Rheumatoid arthritis (RA), a chronic multisystem
disease associated with autoimmunity, is typically characterized by
the excessive proliferation of synovial cells (1,2).
The global incidence of RA is approximately 0.5 to 1.0%, and the
incidence among the female population is significantly higher than
among the male population (3,4).
Without timely diagnosis and treatment, RA may further lead to
joint damage and disability, severely affecting health and the
quality of life of patients (5,6).
Fibroblast-like synoviocytes (FLSs) are the stromal cells of the
joint capsule and play important roles in the development of RA
(7). The abnormal activation and
proliferation of FLSs can cause damage to the joint synovium and
accelerate the disease progression of RA (8). According to existing research, the
pathogenesis of RA is related to a number of factors; however, the
specific etiology remains unclear (9,10). At present, the most common
clinical treatment for RA mainly includes the treatment of symptoms
to reduce pain experienced by patients; however, the effects of
these treatments are not ideal, and they often lead to certain
side-effects. Therefore, the development of novel treatment
strategies for RA is of utmost importance.
Traditional Chinese medicine has been applied as a
means of RA treatment for thousands of years; some of these
medicines are still used frequently and have been proven effective
(11,12). Huang Qi (Astragalus
membranaceous), one of the most common traditional Chinese
medicinal herbs, bearing a long history of medicinal use, has been
observed to be therapeutically effective in the course of RA and
its related complications. For example, Liu et al (13) found that the total flavonoids of
Astragalus membranaceous reduced adjuvant
arthritis-associated damage in rats by regulating the
OPG/RANKL/NF-κB pathway. Pu et al (14) proved that Astragalus
membranaceous polysaccharides exerted therapeutic effects on
inflammation and synovial cell apoptosis in rats with adjuvant
arthritis. Astragaloside (AST) is the main active ingredient of
Huang Qi, including Astragaloside I-VIII. AST has been shown to
exert antioxidative, anti-inflammatory, and immune regulatory
effects (15-17). In a previous study by the
authors, it was found that AST alleviated RA-associated
pathological injury and regulated long non-coding RNA (lncRNA)
abnormal expression (18).
An increasing number of studies have shown that
lncRNAs, which are >200 nucleotide (nt) in length and lack an
open reading frame, play important roles in a variety of biological
processes and participate in the pathogenesis of a number of
autoimmune diseases (19,20).
As the authors have previously demonstrated, lncRNAs with a
differential expression in RA can regulate microRNAs (miRNAs or
miRs), as competitive endogenous RNAs (ceRNAs) and can subsequently
affect RA occurrence and development (21). Moreover, as also previously
demonstrated by Bi et al (22), lncRNA PICSAR promoted the
proliferation, migration and invasion of FLSs by sponging
miRNA-4701-5p in RA. Therefore, with lncRNAs and miRNAs as the
starting point, research on novel therapeutic targets of RA may
prove helpful for identifying new clinical diagnosis and treatment
strategies.
In a previous study by the authors, it was
demonstrated that lncRNA LOC100912373 was critical for RA
pathogenesis (23). In
subsequent studies, was further validated that lncRNA LOC100912373
upregulated the expression of 3-phosphoinositide-dependent protein
kinase-1 (PDK1) by sponging miR-17-5p, and thus enhancing FLS
proliferation (24). In the
present study, the effects of AST on FLS proliferation and cell
cycle progression, and on lncRNA LOC100912373/miR-17-5p/PDK1 axis
component expression levels were investigated and the mechanisms
through which AST funcitons in the treatment of RA were
elucidated.
Materials and methods
Materials and reagents
AST was purchased from Zhibaicui Biotechnology Co.,
Ltd., and its purity was >98.5%. Complete Freund's adjuvant was
purchased from Sigma-Aldrich; Merck KGaA. DMEM was purchased from
Gibco; Thermo Fisher Scientific, Inc. Anti-Vimentin antibody (cat.
no. ab92547) was purchased from Abcam. PI staining solution was
purchased from Beijing Solarbio Science & Technology Co. An MTT
assay kit was purchased from Shanghai, BestBio. EZ-10 Total RNA
Mini-Preps kit reagent was purchased from Sangon Biotech Co., Ltd.
A reverse transcription kit was obtained from ABclonal Biotech Co.,
Ltd. Antibodies against PDK1 (cat. no. ab110025), AKT (cat. no.
ab18785) and phosphorylated (p-)AKT (cat. no. ab38449) were
purchased from Abcam. Secondary antibodies (anti-mouse or
anti-rabbit; cat. nos. ZB-2301 and ZB-2305) were purchased from
Beijing ZSGB-BIO. Cy3-labeled goat anti-rabbit antibody (cat. no.
A0516) was purchased from Beyotime Institute of Biotechnology. The
pcDNA3.1 plasmid was synthesized by Shanghai GenePharma Co., Ltd.
Lipofectamine® 2000 Transfection Reagent was purchased
from Invitrogen; Thermo Fisher Scientific, Inc.
Animals
In the present study, male Sprague-Dawley SPF grade
rats (6-8 weeks old, weighing 200±20 g) were purchased from the
Experimental Animal Center of Anhui Province. All rats were raised
in the animal facility of the First Affiliated Hospital of Anhui
University of Chinese Medicine with an indoor temperature of
18-22°C and a humidity of 40-60%, under 12-h alternate dark/light
cycle. Regular rat feed and tap water were provided ad
libitum. The experimental design was approved by the Animal
Ethics Committee of Anhui University of Chinese Medicine
(AHUCM-rats-004). Following one week of adaptive feeding, a model
of adjuvant arthritis (AA) was established by a single injection of
Freund's adjuvant into the left hind foot of the rats, as
previously described by Jiang et al (25).
Cell culture and identification
Following 20 days of AA modeling (25), the rats were anesthetized by an
intraperitoneal injection of 1.0% pentobarbital sodium (60 mg/kg
body weight) and sacrificed by cervical dislocation after blood
sampling from the abdominal aorta. A quantity of 8-10 ml blood
volume was collected, and the synovial tissue of the knee joint was
then separated, as previously described (26). The animal remains were
subsequently sent to the Anhui Experimental Animal Carcass
Management Organization for unified processing. The FLSs were
cultured in complete DMEM containing 20% fetal bovine serum at 37°C
and 5% CO2 through tissue mass culture method (27). At different time points, the FLSs
were evaluated under an inverted phase contrast microscope (Olympus
Corporation), and passaged synoviocytes were identified by vimentin
immunofluorescence staining. The standard method of
immunofluorescence staining was then applied as described
below.
MTT assay
The cells were digested with trypsin and seeded in a
96-well plate (5,000 cells per well). AST was diluted in culture
medium to a desired concentration (7.8125, 15.625, 31.25, 62.5,
125, 250 and 500 mg/l), added to the cells and then incubated for
24, 48 and 72 h at 37°C. Subsequently, 10 µl MTT were added
4 h before the end of culture. Following 4 h of continuous culture,
100 µl formazan solution were added to each well, and the
plates were oscillated at a low speed for 10 min, in order to fully
dissolve the crystals. The 96-well plates were removed when all the
purple crystals were dissolved. The optical density was measured
using a microplate reader (Multiskan™ GO; Thermo Fisher Scientific,
Inc.) at 570 nm (28).
The IC50 values were also calculated according to
the following formula: lgIC50=Xm-I [P-(3-Pm-Pn)/4],
where Xm represents the lg maximum dose; I represents the lg
(maximum dose/relative dose); P represents the sum of the positive
reaction rate; Pm represents the maximum positive reaction rate;
and Pn represents the minimum positive reaction rate. Inhibition
rate=(1-OD experimental group/OD control group) ×100.
Cell cycle assay
The cells were cultured with AST (50 mg/ml), and
were then collected at the point when a density of 1×106
cells/ml was reached. Subsequently, 100 µg/ml PI staining
solution (100 ml) were added, followed by a 30-min incubation in
the dark. The percentage of synovial cells in each stage was
detected by flow cytometry (Beckman Coulter, Inc.), in order to
observe the effect of AST on FLS cell cycle progression.
Cell ultrastructure
Cells treated with AST were collected, fixed with
2.5% glutaraldehyde and 1% ozone acid, dehydrated with acetone and
infused with an epoxy resin embedding agent. Finally, the samples
were sectioned and stained, and the ultrastructure of the cells was
observed through transmission electron microscopy (Hitachi,
Ltd.).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from the FLSs using an EZ-10
Total RNA Mini-Preps kit, and RNA was then reverse transcribed into
cDNA with the use of a reverse transcription kit. PCR analyses were
performed with TB Green™ Premix Ex Taq™ II (Takara Bio, Inc.). The
reaction conditions were as follows: Pre-denaturation at 95°C for
30 sec, 40 cycles at 95°C for 5 sec and 60°C for 1 min. β-actin was
used as the endogenous control. All the results were calculated
using the 2−ΔΔCq method (29) and all primers were synthesized by
Shanghai Sangon Biotech Technology Service Co., Ltd. Primer
sequences are listed in Table
SI.
Western blot analysis
The cells were washed with cold PBS three times and
lysed with 100 µl lysis buffer. The lysates were then
separated by sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE; concentrated gel, 5%; separating gel,
10%) and then transferred to a polyvinylidene fluoride (PVDF)
membrane. The PVDF membrane was washed with 1X TBST for 2 min,
blocked with 5% skim milk at room temperature for 2 h, and
incubated with a primary anti-PDK1 antibody (1:500), an anti-AKT
antibody (1:1,000) and an anti-p-AKT antibody (1:1,000) overnight
at 4°C. The cells were then washed three times with TBST and
incubated with a secondary HRP-conjugated anti-rabbit antibody HRP
(1:10,000) for 2 h at room temperature. All blotting experiments
were performed three times. The proteins were detected with an
enhanced chemiluminescence kit (Super Signal WestFemto kit; 34094;
Thermo Fisher Scientific, Inc.). The western blot analysis data
were quantified using ImageJ software (version 1.52; National
Institutes of Health).
Immunofluorescence
Cells were fixed in 4% paraformaldehyde for 30 min.
A total of 50-100 µl 0.25% Triton X-100 was added and the
sections were incubated for 10 min at room temperature. The
sections were washed with PBS three times, primary antibodies were
added, and the sections were immunostained overnight in a wet
chamber at 4°C (antibody dilutions: PDK1, 1:200; p-AKT, 1:200). All
sections were then stained with the corresponding secondary
antibody [goat anti-rabbit IgG H&L (FITC); dilution, 1:1,000;
cat. no. ab6717, Abcam] and incubated at room temperature for 50
min. In the end, nuclei were stained with DAPI at room temperature
for 5 min. Immunofluorescence was observed under a fluorescence
microscope (Nikon Corporation).
Rescue experiment
To further confirm whether AST interferes with FLS
proliferation and cell cycle progression by regulating lncRNA
LOC100912373 expression, a rescue experiment was conducted.
Full-length lncRNA LOC100912373 was cloned into the pcDNA3.1
plasmid to construct an overexpression plasmid; this construct was
labeled lncRNA LOC100912373 overexpression (Over). In addition, the
empty pcDNA3.1 vector was labeled as the negative control (NC). The
cells were seeded in a 6-well plate in advance, in order to ensure
that cell confluence was 60 to 80% per well at the time of
transfection. The lncRNA LOC100912373 overexpression and NC
plasmids were transfected into the FLSs using
Lipofectamine® 2000. Specific transfection operation and
screening of the optimum overexpression concentration of lncRNA
LOC100912373 (2.5 µg/5 µl) were performed according
to a published method by the authors (24), and the FLSs were treated with AST
(50 mg/ml) simultaneously. At 48 h after transfection, the cells
were collected and cell proliferation and cell cycle progression
was detected by MTT assay and flow cytometry.
Statistical analysis
All data were analyzed using SPSS 17.0 software
(SPSS, Inc.), and results are expressed as the mean ± SD (standard
deviation). One-way analysis of variance (ANOVA) was used for
comparisons among multiple groups and Tukey's multiple comparisons
test was used to define differences between groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
Identification of FLSs
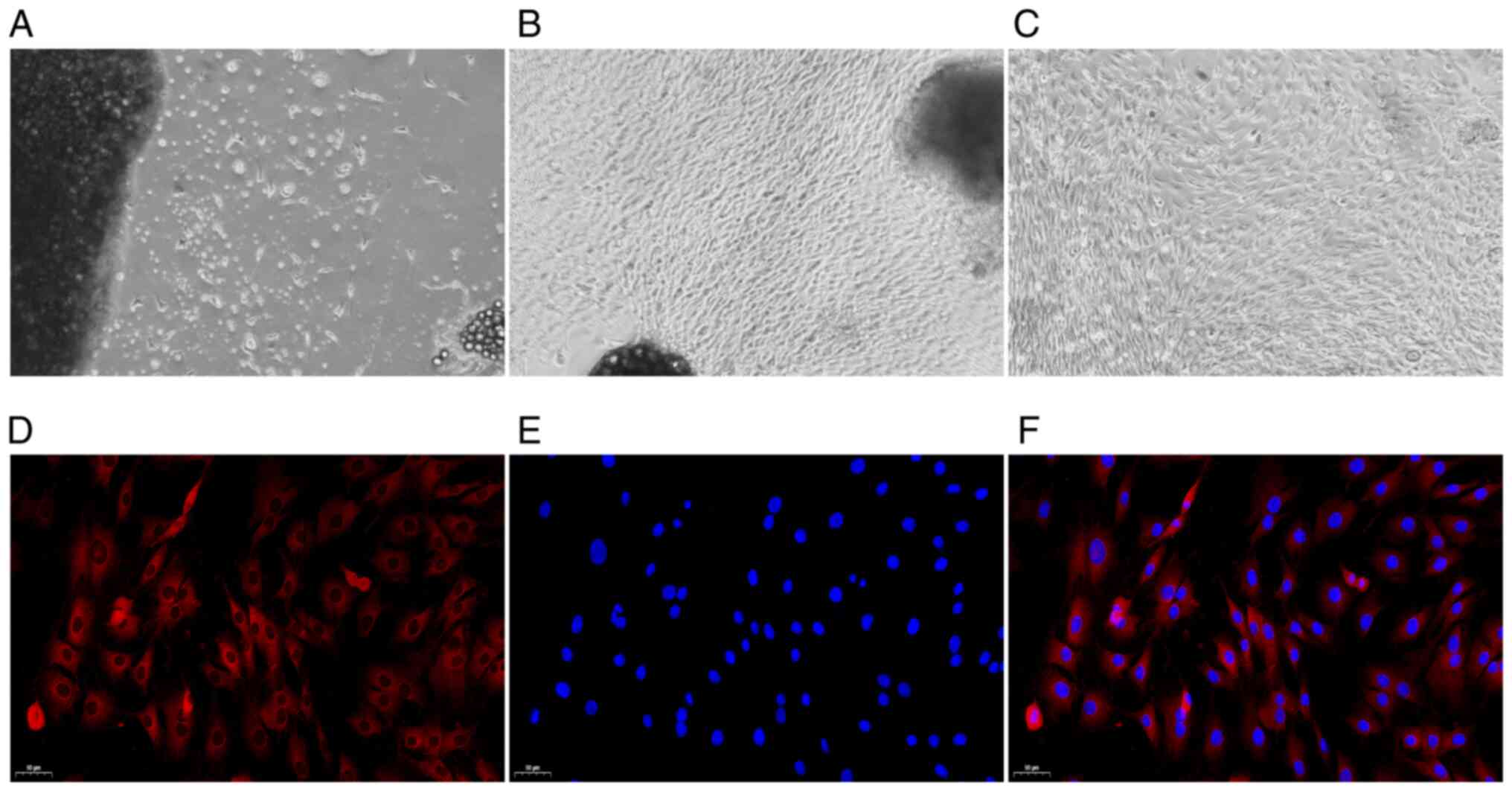
The morphology of the FLSs was observed under a
microscope. Following three days of culture, the primary synovial
cells were gradually separated from the edge of the synovial tissue
(Fig. 1A). After the primary
synovial cells were cultured for 14 days, the synovial tissue
mainly disappeared, a large number of synovial cells were
dissociated, and the cells were oval and spindle-shaped (Fig. 1B). After undergoing three
passages, the synovial cells were uniform and spindle-shaped. Those
cells were then used in the follow-up experiment (Fig. 1C). Under a fluorescence
microscope, the CY3-labeled cytoplasm appeared red in colour, and
the DAPI-labeled nucleus, in blue. As depicted in Fig. 1D-F, all cells were fusiform and
homogeneous in shape, which was consistent with standard FLS cell
morphology (30).
Effects of AST on FLS proliferation, cell
cycle progression and cell ultrastructure
As shown in Fig.
2A, the FLS inhibition rate increased further with the
increasing AST concentration. On the other hand, in comparison with
the inhibition rate at 24 h, the 48- and 72-h inhibition rates were
significantly increased in response to the various AST
concentrations. However, there was no significant difference
between the increase rate at 48 and 72 h. Therefore, the 48-h
incubation time was selected for use in follow-up experiments.
According to the calculations, the IC50 value of AST at
48 h was 48.59 mg/l; thus, 50 mg/l was selected as the drug
concentration for stimulation in the follow-up experiments
(Fig. 2B).
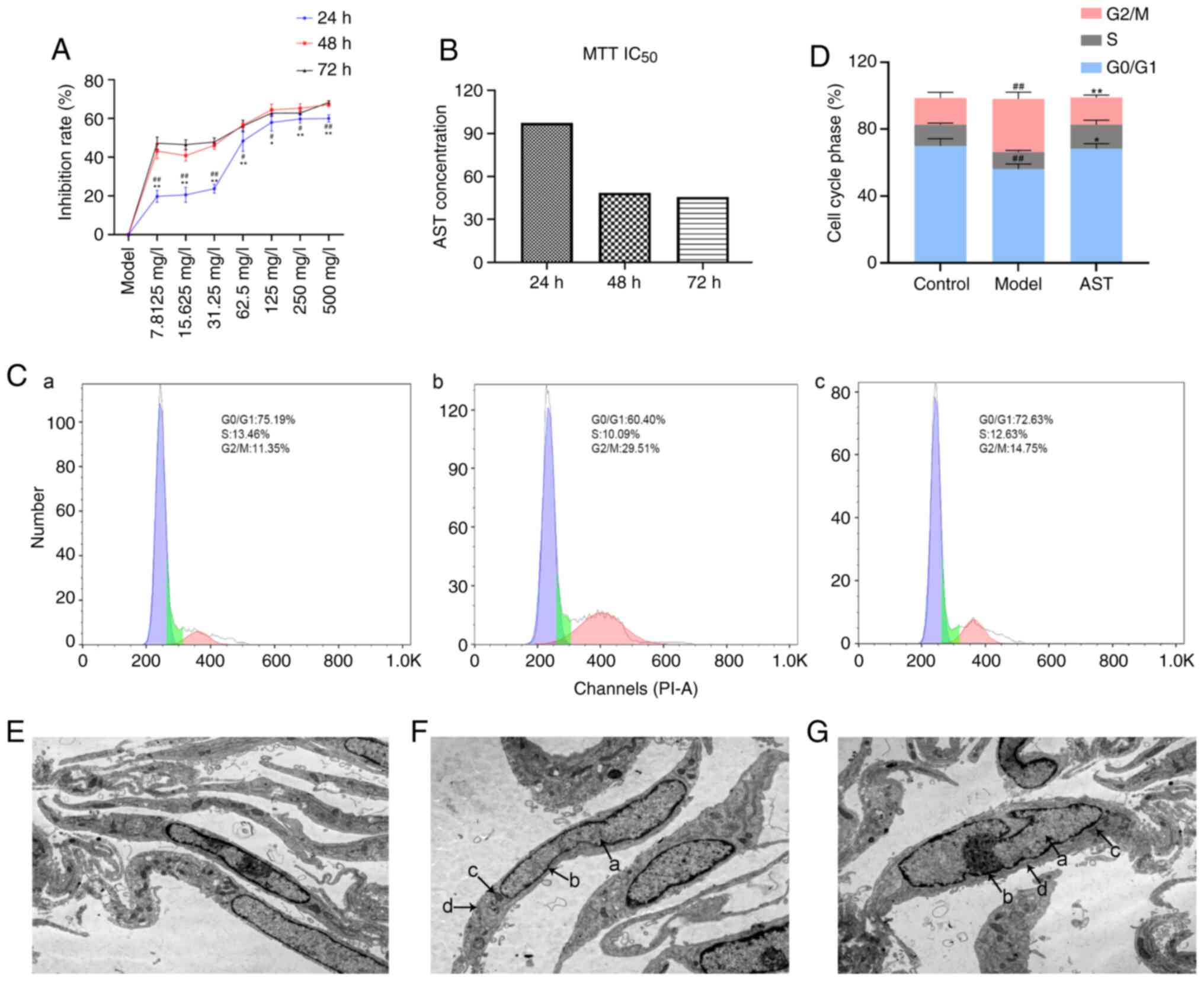 | Figure 2Effect of AST intervention on FLS
proliferation, cell cycle progression and cell ultrastructure. (A)
Inhibition rate in the groups treated with various concentrations
of AST under different reaction times. #P<0.05 and
##P<0.01, compared with the 72-h intervention;
*P<0.05 and **P<0.01, compared with the
48-h intervention. (B) IC50 value calculated by MTT assay. (C) Cell
cycle phase was detected by flow cytometry. (a) Control group; (b)
model group; (c) AST group. (D) Quantification of the cell cycle
results. ##P<0.01, compared with the control group;
**P<0.01, compared with the model group. (E) Control
group (magnification, ×12,000). (F) Model group (magnification,
×12,000). (G) AST group (magnification, ×12,000). a, nucleus; b,
nuclear membrane; c, mitochondria; d, endoplasmic reticulum; ALS,
astragaloside; FLS, fibroblast-like synoviocyte. |
As shown in Fig. 2C
and D, the model group exhibited significantly decreased
numbers of FLSs in the G0/G1 phase, whereas it exhibited
significantly increased numbers of FLSs in the G2/M phase, in
comparison with the control group. In addition, the AST group
exhibited significantly increased numbers of FLSs in the G0/G1
phase, whereas it exhibited significantly decreased numbers of FLSs
in the G2/M phase, in comparison with the control group.
Under a transmission electron microscope, the FLSs
were evidently fusiform. In the control group, the cell membranes
were intact, the nuclei were oval and clearly visible, and a large
number of endoplasmic reticulum and mitochondria could be seen in
the cytoplasm (Fig. 2E). The
model group cell morphology was apparently aberrant compared with
that of the control group, and the model group mainly included FLSs
that were swollen and had damaged, dilated and incomplete cell
membranes. Moreover, the cell nuclear shape was irregular, the
nuclear membrane was blurred and the nucleolus could not be clearly
distinguished in the model group. Additionally, the endoplasmic
reticulum in the cytoplasm was significantly expanded and the
number of mitochondria was significantly decreased in the model
group, when compared with the control group (Fig. 2F). These changes suggested that
the model group FLSs were in an activated state. Following AST
treatment, there was an improvement in the above-mentioned
pathological phenomena in FLSs (Fig.
2G).
Effects of AST on FLS lncRNA LOC100912373
and miR-17-5p expression
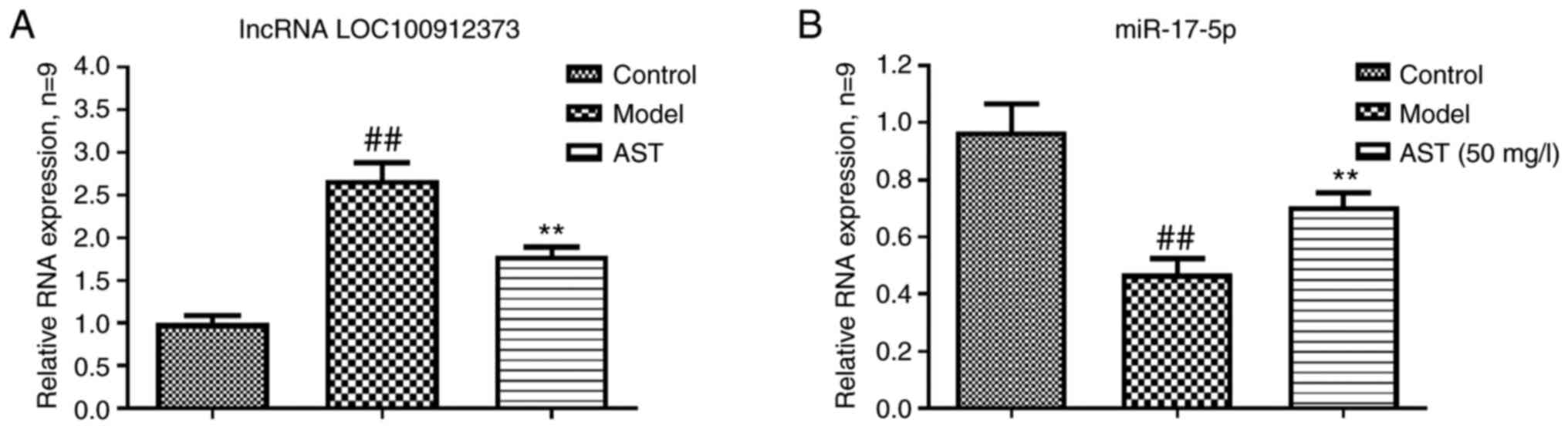
As shown in Fig.
3A, the lncRNA LOC100912373 expression levels in the model
group were significantly higher in contrast to the control group
expression levels. However, the AST group lncRNA LOC100912373
expression levels were significantly lower. As presented in
Fig. 3B, compared with the
control group, the model group exhibited a significantly decreased
miR-17-5p expression. The miR-17-5p expression levels were
significantly increased in the AST group, in comparison to the
model group expression levels.
Effect of AST on FLS PDK1 and AKT
expression
PDK1 controls cell growth, differentiation,
survival, protein translation and glucose metabolism by activating
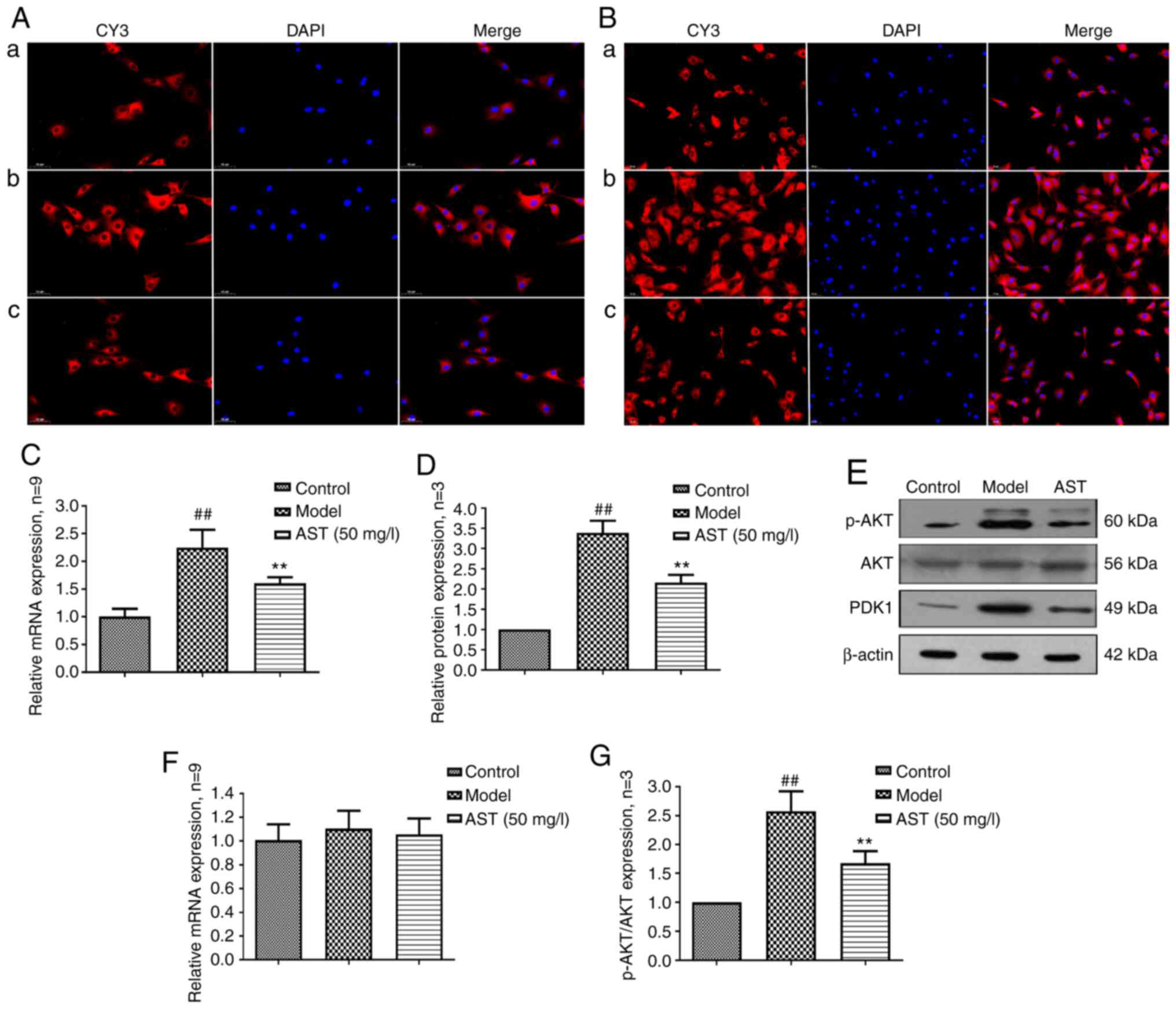
related protein kinases (31,32). As shown in the Fig. 4A, PDK1 was mainly expressed in
the cytoplasm, and PDK1 mRNA and protein expression levels were
significantly increased in the model group. Following AST
treatment, PDK1 mRNA and protein expression was significantly
decreased (Fig. 4B-D).
AKT, a protein kinase that regulates cell survival
and apoptosis, is phosphorylated by PDK1 (33,34). As shown in Fig. 4E, p-AKT was also mainly expressed
in the cytoplasm, and there were no significant differences in the
mRNA and protein expression of AKT among the three groups. However,
when compared with the control group, the model group exhibited a
significantly increased mRNA and protein expression of p-AKT.
Conversely, following AST treatment, p-AKT mRNA and protein
expression was significantly decreased (Fig. 4F-H).
AST reverses the effects of lncRNA
LOC100912373 overexpression on FLSs
To determine whether the effect of AST on FLSs was
achieved by regulating lncRNA LOC100912373 expression, a rescue
experiment was performed. As depicted in Fig. 5A, following transfection with
overexpression plasmid, the expression levels of lncRNA
LOC100912373 were significantly increased according to the
corresponding RT-qPCR results, indicating that the plasmid was
successfully transfected into the FLSs. As shown in Fig. 5B, the rate of FLS inhibition was
significantly decreased following lncRNA LOC100912373
overexpression, in contrast to impact upon the model group FLS
inhibition rate. Of note, after AST treatment was applied to the
lncRNA LOC100912373 overexpression group, the rate of FLS
inhibition was significantly reversed, to some extent.
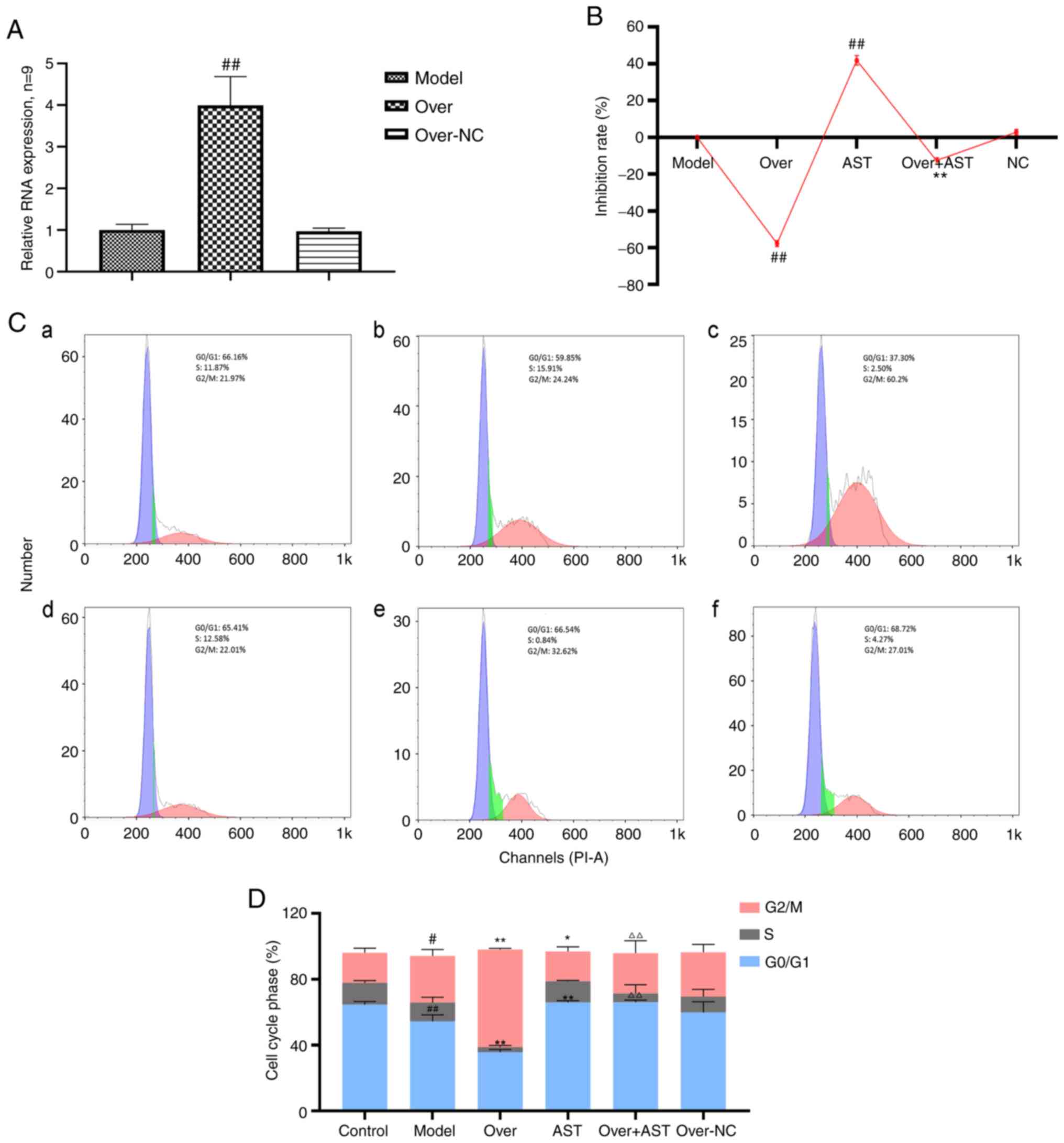 | Figure 5Effect of AST on FLS proliferation
and cell cycle with lncRNA LOC100912373 overexpression. (A) lncRNA
LOC100912373 expression levels following transfection with
overexpression plasmid was detected by RT-qPCR.
##P<0.01, compared with the model group. (B) FLS
inhibition rate between groups, under a 48-h intervention time.
##P<0.01, compared with the model group;
**P<0.01, compared with the lncRNA LOC100912373
overexpression group. (C) Cell cycle phase was detected by flow
cytometry. (a) Control group; (b) model group; (c) lncRNA
LOC100912373 overexpression group; (d) AST group; (e) lncRNA
LOC100912373 overexpression and AST intervention group; (f) NC
group. (D) Cell cycle quantification. #P<0.05,
compared with the control group; ##P<0.01, compared
with the control group; *P<0.05, compared with the
control group; **P<0.01, compared with the model
group; ΔΔP<0.01, compared with the lncRNA
LOC100912373 overexpression group. ALS, astragaloside; FLS,
fibroblast-like synoviocyte. |
As shown in Fig. 5C
and D, the cell cycle progression results were also consistent
with the aforementioned results. The number of FLSs in the G0/G1
phase significantly decreased and the number of FLSs in the G2/M
phase significantly increased following the overexpression of
lncRNA LOC100912373, while AST intervention restored the effects of
lncRNA LOC100912373 overexpression on the FLS cell cycle.
Discussion
Currently, RA treatments mainly focus on mitigating
symptoms and preventing complications rather than reversing or
curing RA. The pathogenesis of RA is extremely complex and involves
environmental, immune, genetic and other factors (35,36), thus rendering the achievement of
RA targeted treatment development a difficult research target.
Currently, increasing attention has been paid to the role of
natural herbal extracts with a probable therapeutic effect on the
treatment of several diseases, including RA (37,38). For example, Astragalus
membranaceus, Tripterygium wilfordii and Radix
Paeoniae Alba have been proven to have certain therapeutic
effects on RA (39-41). The therapeutic effect of
traditional Chinese herbs on diseases mainly depends on a variety
of active components. For example, AST and other ingredients are
components of Astragalus membranaceus. Some studies have
proven that AST may exert beneficial therapeutic effects on a
variety of diseases, through the modulation of immune function and
the inhibition of inflammatory factor production (42,43). In a previously published study by
the authors, it was elucidated that AST may exert a regulatory
effect on differentially expressed lncRNAs during the development
of RA (18). In the present
study, the above-mentioned results were further validated, and it
was attested that AST regulated the expression of lncRNA
LOC100912373, miR-17-5p and PDK1 in the FLSs of rats with AA.
Being a type of RNA that has been linked to cell
biological activities, lncRNAs are indispensable cell function
regulators. In addition to directly regulating cell function,
lncRNAs can also act as a ceRNAs, that regulate miRNA and mRNA
downstream expression, thus affecting RA disease occurrence and
development (44). Zhao et
al (45) confirmed that
lncRNASMAD5-AS1, a ceRNA, inhibits the proliferation of diffuse
large B cell lymphoma via the Wnt/β-catenin pathway by sponging
miR-135b-5p to increase the expression of APC. Similarly, lncRNAs
are also inextricably linked to RA. In contemporary RA research,
lncRNAs have received extensive attention as diagnostic biomarkers
and as potential therapeutic targets for RA (46,47). Some studies have proven that
lncRNA expression is related to the risk and activity of RA. In
addition, lncRNAs can participate in the regulation of relevant
signaling pathways in order to affect FLS proliferation (48,49). In a previously published study,
it was verified that lncRNA LOC100912373 is a differentially
expressed gene that is crucial for the occurrence and development
of RA; in addition, lncRNA LOC100912373 can induce FLS
proliferation by competing with miR-17-5p, so as to promote
PDK1/AKT signaling pathway activation, thereby promoting RA
progression (24). Notably, in
the present study, it was revealed that AST reduced lncRNA
LOC10091237 expression and increased miR-17-5p levels, in order to
affect PDK1 axis activation in the FLSs of rats with AA.
PDK1 is a serine/threonine kinase, which belongs to
the AGC kinase family. AKT, also known as protein kinase B (PKB),
is the downstream target gene of PDK1. Some studies have
demonstrated that the PDK1/AKT signaling pathway is involved in the
occurrence and development of RA, through the regulation of several
cell biological processes, such as cell proliferation, cell cycle
progression and differentiation (50,51). More specifically, as an important
protein of the PI3K/AKT signaling pathway, PDK1 can bind to the
product of activated PI3K, PIP3. Afterwards, PDK1/PIP3 complex
transfers to the cell membrane, in order to phosphorylate AKT and
activate AKT signaling pathway (52). AKT signaling pathway activation
can accelerate the release of pro-inflammatory factors and thus may
lead to abnormal cell biological processes, such as excessive cell
proliferation, decrease in cell proliferation and increase in cell
necrosis (53,54). In general, it is known that
excessive FLS proliferation is an important principal pathological
feature of RA. Because cell proliferation is controlled through the
regulation of the cell cycle, abnormal cell cycle progression must
occur during the development of RA. In a previously published
research by Fu et al (55), PDK1 promoted proliferation and
inhibited apoptosis in human spermatogonial stem cells via the
PDK1/KDR/ZNF367, ERK1/2 and AKT pathway. In the present study, it
was demonstrated that AST can suppress excessive proliferation and
restore the normal cell cycle progression of the FLSs in AA rats.
In addition, PDK1 and p-AKT mRNA and protein expression levels were
significantly decreased in FLSs following AST treatment. Moreover,
in order to further validate the above-mentioned results, a rescue
assay was carried. Thus, it was confirmed that the excessive
proliferation and the establishment of abnormal cell cycle
progression caused by lncRNA LOC100912373 overexpression in FLSs
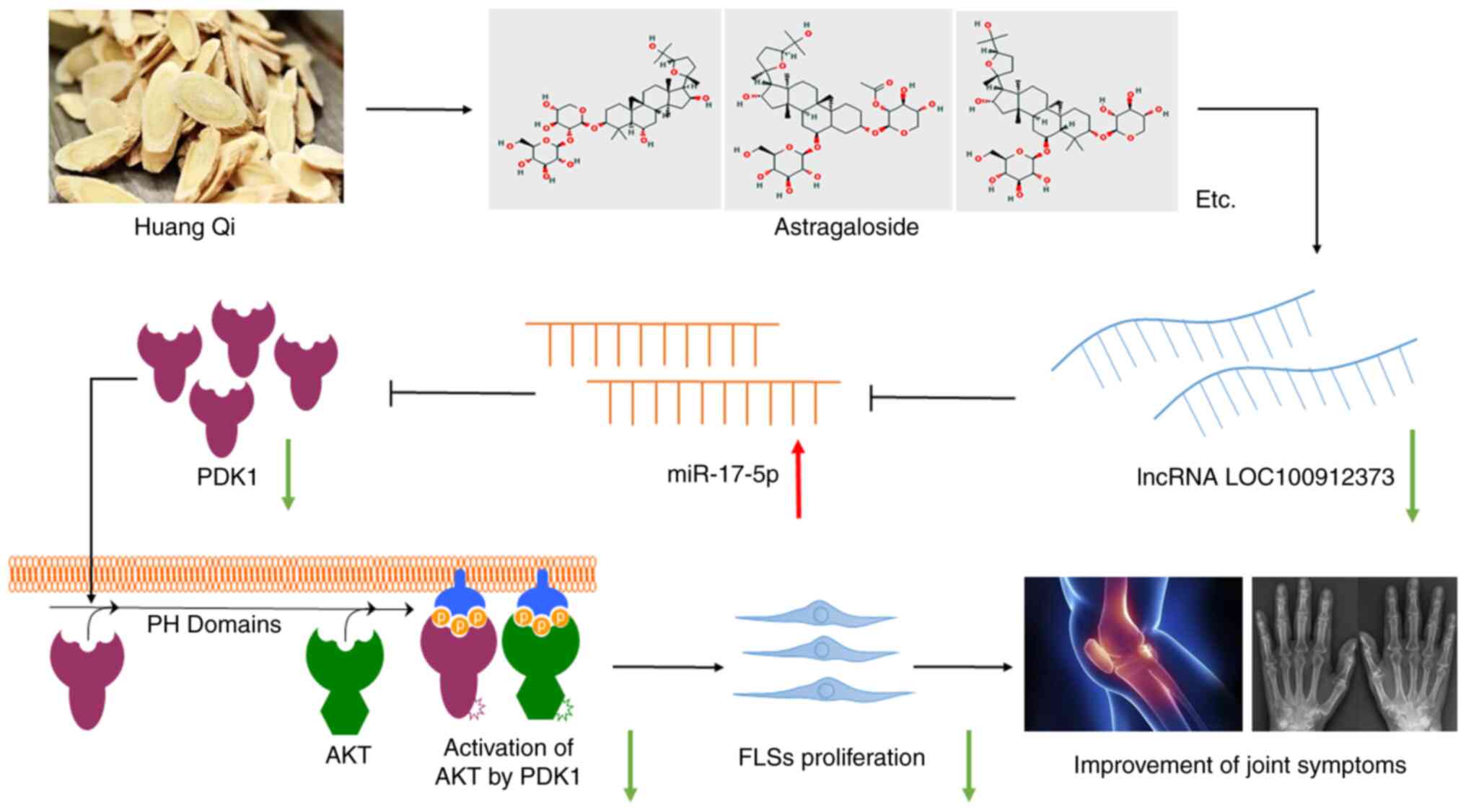
could be reversed by AST treatment (Fig. 6).
In conclusion, the results of the present study
revealed that AST suppressed the excessive proliferation of FLSs in
rats with AA, through the inhibition of lncRNA LOC100912373
expression. miR-17-5p binds to PDK1, and thereby prevents PDK1/AKT
signaling pathway activation. Therefore, AST may serve as a
potential therapeutic for RA, and the results of the present study
may provide novel insight into future research for the use of
traditional Chinese medicines for the prevention and treatment of
RA.
Supplementary Data
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
JL made substantial contributions to the conception
and design of the study. HJ, CF, YL and XC performed the
experiments. HJ and CF contributed to data acquisition, and data
analysis and interpretation. HJ revised the manuscript critically
for important intellectual content. HJ and JL confirm the
authenticity of all the raw data. All authors agreed to be
accountable for all aspects of the work in ensuring that questions
related to the accuracy or integrity of the work are appropriately
investigated and resolved. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
All animal experiments were approved by the Animal
Ethics committee of Anhui University of Chinese Medicine (Hefei,
China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
The authors are grateful to Mr. Qiang Fan (Ao Ji
Bio-tech Co. Ltd., Shanghai, China) for providing assistance with
the data analysis.
Funding
The present study was supported by the National Natural Science
Foundation of China (grant no. 81873139), the 12th Batch of '115'
Innovation Team of Anhui Province [Anhui Talent Office (2019) no.
1], the Key Research and Development Program of Anhui Province,
China (no. 201904a07020004) and the Special Funds for Provincial
TCM Development (no. 2016ZYZJ01).
References
|
1
|
Smolen JS, Aletaha D and McInnes IB:
Rheumatoid arthritis. Lancet. 388:2023–2038. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ham S, Bae JB, Lee S, Kim BJ, Han BG, Kwok
SK and Roh TY: Epigenetic analysis in rheumatoid arthritis
synoviocytes. Exp Mol Med. 51:1–13. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li X, Yuan K, Zhu Q, Lu Q, Jiang H, Zhu M,
Huang G and Xu A: Andrographolide ameliorates rheumatoid arthritis
by regulating the apoptosis-NETosis balance of neutrophils. Int J
Mol Sci. 20:50352019. View Article : Google Scholar
|
|
4
|
Blunk I, Thomsen H, Reinsch N, Mayer M,
Försti A, Sundquist J, Sundquist K and Hemminki K: Genomic
imprinting analyses identify maternal effects as a cause of
phenotypic variability in type 1 diabetes and rheumatoid arthritis.
Sci Rep. 10:115622020. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mameli G, Erre GL, Caggiu E, Mura S, Cossu
D, Bo M, Cadoni ML, Piras A, Mundula N, Colombo E, et al:
Identification of a HERV-K env surface peptide highly recognized in
rheumatoid arthritis (RA) patients: A cross-sectional case-control
study. Clin Exp Immunol. 189:127–131. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Muschter D, Schäfer N, Stangl H, Straub RH
and Grässel S: Sympathetic neurotransmitters modulate
osteoclastogenesis and osteoclast activity in the context of
collagen-induced arthritis. PLoS One. 10:e01397262015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lowin T, Anssar TM, Bäuml M, Classen T,
Schneider M and Pongratz G: Positive and negative cooperativity of
TNF and Interferon-γ in regulating synovial fibroblast function and
B cell survival in fibroblast/B cell co-cultures. Sci Rep.
10:7802020. View Article : Google Scholar
|
|
8
|
Zhao J, Chen B, Peng X, Wang C, Wang K,
Han F and Xu J: Quercetin suppresses migration and invasion by
targeting miR-146a/GATA6 axis in fibroblast-like synoviocytes of
rheumatoid arthritis. Immunopharmacol Immunotoxicol. 42:221–227.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lucchino B, Spinelli FR, Iannuccelli C,
Guzzo MP, Conti F and Di Franco M: Mucosa-environment interactions
in the pathogenesis of rheumatoid arthritis. Cells. 8:7002019.
View Article : Google Scholar
|
|
10
|
Cappelli LC, Thomas MA, Bingham CO III,
Shah AA and Darrah E: Immune checkpoint inhibitor-induced
inflammatory arthritis as a model of autoimmune arthritis. Immunol
Rev. 294:106–123. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Matta R, Wang X, Ge H, Ray W, Nelin LD and
Liu Y: Triptolide induces anti-inflammatory cellular responses. Am
J Transl Res. 1:267–282. 2009.PubMed/NCBI
|
|
12
|
Guo Q, Mao X, Zhang Y, Meng S, Xi Y, Ding
Y, Zhang X, Dai Y, Liu X, Wang C, et al: Guizhi-Shaoyao-Zhimu
decoction attenuates rheumatoid arthritis partially by reversing
inflammation-immune system imbalance. J Transl Med. 14:1652016.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu XY, Xu L, Wang Y, Li JX, Zhang Y,
Zhang C, Wang SS and Zhang XM: Protective effects of total
flavonoids of astragalus against adjuvant-induced arthritis in rats
by regulating OPG/RANKL/NF-κB pathway. Int Immunopharmacol.
44:105–114. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pu X, Ma X, Liu L, Ren J, Li H, Li X, Yu
S, Zhang W and Fan W: Structural characterization and antioxidant
activity in vitro of polysaccharides from angelica and astragalus.
Carbohydr Polym. 137:154–164. 2016. View Article : Google Scholar
|
|
15
|
Dong Z, Zhao P, Xu M, Zhang C, Guo W, Chen
H, Tian J, Wei H, Lu R and Cao T: Astragaloside IV alleviates heart
failure via activating PPARα to switch glycolysis to fatty acid
β-oxidation. Sci Rep. 7:26912017. View Article : Google Scholar
|
|
16
|
Chen L, Xie ZY, Liu L, Zhu L, Wang F, Fan
P, Sinkemani A, Zhang C, Hong X and Wu XT: Nuclear factor-kappa
B-dependent X-box binding protein 1 signalling promotes the
proliferation of nucleus pulposus cells under tumour necrosis
factor alpha stimulation. Cell Prolif. 52:e125422019. View Article : Google Scholar
|
|
17
|
Chen X, Chen X, Gao J, Yang H, Duan Y,
Feng Y, He X, Gong X, Wang H, Wu X and Chang J: Astragaloside III
enhances anti-tumor response of NK cells by elevating NKG2D and
IFN-γ. Front Pharmacol. 10:8982019. View Article : Google Scholar
|
|
18
|
Jiang H, Wu FR, Liu J, Qin XJ, Jiang NN
and Li WP: Effect of astragalosides on long non-coding RNA
expression profiles in rats with adjuvant-induced arthritis. Int J
Mol Med. 44:1344–1356. 2019.PubMed/NCBI
|
|
19
|
Xu F, Jin L, Jin Y, Nie Z and Zheng H:
Long noncoding RNAs in autoimmune diseases. J Biomed Mater Res A.
107:468–475. 2019. View Article : Google Scholar
|
|
20
|
Zhang TP, Zhu BQ, Tao SS, Fan YG, Li XM,
Pan HF and Ye DQ: Long non-coding RNAs genes polymorphisms and
their expression levels in patients with rheumatoid arthritis.
Front Immunol. 10:25292019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jiang H, Liu J, Fan C, Wang J and Li W:
lncRNAS56464.1 as a ceRNA promotes the proliferation of
fibroblast-like synoviocytes in experimental arthritis via the Wnt
signaling pathway and sponges miR-152-3p. Int J Mol Med. 47:172021.
View Article : Google Scholar :
|
|
22
|
Bi X, Guo XH, Mo BY, Wang ML, Luo XQ, Chen
YX, Liu F, Olsen N, Pan YF and Zheng SG: LncRNA PICSAR promotes
cell proliferation, migration and invasion of fibroblast-like
synoviocytes by sponging miRNA-4701-5p in rheumatoid arthritis.
EBioMedicine. 50:408–420. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jiang H, Qin XJ, Li WP, Ma R, Wang T and
Li ZQ: LncRNAs expression in adjuvant-induced arthritis rats
reveals the potential role of LncRNAs contributing to rheumatoid
arthritis pathogenesis. Gene. 593:131–142. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fan C, Cui X, Chen S, Huang S and Jiang H:
LncRNA LOC100912373 modulates PDK1 expression by sponging miR-17-5p
to promote the proliferation of fibroblast-like synoviocytes in
rheumatoid arthritis. Am J Transl Res. 12:7709–7723. 2020.
|
|
25
|
Jiang H, Liu J, Wang T, Gao JR, Sun Y,
Huang CB, Meng M and Qin XJ: Urinary metabolite profiling provides
potential differentiation to explore the mechanisms of
adjuvant-induced arthritis in rats. Biomed Chromatogr.
30:1397–1405. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhu L, Li J, Guo L, Yu X, Wu D, Luo L, Zhu
L, Chen W, Chen C, Ye C and Zhang D: Activation of NALP1
inflammasomes in rats with adjuvant arthritis; a novel therapeutic
target of carboxyamidotriazole in a model of rheumatoid arthritis.
Br J Pharmacol. 172:3446–3459. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Luo C, Liang JS, Gong J, Zhang HL, Feng
ZJ, Yang HT, Zhang HB and Kong QH: miRNA-31 over-expression improve
synovial cells apoptosis induced by RA. Bratisl Lek Listy.
119:355–360. 2018.PubMed/NCBI
|
|
28
|
Chen L, Feng L, Wang X, Du J, Chen Y, Yang
W, Zhou C, Cheng L, Shen Y, Fang S, et al: Mesencephalic
astrocyte-derived neurotrophic factor is involved in inflammation
by negatively regulating the NF-κB pathway. Sci Rep. 5:81332015.
View Article : Google Scholar
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
30
|
Yang Y, Dong Q and Li R: Matrine induces
the apoptosis of fibroblast-like synoviocytes derived from rats
with collagen-induced arthritis by suppressing the activation of
the JAK/STAT signaling pathway. Int J Mol Med. 39:307–316. 2017.
View Article : Google Scholar :
|
|
31
|
Siu MKY, Jiang YX, Wang JJ, Leung THY, Ngu
SF, Cheung ANY, Ngan HYS and Chan KKL: PDK1 promotes ovarian cancer
metastasis by modulating tumor-mesothelial adhesion, invasion, and
angiogenesis via α5β1 integrin and JNK/IL-8 signaling. Oncogenesis.
9:242020. View Article : Google Scholar
|
|
32
|
He J, Wang Y, Liu T, Liu G, Chen S, Li Q,
Quan Y, Yang H, Feng J, Wang S, et al: Stage-specific requirement
of kinase PDK1 for NK cells development and activation. Cell Death
Differ. 26:1918–1928. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Song X, Wang Z, Liang H, Zhang W, Ye Y, Li
H, Hu Y, Zhang Y, Weng H, Lu J, et al: Dioscin induces gallbladder
cancer apoptosis by inhibiting ROS-mediated PI3K/AKT signalling.
Int J Biol Sci. 13:782–793. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Saji M, Kim CS, Wang C, Zhang X, Khanal T,
Coombes K, La Perle K, Cheng SY, Tsichlis PN and Ringel MD: Akt
isoform-specific effects on thyroid cancer development and
progression in a murine thyroid cancer model. Sci Rep.
10:183162020. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bunte K and Beikler T: Th17 cells and the
IL-23/IL-17 axis in the pathogenesis of periodontitis and
immune-mediated inflammatory diseases. Int J Mol Sci. 20:33942019.
View Article : Google Scholar :
|
|
36
|
Bodkhe R, Balakrishnan B and Taneja V: The
role of microbiome in rheumatoid arthritis treatment. Ther Adv
Musculoskelet Dis. 11:1759720X198446322019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang Y, Zhou J, Tang C, Yu J, Zhu W, Guo J
and Wang Y: Positive effect of astragaloside IV on neurite
outgrowth via talin-dependent integrin signaling and microfilament
force. J Cell Physiol. 236:2156–2168. 2021. View Article : Google Scholar
|
|
38
|
Li N, Wu K, Feng F, Wang L, Zhou X and
Wang W: Astragaloside IV alleviates silica-induced pulmonary
fibrosis via inactivation of the TGF-β1/Smad2/3 signaling pathway.
Int J Mol Med. 47:162021. View Article : Google Scholar
|
|
39
|
Wang B and Chen MZ: Astragaloside IV
possesses antiarthritic effect by preventing interleukin 1β-induced
joint inflammation and cartilage damage. Arch Pharm Res.
37:793–802. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wang S, Zuo S, Liu Z, Ji X, Yao Z and Wang
X: Study on the efficacy and mechanism of triptolide on treating
TNF transgenic mice with rheumatoid arthritis. Biomed Pharmacother.
106:813–820. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Xie YF, Feng WW, Liu MC, Xie J, Yu L, Gong
XH, Li YX and Peng C: Investigation of efficacy enhancing and
toxicity reducing mechanism of combination of aconiti lateralis
radix praeparata and paeoniae radix alba in adjuvant-induced
arthritis rats by metabolomics. Evid Based Complement Alternat Med.
2019:98648412019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Jia Q, Wang T, Wang X, Xu H, Liu Y, Wang
Y, Shi Q and Liang Q: Astragalin suppresses inflammatory responses
and bone destruction in mice with collagen-induced arthritis and in
human fibroblast-like synoviocytes. Front Pharmacol. 10:942019.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Yan MM, Chen CY, Zhao BS, Zu YG, Fu YJ,
Liu W and Efferth T: Enhanced extraction of astragalosides from
radix astragali by negative pressure cavitation-accelerated enzyme
pretreatment. Bioresour Technol. 101:7462–7471. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Salmena L, Poliseno L, Tay Y, Kats L and
Pandolfi PP: A ceRNA hypothesis: The rosetta stone of a hidden RNA
language? Cell. 146:353–358. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhao CC, Jiao Y, Zhang YY, Ning J, Zhang
YR, Xu J, Wei W and Kang-Sheng G: Lnc SMAD5-AS1 as ceRNA inhibit
proliferation of diffuse large B cell lymphoma via Wnt/β-catenin
pathway by sponging miR-135b-5p to elevate expression of APC. Cell
Death Dis. 10:2522019. View Article : Google Scholar
|
|
46
|
Mo BY, Guo XH, Yang MR, Liu F, Bi X, Liu
Y, Fang LK, Luo XQ, Wang J, Bellanti JA, et al: Long non-coding RNA
GAPLINC promotes tumor-like biologic behaviors of fibroblast-like
synoviocytes as MicroRNA sponging in rheumatoid arthritis patients.
Front Immunol. 9:7022018. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yan S, Wang P, Wang J, Yang J, Lu H, Jin
C, Cheng M and Xu D: Long non-coding RNA HIX003209 promotes
inflammation by sponging miR-6089 via TLR4/NF-κB signaling pathway
in rheumatoid arthritis. Front Immunol. 10:22182019. View Article : Google Scholar
|
|
48
|
Li G, Liu Y, Meng F, Xia Z, Wu X, Fang Y,
Zhang C, Zhang Y and Liu D: LncRNA MEG3 inhibits rheumatoid
arthritis through miR-141 and inactivation of AKT/mTOR signalling
pathway. J Cell Mol Med. 23:7116–7120. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Sun L, Tu J, Liu C, Pan A, Xia X and Chen
X: Analysis of lncRNA expression profiles by sequencing reveals
that lnc-AL928768.3 and lnc-AC091493.1 are novel biomarkers for
disease risk and activity of rheumatoid arthritis.
Inflammopharmacology. 28:437–450. 2020. View Article : Google Scholar
|
|
50
|
Sun C, Sun Y, Jiang D, Bao G, Zhu X, Xu D,
Wang Y and Cui Z: PDK1 promotes the inflammatory progress of
fibroblast-like synoviocytes by phosphorylating RSK2. Cell Immunol.
315:27–33. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Xu H, He Y, Yang X, Liang L, Zhan Z, Ye Y,
Yang X, Lian F and Sun L: Anti-malarial agent artesunate inhibits
TNF-alpha-induced production of proinflammatory cytokines via
inhibition of NF-kappaB and PI3 kinase/Akt signal pathway in human
rheumatoid arthritis fibroblast-like synoviocytes. Rheumatology
(Oxford). 46:920–926. 2007. View Article : Google Scholar
|
|
52
|
Ichikawa R, Kawasaki R, Iwata A, Otani S,
Nishio E, Nomura H and Fujii T: MicroRNA-126-3p suppresses HeLa
cell proliferation, migration and invasion, and increases apoptosis
via the PI3K/PDK1/AKT pathway. Oncol Rep. 43:1300–1308.
2020.PubMed/NCBI
|
|
53
|
Shen X, Yu Y, Ma P, Luo Z, Hu Y, Li M, He
Y, Zhang Y, Peng Z, Song G and Cai K: Titania nanotubes promote
osteogenesis via mediating crosstalk between macrophages and MSCs
under oxidative stress. Colloids Surf B Biointerfaces. 180:39–48.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Chen D, Zeng S, Huang M, Xu H, Liang L and
Yang X: Role of protein arginine methyltransferase 5 in
inflammation and migration of fibroblast-like synoviocytes in
rheumatoid arthritis. J Cell Mol Med. 21:781–790. 2017. View Article : Google Scholar
|
|
55
|
Fu H, Zhang W, Yuan Q, Niu M, Zhou F, Qiu
Q, Mao G, Wang H, Wen L, Sun M, et al: PAK1 promotes the
proliferation and inhibits apoptosis of human spermatogonial stem
cells via PDK1/KDR/ZNF367 and ERK1/2 and AKT pathways. Mol Ther
Nucleic Acids. 12:769–786. 2018. View Article : Google Scholar : PubMed/NCBI
|