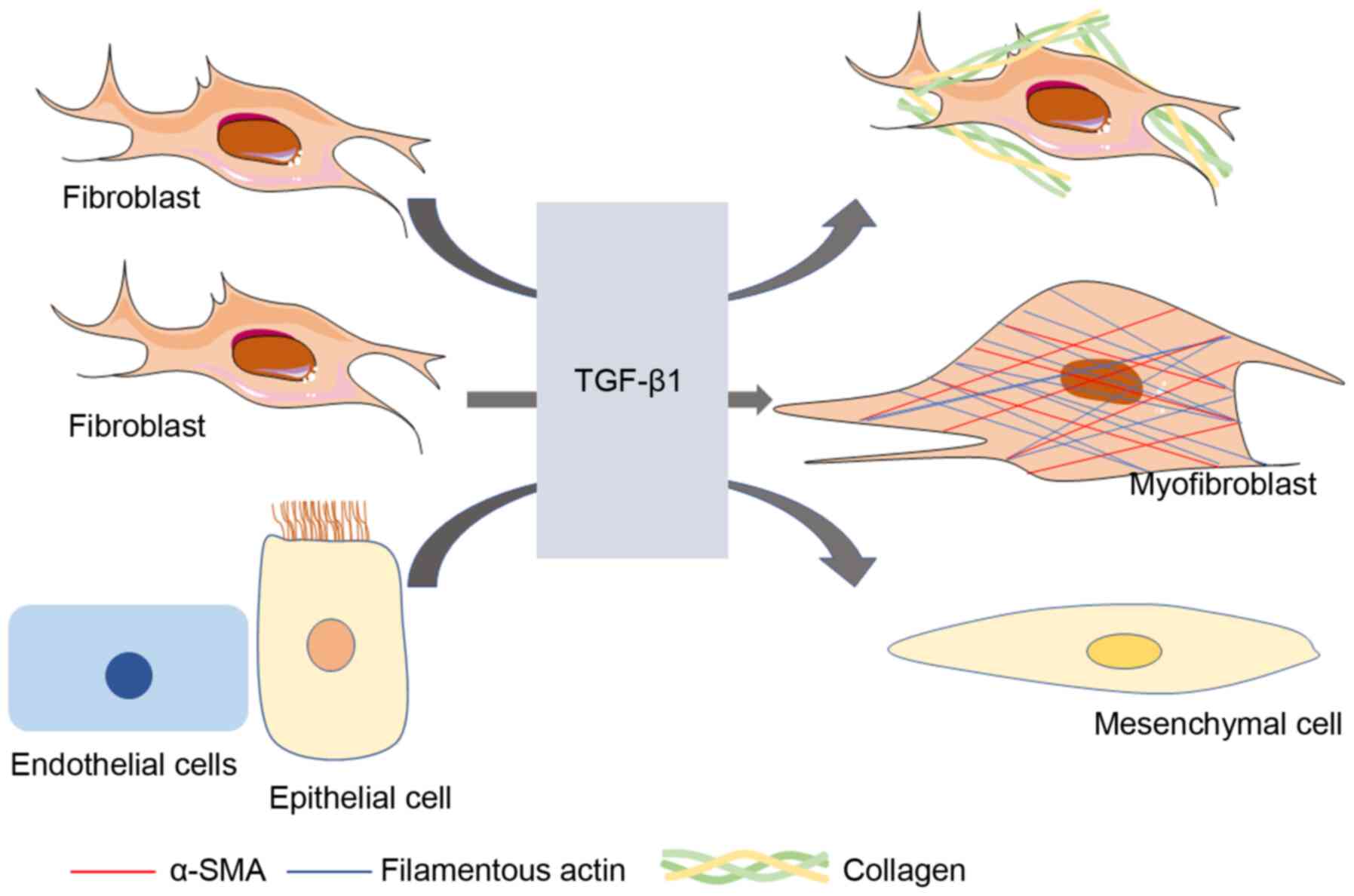
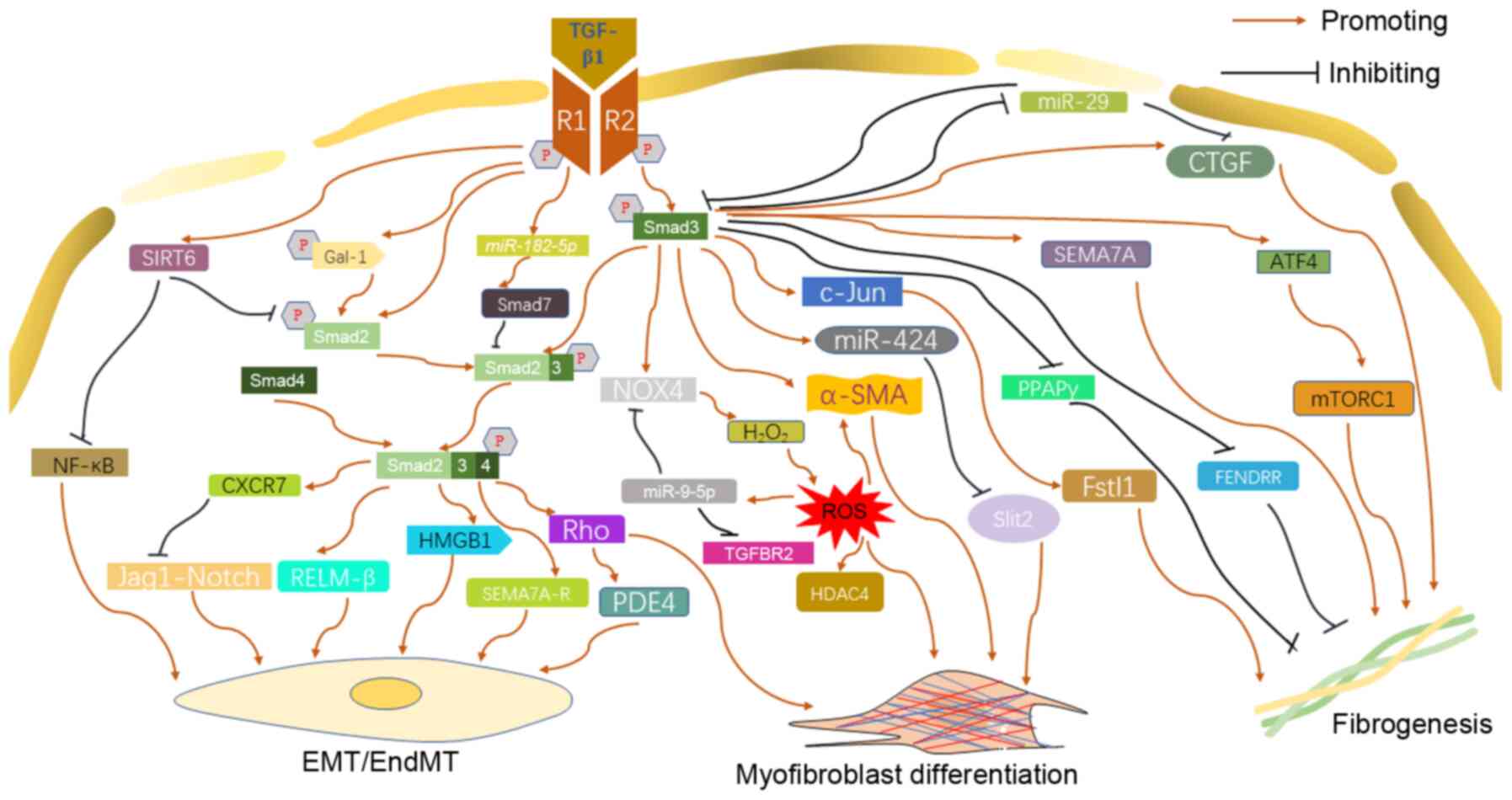
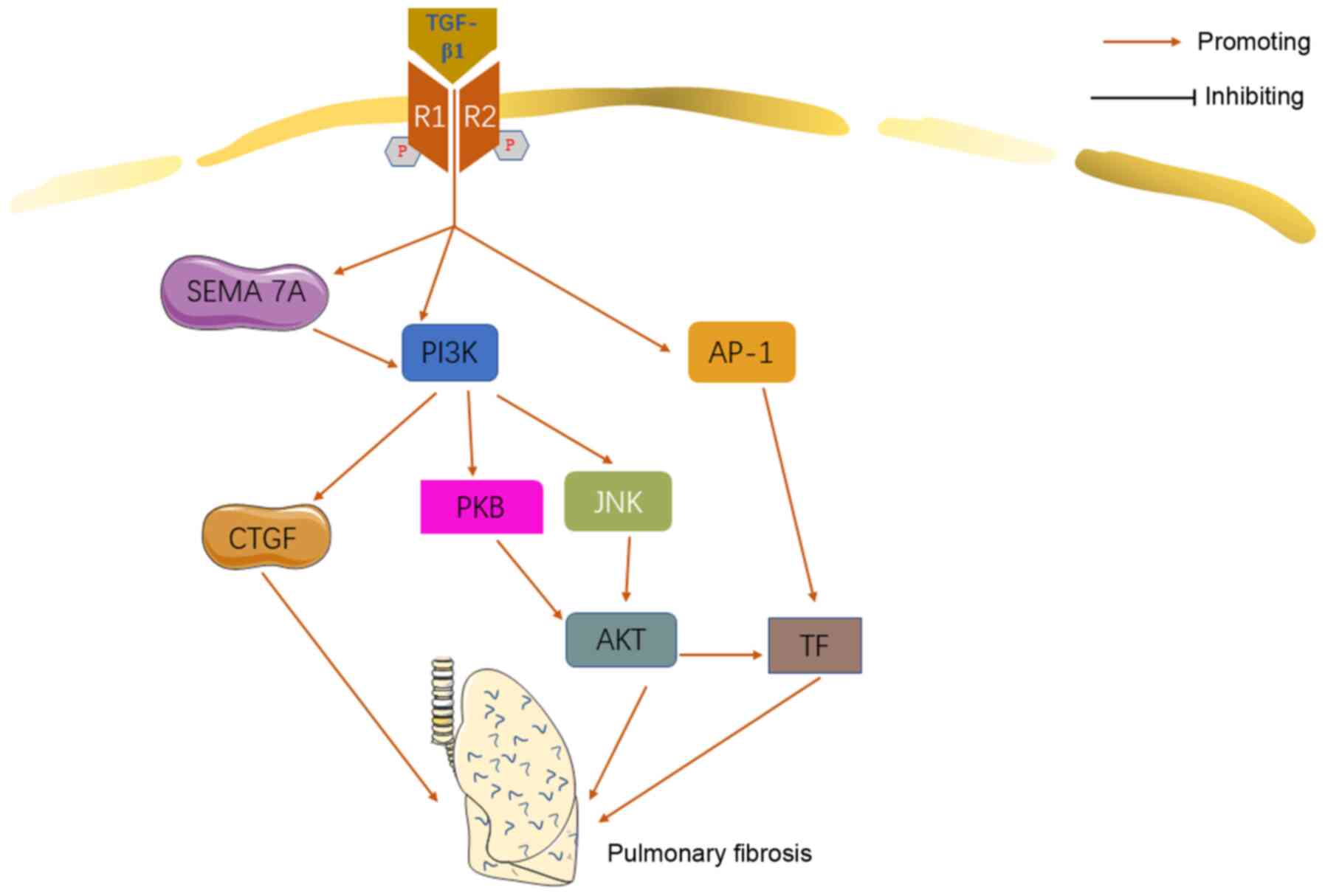
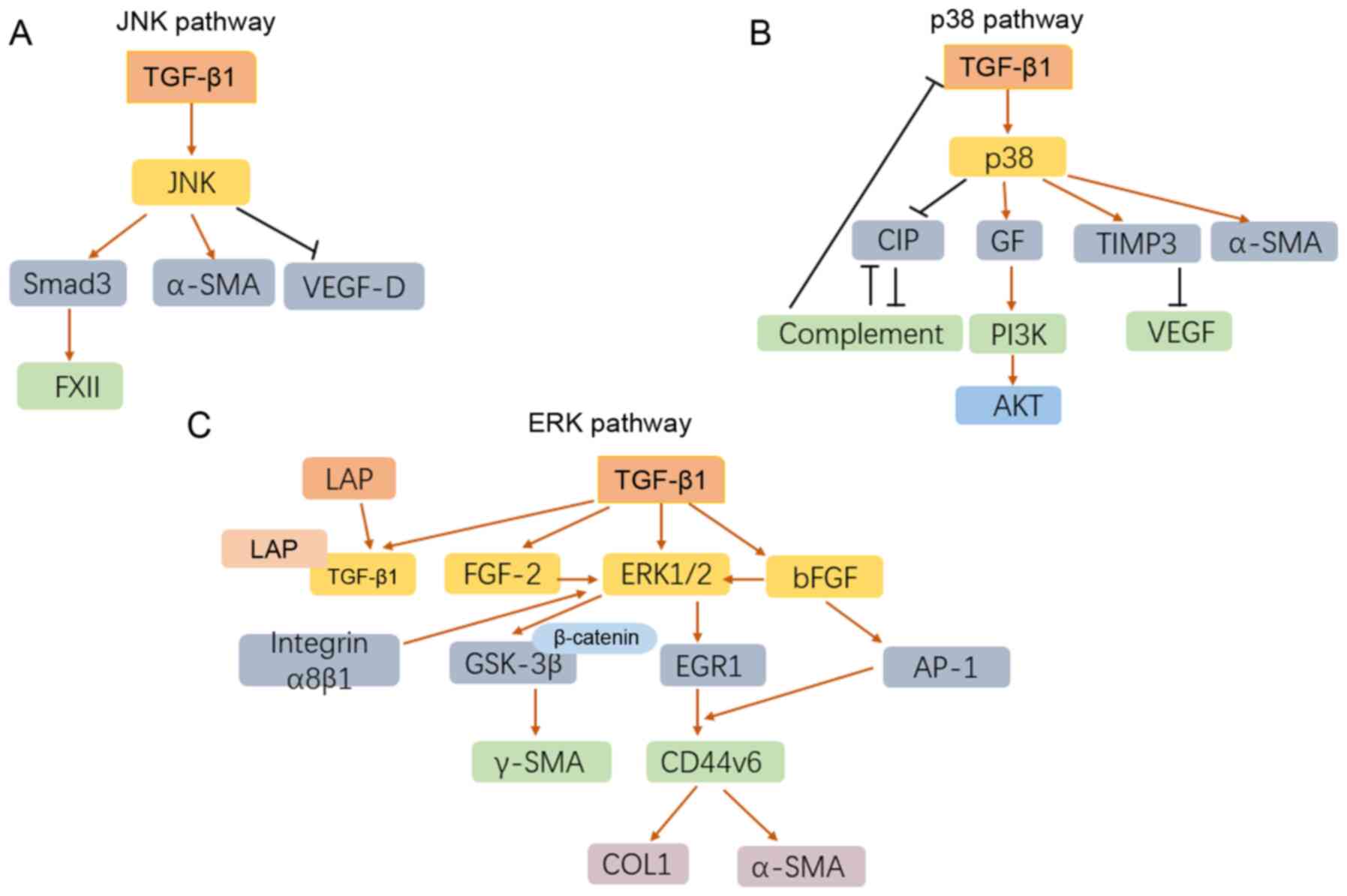
Idiopathic pulmonary fibrosis (IPF) is a chronic,
lethal and irreversible disease, which is characterized by
fibroblast proliferation and excessive deposition of extracellular
matrix in the lung (1,2). It was reported that the overall
survival of the patients who were diagnosed with IPF was 3-5 years
(3). The annual incidence of IPF
is between 0.22 and 7.4 per 100,000 individuals in Europe and North
America, but is lower in East Asia and South American (4). The incidence and prevalence of IPF
increase with age and are higher in men (Tables I and II), which have been on the increase in
recent years (1,5,6).
Smoking, silica, and lampblack may be high risk factors for IPF
(7). IPF can cause many symptoms
such as dyspneal breathlessness, and chest discomfort, which does
great harm to human and induces tremendous economic burden
(8).
At present, many studies have focused on the
pathogenesis mechanisms, which mainly include the Smad, MAPK, and
ERK signaling pathways (9). Of
these mechanisms TGF-β1 is of critical significance (10). Researchers have conducted
pharmacological studies on TGF-β1 in IPF, and some new drugs
targeting TGF-β1-relevant signaling pathways have been developed.
Such drugs include Nimbolide (11), Tanshinone IIA (Tan IIA) (12), methylsulfonylmethane (13) and Isoliquiritigenin (ISL)
(14). However, since none of
these medicines can successfully treat IPF, lung transplantation
remains the primary method of treatment (15).
Both basic research and clinical research have
proven that TGF-β1 plays an important role in the pathogenesis of
IPF (Table III). However, no
review systematically summarizing and discussing the role of TGF-β1
and relevant pathways in IPF has currently been published. The aim
of the present review was to summarize the studies concerning the
role of TGF-β1 in the development of IPF in recent decades
(16) (Fig. 1). The findings may help
researchers to grasp the latest progress in the pathogenesis of IPF
related to TGF-β1 and to provide novel targets and a theoretical
basis for the development of IPF clinical drugs.
The Smads family comprises three subfamilies,
including five receptor-activated Smads (R-Smads), one common
mediator Smad (Co-Smad) and two inhibitory Smads (I-Smads). Smad6
and Smad7 are the third type of Smads known as 'inhibitory Smads'
or 'anti-Smads'. They are structurally different from other members
of the family, and have been proven to be inhibitors of the Smad
signaling pathway by disturbing the activation of R-Smads (17). Usually, TGF-β1 activates Smads
through the transmembrane receptor serine/threonine kinase,
successively regulating the transcription of target genes (18).
When TGF-β type I receptor kinase was activated by
TGF-β1 signal, R-Smads (Smad2 and Smad3) were phosphorylated; of
note is that Smad3 is more sensitive to TGF-β1 than Smad2 (19). Activated Smad2 and Smad3 form a
complex, which combines with the Co-Smad (Smad4) and transfers into
the nucleus to regulate the expression of target genes (20). The contribution of TGF-β1/Smad
signaling pathway to IPF is mainly dependent on the following three
processes: Myofibroblast differentiation, EMT/EndMT, and
fibrogenesis.
TGF-β1 regulates the terminal differentiation of
human lung fibroblasts (HLF) and promotes the synthesis of
fibroblast extracellular matrix (21). Additionally, TGF-β1/Smad3 is the
chief signaling pathway that regulates fibroblast differentiation
(22,23). Transcription of α-smooth muscle
actin (α-SMA), a target of myofibroblasts, was stimulated by TGF-β1
via a Smad3-, but not Smad2, dependent manner, resulting in the
increased expression of α-SMA protein in human fetal lung
fibroblasts (HFLF) (22).
However, Deng et al (24)
demonstrated that although Smad3 can be activated by TGF-β1 in HLF,
the former did not affect the expression of collagen I or α-SMA.
Treating fibroblasts with TGF-β1 could increase the expression of
galectin-1 (Gal-1), which phosphorylated Smad2 and enhanced the
nuclear retention of Smad2, promoting myofibroblast differentiation
and accelerating fibrosis (25).
TGF-β1 induced upregulation of miR-424 through the Smad3-denpendent
signaling pathway, which inhibited the expression of Slit2, an
inhibitory protein on TGF-β1 profibrogenic signaling. As a result,
miR-424 acts as a positive feedback regulator of the TGF-β1
signaling pathway, promoting the myofibroblast differentiation of
HLF (26). Interestingly, with
the treatment of miR-424 inhibitor, Smad3 phosphorylation by TGF-β1
was reduced in HLFs, indicating miR-424 as a positive feedback
regulator of TGF-β1/Smad3 synergistically (26). Previous findings demonstrated
TGF-β1/Smad3-induced NADPH oxidase 4 (NOX4) mediated the production
of H2O2, which was necessary for
myofibroblast differentiation of lung mesenchymal cells, providing
novel insight into the therapeutic targeting in IPF (27,28). In addition, TGF-β1 was reported
to accelerate lung fibrosis by stimulating the production of ROS
depending on NOX-4, and the produced ROS promoted the nuclear
export of histone deacetylase 4 (HDAC4) and formation of α-SMA
fiber in normal human lung fibroblasts (NHLFs) (29). Furthermore, following exposure to
ROS, the expression of miR-9-5p, which inhibits the transformation
from mesothelial cells to myofibroblast and reduces fibrogenesis
via targeting TGF-β receptor type II (TGFBR2) and NOX4, was also
upregulated, demonstrating that there may be a self-limiting
homeostatic mechanism (28).
Moreover, TGF-β1 can upregulate the level of Sirtuin 6 (SIRT6)
protein in HFLF. The overexpression of SIRT6 inhibits
TGF-β1-induced myofibroblast differentiation by suppressing
TGF-β1/Smad2 and NF-κB signaling pathways (30). Inhibition of TGF-β1/Smad signal
downregulated the expression of Rock1, RhoC and RhoA, demonstrating
Rho kinase was a key mediator in myofibroblast differentiation
induced by TGF-β1/Smad (31).
It was also reported that TGF-β1 stimulated primary
human bronchial epithelial cells (HBEC) to the status of EMT in
vitro mainly through Smad2/3-dependent mechanism (32). TGF-β1 induces alveolar epithelial
cells (AEC) to EMT in a time- and concentration-dependent manner
through Smad2 activation, and this event induced by TGF-β1 was not
relevant to the ERK1/2 signaling pathway (33). In addition, TGF-β1/Smad2/3
signaling mediated the EMT induced by the high mobility group box 1
(HMGB1) released from injured lung in A549 cells (34). There was a negative feedback
mechanism in the TGF-β1/Smad-involved pulmonary fibrosis. TGF-β1
upregulates the expression of CXCR7, a seven transmembrane G
protein-coupled receptor in endothelial cells, in a
Smad2/3-dependent pattern. Overexpression of CXCR7 impeded
endothelial-to-mesenchymal transition (EndMT) and lung fibrosis
induced by TGF-β1 through inhibition of the Jag1-Notch pathway
(35). TGF-β1 stimulation
significantly upregulated the expression of Resistin-like
molecule-β (RELM-β) through the Smad2/3/4 pathway, which was
reported to enhance TGF-β1-induced cell proliferation and EndMT
(36). Rho kinase signal
transduction activated by TGF-β1 in EMT was a positive regulator of
phosphodiesterase 4 (PDE4), which promoted EMT of AEC (37).
The expression of peroxisome proliferator-activated
receptor γ (PPAPγ), a negative regulator of TGF-β1-induced
fibrosis, is mainly controlled by TGF-β1. Cells lacking Smad3
showed that the down-regulation effect of TGF-β1 on PPARγ was
weakened, suggesting that TGF-β1 regulates the PPARγ in a
Smad3-dependent manner (38).
TGF-β1 exerted a pro-fibrosis effect by regulating the expression
of connective tissue growth factor (CTGF), which was attributed to
activation of the TGF-β1/Smad3 signaling pathway (39). Follistatin-like protein 1 (Fstl1)
was a glycoprotein that plays a crucial role in promoting
fibrogenesis. At the transcriptional and translational level, the
expression of Fstl1 was upregulated by TGF-β1 via the Smad3-c-Jun
signaling pathway in mouse pulmonary fibroblasts, suggesting that
TGF-β1 may contribute to the IPF through a Smad3/c-Jun/Fstl1 axis
(40). Huang et al
(41) reported that TGF-β1/Smad3
signal inhibited the expression of long noncoding RNA fetal-lethal
noncoding developmental regulatory RNA (FENDRR) which can reduce
fibrogenesis and inhibit the process of pulmonary fibrosis. The
TGF-β1/Smad3 signal upregulates the phosphorylation level of ERK5
and further leads to the contraction and migration of collagen gel
induced by TGF-β1 (42). miR-29,
a downstream target gene of TGF-β/Smad, was capable of inhibiting
numerous fibrosis-related genes upregulated by TGF-β1 including
CTGF, Smad3 and TGF-β1 (43).
However, in fibroblasts, the expression of miR-29 was negatively
regulated by TGF-β1/Smad3 signal (43-45). Similarly, Smad7, a negative
regulator of TGF-β1, is suppressed by miR-182-5p which is induced
by TGF-β1, resulting in the development of IPF (46). TGF-β1 activates Semaphorin (SEMA)
7A and its receptors through a Smad3-independent and Smad
2/3-independent mechanism, respectively, promoting pulmonary
fibrosis (47) Activating
transcription factor 4 (ATF4) was a pivotal transcriptional
regulator for the metabolism of amino acid (48). TGF-β1/Smad3 signaling could
increase the expression of the ATF4 through initiating the
mechanistic target of rapamycin complex 1 (mTORC1) and its
downstream translation initiation factor 4E binding protein 1
(4E-BP1), promoting collagen biosynthesis (49). This is one of the key pathways
through which TGF-β1 stimulates collagen synthesis and IPF in HLF
(50) (Fig. 2).
A great number of studies indicated that
phosphatidylinositol-3-kinase (PI3K) was involved in the
pathomechanism of pulmonary fibrosis (51-54). It was also revealed that PI3K may
play an important role in TGF-β1-relevant IPF.
As mentioned previously, CTGF is a functional
intermediate product between TGF-β1 and ECM protein. CTGF derived
from epithelial cells can activate fibroblasts and further
accelerate the fibrosis process in an autocrine manner (55). It was reported that TGF-β1 may
induce the EMT and synthesis of ECM in lung epithelial cells
through the TGF-β1/PI3K/CTGF signaling pathway (56). Treating human lung epithelial
cells with PI3K inhibitor can, not only inhibit the synthesis of
CTGF and type I collagen, but also reverse the EMT and fibrogenesis
stimulated by TGF-β1. TGF-β1 activated PI3K and protein kinase B
(PKB)/AKT via SEMA 7A-dependent mechanisms. SEMA 7A plays a central
role in the PI3K/PKB/AKT pathway, which contributes to
TGF-β1-induced fibrosis and remodeling (47). TGF-β1 activated the
PI3K/Jun-NH2-terminal kinase (JNK)/AKT and AP-1 synergistically to
induce tissue factor (TF) expression in HLF, promoting the process
of IPF (57) (Fig. 3).
Mitogen-activated protein kinase (MAPK), mainly
consisting of three distinctive cascades, the JNK, p38 and ERK
pathways, is a well-known and crucial signaling pathway in multiple
diseases (58-61). In the past decades, the role of
MAPK cascade in the TGF-β1-relevant IPF has been gradually
elucidated.
Coagulation factor XII (FXII) is a serine protease
relevant to fibrinolysis, it was demonstrated that the production
of FXII induced by TGF-β1 in HLF was mediated with JNK/Smad3
signaling pathways (62). With
the stimulation of TGF-β1, the expression of phosphorylated p38,
phosphorylated JNK, and interstitial phenotypic markers including
desmin, vimentin and a-SMA were significantly increased (63). TGF-β1-induced primary lung
fibroblasts immediately release extracellular fibroblast growth
factor-2 (FGF-2), p38 MAPK and JNK phosphorylation. As a result,
lung fibroblasts proliferated in response to TGF-β1 indirectly
(64). TGF-β1 can induce the
phenotype of HLF to myofibroblasts in a dose- and time-dependent
manner. Although the activity and phosphorylation of c-JNK, p38
MAPK, and ERK increased in response to TGF-β1, phenotypic
modulation from HLF to myofibroblast was only regulated by c-JNK,
suggesting that TGF-β1 induced HLF to myofibroblast via a
c-JNK-mediated pathway (65).
TGF-β1 was also reported to contribute to pulmonary fibrosis
through down-regulation of the expression of vascular endothelial
growth factor-D (VEGF-D) in HLF via the JNK signaling pathway,
providing a speculative mechanism in the tissue remodeling of IPF
(66). Notably, this protective
effect of TGF-β1 on fibroblasts was independent on endothelin
(ET)-1, which also endows fibroblast resistance to apoptosis.
TGF-β1 could induce the deposition of extracellular matrix derived
from tracheal basal cells, and the latter promoted EMT via a c-JNK1
involved pathway, which impairs the homeostasis of epithelial cell
and the occurrence of IPF (67).
Notably, TGF-β1/MAPK signal not only contributed to
the phenotypic modulation to myofibroblast, but also showed a
protective effect on myofibroblasts. For example, TGF-β1 attenuates
the apoptosis of fibroblast by inducing the production of a
p38-dependent growth factor, which activates PI3K/AKT successively
(68). It is noteworthy that
activation of p38 MAPK induced by TGFβ1 was able to induce α-SMA
but not collagen I in HLF (24).
Tissue inhibitors of matrix metalloproteinases 3 (TIMP3), an
effective angiogenesis inhibitor blocking the binding of VEGF to
VEGF receptor 2, may be an important mediator of TGF-β1-mediated
IPF (69). As TGF-β1 strongly
upregulates the expression of TIMP3 in HLF, this process is
relevant to p38 but not ERK pathway. The p38-mediated loss of
epithelial complement inhibitory protein (CIP) caused by TGF-β1 led
to the expansion of IPF epithelial damage, which in turn led to
complement activation, further downregulated CIPs and induced the
expression of TGF-β1 in feedback (70).
TGF-β1 regulates the autocrine of basic fibroblast
growth factor (bFGF) in HLF, which activated the expression of ERK
pathway and the induction of activator protein-1 (AP-1),
accelerating pulmonary fibrogenesis (71). It was also reported that TGF-β1
induces GSK-3β inhibition and nuclear β-catenin translocation in
HLF through ERK1/2 activation, which successively led to the
production of γ-SMA and collagen (72). CD44v6 regulates the synthesis of
COL1 and α-SMA in fibroblasts, and it is a potential activation
target of TGF-β1 in lung fibroblasts (73). The induction of CD44v6 by TGF-β1
not only depends on ERK-induced early growth response-1 (EGR1)
signaling, but also requires abundant AP-1 involvement, suggesting
that there is a TGFβ1-ERK-EGR1/AP-1-CD44v6 axis (73). TGF-β1 can induce the expression
of FGF-2 and its release from type II AEC. In addition, the FGF-2
signaling is responsible for the fibroblast proliferation and
fibrotic activation through the ERK pathway (74). TGF-β1 binds non-covalently to the
latency-related peptide (LAP) to form a complex. Consequently, the
interaction of integrin α8β1 and LAPT-TGF-β1 complex induces FAK
and ERK phosphorylation and promotes cell proliferation (75) (Fig. 4).
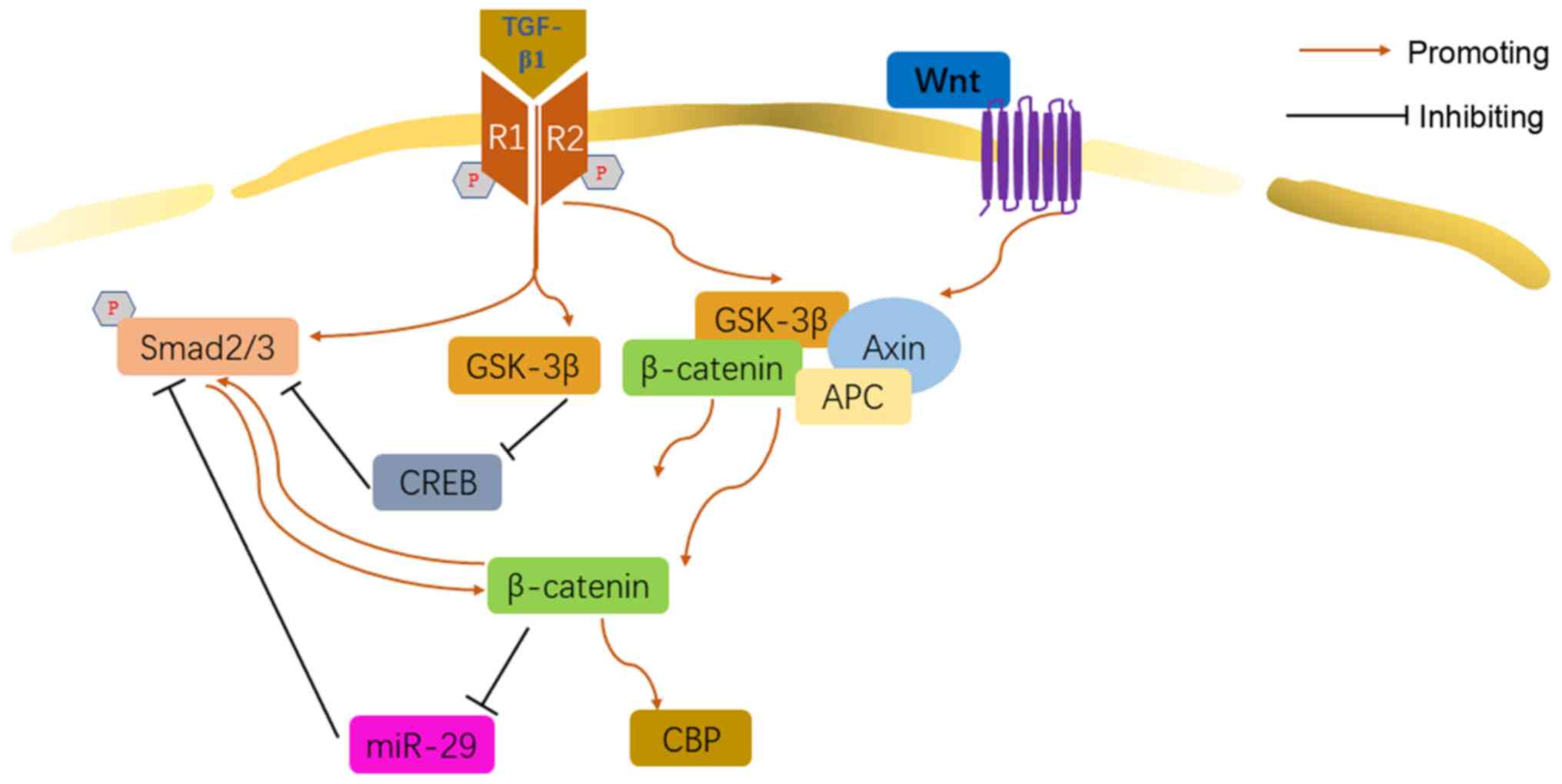
The Wnt/β-catenin pathway is the canonical Wnt
signaling pathway, also known as the 'β-catenin-dependent' Wnt
pathway. Wnt/β-catenin has been proven to play an important role in
body development and growth, tumor, cardiovascular disease,
musculoskeletal diseases, and also respiratory disease (76-78). In normal conditions, the glycogen
synthase kinase-3β (GSK-3β) combines with the β-catenin, axis
inhibition protein (Axin) and adenomatous polyposis coli (APC) to
form a complex. When the Wnt/β-catenin was activated, the complex
degraded, while β-catenin was not degraded and translocated into
the nucleus (77).
Increasing evidence suggested that Wnt/β-catenin was
involved in the TGF-β1-relevant IPF. TGF-β1 initiated the
Wnt/β-catenin cascade via upregulating β-catenin and GSK-3β,
promoting the fibrotic differentiation of lung resident mesenchymal
stem cells (LR-MSCs) (79). In
addition, it was found that, Wnt/β-catenin was required for the
initiation of Smad2/3 induced by TGF-β1, suggesting that there may
be a crosstalk between the two mechanisms in the myofibroblast
differentiation (80). GSK-3
signaling decreases the phosphorylation of cAMP-response element
binding protein (CREB) and attenuates its antagonism function on
TGF-β/Smad signaling, promoting the myofibroblast differentiation
in HLF (81). However, Liu et
al suggested that in the transition of human normal skin
fibroblast to myofibroblast induced by TGF-β1, Wnt/β-catenin played
the role of negative regulator (82). TGF-β1 was capable of inducing the
accumulation of β-catenin in the nuclear, facilitating EMT in a
CREB-binding protein (CBP)-depending pattern in AEC (83). This revealed a potential cascade
of TGF-β1/β-catenin/CBP. miR-29 negatively regulated the
proliferation of IMR-90 cells induced by TGF-β1, but TGF-β1
inhibited the expression of all three members of the miR-29 family
via Wnt3a/β-catenin pathway (84) (Fig. 5).
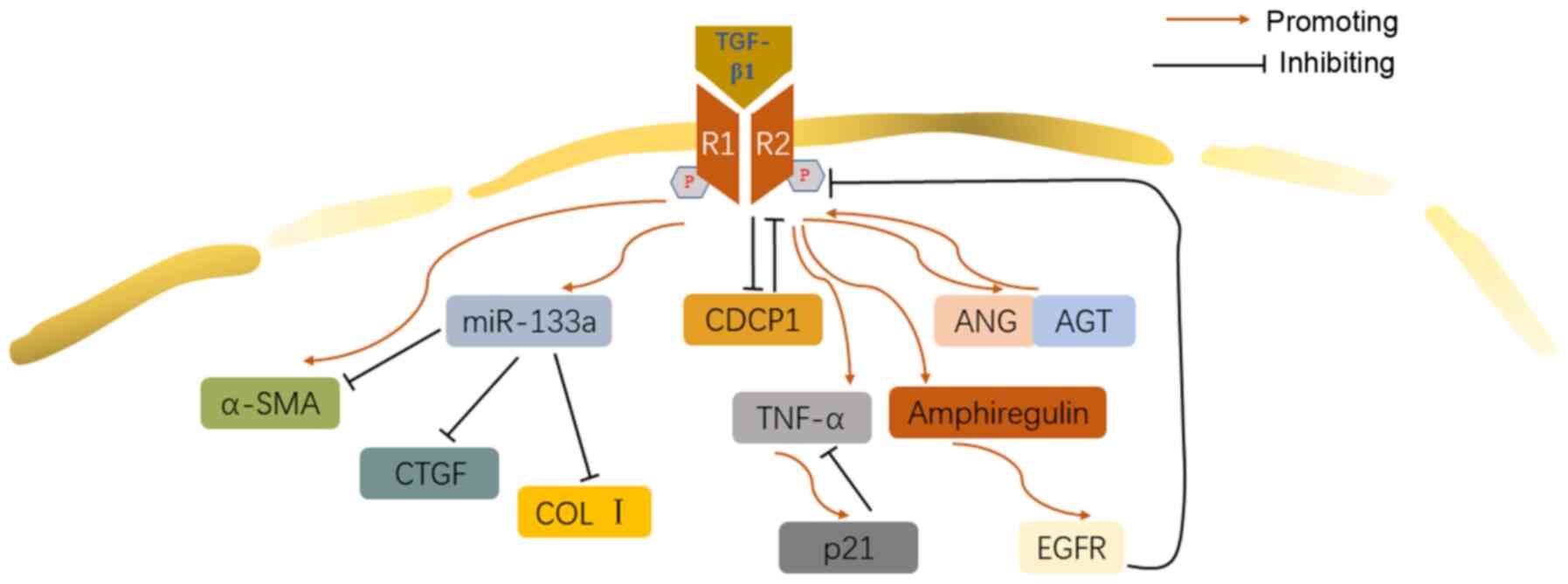
Feedback regulation is a crucial aspect in molecule
cascades. Both positive and negative feedback are revealed in
TGF-β1-involved pathway in IPF.
TGF-β1 strongly downregulated Cub domain-containing
protein 1 (CDCP1), which promoted myofibroblast differentiation
through inhibition of the potential negative feedback effect of
CDCP1 expression on TGF-β1 stimulation (85). Similarly, TGF-β1 activated the
autocrine mechanism of angiotensin (ANG) and angiotensinogen (AGT)
peptide, which upregulated the expression of TGF-β1 to form an
'autocrine loop', promoting the development of IPF (86). miR-133a was reported to attenuate
the differentiation of myofibroblasts by targeting many components
of the TGF-β1 pro-fibrosis pathway, including α-SMA, CTGF and
collagen. There seems to be a negative-feedback loop in the TGF-β1
pro-fibrogenesis pathway, because TGF-β1 upregulates the expression
of miR-133a (87). Additionally,
p21, a key regulator of apoptosis induced by TGF-β1 through tumor
necrosis factor-α (TNF-α) signaling pathway, negatively regulates
TNF-α expression induced by TGF-β1, participating in the fibrosis
and alveolar remodeling induced by TGF-β1 (88). TNF-α could enhance the process of
EMT induced by TGF-β1 in A549 cells through combination with TGF-β1
(89). However, TGF-β1 was also
reported to inhibit the release of TNF-α from mast cells (90). TGF-β1 stimulates the EGFR ligand,
amphiregulin, which regulates the classical and non-classical
TGF-β1 signaling pathway through the activation of EGFR (91) (Fig. 6).
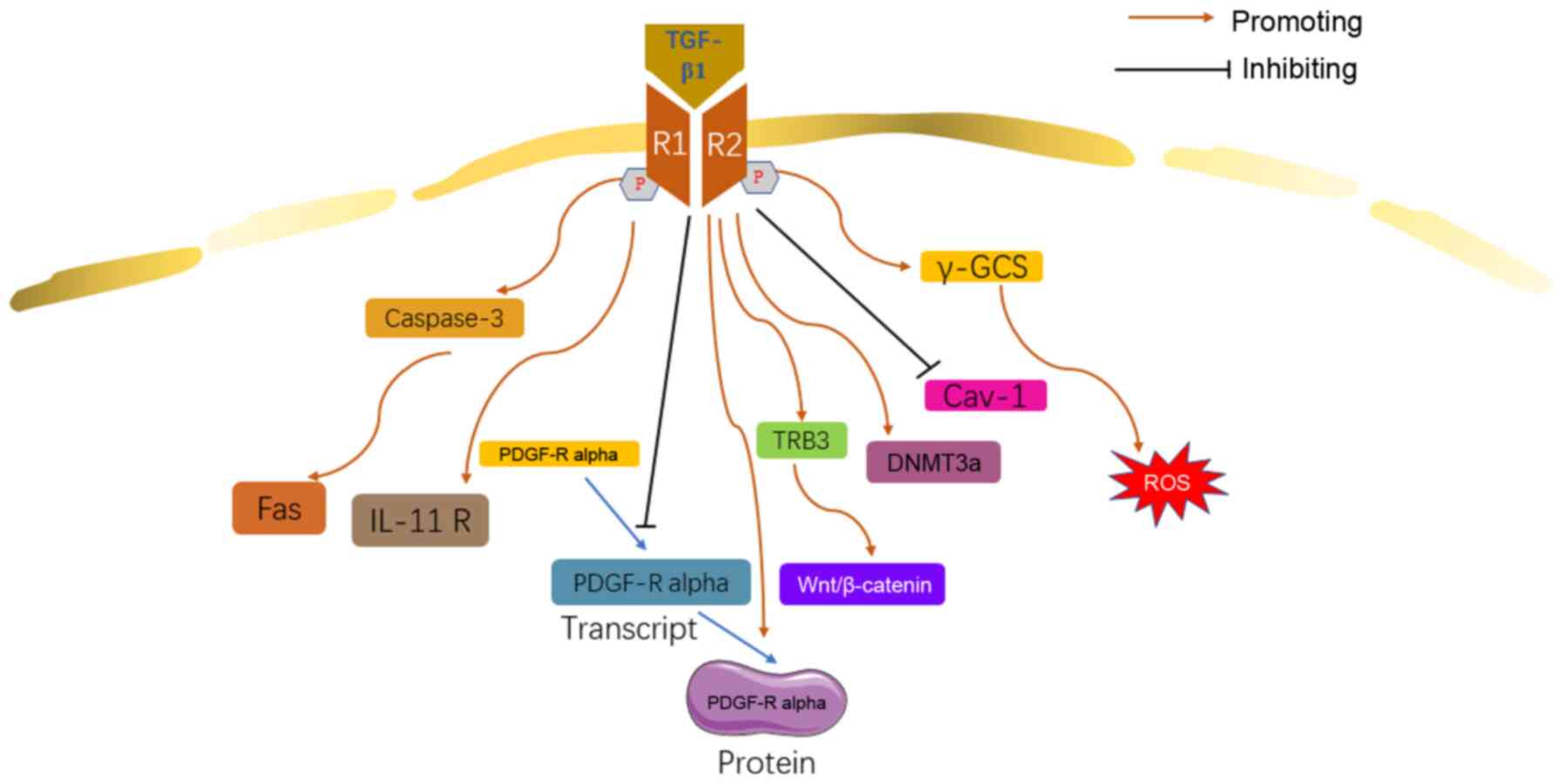
Besides the signaling pathways discussed above,
other molecules cascades were also revealed to be involved in the
TGF-β1 relevant mechanisms of IPF.
The proliferation of fibroblasts is mainly mediated
by platelet-derived growth factor (PDGF) isoforms, whose activity
was potentially regulated by TGF-β1 (92). It was reported that TGF-β1
downregulated the expression of PDGF-α receptor (PDGF-Rα)
transcript. However, TGF-β1 facilitated the transcription of
PDGF-Rα in HLF, suggesting that TGF-β1 may contribute to IPF
through a PDGF-Rα-involved complex network (92). It was reported that the IL-11
secreted by fibroblasts in the lungs of patients with IPF was
significantly upregulated (93),
and results demonstrated that TGF-β1 significantly increases IL-11
receptor expression in mouse fibroblasts (94), suggesting that IL-11 may be an
important mediator of TGF-β1 involved IPF. Fas pathway-mediated
apoptosis of lung epithelial cells is involved in the pathogenesis
of pulmonary fibrosis (95). In
lung tissues of patients with IPF, Fas- and FasL-induced apoptosis
occurs in AEC and infiltrated inflammatory cells. TGF-β1 enhances
the Fas-mediated pulmonary epithelial cell apoptosis through
caspase-3, resulting in lung injury and pulmonary fibrosis
(96). TGF-β1 induces the
expression of exogenous tribbles homolog 3 (TRB3), which stimulates
EMT and promotes the onset of IPF. In addition, TRB3 may
participate in the regulation of EMT in MLE-12 cells induced by
TGF-β1 through the Wnt/β-catenin signaling pathway (97). Insulin-like growth factor-1
(IGF-I) can co-operate with TGF-β1 to enhance the proliferation of
lung fibroblast (98).
Currently, findings have shown that TGF-β1 may
contribute to the development of IPF through epigenetic regulation.
In fibroblasts from patients with IPF, TGF-β1 induces the
upregulation of DNA methyltransferase (DNMT3a) and
tetmethylcytosine dioxygenase 3 (TET3) (99). TGF-β1 inhibits Caveolin (Cav)-1
gene via histone modifications, contributing to fibroblast
proliferation and apoptosis resistance (100).
TGF-β1 may promote IPF by reducing the production of
antioxidant substance and inducing oxidative stress. TGF-β1
disturbs the homeostasis of the messenger RNA (mRNA) of the
γ-glutamylcysteine synthase (γ-GCS) gene and downregulates
the transcription of the gene, inducing the production of ROS in
epithelial cells (101,102). It was also reported that TGF-β1
reduced the production of glutathione by downregulating precursor
amino acid transport and synthesis rate (103). These results are consistent
with previous reports of Guo et al (29) and Hecker et al (27) (Fig. 7).
TGF-β1 activates Smads through the transmembrane
receptor serine/threonine kinase, thereby continuously regulating
the transcription of target genes (110), The TGF-β1/Smad signaling
pathway functions in IPF mainly through the following three
processes: Myofibroblast differentiation, EMT/EndMT and
fibrogenesis (111). TGF-β1
activates PI3K and protein kinase B (PKB)/AKT through a SEMA
7A-dependent mechanism, thereby inducing the formation of EMT and
ECM in lung epithelial cells (47). TGF-β1 mediates the production of
FXII through the JNK/Smad3 signaling pathway (62). It also attenuates the apoptosis
of fibroblasts by inducing the production of p38-dependent growth
factor, which continuously activates PI3K/AKT. At the same time, it
also initiates the Wnt/β-catenin cascade by upregulating β-catenin
and GSK-3β (79). TGF-β1, not
only regulates various mechanism pathways, but also affects IPF by
regulating epigenetics, oxidative stress, and miRNA (112-115). Some research suggested that
Smad3 activation has no effect on collagen I or α-SMA (24). However, Liu et al
suggested that in the transition of human normal skin fibroblast to
myofibroblast induced by TGF-β1, Wnt/β-catenin played a role of
negative regulator, but had different functions in the lung,
thereby promoting the hypothesis that Wnt/β-catenin is
tissue-specific (82).
There are crosstalks and self-regulating loop in
different pathways involved in TGF-β1-induced IPF. The Rho/Rock and
Smad signaling pathways may cross talk in lung fibroblast
differentiation (31). The
Rho/Rock inhibitor downregulated Smad2 expression and the
TGF-β/Smad inhibitor down-regulated RhoA, RhoC and Rock1
expression. There may be a complex network between the Rho/Rock
pathway and Smad signaling in the process of lung fibroblasts to
myofibroblasts induced by TGF-β1. TGF-β1 mainly promotes IPF, but
there are also some self-regulating mechanisms that can induce
miR-133a expression which acts as an antifibrosis regulator of
TGF-β1, which induces IPF (87).
Activation of the MAPK family is mediated by TGF-β1, which affects
Smad signaling. ERK1/2 activation directly phosphorylates and
activates p90RSK, which is a set of serine/threonine kinases that
play a key role in the MAPK signaling pathway (116).
However, some mechanisms and pathways involved in
TGF-β1 have not been clarified; thus, greater efforts to identify
these should be made with regard to TGF-β1. Although some pathways
have been proven, fewer drugs are actually converted into clinical
applications. As for further studies on TGF-β1 in IPF, the focus
should be on the intersection of various pathways, to facilitate
the development of more effective drugs. At the same time, in
addition to study on the various signal pathways involved in
TGF-β1, an in-depth study of its role in epigenetics, and oxidative
stress should also be conducted. After all, the purpose of research
is to serve the clinic and solve the problem of clinical IPF
treatment.
TGF-β1 plays a crucial role in the development of
IPF as it regulates the pathomechanism of IPF through a number of
signaling pathways, including Smad, MAPK, Wnt, and ERK pathways.
The effect of TGF-β1 on IPF is one of stimulation. Nevertheless,
there are some self-limiting mechanisms. Furthermore, some
TGF-β1-relevant mechanisms in IPF remain to be elucidated.
Not applicable.
ZY substantially contributed to the conception and
design of the work and wrote the manuscript. YH revised the
manuscript critically for important intellectual content. Both
authors approved the final version of the manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
Not applicable.
This review was funded by the National Natural Science
Foundation of China (grant no. 81673120).
|
1
|
Martinez FJ, Collard HR, Pardo A, Raghu G,
Richeldi L, Selman M, Swigris JJ, Taniguchi H and Wells AU:
Idiopathic pulmonary fibrosis. Nat Rev Dis Primers. 3:170742017.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
George PM, Spagnolo P, Kreuter M,
Altinisik G, Bonifazi M, Martinez FJ, Molyneaux PL, Renzoni EA,
Richeldi L, Tomassetti S, et al: Progressive fibrosing interstitial
lung disease: Clinical uncertainties, consensus recommendations,
and research priorities. Lancet Respir Med. 8:925–934. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chanda D, Otoupalova E, Smith SR,
Volckaert T, De Langhe SP and Thannickal VJ: Developmental pathways
in the pathogenesis of lung fibrosis. Mol Aspects Med. 65:56–69.
2019. View Article : Google Scholar :
|
|
4
|
Hutchinson J, Fogarty A, Hubbard R and
McKeever T: Global incidence and mortality of idiopathic pulmonary
fibrosis: A systematic review. Eur Respir J. 46:795–806. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nalysnyk L, Cid-Ruzafa J, Rotella P and
Esser D: Incidence and prevalence of idiopathic pulmonary fibrosis:
Review of the literature. Eur Respir Rev. 21:355–361. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Park Y, Ahn C and Kim TH: Occupational and
environmental risk factors of idiopathic pulmonary fibrosis: A
systematic review and meta-analyses. Sci Rep. 11:43182021.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lv M, Liu Y, Ma S and Yu Z: Current
advances in idiopathic pulmonary fibrosis: The pathogenesis,
therapeutic strategies and candidate molecules. Future Med Chem.
11:2595–2620. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hadjicharalambous MR and Lindsay MA:
Idiopathic pulmonary fibrosis: Pathogenesis and the emerging role
of long non-coding RNAs. Int J Mol Sci. 21:5242020. View Article : Google Scholar :
|
|
9
|
Hewlett JC, Kropski JA and Blackwell TS:
Idiopathic pulmonary fibrosis: Epithelial-mesenchymal interactions
and emerging therapeutic targets. Matrix Biol. 71-72:112–127. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hu HH, Chen DQ, Wang YN, Feng YL, Cao G,
Vaziri ND and Zhao YY: New insights into TGF-β/Smad signaling in
tissue fibrosis. Chem Biol Interact. 292:76–83. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Prashanth Goud M, Bale S, Pulivendala G
and Godugu C: Therapeutic effects of Nimbolide, an autophagy
regulator, in ameliorating pulmonary fibrosis through attenuation
of TGF-β1 driven epithelial-to-mesenchymal transition. Int
Immunopharmacol. 75:1057552019. View Article : Google Scholar
|
|
12
|
Feng F, Cheng P, Xu S, Li N, Wang H, Zhang
Y and Wang W: Tanshinone IIA attenuates silica-induced pulmonary
fibrosis via Nrf2-mediated inhibition of EMT and TGF-β1/Smad
signaling. Chem Biol Interact. 319:1090242020. View Article : Google Scholar
|
|
13
|
Moustafa EM, Ibrahim SI and Salem FAF:
Methylsulfonylmethane inhibits lung fibrosis progression,
inflammatory response, and epithelial-mesenchymal transition via
the transforming growth factor-Beta 1/SMAD2/3 pathway in rats
exposed to both γ-radiation and Bisphenol-A. Toxin Rev. 1–10.
2020.
|
|
14
|
He J, Peng H, Wang M, Liu Y, Guo X, Wang
B, Dai L, Cheng X, Meng Z, Yuan L, et al: Isoliquiritigenin
inhibits TGF-β1-induced fibrogenesis through activating autophagy
via PI3K/AKT/mTOR pathway in MRC-5 cells. Acta Biochim Biophys Sin
(Shanghai). 52:810–820. 2020. View Article : Google Scholar
|
|
15
|
Sgalla G, Iovene B, Calvello M, Ori M,
Varone F and Richeldi L: Idiopathic pulmonary fibrosis:
Pathogenesis and management. Respir Res. 19:322018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kim KK, Sheppard D and Chapman HA:
TGF-beta 1 signaling and tissue fibrosis. Cold Spring Harb Perspect
Biol. 10:a0222932018. View Article : Google Scholar
|
|
17
|
Werner F, Jain MK, Feinberg MW, Sibinga
NE, Pellacani A, Wiesel P, Chin MT, Topper JN, Perrella MA and Lee
ME: Transforming growth factor-beta 1 inhibition of macrophage
activation is mediated via Smad3. J Biol Chem. 275:36653–36658.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Flanders KC: Smad3 as a mediator of the
fibrotic response. Int J Exp Pathol. 85:47–64. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zheng R, Xiong Q, Zuo B, Jiang S, Li F,
Lei M, Deng C and Xiong Y: Using RNA interference to identify the
different roles of SMAD2 and SMAD3 in NIH/3T3 fibroblast cells.
Cell Biochem Funct. 26:548–556. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Roberts AB, Piek E, Bottinger EP, Ashcroft
G, Mitchell JB and Flanders KC: Is Smad3 a major player in signal
transduction pathways leading to fibrogenesis? Chest. 120(1 Suppl):
43S–47S. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Evans RA, Tian YC, Steadman R and Phillips
AO: TGF-beta1-mediated fibroblast-myofibroblast terminal
differentiation-the role of Smad proteins. Exp Cell Res.
282:90–100. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gu L, Zhu YJ, Yang X, Guo ZJ, Xu WB and
Tian XL: Effect of TGF-beta/Smad signaling pathway on lung
myofibroblast differentiation. Acta Pharmacol Sin. 28:382–391.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kobayashi T, Liu X, Wen FQ, Kohyama T,
Shen L, Wang XQ, Hashimoto M, Mao L, Togo S, Kawasaki S, et al:
Smad3 mediates TGF-beta1-induced collagen gel contraction by human
lung fibroblasts. Biochem Biophys Res Commun. 339:290–295. 2006.
View Article : Google Scholar
|
|
24
|
Deng X, Jin K, Li Y, Gu W, Liu M and Zhou
L: Platelet-derived growth factor and transforming growth factor β1
Regulate ARDS-associated lung fibrosis through distinct signaling
pathways. Cell Physiol Biochem. 36:937–946. 2015. View Article : Google Scholar
|
|
25
|
Lim MJ, Ahn J, Yi JY, Kim MH, Son AR, Lee
SL, Lim DS, Kim SS, Kang MA, Han Y and Song JY: Induction of
galectin-1 by TGF-β1 accelerates fibrosis through enhancing nuclear
retention of Smad2. Exp Cell Res. 326:125–135. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Huang Y, Xie Y, Abel PW, Wei P, Plowman J,
Toews ML, Strah H, Siddique A, Bailey KL and Tu Y: TGF-β1-induced
miR-424 promotes pulmonary myofibroblast differentiation by
targeting Slit2 protein expression. Biochem Pharmacol.
180:1141722020. View Article : Google Scholar
|
|
27
|
Hecker L, Vittal R, Jones T, Jagirdar R,
Luckhardt TR, Horowitz JC, Pennathur S, Martinez FJ and Thannickal
VJ: NADPH oxidase-4 mediates myofibroblast activation and
fibrogenic responses to lung injury. Nat Med. 15:1077–1081. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fierro-Fernández M, Busnadiego Ó, Sandoval
P, Espinosa-Díez C, Blanco-Ruiz E, Rodríguez M, Pian H, Ramos R,
López-Cabrera M, García-Bermejo ML and Lamas S: miR-9-5p suppresses
pro-fibrogenic transformation of fibroblasts and prevents organ
fibrosis by targeting NOX4 and TGFBR2. EMBO Rep. 16:1358–1377.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Guo W, Saito S, Sanchez CG, Zhuang Y,
Gongora Rosero RE, Shan B, Luo F and Lasky JA: TGF-β1
stimulates HDAC4 nucleus-to-cytoplasm translocation and NADPH
oxidase 4-derived reactive oxygen species in normal human lung
fibroblasts. Am J Physiol Lung Cell Mol Physiol. 312:L936–L944.
2017. View Article : Google Scholar
|
|
30
|
Zhang Q, Tu W, Tian K, Han L, Wang Q, Chen
P and Zhou X: Sirtuin 6 inhibits myofibroblast differentiation via
inactivating transforming growth factor-β1/Smad2 and nuclear
factor-κB signaling pathways in human fetal lung fibroblasts. J
Cell Biochem. 120:93–104. 2019. View Article : Google Scholar
|
|
31
|
Ji H, Tang H, Lin H, Mao J, Gao L, Liu J
and Wu T: Rho/Rock cross-talks with transforming growth
factor-β/Smad pathway participates in lung fibroblast-myofibroblast
differentiation. Biomed Rep. 2:787–792. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Câmara J and Jarai G:
Epithelial-mesenchymal transition in primary human bronchial
epithelial cells is Smad-dependent and enhanced by fibronectin and
TNF-alpha. Fibrogenesis Tissue Repair. 3:22010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kasai H, Allen JT, Mason RM, Kamimura T
and Zhang Z: TGF-beta1 induces human alveolar epithelial to
mesenchymal cell transition (EMT). Respir Res. 6:562005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li LC, Li DL, Xu L, Mo XT, Cui WH, Zhao P,
Zhou WC, Gao J and Li J: High-mobility group box 1 mediates
epithelial-to-mesenchymal transition in pulmonary fibrosis
involving transforming growth factor-β1/Smad2/3 signaling. J
Pharmacol Exp Ther. 354:302–309. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Guan S and Zhou J: CXCR7 attenuates the
TGF-β-induced endothelial-to-mesenchymal transition and pulmonary
fibrosis. Mol Biosyst. 13:2116–2124. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Jiang Y, Zhou X, Hu R and Dai A:
TGF-β1-induced SMAD2/3/4 activation promotes RELM-β transcription
to modulate the endothelium-mesenchymal transition in human
endothelial cells. Int J Biochem Cell Biol. 105:52–60. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kolosionek E, Savai R, Ghofrani HA,
Weissmann N, Guenther A, Grimminger F, Seeger W, Banat GA,
Schermuly RT and Pullamsetti SS: Expression and activity of
phosphodiesterase isoforms during epithelial mesenchymal
transition: The role of phosphodiesterase 4. Mol Biol Cell.
20:4751–4765. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ramirez A, Ballard EN and Roman J: TGFβ1
controls PPARγ expression, transcriptional potential, and activity,
in part, through Smad3 signaling in murine lung fibroblasts. PPAR
Res. 2012:3758762012. View Article : Google Scholar
|
|
39
|
Li HH, Cai Q, Wang YP, Liu HR and Huang M:
The role of transforming growth factor-β1/connective
tissue growth factor signaling pathway in paraquat-induced
pulmonary fibrosis. Zhonghua Lao Dong Wei Sheng Zhi Ye Bing Za Zhi.
34:484–488. 2016.In Chinese. PubMed/NCBI
|
|
40
|
Zheng X, Qi C, Zhang S, Fang Y and Ning W:
TGF-β1 induces Fstl1 via the Smad3-c-Jun pathway in lung
fibroblasts. Am J Physiol Lung Cell Mol Physiol. 313:L240–L251.
2017. View Article : Google Scholar
|
|
41
|
Huang C, Liang Y, Zeng X, Yang X, Xu D,
Gou X, Sathiaseelan R, Senavirathna LK, Wang P and Liu L: Long
noncoding RNA FENDRR exhibits antifibrotic activity in pulmonary
fibrosis. Am J Respir Cell Mol Biol. 62:440–453. 2020. View Article : Google Scholar :
|
|
42
|
Kadoya K, Togo S, Tulafu M, Namba Y, Iwai
M, Watanabe J, Okabe T, Jin J, Kodama Y, Kitamura H, et al:
Specific features of fibrotic lung fibroblasts highly sensitive to
fibrotic processes mediated via TGF-β-ERK5 interaction. Cell
Physiol Biochem. 52:822–837. 2019. View Article : Google Scholar
|
|
43
|
Cushing L, Kuang PP, Qian J, Shao F, Wu J,
Little F, Thannickal VJ, Cardoso WV and Lü J: miR-29 is a major
regulator of genes associated with pulmonary fibrosis. Am J Respir
Cell Mol Biol. 45:287–294. 2011. View Article : Google Scholar :
|
|
44
|
Yang T, Liang Y, Lin Q, Liu J, Luo F, Li
X, Zhou H, Zhuang S and Zhang H: miR-29 mediates TGFβ1-induced
extracellular matrix synthesis through activation of PI3K-AKT
pathway in human lung fibroblasts. J Cell Biochem. 114:1336–1342.
2013. View Article : Google Scholar
|
|
45
|
Xiao J, Meng XM, Huang XR, Chung AC, Feng
YL, Hui DS, Yu CM, Sung JJ and Lan HY: miR-29 inhibits
bleomycin-induced pulmonary fibrosis in mice. Mol Ther.
20:1251–1260. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Chen Y, Zhang Q, Zhou Y, Yang Z and Tan M:
Inhibition of miR-182-5p attenuates pulmonary fibrosis via
TGF-β/Smad pathway. Hum Exp Toxicol. 39:683–695. 2020. View Article : Google Scholar
|
|
47
|
Kang HR, Lee CG, Homer RJ and Elias JA:
Semaphorin 7A plays a critical role in TGF-beta1-induced pulmonary
fibrosis. J Exp Med. 204:1083–1093. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Mukherjee D, Bercz LS, Torok MA and Mace
TA: Regulation of cellular immunity by activating transcription
factor 4. Immunol Lett. 228:24–34. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Selvarajah B, Azuelos I, Platé M,
Guillotin D, Forty EJ, Contento G, Woodcock HV, Redding M, Taylor
A, Brunori G, et al: mTORC1 amplifies the ATF4-dependent de novo
serine-glycine pathway to supply glycine during
TGF-β1-induced collagen biosynthesis. Sci Signal.
12:eaav30482019. View Article : Google Scholar
|
|
50
|
Woodcock HV, Eley JD, Guillotin D, Platé
M, Nanthakumar CB, Martufi M, Peace S, Joberty G, Poeckel D, Good
RB, et al: The mTORC1/4E-BP1 axis represents a critical signaling
node during fibrogenesis. Nat Commun. 10:62019. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Cong LH, Li T, Wang H, Wu YN, Wang SP,
Zhao YY, Zhang GQ and Duan J: IL-17A-producing T cells exacerbate
fine particulate matter-induced lung inflammation and fibrosis by
inhibiting PI3K/Akt/mTOR-mediated autophagy. J Cell Mol Med.
24:8532–8544. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Fang L, Chen H, Kong R and Que J:
Endogenous tryptophan metabolite 5-methoxytryptophan inhibits
pulmonary fibrosis by downregulating the TGF-β/SMAD3 and PI3K/AKT
signaling pathway. Life Sci. 260:1183992020. View Article : Google Scholar
|
|
53
|
Hettiarachchi SU, Li YH, Roy J, Zhang F,
Puchulu-Campanella E, Lindeman SD, Srinivasarao M, Tsoyi K, Liang
X, Ayaub EA, et al: Targeted inhibition of PI3 kinase/mTOR
specifically in fibrotic lung fibroblasts suppresses pulmonary
fibrosis in experimental models. Sci Transl Med. 12:eaay37242020.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Hu X, Xu Q, Wan H, Hu Y, Xing S, Yang H,
Gao Y and He Z: PI3K-Akt-mTOR/PFKFB3 pathway mediated lung
fibroblast aerobic glycolysis and collagen synthesis in
lipopolysaccharide-induced pulmonary fibrosis. Lab Invest.
100:801–811. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Graves DT and Milovanova TN: Mucosal
immunity and the FOXO1 transcription factors. Front Immunol.
10:25302019. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Shi L, Dong N, Fang X and Wang X:
Regulatory mechanisms of TGF-β1-induced fibrogenesis of human
alveolar epithelial cells. J Cell Mol Med. 20:2183–2193. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Wygrecka M, Zakrzewicz D, Taborski B,
Didiasova M, Kwapiszewska G, Preissner KT and Markart P: TGF-β1
induces tissue factor expression in human lung fibroblasts in a
PI3K/JNK/Akt-dependent and AP-1-dependent manner. Am J Respir Cell
Mol Biol. 47:614–627. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Bengal E, Aviram S and Hayek T: p38 MAPK
in glucose metabolism of skeletal muscle: Beneficial or harmful?
Int J Mol Sci. 21:64802020. View Article : Google Scholar :
|
|
59
|
Guo YJ, Pan WW, Liu SB, Shen ZF, Xu Y and
Hu LL: ERK/MAPK signalling pathway and tumorigenesis. Exp Ther Med.
19:1997–2007. 2020.PubMed/NCBI
|
|
60
|
He X and Wang C, Wang H, Li L and Wang C:
The function of MAPK cascades in response to various stresses in
horticultural plants. Front Plant Sci. 11:9522020. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Magnelli L, Schiavone N, Staderini F,
Biagioni A and Papucci L: MAP kinases pathways in gastric cancer.
Int J Mol Sci. 21:28932020. View Article : Google Scholar :
|
|
62
|
Jablonska E, Markart P, Zakrzewicz D,
Preissner KT and Wygrecka M: Transforming growth factor-β1 induces
expression of human coagulation factor XII via Smad3 and JNK
signaling pathways in human lung fibroblasts. J Biol Chem.
285:11638–11651. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Chen HH, Zhou XL, Shi YL and Yang J: Roles
of p38 MAPK and JNK in TGF-β1-induced human alveolar epithelial to
mesenchymal transition. Arch Med Res. 44:93–98. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Khalil N, Xu YD, O'Connor R and Duronio V:
Proliferation of pulmonary interstitial fibroblasts is mediated by
transforming growth factor-beta1-induced release of extracellular
fibroblast growth factor-2 and phosphorylation of p38 MAPK and JNK.
J Biol Chem. 280:43000–43009. 2005. View Article : Google Scholar
|
|
65
|
Hashimoto S, Gon Y, Takeshita I, Matsumoto
K, Maruoka S and Horie T: Transforming growth factor-beta1 induces
phenotypic modulation of human lung fibroblasts to myofibroblast
through a c-Jun-NH2-terminal kinase-dependent pathway. Am J Respir
Crit Care Med. 163:152–157. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Cui Y, Osorio JC, Risquez C, Wang H, Shi
Y, Gochuico BR, Morse D, Rosas IO and El-Chemaly S: Transforming
growth factor-β1 downregulates vascular endothelial growth factor-D
expression in human lung fibroblasts via the Jun NH2-terminal
kinase signaling pathway. Mol Med. 20:120–134. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
van der Velden JL, Wagner DE, Lahue KG,
Abdalla ST, Lam YW, Weiss DJ and Janssen-Heininger YMW:
TGF-β1-induced deposition of provisional extracellular matrix by
tracheal basal cells promotes epithelial-to-mesenchymal transition
in a c-Jun NH2-terminal kinase-1-dependent manner. Am J Physiol
Lung Cell Mol Physiol. 314:L984–L997. 2018. View Article : Google Scholar
|
|
68
|
Kulasekaran P, Scavone CA, Rogers DS,
Arenberg DA, Thannickal VJ and Horowitz JC: Endothelin-1 and
transforming growth factor-beta1 independently induce fibroblast
resistance to apoptosis via AKT activation. Am J Respir Cell Mol
Biol. 41:484–493. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
García-Alvarez J, Ramirez R, Checa M,
Nuttall RK, Sampieri CL, Edwards DR, Selman M and Pardo A: Tissue
inhibitor of metalloproteinase-3 is up-regulated by transforming
growth factor-beta1 in vitro and expressed in fibroblastic foci in
vivo in idiopathic pulmonary fibrosis. Exp Lung Res. 32:201–214.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Gu H, Mickler EA, Cummings OW, Sandusky
GE, Weber DJ, Gracon A, Woodruff T, Wilkes DS and Vittal R:
Crosstalk between TGF-β1 and complement activation augments
epithelial injury in pulmonary fibrosis. FASEB J. 28:4223–4234.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Finlay GA, Thannickal VJ, Fanburg BL and
Paulson KE: Transforming growth factor-beta 1-induced activation of
the ERK pathway/activator protein-1 in human lung fibroblasts
requires the autocrine induction of basic fibroblast growth factor.
J Biol Chem. 275:27650–27656. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Caraci F, Gili E, Calafiore M, Failla M,
La Rosa C, Crimi N, Sortino MA, Nicoletti F, Copani A and Vancheri
C: TGF-beta1 targets the GSK-3beta/beta-catenin pathway via ERK
activation in the transition of human lung fibroblasts into
myofibroblasts. Pharmacol Res. 57:274–282. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Ghatak S, Markwald RR, Hascall VC, Dowling
W, Lottes RG, Baatz JE, Beeson G, Beeson CC, Perrella MA,
Thannickal VJ and Misra S: Transforming growth factor β1 (TGFβ1)
regulates CD44V6 expression and activity through extracellular
signal-regulated kinase (ERK)-induced EGR1 in pulmonary fibrogenic
fibroblasts. J Biol Chem. 292:10465–10489. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Xiao L, Du Y, Shen Y, He Y, Zhao H and Li
Z: TGF-beta 1 induced fibroblast proliferation is mediated by the
FGF-2/ERK pathway. Front Biosci (Landmark Ed). 17:2667–2674. 2012.
View Article : Google Scholar
|
|
75
|
Lu M, Munger JS, Steadele M, Busald C,
Tellier M and Schnapp LM: Integrin alpha8beta1 mediates adhesion to
LAP-TGFbeta1. J Cell Sci. 115:4641–4648. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Bugter JM, Fenderico N and Maurice MM:
Mutations and mechanisms of WNT pathway tumour suppressors in
cancer. Nat Rev Cancer. 21:5–21. 2021. View Article : Google Scholar
|
|
77
|
Rapetti-Mauss R, Berenguier C, Allegrini B
and Soriani O: Interplay between ion channels and the Wnt/β-catenin
signaling pathway in cancers. Front Pharmacol. 11:5250202020.
View Article : Google Scholar
|
|
78
|
Söderholm S and Cantù C: The WNT/β-catenin
dependent transcription: A tissue-specific business. Wiley
Interdiscip Rev Syst Biol Med. Oct 21–2020.Epub ahead of print.
View Article : Google Scholar
|
|
79
|
Lu Y, Zhang T, Shan S, Wang S, Bian W, Ren
T and Yang D: MiR-124 regulates transforming growth factor-β1
induced differentiation of lung resident mesenchymal stem cells to
myofibroblast by repressing Wnt/β-catenin signaling. Dev Biol.
449:115–121. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Xu L, Cui WH, Zhou WC, Li DL, Li LC, Zhao
P, Mo XT, Zhang Z and Gao J: Activation of Wnt/β-catenin signalling
is required for TGF-β/Smad2/3 signalling during myofibroblast
proliferation. J Cell Mol Med. 21:1545–1554. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Baarsma HA, Engelbertink LH, van Hees LJ,
Menzen MH, Meurs H, Timens W, Postma DS, Kerstjens HA and Gosens R:
Glycogen synthase kinase-3 (GSK-3) regulates TGF-β1-induced
differentiation of pulmonary fibroblasts. Br J Pharmacol.
169:590–603. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Liu J, Wang Y, Pan Q, Su Y, Zhang Z, Han
J, Zhu X, Tang C and Hu D: Wnt/β-catenin pathway forms a negative
feedback loop during TGF-β1 induced human normal skin
fibroblast-to-myofibroblast transition. J Dermatol Sci. 65:38–49.
2012. View Article : Google Scholar
|
|
83
|
Zhou B, Liu Y, Kahn M, Ann DK, Han A, Wang
H, Nguyen C, Flodby P, Zhong Q, Krishnaveni MS, et al: Interactions
between β-catenin and transforming growth factor-β signaling
pathways mediate epithelial-mesenchymal transition and are
dependent on the transcriptional co-activator cAMP-response
element-binding protein (CREB)-binding protein (CBP). J Biol Chem.
287:7026–7038. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Wang Y, Liu J, Chen J, Feng T and Guo Q:
MiR-29 mediates TGFβ 1-induced extracellular matrix synthesis
through activation of Wnt/β-catenin pathway in human pulmonary
fibroblasts. Technol Health Care. 23(Suppl 1): S119–S125. 2015.
View Article : Google Scholar
|
|
85
|
Noskovičová N, Heinzelmann K, Burgstaller
G, Behr J and Eickelberg O: Cub domain-containing protein 1
negatively regulates TGF-β signaling and myofibroblast
differentiation. Am J Physiol Lung Cell Mol Physiol. 314:L695–L707.
2018. View Article : Google Scholar
|
|
86
|
Uhal BD, Kim JK, Li X and Molina-Molina M:
Angiotensin-TGF-beta 1 crosstalk in human idiopathic pulmonary
fibrosis: Autocrine mechanisms in myofibroblasts and macrophages.
Curr Pharm Des. 13:1247–1256. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Wei P, Xie Y, Abel PW, Huang Y, Ma Q, Li
L, Hao J, Wolff DW, Wei T and Tu Y: Transforming growth factor
(TGF)-β1-induced miR-133a inhibits myofibroblast differentiation
and pulmonary fibrosis. Cell Death Dis. 10:6702019. View Article : Google Scholar
|
|
88
|
Yamasaki M, Kang HR, Homer RJ, Chapoval
SP, Cho SJ, Lee BJ, Elias JA and Lee CG: P21 regulates
TGF-beta1-induced pulmonary responses via a TNF-alpha-signaling
pathway. Am J Respir Cell Mol Biol. 38:346–353. 2008. View Article : Google Scholar
|
|
89
|
Yamauchi Y, Kohyama T, Takizawa H,
Kamitani S, Desaki M, Takami K, Kawasaki S, Kato J and Nagase T:
Tumor necrosis factor-alpha enhances both epithelial-mesenchymal
transition and cell contraction induced in A549 human alveolar
epithelial cells by transforming growth factor-beta 1. Exp Lung
Res. 36:12–24. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Bissonnette EY, Enciso JA and Befus AD:
TGF-beta1 inhibits the release of histamine and tumor necrosis
factor-alpha from mast cells through an autocrine pathway. Am J
Respir Cell Mol Biol. 16:275–282. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Zhou Y, Lee JY, Lee CM, Cho WK, Kang MJ,
Koff JL, Yoon PO, Chae J, Park HO, Elias JA and Lee CG:
Amphiregulin, an epidermal growth factor receptor ligand, plays an
essential role in the pathogenesis of transforming growth
factor-β-induced pulmonary fibrosis. J Biol Chem. 287:41991–42000.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Bonner JC, Badgett A, Lindroos PM and
Osornio-Vargas AR: Transforming growth factor beta 1 downregulates
the platelet-derived growth factor alpha-receptor subtype on human
lung fibroblasts in vitro. Am J Respir Cell Mol Biol. 13:496–505.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Ng B, Dong J, D'Agostino G, Viswanathan S,
Widjaja AA, Lim WW, Ko NSJ, Tan J, Chothani SP, Huang B, et al:
Interleukin-11 is a therapeutic target in idiopathic pulmonary
fibrosis. Sci Transl Med. 11:eaaw12372019. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Zhang L, Zhang J, Zhang Y and Yi Z:
Expression of interleukin-11 and its receptor in lung of mice with
idiopathic pulmonary fibrosis. Zhong Nan Da Xue Xue Bao Yi Xue Ban.
43:1083–1088. 2018.In Chinese. PubMed/NCBI
|
|
95
|
Otsuki T, Hayashi H, Nishimura Y, Hyodo F,
Maeda M, Kumagai N, Miura Y, Kusaka M and Uragami K: Dysregulation
of autoimmunity caused by silica exposure and alteration of
Fas-mediated apoptosis in T lymphocytes derived from silicosis
patients. Int J Immunopathol Pharmacol. 24(Suppl): 11S–16S.
2011.PubMed/NCBI
|
|
96
|
Hagimoto N, Kuwano K, Inoshima I, Yoshimi
M, Nakamura N, Fujita M, Maeyama T and Hara N: TGF-beta 1 as an
enhancer of Fas-mediated apoptosis of lung epithelial cells. J
Immunol. 168:6470–6478. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Yu W, Mi L and Wang F: Effect of the
alteration of Tribbles homologue 3 expression on
epithelial-mesenchymal transition of transforming growth factor
β1-induced mouse alveolar epithelial cells through the
Wnt/β-catenin signaling pathway. Mol Med Rep. 21:615–622.
2020.PubMed/NCBI
|
|
98
|
Andonegui G, Ni A, Leger C, Kelly MM, Wong
JF, Jalloul A and Winston BW: Sequential expression of IGF-IB
followed by active TGF-β1 induces synergistic pulmonary
fibroproliferation in vivo. Am J Physiol Lung Cell Mol Physiol.
303:L788–L798. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Negreros M, Hagood JS, Espinoza CR,
Balderas-Martinez YI, Selman M and Pardo A: Transforming growth
factor beta 1 induces methylation changes in lung fibroblasts. PLoS
One. 14:e02235122019. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Sanders YY, Liu H, Scruggs AM, Duncan SR,
Huang SK and Thannickal VJ: Epigenetic regulation of caveolin-1
gene expression in lung fibroblasts. Am J Respir Cell Mol Biol.
56:50–61. 2017. View Article : Google Scholar :
|
|
101
|
Arsalane K, Dubois CM, Muanza T, Bégin R,
Boudreau F, Asselin C and Cantin AM: Transforming growth
factor-beta1 is a potent inhibitor of glutathione synthesis in the
lung epithelial cell line A549: Transcriptional effect on the GSH
rate-limiting enzyme gamma-glutamylcysteine synthetase. Am J Respir
Cell Mol Biol. 17:599–607. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Jardine H, MacNee W, Donaldson K and
Rahman I: Molecular mechanism of transforming growth factor
(TGF)-beta1-induced glutathione depletion in alveolar epithelial
cells. Involvement of AP-1/ARE and Fra-1. J Biol Chem.
277:21158–21166. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Boustani MR, Hertig IA, Maloney EK,
Fanburg BL and White AC: Transforming growth factor B1 decreases
uptake of glutathione precursor amino acids in bovine pulmonary
artery endothelial cells. Endothelium. 5:1–10. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Cho SJ and Stout-Delgado HW: Aging and
lung disease. Annu Rev Physiol. 82:433–459. 2020. View Article : Google Scholar
|
|
105
|
Wakwaya Y and Brown KK: Idiopathic
pulmonary fibrosis: Epidemiology, diagnosis and outcomes. Am J Med
Sci. 357:359–369. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Abramson MJ, Murambadoro T, Alif SM, Benke
GP, Dharmage SC, Glaspole I, Hopkins P, Hoy RF, Klebe S, Moodley Y,
et al: Occupational and environmental risk factors for idiopathic
pulmonary fibrosis in Australia: Case-control study. Thorax.
75:864–869. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Somogyi V, Chaudhuri N, Torrisi SE, Kahn
N, Müller V and Kreuter M: The therapy of idiopathic pulmonary
fibrosis: What is next? Eur Respir Rev. 28:1900212019. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Amor MS, Rosengarten D, Shitenberg D,
Pertzov B, Shostak Y and Kramer MR: Lung transplantation in
idiopathic pulmonary fibrosis: Risk factors and outcome. Isr Med
Assoc J. 22:741–746. 2020.
|
|
109
|
Yang S, Liu P, Jiang Y, Wang Z, Dai H and
Wang C: Therapeutic applications of mesenchymal stem cells in
idiopathic pulmonary fibrosis. Front Cell Dev Biol. 9:6396572021.
View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Massagué J, Seoane J and Wotton D: Smad
transcription factors. Genes Dev. 19:2783–2810. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Chang X, Tian M, Zhang Q, Gao J, Li S and
Sun Y: Nano nickel oxide promotes epithelial-mesenchymal transition
through transforming growth factor β1/smads signaling pathway in
A549 cells. Environ Toxicol. 35:1308–1317. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Rosell-García T, Palomo-Álvarez O and
Rodríguez-Pascual F: A hierarchical network of hypoxia-inducible
factor and SMAD proteins governs procollagen lysyl hydroxylase 2
induction by hypoxia and transforming growth factor β1. J Biol
Chem. 294:14308–14318. 2019. View Article : Google Scholar
|
|
113
|
Ko J, Mills T, Huang J, Chen NY, Mertens
TCJ, Collum SD, Lee G, Xiang Y, Han L, Zhou Y, et al: Transforming
growth factor β1 alters the 3′-UTR of mRNA to promote lung
fibrosis. J Biol Chem. 294:15781–15794. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Senavirathna LK, Huang C, Pushparaj S, Xu
D and Liu L: Hypoxia and transforming growth factor β1 regulation
of long non-coding RNA transcriptomes in human pulmonary
fibroblasts. Physiol Rep. 8:e143432020. View Article : Google Scholar
|
|
115
|
Neveu WA, Mills ST, Staitieh BS and
Sueblinvong V: TGF-β1 epigenetically modifies Thy-1 expression in
primary lung fibroblasts. Am J Physiol Cell Physiol. 309:C616–C626.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Kim S, Han JH, Kim S, Lee H, Kim JR, Lim
JH and Woo CH: p90RSK inhibition ameliorates TGF-β1 signaling and
pulmonary fibrosis by inhibiting Smad3 transcriptional activity.
Cell Physiol Biochem. 54:195–210. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Miyake Y, Sasaki S, Yokoyama T, Chida K,
Azuma A, Suda T, Kudoh S, Sakamoto N, Okamoto K, Kobashi G, et al:
Occupational and environmental factors and idiopathic pulmonary
fibrosis in Japan. Ann Occup Hyg. 49:259–265. 2005.PubMed/NCBI
|
|
118
|
Kim SY, Kang DM, Lee HK, Kim KH and Choi
J: Occupational and environmental risk factors for chronic
fibrosing idiopathic interstitial pneumonia in South Korea. J Occup
Environ Med. 59:e221–e226. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Baumgartner KB, Samet JM, Coultas DB,
Stidley CA, Hunt WC, Colby TV and Waldron JA: Occupational and
environmental risk factors for idiopathic pulmonary fibrosis: A
multicenter case-control study. Collaborating centers. Am J
Epidemiol. 152:307–315. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
García-Sancho Figueroa MC, Carrillo G,
Pérez-Padilla R, Fernández-Plata MR, Buendía-Roldán I, Vargas MH
and Selman M: Risk factors for idiopathic pulmonary fibrosis in a
Mexican population. A case-control study. Respir Med. 104:305–309.
2010. View Article : Google Scholar
|
|
121
|
Awadalla NJ, Hegazy A, Elmetwally RA and
Wahby I: Occupational and environmental risk factors for idiopathic
pulmonary fibrosis in Egypt: A multicenter case-control study. Int
J Occup Environ Med. 3:107–116. 2012.PubMed/NCBI
|
|
122
|
Koo JW, Myong JP, Yoon HK, Rhee CK, Kim Y,
Kim JS, Jo BS, Cho Y, Byun J, Choi M, et al: Occupational exposure
and idiopathic pulmonary fibrosis: A multicentre case-control study
in Korea. Int J Tuberc Lung Dis. 21:107–112. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Paolocci G, Folletti I, Torén K, Ekström
M, Dell'Omo M, Muzi G and Murgia N: Occupational risk factors for
idiopathic pulmonary fibrosis in Southern Europe: A case-control
study. BMC Pulm Med. 18:752018. View Article : Google Scholar : PubMed/NCBI
|