Introduction
White matter injury (WMI) is a major form of preterm
brain injury induced by hypoxic ischemia (HI), particularly between
23 and 32 weeks of gestational age, a period that corresponds to
the peak of myelination (1). WMI
impairs the differentiation ability of oligodendrocyte progenitor
cells (OPCs), resulting in oligodendrocyte (OL) deficiency and
deficits in myelination, leading to cognitive and behavioral
disorders that negatively impact survival and quality of life in
children (2,3). In this regard, impaired
differentiation ability of OPCs constitutes a key mechanism of WMI
pathogenesis. Currently, clinical treatments for mature OL
deficiency are limited. Hence, there is an urgent need to explore
strategies to promote the differentiation and maturation of OPCs,
which may provide novel therapeutic approaches for WMI.
The myelin sheath is a major component of white
matter and is formed by mature OLs that differentiate from OPCs
(4). The myelinating
microenvironment is a key factor that hinders differentiation and
maturation of OPCs after WMI (5).
Therefore, optimizing the myelinating microenvironment, which is
formed by a network of neuronal astrocytes and microglial blood
vessels, is an important means to promote the differentiation and
maturation of OPCs (5). Based on
extant literature and previous in vitro experiments by the
authors, several endogenous molecules have been identified to
directly or indirectly improve the myelinating microenvironment and
promote the differentiation of human OPCs (5). Among them, Activin A (Act A) plays a
key role (6). Act A is a widely
expressed homodimer composed of two βA chains. Sequence
analysis has revealed that the β subunit of Act A possesses the
typical structural features of the transforming growth factor-β
superfamily, and the mature human βA chain of Act A has
100% amino acid sequence identity in cattle, cats, mice, and pigs,
highlighting its highly conserved structure (7). In the nervous system, Act A is
secreted by neurons and glial cells, which exert neuroprotective
effects. A previous study reported that treatment of OPCs cultured
in vitro with recombinant Act A protein promoted OPC
proliferation and differentiation (6). Further, another study reported that
Act A improved neurological outcomes by regulating OPC function in
adult male mice (8).
Collectively, these reports suggested that Act A may be used in the
treatment of myelination disorders. However, the roles and
mechanisms of action of Act A in preterm brain injury remain
unclear. Given the considerable differences between the adult and
preterm brains in responses to external stimuli such as HI, it is
essential to examine the effects of Act A in the preterm brain.
Therefore, the present study aimed to investigate whether exogenous
Act A treatment could enhance OPC differentiation in WMI and to
explore the underlying mechanisms.
Materials and methods
Animals
Brain development of newborn Sprague-Dawley (SD)
rats aged 2-5 days has been reported to be consistent with that of
human fetuses aged 23-32 weeks (9,10).
In order to avoid the effects of high mortality on the modeling
process of young rats, postnatal day 5 (P5) SD rats were used for
modeling.
In the present study, a total of 1,335
specific-pathogen-free, P5 healthy male (SD) rats (average weight:
10-15 g) were purchased from Sichuan Dashuo Animal Science and
Technology Co., Ltd. (Chengdu, China). A total of six rats were
used for each group and each group was maintained with one cage
under a 12-h light/dark cycle. The relative humidity was controlled
at 40-70% and the temperature at 23±2°C, with ad libitum access to
food and water. During the experimental period (from P5 to P35), if
any rat began to show signs of inability to move or eat, ruffled
fur or self-mutilation, they were immediately sacrificed. In
addition, animals were euthanized to prevent further suffering if
they were unable to stand or displayed agonal breathing, severe
muscular atrophy, severe ulcers or uncontrolled bleeding. The rats
with unsuccessful modeling were euthanized by cervical dislocation
under anesthesia for euthanasia. Complete cardiac and respiratory
arrest were observed to verify animal death. It was confirmed that
the animal studies abided to all of the animal welfare, including
efforts to minimize suffering and distress, use of analgesics or
anesthetics (including the dose and duration), or special housing
conditions. All animal experiments were approved (approval no.
WCSUH21-2018-034) by the Sichuan University Committee on Animal
Research (Chengdu, China) and complied with the ARRIVE
guidelines.
WMI modeling
The rats were randomly divided into WMI and Sham
groups. The WMI group was established using the following procedure
as previously described (11).
First, P5 neonatal rats were fixed on their backs after general
anesthesia. A 1-cm longitudinal incision was made in the neck, and
the right carotid artery was exposed and ligated after separation
from glands and muscle tissue. After surgery, the rats were placed
in an incubator for 30 min to recover. Subsequently, the rats were
placed in an 8%-oxygen and 92%-nitrogen cabin (8% O2 and
92% N2) with a gas flow rate of 3 l/min for 2 h to
induce WMI. Rats were maintained on a heating pad during surgical
procedures to maintain body temperature at 36-37°C. Rats in the
Sham group were only subjected to neck incision to dissociate the
right carotid artery, without ligation or hypoxia. Following
surgery, all neonatal rat pups were returned to their cages.
Establishment of testing and control
groups
After 24 h of WMI modeling, the WMI group was
treated with Act A or PBS to establish the Act A and PBS groups as
testing and control groups, respectively. To establish the Act A
and PBS groups, rats were injected with 5 µl of Act A (12.5,
25, and 50 mg/kg) or PBS, respectively, using a Hamilton syringe
needle via the lateral ventricle (LV), located 2 mm posterior and 2
mm lateral (right) from bregma to a needle depth of 2 mm. Next, the
Act A group was treated with Id2-overexpressing lentiviral vector
and mock-vehicle to establish the Id2 and V groups as the testing
and control groups, respectively. To establish the Id2 and V
groups, rats in the Act A group were injected with 4 µl of
Id2 (1×109 TU/ml)-overexpressing lentiviral vector or
mock-vehicle into the lateral ventricle using a Hamilton syringe
needle 6 h after Act A treatment.
Hematoxylin & eosin staining
On P7, the rats were sequentially perfused with 0.9%
normal saline and 4% paraformaldehyde (100 ml each), after which
tissues were extracted and post-fixed in a 4% paraformaldehyde
solution for 24-36 h at 4°C. Then, the tissues were
paraffin-embedded and 5-mm thick serial sections were made in the
coronal plane. A total three 3 sections containing the corpus
callosum (CC) (0.26-1.80 mm behind the anterior fontanelle
according to the rat brain atlas) were selected for analysis.
Finally, the sectioned tissues were stained with hematoxylin &
eosin (H&E) and were observed using an inverted optical
microscope (Leica Microsystems GmbH). Randomly selected fields
(n=4) were examined in each animal. A total of six animals per
group were analyzed.
Immunofluorescence staining
The rat brains were obtained at set time points (P7,
P14, P21, P28, and P35) and post-fixed in 4% paraformaldehyde at
4°C for at least 48 h, then embedded in 2-3% agarose. Coronal brain
sections were cut using an oscillating tissue slicer (Leica
Microsystems GmbH). A total of three sections containing the CC
were selected for analysis. The sections were first washed in PBS
and incubated in 0.3% Triton X-100 at room temperature for 30 min
and then incubated for 1 h in fetal calf serum (Gibco; Thermo
Fisher Scientific, Inc.) to inhibit non-specific binding. Next, the
brain sections were incubated with primary antibodies (Table I) at 4°C overnight, then incubated
for 2 h at room temperature with secondary antibodies (Table I). Finally, fluorescence imaging
was performed using a confocal laser scanning microscope (Olympus
Corporation) and FV-ASW-3.1 software (Olympus Corporation), and
mean fluorescence intensity or positive cells were quantified. Mean
fluorescence intensity was defined as the ratio between the sum of
the integral optical density of the target protein and area.
Positive cells and mean fluorescence intensity were quantified for
each field with a ×40 objective lens (field size, 0.24
mm2) using ImageJ 1.8.0.345 software (National
Institutes of Health). Randomly selected fields (n=6) from the CC
were examined in each animal. A total of six animals per group were
analyzed.
 | Table IInformation of the antibodies used in
immunofluorescence and western blot experiments. |
Table I
Information of the antibodies used in
immunofluorescence and western blot experiments.
| | Antibody name | Host species | Dilution | Cat.
no./Supplier |
|---|
|
Immunofluorescence | Primary
antibody | Id2 | Rabbit | 1:500 | NBP2-27194/Novus
Biologicals, LLC |
| BMP4 | Rabbit | 1:500 | ab39973/Abcam |
| Olig2 | Rabbit | 1:500 | AB9610/Abcam |
| Ki67 | Rabbit | 1:500 | ab15580/Abcam |
| Vimentin | Mouse | 1:200 | ab8978/Abcam |
| CC3 | Rabbit | 1:500 |
PA5-77887/Invitrogen; Thermo Fisher
Scientific, Inc. |
| NG2 | Rabbit | 1:200 | AB5320/Abcam |
| O4 | Mouse | 1:25 |
MAB345/MilliporeSigma |
| CC-1 | Mouse | 1:200 | ab16794/Abcam |
| MBP | Mouse | 1:1,000 | ARG10722/Arigo
Biolaboratories, Inc. |
| MAG | Rabbit | 1:100 | ab89780/Abcam |
| PLP | Rabbit | 1:1,000 | ab28486/Abcam |
| Tau1 | Mouse | 1:1,000 |
MAB3420/MilliporeSigma |
| SMI31 | Mouse | 1:1,000 | SMI31P/BioLegend,
Inc. |
| SMI312 | Mouse | 1:1,000 | 837904/BioLegend,
Inc. |
| Secondary
antibody |
Cy3/488-conjugated | Donkey
anti-rabbit/mouse IgG | 1:500 |
712-166-150/715-165-150
711-545-152/715-545-150/Jackson ImmunoResearch Laboratories,
Inc. |
| Western blot
analysis | Primary
antibody | Act A | Rabbit | 1:500 | NBP1-30928/Novus
Biologicals, LLC |
| MBP | Mouse | 1:500 | ARG10722/Arigobio
Biolaboratories, Inc. |
| MAG | Rabbit | 1:100 | ab89780/Abcam |
| PLP | Rabbit | 1:500 | ab28486/Abcam |
| Tau1 | Mouse | 1:500 |
MAB3420/MilliporeSigma |
| SMI31 | Mouse | 1:500 | SMI31P/BioLegend,
Inc. |
| SMI312 | Mouse | 1:500 | 837904/BioLegend,
Inc. |
| Noggin | Mouse | 1:200 | ab239520/Abcam |
| BMP4 | Rabbit | 1:500 | ab39973/Abcam |
| Id2 | Rabbit | 1:500 | NBP2-27194/Novus
Biologicals, LLC |
| Actin | Mouse | 1:5,000 | sc-2357/Santa Cruz
Biotechnology, Inc. |
| Secondary
antibody | HRP-conjugated | Goat
anti-rabbit/mouse IgG | 1:5,000 |
sc-2004/sc-2005/Santa Cruz Biotechnology,
Inc. |
Western blot analysis
Isolated CC tissues were treated with a brain tissue
protein extraction kit (cat. no. BB-31227-1; Chengdu beibo;
http://beibokit.com/). Lysates were centrifuged
at 12,000 × g for 30 min at 4°C. Protein concentration was
determined using a BCA protein assay kit (Pierce; Thermo Fisher
Scientific, Inc.). Protein samples (50 µg per lane) were
separated on sodium dodecyl sulfate-polyacrylamide gels (12%).
Next, the proteins were transferred to polyvinylidene fluoride or
polyvinylidene difluoride membranes, blocked [2.5 g skim milk
powder dissolved in 50 ml TBST (0.1% Tween 20)] at room temperature
for 1 h, and incubated overnight at 4°C with primary antibodies
(Table I). The next day, the
membranes were washed and then incubated with secondary antibodies
(Table I) in blocking solution
for 1 h. Finally, the membranes were exposed to obtain signals of
the bound antibody signals. Quantification was performed using
ImageJ software (National Institutes of Health). The relative
expression levels of target proteins were calculated as the target
protein integrated density values (IDVs) relative to actin
IDVs.
Electron microscopy (EM)
On P35, rat brains were obtained and sectioned into
~1 mm3 blocks containing the CC area. The sectioned
tissue was pre-fixed with a mixed solution of 3% glutaraldehyde at
4°C for 48 h, post-fixed in 1% osmium tetroxide, dehydrated in an
acetone series 2 h at room temperature, filtrated in Epox 812, and
embedded with EMBed 812 (cat. no. 90529-77-4; SPI; https://www.2spi.com/category/chemicals/). Next,
semi-thin sections were stained with methylene blue for 8 min at
room temperature, and ultrathin sections were stained with uranyl
acetate and lead citrate for 8 min at room temperature. Finally,
the ultrathin sections were examined using a transmission electron
microscope (H-600IV; Hitachi, Ltd.). Myelinated axons in each field
were quantified using Image-Pro Plus 6.0 software (Media
Cybernetics, Inc.). A total of four randomly selected fields from
the CC were examined in each animal. A total of six animals per
group were analyzed.
Morris water maze (MWM) test
Neurological performance was verified using the MWM
test from P29 to P35. The testing apparatus comprised a circular
tank (1.5 m in diameter), location-constant platform (14 cm in
diameter) placed 1.5 cm under the surface of the water, and an
overhead camera. The water temperature was maintained at 25±1°C
during testing. The test consists of two parts, namely place
navigation training and space exploration, both of which are aimed
to test spatial learning and memory ability. Place navigation
training was conducted during the first 6 days (P29-P34), for which
the rats were trained to swim in the four alternating quadrants.
The rats were allowed to swim in the water from each quadrant for
120 sec. If the platform was successfully found during this period,
the escape latency was recorded as the time taken for rats to find
the platform. Rats that failed to find the platform within 120 sec
were guided to it by a researcher and allowed to stay on the
platform for 30 sec, and the escape latency was recorded as 120
sec. The time in which the rats found the platform in each training
session was recorded. The average latency period for the four
quadrants was computed as a daily final score representing the
ability to acquire spatial information. The platform was removed,
and a space navigation test was conducted on P35 to assess memory
retention ability 24 h after the final place navigation training.
The rats were allowed to swim freely in the tank for 120 sec from
the third quadrant starting point. The trials were recorded using a
video camera on the ceiling, and the platform crossing time was
calculated and analyzed using a tracking system (Mengtai,
China).
Quantification and statistical
analysis
All images were acquired from the same CC area. All
data are presented as mean ± standard deviation (SD). All graphs
were generated using GraphPad Prism 8.0 (GraphPad Software, Inc.).
Comparison between two groups was performed using an unpaired
Student's t-test. Analysis of variance (ANOVA) was used to compare
more than two groups, followed by the Student's t-test if
homogeneity of variance was assumed or by Dunnett's test if
homogeneity of variance was not assumed. A total of six animals
were used for each group, each experiment was conducted for three
times, and totally ~1,335 rats were used in the present study. All
statistical analyses were performed using SPSS 23.0 (IBM Corp.).
P<0.05 was considered to indicate a statistically significant
difference.
Results
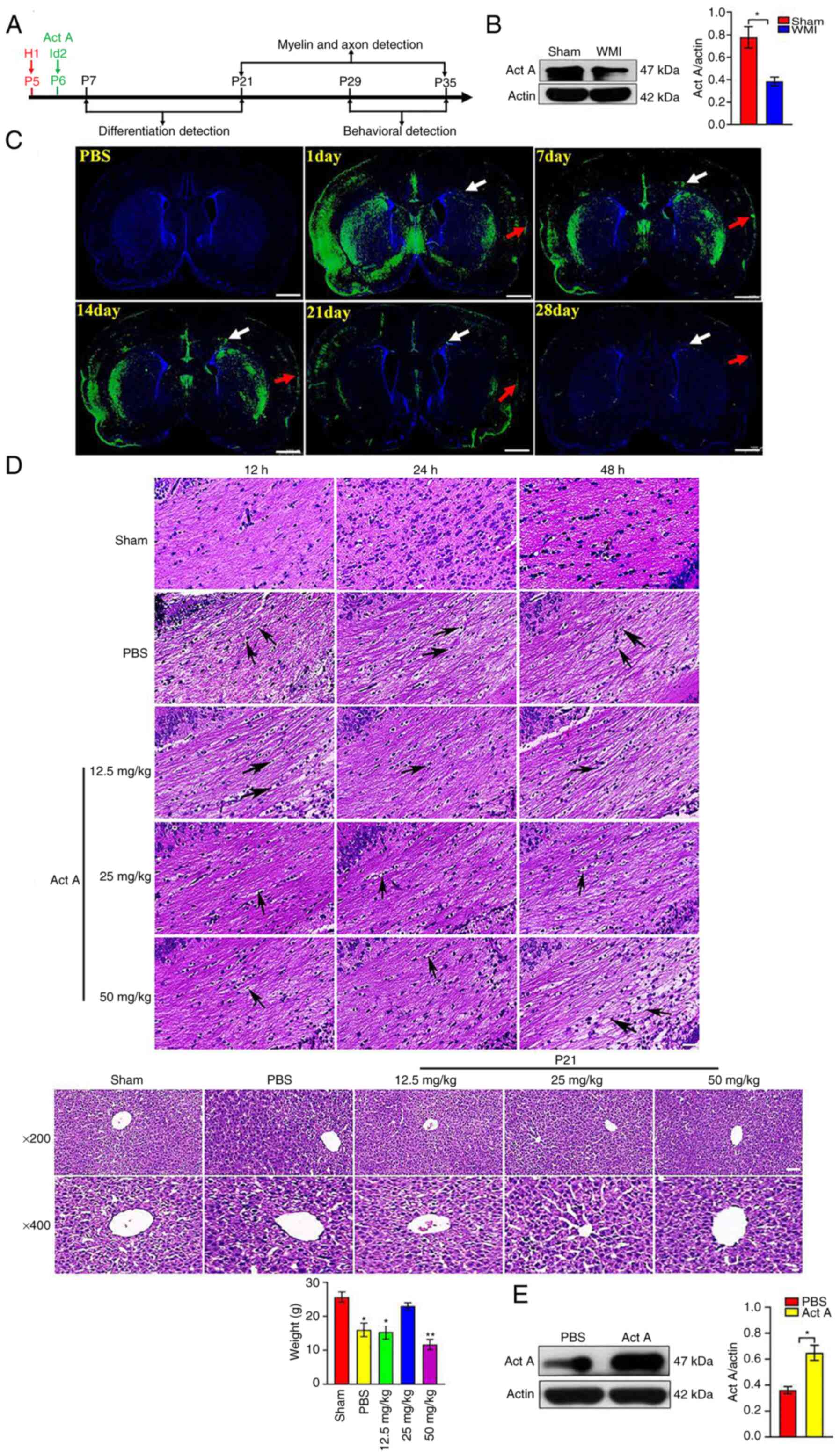
HI attenuates Act A expression in
neonatal rat brains
Based on brain developmental characteristics in
rats, a time course for each assay was set, which is presented as a
schematic diagram in Fig. 1A.
Western blot experiments were first conducted to detect the
endogenous expression of Act A after HI at P21 by using the
isolated CC tissues. Western blotting revealed that Act A
expression was lower in the WMI group than in the Sham group
(Fig. 1B), indicating that HI
reduced Act A expression in neonatal rats.
Act A treatment alleviates pathological
damage after WMI
To detect the distribution of Act A after injection
via the LV, Act A-enhanced green fluorescent protein (EGFP) protein
was constructed and immunofluorescence tracing was performed.
Fluorescence scanning revealed that Act A-EGFP was distributed in
the cortex and white matter (including CC) from days 1 to 28 after
LV injection (Fig. 1C). To select
the optimal usage of Act A for WMI therapy, three dose
concentrations were used (low, 12.5 mg/kg; medium, 25 mg/kg; and
high, 50 mg/kg) and three time points (12, 24 and 48 h) for Act A
administration after HI. Pathological changes in the brain white
matter and liver were examined via H&E staining and body
weights on P14 were analyzed. H&E staining revealed that Act A
treatment decreased the loosely arranged nerve fibers and cell
edema in the CC area, which exhibited with markedly less tissue
vacuolization and nuclear fragmentation and liquefaction 24 h after
HI, suggesting that Act A treatment ameliorated the pathological
characteristics of WMI (Fig. 1D).
Further analysis indicated that the medium and high doses of Act A
treatment showed more effective pathological improvement when
compared with that treated with the low dose, though there was no
significant difference in pathological improvement effects between
the medium and high doses (Fig.
1D). However, hepatic H&E staining on P21 and body weight
analysis revealed that the high dose of Act A treatment led to less
hepatic lobule and had lower body weight and poor state in rats
(Fig. 1D). Therefore, subsequent
experiments were performed with a medium dose of Act A (25 mg/kg).
To detect the overall expression levels of Act A in the brain after
exogenous Act A injection, western blotting was conducted on P6.
Act A was abundantly expressed in the Act A group than in the PBS
group (Fig. 1E). Collectively,
these results suggested that exogenous Act A supplementation
alleviated pathological damage in WMI.
Act A treatment promotes OPC
differentiation after WMI
Next, it was examined whether Act A contributes to
the differentiation of OPCs to OLs. On P7 and P14, the number of
OPCs in white matter was quantified via double immunostaining with
mature-oligodendrocyte marker oligodendrocyte transcription factor
(Olig2) and OPC-specific marker neural/glial antigen 2 (NG2) in the
experimental groups. The number of NG2/Olig2-positive cells and
mean NG2/Olig2 fluorescence intensity at both time points were
significantly higher in the Act A group than in the PBS group
(Fig. 2A). On P14 and P21, the
number of pre-OLs was quantified in the CC by double immunostaining
with Olig2 and the pre-OL-specific marker O4 in the experimental
groups. The number of O4/Olig2-positive cells and mean O4/Olig2
fluorescence intensity at both time points were significantly
higher in the Act A group than in the PBS group (Fig. 2B). Further, on P21 and P28, double
immunofluorescence staining was performed with the mature OL marker
CC1 with Olig2 in the CC. The number of CC1/Olig2-positive cells
and mean CC1/Olig2 fluorescence intensity were significantly higher
in the Act A group than in the PBS group (Fig. 2C). Collectively, these results
indicated that Act A promoted OPC differentiation.
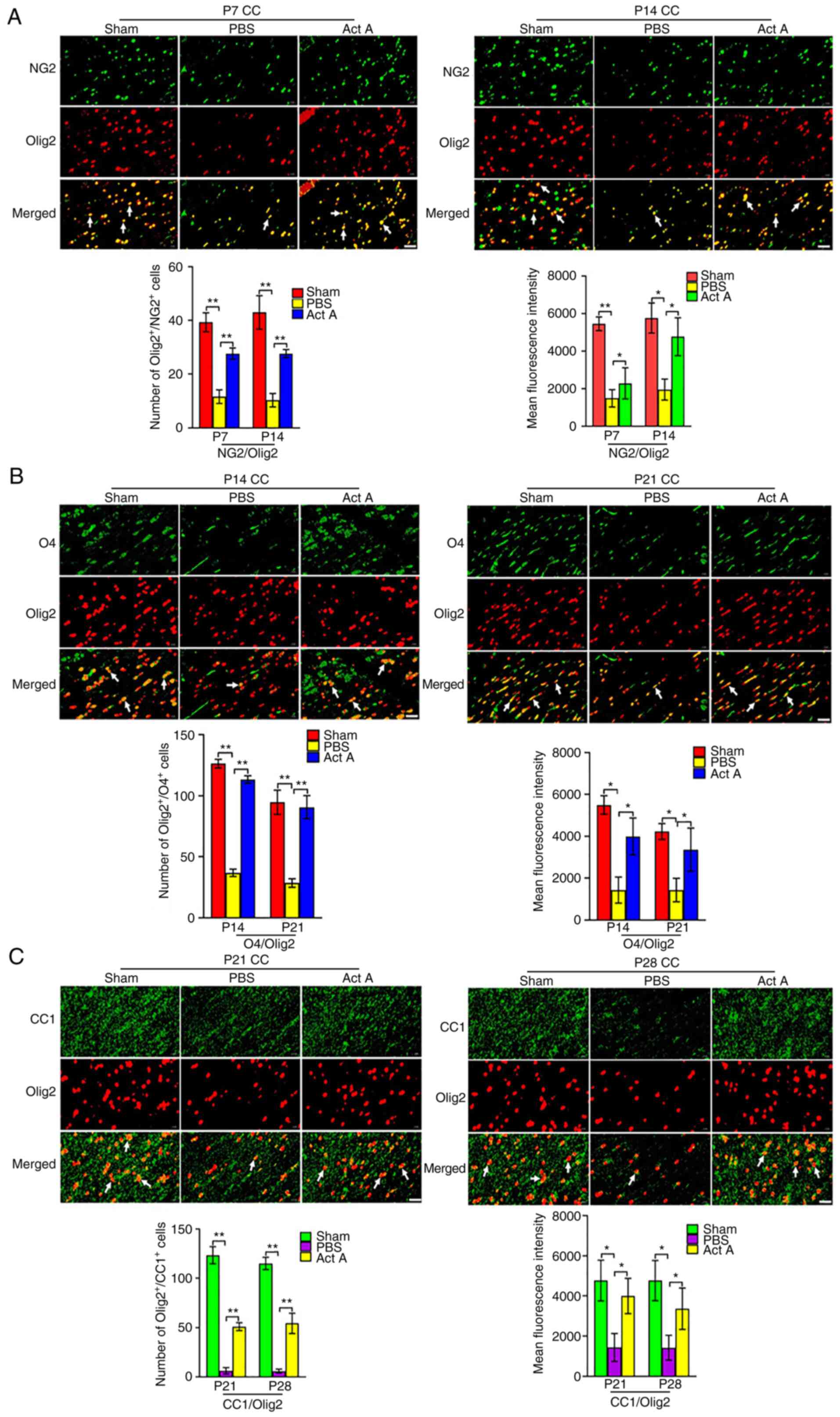 | Figure 2Act A treatment promotes OPC
differentiation in WMI. (A) Representative immunofluorescence
images and quantification of NG2 (green) expression via double
staining with Olig2 (red) at P7 and P14. Arrows indicate the
NG2/Olig2 (yellow) positive cells. NG2, a OPCs marker. Scale bar,
20 µm. (B) Representative immunofluorescence images and
quantification of O4 (green) expression via double staining with
Olig2 (red) at P14 and P21. Arrows indicate the O4/Olig2 (yellow)
positive cells. O4, a marker of pre-oligodendrocytes. Scale bar, 20
µm. (C) Representative immunofluorescence images and
quantification of CC1 (green) expression via double staining with
Olig2 (red) at P21 and P28. Arrows indicate the CC1/Olig2 (yellow)
positive cells. CC1, Anti-APC (Activated protein C), a marker of
mature oligodendrocytes. Scale bar, 20 µm.
*P<0.05 and **P<0.001. OPC,
oligodendrocyte progenitor cell; WMI, white matter injury; Olig2,
oligodendrocyte transcription factor; Act A, Activin A; NG2,
neural/glial antigen 2; CC, corpus callosum. |
Act A treatment promotes myelination and
axon formation after WMI
CC myelination was then examined by assessing the
expression of myelin basic protein (MBP), proteolipid protein
(PLP), and myelin-associated glycoprotein (MAG) using
immunofluorescence and western blot analysis. On P28, the
expression of MBP, PLP, and MAG was significantly higher in the Act
A group than in the PBS group (Fig.
3A and B). Consistent with this, on P35, expression of the axon
markers Tau1, SMI31, and SMI312 was significantly higher in the Act
A group than in the PBS group (Fig.
4A and B). On P35, EM revealed more myelinated axons in the CC
of brains in the Act A group than in the CC of brains in the PBS
group (Fig. 4C). Collectively,
these results indicated that exogenous Act A supplementation after
WMI contributed to myelination and axon formation.
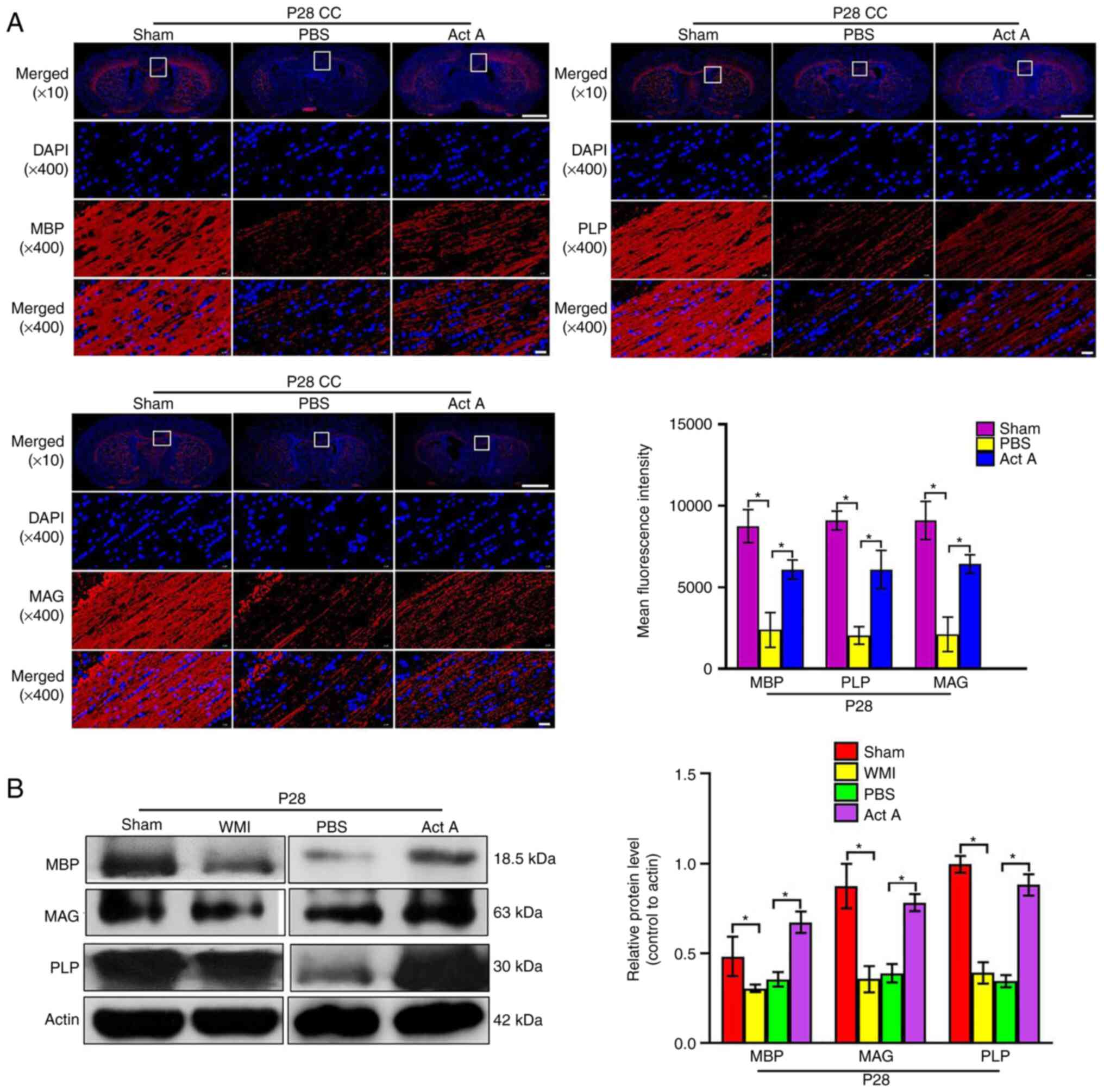 | Figure 3Act A treatment promotes the
myelination in WMI. (A) Representative immunofluorescence images
and quantification of the expression of the myelin sheath markers
MBP, PLP and MAG (red) at P28. Cell nuclei were labeled with DAPI
(blue). The mean fluorescence intensity of MBP, PLP, and MAG was
quantified. Scale bar, 1,000 or 20 µm. (B) Western blot
analysis and corresponding quantification were conducted to measure
the expression of MBP, PLP and MAG at P28. It showed that the
expression of MBP, PLP and MAG was significantly enhanced in the
Act A group compared with the PBS group. *P<0.05. Act
A, Activin A; WMI, white matter injury; MBP, myelin basic protein;
PLP, proteolipid protein; MAG, myelin-associated glycoprotein. |
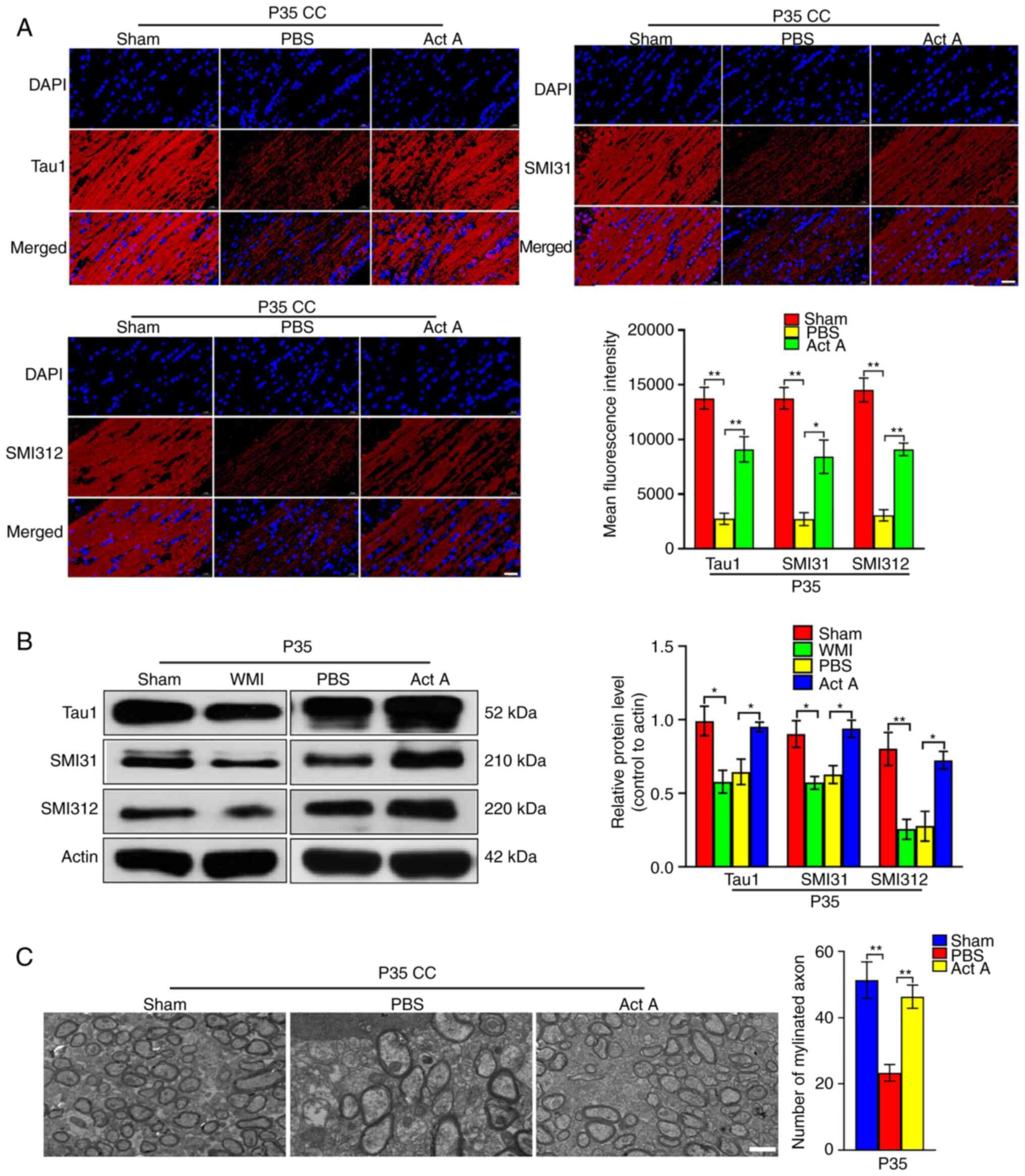 | Figure 4Act A treatment enhances the axon
formation in white matter injury. (A) Representative
immunofluorescence images and quantification of axons markers Tau1,
SMI31, and SMI312 (red) at P35. The mean fluorescence intensity of
Tau1, SMI31 and SMI312 was quantified. Scale bar, 20 µm. (B)
Western blot analysis and corresponding quantification were
performed to measure the expression of Tau1, SMI31, and SMI312 at
P35. It showed that the expression of Tau1, SMI31 and SMI312 was
significantly increased in the Act A group compared with the PBS
group. (C) Representative EM images in the CC at P35. The number of
myelinated axons was counted per field using the Image-Pro Plus 6.0
software. It showed that the Act A group with more myelinated axons
compared with the PBS group. Scale bar, 2 µm.
*P<0.05 and **P<0.001. Act A, Activin
A; WMI, white matter injury; CC, corpus callosum. |
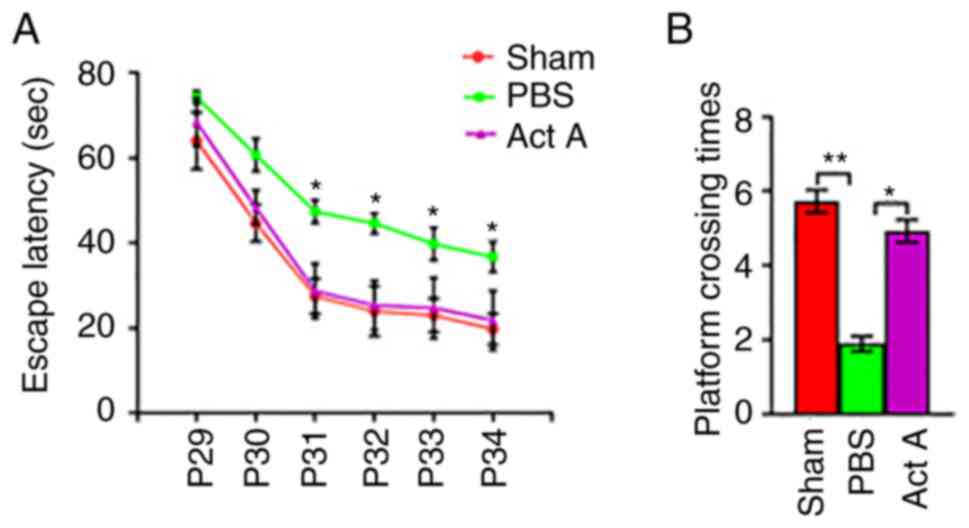
Act A treatment improves neurological
performance after WMI
The MWM test was conducted to compare learning and
memory abilities among the experimental groups from P29 to P35 by
calculating average escape latency and platform crossing times. The
average escape latency from P31 was significantly lower in the Act
A group than in the PBS group (Fig.
5A). The frequency of platform crossings at P35 was
significantly higher in the Act A group than in the PBS group,
whereas no significant difference was observed between the Act A
and Sham groups (Fig. 5B). These
results suggested that exogenous Act A treatment contributed to
improvements in learning and memory.
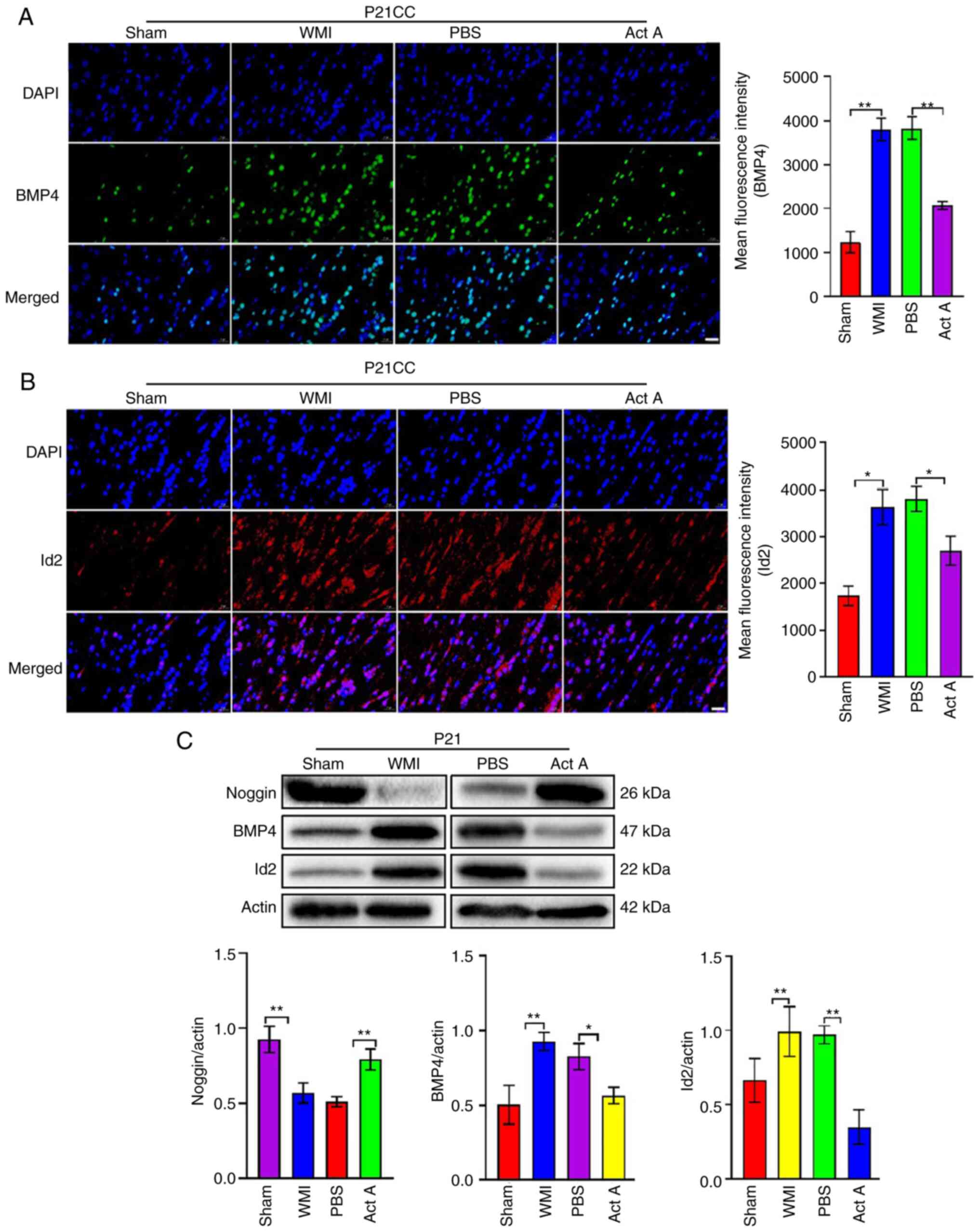
Act A treatment increases noggin
expression and inhibits BMP4/Id2 expression after WMI
Immunofluorescence was performed to detect the
expression of bone morphogenetic protein 4 (BMP4) and inhibitor of
DNA binding 2 (Id2) in Sham, PBS and Act A groups. Expression of
these proteins was significantly higher in the PBS group than in
the Sham group (Fig. 6A and B)
but was significantly decreased after Act A treatment (Fig. 6A and B). Western blot analysis
revealed that noggin expression was significantly higher in the Act
A group than in the PBS group, whereas both BMP4/Id2 proteins were
significantly lower in the Act A group than in the PBS group
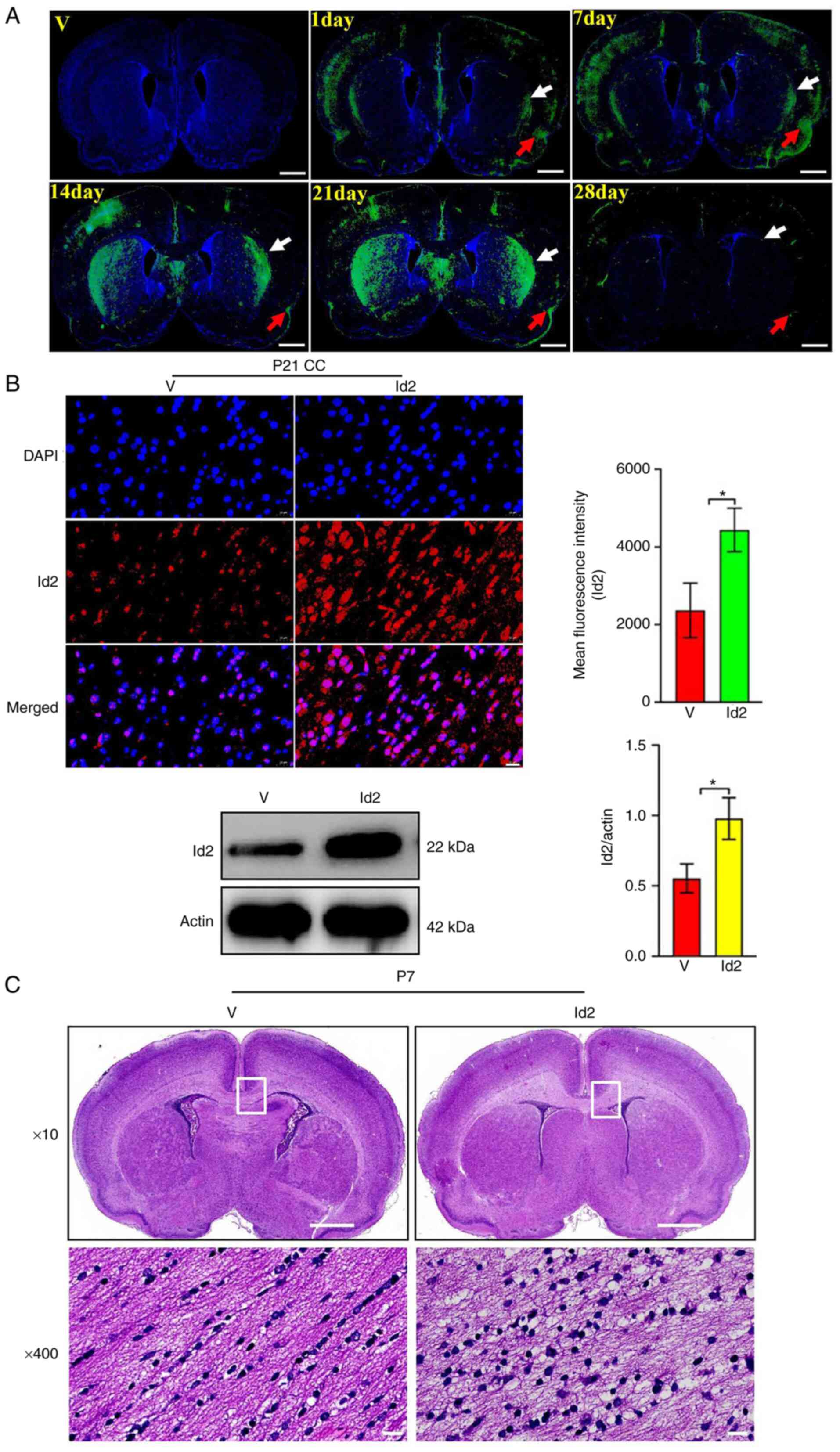
(Fig. 6C). Next, Id2 was
upregulated in the Act A group using an Id2-overexpressing
lentiviral vector (1×109 TU/ml). Fluorescence imaging
revealed that Id2-EGFP was distributed in the cortex and white
matter (including CC) for up to 4 weeks. On the first day
post-injection, Id2-EGFP was observed in the CC (white arrow) and
cortex (red arrow). From the 7th to 21st day post-injection,
Id2-EGFP fluorescence intensity in the CC and cortex was
significantly increased, whereas on the 28th day post-injection,
Id2-EGFP fluorescence intensity in the CC and cortex was
significantly decreased (Fig.
7A). In addition, immunofluorescence and western blot
experiments were performed to detect Id2 expression in the V and
Id2 groups. The analysis revealed that Id2 expression was
significantly higher in the Id2 group than in the V group (Fig. 7B). H&E staining revealed more
white matter vacuolization and nuclear fragmentation in the Id2
group than in the V group (Fig.
7C). Collectively, these results suggested that the beneficial
effects of Act A on WMI involved the Noggin/BMP4/Id2 signaling
pathway.
Id2 overexpression attenuates the
therapeutic effects of Act A on WMI via the Noggin/BMP4/Id2
signaling pathway
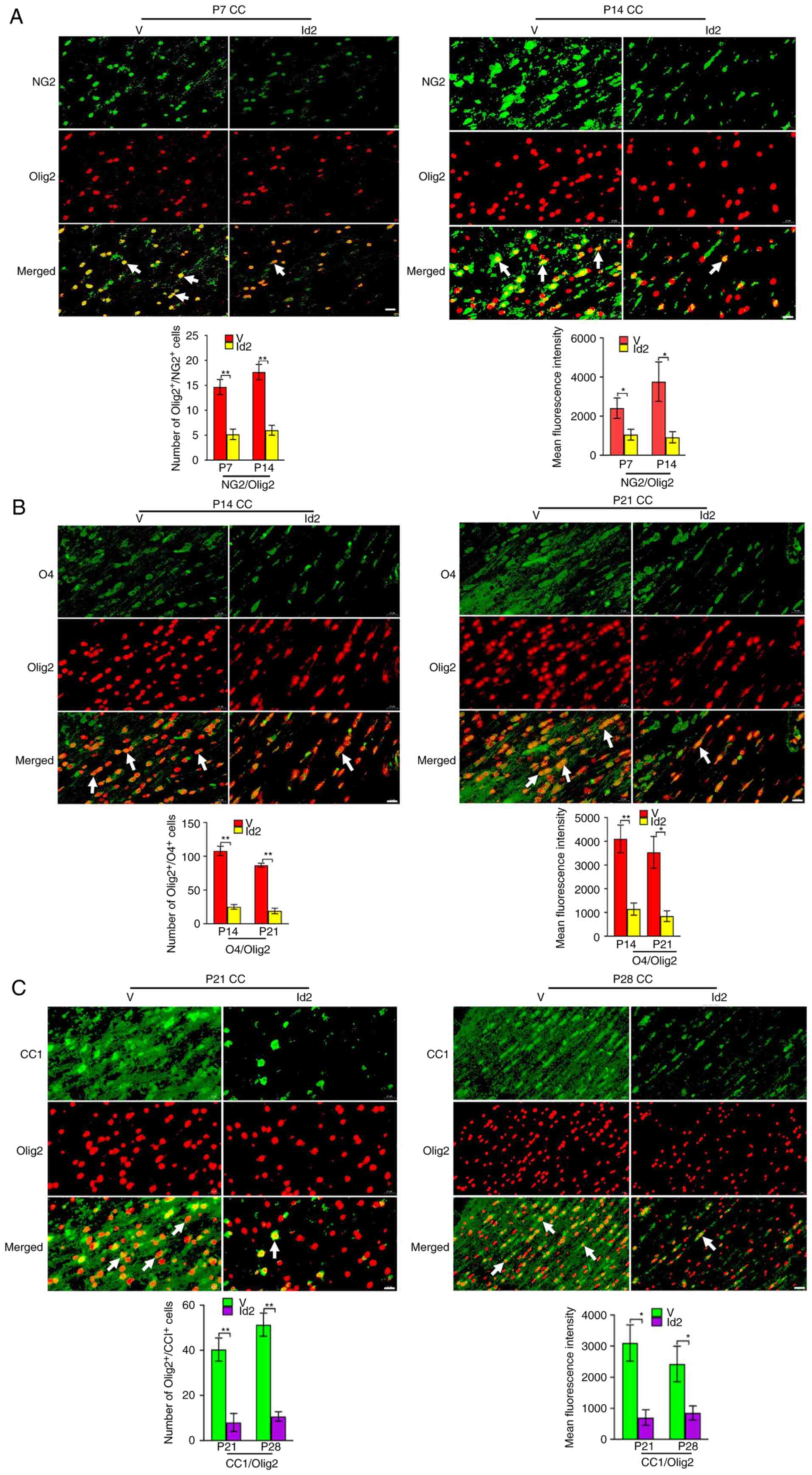
The number of immunopositive cells and mean
fluorescence intensity for NG2/Olig2 (Fig. 8A), O4/Olig2 (Fig. 8B), and CC1/Olig2 (Fig. 8C) were significantly lower in the
Id2 group than in the V group. Furthermore, Id2 upregulation
attenuated the expression of MBP, PLP, MAG (Fig. 9A), Tau1, SMI31 and SMI312
(Fig. 9B) after Act A treatment,
as indicated by immunofluorescence. Similarly, on P35, EM revealed
fewer myelinated axons in the Id2 group than in the V group
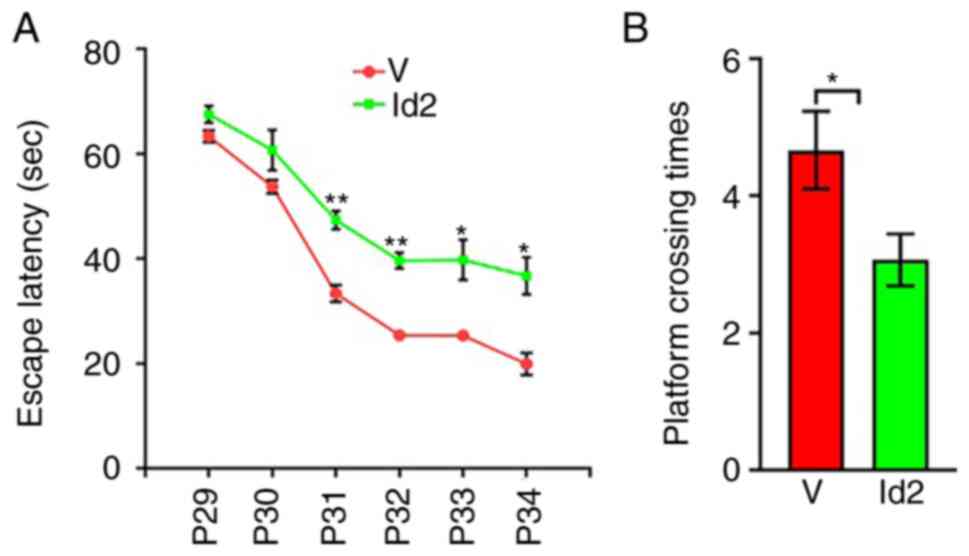
(Fig. 9C). The MWM test revealed
that the average escape latency from P31 was significantly higher
in the Id2 group than in the V group (Fig. 10A), and the frequency of platform
crossings at P35 was significantly lower in the Id2 group than in
the V group (Fig. 10B).
Performance was similar between the Id2 and PBS groups, suggesting
that Id2 upregulation rescued behavioral dysfunction. Collectively,
these results verified that Act A exerted therapeutic effects on
WMI via the Noggin/BMP4/Id2 signaling pathway.
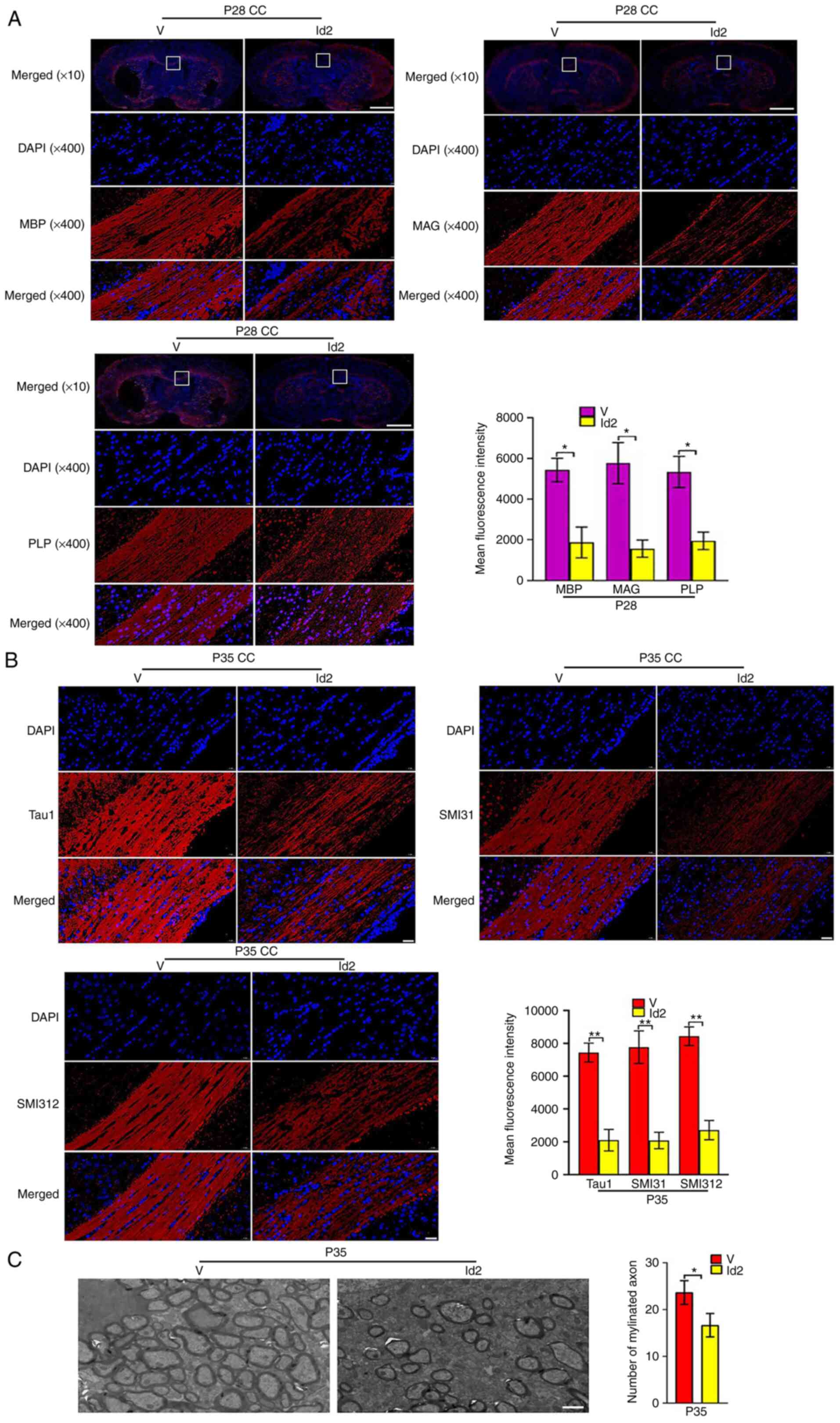 | Figure 9Upregulation of Id2 attenuates the
roles of Activin A on myelination and axon formation. (A)
Representative immunofluorescence staining images of the expression
of myelin sheath markers MBP, PLP and MAG (red) at P28. Scale bar,
1,000 or 20 µm. (B) Representative immunofluorescence
staining images of the expression of axon markers Tau1, SMI31 and
SMI312 (red) at P35. Scale bar, 20 µm. (C) Representative
electron microscopy images at P35. It showed that the Id2 group
with less myelinated axons compared with the V group. Scale bar, 2
µm. *P<0.05 and **P<0.001. Id2,
inhibitor of DNA binding 2; MBP, myelin basic protein; PLP,
proteolipid protein; MAG, myelin-associated glycoprotein. |
Discussion
OPCs constitute the major cell population that is
injured in WMI. As such, protecting OPCs constitutes a key strategy
for WMI treatment (12). Previous
studies have reported the involvement of Act A in the regulation of
OPC maturation in vitro, which led us to examine its role in
WMI in vivo (13,14). In an adult rat model of focal
cerebral ischemia simulating stroke, expression of Act A around the
infarction was higher than that in control rats (15). However, in the current study, it
was observed that the expression of endogenous Act A was
significantly lower after WMI in newborn rats. This discrepancy may
be due to the differences in the age of rats and injury models
used. Further, the reduction in Act A expression suggests the
involvement of Act A in WMI pathophysiology. Act A is secreted by
both neurons and glial cells, which exert neuroprotective effects.
In the present study, it was attempted to partially replenish Act A
dosage via exogenous administration to compensate for the
WMI-induced decrease in Act A levels. Given the presence of the
blood-brain barrier (BBB), Act A was injected via the LV in a rat
model of WMI. Act A-EGFP tracing experiments revealed that Act
A-EGFP protein was distributed in the cerebral cortex and white
matter (including the CC) from days 1 to 28 after injection. This
indicated that Act A could enter the brain and persist for up to 4
weeks after intra-LV injections, supporting its efficacy after a
single administration. To explore the therapeutic time window of
Act A, injections were performed before and after modeling, as well
as single or continuous multiple injections in a preliminary study.
It was observed that the effects of injection before modeling were
improved compared with those after modeling. Nevertheless, no
differences were observed in the effects of single vs. multiple
injections. Given that injections before modeling do not fully
recapitulate clinical settings, injections were performed after
model establishment. Next, three different concentrations of Act A
were analyzed to determine the optimal dosage. H&E staining for
the CC showed that three doses of Act A improved the injury when
compared with the PBS-treated groups. Further H&E staining for
hepatic tissues and body weight analysis demonstrated that the
highest dose (50 mg/kg) led to less hepatic lobule and with poor
body weight improvement. Furthermore, the body wight in the lowest
dosage (12.5 mg/kg) decrease obviously than that in the medium dose
(25 mg/kg). Collectively, after careful consideration, it was
decided to use the medium dose of Act A to treat the WMI rats. In
this regard, exogenous administration of a certain dose of Act A
may have rescued vulnerable cell populations, such as OPCs, which,
in turn, increased myelin sheath and neural network formation,
improved the white matter microenvironment in WMI, and ameliorated
WMI-induced diffuse damage. Although LV injections are a useful
experimental approach, the invasive nature of this operation limits
its clinical applicability. Recently, several non-invasive methods
to deliver drugs to the brain and overcome the BBB have emerged,
predominantly employing material-based deliveries. For example,
Wang et al (16)
effectively delivered glial cell-derived neurotrophic factor to the
brain of rats via conjugated-biotinylated lipid-coated
microbubbles. Other feasible pathways to deliver Act A into the
brain shall be investigated in future studies by the authors.
Axons in the vertebrate central nervous system (CNS)
are generally ensheathed by myelin, a tight spiral wrapping of
plasma membrane generated by OLs (13). Myelin-wrapped axons are the major
mediators of signal transduction in the CNS, and their formation is
fundamental for brain development and function (13). In accordance with the
developmental characteristics of the rat brain, the formation of
myelin sheath wrapping axons involves several successive stages,
starting from OPC differentiation (17,18). In the present study, different
time periods were set to detect the progressive differentiation and
maturation of OLs: P7 and P14 to detect OPCs, P14 and P21 to detect
pre-OLs, P21 and P28 to detect OLs, P28 to detect myelin formation,
and P35 to detect myelin-wrapped axons. This experimental design
allowed the authors to obtain an overall view of the effects of Act
A on WMI progression. The results of the present study indicated
that Act A treatment promoted myelination and axon formation after
WMI. It was concluded that this was owing to the alleviation of the
OPC differentiation barrier in WMI by Act A treatment, which, in
turn, increased the formation of mature OLs to support the
formation of myelin and myelinated axons. Further, the MWM test was
used to detect behavioral performance reflecting the long-time
effects of Act A after WMI. The MWM aims to assess learning and
memory ability by analyzing the average escape latency and
frequency of platform crossings. The average escape latency was
significantly lower whereas the frequency of platform crossings was
significantly higher in the Act A group than in the PBS group.
These results indicated that Act A treatment improved learning and
memory ability in WMI rats, illustrating that Act A treatment
enhanced long-time behavioral performance after WMI. The persistent
positive effects of Act A after WMI may involve a cascade of
events, as follows. First, Act A alleviated the differentiation
barrier of OPCs in WMI, which, in turn, increased the formation of
mature OLs. After reaching a sufficient number, OLs contributed to
myelination and formation of myelinated axons, thereby alleviating
pathological damage caused by WMI-induced OPC damage. This
ultimately resulted in signal transduction in myelinated axons
returning to a normal state reflecting behavioral performance.
A previous study revealed that Act A exerted its
neuroprotection roles mainly through the Smad-dependent pathways
(19). By contrast, a previous
study stated that Act A exerted its effects through
Smad-independent pathways, such as nuclear factor-κB, extracellular
signal-regulated kinase, ubiquitin-proteolytic, mitogen-activated
protein kinase, AKT, and TGF signaling pathways (20). The present study revealed that the
reparative effects of exogenous Act A after WMI in newborn rats was
predominantly achieved by promoting the differentiation and
maturation of OLs. Previous RNA-sequencing experiments on WMI,
which indicated that Id2 was a negative regulator of OL maturation,
provided clues to explore potential pathways for the roles of Act A
in WMI (SU et al, unpublished data). Moreover, studies have
reported that Id2 participates in different stages of OL
differentiation and is a key regulator of OL differentiation and
maturation (21,22). Id2 inhibits the expression of
myelin formation genes and maintains OPCs in an undifferentiated
state, thereby inhibiting OPC differentiation and production of
mature OLs (23). Based on this
evidence, the KEGG pathway website was searched to elucidate the
relationship between Act A and Id2. It was identified that Act A
regulated cell differentiation and neurogenesis via the
Noggin/BMP/Id signaling pathway. Indeed, several studies have
reported that Act A regulates cell differentiation by interacting
with BMP4 (24-27), and BMP4, in turn, regulates the
differentiation and maturation of OLs by regulating its downstream
target molecule Id2. BMP4/Id2 signaling hinders the differentiation
of OPCs into OLs (28). Noggin is
a key upstream molecule regulated by BMP4. Increased noggin
expression inhibits BMP4 expression, whereas Act A enhances noggin
expression (29). Based on these
data, it was hypothesized that the effects of Act A on WMI may be
achieved via Noggin/BMP4/Id2 signaling.
To test the aforementioned hypothesis, the
expression of Noggin/BMP4/Id2 after HI or Act A treatment was
analyzed and it was observed that noggin expression was inhibited,
whereas BMP4 and Id2 expression was increased after HI. After Act A
treatment, noggin expression was significantly upregulated, whereas
BMP4 and Id2 expression was significantly downregulated. Moreover,
Id2 upregulation blocked the rescuing effects of Act A after WMI.
Collectively, these results suggested that Act A rescues WMI via
the Noggin/BMP4/Id2 signaling pathway. Mechanistically, it was
hypothesized that Act A enters the intercellular space through
diffusion after injection into the LV and binds to Act A receptors
on the surface of OPCs. Subsequently, Act A activates noggin
expression and inhibits BMP4 and Id2 expression. This relieves the
negative regulatory factors that modulate OPC differentiation,
promotes myelin sheath formation, reduces pathological white matter
damage in the brain, and rescues neurobehavioral defects in rats.
However, the effects of blocking or overexpressing noggin and BMP4
have not been verified. Hence, this remains speculative, and more
research is warranted to verify the causal relationship between Act
A-mediated regulation of Id2 and Noggin/BMP4 signaling.
In summary, the present study demonstrated that
exogenous Act A treatment rescued WMI via the Noggin/BMP4/Id2
signaling pathway. Although Act A has been used as a diagnostic and
prognostic biomarker for several brain diseases (30), it has not been used to treat brain
damage in clinical practice. The present findings demonstrated for
the first time, to the best of our knowledge, that exogenous Act A
treatment may alleviate WMI in the neonatal rat brain, highlighting
the potential of using Act A as a therapeutic agent to treat
neonatal WMI. Besides, the encephalopathy of prematurity conceived
by Volpe (31) indicated that it
is a complex amalgam of primary destructive disease and secondary
maturational and trophic disturbances. The aforementioned study
claimed that the neuropathology of brain injury in the premature
infant occurs against a background of multiple active developmental
events that take place at 24-40 weeks of gestation and involve
pre-OLs, microglia, axons, subplate neurons, the proliferative
cerebral dorsal subventricular zone and ventral germinative
epithelium of the ganglionic eminence, thalamus, cortex and
cerebellum (31). In the present
study, the treatment roles and mechanisms of Act A against the
background of human fetuses aged 23-32 weeks that within the
duration of the preterm infants described in Volpe (31) were explored; the main
characteristics of the aforementioned fetuses are the obstacle of
OPCs differentiation, which finally leads to the behavioral and
cognitive dysfunction. Thus, the pathological characteristics,
oligodendrocyte lineage cells, the myelination, axon formation and
the behavioral and cognitive ability were established as the
primary study index. The present findings indicated that Act A
ameliorated the pathological damages, promoted the differentiation
of OPCs, improved the myelination and axon formation, and finally
rescued the learning and memory abilities, of which hinted that Act
A not only influence the primary destructive disease but also the
secondary developmental process. Collectively, the current study
concluded that Act A shows rescue effects in premature
encephalopathy of 23-32 weeks. Regarding the effective roles of Act
A in the primary destructive disease and the secondary maturational
process, it is hypothesized that Act A may also play roles during
the encephalopathy of prematurity of 24-40 weeks. Besides, it is
considered that the types of pathological changes, the involved
tissues and severity of neuro injury are more complex between the
large extents of 24-40 weeks, which needs further validation.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
XS and JY contributed to the conception and design
of the research and drafting of the present study. DX and XQ
performed the MWM test. SL and FZ participated in data acquisition.
JT made substantial contributions to the conception, design,
acquisition, analysis and interpretation of data and revised the
manuscript. XS and JT confirm the authenticity of all the raw data.
All authors read and approved the final version of the
manuscript.
Ethics approval and consent to
participate
All animal experiments were approved (approval no.
WCSUH21-2018-034) by the Sichuan University Committee on Animal
Research (Chengdu, China) and complied with the ARRIVE
guidelines.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Funding
The present study was supported by the National Key R&D
Program of China (grant nos. 2021YFC2701704 and 2017YFA0104200),
the National Natural Science Foundation of China (grant nos.
81971433 and 81971428), the Science and Technology Bureau of
Sichuan Province (grant nos. 2021YJ0017 and 2020YFS0041), the
Fundamental Research Funds for Central University (grant no.
SCU2021D009) and the National Key Project of Neonatal Children
(grant no. 1311200003303).
References
|
1
|
Alexandrou G, Mårtensson G, Skiöld B,
Blennow M, Adén U and Vollmer B: White matter microstructure is
influenced by extremely preterm birth and neonatal respiratory
factors. Acta Paediatr. 103:48–56. 2014. View Article : Google Scholar
|
|
2
|
Liu XB, Shen Y, Plane JM and Deng W:
Vulnerability of premyelinating oligodendrocytes to white-matter
damage in neonatal brain injury. Neurosci Bull. 29:229–238. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gano D: White matter injury in premature
newborns. Neonatal Netw. 35:73–77. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Suzuki N, Sekimoto K, Hayashi C, Mabuchi
Y, Nakamura T and Akazawa C: Differentiation of oligodendrocyte
precursor cells from Sox10-venus mice to oligodendrocytes and
astrocytes. Sci Rep. 7:141332017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Anand C, Brandmaier AM, Arshad M, Lynn J,
Stanley JA and Raz N: White-matter microstructural properties of
the corpus callosum: Test-retest and repositioning effects in two
parcellation schemes. Brain Struct Funct. 224:3373–3385. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Goebbels S, Wieser GL, Pieper A, Spitzer
S, Weege B, Yan K, Edgar JM, Yagensky O, Wichert SP, Agarwal A, et
al: A neuronal PI(3,4,5)P3-dependent program of
oligodendrocyte precursor recruitment and myelination. Nat
Neurosci. 20:10–15. 2017. View
Article : Google Scholar
|
|
7
|
Wang X, Fischer G and Hyvönen M: Structure
and activation of pro-activin A. Nat Commun. 7:120522016.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zheng J, Zhang T, Han S, Liu C, Liu M, Li
S and Li J: Activin A improves the neurological outcome after
ischemic stroke in mice by promoting oligodendroglial
ACVR1B-mediated white matter remyelination. Exp Neurol.
337:1135742021. View Article : Google Scholar
|
|
9
|
Clancy B, Darlington RB and Finlay BL:
Translating developmental time across mammalian species.
Neuroscience. 105:7–17. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Huang L, Zhao F, Qu Y, Zhang L, Wang Y and
Mu D: Animal models of hypoxic-ischemic encephalopathy: Optimal
choices for the best outcomes. Rev Neurosci. 28:31–43. 2017.
View Article : Google Scholar
|
|
11
|
Huang Z, Liu J, Cheung PY and Chen C:
Long-term cognitive impairment and myelination deficiency in a rat
model of perinatal hypoxic-ischemic brain injury. Brain Res.
1301:100–109. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang F, Yang YJ, Yang N, Chen XJ, Huang
NX, Zhang J, Wu Y, Liu Z, Gao X, Li T, et al: Enhancing
oligodendrocyte myelination rescues synaptic loss and improves
functional recovery after chronic hypoxia. Neuron. 99:689–701.e5.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Snaidero N, Möbius W, Czopka T, Hekking
LH, Mathisen C, Verkleij D, Goebbels S, Edgar J, Merkler D, Lyons
DA, et al: Myelin membrane wrapping of CNS axons by
PI(3,4,5)P3-dependent polarized growth at the inner tongue. Cell.
156:277–290. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
De Berdt P: Bottemanne P, Bianco J,
Alhouayek M, Diogenes A, Lloyd A, Llyod A, Gerardo-Nava J, Brook
GA, Miron V, et al Stem cells from human apical papilla decrease
neuro-inflammation and stimulate oligodendrocyte progenitor
differentiation via activin-A secretion. Cell Mol Life Sci.
75:2843–2856. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nishio S, Yunoki M, Chen ZF, Anzivino MJ
and Lee KS: Ischemic tolerance in the rat neocortex following
hypothermic preconditioning. J Neurosurg. 93:845–851. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang F, Shi Y, Lu L, Liu L, Cai Y, Zheng
H, Liu X, Yan F, Zou C, Sun C, et al: Targeted delivery of GDNF
through the blood-brain barrier by MRI-guided focused ultrasound.
PLoS One. 7:e529252012. View Article : Google Scholar
|
|
17
|
Simons M and Nave KA: Oligodendrocytes:
Myelination and axonal support. Cold Spring Harb Perspect Biol.
8:a0204792015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Back SA: White matter injury in the
preterm infant: Pathology and mechanisms. Acta Neuropathol.
134:331–349. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang Y, Zhang J, Navrazhina K, Argaw AT,
Zameer A, Gurfein BT, Brosnan CF and John GR: TGFbeta1 induces
Jagged1 expression in astrocytes via ALK5 and Smad3 and regulates
the balance between oligodendrocyte progenitor proliferation and
differentiation. Glia. 58:964–974. 2010.PubMed/NCBI
|
|
20
|
Derynck R and Zhang YE: Smad-dependent and
Smad-independent pathways in TGF-beta family signalling. Nature.
425:577–584. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen XS, Zhang YH, Cai QY and Yao ZX: ID2:
A negative transcription factor regulating oligodendroglia
differentiation. J Neurosci Res. 90:925–932. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gou X, Tang Y, Qu Y, Xiao D, Ying J and Mu
D: Could the inhibitor of DNA binding 2 and 4 play a role in white
matter injury? Rev Neurosci. 30:625–638. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Samanta J and Kessler JA: Interactions
between ID and OLIG proteins mediate the inhibitory effects of BMP4
on oligodendroglial differentiation. Development. 131:4131–4142.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kim MS, Horst A, Blinka S, Stamm K, Mahnke
D, Schuman J, Gundry R, Tomita-Mitchell A and Lough J: Activin-A
and Bmp4 levels modulate cell type specification during
CHIR-induced cardiomyogenesis. PLoS One. 10:e01186702015.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Olsen OE, Wader KF, Hella H, Mylin AK,
Turesson I, Nesthus I, Waage A, Sundan A and Holien T: Activin A
inhibits BMP-signaling by binding ACVR2A and ACVR2B. Cell Commun
Signal. 13:272015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yang S, Yuan Q, Niu M, Hou J, Zhu Z, Sun
M, Li Z and He Z: BMP4 promotes mouse iPS cell differentiation to
male germ cells via Smad1/5, Gata4, Id1 and Id2. Reproduction.
153:211–220. 2017. View Article : Google Scholar
|
|
27
|
Lepletier A, Hun ML, Hammett MV, Wong K,
Naeem H, Hedger M, Loveland K and Chidgey AP: Interplay between
follistatin, activin A, and BMP4 signaling regulates postnatal
thymic epithelial progenitor cell differentiation during aging.
Cell Rep. 27:3887–3901.e4. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Miyazono K and Miyazawa K: Id: A target of
BMP signaling. Sci STKE. 2002:pe402002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Koyano S, Fukui A, Uchida S, Yamada K,
Asashima M and Sakuragawa N: Synthesis and release of activin and
noggin by cultured human amniotic epithelial cells. Dev Growth
Differ. 44:103–112. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bergestuen DS, Edvardsen T, Aakhus S,
Ueland T, Oie E, Vatn M, Aukrust P and Thiis-Evensen E: Activin A
in carcinoid heart disease: A possible role in diagnosis and
pathogenesis. Neuroendocrinology. 92:168–177. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Volpe JJ: Brain injury in premature
infants: A complex amalgam of destructive and developmental
disturbances. Lancet Neurol. 8:110–124. 2009. View Article : Google Scholar :
|