Age-related macular degeneration (AMD) is a widely
reported cause of blindness in elderly adults worldwide (1). The progression of AMD is initially
marked by the accumulation of debris during the early stages, while
later stages are characterized by the accumulation of retinal
epithelial dysregulations. AMD is classified into two distinct
subtypes, known as 'non-exudative' and 'exudative' AMD.
The early stages of AMD are characterized by the
presence of drusen in the retina eyeground and the dysregulation of
the retinal pigment epithelium (RPE). Geographical atrophy and
choroidal neovascularization occur during the late atrophic and
exudative phases. Exudative AMD is characterized by choroidal
neovascularization (CNV) invading the subretinal space,
concurrently enhancing the appearance of exudation and
hemorrhaging, and it has been reported to be caused by angiogenesis
(2–4). The exudative AMD subtype
corresponds 10 to 15% of AMD cases. It has been reported as the
major cause of vision loss and blindness (5,6).
In AMD, CNV is responsible for 80% of the cases presenting vision
loss (7). The development of CNV
has been shown to be associated with the involvement of the
vascular endothelial growth factor (VEGF) (8). However, the implication of cellular
signaling in exudative AMD have not yet been fully elucidated.
However, the aging mechanism is considered one of the major
exudative AMD risk markers. This mechanism can dysregulate cellular
signaling, which controls homeostatic processes (9).
The use of curcumin has been revealed to possibly
have major therapeutic benefits for disease treatment in clinical
practice, including cancer and cardiovascular diseases (10–12). However, the number of available
published studies concerning the possible therapeutic effects of
curcumin in ophthalmological disorders, and particularly in
exudative AMD, remains limited. Since combating avoidable visual
impairment and blindness is of utmost importance for public health,
the application of curcumin for the treatment of ophthalmological
disorders (age-related cataracts, glaucoma, AMD, diabetic
retinopathy) may bear promising results (13). The present review article focuses
on the presentation of the possible effects of curcumin on
exudative AMD by targeting oxidative stress, inflammation and
angiogenesis through its mediation of Wingless/Int (Wnt)/β-catenin
signaling.
Exudative AMD has been shown to be associated with
choriocapillaris changes, whereas the RPE monolayer remains intact
(14), ultimately leading to
hypoxia stimulation in the overlying of RPE cells (15). The loss of choriocapillaris may
result in the initiation of CNV. The mechanism of CNV enhances
immature new blood vessels, which may invade Bruch's membrane from
the choriocapillaris to extend in the subretinal or sub-RPE space
(16).
During ocular development, Wnt/β-catenin is mainly
activated. Wnt/β-catenin signaling dysregulation enhances numerous
ocular dysregulations, due to defects in cell fate differentiation
(40). During lens development,
Wnt/β-catenin signaling is activated in the periocular surface
ectoderm and lens epithelium (41,42). For the development of the retinal
epithelium, Wnt/β-catenin signaling is activated in the dorsal
optic vesicle, and is also involved in the stimulation of the RPE
at the optic vesicle stage. At this stage, Wnt/β-catenin signaling
is localized in the peripheral RPE (43). The retinal vascular development
is controlled by the regulation of the Wnt/β-catenin signaling
(40). In the retinal vascular
process, Wnt/β-catenin signaling is modulated by the erythroblast
transformation-specific transcription factor, Erg, which plays a
key role in the angiogenic process (44). Erg regulates the activation of
the Wnt/β-catenin signaling pathway through the concurrent
enhancement of β-catenin and Frizzled 4 (FZD4) transcription
((44). The formation of the
low-density lipoprotein receptor-related protein 5 (LRP5)/LRP6
complex is required for the activation of FZD4/β-catenin signaling
(45). LRP5 has been reported to
play a crucial role through the formation of a complex with LRP6;
however, it has a minimal effect on retinal vascularization
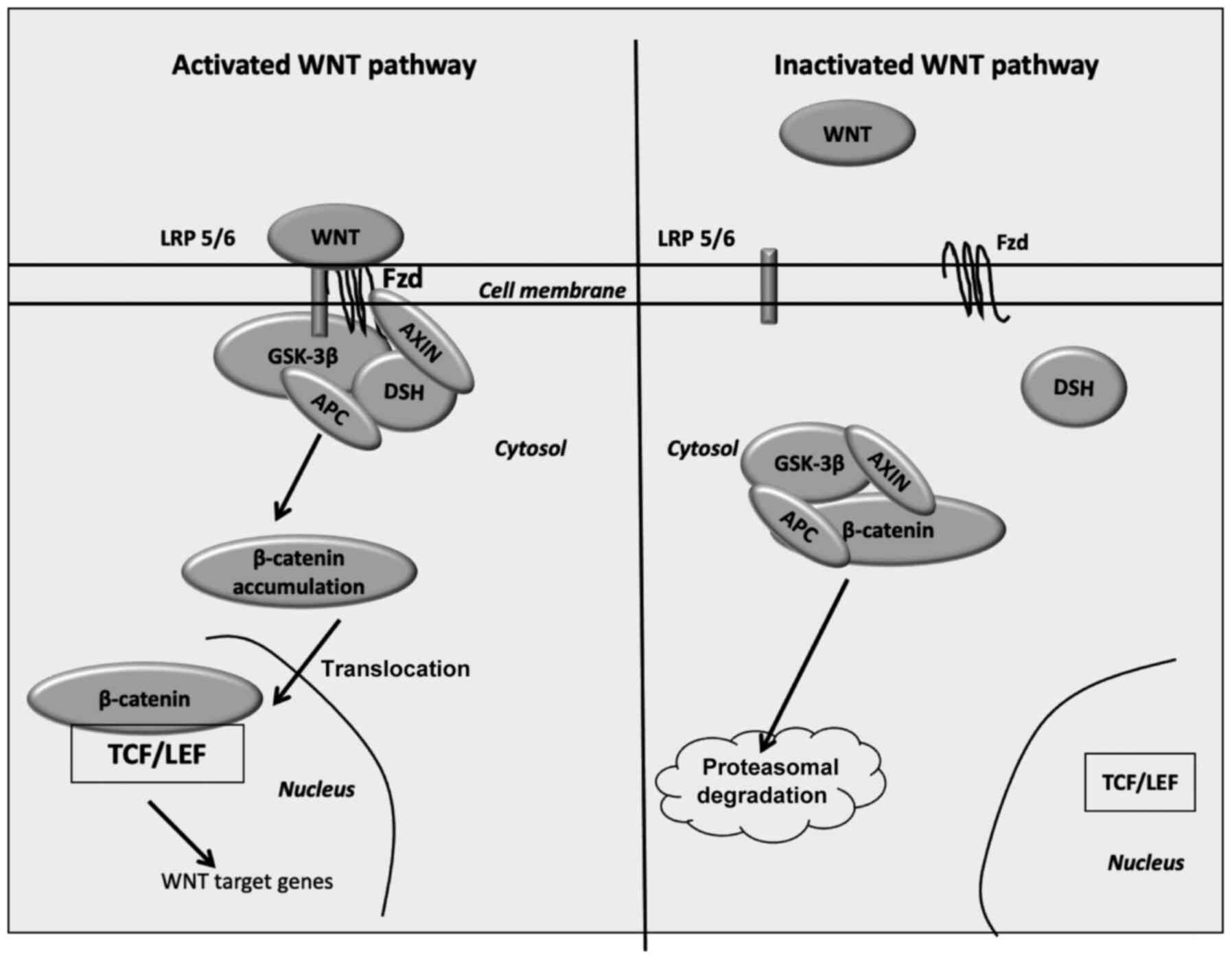
(46,47). Disheveled forms a complex with
AXIN1, in order to prevent β-catenin phosphorylation by glycogen
synthase kinase-3β (GSK-3β). β-catenin accumulates into the
cytoplasm, subsequently translocating to the nucleus and binding to
the T-cell factor/lymphoid enhancer factor (TCF/LEF)
co-transcription factors. The nuclear link enhances the activation
of Wnt-response genes, including cyclin D1, c-Myc, pyruvate
dehydrogenase kinase (PDK)1 and monocarboxylate transporter 1
(MCT-1) (48–52).
Following the inactivation of Wnt ligands, GSK-3β is
activated and then phosphorylates cytoplasmic β-catenin. The
destruction complex is formed by the tumor suppressor adenomatous
polyposis coli (APC), AXIN, GSK-3β and ultimately, β-catenin. The
disintegration of phosphorylated β-catenin is performed in the
proteasome (53). Wnt
inhibitors, including Dickkopf (DKK) family proteins and secreted
Frizzled-related proteins (SFRPs), modulate the Wnt/β-catenin
signaling through the prevention of its ligand-receptor actions
(54) (Fig. 1).
GSK-3β, an intracellular serin-threonine kinase, is
an important regulator of the Wnt/β-catenin signaling pathway
(55), and controls various cell
signaling routes, including cell membrane, neuronal polarity and
inflammatory processes (56–58). GSK-3β concurrently decreases
β-catenin cytoplasmic expression and β-catenin nuclear
translocation (56). β-catenin,
mTOR signaling, hypoxia-inducible factor 1-α (HIF-1α) and VEGF are
downregulated, due to the increased activity of GSK-3β (59).
Various animal models (models of oxygen-induced
retinopathy, streptozotocin rat model, rat model of CNV, rat,
mouse, pig, primate, rabbit) have been utilized for the
investigation of AMD (60), and
previous studies have revealed that aberrantly activated
Wnt/β-catenin signaling may be a pathogenic marker for AMD
(33,61). Stimulated Wnt/β-catenin signaling
has been observed in both human AMD macular tissues (21), and in murine laser-induced CNV
models (20), which are mainly
utilized to investigate the angiogenic form of AMD. The
phosphorylation of LRP6 and the stimulation of β-catenin have been
observed in a laser-induced CNV animal model (20) and in very-low-density lipoprotein
(VLDL) receptor gene knockout (VLDLR−/−) mice with
abnormal intraretinal vessels (62,63). The downregulation of
Wnt/β-catenin signaling with the use of an anti-LRP6 antibody or a
DKK-1 agonist have been reported to impede the formation of
neovascular lesions in murine CNV and VLDLR−/− models
(20). The decrease in Wnt gene
expression in mouse choroidal explants is associated with the
limitation of laser-induced CNV severity (64).
The stimulation of the Wnt/β-catenin signaling has
been shown to be associated with the degeneration of the focal
retina and the subsequent formation of exudative lesions (21). Kallistatin, an endogenous
inhibitor of the Wnt/β-catenin signaling pathway and a member of
the serine proteinase inhibitor (SERPIN) family, has been reported
to be decreased in patients with AMD (21). Kallistatin exerts anti-angiogenic
and anti-inflammatory actions (33,65–69). Kallistatin forms a complex with
LRP6, decreasing Wnt/β-catenin signaling activation (68,69). In murine models with focal
retinal AMD-like lesions, the use of anti-LRP6 antibody has been
found to decrease the Wnt/β-catenin signaling and arrest the
initiation of lesions of the retina (21) (Table I).
Tissue factor (TF), a transmembrane cell-surface
receptor for plasma coagulation factor VII, is one of the main
regulators of the extrinsic coagulation signaling pathway (70). TF exerts angiogenic effects
during the different stages of CNV development (71–73). The stimulation of TF has been
found to be associated with exudative AMD retina (72), with its increase leading to the
development of exudative AMD due to the inflammatory (72,74–76) and angiogenetic processes
(76,77). TF stimulates VEGF activity and
leads to the formation of vascular vessels, through the stimulation
of the Wnt/β-catenin signaling (78). Mab2F1, a monoclonal antibody
specific for LRP6, has been reported to directly inactivate
Wnt/β-catenin signaling, in exudative AMD. In particular, the
inhibition of the Wnt/β-catenin signaling in CNV by Mab2F1 leads to
the reduction of the retinal vascular leakage (20,63). Moreover, the decrease in DKK-1
circulating levels has been shown to be associated with the
initiation of exudative AMD (79).
The downregulation in the levels of DKK-1 has been
found to be associated with the severity of exudative AMD and CNV
development (79). Nevertheless,
the cause for the decrease in the DKK-1 expression level remains
unclear; however, previous research has revealed that circulating
DKK-1 expression is produced from platelets (80). Wnt/β-catenin signaling stimulates
the process of aerobic glycolysis (also known as the Warburg
effect), by the simultaneous stimulation of PI3K/Akt signaling and
HIF-1α, two crucial regulators of the Warburg effect (81–83).
The activation of PI3K/Akt signaling leads to the
stimulation of glucose metabolism and the prevention of reactive
oxygen species (ROS) production through the activation of HIF-1α,
which diverts the glucose from the tricarboxylic acid cycle and the
production of lactate (84).
ROS, a production of normal cell metabolism, can act
either favorably or negatively for cells, depending mainly on the
concentration. The principal source of ROS production is oxidative
mechanisms in the mitochondria and several enzymatic interactions
catalyzed by the oxidoreductase enzymes (85). Decreased concentrations of ROS
interact as cell proliferation enhancers and subsequently
pro-apoptotic enhancers. ROS stimulate a number of transcription
factors, including NF-κB and activator protein 1 (AP-1) (86). ROS have also been reported to
enhance angiogenic process and inflammation (87,88). However, increased ROS
concentrations may be toxic and mutagenic, damaging lipids,
proteins, DNA and ultimately enhancing apoptosis. The endogenous
antioxidant defense mechanism is composed of antioxidant enzymes,
including superoxide dismutase (SOD), catalase, glutathione
peroxidase, heme oxygenase (HO-1) and non-enzymatic antioxidants,
including decreased molecular weight scavengers [glutathione (GSH),
uric acid, lipoic acid, ascorbic acid, tocopherol. Exogenous
antioxidant defense system consists of antioxidants grouped into
natural products and identical to natural ones but synthesized by
the industry, including vitamins (89). The imbalance between ROS
production and antioxidant processes defines oxidative stress (OS).
OS plays a major role in disease initiation, including AMD, as well
as in physiological processes, including aging (90–92).
HIF-1α is transcriptionally involved through
PI3K/Akt signaling by eukaryotic translation initiation factor
4E-binding protein 1 and STAT3 (93–98). c-Myc has been reported to
activate HIF-1α (99). HIF-1α
stimulates the activity of numerous glycolytic enzymes, including
PDK, responsible for the phosphorylation of pyruvate dehydrogenase
(PDH). This results in PDH inactivation and leads to cytoplasmic
pyruvate being converted into lactate by the activation of lactate
dehydrogenase A (LDH-A) (100).
HIF-1α and c-Myc are the main controlling factors of LDH-A
(101–104). This process is characterized by
increased levels of cytoplasmic lactate production (105), and has also been observed in
exudative AMD (59,106). In exudative AMD, the
overstimulation of the Wnt/β-catenin signaling pathway results in
the activation of the Warburg effect and the subsequent enhancement
of photoreceptor protection in retina cells, against OS damage
(107,108).
Previous studies have revealed an association
between Wnt/β-catenin signaling and inflammation, due to their
effects on TNF-α and NF-κB signaling targets (37,109,110). TNF-α has been reported to be
stimulated in AMD (111,112),
while intercellular adhesion molecule (ICAM)-1 is produced in the
RPE and exerts a marked effect on leukocyte adherence (113,114). Inflammation plays a crucial
role in exudative AMD through the activation of the Wnt/β-catenin
signaling pathway, ultimately resulting in the activation of VEGF
(25,33,115,116). Thus, Wnt/β-catenin signaling
activation has been considered to play an integral role in the
development of AMD. Wnt/β-catenin signaling stimulation is
associated with the degeneration of the focal retina and exudative
lesions (21). The stimulation
of Wnt/β-catenin signaling has been shown to be associated with the
initiation of exudative lesions by its associations with
pro-inflammatory markers (33).
The mechanism of inflammation plays a crucial role
in the development of CNV by the stimulation of VEGF (25,33,115,116). The stimulation of NF-κB
signaling, a main inflammatory factor, has been found to be
associated with the activation of Wnt/β-catenin signaling in AMD
(35). The stimulation of
Wnt/β-catenin signaling induces the upregulation of various
factors, including VEGF, TNF-α and ICAM-1 (31,33,117). Subsequently, the stimulation of
VEGF by TNF-α plays a role in CNV (118–121).
The downregulation of Wnt inhibitors, including
DKK-1, has been shown to be associated with exudative lesions and
the severity of CNV (79). In
exudative AMD, VEGF overexpression may be enhanced by the
stimulation of the Wnt/β-catenin signaling (31,33,117) and this occurs by a direct
targeting link (20,122).
The activation of HIF-1α, enhanced by Wnt/β-catenin
signaling, may result in the stimulation of VEGF activity,
ultimately damaging choroid and retinal endothelial cell functions,
subsequently stimulating angiogenesis (123–125).
Curcumin, classified as bis-α, β-unsaturated
β-diketone, is a natural well-known compound. Curcumin is the
active component of Curcuma longa L., which has been
reported to exert a wide range of beneficial effects, including
anticancer properties (132,133). Additionally, curcumin has been
revealed to possess a number of therapeutic properties, including
anti-inflammatory and anti-aging properties (134). In 1815, curcumin was initially
investigated by Vogel and Pelletier from the rhizomes of Curcuma
longa (135). Subsequently,
in 1842, Vogel Jr purified curcumin. In 1910, another study
revealed the chemical structure of curcumin, as diferuloylmethane,
or 1,6-heptadiene-3,5-dione-1,7-bis (4-hydroxy-3-methoxyphenyl)-
(1E,6E) (135). In 1913, for
the first time, a method was developed for curcumin synthesis
(136).
The predicted benefits of curcumin are restricted
due to its reduced oral bioavailability, which can be attributed to
its poor absorption, a high rate of metabolism and a rapid systemic
increase in curcumin levels.
Previous studies have observed that curcumin
pharmacokinetics include a reduced bioavailability (137), and increased pharmacological
and clinical applications (138). However, several potential
processes to overcome this poor bioavailability could be
counteracted through alternative approaches. Various strategies can
improve its bioavailability, including phospholipid complexes,
liposomes and nanoparticles. A number of polymers h utilized to
synthesize nano-formulations for curcumin use to enhance its
biological metabolism (139).
Biocompatible and biodegradable polymers have been utilized in the
administration of therapeutics, due to their low toxicity risk
(140). Previous findings of
liposome formulations have resulted in the improvement of
treatments for drug-resistant cancers and in the reduction of
toxicity (141). Furthermore,
other curcumin delivery processes have been applied, including
nanogels (142), peptide and
protein formulations (143) and
cyclodextrin complexes (144).
As regards AMD, curcumin has been reported to
possibly counteract cell death through the effects on several
cellular signaling pathways (i.e. VEGF, PI3K/Akt, TGF, FGF, COX-2,
I-CAM-1, V-CAM-1) (145). These
processes include the decrease in apoptotic rates of RPE cells and
the diminution of inflammatory mechanisms (146). Curcumin may also reduce free
radical concentrations and oxidative biomarker expression levels,
including superoxide dismutase. Curcumin inhibits apoptosis to
increase the viability of cells (147). It has been previously reported
that, specific microRNAs controlling the antioxidant process, may
be modulated by the administration of curcumin (148). Apart from this, the expression
of HO-1, an enzyme serving cellular defense processes in AMD, is
augmented by the effects of curcumin. Curcumin simultaneously
decreases NF-κB activity and inflammatory gene expression (TNF,
IL-1) (149).
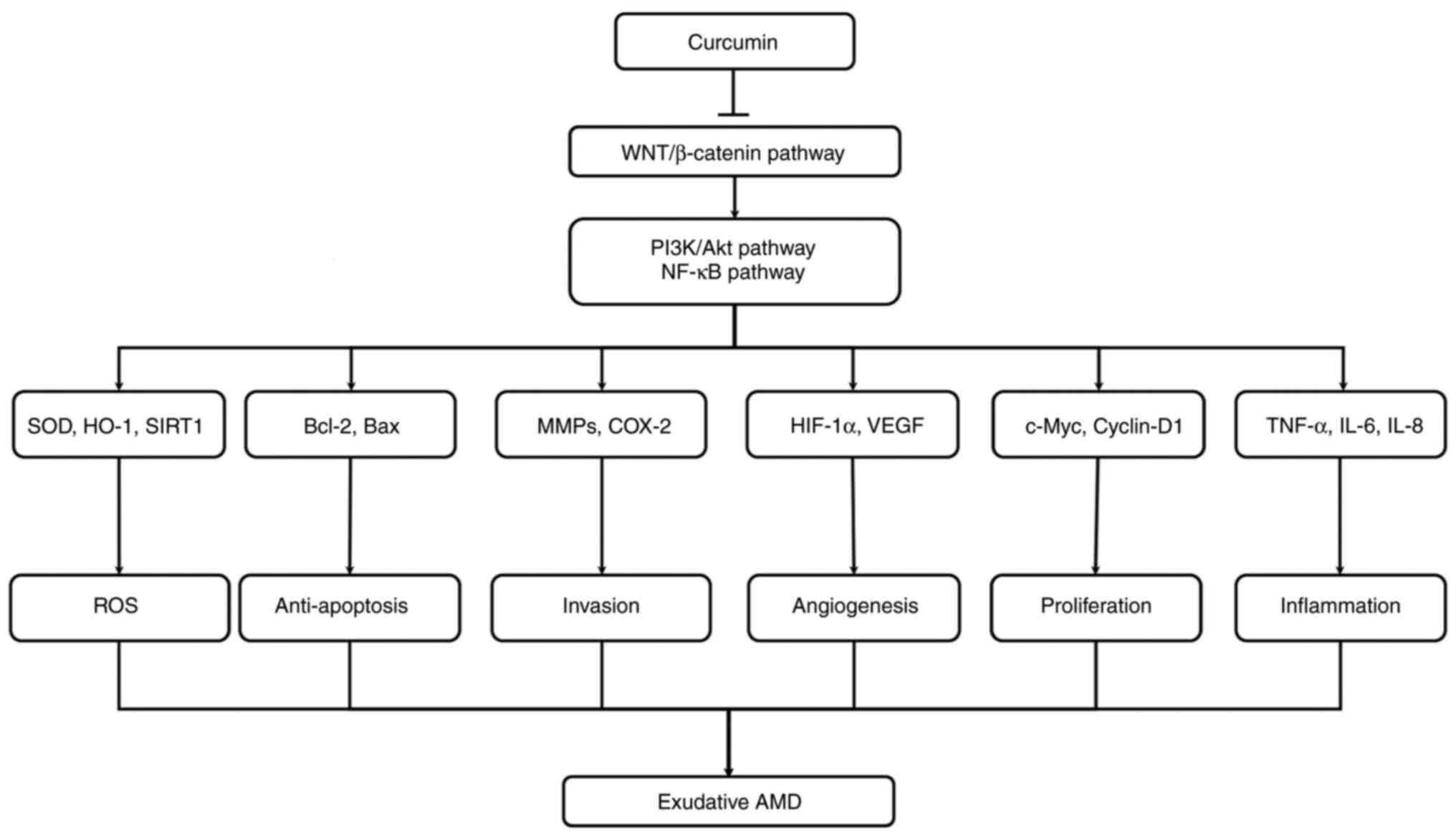
Another protective effect of curcumin has been
observed by counteracting OS induced in ARPE-19 cells (150). In ARPE-19 cells, curcumin can
decrease p44/42 (ERK) apoptotic signaling, with a consecutive
decrease in Bax and Bcl2 levels (Fig. 2). Furthermore, curcumin exerts a
protective effect against OS, which may be a possible therapeutic
approach for AMD (Table I).
The use of curcumin has been reported to lead to
cell cycle arrest in the G2/M stage of tumor cells, due to the
decrease in Wnt/β-catenin signaling (151). Curcumin activates GSK-3β to
decrease nuclear β-catenin translocation and subsequently, to
inhibit the action of cyclin D1. In cancer cells, curcumin analogs
dysregulate the translocation of β-catenin into the nucleus
(152). In xenograft mouse
models, curcumin decreases
12-0-tetradecanoylphorbol-13-acetate-induced Wnt signaling
(153). Additionally, curcumin
and its analog, CHC007, may decrease complex β-catenin/TCF/LEF
levels in various tumor cells (154). Furthermore, curcumin increased
the GSK-3β mRNA level in DAOY medulloblastoma cells to decrease
Wnt/β-catenin signaling (155).
By the decrease in Wnt/β-catenin signaling, curcumin diminishes
cyclin D1 and is responsible for the diminution of brain tumor
growth (155) (Fig. 2).
Curcumin belongs to natural antioxidants. The
effects of curcumin on OS involve numerous processes. Curcumin may
scavenge different forms of OS, including the production of ROS and
reactive nitrogen species (156). It can directly regulate GSH
activity (157). Moreover,
curcumin may decrease ROS-generating enzymes, including
cyclooxygenase 2 (COX-2) (158).
Moreover, curcumin is a chain-breaking antioxidant
and a lipophilic component; this renders it an efficient scavenger
of peroxyl radicals (136,167). Curcumin can enhance the levels
of GSH (168), but can decrease
the activity of nitric oxide synthase in murine macrophages and can
enhance the HO expression in several cell subtypes (169). A decrease in sirtuin-1 (SIRT1)
levels has been shown to be associated with a reduction in SOD
levels. SIRT1 deacetylates SOD (170). SIRT1 is an NAD-dependent enzyme
deacetylating several substrates and regulating metabolism,
including aging. The main role of SIRT1 is the alleviation of
inflammatory process by the decrease in NF-κB signaling and by the
reduction of OS. Previous findings have observed that the
inhibition of SIRT1 is associated AMD (171). SIRT1 has also been reported to
decrease OS by possessing neuroprotective action in mice with optic
nerve crush injury (172).
Moreover, a recent study observed that curcumin activated SIRT1 to
decrease OS (173) (Fig. 2).
Curcumin has been shown to inhibit angiogenesis
through the suppression of VEGF production in U937 and Raji cells
(179). Moreover, COX-2 and
VEGF have been found to be directly suppressed by curcumin in HepG2
hepatoma cells (180).
Curcumin (3,000 mg/kg body weight) administration
has been also associated with a decrease in tumor angiogenesis,
through the inhibition of VEGF and COX-2 expression (180). These effects have been also
reported to be exerted through the liposomal availability of
curcumin and by attenuating the NF-κB signaling pathway (181). Moreover, curcumin may decrease
angiogenesis in basic fibroblast growth factor (bFGF)-induced
corneal neovascularization (182). Furthermore, curcumin may
decrease the activity of FGF-induced neovascularization (183). Previous studies have revealed
the angiogenic synergy between bFGF and VEGF pathway (184–186). bFGF may also increase the
expression of pro-angiogenic factors, including VEGF, to regulate
the angiogenic processes (187,188). As a result, curcumin may
decrease the expression of VEGF through the inhibition of bFGF
expression.
Curcumin may inhibit the activity of the urokinase
plasminogen activator system (uPA; (189). uPA complexes with a specific
receptor (uPAR), through the EGF-like domain in the urokinase
amino-terminal fragment (ATF). This effect has been reported to
result in a decrease in endothelial cell migration and a decrease
in bFGF, TGF, TNF-α, hepatocyte growth factor and VEGF release
(190). Additionally, curcumin
may inhibit MMP-2 expression by interacting via FGF-2 angiogenic
signaling (191) (Fig. 2).
Different properties of curcumin confer
anti-inflammatory and antioxidant activities. Curcumin has been
investigated in congenital and degenerative eye disorders of both
the anterior and posterior segments, and has been previously
utilized as a possible therapeutic (192–194). However, the major issue
concerning the oral use of curcumin remains the reduced curcumin
bioavailability, due to a low gastrointestinal absorption with a
rapid hepatic and intestinal metabolism. Therefore, to counteract
these limitations, numerous methods are investigated, including
curcumin analogues, enhancers and delivery systems. Promising
substances are the pro-drug diphosphorylated curcumin, marked by a
high molecular stability in the aqueous media (195) and the curcumin pro-drug
curcumin diethyl disuccinate (196). Bioavailability enhancers have
been considered, with the use of piperine being highly promising,
having the ability to diminish curcumin hepatic and intestinal
glucuronidation (197), leading
to increase curcumin bioavailability (198). Nanoparticles and liposomes
present high interest to also enhance curcumin bioavailability
(199). Nevertheless, to the
best of our knowledge, the aforementioned strategies have not yet
been investigated for ocular disorder treatment, with the sole
exception of the use of a biodegradable curcumin-loaded scleral
plug for therapy of posterior ocular diseases in rabbit ocular
model (200). Furthermore, a
curcumin-phospholipid lecithin formulation, known as
Meriva®, has been reported to enhance visual acuity and
can diminish macular edema among diabetic retinopathy patients
(201). Nevertheless, in the
therapy of chronic anterior uveitis with complications, curcumin
has demonstrated promising results (202).
Curcumin presents a wide range of pharmacological
actions, including antioxidant, anti-inflammatory and
anti-angiogenic activities in exudative AMD. The role of curcumin
in OS, angiogenesis and inflammatory mechanisms, through its action
of de-activating the Wnt/β-catenin signaling pathway, may indicate
that it can decrease these pathological conditions and may prove to
be an interesting pharmacological agent in exudative AMD. However,
future clinical and pre-clinical studies are warranted to
investigate the role of curcumin as a therapeutic agent in AMD.
Not applicable.
Conceptualization, writing and the preparation of
the original draft were performed by AV. The author has read and
approved the final manuscript. Data authentication is not
applicable.
Not applicable.
Not applicable.
The author declares that he has no competing
interests.
The author would like to thank Polly Gobin (DRCI),
Foch Hospital for her help in reviewing and English editing.
No funding was received.
|
1
|
Bird AC: Therapeutic targets in
age-related macular disease. J Clin Invest. 120:3033–3041. 2010.
View Article : Google Scholar :
|
|
2
|
Radomska-Leśniewska DM, Skopiński P, Bałan
BJ, Białoszewska A, Jóźwiak J, Rokicki D, Skopińska-Różewska E,
Borecka A and Hevelke A: Angiomodulatory properties of Rhodiola
spp. and other natural antioxidants. Cent Eur J Immunol.
40:249–262. 2015. View Article : Google Scholar
|
|
3
|
Radomska-Leśniewska DM, Bałan BJ and
Skopiński P: Angiogenesis modulation by exogenous antioxidants.
Cent Eur J Immunol. 42:370–376. 2017. View Article : Google Scholar
|
|
4
|
Vallée A, Lecarpentier Y, Guillevin R and
Vallée JN: PPARγ agonists: Potential treatments for exudative
age-related macular degeneration. Life Sci. 188:123–130. 2017.
View Article : Google Scholar
|
|
5
|
Coffe V, Carbajal RC and Salceda R:
Glucose metabolism in rat retinal pigment epithelium. Neurochem
Res. 31:103–108. 2006. View Article : Google Scholar
|
|
6
|
Kaur C, Foulds WS and Ling EA:
Hypoxia-ischemia and retinal ganglion cell damage. Clin Ophthalmol.
2:879–889. 2008. View Article : Google Scholar
|
|
7
|
Ferris FL, Fine SL and Hyman L:
Age-related macular degeneration and blindness due to neovascular
maculopathy. Arch Ophthalmol. 102:164016421984. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Barchitta M and Maugeri A: Association
between vascular endothelial growth factor polymorphisms and
age-related macular degeneration: An updated meta-analysis. Dis
Markers. 2016:84864062016. View Article : Google Scholar
|
|
9
|
Yin F, Boveris A and Cadenas E:
Mitochondrial energy metabolism and redox signaling in brain aging
and neurodegeneration. Antioxid Redox Signal. 20:353–371. 2014.
View Article : Google Scholar
|
|
10
|
Vallée A, Lecarpentier Y, Guillevin R and
Vallée JN: opposite interplay between the canonical WNT/β-catenin
pathway and PPAR Gamma: A potential therapeutic target in gliomas.
Neurosci Bull. 34:573–588. 2018. View Article : Google Scholar
|
|
11
|
Vallée A, Lecarpentier Y and Vallée JN:
Curcumin: A therapeutic strategy in cancers by inhibiting the
canonical WNT/β-catenin pathway. J Exp Clin Cancer Res. 38:3232019.
View Article : Google Scholar
|
|
12
|
Yeung AWK, Horbańczuk M, Tzvetkov NT,
Mocan A, Carradori S, Maggi F, Marchewka J, Sut S, Dall'Acqua S,
Gan RY, et al: Curcumin: Total-scale analysis of the scientific
literature. Molecules. 24:13932019. View Article : Google Scholar
|
|
13
|
Kao YW, Hsu SK, Chen JY, Lin IL, Chen KJ,
Lee PY, Ng HS, Chiu CC and Cheng KC: Curcumin metabolite
tetrahydrocurcumin in the treatment of eye diseases. Int J Mol Sci.
22:2122020. View Article : Google Scholar
|
|
14
|
Bhutto I and Lutty G: Understanding
age-related macular degeneration (AMD): Relationships between the
photoreceptor/retinal pigment epithelium/Bruch's
membrane/choriocapillaris complex. Mol Aspects Med. 33:295–317.
2012. View Article : Google Scholar
|
|
15
|
McLeod DS, Grebe R, Bhutto I, Merges C,
Baba T and Lutty GA: Relationship between RPE and choriocapillaris
in age-related macular degeneration. Invest Ophthalmol Vis Sci.
50:4982–4991. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Takata S, Masuda T, Nakamura S, Kuchimaru
T, Tsuruma K, Shimazawa M, Nagasawa H, Kizaka-Kondoh S and Hara H:
The effect of triamcinolone acetonide on laser-induced choroidal
neovascularization in mice using a hypoxia visualization
bio-imaging probe. Sci Rep. 5:98982015. View Article : Google Scholar
|
|
17
|
Sakurai E, Anand A, Ambati BK, van Rooijen
N and Ambati J: Macrophage depletion inhibits experimental
choroidal neovascularization. Invest Ophthalmol Vis Sci.
44:3578–3585. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Indaram M, Ma W, Zhao L, Fariss RN,
Rodriguez IR and Wong WT: 7-Ketocholesterol increases retinal
microglial migration, activation, and angiogenicity: A potential
pathogenic mechanism underlying age-related macular degeneration.
Sci Rep. 5:91442015. View Article : Google Scholar
|
|
19
|
Terasaki H, Kase S, Shirasawa M, Otsuka H,
Hisatomi T, Sonoda S, Ishida S, Ishibashi T and Sakamoto T: TNF-α
decreases VEGF secretion in highly polarized RPE cells but
increases it in non-polarized RPE cells related to crosstalk
between JNK and NF-κB pathways. PLoS One. 8:e699942013. View Article : Google Scholar
|
|
20
|
Hu Y, Chen Y, Lin M, Lee K, Mott RA and Ma
J: Pathogenic role of the Wnt signaling pathway activation in
laser-induced choroidal neovascularization. Invest Ophthalmol Vis
Sci. 54:141–154. 2013. View Article : Google Scholar
|
|
21
|
Tuo J, Wang Y, Cheng R, Li Y, Chen M, Qiu
F, Qian H, Shen D, Penalva R, Xu H, et al: Wnt signaling in
age-related macular degeneration: Human macular tissue and mouse
model. J Transl Med. 13:3302015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nussenblatt RB and Ferris F: Age-related
macular degeneration and the immune response: Implications for
therapy. Am J Ophthalmol. 144:618–626. 2007. View Article : Google Scholar
|
|
23
|
Radeke MJ, Radeke CM, Shih YH, Hu J, Bok
D, Johnson LV and Coffey PJ: Restoration of mesenchymal retinal
pigmented epithelial cells by TGFβ pathway inhibitors: Implications
for age-related macular degeneration. Genome Med. 7:582015.
View Article : Google Scholar
|
|
24
|
Lin CH, Li CH, Liao PL, Tse LS, Huang WK,
Cheng HW and Cheng YW: Silibinin inhibits VEGF secretion and
age-related macular degeneration in a hypoxia-dependent manner
through the PI-3 kinase/Akt/mTOR pathway. Br J Pharmacol.
168:920–931. 2013. View Article : Google Scholar
|
|
25
|
Ambati J: Age-related macular degeneration
and the other double helix. The Cogan lecture. Invest Ophthalmol
Vis Sci. 52:2165–2169. 2011. View Article : Google Scholar
|
|
26
|
Blasiak J, Petrovski G, Veréb Z, Facskó A
and Kaarniranta K: Oxidative stress, hypoxia, and autophagy in the
neovascular processes of age-related macular degeneration. Biomed
Res Int. 2014:7680262014. View Article : Google Scholar
|
|
27
|
Jaffe GJ, Eliott D, Wells JA, Prenner JL,
Papp A and Patel S: A Phase 1 study of Intravitreous E10030 in
combination with ranibizumab in neovascular age-related macular
degeneration. Ophthalmology. 123:78–85. 2016. View Article : Google Scholar
|
|
28
|
Kwak N, Okamoto N, Wood JM and Campochiaro
PA: VEGF is major stimulator in model of choroidal
neovascularization. Invest Ophthalmol Vis Sci. 41:3158–3164.
2000.
|
|
29
|
Brown DM, Kaiser PK, Michels M, Soubrane
G, Heier JS, Kim RY, Sy JP and Schneider S; ANCHOR Study Group:
Ranibizumab versus verteporfin for neovascular age-related macular
degeneration. N Engl J Med. 355:1432–1444. 2006. View Article : Google Scholar
|
|
30
|
Rosenfeld PJ, Brown DM, Heier JS, Boyer
DS, Kaiser PK, Chung CY and Kim RY; MARINA Study Group: Ranibizumab
for neovascular age-related macular degeneration. N Engl J Med.
355:1419–1431. 2006. View Article : Google Scholar
|
|
31
|
Zhang X, Gaspard JP and Chung DC:
Regulation of vascular endothelial growth factor by the Wnt and
K-ras pathways in colonic neoplasia. Cancer Res. 61:6050–6054.
2001.PubMed/NCBI
|
|
32
|
Katoh Y and Katoh M: Comparative
integromics on VEGF family members. Int J Oncol. 28:1585–1589.
2006.
|
|
33
|
Zhou T, Hu Y, Chen Y, Zhou KK, Zhang B,
Gao G and Ma J: The pathogenic role of the canonical Wnt pathway in
age-related macular degeneration. Invest Ophthalmol Vis Sci.
51:4371–4379. 2010. View Article : Google Scholar :
|
|
34
|
Ma B and Hottiger MO: Crosstalk between
Wnt/β-catenin and NF-κB signaling pathway during inflammation.
Front Immunol. 7:3782016. View Article : Google Scholar
|
|
35
|
Wang H and Hartnett ME: Regulation of
signaling events involved in the pathophysiology of neovascular
AMD. Mol Vis. 22:189–202. 2016.
|
|
36
|
Al-Harthi L: Wnt/β-catenin and its diverse
physiological cell signaling pathways in neurodegenerative and
neuropsychiatric disorders. J Neuroimmune Pharmacol. 7:725–730.
2012. View Article : Google Scholar
|
|
37
|
Logan CY and Nusse R: The Wnt signaling
pathway in development and disease. Annu Rev Cell Dev Biol.
20:781–810. 2004. View Article : Google Scholar
|
|
38
|
Klaus A and Birchmeier W: Wnt signalling
and its impact on development and cancer. Nat Rev Cancer.
8:387–398. 2008. View Article : Google Scholar
|
|
39
|
Fuhrmann S: Wnt signaling in eye
organogenesis. Organogenesis. 4:60–67. 2008. View Article : Google Scholar
|
|
40
|
Fujimura N: WNT/β-catenin signaling in
vertebrate eye development. Front Cell Dev Biol. 4:1382016.
View Article : Google Scholar
|
|
41
|
Machon O, Kreslova J, Ruzickova J, Vacik
T, Klimova L, Fujimura N, Lachova J and Kozmik Z: Lens
morphogenesis is dependent on Pax6-mediated inhibition of the
canonical Wnt/beta-catenin signaling in the lens surface ectoderm.
Genesis. 48:86–95. 2010.
|
|
42
|
Carpenter AC, Smith AN, Wagner H,
Cohen-Tayar Y, Rao S, Wallace V, Ashery-Padan R and Lang RA: Wnt
ligands from the embryonic surface ectoderm regulate 'bimetallic
strip' optic cup morphogenesis in mouse. Development. 142:972–982.
2015. View Article : Google Scholar
|
|
43
|
Hägglund AC, Berghard A and Carlsson L:
Canonical Wnt/β-catenin signalling is essential for optic cup
formation. PLoS One. 8:e811582013. View Article : Google Scholar
|
|
44
|
Birdsey GM, Shah AV, Dufton N, Reynolds
LE, Osuna Almagro L, Yang Y, Aspalter IM, Khan ST, Mason JC, Dejana
E, et al: The endothelial transcription factor ERG promotes
vascular stability and growth through Wnt/β-catenin signaling. Dev
Cell. 32:82–96. 2015. View Article : Google Scholar
|
|
45
|
Ye X, Wang Y, Cahill H, Yu M, Badea TC,
Smallwood PM, Peachey NS and Nathans J: Norrin, frizzled-4, and
Lrp5 signaling in endothelial cells controls a genetic program for
retinal vascularization. Cell. 139:285–298. 2009. View Article : Google Scholar
|
|
46
|
Zhou Y, Wang Y, Tischfield M, Williams J,
Smallwood PM, Rattner A, Taketo MM and Nathans J: Canonical WNT
signaling components in vascular development and barrier formation.
J Clin Invest. 124:3825–3846. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Huang W, Li Q, Amiry-Moghaddam M, Hokama
M, Sardi SH, Nagao M, Warman ML and Olsen BR: Critical endothelial
regulation by LRP5 during retinal vascular development. PLoS One.
11:e01528332016. View Article : Google Scholar
|
|
48
|
Shtutman M, Zhurinsky J, Simcha I,
Albanese C, D'Amico M, Pestell R and Ben-Ze'ev A: The cyclin D1
gene is a target of the beta-catenin/LEF-1 pathway. Proc Natl Acad
Sci USA. 96:5522–5527. 1999. View Article : Google Scholar
|
|
49
|
Nusse R: Wnt signaling. Cold Spring Harb
Perspect Biol. 4:a0111632012. View Article : Google Scholar
|
|
50
|
Clevers H and Nusse R: Wnt/β-catenin
signaling and disease. Cell. 149:1192–1205. 2012. View Article : Google Scholar
|
|
51
|
Sprowl-Tanio S, Habowski AN, Pate KT,
McQuade MM, Wang K, Edwards RA, Grun F, Lyou Y and Waterman ML:
Lactate/pyruvate transporter MCT-1 is a direct Wnt target that
confers sensitivity to 3-bromopyruvate in colon cancer. Cancer
Metab. 4:202016. View Article : Google Scholar :
|
|
52
|
Pate KT, Stringari C, Sprowl-Tanio S, Wang
K, TeSlaa T, Hoverter NP, McQuade MM, Garner C, Digman MA, Teitell
MA, et al: Wnt signaling directs a metabolic program of glycolysis
and angiogenesis in colon cancer. EMBO J. 33:1454–1473. 2014.
View Article : Google Scholar :
|
|
53
|
Gao C, Xiao G and Hu J: Regulation of
Wnt/β-catenin signaling by posttranslational modifications. Cell
Biosci. 4:132014. View Article : Google Scholar
|
|
54
|
Cruciat CM and Niehrs C: Secreted and
transmembrane Wnt inhibitors and activators. Cold Spring Harb
Perspect Biol. 5:a0150812013. View Article : Google Scholar
|
|
55
|
Aberle H, Bauer A, Stappert J, Kispert A
and Kemler R: Beta-catenin is a target for the ubiquitin-proteasome
pathway. EMBO J. 16:3797–3804. 1997. View Article : Google Scholar
|
|
56
|
Wu D and Pan W: GSK3: A multifaceted
kinase in Wnt signaling. Trends Biochem Sci. 35:161–168. 2010.
View Article : Google Scholar :
|
|
57
|
Hur EM and Zhou FQ: GSK3 signalling in
neural development. Nat Rev Neurosci. 11:539–551. 2010. View Article : Google Scholar
|
|
58
|
Ambacher KK, Pitzul KB, Karajgikar M,
Hamilton A, Ferguson SS and Cregan SP: The JNK- and
AKT/GSK3β-signaling pathways converge to regulate puma induction
and neuronal apoptosis induced by trophic factor deprivation. PLoS
One. 7:e468852012. View Article : Google Scholar
|
|
59
|
Yokosako K, Mimura T, Funatsu H, Noma H,
Goto M, Kamei Y, Kondo A and Matsubara M: Glycolysis in patients
with age-related macular degeneration. Open Ophthalmol J. 8:39–47.
2014. View Article : Google Scholar
|
|
60
|
Grossniklaus HE, Kang SJ and Berglin L:
Animal models of choroidal and retinal neovascularization. Prog
Retin Eye Res. 29:500–519. 2010. View Article : Google Scholar
|
|
61
|
Wang Z, Liu CH, Huang S and Chen J: Wnt
Signaling in vascular eye diseases. Prog Retin Eye Res. 70:110–133.
2019. View Article : Google Scholar
|
|
62
|
Wang Z, Cheng R, Lee K, Tyagi P, Ding L,
Kompella UB, Chen J, Xu X and Ma JX: Nanoparticle-mediated
expression of a wnt pathway inhibitor ameliorates ocular
neovascularization. Arterioscler Thromb Vasc Biol. 35:855–864.
2015. View Article : Google Scholar
|
|
63
|
Chen Y, Hu Y, Lu K, Flannery JG and Ma JX:
Very low density lipoprotein receptor, a negative regulator of the
wnt signaling pathway and choroidal neovascularization. J Biol
Chem. 282:34420–34428. 2007. View Article : Google Scholar
|
|
64
|
Lin JB, Sene A, Wiley LA, Santeford A,
Nudleman E, Nakamura R, Lin JB, Moolani HV and Apte RS: WNT7A/B
promote choroidal neovascularization. Exp Eye Res. 174:107–112.
2018. View Article : Google Scholar
|
|
65
|
Park K, Lee K, Zhang B, Zhou T, He X, Gao
G, Murray AR and Ma JX: Identification of a novel inhibitor of the
canonical Wnt pathway. Mol Cell Biol. 31:3038–3051. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Dai Z, Lu L, Yang Z, Mao Y, Lu J, Li C, Qi
W, Chen Y, Yao Y, Li L, et al: Kallikrein-binding protein inhibits
LPS-induced TNF-α by upregulating SOCS3 expression. J Cell Biochem.
114:1020–1028. 2013. View Article : Google Scholar
|
|
67
|
Zhang J, Yang Z, Li P, Bledsoe G, Chao L
and Chao J: Kallistatin antagonizes Wnt/β-catenin signaling and
cancer cell motility via binding to low-density lipoprotein
receptor-related protein 6. Mol Cell Biochem. 379:295–301. 2013.
View Article : Google Scholar
|
|
68
|
Lu SL, Tsai C Y, Luo YH, Kuo C F, Lin WC,
Chang YT, Wu JJ, Chuang WJ, Liu CC, Chao L, et al: Kallistatin
modulates immune cells and confers anti-inflammatory response to
protect mice from Group A streptococcal infection. Antimicrob
Agents Chemother. 57:5366–5372. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
McBride JD, Jenkins AJ, Liu X, Zhang B,
Lee K, Berry WL, Janknecht R, Griffin CT, Aston CE, Lyons TJ, et
al: Elevated circulation levels of an antiangiogenic SERPIN in
patients with diabetic microvascular complications impair wound
healing through suppression of Wnt signaling. J Invest Dermatol.
134:1725–1734. 2014. View Article : Google Scholar
|
|
70
|
Bach RR: Initiation of coagulation by
tissue factor. CRC Crit Rev Biochem. 23:339–368. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Tuo J, Bojanowski CM, Zhou M, Shen D, Ross
RJ, Rosenberg KI, Cameron DJ, Yin C, Kowalak JA, Zhuang Z, et al:
Murine ccl2/cx3cr1 deficiency results in retinal lesions mimicking
human age-related macular degeneration. Invest Ophthalmol Vis Sci.
48:3827–3836. 2007. View Article : Google Scholar
|
|
72
|
Chan CC, Ross RJ, Shen D, Ding X, Majumdar
Z, Bojanowski CM, Zhou M, Salem N Jr, Bonner R and Tuo J:
Ccl2/Cx3cr1-deficient mice: An animal model for age-related macular
degeneration. Ophthalmic Res. 40:124–128. 2008. View Article : Google Scholar
|
|
73
|
Chu XK, Wang Y, Ardeljan D, Tuo J and Chan
CC: Controversial view of a genetically altered mouse model of
focal retinal degeneration. Bioengineered. 4:130–135. 2013.
View Article : Google Scholar :
|
|
74
|
Tuo J, Ross RJ, Herzlich AA, Shen D, Ding
X, Zhou M, Coon SL, Hussein N, Salem N Jr and Chan CC: A high
omega-3 fatty acid diet reduces retinal lesions in a murine model
of macular degeneration. Am J Pathol. 175:799–807. 2009. View Article : Google Scholar
|
|
75
|
Tuo J, Pang JJ, Cao X, Shen D, Zhang J,
Scaria A, Wadsworth SC, Pechan P, Boye SL, Hauswirth WW and Chan
CC: AAV5-mediated sFLT01 gene therapy arrests retinal lesions in
Ccl2(-/-)/Cx3cr1(-/-) mice. Neurobiol Aging. 33. pp. 433.e1–e10.
2012, View Article : Google Scholar
|
|
76
|
Zhang J, Tuo J, Cao X, Shen D, Li W and
Chan CC: Early degeneration of photoreceptor synapse in
Ccl2/Cx3cr1-deficient mice on Crb1(rd8) background. Synapse.
67:515–531. 2013. View Article : Google Scholar
|
|
77
|
Clemons TE, Milton RC, Klein R and Seddon
JM: Risk factors for the incidence of advanced age-related macular
degeneration in the age-related eye disease study (AREDS) AREDS
report no. 19. Ophthalmology. 112:533–539. 2005. View Article : Google Scholar
|
|
78
|
Wang Y, Sang A, Zhu M, Zhang G, Guan H, Ji
M and Chen H: Tissue factor induces VEGF expression via activation
of the Wnt/β-catenin signaling pathway in ARPE-19 cells. Mol Vis.
22:886–897. 2016.
|
|
79
|
Qiu F, Liu Z, Zhou Y, He J, Gong S, Bai X,
Zeng Y, Liu Z and Ma JX: Decreased circulating levels of dickkopf-1
in patients with exudative age-related macular degeneration. Sci
Rep. 7:12632017. View Article : Google Scholar
|
|
80
|
Voorzanger-Rousselot N, Goehrig D, Facon
T, Clézardin P and Garnero P: Platelet is a major contributor to
circulating levels of Dickkopf-1: Clinical implications in patients
with multiple myeloma. Br J Haematol. 145:264–266. 2009. View Article : Google Scholar
|
|
81
|
Esen E, Chen J, Karner CM, Okunade AL,
Patterson BW and Long F: WNT-LRP5 signaling induces Warburg effect
through mTORC2 activation during osteoblast differentiation. Cell
Metab. 17:745–755. 2013. View Article : Google Scholar
|
|
82
|
Pate KT, Stringari C, Sprowl-Tanio S, Wang
K, TeSlaa T, Hoverter NP, McQuade MM, Garner C, Digman MA, Teitell
MA, et al: Wnt signaling directs a metabolic program of glycolysis
and angiogenesis in colon cancer. EMBO J. 33:1454–1473. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Thompson CB: Wnt meets Warburg: Another
piece in the puzzle? EMBO J. 33:1420–1422. 2014. View Article : Google Scholar
|
|
84
|
Lum JJ, Bui T, Gruber M, Gordan JD,
DeBerardinis RJ, Covello KL, Simon MC and Thompson CB: The
transcription factor HIF-1alpha plays a critical role in the growth
factor-dependent regulation of both aerobic and anaerobic
glycolysis. Genes Dev. 21:1037–1049. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Manea A: NADPH oxidase-derived reactive
oxygen species: Involvement in vascular physiology and pathology.
Cell Tissue Res. 342:325–339. 2010. View Article : Google Scholar
|
|
86
|
Radomska-Leśniewska DM, Hevelke A,
Skopiński P, Bałan B, Jóźwiak J, Rokicki D, Skopińska-Różewska E
and Białoszewska A: Reactive oxygen species and synthetic
antioxidants as angiogenesis modulators: Clinical implications.
Pharmacol Rep. 68:462–471. 2016. View Article : Google Scholar
|
|
87
|
Mittal M, Siddiqui MR, Tran K, Reddy SP
and Malik AB: Reactive oxygen species in inflammation and tissue
injury. Antioxid Redox Signal. 20:1126–1167. 2014. View Article : Google Scholar :
|
|
88
|
Kim YW, West XZ and Byzova TV:
Inflammation and oxidative stress in angiogenesis and vascular
disease. J Mol Med (Berl). 91:323–328. 2013. View Article : Google Scholar
|
|
89
|
Brambilla D, Mancuso C, Scuderi MR, Bosco
P, Cantarella G, Lempereur L, Di Benedetto G, Pezzino S and
Bernardini R: The role of antioxidant supplement in immune system,
neoplastic, and neurodegenerative disorders: A point of view for an
assessment of the risk/benefit profile. Nutr J. 7:292008.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Carmeliet P and Jain RK: Molecular
mechanisms and clinical applications of angiogenesis. Nature.
473:298–307. 2011. View Article : Google Scholar
|
|
91
|
Kim YW and Byzova TV: Oxidative stress in
angiogenesis and vascular disease. Blood. 123:625–631. 2014.
View Article : Google Scholar :
|
|
92
|
Vallée A, Lecarpentier Y, Vallée R,
Guillevin R and Vallée JN: Circadian rhythms in exudative
age-related macular degeneration: The key role of the canonical
WNT/β-catenin pathway. Int J Mol Sci. 21:8202020. View Article : Google Scholar
|
|
93
|
Brugarolas JB, Vazquez F, Reddy A, Sellers
WR and Kaelin WG: TSC2 regulates VEGF through mTOR-dependent and
-independent pathways. Cancer Cell. 4:147–158. 2003. View Article : Google Scholar
|
|
94
|
Düvel K, Yecies JL, Menon S, Raman P,
Lipovsky AI, Souza AL, Triantafellow E, Ma Q, Gorski R, Cleaver S,
et al: Activation of a metabolic gene regulatory network downstream
of mTOR complex 1. Mol Cell. 39:171–183. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Jung JE, Lee HG, Cho IH, Chung DH, Yoon
SH, Yang YM, Lee JW, Choi S, Park JW, Ye SK and Chung MH: STAT3 is
a potential modulator of HIF-1-mediated VEGF expression in human
renal carcinoma cells. FASEB J. 19:1296–1298. 2005. View Article : Google Scholar
|
|
96
|
Land SC and Tee AR: Hypoxia-inducible
factor 1alpha is regulated by the mammalian target of rapamycin
(mTOR) via an mTOR signaling motif. J Biol Chem. 282:20534–20543.
2007. View Article : Google Scholar
|
|
97
|
Toschi A, Lee E, Gadir N, Ohh M and Foster
DA: Differential dependence of hypoxia-inducible factors 1 alpha
and 2 alpha on mTORC1 and mTORC2. J Biol Chem. 283:34495–34499.
2008. View Article : Google Scholar
|
|
98
|
Xu Q, Briggs J, Park S, Niu G, Kortylewski
M, Zhang S, Gritsko T, Turkson J, Kay H, Semenza GL, et al:
Targeting Stat3 blocks both HIF-1 and VEGF expression induced by
multiple oncogenic growth signaling pathways. Oncogene.
24:5552–5560. 2005. View Article : Google Scholar
|
|
99
|
Kim J, Gao P, Liu YC, Semenza GL and Dang
CV: Hypoxia-inducible factor 1 and dysregulated c-Myc cooperatively
induce vascular endothelial growth factor and metabolic switches
hexokinase 2 and pyruvate dehydrogenase kinase 1. Mol Cell Biol.
27:7381–7393. 2007. View Article : Google Scholar
|
|
100
|
Suda T, Takubo K and Semenza GL: Metabolic
regulation of hematopoietic stem cells in the hypoxic niche. Cell
Stem Cell. 9:298–310. 2011. View Article : Google Scholar
|
|
101
|
Firth JD, Ebert BL and Ratcliffe PJ:
Hypoxic regulation of lactate dehydrogenase A. Interaction between
hypoxia-inducible factor 1 and cAMP response elements. J Biol Chem.
270:21021–21027. 1995. View Article : Google Scholar
|
|
102
|
Lewis BC, Shim H, Li Q, Wu CS, Lee LA,
Maity A and Dang CV: Identification of putative c-Myc-responsive
genes: Characterization of rcl, a novel growth-related gene. Mol
Cell Biol. 17:4967–4978. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Semenza GL, Jiang BH, Leung SW, Passantino
R, Concordet JP, Maire P and Giallongo A: Hypoxia response elements
in the Aldolase A, Enolase 1, and lactate dehydrogenase A gene
promoters contain essential binding sites for hypoxia-inducible
factor 1. J Biol Chem. 271:32529–32537. 1996. View Article : Google Scholar
|
|
104
|
Shim H, Dolde C, Lewis BC, Wu CS, Dang G,
Jungmann RA, Dalla-Favera R and Dang CV: c-Myc transactivation of
LDH-A: implications for tumor metabolism and growth. Proc Natl Acad
Sci USA. 94:6658–6663. 1997. View Article : Google Scholar
|
|
105
|
Warburg O: On the origin of cancer cells.
Science. 123:309–314. 1956. View Article : Google Scholar
|
|
106
|
Koukourakis MI, Giatromanolaki A, Sivridis
E, Bougioukas G, Didilis V, Gatter KC and Harris AL; Tumour and
Angiogenesis Research Group: Lactate dehydrogenase-5 (LDH-5)
overexpression in non-small-cell lung cancer tissues is linked to
tumour hypoxia, angiogenic factor production and poor prognosis. Br
J Cancer. 89:877–885. 2003. View Article : Google Scholar
|
|
107
|
Vallée A, Lecarpentier Y, Guillevin R and
Vallée JN: Aerobic glycolysis hypothesis through WNT/beta-catenin
pathway in exudative age-related macular degeneration. J Mol
Neurosci. 62:368–379. 2017. View Article : Google Scholar
|
|
108
|
Léveillard T and Sahel JA: Metabolic and
redox signaling in the retina. Cell Mol Life Sci. 74:3649–3665.
2017. View Article : Google Scholar
|
|
109
|
Oguma K, Oshima H and Oshima M:
Inflammation, tumor necrosis factor and Wnt promotion in gastric
cancer development. Future Oncol. 6:515–526. 2010. View Article : Google Scholar
|
|
110
|
Schön S, Flierman I, Ofner A, Stahringer
A, Holdt LM, Kolligs FT and Herbst A: β-catenin regulates NF-κB
activity via TNFRSF19 in colorectal cancer cells. Int J Cancer.
135:1800–1811. 2014. View Article : Google Scholar
|
|
111
|
Oh H, Takagi H, Takagi C, Suzuma K, Otani
A, Ishida K, Matsumura M, Ogura Y and Honda Y: The potential
angiogenic role of macrophages in the formation of choroidal
neovascular membranes. Invest Ophthalmol Vis Sci. 40:1891–1898.
1999.
|
|
112
|
Cousins SW, Espinosa-Heidmann DG and Csaky
KG: Monocyte activation in patients with age-related macular
degeneration: A biomarker of risk for choroidal neovascularization?
Arch Ophthalmol. 122:1013–1018. 2004. View Article : Google Scholar
|
|
113
|
Duguid IG, Boyd AW and Mandel TE: Adhesion
molecules are expressed in the human retina and choroid. Curr Eye
Res. 11(Suppl): S153–S159. 1992. View Article : Google Scholar
|
|
114
|
Elner SG, Elner VM, Pavilack MA, Todd RF
III, Mayo-Bond L, Franklin WA, Strieter RM, Kunkel SL and Huber AR:
Modulation and function of intercellular adhesion molecule-1 (CD54)
on human retinal pigment epithelial cells. Lab Invest. 66:200–211.
1992.
|
|
115
|
Anderson DH, Mullins RF, Hageman GS and
Johnson LV: A role for local inflammation in the formation of
drusen in the aging eye. Am J Ophthalmol. 134:411–431. 2002.
View Article : Google Scholar
|
|
116
|
Donoso LA, Kim D, Frost A, Callahan A and
Hageman G: The role of inflammation in the pathogenesis of
age-related macular degeneration. Surv Ophthalmol. 51:137–152.
2006. View Article : Google Scholar
|
|
117
|
Easwaran V, Lee SH, Inge L, Guo L,
Goldbeck C, Garrett E, Wiesmann M, Garcia PD, Fuller JH, Chan V, et
al: Beta-Catenin regulates vascular endothelial growth factor
expression in colon cancer. Cancer Res. 63:3145–3153. 2003.
|
|
118
|
Ip MS, Scott IU, Brown GC, Brown MM, Ho
AC, Huang SS and Recchia FM; American Academy of Ophthalmology:
Anti-vascular endothelial growth factor pharmacotherapy for
age-related macular degeneration: A report by the American Academy
of Ophthalmology. Ophthalmology. 115:1837–1846. 2008. View Article : Google Scholar
|
|
119
|
Wolf S: Current status of anti-vascular
endothelial growth factor therapy in Europe. Jpn J Ophthalmol.
52:433–439. 2008. View Article : Google Scholar
|
|
120
|
Menon G and Walters G: New paradigms in
the treatment of wet AMD: The impact of anti-VEGF therapy. Eye
(Lond). 23(Suppl 1): S1–S7. 2009. View Article : Google Scholar
|
|
121
|
Grisanti S, Zhu Q, Tatar O, Lueke J, Lueke
M, Tura A and Grisanti S: Differential expression of vascular
endothelial growth factor-a isoforms in neovascular age-related
macular degeneration. Retina. 35:764–772. 2015. View Article : Google Scholar
|
|
122
|
Liu X: Overstimulation can create health
problems due to increases in PI3K/Akt/GSK3 insensitivity and GSK3
activity. Springerplus. 3:3562014. View Article : Google Scholar
|
|
123
|
Zhang P, Wang Y, Hui Y, Hu D, Wang H, Zhou
J and Du H: Inhibition of VEGF expression by targeting HIF-1 alpha
with small interference RNA in human RPE cells. Ophthalmologica.
221:411–417. 2007. View Article : Google Scholar
|
|
124
|
Zhang P, Zhang X, Hao X, Wang Y, Hui Y,
Wang H, Hu D and Zhou J: Rac1 activates HIF-1 in retinal pigment
epithelium cells under hypoxia. Graefes Arch Clin Exp Ophthalmol.
247:633–639. 2009. View Article : Google Scholar
|
|
125
|
Arjamaa O, Nikinmaa M, Salminen A and
Kaarniranta K: Regulatory role of HIF-1alpha in the pathogenesis of
age-related macular degeneration (AMD). Ageing Res Rev. 8:349–358.
2009. View Article : Google Scholar
|
|
126
|
Koukourakis MI, Giatromanolaki A, Sivridis
E, Gatter KC, Trarbach T, Folprecht G, Shi MM, Lebwohl D, Jalava T,
Laurent D, et al: Prognostic and predictive role of lactate
dehydrogenase 5 expression in colorectal cancer patients treated
with PTK787/ZK 222584 (vatalanib) antiangiogenic therapy. Clin
Cancer Res. 17:4892–4900. 2011. View Article : Google Scholar
|
|
127
|
Giatromanolaki A, Sivridis E, Gatter KC,
Turley H, Harris AL and Koukourakis MI; Tumour and Angiogenesis
Research Group: Lactate dehydrogenase 5 (LDH-5) expression in
endometrial cancer relates to the activated VEGF/VEGFR2(KDR)
pathway and prognosis. Gynecol Oncol. 103:912–918. 2006. View Article : Google Scholar
|
|
128
|
Kolev Y, Uetake H, Takagi Y and Sugihara
K: Lactate dehydrogenase-5 (LDH-5) expression in human gastric
cancer: Association with hypoxia-inducible factor (HIF-1alpha)
pathway, angiogenic factors production and poor prognosis. Ann Surg
Oncol. 15:2336–2344. 2008. View Article : Google Scholar
|
|
129
|
Dhup S, Dadhich RK, Porporato PE and
Sonveaux P: Multiple biological activities of lactic acid in
cancer: Influences on tumor growth, angiogenesis and metastasis.
Curr Pharm Des. 18:1319–1330. 2012. View Article : Google Scholar
|
|
130
|
Polet F and Feron O: Endothelial cell
metabolism and tumour angiogenesis: Glucose and glutamine as
essential fuels and lactate as the driving force. J Intern Med.
273:156–165. 2013. View Article : Google Scholar
|
|
131
|
San-Millán I and Brooks GA: Reexamining
cancer metabolism: Lactate production for carcinogenesis could be
the purpose and explanation of the Warburg Effect. Carcinogenesis.
38:119–133. 2017.
|
|
132
|
Liu W, Zhai Y, Heng X, Che FY, Chen W, Sun
D and Zhai G: Oral bioavailability of curcumin: problems and
advancements. J Drug Target. 24:694–702. 2016. View Article : Google Scholar
|
|
133
|
Vallée A and Lecarpentier Y: Curcumin and
endometriosis. Int J Mol Sci. 21:24402020. View Article : Google Scholar :
|
|
134
|
Kotha RR and Luthria DL: Curcumin:
Biological, Pharmaceutical, Nutraceutical, and Analytical Aspects.
Molecules. 24:29302019. View Article : Google Scholar
|
|
135
|
Prasad S, Gupta SC, Tyagi AK and Aggarwal
BB: Curcumin, a component of golden spice: From bedside to bench
and back. Biotechnol Adv. 32:1053–1064. 2014. View Article : Google Scholar
|
|
136
|
Priyadarsini KI: The chemistry of
curcumin: From extraction to therapeutic agent. Molecules.
19:20091–20112. 2014. View Article : Google Scholar
|
|
137
|
Zhang L, Zhu W, Yang C, Guo H, Yu A, Ji J,
Gao Y, Sun M and Zhai G: A novel folate-modified
self-microemulsifying drug delivery system of curcumin for colon
targeting. Int J Nanomedicine. 7:151–162. 2012.
|
|
138
|
Shen L, Liu CC, An CY and Ji HF: How does
curcumin work with poor bioavailability? Clues from experimental
and theoretical studies. Sci Rep. 6:208722016. View Article : Google Scholar :
|
|
139
|
Sun M, Su X, Ding B, He X, Liu X, Yu A,
Lou H and Zhai G: Advances in nanotechnology-based delivery systems
for curcumin. Nanomedicine (Lond). 7:1085–1100. 2012. View Article : Google Scholar
|
|
140
|
Naksuriya O, Okonogi S, Schiffelers RM and
Hennink WE: Curcumin nanoformulations: A review of pharmaceutical
properties and preclinical studies and clinical data related to
cancer treatment. Biomaterials. 35:3365–3383. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Malam Y, Loizidou M and Seifalian AM:
Liposomes and nanoparticles: Nanosized vehicles for drug delivery
in cancer. Trends Pharmacol Sci. 30:592–599. 2009. View Article : Google Scholar
|
|
142
|
Lee WH, Loo CY, Young PM, Traini D, Mason
RS and Rohanizadeh R: Recent advances in curcumin nanoformulation
for cancer therapy. Expert Opin Drug Deliv. 11:1183–1201. 2014.
View Article : Google Scholar
|
|
143
|
Hatefi A and Amsden B: Biodegradable
injectable in situ forming drug delivery systems. J Control
Release. 80:9–28. 2002. View Article : Google Scholar
|
|
144
|
Yallapu MM, Jaggi M and Chauhan SC:
Beta-Cyclodextrin-curcumin self-assembly enhances curcumin delivery
in prostate cancer cells. Colloids Surf B Biointerfaces.
79:113–125. 2010. View Article : Google Scholar
|
|
145
|
Radomska-Leśniewska DM, Osiecka-Iwan A,
Hyc A, Góźdź A, Dąbrowska AM and Skopiński P: Therapeutic potential
of curcumin in eye diseases. Cent Eur J Immunol. 44:181–189. 2019.
View Article : Google Scholar
|
|
146
|
Zhu W, Wu Y, Meng YF, Wang JY, Xu M, Tao
JJ and Lu J: Effect of curcumin on aging retinal pigment epithelial
cells. Drug Des Devel Ther. 9:5337–5344. 2015.
|
|
147
|
Jiang X, Li S, Qiu X, Cong J, Zhou J and
Miu W: Curcumin inhibits cell viability and increases apoptosis of
SW620 human colon adenocarcinoma cells via the caudal type
homeobox-2 (CDX2)/Wnt/β-catenin pathway. Med Sci Monit.
25:7451–7458. 2019. View Article : Google Scholar
|
|
148
|
Howell JC, Chun E, Farrell AN, Hur EY,
Caroti CM, Iuvone PM and Haque R: Global microRNA expression
profiling: Curcumin (diferuloylmethane) alters oxidative
stress-responsive microRNAs in human ARPE-19 cells. Mol Vis.
19:544–560. 2013.
|
|
149
|
Mandal MNA, Patlolla JMR, Zheng L, Agbaga
MP, Tran JT, Wicker L, Kasus-Jacobi A, Elliott MH, Rao CV and
Anderson RE: Curcumin protects retinal cells from light-and oxidant
stress-induced cell death. Free Radic Biol Med. 46:672–679. 2009.
View Article : Google Scholar
|
|
150
|
Muangnoi C, Sharif U, Ratnatilaka Na,
Bhuket P, Rojsitthisak P and Paraoan L: Protective effects of
curcumin ester prodrug, curcumin diethyl disuccinate against
H2O2-Induced oxidative stress in human
retinal pigment epithelial cells: Potential therapeutic avenues for
age-related macular degeneration. Int J Mol Sci. 20:33672019.
View Article : Google Scholar
|
|
151
|
Kim HJ, Park SY, Park OJ and Kim YM:
Curcumin suppresses migration and proliferation of Hep3B
hepatocarcinoma cells through inhibition of the Wnt signaling
pathway. Mol Med Rep. 8:282–286. 2013. View Article : Google Scholar
|
|
152
|
Leow PC, Bahety P, Boon CP, Lee CY, Tan
KL, Yang T and Ee PL: Functionalized curcumin analogs as potent
modulators of the Wnt/β-catenin signaling pathway. Eur J Med Chem.
71:67–80. 2014. View Article : Google Scholar
|
|
153
|
Kolb TM and Davis MA: The tumor promoter
12-O-tetradecanoylphorbol 13-acetate (TPA) provokes a prolonged
morphologic response and ERK activation in Tsc2-null renal tumor
cells. Toxicol Sci. 81:233–242. 2004. View Article : Google Scholar
|
|
154
|
Balasubramanyam K, Varier RA, Altaf M,
Swaminathan V, Siddappa NB, Ranga U and Kundu TK: Curcumin, a novel
p300/CREB-binding protein-specific inhibitor of acetyltransferase,
represses the acetylation of histone/nonhistone proteins and
histone acetyltransferase-dependent chromatin transcription. J Biol
Chem. 279:51163–51171. 2004. View Article : Google Scholar
|
|
155
|
He M, Li Y, Zhang L, Li L, Shen Y, Lin L,
Zheng W, Chen L, Bian X, Ng HK and Tang L: Curcumin suppresses cell
proliferation through inhibition of the Wnt/β-catenin signaling
pathway in medulloblastoma. Oncol Rep. 32:173–180. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Menon VP and Sudheer AR: Antioxidant and
anti-inflammatory properties of curcumin. Adv Exp Med Biol.
595:105–125. 2007. View Article : Google Scholar
|
|
157
|
Marchiani A, Rozzo C, Fadda A, Delogu G
and Ruzza P: Curcumin and curcumin-like molecules: From spice to
drugs. Curr Med Chem. 21:204–222. 2014. View Article : Google Scholar
|
|
158
|
Schneider C, Boeglin WE, Yin H, Stec DF
and Voehler M: Convergent oxygenation of arachidonic acid by
5-lipoxygenase and cyclooxygenase-2. J Am Chem Soc. 128:720–721.
2006. View Article : Google Scholar
|
|
159
|
Giménez-Bastida JA, González-Sarrías A,
Laparra-Llopis JM, Schneider C and Espín JC: Targeting mammalian
5-lipoxygenase by dietary phenolics as an anti-inflammatory
mechanism: A systematic review. Int J Mol Sci. 22:79372021.
View Article : Google Scholar :
|
|
160
|
Othman A, Ahmad S, Megyerdi S, Mussell R,
Choksi K, Maddipati KR, Elmarakby A, Rizk N and Al-Shabrawey M:
12/15-Lipoxygenase-derived lipid metabolites induce retinal
endothelial cell barrier dysfunction: Contribution of NADPH
oxidase. PLoS One. 8:e572542013. View Article : Google Scholar
|
|
161
|
Subramanian P, Mendez EF and Becerra SP: A
novel inhibitor of 5-Lipoxygenase (5-LOX) prevents oxidative
stress-induced cell death of retinal pigment epithelium (RPE)
cells. Invest Ophthalmol Vis Sci. 57:4581–4588. 2016. View Article : Google Scholar
|
|
162
|
Yadav UCS and Ramana KV: Regulation of
NF-κB-induced inflammatory signaling by lipid peroxidation-derived
aldehydes. Oxid Med Cell Longev. 2013:6905452013. View Article : Google Scholar
|
|
163
|
Ruan Y, Jiang S and Gericke A: Age-related
macular degeneration: Role of oxidative stress and blood vessels.
Int J Mol Sci. 22:12962021. View Article : Google Scholar :
|
|
164
|
Yabas M, Orhan C, Er B, Tuzcu M, Durmus
AS, Ozercan IH, Sahin N, Bhanuse P, Morde AA, Padigaru M and Sahin
K: A next generation formulation of curcumin ameliorates
experimentally induced osteoarthritis in rats via regulation of
inflammatory mediators. Front Immunol. 12:6096292021. View Article : Google Scholar :
|
|
165
|
Li X, Lu Y, Jin Y, Son JK, Lee SH and
Chang HW: Curcumin inhibits the activation of immunoglobulin
e-mediated mast cells and passive systemic anaphylaxis in mice by
reducing serum eicosanoid and histamine levels. Biomol Ther
(Seoul). 22:27–34. 2014. View Article : Google Scholar
|
|
166
|
Manjunatha H and Srinivasan K: Protective
effect of dietary curcumin and capsaicin on induced oxidation of
low-density lipoprotein, iron-induced hepatotoxicity and
carrageenan-induced inflammation in experimental rats. FEBS J.
273:4528–4537. 2006. View Article : Google Scholar
|
|
167
|
Priyadarsini KI, Maity DK, Naik GH, Kumar
MS, Unnikrishnan MK, Satav JG and Mohan H: Role of phenolic O-H and
methylene hydrogen on the free radical reactions and antioxidant
activity of curcumin. Free Radic Biol Med. 35:475–484. 2003.
View Article : Google Scholar
|
|
168
|
Piwocka K, Jaruga E, Skierski J, Gradzka I
and Sikora E: Effect of glutathione depletion on caspase-3
independent apoptosis pathway induced by curcumin in Jurkat cells.
Free Radic Biol Med. 31:670–678. 2001. View Article : Google Scholar
|
|
169
|
Motterlini R, Foresti R, Bassi R and Green
CJ: Curcumin, an antioxidant and anti-inflammatory agent, induces
heme oxygenase-1 and protects endothelial cells against oxidative
stress. Free Radic Biol Med. 28:1303–1312. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
170
|
Cao K, Dong YT, Xiang J, Xu Y, Hong W,
Song H and Guan ZZ: Reduced expression of SIRT1 and SOD-1 and the
correlation between these levels in various regions of the brains
of patients with Alzheimer's disease. J Clin Pathol. 71:1090–1099.
2018. View Article : Google Scholar
|
|
171
|
Golestaneh N, Chu Y, Cheng SK, Cao H,
Poliakov E and Berinstein DM: Repressed SIRT1/PGC-1α pathway and
mitochondrial disintegration in iPSC-derived RPE disease model of
age-related macular degeneration. J Transl Med. 14:3442016.
View Article : Google Scholar
|
|
172
|
Zuo L, Khan RS, Lee V, Dine K, Wu W and
Shindler KS: SIRT1 promotes RGC survival and delays loss of
function following optic nerve crush. Invest Ophthalmol Vis Sci.
54:5097–5102. 2013. View Article : Google Scholar
|
|
173
|
Li K, Zhai M, Jiang L, Song F, Zhang B, Li
J, Li H, Li B, Xia L, Xu L, et al: Tetrahydrocurcumin ameliorates
diabetic cardiomyopathy by attenuating high glucose-induced
oxidative stress and fibrosis via activating the SIRT1 Pathway.
Oxid Med Cell Longev. 2019:67469072019. View Article : Google Scholar
|
|
174
|
Ghasemi F, Shafiee M, Banikazemi Z,
Pourhanifeh MH, Khanbabaei H, Shamshirian A, Amiri Moghadam S,
ArefNezhad R, Sahebkar A, Avan A and Mirzaei H: Curcumin inhibits
NF-kB and Wnt/β-catenin pathways in cervical cancer cells. Pathol
Res Pract. 215:1525562019. View Article : Google Scholar
|
|
175
|
Olivera A, Moore TW, Hu F, Brown AP, Sun
A, Liotta DC, Snyder JP, Yoon Y, Shim H, Marcus AI, et al:
Inhibition of the NF-κB signaling pathway by the curcumin analog,
3,5-Bis(2-pyridinylmethylidene)-4-piperidone (EF31): Anti-inf
lammatory and anti-cancer properties. Int Immunopharmacol.
12:368–377. 2012. View Article : Google Scholar
|
|
176
|
da Cruz BO, Cardozo LFM de F, Magliano DC
and Stockler-Pinto MB: Nutritional strategies to modulate
inflammation pathways via regulation of peroxisome
proliferator-activated receptor β/δ. Nutr Rev. 78:207–214.
2020.
|
|
177
|
Lin YG, Kunnumakkara AB, Nair A, Merritt
WM, Han LY, Armaiz-Pena GN, Kamat AA, Spannuth WA, Gershenson DM,
Lutgendorf SK, et al: Curcumin inhibits tumor growth and
angiogenesis in ovarian carcinoma by targeting the nuclear
factor-kappaB pathway. Clin Cancer Res. 13:3423–3430. 2007.
View Article : Google Scholar
|
|
178
|
Zhang ZB, Luo DD, Xie JH, Xian YF, Lai ZQ,
Liu YH, Liu WH, Chen JN, Lai XP, Lin ZX and Su ZR: Curcumin's
metabolites, tetrahydrocurcumin and octahydrocurcumin, possess
superior anti-inflammatory effects in vivo through suppression of
TAK1-NF-κB pathway. Front Pharmacol. 9:11812018. View Article : Google Scholar
|
|
179
|
Chen W, Chen Y and Cui G: Effects of
TNF-alpha and curcumin on the expression of VEGF in Raji and U937
cells and on angiogenesis in ECV304 cells. Chin Med J (Engl).
118:2052–2057. 2005.
|
|
180
|
Yoysungnoen P, Wirachwong P, Bhattarakosol
P, Niimi H and Patumraj S: Effects of curcumin on tumor
angiogenesis and biomarkers, COX-2 and VEGF, in hepatocellular
carcinoma cell-implanted nude mice. Clin Hemorheol Microcirc.
34:109–115. 2006.PubMed/NCBI
|
|
181
|
Li L, Braiteh FS and Kurzrock R:
Liposome-encapsulated curcumin: In vitro and in vivo effects on
proliferation, apoptosis, signaling, and angiogenesis. Cancer.
104:1322–1331. 2005. View Article : Google Scholar
|
|
182
|
Arbiser JL, Klauber N, Rohan R, van
Leeuwen R, Huang MT, Fisher C, Flynn E and Byers HR: Curcumin is an
in vivo inhibitor of angiogenesis. Mol Med. 4:376–383. 1998.
View Article : Google Scholar
|
|
183
|
Gururaj AE, Belakavadi M, Venkatesh DA,
Marmé D and Salimath BP: Molecular mechanisms of anti-angiogenic
effect of curcumin. Biochem Biophys Res Commun. 297:934–942. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
184
|
Zahra FT, Sajib MS and Mikelis CM: Role of
bFGF in acquired resistance upon Anti-VEGF therapy in cancer.
Cancers (Basel). 13:14222021. View Article : Google Scholar
|
|
185
|
Compagni A, Wilgenbus P, Impagnatiello MA,
Cotten M and Christofori G: Fibroblast growth factors are required
for efficient tumor angiogenesis. Cancer Res. 60:7163–7169.
2000.
|
|
186
|
Nissen LJ, Cao R, Hedlund EM, Wang Z, Zhao
X, Wetterskog D, Funa K, Bråkenhielm E and Cao Y: Angiogenic
factors FGF2 and PDGF-BB synergistically promote murine tumor
neovascularization and metastasis. J Clin Invest. 117:2766–2777.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
187
|
Casanovas O, Hicklin DJ, Bergers G and
Hanahan D: Drug resistance by evasion of antiangiogenic targeting
of VEGF signaling in late-stage pancreatic islet tumors. Cancer
Cell. 8:299–309. 2005. View Article : Google Scholar
|
|
188
|
Choi HJ, Armaiz Pena GN, Pradeep S, Cho
MS, Coleman RL and Sood AK: Anti-vascular therapies in ovarian
cancer: Moving beyond anti-VEGF approaches. Cancer Metastasis Rev.
34:19–40. 2015. View Article : Google Scholar :
|
|
189
|
Aggarwal BB and Natarajan K: Tumor
necrosis factors: Developments during the last decade. Eur Cytokine
Netw. 7:93–124. 1996.PubMed/NCBI
|
|
190
|
Li H, Soria C, Griscelli F, Opolon P,
Soria J, Yeh P, Legrand C, Vannier JP, Belin D, Perricaudet M and
Lu H: Amino-terminal fragment of urokinase inhibits tumor cell
invasion in vitro and in vivo: Respective contribution of the
urokinase plasminogen activator receptor-dependent or -independent
pathway. Hum Gene Ther. 16:1157–1167. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
191
|
Aggarwal BB, Kumar A and Bharti AC:
Anticancer potential of curcumin: Preclinical and clinical studies.
Anticancer Res. 23:363–398. 2003.
|
|
192
|
Wang LL, Sun Y, Huang K and Zheng L:
Curcumin, a potential therapeutic candidate for retinal diseases.
Mol Nutr Food Res. 57:1557–1568. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
193
|
Pittalà V, Fidilio A, Lazzara F, Platania
CBM, Salerno L, Foresti R, Drago F and Bucolo C: Effects of novel
nitric oxide-releasing molecules against oxidative stress on
retinal pigmented epithelial cells. Oxid Med Cell Longev.
2017:14208922017. View Article : Google Scholar : PubMed/NCBI
|
|
194
|
Bucolo C, Drago F, Maisto R, Romano GL,
D'Agata V, Maugeri G and Giunta S: Curcumin prevents high glucose
damage in retinal pigment epithelial cells through ERK1/2-mediated
activation of the Nrf2/HO-1 pathway. J Cell Physiol.
234:17295–17304. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
195
|
Vyas A, Dandawate P, Padhye S, Ahmad A and
Sarkar F: Perspectives on New Synthetic curcumin analogs and their
potential anticancer properties. Curr Pharm Des. 19:2047–2069.
2013.
|
|
196
|
Muangnoi C, Ratnatilaka Na Bhuket P,
Jithavech P, Supasena W, Paraoan L, Patumraj S and Rojsitthisak P:
Curcumin diethyl disuccinate, a prodrug of curcumin, enhances
anti-proliferative effect of curcumin against HepG2 cells via
apoptosis induction. Sci Rep. 9:117182019. View Article : Google Scholar :
|
|
197
|
Ohori H, Yamakoshi H, Tomizawa M, Shibuya
M, Kakudo Y, Takahashi A, Takahashi S, Kato S, Suzuki T, Ishioka C,
et al: Synthesis and biological analysis of new curcumin analogues
bearing an enhanced potential for the medicinal treatment of
cancer. Mol Cancer Ther. 5:2563–2571. 2006. View Article : Google Scholar
|
|
198
|
Shoba G, Joy D, Joseph T, Majeed M,
Rajendran R and Srinivas PS: Influence of piperine on the
pharmacokinetics of curcumin in animals and human volunteers.
Planta Med. 64:353–356. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
199
|
Sai N, Dong X, Huang P, You L, Yang C, Liu
Y, Wang W, Wu H, Yu Y, Du Y, et al: A novel gel-forming solution
based on PEG-DSPE/Solutol HS 15 Mixed Micelles and Gellan Gum for
ophthalmic delivery of curcumin. Molecules. 25:812019. View Article : Google Scholar
|
|
200
|
Zhang J, Sun H, Zhou N, Zhang B and Ma J:
Preparation and evaluation of biodegradable scleral plug containing
curcumin in rabbit eye. Curr Eye Res. 42:1597–1603. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
201
|
Mazzolani F, Togni S, Giacomelli L,
Eggenhoffner R and Franceschi F: Oral administration of a
curcumin-phospholipid formulation (Meriva®) for treatment of
chronic diabetic macular edema: A pilot study. Eur Rev Med
Pharmacol Sci. 22:3617–3625. 2018.PubMed/NCBI
|
|
202
|
Allegri P, Mastromarino A and Neri P:
Management of chronic anterior uveitis relapses: Efficacy of oral
phospholipidic curcumin treatment. Long-term follow-up. Clin
Ophthalmol. 4:1201–1206. 2010.
|
|
203
|
Chen J and Smith LEH: Retinopathy of
prematurity. Angiogenesis. 10:133–140. 2007. View Article : Google Scholar
|