Introduction
The treatment of gliomas remains a challenge for
neurosurgeons (1). There remains
a need to explore the molecular mechanisms of malignant gliomas and
to find new therapeutic strategies and drug targets. Metastasis and
invasion are important causes of cancer mortality. Numerous studies
have shown that voltage-gated sodium channels (VGSCs) are
aberrantly expressed in a variety of metastatic tumors (2-7).
High expression of VGSCs has the potential to enhance cancer cell
invasion and metastasis. It was found that several types of ion
channels are distributed on the nervous system and glial cells, and
certain sodium channels were found to play a role in the
physiological progression of glial cells (8,9).
Previous studies have shown that neonatal isoform Nav1.5 is highly
expressed in high-grade gliomas but is also intimately linked to
their biological behavior, including invasion, proliferation and
migration (10-12). This suggests that VGSCs are
associated with the development of gliomas.
VGSCs are directly or indirectly involved in various
physiological processes by regulating the transport of sodium ions
across membranes (13). VGSCs
consist of a core α subunit and auxiliary β subunits (14). A total of 10 different isoforms
have been identified depending on the α subunit. The distribution
and function of the Navs varies among the different isoforms
(15). The α-subunit is the main
determinant of the physiological activity of VGSC and can function
partially on its own, but is only fully functional when linked to
the auxiliary subunit β-subunit by disulfide or non-covalent bonds.
The sodium channel is widely expressed in the nervous system, but
the encoded proteins, structures and electrophysiological
properties vary between isoforms (16). The Nav1.6 gene encoded by sodium
voltage-gated channel alpha subunit 8 (SCN8A) is widely expressed
in the central and peripheral nervous systems. The expression and
role of Nav1.6 have been verified in various mouse neurological
diseases and animal models (17-20), and it has been reported that
Nav1.6 is highly expressed in human colorectal and oral squamous
carcinomas, and its knockdown significantly affects the invasive
and metastatic ability of cancer cells (7). However, the specific mechanisms
affecting tumor invasion and metastasis remain unclear (21,22).
Currently, the expression of Nav1.6 in glioma and
its effects and mechanisms on glioma development have not been
reported. In the present study, the expression of Nav1.6 in
different grades of gliomas was firstly described and the role and
mechanism of Nav1.6 isoforms in gliomas were explored. Finally,
certain FDA-approved drugs were identified based on the expression
and structure of Nav1.6.
Materials and methods
Cells and tissues
The glioma cell lines HEB, U87 MG (cat. no. HTB-14;
glioblastoma of unknown origin; verified by STR analysis), A172 and
U251 were obtained from the Shanghai Cell Bank and glioma
pathological tissues were obtained from the Department of
Neurosurgery, The First Affiliated Hospital of China Medical
University (Shenyang, China). Ages ranged from 33 to 76, including
six male and six female patients. Inclusion criteria were patients
diagnosed with glioma and sufficient tumor tissue available for
study. All tissues and specimens were obtained with the approval
(approval no. 20190142) and endorsement of the Ethics Committee of
China Medical University (Shenyang, China). The date range of
tissue collection was between July 17, 2019 and August 12, 2021.
Written informed consent was obtained from each tumor tissue donor
for the use of the tumor tissue and clinical data for future
research.
Cell culture and transfection
Glioma cells A172 and U251 were cultured in DMEM
sugar-free medium (Gibco; Thermo Fisher Scientific, Inc.)
containing 10% fetal bovine serum (Gibco; Thermo Fisher Scientific,
Inc.) and 1% double anti-bodies (Gibco; Thermo Fisher Scientific,
Inc.). All cells were cultured at 37°C in an atmosphere containing
5% CO2. Glioma A172 and U251 cell lines in logarithmic
growth phase were received and transfection was performed at 37°C
for 24 h using Lipofectamine® 3000 reagent according to
the manufacturer's instructions (Invitrogen; Thermo Fisher
Scientific, Inc.). The concentration of transfected small
interfering (si)RNAs was 50 nM. The efficiency of transfection was
verified by qPCR and western blotting (WB). The sequences of the
siRNAs used are shown in Table
SI.
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
The corresponding tissues or cells
(1×107) were received lysed by TRIzol®
reagent (Invitrogen; Thermo Fisher Scientific, Inc.); RNA was
extracted by chloroform, precipitated by isopropanol and lysed in
the presence of DEPC in water to inactivate RNAse enzymes. RNA
purification was performed according to the instructions of the
corresponding reverse transcription kit Shanghai Yeasen
Biotechnology Co., Ltd. Hifair® III 1st Strand cDNA
Synthesis SuperMix (cat. no. 11141ES60; Shanghai Yeasen
Biotechnology Co., Ltd.) was used for qPCR. The thermocycling
conditions for the RT-qPCR reaction were as follows: 25°C for 5
min, 55°C for 15 min, and 85°C for 5 min for the reverse
transcription step, and 95°C for 5 min for the pre-denaturation
step followed by one cycle; 95°C for 10 sec for the denaturation
step followed by 40 cycles; and 60°C for 30 sec for the extension
step followed by 40 cycles. The reference gene was β-actin,
quantified by 2−ΔΔCq method (23). The sequences of the primers used
are shown in Table SI.
WB
Protein quantification was performed using a BCA kit
(cat. no. WB6501; NCM Biotech; http://www.ncmbio.com/) according to the
manufacturer's instructions. Phosphatase inhibitor (1%) and
protease inhibitor were added during cell lysis (cat. no. P002; NCM
Biotech). Proteins were separated using SDS-PAGE. A 7.5% gel (cat.
no. P2011; NCM Biotech) was used, and 20 µg of protein was
loaded per lane. This was performed in Tris-glycine buffer system.
The separated proteins were subsequently transferred to
0.45-µm PVDF membrane (Merck KGaA) and Skim milk powder
closed at room temperature for two h. Next, the membranes were
rinsed three times with TBST prepared with 0.1% Tween 20 for 8 min
each. The membranes were incubated overnight at 4°C with primary
antibodies against SCN8A (1:800; cat. no. ab230654; Abcam) and
β-actin (1:5,000; cat. no. bs-0061R; BIOSS). Following the primary
incubation, membranes were incubated at room temperature with
HRP-labeled goat anti-rabbit IgG secondary antibody (1:10,000; cat.
no. bs-0295G; BIOSS). Membranes were washed with TBST for 10 min
each time. Luminescence was performed, developed and images of the
protein bands were captured for analysis using the ECL
chemiluminescence kit (NCM). Densitometric analysis was performed
using ImageJ software version 1.52a (National Institutes of
Health).
Immunohistochemistry
Paraffin sections of 5-µm thickness were
first dewaxed with xylene for 30 min after 1 h in an oven at 60°C.
Subsequently, the sections were placed sequentially in a gradient
alcohol and the tissue sections were washed in water to remove
residual alcohol and dewaxing agent while the sections were
hydrated. Sections were immersed in citrate buffer and placed in a
microwave oven for antigen repair, then washed in PBS. Blocking
treatment was performed using 5% bovine serum albumin for 1 h at
room temperature. Sections were washed with PBS and incubated with
3% hydrogen peroxide. Deparaffinized sections were incubated
overnight at 4°C with primary antibodies against SCN8A (1:100) and
subsequently with HRP-conjugated goat anti-rabbit IgG secondary
antibody (1:1,000) for 1 h at room temperature. The slides were
washed and stained with DAB for color development. Color
development was terminated by tap water. Hematoxylin
counterstaining followed, using hydrochloric acid for 1% alcohol
fractionation. Tap water was used to reverse the blue, followed by
alcohol dehydration (gradient concentration) and dewaxing with 95%
alcohol. Next, xylene dehydration, sealing and drying of the film
was performed. Tissues were observed under a light microscope and
images were captured.
Cellular experiments
Cell Counting Kit-8 (CCK-8) cell
proliferation assay
A172 and U251 cells were inoculated into 96-well
plates at a density of 1,500 and 2,500 per well, respectively.
CCK-8 assay was performed following the manufacturer's protocol
(BIOSS). Each well was spiked with 10 µl of CCK-8 reagent
and incubated at 37°C for 2 h, followed by detection at 450 nm
using a multifunctional enzyme marker (BioTek Instruments,
Inc.).
Wound healing assay
A172 and U251 cells were spread into six-well plates
at a density of 2×105 and 3×105 per well and
treated with mitomycin (1 µg/ml) after spreading. Cells were
scratched with a sterile gun tip. Cells were serum-starved (FBS
<5%) during the assay. Images were captured at different time
points (0, 12 and 24 h) using an inverted fluorescence microscope
ECLIPSE Ts2 (Nikon Corporation). Data analysis was performed using
GraphPad Prism 7.
Transwell cell invasion assay
Matrigel (cat. no. 356234; Corning, Inc.) was placed
overnight at 4°C and then diluted 1:8 and wrapped on the surface of
the upper chamber wrapped in Transwell bottom membrane. Cells
(4×105) were seeded in the upper chamber containing 200
µl of serum-free medium and the lower chamber was filled
with 600 µl of medium containing 10% FBS. Cells were fixed
with 4% paraformaldehyde for 20 min, followed by staining with 0.1%
crystal violet for 15 min at 25°C and inverted microscopy
observation.
Apoptosis measurement by flow
cytometry
Analysis of apoptosis was conducted using the PE
Annexin V Apoptosis Detection kit (cat. no. 559763; BD Biosciences)
according to the manufacturer's instructions. Apoptotic cells were
subsequently detected on LSRFortessa (BD Biosciences) and analyzed
using FACSDiva software (Version 6.2; BD Biosciences). Early
apoptosis plus late apoptosis was used to assess the apoptosis of
cells.
Virtual screening and drug sensitivity
analysis
The FDA-approved and pharmacopeial drugs were
downloaded from TargetMol (24),
which contains 2858 drugs and their structures, with full drug
names and drug IDs shown in Table
SII. The protein structure for SCN8A was obtained from the
AlphaFold Protein Structure Database (accession no. Q9UQD0)
(25,26), and the binding sites were
identified by the GHECOM algorithm (27); virtual screening and molecular
docking were performed using UCSF DOCK 6.9 (28). Finally, docking conformations were
demonstrated using PyMol (Version 2.3) and interaction forces were
analysed and demonstrated by Ligplus (29). A total of 23808 genetic data (RNA:
RNA-seq) and 24359 compounds' data (Compound activity: DTP NCI-60,
which includes 218 drugs that have been approved by the FDA, and
574 drug molecules in clinical trials) have been identified in
NCI-60 cell lines, and were obtained from the CellMiner database
(30). The results of drugs that
have undergone clinical trials and FDA approval were selected to
ensure the reliability of the analysis. P<0.05 was used as a
cut-off in screening the results based on Pearson's correlation
coefficients between each gene expression and each drug.
Statistical analysis
All data are presented as the mean ± standard error
of the mean (M ± SEM). Unpaired Student's t-test was used to
determine the statistical significance between the two groups. The
results were analyzed using GraphPad Prism (version 7.0;
Dotmatics). Depending on the design of the experiment, the data
were analyzed using one-way ANOVA followed by Tukey's post hoc
test. P<0.05 was considered to indicate a statistically
significant difference.
Results
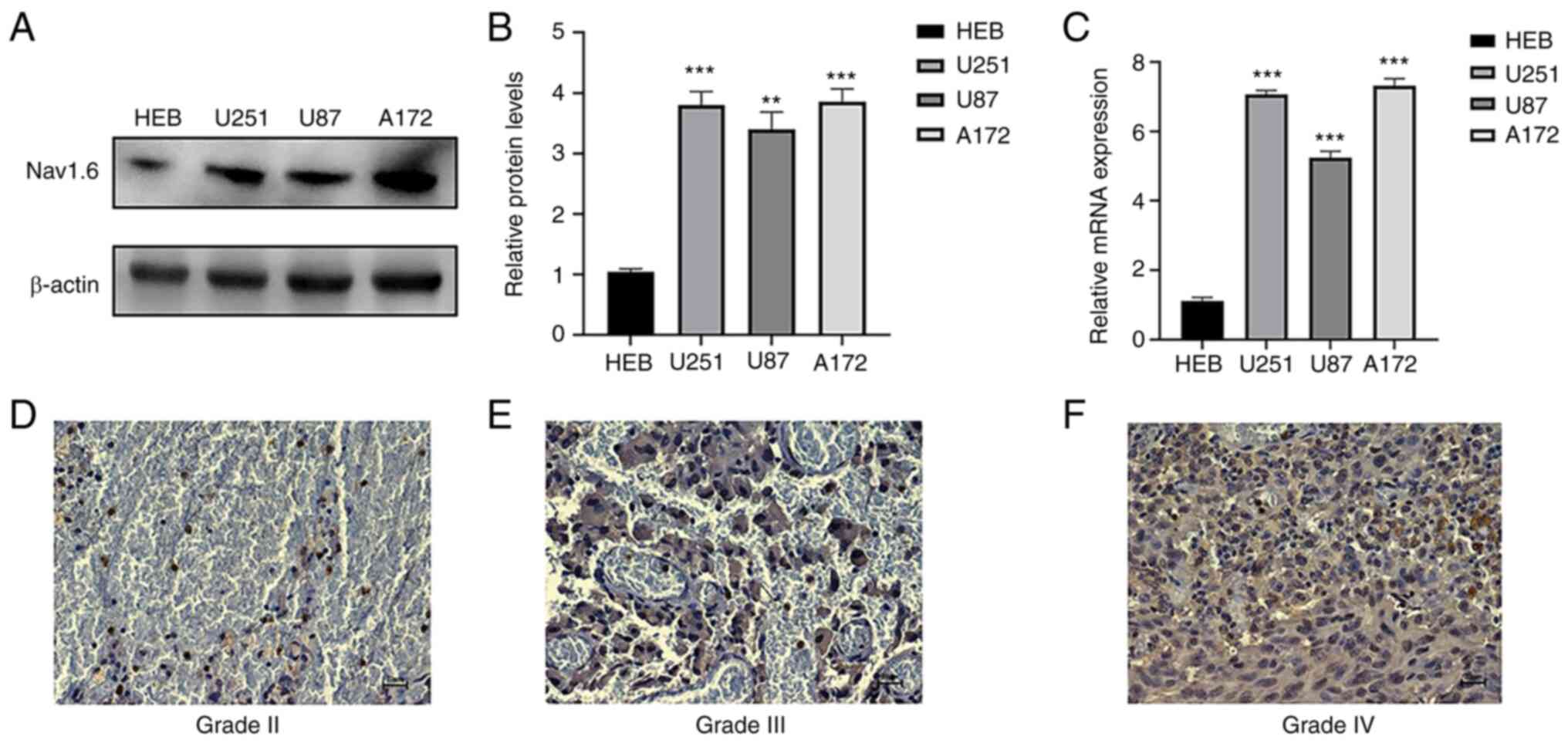
Nav1.6 mRNA and protein expression are
higher in human glioma cells and tissues than in normal glial
cells
The expression of Nav1.6 mRNA and protein in glioma
cell lines U251, U87 A172 and normal glial cells HEB was detected
by RT-qPCR and WB. It was demonstrated that Nav1.6 was expressed in
both normal and glioma cell lines, and the expression in glioma
cell lines U251 and A172 was significantly higher than that in
normal glial cells (Fig. 1A-C),
therefore U251 and A172 cell lines were subsequently used for
knockdown experiments. WB results also showed that the relative
protein expression of Nav1.6 in glioma cell lines U251, U87, and
A172 was several times higher than that in normal glial tissues. To
investigate whether the expression level of Nav1.6 correlated with
the malignant progression of glioma, immunohistochemical
experiments were further performed using paraffin sections of
clinically collected glioma tissues of different grades. The
results revealed that Nav1.6 was mainly expressed on the cytoplasm
and cell membrane, and the expression level of Nav1.6 was
significantly higher in high-grade tissues compared with
lower-grade glioma tissues (Fig.
1D-F). In summary, it was evident that the expression level of
Nav1.6 is increased in glioma and positively correlates with the
pathological grade of glioma.
Knockdown of Nav1.6 inhibits the
proliferation, migration and invasive ability of glioma cells and
promotes apoptosis of glioma cells
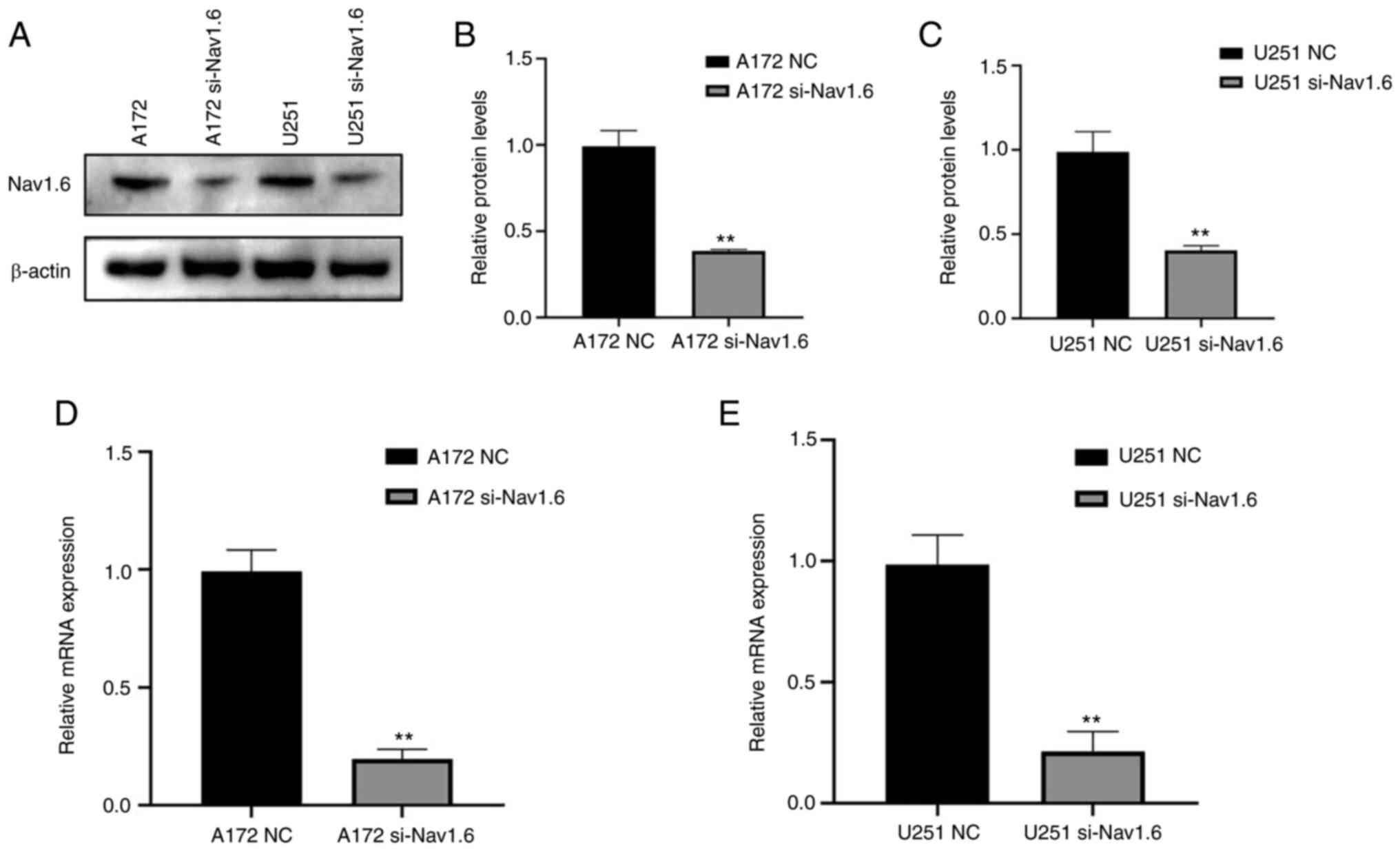
The glioma U251 and A172 cell lines were transfected
with si-Nav1.6 and si-NC, and the efficiency of transfection was
verified by WB (Fig. 2A-C) and
RT-qPCR (Fig. 2D and E). It was
identified that the mRNA and protein levels of Nav1.6 in both U251
and A172 cells were significantly decreased after transfection with
si-Nav1.6 compared with the NC group, and the knockdown effect was
significant.
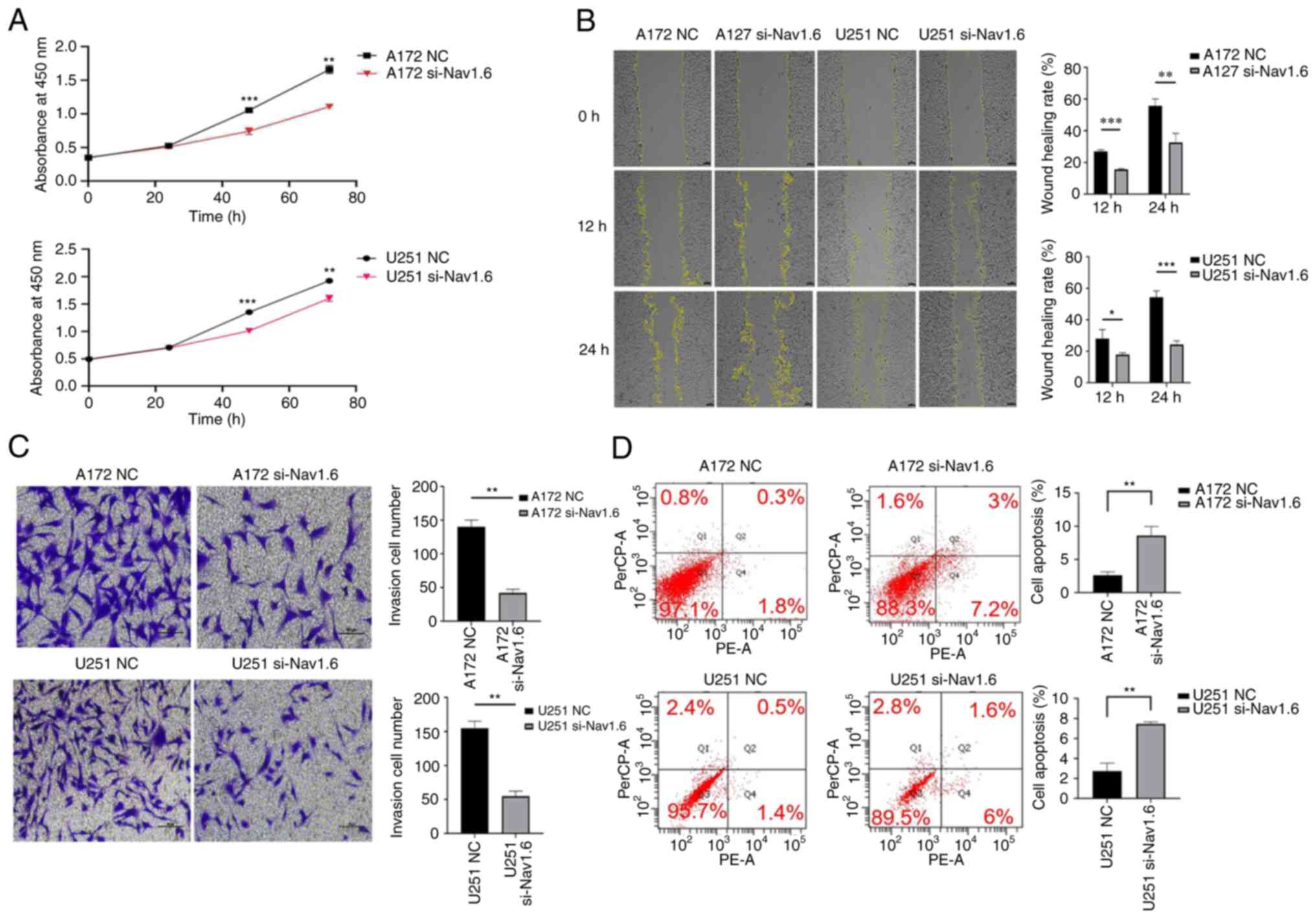
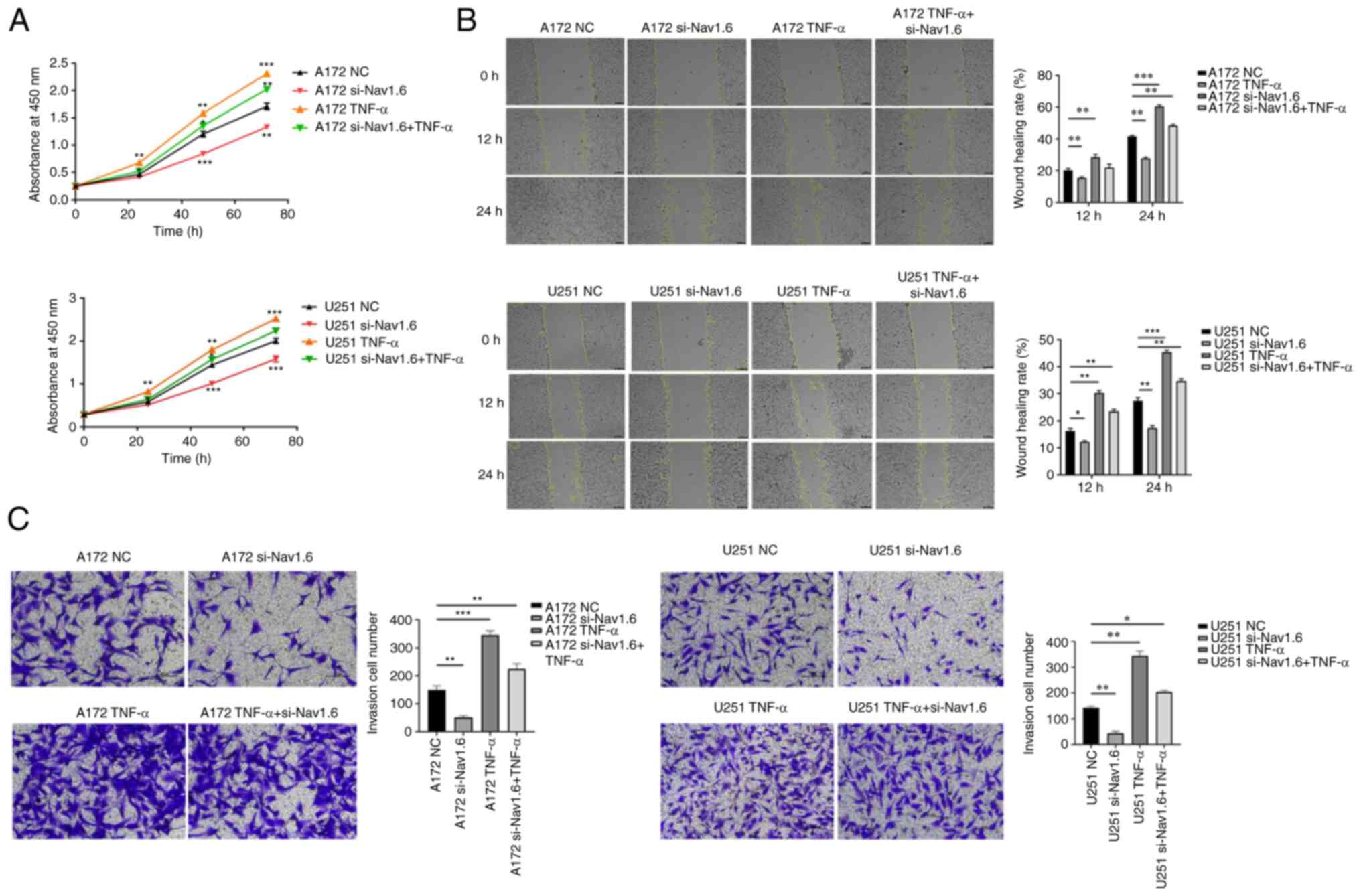
Several assays were conducted to detect the effect
of si-Nav1.6 on glioma cells, including proliferation, wound
healing, cell invasion and flow cytometric analysis apoptosis
assay. The experimental data demonstrated that there was no
significant difference in cell proliferation between A172 si-Nav1.6
and U251 si-Nav1.6 within 24 h compared with the control group, and
transfection of si-Nav1.6 significantly inhibited the proliferation
of glioma A172 and U251 cells after 24 h (P<0.01, Fig. 3A). In addition, it was revealed
that si-Nav1.6 wound healing and invasive abilities of A172 and
U251 cells (Fig. 3B and C,
respectively). Flow cytometric analysis of apoptosis assay showed
that the transfection of si-Nav1.6 promoted the apoptotic rate of
glioma A172 and U251 cells (Fig.
3D).
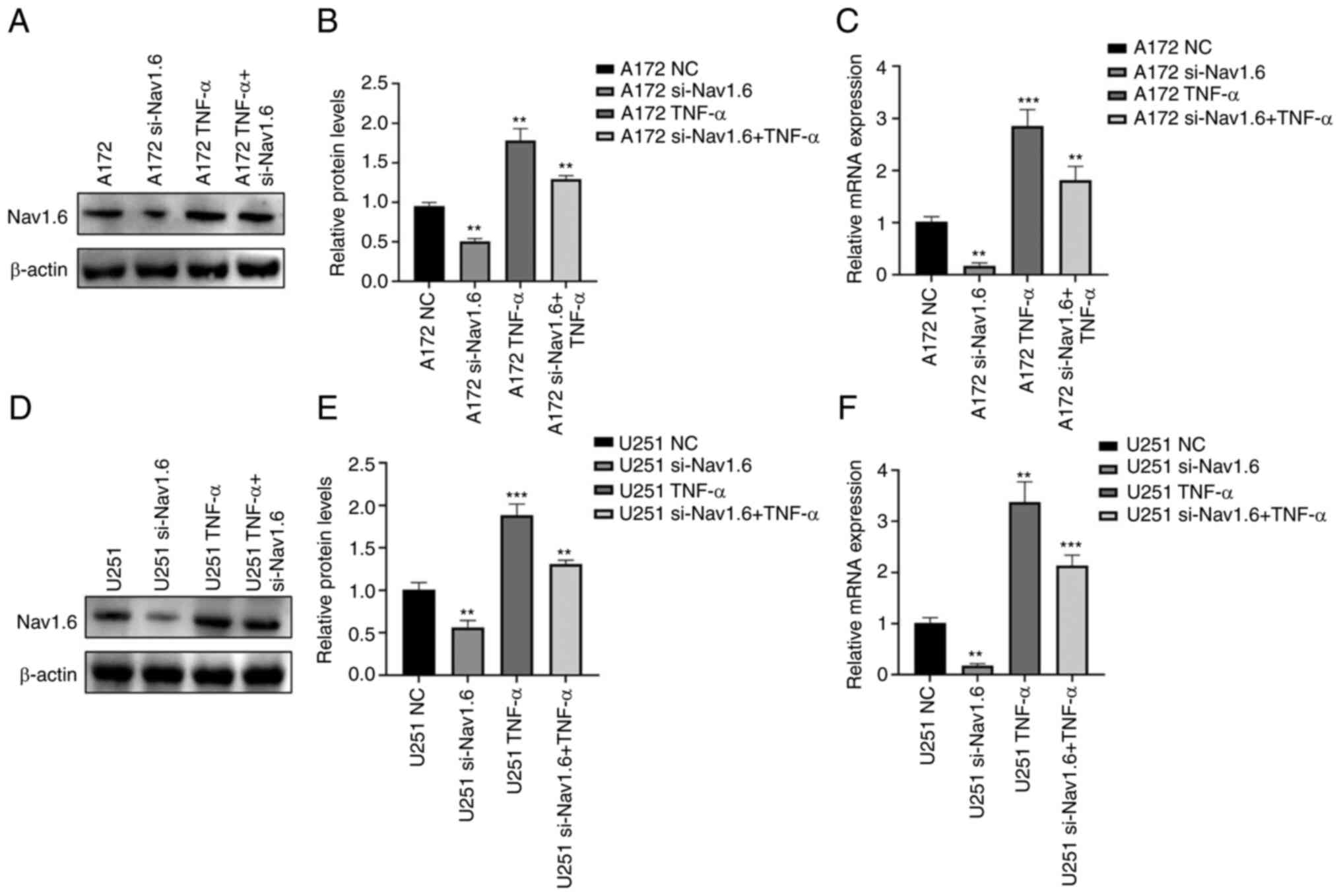
TNF-α induces the expression of Nav1.6
and induces the proliferation, migration and invasion of glioma
cells
Through literature review and analysis of relevant
data, it was found that TNF-α can affect Nav1.6 expression and has
been verified at the electrophysiological level (31), but it has still not been studied
in tumor cells. Therefore, the effect of TNF-α on Nav1.6 expression
was studied at the cellular level. First, the optimal concentration
and optimal time of TNF-α action were mapped by pre-experiments.
The effect of TNF-α on Nav1.6 was detected at concentrations of 1,
10, 100 and 1,000 pg/ml, and the final treatment was selected at an
action concentration of 100 pg/ml. The relative expression levels
of mRNA and protein of Nav1.6 were significantly increased after
the action of TNF-α (100 pg/ml) on A172 and U251 cells. TNF-α
action in cell lines transfected with si-Nav1.6 significantly
increased the protein and mRNA expression of Nav1.6, but was
slightly lower than the direct action of TNF-α (Fig. 4A-F). This suggested that TNF-α
induced the protein and mRNA expression of Nav1.6 in glioma cells
and that si-Nav1.6 partially blocked the induction of TNF-α.
To determine the effects of TNF-α on glioma cells,
CCK-8, wound healing and Transwell invasion assays were performed.
Proliferation, migration, and invasion of glioma cells were
significantly enhanced, and TNF-α also resisted the effect of
si-Nav1.6 and increased the level of Nav1.6 expression after
transfection with siRNA, but si-Nav1.6 also partially blocked the
induction of TNF-α (Fig. 5A-C).
The association of Nav1.6 with the development and malignant
progression of gliomas was also further confirmed by the action of
TNF-α.
A virtual screening and sensitivity
analysis of drugs
Based on the aforementioned analysis, in order to
explore the drugs targeting SCN8A, molecular docking and virtual
screening against FDA-approved drugs was conducted based on the 3D
structure of the protein. A list of the top 10 scoring drug
molecules is shown in Table I and
a summary of the scores is presented in Table SII. The conformation and
interaction force analysis of the top six scoring drugs can be
observed in Fig. 6, and the
results indicated that Neomycin Sulphate B (T2096), Paromomycin
Sulfate (T1104), Neomycin sulfate (T0950), Plerixafor 8HCl (T1776
and T1776L) and Isavuconazonium sulfate (T3934) showed stronger
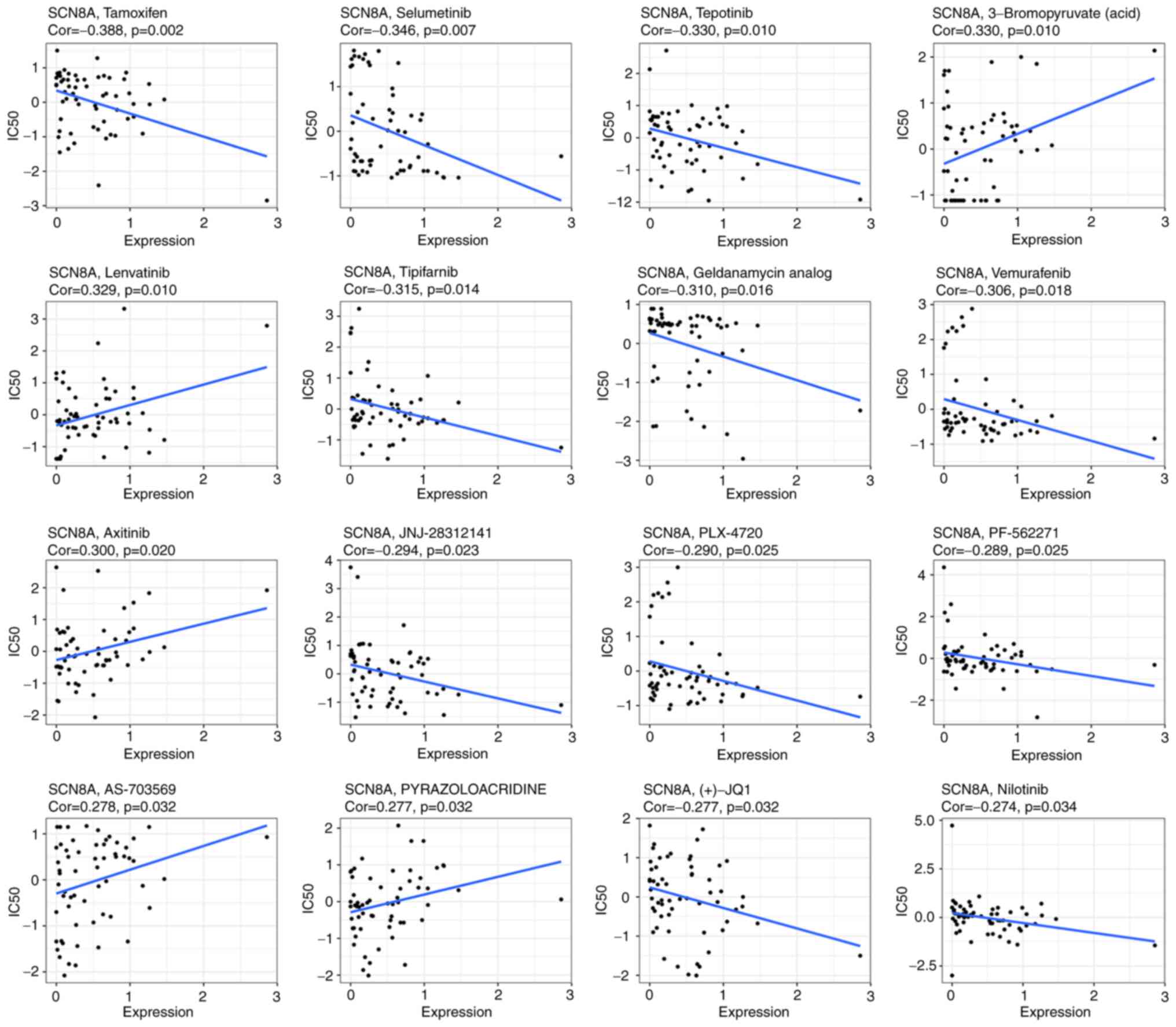
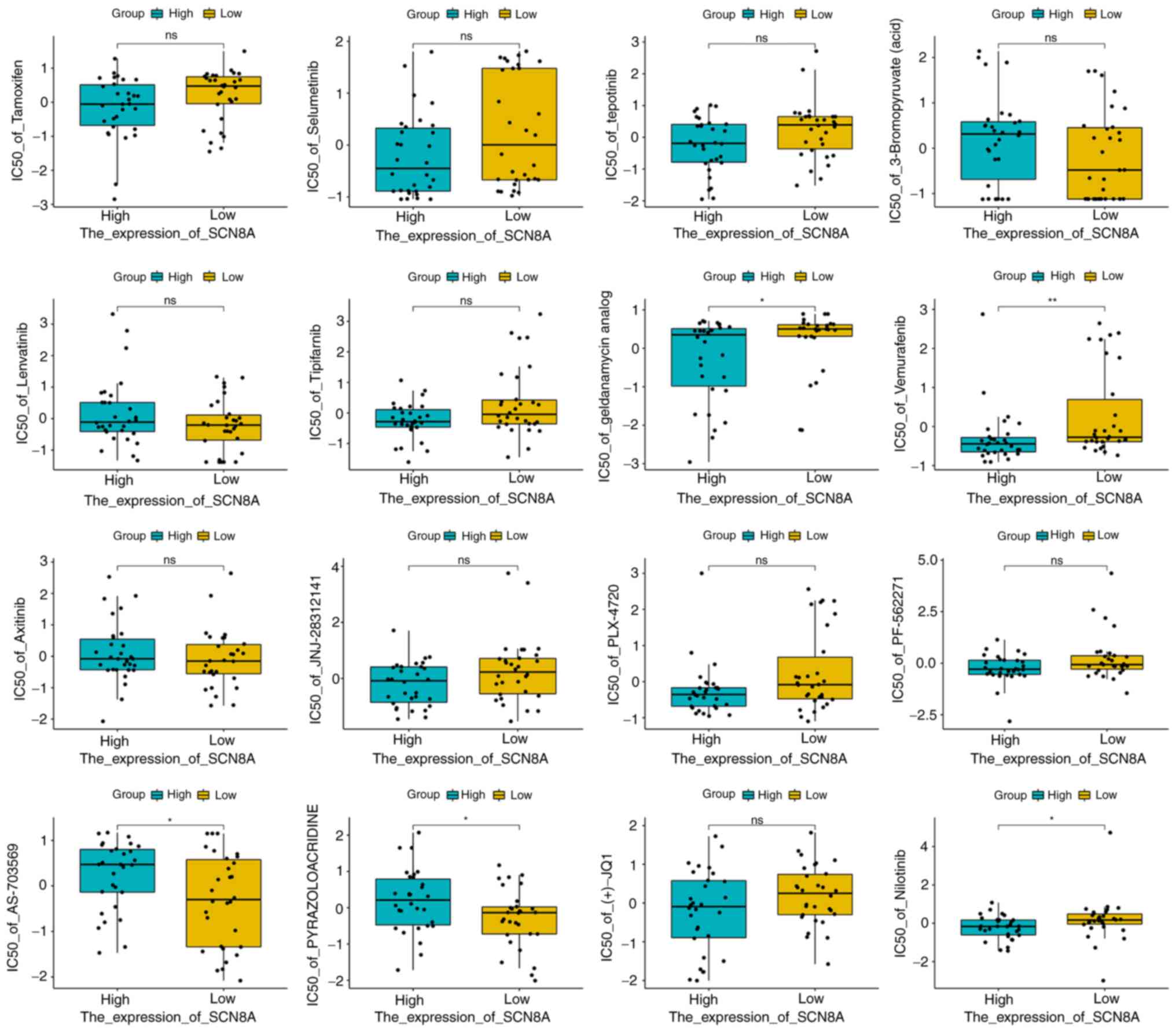
binding to SCN8A. The results of NCI-60 drug sensitivity analysis
similarly revealed that the sensitivity of 23 drugs correlated with
the expression of SCN8A. A total of 3 of these drugs [Tamoxifen
(TAM), Selumetinib and tepotinib] demonstrated a significant
correlation (Table II, Fig. 7). In addition, different
expression levels of SCN8A similarly affected the half maximal
inhibitory concentration values of the drugs (Fig. 8). Finally, it is worth mentioning
that numerous of the screened drugs already have therapeutic value
in non-tumor and other oncologic diseases, and based on the
analysis of the aforementioned results, they could also be
potential drugs for the treatment of glioma.
 | Table IThe 10 drugs with the highest docking
scores to Nav1.6. |
Table I
The 10 drugs with the highest docking
scores to Nav1.6.
| ID | Grid Score | Grid_vdw | Grid_es | Int_energy |
|---|
| T2096 | −122.5961 | −67.5617 | −55.0345 | 14.1862 |
| T1104 | −122.1724 | −69.007 | −53.1654 | 19.9233 |
| T0950 | −122.1169 | −70.7967 | −51.3203 | 15.3193 |
| T1776 | −121.5208 | −71.2886 | −50.2322 | 4.5184 |
| T1776L | −120.1207 | −69.3475 | −50.7733 | 4.7753 |
| T3934 | −112.7299 | −102.7981 | −9.9318 | 31.7494 |
| T1013 | −111.8229 | −68.5774 | −43.2455 | 12.3296 |
| T4509 | −111.4996 | −110.4062 | −1.0934 | 24.5032 |
| T2119 | −111.4223 | −106.4619 | −4.9605 | 22.766 |
| T1670 | −110.7266 | −107.2315 | −3.4951 | 54.6025 |
| Table IIAn analysis of correlations between
SCN8A expression and the sensitivity of FDA-approved drugs. |
Table II
An analysis of correlations between
SCN8A expression and the sensitivity of FDA-approved drugs.
| Gene | Drug | Correlation | P-value |
|---|
| SCN8A | Tamoxifen | −0.3877366 | 0.00220652 |
| SCN8A | Selumetinib | −0.3460751 | 0.0067579 |
| SCN8A | tepotinib | −0.3302933 | 0.00995474 |
| SCN8A | 3-Bromopyruvate
(acid) | 0.32957993 | 0.01012599 |
| SCN8A | Lenvatinib | 0.32851997 | 0.01038515 |
| SCN8A | Tipifarnib | −0.3153499 | 0.0141166 |
| SCN8A | geldanamycin
analog | −0.3099119 | 0.01596459 |
| SCN8A | Vemurafenib | −0.3057577 | 0.01751239 |
| SCN8A | Axitinib | 0.30044955 | 0.01967502 |
| SCN8A | JNJ-28312141 | −0.2939791 | 0.02261459 |
| SCN8A | PLX-4720 | −0.2898328 | 0.02468725 |
| SCN8A | PF-562271 | −0.2885017 | 0.02538575 |
| SCN8A | AS-703569 | 0.27787558 | 0.03158141 |
| SCN8A |
PYRAZOLOACRIDINE | 0.27702531 | 0.03212751 |
| SCN8A | (+)-JQ1 | −0.2765262 | 0.03245173 |
| SCN8A | Nilotinib | −0.2744798 | 0.0338097 |
| SCN8A | VINORELBINE | −0.273364 | 0.0345699 |
| SCN8A | ARRY-162 | −0.2731846 | 0.03469338 |
| SCN8A | RO-5126766 | −0.2696654 | 0.03719198 |
| SCN8A | EMD-534085 | −0.2692616 | 0.037488 |
| SCN8A | Amonafide | 0.26430171 | 0.04128567 |
| SCN8A | HYPOTHEMYCIN | −0.2622113 | 0.04297867 |
| SCN8A | Pimasertib | −0.2599519 | 0.04487232 |
Discussion
Metastatic and aggressive progression of tumors is
the main reason for the poor prognosis and low survival rate of
most patients with malignant tumors. Therefore, it is particularly
important to explore the specific mechanisms of metastasis and
invasion of malignant tumors. In 2017, the electron microscopic
structure of eukaryotic VGSCs was first reported by Pan et
al (32) using cryo-electron
microscopy. This also provided a basis for further understanding of
the VGSCs. VGSCs only started to be studied in gliomas in 2002, and
researchers initially detected and elaborated the expression of
different ion isoforms in gliomas, but no systematic study of tumor
cells has been performed yet (33). The aberrant expression of ion
channels in gliomas also suggests that VGSCs have the potential to
become glioma-related biomarkers.
VGSCs are composed of core α and β subunits
(34). The α-subunit is a key
determinant of the activity and function of VGSCs, whereas the
β-subunit can regulate channel expression, homeostasis and kinetic
properties by binding to the α-subunit. α-subunit can function
intact by binding to the β-subunit. α-subunit and β-subunit
interactions can enhance the internal repair capacity of the
channel, improve channel stability and kinetic properties, and
increase the expression of the channel. Even the α subunits of two
VGSCs can interact to regulate the activity of ion channels, with
the direct mutual region located at the N terminus (35). Sodium channels are key initiators
of electrical signals in all animals, and electrical signals play a
crucial importance in a range of basic physiological activities
including neural activity and muscle contraction. In humans, there
are nine known subtypes of VGSCs that function in different organs
and physiological processes. In the nervous system, sodium channels
are widely distributed. Abnormalities in sodium channels contribute
to the progression of neurological disorders, muscular disorders
and cardiac ailments. This also appears to be consistent with the
currently known distribution of ion channel subtypes, further
validating the association of VGSCs with tumor progression.
In the present experiment, it was verified that
Nav1.6 is highly expressed in gliomas. This is also consistent with
previous studies and the previous experimental results (Wang et
al, unpublished data) of the authors' group. Previous studies
also showed that Nav1.6 was associated with malignant progression
of glioma, and inhibition of its expression reduced the metastatic
and invasive potential of glioma, which was also consistent with
the hypothesis of the current study. There are few studies on
Nav1.6 in tumors, and certain of the rare studies are in colorectal
and cervical cancers (7,36). The majority of the studies involve
disease models, mouse-rat models and electrophysiology. Extending
the studies in colorectal and cervical cancer to other cellular and
cancer models will also help advance the understanding and
mechanistic studies of sodium channels. In addition, Nav1.6
isoforms have also been proposed as a therapeutic target for the
treatment of cancer, as they have been shown to play a role in
promoting tumor growth and migration. Targeting Nav1.6 isoforms
with specific inhibitors, such as siRNAs or drugs, has been shown
to reduce the growth and spread of cancer cells in vitro and
in animal models (37). However,
while these findings are promising, further research is needed to
determine the usefulness of Nav1.6 isoforms as diagnostic markers
and therapeutic targets in the treatment of clinical metastatic
tumors. This may involve further studies in larger animal models
and clinical trials in humans to determine the safety and efficacy
of targeting Nav1.6 isoforms for the treatment of cancer. Overall,
Nav1.6 isoforms appear to be promising as diagnostic markers and
therapeutic targets in the treatment of clinical metastatic tumors,
but further research is required to determine their usefulness in
clinical practice.
Through an extensive literature review, it was found
that Cheng et al (31)
indicated that TNF-α upregulated Nav1.6 currents in retinal cells;
Ding et al (38) also
found that the TNF-α/STAT3 signalling pathway upregulated Nav1.6
expression in the dorsal root ganglion. However, the regulation
within the tumor remains unclear, thus the role of TNF-α in this
regard was investigated in the present study. As a classical
inflammatory cytokine, TNF-α binds to multiple receptors within the
nervous system and exerts multiple functions. TNF-α does not have a
single effect on tumor cells, but can both promote and inhibit cell
proliferation and induce apoptosis (39). In the present study, it was
demonstrated that TNF-α promotes the production of Nav1.6 protein
and mRNA, and si-Nav1.6 could partially block the induction of
TNF-α in glioma, suggesting that TNF-α is associated with malignant
progression of tumors in glioma.
Subsequently, a number of drugs that have potential
therapeutic value for gliomas were screened. After the initial
screening was completed, there were certain interesting and
surprising findings during the further analysis of drug feasibility
and possible relevant mechanisms. The available literature on
several drugs is consistent with the current experimental results,
and this has greatly increased the authors' confidence in these
drugs. Among the numerous studies on the mechanisms of VGSCs
regulating tumor metastasis, the most likely mechanisms are the
following: The inward flow of sodium ions affects the PH and
calcium ion concentrations, which carry hydrogen ions out of the
cell into the extracellular matrix by the action of the
sodium-hydrogen exchanger, and the activation of cysteine proteases
during acidification accelerates the degradation of the
extracellular matrix and thus promotes the PH-dependent tumor cell
metastasis (40). There is also
another sodium-calcium exchanger (NCX) where ion exchange leads to
changes in intracellular calcium ion concentration and PH as well
as changes in the cellular microenvironment. The altered calcium
ion concentration promotes the formation of pedicles/endopods,
which indirectly affects the cell migration and metastasis process
(41-43). Among the results of virtual
screening based on the structure of SCN8A, T0950 (trade name
Neomycin sulfate) is an FDA-approved aminoglycoside antibiotic and
phospholipase C inhibitor known to be commercially available. It
can inhibit bacteria, cell proliferation and angiogenesis, and more
importantly, it is also a commercially available calcium channel
blocker (44). As a drug
targeting Nav1.6, it also plays a role in the blockade of calcium
ions. This appears to be consistent with the mechanistic hypothesis
that currently exists. Blocking the inward flow of sodium ions
affects the action of NCX, somehow blocking calcium ions as well.
And as early as 1999, Mikkelsen et al (45) found that it also has an effect on
pH. Perhaps there is also a relationship with the inhibition of the
action of the sodium-hydrogen exchanger. These previous studies
suggested that the drug has far-reaching research prospects and the
reliability of drug screening is further enhanced. The specific
mechanism is to be verified and explored in depth in the authors'
future study.
Meanwhile, it was found that T1716 (Trade name
Echinacoside), possesses a variety of biological activities. In
vitro experiments have revealed that Echinacoside behaves as an
antioxidant and an effective free radical scavenger in
vitro. TNF-α-induced apoptosis in SH-SY5Y neuronal cells is
reversed by this agent. This also supports the relationship between
Nav1.6 and TNF-α in the current study. Echinacoside can block
Nav1.6 expression and act on TNF-α-induced apoptosis. In addition,
it was recently reported that Echinacoside can also effectively
inhibit the Wnt/β-catenin signaling pathway (46). Echinacoside plays a
neuroprotective role by activating Trk receptor and its downstream
signaling pathway (47). This
provides a pathway for further research on the pathway and
mechanism. It also raises the authors' expectations that
Echinacoside can play a key role in neurological tumors as
well.
In the subsequent NCI-60 drug sensitivity analysis,
the sensitivity of 23 drugs was correlated with the expression of
SCN8A. A total of 3 of these drugs (TAM, Selumetinib and tepotinib)
exhibited significant correlations (P<0.01). Of particular
interest was the strongest correlation, TAM, which is a selective
estrogen receptor modulator, widely used as a treatment and
prevention of breast cancer. Different concentrations of TAM exert
different effects, with low concentrations of (0.1-1 µM)
inducing cell cycle arrest and drug concentrations (>5
µM) inducing apoptosis in breast cancer cells. A total of 5
µM TAM rapidly induces sustained ERK1/2 activation in
ER+ breast cancer cells (MCF-7 and T47D) (48). It has also been identified that
TAM can also induce knockout in CreER (T2) transgenic mice
(49). By contrast, it is
carcinogenic in the human uterus and rat liver (50). These experiments have demonstrated
that TAM exerts different effects in different tumors and diseases
and has a large application potential. This has been confirmed by
extensive clinical trial studies. Certain studies that completed
clinical phase IV have fully demonstrated the clinical promise of
TAM in the treatment of breast cancer (NCT00537771), infertility
(NCT02690870), vaginal bleeding (NCT04933240) and endometrium
(NCT03060304); several studies in clinical phase II [NCT00108069
(51), NCT00492687, NCT04765098,
NCT00024336, NCT00541138 and NCT00006388] suggested that TAM has
great potential in the treatment of gliomas. TAM has been shown to
cross the blood-brain barrier, which renders it a promising drug
for the treatment of brain tumors (52). The exact mechanism by which TAM
crosses the blood-brain barrier is not well understood. It is
considered that TAM may cross the blood-brain barrier through
passive diffusion, transcytosis, or facilitated transport (53). The exact absorption percentage of
TAM in the brain is not well characterized, but it is considered to
be low due to the presence of the blood-brain barrier. To further
understand the potential of TAM in the treatment of gliomas, animal
experiments need be performed. These experiments can enable to
determine the efficacy and safety of TAM in treating brain tumors,
as well as to further understand the mechanism by which TAM crosses
the blood-brain barrier. Animal experiments may involve the use of
animal models of gliomas, including mice or rats, and can help to
evaluate the efficacy and safety of TAM in treating brain tumors.
These experiments can also aid to determine the optimal dosing
regimen for TAM, as well as to identify any potential side effects
of the drug. Overall, animal experiments can provide valuable
information about the potential of TAM in the treatment of gliomas
and can help to expand the understanding of the mechanism by which
TAM crosses the blood-brain barrier.
These existing studies demonstrated the therapeutic
value and potential of these screened drugs as treatment for glioma
in non-oncologic diseases and other oncologic diseases. It is
considered that in the next studies the authors' group will be able
to fully refine and fill certain of the gaps of currently unstudied
drugs and provide a new possibility for the treatment of
glioma.
In conclusion, the present study first demonstrated
the expression and role of Nav1.6 in glioma, laying the foundation
for further investigation of specific mechanisms, and several
FDA-approved drugs that may be glioma treatment candidate drugs
were screened.
Supplementary Data
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
AY, WJ, LJY, OSW and ZXD designed the study. AY,
HXD, GJT and ZXD performed the experimental work. AY, ZXD, WJ, OSW
and LJY performed the data analysis. AY, ZXD, HXD, GJT, LJY, OSW
and WJ produced the main drafts of the text and figures. AY, ZXD
and WJ confirm the authenticity of all raw data. All authors have
reviewed, corrected, read and approved the final manuscript.
Ethics approval and consent to
participate
All tissues and specimens were obtained with the
approval and endorsement of the Ethics Committee of China Medical
University (Shenyang, China). Written informed consent was provided
by all patients for the use of their glioma tissue in the present
study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
The authors would like to thank Mr Zhihong Zong from
the Department of Biochemistry and Molecular Biology, School of
Basic Medical Sciences, China Medical University for providing the
experimental space and technical support.
Funding
The present study was supported by the National Natural Science
Foundation of China (grant no. 31100770) and the Shenyang Natural
Science Foundation (grant no. 18-013-0-60).
References
|
1
|
Lapointe S, Perry A and Butowski NA:
Primary brain tumours in adults. Lancet. 392:432–446. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Luo Q, Wu T, Wu W, Chen G, Luo X, Jiang L,
Tao H, Rong M, Kang S and Deng M: The functional role of
voltage-gated sodium channel Nav1.5 in metastatic breast cancer.
Front Pharmacol. 11:11112020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Altamura C, Greco MR, Carratu MR and
Cardone RA: Desaphy JF. Emerging roles for ion channels in ovarian
cancer: Pathomechanisms and pharmacological treatment. Cancers
(Basel). 13:6682021. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Brummelhuis IS, Fiascone SJ, Hasselblatt
KT and Frendl G: Elias KM. Voltage-gated sodium channels as
potential biomarkers and therapeutic targets for epithelial ovarian
cancer. Cancers (Basel). 13:54372021. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Brackenbury WJ: Voltage-gated sodium
channels and metastatic disease. Channels (Austin). 6:352–361.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liu J, Tan H, Yang W, Yao S and Hong L:
The voltage-gated sodium channel Nav1.7 associated with endometrial
cancer. J Cancer. 10:4954–4960. 2019. View Article : Google Scholar :
|
|
7
|
Lin S, Lv Y, Xu J, Mao X, Chen Z and Lu W:
Over-expression of Nav1.6 channels is associated with lymph node
metastases in colorectal cancer. World J Surg Oncol. 17:1752019.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Black JA, Liu S and Waxman SG: Sodium
channel activity modulates multiple functions in microglia. Glia.
57:1072–1081. 2009. View Article : Google Scholar
|
|
9
|
Takayasu T, Kurisu K, Esquenazi Y and
Ballester LY: Ion channels and their role in the pathophysiology of
gliomas. Mol Cancer Ther. 19:1959–1969. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang J, Ou SW, Wang YJ, Kameyama M,
Kameyama A and Zong ZH: Analysis of four novel variants of
Nav1.5/SCN5A cloned from the brain. Neurosci Res. 64:339–347. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Guan G, Zhao M, Xu X, Boczek T, Mao X, Li
Z, Gao Q, Li J, Zhao D, Niu W, et al: Abnormal changes in
voltage-gated sodium channels subtypes NaV1.1, NaV1.2, NaV1.3,
NaV1.6 and CaM/CaMKII pathway in low-grade astrocytoma. Neurosci
Lett. 674:148–155. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xing D, Wang J, Ou S, Wang Y, Qiu B, Ding
D, Guo F and Gao Q: Expression of neonatal Nav1.5 in human brain
astrocytoma and its effect on proliferation, invasion and apoptosis
of astrocytoma cells. Oncol Rep. 31:2692–2700. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mao W, Zhang J, Korner H, Jiang Y and Ying
S: The emerging role of voltage-gated sodium channels in tumor
biology. Front Oncol. 9:1242019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xu L, Ding X, Wang T, Mou S, Sun H and Hou
T: Voltage-gated sodium channels: Structures, functions, and
molecular modeling. Drug Discov Today. 24:1389–1397. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang J, Ou SW and Wang YJ: Distribution
and function of voltage-gated sodium channels in the nervous
system. Channels (Austin). 11:534–554. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Diaz-Garcia A and Varela D: Voltage-Gated
K(+)/Na(+) channels and scorpion venom toxins in cancer. Front
Pharmacol. 11:9132020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Schreiber JM, Tochen L, Brown M, Evans S,
Ball LJ, Bumbut A, Thewamit R, Whitehead MT, Black C, Boutzoukas E,
et al: A multi-disciplinary clinic for SCN8A-related epilepsy.
Epilepsy Res. 159:1062612020. View Article : Google Scholar
|
|
18
|
Bennett DL, Clark AJ, Huang J, Waxman SG
and Dib-Hajj SD: The role of Voltage-gated sodium channels in pain
signaling. Physiol Rev. 99:1079–1151. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Alrashdi B, Dawod B, Schampel A, Tacke S,
Kuerten S, Marshall JS and Côté PD: Nav1.6 promotes inflammation
and neuronal degeneration in a mouse model of multiple sclerosis. J
Neuroinflammation. 16:2152019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Menezes LFS, Sabia Junior EF, Tibery DV,
Carneiro LDA and Schwartz EF: Epilepsy-Related Voltage-gated sodium
channelopathies: A review. Front Pharmacol. 11:12762020. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Angus M and Ruben P: Voltage gated sodium
channels in cancer and their potential mechanisms of action.
Channels (Austin). 13:400–409. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Besson P, Driffort V, Bon E, Gradek F,
Chevalier S and Roger S: How do voltage-gated sodium channels
enhance migration and invasiveness in cancer cells? Biochim Biophys
Acta. 1848:2493–2501. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
24
|
FDA-Approved & Pharmacopeia Drug
Library. https://www.targetmol.com/compound-library/fda_approved_&_pharmacopeia_drug_library
Accessed March 8, 2022.
|
|
25
|
Jumper J, Evans R, Pritzel A, Green T,
Figurnov M, Ronneberger O, Tunyasuvunakool K, Bates R, Žídek A,
Potapenko A, et al: Highly accurate protein structure prediction
with AlphaFold. Nature. 596:583–589. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Varadi M, Anyango S, Deshpande M, Nair S,
Natassia C, Yordanova G, Yuan D, Stroe O, Wood G, Laydon A, et al:
AlphaFold protein structure database: Massively expanding the
structural coverage of protein-sequence space with high-accuracy
models. Nucleic Acids Res. 50:D439–D444. 2022. View Article : Google Scholar :
|
|
27
|
Kawabata T: Detection of multiscale
pockets on protein surfaces using mathematical morphology.
Proteins. 78:1195–1211. 2010. View Article : Google Scholar
|
|
28
|
Allen WJ, Fochtman BC, Balius TE and Rizzo
RC: Customizable de novo design strategies for DOCK: Application to
HIVgp41 and other therapeutic targets. J Comput Chem. 38:2641–2663.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wallace AC, Laskowski RA and Thornton JM:
LIGPLOT: A program to generate schematic diagrams of protein-ligand
interactions. Protein Eng. 8:127–134. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Reinhold WC, Sunshine M, Liu H, Varma S,
Kohn KW, Morris J, Doroshow J and Pommier Y: CellMiner: A web-based
suite of genomic and pharmacologic tools to explore transcript and
drug patterns in the NCI-60 cell line set. Cancer Res.
72:3499–3511. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cheng S, Wang HN, Xu LJ, Li F, Miao Y, Lei
B, Sun X and Wang Z: Soluble tumor necrosis factor-alpha-induced
hyperexcitability contributes to retinal ganglion cell apoptosis by
enhancing Nav1.6 in experimental glaucoma. J Neuroinflammation.
18:1822021. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Pan X, Li Z, Zhou Q, Shen H, Wu K, Huang
X, Chen J, Zhang J, Zhu X, Lei J, et al: Structure of the human
voltage-gated sodium channel Nav1.4 in complex with β1.
Science. 362:eaau24862018. View Article : Google Scholar
|
|
33
|
Schrey M, Codina C, Kraft R, Beetz C,
Kalff R, Wölfl S and Patt S: Molecular characterization of
voltage-gated sodium channels in human gliomas. Neuroreport.
13:2493–2498. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Catterall WA, Lenaeus MJ and Gamal El-Din
TM: Structure and pharmacology of voltage-gated sodium and calcium
channels. Annu Rev Pharmacol Toxicol. 60:133–154. 2020. View Article : Google Scholar
|
|
35
|
Clatot J, Ziyadeh-Isleem A, Maugenre S,
Denjoy I, Liu H, Dilanian G, Hatem SN, Deschênes I, Coulombe A,
Guicheney P and Neyroud N: Dominant-negative effect of SCN5A
N-terminal mutations through the interaction of Na(v)1.5
α-subunits. Cardiovasc Res. 96:53–63. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hernandez-Plata E, Ortiz CS,
Marquina-Castillo B, Medina-Martinez I, Alfaro A, Berumen J, Rivera
M and Gomora JC: Overexpression of NaV 1.6 channels is associated
with the invasion capacity of human cervical cancer. Int J Cancer.
130:2013–2023. 2012. View Article : Google Scholar
|
|
37
|
Yang Y, Luo Z, Hao Y, Ba W, Wang R, Wang
W, Ding X and Li C: mTOR-mediated Na+/Ca2+
exchange affects cell proliferation and metastasis of melanoma
cells. Biomed Pharmacother. 92:744–749. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ding HH, Zhang SB, Lv YY, Ma C, Liu M,
Zhang KB, Ruan XC, Wei JY, Xin WJ and Wu SL: TNF-α/STAT3 pathway
epigenetically upregulates Nav1.6 expression in DRG and contributes
to neuropathic pain induced by L5-VRT. J Neuroinflammation.
16:292019. View Article : Google Scholar
|
|
39
|
Lei Q, Gu H, Li L, Wu T, Xie W, Li M and
Zhao N: TNIP1-mediated TNF-α/NF-κB signalling cascade sustains
glioma cell proliferation. J Cell Mol Med. 24:530–538. 2020.
View Article : Google Scholar
|
|
40
|
Onganer PU and Djamgoz MB: Small-cell lung
cancer (human): Potentiation of endocytic membrane activity by
voltage-gated Na(+) channel expression in vitro. J Membr Biol.
204:67–75. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Reddy Chichili VP, Xiao Y, Seetharaman J,
Cummins TR and Sivaraman J: Structural basis for the modulation of
the neuronal voltage-gated sodium channel NaV1.6 by calmodulin. Sci
Rep. 3:24352013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Bulling A, Brucker C, Berg U, Gratzl M and
Mayerhofer A: Identification of voltage-activated Na+
and K+ channels in human steroid-secreting ovarian
cells. Ann N Y Acad Sci. 868:77–79. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Anderson KJ, Cormier RT and Scott PM: Role
of ion channels in gastrointestinal cancer. World J Gastroenterol.
25:5732–5772. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Huang K, Wang X, Liu Y and Zhao Y: CRAC
channel is inhibited by neomycin in a Ptdlns(4,5)P2-independent
manner. Cell Biochem Funct. 33:97–100. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Mikkelsen NE, Brännvall M, Virtanen A and
Kirsebom LA: Inhibition of RNase P RNA cleavage by aminoglycosides.
Proc Natl Acad Sci USA. 96:6155–6160. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Tang C, Gong L, Lvzi X, Qiu K, Zhang Z and
Wan L: Echinacoside inhibits breast cancer cells by suppressing the
Wnt/β-catenin signaling pathway. Biochem Biophys Res Commun.
526:170–175. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zhu M, Lu C and Li W: Transient exposure
to echinacoside is sufficient to activate Trk signaling and protect
neuronal cells from rotenone. J Neurochem. 124:571–580. 2013.
View Article : Google Scholar
|
|
48
|
Zheng A, Kallio A and Härkönen P:
Tamoxifen-induced rapid death of MCF-7 breast cancer cells is
mediated via extracellularly signal-regulated kinase signaling and
can be abrogated by estrogen. Endocrinology. 148:2764–2777. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Feil S, Valtcheva N and Feil R: Inducible
Cre mice. Methods Mol Biol. 530:343–363. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Rondón-Lagos M, Rangel N, Di Cantogno LV,
Annaratone L, Castellano I, Russo R, Manetta T, Marchiò C and
Sapino A: Effect of low doses of estradiol and tamoxifen on breast
cancer cell karyotypes. Endocr Relat Cancer. 23:635–650. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Odia Y, Kreisl TN, Aregawi D, Innis EK and
Fine HA: A phase II trial of tamoxifen and bortezomib in patients
with recurrent malignant gliomas. J Neurooncol. 125:191–195. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Lien EA, Solheim E and Ueland PM:
Distribution of tamoxifen and its metabolites in rat and human
tissues during steady-state treatment. Cancer Res. 51:4837–4844.
1991.PubMed/NCBI
|
|
53
|
Novick AM, Scott AT, Neill Epperson C and
Schneck CD: Neuropsychiatric effects of tamoxifen: Challenges and
opportunities. Front Neuroendocrinol. 59:1008692020. View Article : Google Scholar : PubMed/NCBI
|