With an ever-increasing aging population, the
incidence of cancer is rising. Malignant tumors have high mortality
rates and severely endanger human health. The drug resistance and
metastasis of malignant tumors are the primary causes of mortality
among cancer patients and are the major obstacles to cancer
treatment at present (1). The
initial solution to the problem of resistance to single-agent
chemotherapy was a combination of drugs with non-overlapping
mechanisms of action or combination chemotherapy. After almost 50
years of the application of combination chemotherapeutic regimens,
the role of combination chemotherapy has largely stalled. Surgery,
radiotherapy and combination chemotherapy are not sufficient to
cure multiple tumors (2).
Targeted therapies and immunotherapies represent a major
advancement and breakthrough in the field of chemotherapy. However,
studies have demonstrated that despite the notable efficacy of
targeted therapies and immunotherapy in the treatment of tumors,
drug resistance still emerges (3-5).
Drug resistance remains a major limiting factor in achieving a cure
for patients with cancer (6).
That is, overcoming multidrug resistance (MDR) in patients with
cancer remains a significant challenge.
MDR occurs in tumor cells during chemotherapy, and
is a phenomenon in which tumor cells become resistant to one
chemotherapeutic drug and then become resistant to other unexposed
chemotherapeutic drugs of different origins, structures and
mechanisms of action. Tumor MDR is mediated by multiple genes and
factors, the mechanisms of which are complex and diverse. The MDR
phenotype is characterized by cross-resistance to multiple
anticancer drugs with different structures and mechanisms of
action. The multiple factors mediating MDR may include host
factors, tumor factors as well as tumor-host interactions (7). More accurately, tumor cells develop
MDR during chemotherapy. MDR can occur as both primary and acquired
resistance, with the former being the phenomenon of insensitivity
when first exposed to the drug, while acquired resistance refers to
the phenomenon of a tumor being sensitive to the drug and only
becoming insensitive following a period of treatment with
chemotherapy (8). The acquisition
of drug resistance is mediated by mutations, changes in gene
expression, selective splicing and post-translational protein
modifications, etc. (9). Acquired
drug resistance is a common type of MDR. The most common mechanism
of drug resistance relies on drug efflux from cancer cells mediated
by ATP-binding cassette (ABC) transport proteins (10). Non-transport protein mechanisms,
such as the persistence of cancer stem cells (CSCs) (11), interactions of the tumor
microenvironment and epithelial-mesenchymal transition (12), enhanced DNA damage repair
(13), epigenetic alterations
(14), and the suppression of
apoptosis (15) are also common
causes of treatment failure.
A large number of discovered drugs and methods have
the potential to reverse or antagonize resistance to MDR by various
resistance mechanisms. These reversal agents include both
chemically synthesized small molecules and large polymers, as well
as natural products such as traditional Chinese medicines (TCMs)
(16). Inhibitors studied for
P-glycoprotein (ABCB1) include verapamil, dexverapamil, zosuquidar
and elaquidar amongst others (17,18), although with limited efficacy.
TCMs, including compound preparations and the screening of natural
active ingredients with resistance to MDR, primarily including
flavonoids (19,20), saponins (21,22) and alkaloids (23) have exhibited some success in
overcoming tumor drug resistance, but have not been widely adopted.
Nanodrug delivery systems are discoveries in technology for
overcoming MDR. Nanoparticles (NPs) can enhance the therapeutic
efficacy of anticancer drugs at the target site of action, which
can limit the adverse systemic effects of chemotherapeutic drugs
and reduce drug resistance. In addition, other innovative
strategies include RNA interference as a biological process used to
inhibit or knockdown the expression of specific genes, natural
products such as MDR modulators with minimal systemic toxic
effects, interference with the function of proteins involved in
drug efflux, and physical approaches such as combinations also
highlighting conventional drug administration using thermal,
ultrasound and photodynamic strategies (24). Due to their tumor-targeting
ability, NPs are a novel means of enhancing the therapeutic effects
of anticancer drugs at the target site of action, which can limit
the adverse systemic effects of chemotherapeutic drugs and reduce
drug resistance. NPs that allow multi-target and multi-pathway
combination therapies, the delivery of a stable dosage, or easy
utilization by the organism are considered promising means of
overcoming drug resistance and are under further investigation.
High mobility group box 1 (HMGB1) has been shown to
be involved in inflammation, angiogenesis, DNA repair, tumor
invasion, progression, metastasis and resistance to treatment
(25-27). In the present review, the
association between HMGB1 and MDR is described with an emphasis on
MDR mechanisms, in an aim to provide a theoretical foundation for
exploring novel strategies with which to overcome drug
resistance.
HMGB1 is a non-histone chromatin-associated protein
that is widely distributed in eukaryotic cells and consists of 215
amino acids, including the DNA-binding region A-box (amino acid
sequence 1-79), B-box (amino acid sequence 89-163) and the
hydroxyl-terminal domain (C-tail) (amino acid sequence 186-215).
Residues 7-74 bind to the suppressor gene p53 to regulate the
transcription of target genes; residues 89-108 bind to Toll-like
receptor (TLR)4 to promote the inflammatory response of the body;
and residues 150-183 bind to the receptor for advanced glycation
end products (RAGE) to promote cell migration. The overexpression
of HMGB1 in cells with C-tail deletion can affect the
transcription, translation and expression of downstream genes
(45), through the regulation of
DNA damage repair and genome stability maintenance (46). HMGB1 is active both
intracellularly and extracellularly and the biological functions of
the pro- and antitumor effects of HMGB1 are closely related to its
expression levels and subcellular distribution. In the nucleus,
HMGB1 can function as a DNA molecular chaperone to participate in
DNA replication, binding and damage repair, maintaining nuclear
homeostasis and exerting antitumor effects through a series of
regulatory transcriptional pathways. In the cytoplasm, HMGB1
participates in the immune response by increasing autophagy,
inhibiting apoptosis and regulating mitochondrial function
(47). HMGB1 is secreted
extracellularly as an immune signal; through interactions with
ligand-receptors, extracellular HMGB1 functions as a typical
damage-associated molecular pattern (DAMP) that functions as a
cytokine directly or indirectly on TLRs, the major receptors of
HMGB1, and on RAGE, thereby regulating immune cells and specific
signaling pathways in the tumor microenvironment, including
inflammation, proliferation, metastasis, autophagy and specific
signaling pathways (such as RAGE-PI3K/AKT, HMGB1/RAGE/IL-8 pathway,
etc.) (41,46,48-50). The aberrant expression of HMGB1
can function as a double-edged sword, exerting both pro- and
antitumor effects during tumorigenesis and development.
HMGB1 is involved in tumorigenesis and development.
A high expression of HMGB1 has been detected in several types of
tumors, suggesting that HMGB1 is closely associated with tumor
progression. With the aid of the GEPIA database (http://GEPIA.cancer-pku.cn), HMGB1 has been found to
be highly expressed in colon adenocarcinoma, diffuse large B cell
lymphoma, glioblastoma (GBM), low-grade glioma, pancreatic
adenocarcinoma, rectum adenocarcinoma, stomach adenocarcinoma and
thymoma. HMGB1 has been found to be involved in a variety of
malignancies, including liver cancer (37,51,52), breast cancer (50,53), multiple myeloma (MM) (25), thyroid cance (38), rectal cancer (54), cutaneous melanoma (55), lung cancer (56), gastric cancer (57) and leukemia (58). In studies on hepatocellular
carcinoma (HCC), HMGB1 and the RICTOR 3′untranslated region (UTR)
epigenetically promote malignant proliferation, self-renewal and
tumorigenesis (37), and
hypoxia-induced HMGB1 and mitochondrial DNA mediate tumor growth
via TLRs (59). In breast cancer,
HMGB1 expression levels have been shown to be associated with
sensitivity to chemotherapy, with a higher HMGB1 expression in
adriamycin-resistant breast cancer cells than in parental cells
(60). HMGB1 has also been found
to activate fibroblast-promoted breast cancer cell metastasis
through RAGE/aerobic glycolysis (61). Lee et al (62) found that cytoplasmic HMGB1
expression was associated with the levels of tumor-infiltrating
lymphocytes, and it has also been found to be a key factor in
tamoxifen treatment resistance (53). HMGB1 and RAGE may be used to
predict and assess the efficacy of breast cancer treatment
(62). It has been suggested that
HMGB1 is a key regulator of hematopoietic malignancies and that the
dysfunction of HMGB1 may contribute to the development of
hematological malignancies by interfering with the hematopoietic
function of bone marrow (63). MM
cell lines and primary MM samples express high levels of HMGB1,
which is negatively associated with the 3-year survival of patients
with MM (25). During the
development of acute myeloid leukemia (AML), HMGB1 is secreted to
induce the production of TNF-α and subsequently, IL-1β, which
stimulates the release of stem cell factor from endothelial cells,
and also induces the secretion of angiogenic vascular endothelial
growth factor (VEGF), which further promotes the proliferation of
AML cells (64). Another study
found that HMGB1 induced cell proliferation and myeloid
differentiation blockade, inhibited the apoptosis of AML cells, and
that chidamide, a selective histone deacetylase inhibitor,
exhibited good therapeutic efficacy in AML by downregulating HMGB1
expression (65). In chronic
myeloid leukemia (CML), cytoplasmic HMGB1 has been found to reduce
the sensitivity of CML cells to anticancer drugs by upregulating
the autophagic pathway. HMGB1 overexpression increases the
transcriptional activity of JNK, ERK and Beclin-1 to regulate
autophagy (66).
The upregulation of HMGB1 is commonly observed in
thyroid cancer tissues and is strongly associated with worse
overall lymph node metastasis and clinical staging. HMGB1 knockdown
significantly inhibits autophagy, sodium/iodide symporter (NIS)
degradation and iodine uptake in Hank's balanced salt solution
(HBSS)-treated cells. In addition, HBSS enhances reactive oxygen
species (ROS)-maintained autophagy and promotes the cytoplasmic
translocation of HMGB1. HMGB1 knockdown inhibits LC3-II
transactivation and NIS degradation via AMP-activated protein
kinase (AMPK)/mTOR-dependent signaling pathways by regulating ROS
production, but not adenosine triphosphate (ATP) (38), and HMGB1 knockdown has been shown
to inhibit cell migration and invasion (67). The nuclear expression of HMGB1 has
been found to be significantly higher in colorectal cancer (CRC)
and colorectal adenoma tissue samples (84.0 and 92.6%,
respectively) than in normal colorectal tissue (15.0%), and a
positive cytoplasmic expression of HMGB1 is significantly higher in
CRC tissues (25.2%) compared with colorectal adenoma (11.8%) and
normal colorectal tissue (0.0%). A positive cytoplasmic expression
of HMGB1 is associated with high-grade CRC and a poor prognosis,
and negatively associated with a strong positive nuclear HMGB1
expression in CRC tissue specimens. Patients with CRC with a strong
positive nuclear HMGB1 expression exhibited an improved prognosis
compared with other patients with CRC (68). HMGB1 is involved in the
tumorigenesis of colitis-associated CRC via the ERK1/2 pathway
(54). Furthermore, HMGB1
exposure has been shown to lead to CRC cell proliferation and CRC
stem cell expansion (69). The
low expression of HMGB1 inhibits cell proliferation and migration
in CRC (70). In lung cancer, the
overexpression of HMGB1 has been shown to promote lung cancer
invasion and metastasis by promoting matrix metalloproteinase
(MMP)-2 mediated-invasion and metastasis through an NF-κB-dependent
mechanism; thus, HMGB1 is a potential prognostic marker for lung
cancer (71). HMGB1 expression
has also been shown to be upregulated in gastric cancer tissues,
and an increased HMGB1 expression is associated with a poorer
prognosis of patients with gastric cancer (57). Gastric cancer cell-derived
exosomes induce autophagy and pro-tumor activation via
HMGB1/TLR4/NF-κB signaling, and induce neutrophil autophagy and
pro-tumor activation (57). RAGE
and HMGB1 may regulate polypyrimidine tract-binding protein 1
expression and inhibit cellular glycolysis, thereby affecting
gastric cancer cell proliferation and migration (72). Taken together, HMGB1 is closely
related to the development and progression of a variety of
tumors.
In a review summary study on the biological
functions and therapeutic potential of HMGB family members in human
cancer (32), several key points
of the role of HMGB1 in carcinogenesis were summarized, including
two primary aspects: Angiogenesis and migration. HMGB1 promotes the
expression of neurofibrillin-1, VEGFA, VEGF receptors-1 and -2 and
platelet-derived growth factor (73), which induces angiogenesis, and
also binds to RAGE to activate NF-κB (74). In terms of tumor migration, HMGB1
binds to RAGE and mediates epithelial-mesenchymal transition (EMT)
via MMP-7, p-NF-κB and Snail (75); HMGB1 binds to RAGE and mediates
EMT via NF-κB, p65, inducible nitric oxide synthase and MMP-9
production, and the phosphorylation of Rac-1, ERK1 and AKT
(76); HMGB1 binds to TLR4 and
upregulates downstream signaling and PI3K signaling pathways
(77); secreted HMGB1 targets
other stromal cells and release factors, which leads to the
hydrolytic degradation of ECM proteins (78). In addition, a number of microRNAs
(miRNAs/miRs), such as miR-200-c (79) and miR-325-3p (80) inhibit HMGB1 expression and
translocation. In summary, HMGB1 is involved in the development of
a variety of tumors by regulating tumorigenesis via several
mechanisms.
A study on the role of the induction of secretory
clusterin (sCLU) by HMGB1 in chemoresistance in human prostate
tumor cells found that a variety of chemotherapeutic agents
currently in use can result in the release of HMGB1 from tumor
cells (81). The acquisition of
chemoresistance may be achieved through the regulation of an
HMGB1/TLR4-RAGE/sCLU axis triggered by dead cells, thus providing a
survival advantage to residual living tumor cells. That study
suggested a link between HMGB1 and the development of drug
resistance in tumor cells. Cell death is the outcome of successful
chemotherapeutic action (82).
The authors then found numerous studies (as described below) to
confirm that HMGB1 is associated with apoptosis, necroptosis,
autophagy, pyroptosis and ferroptosis in tumors.
One of the well-known mechanisms of chemoresistance
is the mitochondrial death pathway, which is activated when cells
are exposed to increased levels of stress, such as during treatment
with chemotherapy. Mitochondrial autophagy or mitochondrial
selective autophagy is essential for cell quality regulation, as it
effectively breaks down, removes and recycles defective or damaged
mitochondria. Cancer cells use mitophagy to rapidly remove damaged
mitochondria to mediate their drug resistance, which affects the
efficacy of tumor chemotherapy and the degree of drug resistance
(83). The activation of
autophagy by chemotherapeutic agents contributes, at least in part,
to the development of MDR (84).
Therefore, the combination of chemotherapeutic drugs with
appropriate autophagy inhibitors has been proposed as an effective
method with which to reverse the transformation of tumor cells from
chemotherapy-resistant to sensitive. Chloroquine is an effective
autophagy blocker that has been shown to enhance chemotherapy
sensitivity in cancer (85).
Apoptosis is a programmed cell death (PCD) process
that usually occurs through one of two pathways, the endogenous and
exogenous pathways. The intrinsic pathway of apoptosis involves the
mitochondria and is activated by a decrease in mitochondrial
membrane potential, the release of cytochrome c from the
mitochondria into the cytoplasm, and the activation of the
performer cysteine protease (86). Cancer chemotherapy generates
stress signals that act on mitochondria to initiate apoptosis.
Significantly reduced apoptosis in MDR tumor cells acquired in
children with relapsed neuroblastoma (87) suggests that
mitochondria-associated apoptosis is involved in the mechanism of
tumor MDR.
Both apoptosis (self-killing) and autophagy
(self-eating) are evolutionarily conserved processes whose
interactions affect anticancer drug sensitivity and cell death. The
inhibition of the PARP1/HMGB1 pathway reduces autophagy and
increases apoptosis. HMGB1 is a significant upstream regulator of
autophagy, maintaining a homeostatic balance between apoptosis and
autophagy (88). The interaction
between apoptosis and autophagy has been shown to affect anticancer
drug sensitivity and cell death in HCC cells and tumor models, and
HepG2 cells exhibit an enhanced sensitivity to cisplatin when HMGB1
inhibitors and autophagy inhibitors are used (89). The REDOX status of HMGB1 can
regulate the autophagy or apoptosis of tumor cells. The reduced
state of HMGB1 induces Beclin-1-dependent autophagy by acting on
the RAGE receptor, thereby promoting the resistance of colon cancer
cells to radiation and chemotherapy. By contrast, the oxidized
state of HMGB1 increases the cytotoxicity of the drug, thereby
inducing apoptosis via the mitochondrial pathway (90). Endogenous HMGB1 is a key
pro-autophagic protein that enhances cell survival and limits
programmed apoptotic cell death. HMGB1 induces resistance to
chemotherapy by upregulating autophagy through three different
mechanisms: Nuclear HMGB1 upregulating autophagy through HSPB1,
cytoplasmic HMGB1 binding Beclin-1 and extracellular HMGB1, leading
to advanced glycosylation end-product specific receptor
(AGER)-mediated class III PI3K activation (91,92).
HMGB1-mediated autophagy and apoptosis have been
demonstrated in several tumor MDR studies, by promoting autophagy
and inhibiting apoptosis induced by anticancer drugs, which has
been identified as a key mechanism in the development of MDR. In
liver cancer, the HMGB1-mediated activation of autophagy has been
found to be involved in the development of cisplatin resistance in
HepG2 cells (93). It has been
found that morin hydrate enhances the sensitivity of HCC cells and
xenografts to cisplatin chemotherapy by downregulating
PARP-1-HMGB1-mediated autophagy (89). In hepatoma cells, HepG2, HMGB1 and
p53 have been found to jointly provide a delicate balance between
autophagy and apoptosis (94). In
breast cancer, the inhibition of autophagy has been found to
increase paclitaxel-induced apoptosis and enhance the sensitivity
of MCF-7 cells to paclitaxel (95). HMGB1 is a target gene and
autophagy regulator of miR-129-5p, and interference with HMGB1
expression enhances the chemosensitivity of paclitaxel by
inhibiting autophagy and inducing the apoptosis of MCF-7 cells
(95). In a previous study, the
overexpression of HMGB1 in doxorubicin (DOX)-resistant MCF-7 breast
cancer cells significantly reversed apoptosis promotion and
autophagy inhibition mediated by miR-142-3p upregulation (60). Zhang et al (96) found that pediatric AML bone marrow
and cell lines exhibited a high expression of HMGB1, and miR-451
exerted tumor suppressive effects by targeting and modulating HMGB1
to enhance cell death and reduce autophagy in AML cells. In a study
on thyroid cancer, the higher expression of HMGB1 was observed in
vemurafenib-resistant thyroid cancer BCPAP-R cells (97). The overexpression of HMGB1
attenuated the sensitivity of BCPAP cells to vemurafenib by
increasing cell viability and decreasing apoptosis and caspase-3
activity. HMGB1 targeting inhibited vemurafenib-induced autophagy.
Blocking the autophagy pathway with the autophagy inhibitor,
3-methyladenine, or knocking out HMGB1, sensitized BCPAP-R cells to
vemurafenib (97). In a study on
skin cancer, autocrine HMGB1 was found to regulate autophagy via a
RAGE/HMGB1/ERK1/2-dependent pathway, thereby protecting
keratinocytes from apoptosis upon UV irradiation (98). GBM exhibits a high autophagic
activity, where YAP overexpression enhances autophagy in glioma
cells, and the downregulation of HMGB1 eliminates the effects of
YAP on autophagy and glioma growth, suggesting that YAP promotes
glioma progression by enhancing HMGB1-mediated autophagy (39). It has been shown that
HMGB1-inducible p62 overexpression promotes EMT in glioblastoma
cells (99). In their study on
the chemosensitization of rectal cancer, Liu et al (100) found that oxaliplatin
administration significantly enhanced the expression of HMGB1,
which regulated the autophagic response and negatively regulated
apoptosis. Following HMGB1 knockdown in rectal cancer cells, the
cells were treated with oxaliplatin, and the autophagy response
decreased, whereas apoptosis increased (100). In a study on pancreatic cancer,
triphorone inhibited autophagy and MDR1 expression in pancreatic
cancer cells through an HMGB1/RAGE/PI3K/Akt axis (101). The results of another study on
the resistance mechanisms of sunitinib, a multi-kinase inhibitor
approved for multiple cancer indications, suggested that HMGB1
controlled TP53 autophagic degradation through the regulation of
its nucleus-to-cytoplasm translocation, thereby inhibiting
sunitinib resistance (102).
In summary, HMGB1 functions as an immune signal and
can be considered an autophagy inducer, regulating various
autophagic pathways to inhibit or promote tumor progression. The
interaction between autophagy and apoptosis affects cell death and
anticancer drug sensitivity in various cancer cell lines, and
HMGB1-mediated autophagy and apoptosis are involved in MDR.
Ferroptosis is a newly discovered form of regulated
cell death that can be induced by small molecule compounds or
drugs. First proposed in 2012 by Dixon et al (103), ferroptosis is an iron-dependent
and peroxide-driven form of cell death associated with multiple
metabolic disorders and imbalances in body homeostasis. The primary
biochemical features of ferroptosis are iron accumulation and lipid
peroxidation. Glutathione peroxidase 4 (GPX4) activity prevents the
accumulation of lipid ROS, and ferroptosis can be triggered by the
inhibition of glutathione (GSH) or GPX4 biosynthesis (104). Ferrous ions generate large
quantities of ROS through the Fenton reaction, leading to lipid
peroxidation. If GPX4 activity decreases, the cellular antioxidant
capacity is reduced and the excessive accumulation of ROS is not
removed in a timely manner, which leads to oxidative stress,
causing oxidative damage to the cell and consequently ferroptosis
(105).
It has been shown that ferroptosis is involved in a
variety of diseases, including neurodegenerative diseases (such as
Alzheimer's disease and Parkinson's disease, amongst others),
strokes, tumors and other diseases, and that ferroptosis plays a
crucial role in the development and progression of these diseases
(106). Compared to normal
cells, cancer cells are more iron-dependent than normal cells for
maintaining cell proliferation and promoting cell expansion, and
this increased need for iron renders cancer cells more susceptible
to ferroptosis (107). A review
article on studies using iron death as a therapeutic target in
oncological disease focused on two primary areas for intervention:
On the one hand, the use of ferroptosis inhibitors (primarily
ferrostatin-1, liproxstatin-1, vitamin E and iron chelators) and
the upregulation of GPX4 are effective in inhibiting ferroptosis to
prevent the accumulation of lipid ROS, attenuating progression and
improving clinical symptoms; on the other hand, the inhibition or
promotion of GPX4 degradation, as well as the reduction of coenzyme
Q10, achieved through peroxide, iron, or polyunsaturated fatty acid
overload, and the induction of lipid peroxidation, improve the
clearance of specific cell populations (e.g., specific tumor types)
(108). In cancer cells,
metabolic rates, the levels of ROS and iron content are higher than
those in normal cells (109).
Tumor cells usually contain large quantities of
H2O2, and ferrous/iron ions react with
excessive H2O2 to produce hydroxyl radicals
in the cells and induce ferroptosis in tumor cells (110). Based on these features,
ferroptosis in cancer cells can inhibit tumor growth (106).
Ferroptosis is associated with HMGB1 in disorders,
including inflammation and neoplasia. Extracellular HMGB1 generally
functions as a DAMP molecule following active secretion or passive
release, controlling inflammatory and immunological responses via
various receptors or direct absorption. A variety of variables
influence HMGB1 secretion and release, including post-translational
changes (acetylation, ADP-ribosylation, phosphorylation and
methylation) and PCD (apoptosis, pyroptosis, necrosis, alkalosis
and ferroptosis) (111).
Ferroptosis activators induce the release of HMGB1, and autophagy
promotes the acetylation of HMGB1 in ferroptosis cells to promote
the release of HMGB1. HMGB1 mediates the inflammatory response in
ferroptosis through the HMGB1/AGER pathway. HMGB1 inhibition or
AGER deficiency attenuates the iron-dependent cell-induced
inflammatory response in macrophages, suggesting that targeting
HMGB1 release can limit the iron inflammatory response during
ferroptosis (112). By
constructing a model of induced ferroptosis, three stages of iron
death have been identified: i) The 'initial' stage related to lipid
peroxidation; ii) the 'intermediate' stage related to ATP release;
and iii) the 'terminal' stage is characterized by the release of
HMGB1 and the loss of plasma membrane integrity (113). In another study, HMGB1 was shown
to be passively released during ferroptosis, where it serves as a
signal associated with early ferroptosis immunogenicity in cancer
cells (114). In a study on skin
inflammation induced by ferroptosis in keratinocytes following UV-B
radiation, the inhibition of ferroptosis prevented the release of
HMGB1 from human epidermal keratinocytes and blocked
necroinflammation in the skin of UV-B radiation-exposed mice
(115). In another study on the
role and mechanisms of dexamethasone in adriamycin-induced
ferroptosis and cardiomyopathy in rats, molecular experiments were
performed to detect ferroptosis in adriamycin-treated rats with the
downregulation or overexpression of the HMGB1 gene (116). The overexpression of HMGB1
promoted adriamycin-induced ferroptosis and cardiotoxicity in rats,
while the silencing of HMGB1 resulted in the opposite effect.
Dexamethasone had a protective effect on ferroptosis and
cardiomyopathy in rats by regulating HMGB1 (116).
It has been found that HMGB1 and ferroptosis are
associated with tumor MDR, and its drug-resistance mechanism has
emerged as a possible therapeutic target. Friedman Angeli et
al (117) argued that
ferroptosis is present at the crossroads of cancer-acquired
resistance and immune evasion, that ferroptosis may act as a
double-edged sword in tumors, and that it is critical to identify
the difference between ferroptosis that inhibits tumor growth and
ferroptosis that drives cancer progression. Metabolic
reorganization is critical for the persistence, dedifferentiation
and expansion of cancer cells, and in certain cases, this metabolic
reorganization is associated with acquired ferroptosis. The
ferroptosis-sensitive state is what allows cancer cells to produce
lipid-derived mediators that regulate intracellular and
intercellular signaling pathways, including receptor tyrosine
kinase (RTK) signaling, leading to cancer cell growth. Furthermore,
the complex interactions between different lipid oxidases (such as
LOXs and PTGS2) highlight novel possibilities to bind and regulate
them at the tumor site, thus allowing an effective immune response
and enhancing the immunogenicity of ferrophilic cells. A study on
leukemia found that HMGB1 plays a critical role in both leukemia
pathogenesis and chemotherapy resistance. Ferroptosis agonists
sensitize chemotherapy by enhancing ROS levels, thereby promoting
the cytoplasmic translocation of HMGB1 and enhancing ferroptosis,
and HMGB1 is a novel modulator of iron death through the
RAS-JNK/p38 pathway and a potential drug target for therapeutic
intervention in patients with leukemia (58). Lipocalin 2 is a
siderophore-binding protein that regulates iron homeostasis.
Lipocalin 2 can promote tumor growth and chemoresistance by
inhibiting ferroptosis in CRC. In a previous study, the inhibition
of Lipocalin 2 function by monoclonal antibodies reduced
chemoresistance in the xenograft mouse model (118). In another study, cell death
significantly increased in paclitaxel-resistant persistent cancer
cells compared to parental cells when exposed to ferroptosis
inducers inhibiting xCT: Erastin, sulfasalazine and cyst(e)ine
deprivation (119). Ferroptosis
is relatively insensitive in cisplatin-resistant head and neck
cancer cells, and ferroptosis agonists are expected to exhibit a
chemosensitizing effect (120).
In summary, ferroptosis as a new mode of PCD, is
closely related to tumor pathogenesis and drug resistance
mechanisms. The release of HMGBI is closely related to ferroptosis.
The interaction mechanism between HMGB1 and ferroptosis is a
potential therapeutic target for tumor therapy, which is worthy of
further investigations.
Pyroptosis is a unique form of PCD characterized by
DNA fragmentation, chromatin condensation, cell swelling with the
appearance of large bubbles and the leakage of cell contents,
accompanied by inflammation and membrane disruption (121). Pyroptosis is divided into a
typical pathway triggered by caspase-1 and an atypical pathway
independent of caspase-2. In general, the pyroptosis process can be
divided into four main stages: i) The capture of the stimulus
signal; ii) transmission of the stimulus signal; iii) the
activation of the pyroptosis actuator; and iv) the execution of
pyroptosis. Pyroptosis plays a crucial role in cancer initiation,
progression and metastasis, and it also influences immunotherapy
outcomes by affecting immune cell infiltration (122).
Pyroptosis is related to the sensitivity of tumor
chemotherapy in liver cancer, gastric cancer, non-small cell lung
cancer (NSCLC) and rectal cancer. Sorafenib is a kinase inhibitor
with direct effects on cancer cells and angiogenesis, inducing the
pyroptosis of macrophages and releasing pro-inflammatory cytokines.
Natural killer cells are then activated to ultimately eliminate HCC
cells (123). As previously
demonstrated, lipopolysaccharide (LPS) from Gram-negative bacterial
outer membranes enhances sensitivity to oxaliplatin in rectal
cancer by inducing the gasdermin D (GSDMD)-mediated pyroptosis of
HT29 cells (124). The
camptothecin analog, FL118, has been shown to inhibit rectal cancer
growth and metastasis by inducing NLR family pyrin domain
containing 3 (NLRP3)/caspase-1-mediated pyroptosis (125). It has been shown that gasdermin
E (GSDME) mediates the lobaplatin-induced pyroptosis downstream of
ROS/JNK/Bax-mitochondrial apoptosis pathway and caspase-3/-9
activation in colon cancer cells (126). The activation of the NLRP3
inflammasome and caspase-1 by simvastatin can stimulate pyroptosis
activation through the canonical pathway and inhibit the migration
of NSCLC (127). It is
hypothesized that pyroptosis plays a 'double-edged sword' role in
tumors. On the one hand, pyroptosis alters the tumor
microenvironment and inhibits tumor growth by releasing
inflammatory factors, such as IL-1β and IL-18 (128). On the other hand, pyroptosis
reduces the immune response of the body to tumor cells and
accelerates the growth rate of different types of cancers (129). In the treatment of malignant
tumors, appropriate chemotherapeutic drugs can be selected
according to the expression levels of DFNA5/GSDME, such that it can
be upregulated in tumor cells, thereby increasing the sensitivity
to chemotherapeutic drugs and reducing drug resistance. Thus,
induced pyroptosis may play a major role in the treatment of cancer
(130). It is important to
determine the balance between the induction or inhibition of
pyroptosis and the precise improvement of chemotherapy sensitivity
in different types of cancer.
HMGBI interacts with pyroptosis and is involved in
the pathogenesis of several diseases. GSDMD indirectly regulates
the release of HMGB1 in cases where HMGB1 is not released when
pyroptosis is suppressed, inflammatory vesicle activation and IL-1β
secretion (131). The depletion
of hepatocyte HMGB1, the inhibition of hepatocyte HMGB1 release,
and the neutralization of extracellular HMGB1 or RAGE deficiency
prevent caspase-11-dependent pyroptosis and death from endotoxemia
and bacterial sepsis (132).
These findings suggest that HMGB1 interacts with LPS to mediate
caspase-11-dependent focal death in lethal sepsis (132). As previously demonstrated, in a
rat model of chronic liver failure aggravation, the percentage of
hepatocyte pyroptotic cells was significantly increased, and the
expression levels of pyroptosis-related genes and proteins in liver
tissue and serum were significantly increased (133). However, these phenomena were
attenuated by the inhibition of HMGB1, and the dual inhibition of
HMGB1 and caspase-1 exerted a more potent effect. GSDME-mediated
cellular pyroptosis promotes CRC development through the release of
HMGB1. Previous in vivo experiments using mice demonstrated
that the number and size of tumors in mice were reduced by
interfering with HMGB1, and in vitro experiments also
revealed that HMGB1 induced CT26 colon cancer cell proliferation
and proliferating cell nuclear antigen expression through the
ERK1/2 pathway (54).
The search for solutions to tumor chemoresistance by
intervening in HMGB1 and pyroptosis has identified partially
positive outcomes. GSDME-enhanced cisplatin sensitivity in NSCLC
(134). miR-556-5p has been
found to be significantly upregulated in tumor tissues of patients
with cisplatin-resistant NSCLC, and its knockdown inhibits tumor
cell viability and induced pyroptosis, while miR-556-5p
downregulation induces NLRP3-mediated cell pyroptosis, which
effectively enhances cisplatin sensitivity in NSCLC, providing a
novel therapeutic strategy to overcome resistance to chemotherapy
in patients with NSCLC (135).
In a previous study on the pyroptosis regulation of the tumor
immune microenvironment, the expression of pyroapoptotic gene sets
was positively associated with the percentage of tumor response to
target therapy, and the pyroptosis signal upregulated during the
process of tumor drug resistance (136). The release of HMGB1 and GSDME
cleavage were inhibited in melanoma BRAFi + MEKi-resistant cells,
and targeting this PCD pathway is a potential strategy for salvage
therapy in patients with BRAFi + MEKi-resistant melanoma (136).
In conclusion, pyroptosis is a form of inflammatory
necrosis of cells triggered by inflammasomes, which are present in
a variety of tumor cells. HMGB1 can affect tumor proliferation,
invasion, metastasis, and MDR through pyroptosis. Therefore, HMGB1
is a promising target therapeutic target for overcoming MDR,
highlighting a novel basis for the diagnosis and treatment of
cancer and the development of new drugs.
ncRNAs are RNAs that do not code for proteins and
include miRNAs, long ncRNAs (lncRNAs), circurlar RNAs (circRNAs),
small nuclear RNAs (snRNAs); miRNAs, lncRNAs and circRNAs are
widely recognized as universal regulators of a variety of cancer
features, such as proliferation, apoptosis, invasion, metastasis
and genomic instability. ncRNAs also play crucial roles in
resisting different cancer treatments by rewiring important
signaling pathways (137). RNAs
in the circulatory system are primarily secreted by cells and can
be indicative of disease and biological processes, including
response to therapy. Exploiting the role of certain ncRNAs in
intrinsic and acquired therapeutic resistance high-lights their
value as biomarkers that can predict the outcome of a particular
patient before, during, and/or following treatment. This
association relies in part on the properties of ncRNAs that play a
role in intercellular communication (138-140).
miRNAs regulate gene expression at the
transcriptional level by binding to the mRNA 3′UTR region of target
genes and inhibiting their translation through degradation of the
mRNA (141). miRNAs in tumors
can be divided into oncogenic and suppressor miRNAs. Oncogenic
miRNAs suppress their target genes through overexpression, leading
to the occurrence of tumors, while suppressor miRNAs can also
induce tumorigenesis when their target genes (oncogenes) are
overexpressed when their expression is decreased or absent. Several
miRNAs are involved in drug resistance in cancer therapy through
complex underlying regulatory mechanisms, and knowledge of these
may assist clinicians in tailoring treatments and thus improve a
patient's prognosis (142).
Research on bladder cancer (BC) has recently identified miRNAs as
potential regulators of the oncogenic potential of BC cells
(143). A variety of miRNAs,
including miR-23a-3p and miR-141-3p, were found to be abnormally
expressed in BC tissues, and in the peripheral, blood, or urine
samples from patients with BC. These miRNAs promote the oncogenic
potential of BC by regulating epithelial-mesenchymal transition or
PI3K/AKT, JAK/STAT and NF-κB/Snail signaling pathways (143). In addition, several tumor
suppressor miRNAs were found to be downregulated in BC samples,
leading to the enhanced proliferation, invasion and metastasis of
these cells (143). miRNAs are
also involved in regulating the response of cancer cells to
chemotherapeutic agents. The role of miR-193a-3p in enhancing
multi-chemoresistance in BC cells was identified in BC treatment;
the miR-193a-3p/LOXL4 axis was found to be a possible target for
enhancing the response of BC tissues to chemotherapy (144). The upregulation of miR-204-5p
and miR-211-5p following treatment with vemurafenib in melanoma
leads to the emergence of drug resistance (145). This indicates that miRNAs are
associated with tumor development, drug resistance and the
assessment of therapeutic efficacy.
circRNAs are single-stranded ncRNAs with an
increased stability and a longer half-life compared to linear RNAs
due to the secondary closed-loop structure of the phosphodiester
bond between the 3′ and 5′ ends and the absence of the 3′
polyadenosine tail (152).
circRNAs can play oncogenic or tumor suppressor roles in a variety
of cancer types and affect cancer phenotypes via a variety of
mechanisms. Singh et al (153) found that circRNAs could be used
as diagnostic and prognostic biomarkers for AML. circRNAs regulate
gene expression and various steps of leukemia development and
progression, including differentiation, proliferation, cell cycle
transition, adhesion and apoptosis. A previous study on gastric
cancer found that circDLG1 was significantly upregulated in distant
metastatic lesions and in anti-programmed cell death protein 1
(PD-1)-treated gastric cancer tissue, and was associated with an
aggressive tumor phenotype and a poorer prognosis of patients with
gastric cancer treated with anti-PD-1 (154). The ectopic expression of
circDLG1 promoted the proliferation, migration, invasion and immune
escape of gastric cancer cells. Mechanistically, circDLG1
interacted with miR-141-3p, acting as a miRNA sponge to increase
CXCL12 expression, thereby promoting gastric cancer progression and
resistance to anti-PD-1-based therapy (154). In a study on HCC, it was found
that the knockdown of circMRPS35 expression in HCC cells inhibited
proliferation, migration, invasion, colony formation and cell cycle
progression in vitro, and tumor growth in vivo
(155). Furthermore, a peptide
encoded by circMRPS35 (circMRPS35-168aa) was significantly induced
by chemotherapeutic agents and promoted cisplatin resistance in HCC
(155).
The regulation of MHGB1 expression by ncRNAs is
frequently observed in studies to overcome chemoresistance in
tumors. For example, miR-451 was found to play an oncogenic role in
pediatric AML (96). Pediatric
AML bone marrow and cell lines exhibited a low miR-451 expression
and a high HMGB1 expression, and HMGB1 was identified as a
functional target of miR-451. miR-451 overexpression significantly
enhanced apoptosis and reduced autophagy in both AML cell lines,
which was reversed by pcDNA-HMGB1 transfection (96). In addition, exogenous miR-451
significantly enhanced the sensitivity of HL-60 cells to
chemotherapeutic agents. miR-451 played a tumor suppressor role by
enhancing cell death and reducing autophagy in AML cells by
targeting HMGB1 (96). In another
study on leukemia, the upregulation of miR-142-3p was found to
improve drug sensitivity in AML by reducing P-glycoprotein and
inhibiting autophagy by targeting HMGB1 (156). Similarly, Liang et al
(60) demonstrated that
miR-142-3p was significantly downregulated in DOX-resistant breast
cancer cell lines, and that miR-142-3p negatively regulated HMGB1
expression. Furthermore, the overexpression of HMGB1 significantly
reversed the increase in apoptosis and inhibition of autophagy
mediated by miR-142-3p upregulation (60). miR-142-3p overexpression may
inhibit autophagy and promote drug sensitivity to DOX in breast
cancer cells by targeting HMGB1. The miR-142-3p/HMGB1 axis may thus
serve as a novel target for regulating drug resistance in breast
cancer patients (60). The most
common type of thyroid cancer is papillary thyroid carcinoma (PTC)
(157). The overexpression of
miR-let-7e or the knockdown of HMGB1 has been found to inhibit cell
migration and invasion (67).
miR-let-7e has been shown to downregulate HMGB1 expression by
directly targeting the HMGB1 3′-UTR. In addition, the
reintroduction of HMGB1 has been found to reversed the
anti-proliferative, anti-migratory and anti-invasive effects of
miR-let-7e. miR-let-7e may thus exert tumor suppressive effects via
HMGB1 in PTC (67). The RNA
interference-mediated knockdown of HMGB1 reduces lung cancer cell
viability, induced apoptosis and partially restores sensitivity to
gefitinib (158).
In summary, ncRNAs have a wide impact on tumor
development, treatment, drug resistance, and efficacy assessment.
ncRNAs can be transmitted between cells as cellular signals,
including ncRNAs which imbue drug resistance. ncRNAs are often
specific for diagnosis, and there is a significant potential that
ncRNAs can be used as biomarkers for clinical applications. Given
the widespread involvement of ncRNAs in tumor and chemotherapy
resistance, the prospect of different ncRNA intervention strategies
to inhibit tumor growth and overcome drug resistance is
promising.
In a previous study, when mouse bone marrow
mesenchymal stem cells pre-treated with oxidized HMGB1 were
co-cultured with homologous cancer cells, the cell proliferation
and stemness of the cancer cells increased, and tumorigenesis and
drug resistance increased (159). It has been found that
extracellular HMGB1 promotes the release of cytokines, such as IL-6
and IL-8 through the activation of MAPK and via MyD88-dependent
NF-κB pathways, which in turn stimulates tumor cell proliferation,
angiogenesis, EMT, invasion and metastasis. Nuclear and cytoplasmic
HMGB1 promotes phagocytosis and inhibits tumor cell apoptosis to
induce chemoresistance (63).
HMGB1 is involved in the tumorigenesis of colitis-associated CRC
via the ERK1/2 pathway (54).
HMGB1-mediated autophagy regulates sodium/iodine transporter
degradation in thyroid cancer cells, and the knockdown of HMGB1
inhibits LC3-II transactivation and sodium/iodine cotransporter
degradation through AMPK/mTOR-dependent signaling pathways by
regulating ROS production, but not ATP (38). Zha et al (41) found that neutrophil extracellular
traps produced tumor-infiltrating neutrophils-mediated crosstalk
between glioma progression and the tumor microenvironment by
regulating the HMGB1/RAGE/IL-8 axis. The autophagic secretion of
HMGB1 by cancer-associated fibroblasts promotes metastasis of NSCLC
cells via NF-κB signaling (160). Cancer-associated fibroblast
autophagy secretes HMGB1 to promote the metastasis of NSCLC cells
via NF-κB signaling (160).
HMGB1 regulates NF-κB in MM; that is, HMGB1 regulates drug
resistance in MM cells by modulating the NF-κB signaling pathway
(161). HMGB1 and the PI3K/Akt
signaling pathways are involved in tumor chemotherapy sensitization
research, and gemcitabine resistance in pancreatic ductal
adenocarcinoma involves HMGB1/RAGE-initiated PI3K/Akt/MDR1
signaling (101). In breast
cancer, HMGB1/RAGE has been found to promote not only breast cancer
cell invasion, but also PD-L1 expression through PI3K/AKT
signaling, leading to the destruction of effector T-cells (50). In gastric cancer, the
HMGB1-mediated activation of the PI3K/Akt/HIF-1α signaling pathway
is associated with gastric cancer growth and metastasis (162). As observed in HCC, the
interaction of VCP with HMGB1 promotes HCC cell proliferation,
migration and invasion through the activation of the PI3K/AKT/mTOR
pathway (163). In esophageal
squamous cell carcinoma, patients with an increased expression of
HMGB1 and p-ATM have been shown to have a poorer prognosis
following radiotherapy; the downregulation of HMGB1 may promote the
radiosensitivity of esophageal cancer cells by regulating the
PI3K/Akt/ATM pathway (164).
In the search for methods with which to overcome
tumor resistance, it has also been found that chemotherapy
sensitization can be achieved by targeted intervention in
HMGB1-related pathways (165).
Autocrine and paracrine active signaling-mediated NF-κB
signaling-mediated PD-L1/PD-1 upregulation is a key response to
stress in lung cancer cells and their tumor-associated macrophages
(TAMs), which can be induced by nanodiamond-doxorubicin coupling
(Nano-DOX). The blockade of Nano-DOX-induced PD-L1 in cancer cells
and TAM enhances the activation of TAM-mediated antitumor responses
(165).
In summary, the signaling pathways regulated by
HMGB1 include PI3K/AKT/mTOR, AMPK/mTOR, NF-κB and ERK1/2, amongst
others. HMGB1 binds to a variety of receptors, such as RAGE and
TLRs to induce downstream biological effects, thereby mediating
drug resistance. The clear division of labor and mutual
relationship between each pathway increases the complexity of drug
resistance mechanisms. Thus, determining the core targets for
intervention is of utmost importance to better understand and
overcome drug resistance.
HMGB1 is involved in tumor resistance to
chemotherapy through the regulation of multiple pathways. Given the
various mechanisms of drug resistance, several methods have been
experimented with. Silencing the HMGB1 gene is the most direct
method; silencing of HMGB1 expression in tumor cells can enhance
the sensitivity of tumor cells to chemotherapeutic drugs. In MM,
HMGB1 knockdown has been shown to increase the vulnerability of MM
cells by regulating autophagy and DNA damage repair. HMGB1
knockdown in MM cells enhances the inhibitory effects of
dexamethasone chemotherapy through the induction of apoptosis. The
downregulation of HMGB1 activates the mTOR pathway by regulating
the expression of related genes, inhibiting autophagy and
increasing dexamethasone-induced DNA damage. In vivo
experiments using xenograft models have demonstrated that HMGB1
knockdown in mice leads to a reduced tumor load following
dexamethasone treatment compared with the control mice (25). HMGB1 has also been found to
regulate drug resistance in MM cells by modulating the NF-κB
signaling pathway, and HMGB1 knockdown inhibits MM cell viability
(161). In leukemia, it has been
found that HMGB1 overexpression attenuates the sensitivity of
leukemia cells to adriamycin, vincristine and cytarabine, while
HMGB1 knockdown enhances this effect. The overexpression of HMGB1
induces autophagy through the PI3K/Akt/mTORC1 pathway to induce
drug resistance in leukemic cells, whereas the knockdown of HMGB1
inhibits autophagy and enhances the drug sensitivity of leukemic
cells by increasing the phosphorylation of Akt and p70S6k (166). The expression levels of HMGB1
positively correlate with NLRP3 mRNA levels in patients with AML.
Chronic stress promotes the progression of AML through an
HMGB1/NLRP3/IL-1β signaling pathway. The knockdown of HMGB1
significantly reduces the expression of NLRP3 and IL-1β in AML cell
lines, and the secretion of IL-1β in AML cell culture supernatant,
while the stimulation of HMGB1 expression results in the opposite
effect (167).
The essence of tumor drug resistance is 'the
stronger the tumor's defensive ability' the more likely the tumor
cells are likely to survive. When the combined application of
multiple antagonists exerts poor therapeutic effects and increases
the toxic and side-effects of drugs, TCMs for antitumor therapy
have been shown to exhibit positive clinical effects. TCMs exhibit
low toxicity, multiple targets, multi-components, and multiple
antitumor effects, overcoming the drug resistance to chemotherapy,
and thus have significant prospects for reversing tumor resistance.
TCMs and modern medicines synergistically inhibit the development
of liver cancer, improve sensitivity after chemoresistance,
ameliorate the adverse effects of chemotherapeutic drugs and
molecularly target drugs, and prolong the survival after surgery or
interventional treatment (168).
TCMs primarily consist of compounded formulas and extract
preparations (monomers, active ingredients, compounding, capsules,
granules and injections). The primary types of monomers in Chinese
medicine are alkaloids, flavonoids and saponins, amongst others.
Wei et al (169) found
that TCMs can overcome tumor MDR primarily by regulating ABC
transporters, regulating autophagy, regulating cell cycle
progression, regulating key signaling pathways, regulating CSC and
regulating hypoxia.
The role of compound formulas in overcoming MDR in
tumors has also been shown. Shenling-baizhu Powder is a classic
Chinese herbal formula that has been used for decades in the
treatment of patients with malignancies of the gastrointestinal
tract (170). When combined with
cytoxan (CTX) in a mouse model of HCC, the combined treatment
reduced NF-κB expression and promoted apoptosis, thereby inhibiting
tumor growth, compared to the untreated or treated group (170). Shengbai decoction, which
consists of Codonopsis, astragalus, licorice and
other Chinese herbs, has been shown to reduce the levels of
inflammatory factors in the serum of mice following treatment with
CTX, promote apoptosis of HCC cells, and prolong survival through
regulation of the p53 pathway (171). A previous study demonstrated
that Baosheng combined with the Lomb II formula enhanced the
antitumor effects of 5-fluorouracil by clearing appetite regulators
in the hypothalamic central nervous system and feeding zone and
improving chemotherapy-induced anorexia and gastrointestinal damage
in ruminant mice (172).
TCM monomers can likewise overcome tumor MDR.
Artesunate (an artemisinin derivative derived from Artemisia
annua) significantly enhances the inhibitory effects of
sorafenib on HCC in vitro and in vivo, attributed to
the synergistic effects of lysosome-activated oxidation, the
induction of iron sagging, and the enhancement of the sensitivity
of HCC cells to sorafenib (173). Fucoxanthin (FX) is a major
bioactive component extracted from kelp. It inhibits the cancerous
behaviors of not only human lung cancer PC9 cells. but also of
gefitinib-resistant PC9/G cells. FX inhibits EMT-related factors
(Snail, Twist, fibronectin, N-cadherin, and MMP-2) and sensitizes
drug-resistant cancer cells to gefitinib (174). Baicalein (BAI) is an herbal
monomer that has been found to improve cognitive impairment and
protect microglia from neuroinflammation induced by LPS via the
SIRT1/HMGB1 pathway in -BV2 cells (175). BAI inhibits LPS-induced
inflammation in RAW264.7 cells via a miR-181b/HMGB1/TRL4/NF-κB axis
(176). Curcumin overcomes
primary resistance to gefitinib in NSCLC cells by inducing
autophagy-associated cell death (177). The proteasomal degradation of
HMGB1 by lignocaine inhibits MEK-ERK activation, thereby decreasing
the phosphorylation of Bcl-2 and leading to the constitutive
binding of Bcl-2 to Beclin-1. It has been found that compounded
herbal preparations or herbal monomers achieve therapeutic effects
by interfering with HMGB1, such as auxin injection and its active
ingredient, lignocaine, prevent sepsis in part by inhibiting the
HMGB1/TLR4/NF-κB/MAPKs signaling pathway (178). Gegen Qinlian pills have been
shown to attenuate carrageenan-induced thrombosis in mice by
modulating the HMGB1/NF-κB/NLRP3 signaling pathway (179). In addition, it has been observed
that the high expression of HMGB1 in bortezomib-resistant cells and
the combination of bortezomib with lithophane is very effective in
in vitro and in vivo myeloma models and
re-sensitizing drug-resistant cells to bortezomib (180).
On the whole, the use of TCM monomers and compounds
by the targeted intervention of HMGB1 can reduce invasion and
metastasis, and reverse tumor drug resistance, improve the
tolerance of patients to chemotherapy, reduce the adverse reactions
to chemotherapy, reduce the recurrence of cancer and prolong the
survival of patients.
RNA interference technology is one of the most
effective post-transcriptional gene suppression methods, as it can
specifically eliminate or suppress gene expression, and has several
advantages, including being cost-effective, fast, efficient,
exhibiting high stability and easy to produce, and has been widely
used to explore gene function and the treatment of tumors and
infectious diseases (181). In
recent years, it has been found that ncRNAs play a critical and
complex role in human carcinogenesis and tumor metastasis. Common
ncRNAs include shRNAs, siRNAs, miRNAs, circRNAs and lncRNAs,
amongst others (182). circRNAs
can function as a competitive endogenous RNAs (ceRNAs) to
specifically bind to miRNAs, which in turn inhibits the function of
miRNAs and thus indirectly regulates the expression of the miRNA
target gene (182). siRNAs have
a defined as the precise mode of action with which to achieve
protein knockdown. siRNA-based therapies are advantageous in breast
cancer treatment for inhibiting metastasis, overcoming drug
resistance and immune evasion (183). In a previous study, shRNA or
miR-142-3p was used to target HMGB3, and the suppression of HMGB3
expression inhibited colony formation and induced apoptosis,
including increased ROS accumulation, and decreased MMP, p-mTOR and
STAT3 (184). In another study,
following the simultaneous targeting of HMGA1, HMGA2, HMGB1 and
HMGB3 using shRNAs or miR-142-3p, apoptosis was induced, and cell
viability and the clonogenic capacity of human cervical cancer
cells were inhibited. miR-142-3p induced apoptosis by binding to
the 3′UTR of HMGA1, HMGA2, HMGB1 and HMGB3, and the proliferation,
migration and invasion of human cervical cancer cells were
inhibited (185). Similar and
experimental findings have been observed in human osteosarcoma
cells; the siRNA-mediated knockdown of HMGB1, HMGB2, or HMGB3 in
human osteosarcoma cells increased their sensitivity to
chemotherapy (186).
Tumor drug resistance to chemotherapy is very
common and the mechanisms by which tumor resistance arises are
complex. The overexpression of drug efflux pumps and MDR-associated
proteins, mutations in drug targets, enhanced cell repair function
and reduced apoptosis can all lead to drug resistance in tumor
cells. The complex mechanisms of drug resistance in tumor cells
have not yet been fully elucidated. PCD is regulated by a complex
mechanism that is closely associated with anticancer therapy and
drug resistance. Members of the HMGB protein family, including
HMGB1, HMGB2, HMGB3 and HMGB4 are extensively involved in cellular
processes in tumor cells, including proliferation, metastasis,
autophagy, apoptosis and drug resistance. HMGB1 is closely
associated with the development and progression of a variety of
tumors. HMGB1 is involved in tumor MDR through a variety of
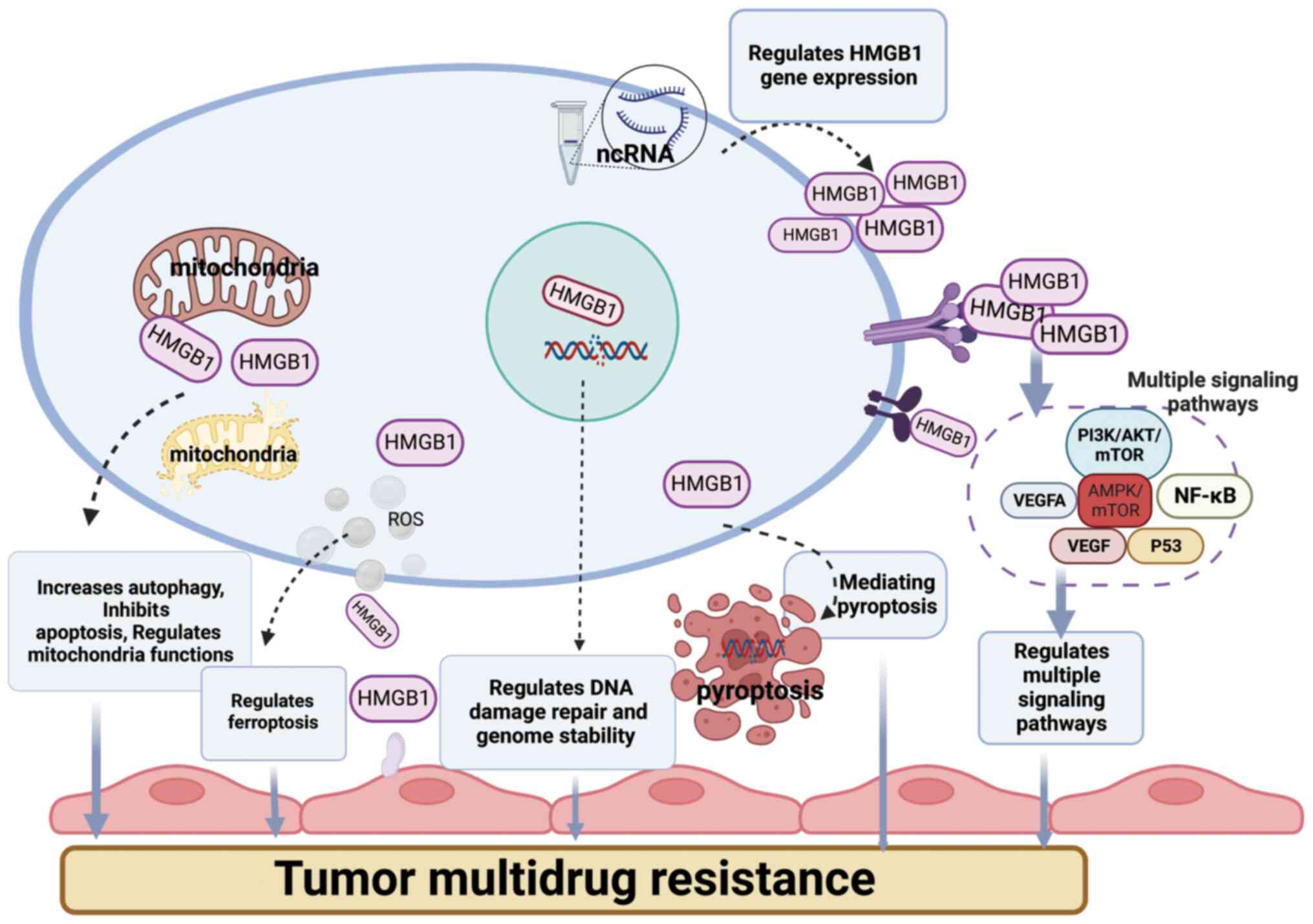
mechanisms (Fig. 1), including
mediating multiple cell death modes and signaling pathways, and
several ncRNAs are involved in tumor drug resistance by regulating
or promoting the function of HMGB1. Several ncRNAs are involved in
tumor drug resistance by regulating or promoting the function of
HMGB1, and various drug resistance mechanisms are clearly regulated
upstream and downstream, but can also crosstalk with each other,
which contributes to the complexity of tumor drug resistance
mechanisms. In a previous meta-analysis on 18 studies from 11
different tumor types, including gastric cancer, CRC, HCC,
pancreatic cancer, nasopharyngeal cancer, head and neck squamous
cell carcinoma, esophageal cancer, malignant pleural mesothelioma,
BC, prostate cancer and cervical cancer, higher HMGB1 protein
levels indicated a lower overall survival and lower
progression-free survival rates (187). Given that the HMGB1 gene is
involved in tumor chemotherapy resistance through multiple
pathways, reducing HMGB1 gene expression is the most direct
strategy with which to address drug resistance and has been
demonstrated in several cellular and animal studies; however, such
experiments cannot be performed in clinical trials. The targeted
intervention of HMGB1 expression can be used in cellular, animal
and clinical trials to identify solutions to unblock drug
resistance. TCM has a long history and has long been widely
accepted. Chinese herbal medicines alone and in combination with
Western medicines can inhibit tumor invasion, metastasis and
reverse drug resistance, and can also be used in combination with
chemical drugs to inhibit tumor cell invasion and metastasis and
thus reverse drug resistance. Research on TCM to interfere with
HMGB1 expression to overcome drug resistance has achieved some
success and is a promising method that warrants further
investigtions. The use of RNA interference technology to
specifically eliminate or inhibit the expression of genes has been
very fruitful in targeting and regulating HMGB1 expression,
chemotherapy sensitization and therapeutic efficacy due to its
several advantages such as economic, speed, efficiency, high
stability and proliferation.
HMGB1 is associated with multiple PCD modalities
including apoptosis, autophagy, apoptosis, pyroptosis and
ferroptosis. Of note, regards tumor development and drug
resistance, it is hypothesized that HMGB1 may also be associated
with the newly discovered cell death modality, cuproptosis. A
previous review of the literature revealed that cuproptosis is a
recently recognized form of cell death driven by intracellular
copper-dependent mitochondrial stress (188). HMGB1, a molecular pattern
associated with injury, is released by copper apoptotic cells to
trigger inflammation. Copper accumulation-induced ATP depletion
activates AMPK to promote HMGB1 phosphorylation, leading to
increased extracellular release (189). By contrast, the genetic (using
RNA interference) or pharmacological (using dorsomorphin)
inhibition of AMPK activation limits copper sagging and HMGB1
release. Functionally, the ability of HMGB1-deficient copper
apoptotic cells to promote late glycosylation end-product-specific
receptor (AGER, also known as RAGE)-dependent inflammatory cytokine
production is significantly reduced. Thus, HMGB1 is a key immune
mediator of aseptic inflammation triggered by copper sagging
(189). As a following step,
exploring whether HMGB1 and cuproptosis are involved in tumor MDR
and identifying chemotherapy sensitization regimens based on their
mechanisms may facilitate a multi-pathway solution to the issue of
tumor chemotherapeutic resistance.
Nanomedicines are the latest promising area of
research and development. Compared to conventional chemotherapeutic
drugs, NPs as drug delivery systems have several unique properties.
NP surface binding to hydrophobic nuclei can overcome drug
solubility issues. NPs can be modified by a variety of molecules
(peptides, small molecule ligands) for effective cell type and
tissue targeting to improve therapeutic efficacy (190). Newly developed iron apoptosis
inducers have now been combined with nanotechnology-based tumor
cell targeting to demonstrate their advantages in overcoming drug
resistance (190), and
NP-mediated iron death enhances natural killer cell antitumor
activity, achieving promising preliminary results in the treatment
of prostate cancer (191). The
combination of conventional chemotherapeutic drugs with other drugs
in the area of nanomedicines has also shown some success in
treatment. The biomimetic co-assembled nanomedicine of DOX and
berberine not only effectively inhibits breast tumor growth with
limited side-effects, but also significantly inhibits lung
metastasis by blocking the HMGB1-TLR4 axis (192). Nanodelivery and self-cellular
drug delivery platforms are considered effective strategies with
which to overcome drug resistance due to their tumor-targeting
ability, controlled release and consistent pharmacokinetic profile,
particularly with the combined application of novel technologies
(including CRISPR systems) (193). Combining ncRNA interference
mechanisms with NP-based technologies has shown promising results
and potential in tumor therapy. siRNA-loaded NPs for GBM, when
loaded with temozolomide (TMZ) as a model, achieve effective
synergistic therapeutic effects by targeting key genes in the TMZ
resistance STAT3 pathway (194).
Given the several advantages of NPs and the understanding of the
numerous mechanisms of targeted intervention of HMGB1 to overcome
drug resistance, the two could be effectively combined in the
future to develop NPs that target the HMGB1 gene. The advantages of
pure drug nanocomponents based on the self-assembly or co-assembly
of pure drug molecules over conventional NPs are promising and
challenging for cancer therapy (195). According to different
tumorigenesis, development and drug resistance mechanisms,
autophagy agonists (or autophagy inhibitors), ferroptosis agonists
(or ferroptosis inhibitors), pyroptosis agonists (or pyroptosis
inhibitors), Chinese herbal medicines with significant therapeutic
effects, and ncRNAs targeting HMGB1 regulation are prepared as pure
drug nano-components, which are precisely delivered to the target
site and are rapidly released to exert their effects in a targeted
manner. The drug can regulate several downstream signaling
pathways, inhibit tumor cell proliferation, induce apoptosis,
inhibit invasion and metastasis, inhibit tumor vascularization,
regulate body immunity and induce programmed tumor cell death, to
overcome tumor MDR. This may serve as a novel approach for the
development of a low-toxicity, high-efficiency and multi-target
reversal strategy for tumor drug resistance using Chinese medicines
and their active ingredients.
Not applicable.
LHS and LZ were involved in the conception of the
review and in the writing of the manuscript. MW, YN and FQC were
involved in the preparation of the figure and in revising the
manuscript. XQG and CTY were involved in the literature search. HWW
and HLL reviewed the manuscript. All authors have read and agreed
to the final version of the manuscript. Data authentication is not
applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
Not applicable.
The present study was funded by the National Natural Science
Foundation of Gansu Province (grant no. 21JR7RA579), and by the
Guiding Projects of the Enshi Tujia and Miao Autonomous Prefecture
Science and Technology Plan of Hubei Province (grant no.
E20220033).
|
1
|
Jolly MK, Somarelli JA, Sheth M, Biddle A,
Tripathi SC, Armstrong AJ, Hanash SM, Bapat SA, Rangarajan A and
Levine H: Hybrid Epithelial/mesenchymal phenotypes promote
metastasis and therapy resistance across carcinomas. Pharmacol
Ther. 194:161–184. 2019. View Article : Google Scholar
|
|
2
|
Phan TG and Croucher PI: The dormant
cancer cell life cycle. Nat Rev Cancer. 20:398–411. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jenkins RW, Barbie DA and Flaherty KT:
Mechanisms of resistance to immune checkpoint inhibitors. Br J
Cancer. 118:9–16. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lopez JS and Banerji U: Combine and
conquer: Challenges for targeted therapy combinations in early
phase trials. Nat Rev Clin Oncol. 14:57–66. 2017. View Article : Google Scholar
|
|
5
|
Wilczyński JR, Wilczyński M and Paradowska
E: Cancer stem cells in ovarian cancer-a source of tumor success
and a challenging target for novel therapies. Int J Mol Sci.
23:24962022. View Article : Google Scholar
|
|
6
|
Vasan N, Baselga J and Hyman DM: A view on
drug resistance in cancer. Nature. 575:299–309. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Assaraf YG, Brozovic A, Gonçalves AC,
Jurkovicova D, Linē A, Machuqueiro M, Saponara S, Sarmento-Ribeiro
AB, Xavier CPR and Vasconcelos MH: The multi-factorial nature of
clinical multidrug resistance in cancer. Drug Resist Updat.
46:1006452019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Schoenfeld AJ and Hellmann MD: Acquired
resistance to immune checkpoint inhibitors. Cancer Cell.
37:443–455. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Aleksakhina SN, Kashyap A and Imyanitov
EN: Mechanisms of acquired tumor drug resistance. Biochim Biophys
Acta Rev Cancer. 1872:1883102019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xie Y, Feng SL, He F, Yan PY, Yao XJ, Fan
XX, Leung EL and Zhou H: Down-regulating Nrf2 by tangeretin
reverses multiple drug resistance to both chemotherapy and EGFR
tyrosine kinase inhibitors in lung cancer. Pharmacol Res.
186:1065142022. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Makena MR, Ranjan A, Thirumala V and Reddy
AP: Cancer stem cells: Road to therapeutic resistance and
strategies to overcome resistance. Biochim Biophys Acta Mol Basis
Dis. 1866:1653392020. View Article : Google Scholar
|
|
12
|
Erin N, Grahovac J, Brozovic A and Efferth
T: Tumor micro-environment and epithelial mesenchymal transition as
targets to overcome tumor multidrug resistance. Drug Resist Updat.
53:1007152020. View Article : Google Scholar
|
|
13
|
Gourley C, Balmaña J, Ledermann JA, Serra
V, Dent R, Loibl S, Pujade-Lauraine E and Boulton SJ: Moving from
poly (ADP-Ribose) polymerase inhibition to targeting DNA repair and
DNA damage response in cancer therapy. J Clin Oncol. 37:2257–2269.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sundar R, Huang KK, Kumar V, Ramnarayanan
K, Demircioglu D, Her Z, Ong X, Bin Adam Isa ZF, Xing M, Tan AL, et
al: Epigenetic promoter alterations in GI tumour immune-editing and
resistance to immune checkpoint inhibition. Gut. 71:1277–1288.
2022. View Article : Google Scholar
|
|
15
|
Carneiro BA and El-Deiry WS: Targeting
apoptosis in cancer therapy. Nat Rev Clin Oncol. 17:395–417. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kumar A and Jaitak V: Natural products as
multidrug resistance modulators in cancer. Eur J Med Chem.
176:268–291. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lai JI, Tseng YJ, Chen MH, Huang CF and
Chang PM: Clinical perspective of FDA approved drugs with
P-Glycoprotein inhibition activities for potential cancer
therapeutics. Front Oncol. 10:5619362020. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Robey RW, Pluchino KM, Hall MD, Fojo AT,
Bates SE and Gottesman MM: Revisiting the role of ABC transporters
in multidrug-resistant cancer. Nat Rev Cancer. 18:452–464. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen Y, Zhang J, Zhang M, Song Y, Zhang Y,
Fan S, Ren S, Fu L, Zhang N, Hui H, et al: Baicalein resensitizes
tamoxifen-resistant breast cancer cells by reducing aerobic
glycolysis and reversing mitochondrial dysfunction via inhibition
of hypoxia-inducible factor-1α. Clin Transl Med. 11:e5772021.
View Article : Google Scholar
|
|
20
|
Zhai K, Mazurakova A, Koklesova L, Kubatka
P and Büsselberg D: Flavonoids synergistically enhance the
anti-glioblastoma effects of chemotherapeutic drugs. Biomolecules.
11:18412021. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
He XL, Xu XH, Shi JJ, Huang M, Wang Y,
Chen X and Lu JJ: Anticancer effects of ginsenoside Rh2: A
systematic review. Curr Mol Pharmacol. 15:179–189. 2022.
|
|
22
|
Wang Y, Xu J, Wang Y, Xiang L and He X:
S-20, a steroidal saponin from the berries of black nightshade,
exerts anti-multidrug resistance activity in K562/ADR cells through
autophagic cell death and ERK activation. Food Funct. 13:2200–2215.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tilaoui M, Ait Mouse H and Zyad A: Update
and new insights on future cancer drug candidates from plant-based
alkaloids. Front Pharmacol. 12:7196942021. View Article : Google Scholar :
|
|
24
|
Majidinia M, Mirza-Aghazadeh-Attari M,
Rahimi M, Mihanfar A, Karimian A, Safa A and Yousefi B: Overcoming
multidrug resistance in cancer: Recent progress in nanotechnology
and new horizons. IUBMB Life. 72:855–871. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Guo X, He D, Zhang E, Chen J, Chen Q, Li
Y, Yang L, Yang Y, Zhao Y, Wang G, et al: HMGB1 knockdown increases
MM cell vulnerability by regulating autophagy and DNA damage
repair. J Exp Clin Cancer Res. 37:2052018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Borde C, Dillard C, L'Honoré A, Quignon F,
Hamon M, Marchand CH, Faccion RS, Costa MGS, Pramil E, Larsen AK,
et al: The C-terminal acidic tail modulates the anti-cancer
properties of HMGB1. Int J Mol Sci. 23:78652022. View Article : Google Scholar
|
|
27
|
Gaikwad S, Puangmalai N, Bittar A,
Montalbano M, Garcia S, McAllen S, Bhatt N, Sonawane M, Sengupta U
and Kayed R: Tau oligomer induced HMGB1 release contributes to
cellular senescence and neuropathology linked to Alzheimer's
disease and frontotemporal dementia. Cell Rep. 36:1094192021.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Stros M: HMGB proteins: Interactions with
DNA and chromatin. Biochim Biophys Acta. 1799:101–113. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wen B, Wei YT and Zhao K: The role of high
mobility group protein B3 (HMGB3) in tumor proliferation and drug
resistance. Mol Cell Biochem. 476:1729–1739. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Voong CK, Goodrich JA and Kugel JF:
Interactions of HMGB Proteins with the Genome and the Impact on
Disease. Biomolecules. 11:14512021. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Catena R, Escoffier E, Caron C, Khochbin
S, Martianov I and Davidson I: HMGB4, a novel member of the HMGB
family, is preferentially expressed in the mouse testis and
localizes to the basal pole of elongating spermatids. Biol Reprod.
80:358–366. 2009. View Article : Google Scholar
|
|
32
|
Niu L, Yang W, Duan L, Wang X, Li Y, Xu C,
Liu C, Zhang Y, Zhou W, Liu J, et al: Biological functions and
theranostic potential of HMGB family members in human cancers. Ther
Adv Med Oncol. 12:17588359209708502020. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Taniguchi N, Kawakami Y, Maruyama I and
Lotz M: HMGB proteins and arthritis. Hum Cell. 31:1–9. 2018.
View Article : Google Scholar
|
|
34
|
Bianchi ME, Crippa MP, Manfredi AA,
Mezzapelle R, Rovere Querini P and Venereau E: High-mobility group
box 1 protein orchestrates responses to tissue damage via
inflammation, innate and adaptive immunity, and tissue repair.
Immunol Rev. 280:74–82. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Cheng KJ, Alshawsh MA, Mejia Mohamed EH,
Thavagnanam S, Sinniah A and Ibrahim ZA: HMGB1: An overview of its
versatile roles in the pathogenesis of colorectal cancer. Cell
Oncol (Dordr). 43:177–193. 2020. View Article : Google Scholar
|
|
36
|
Liu L, Liu S, Luo H, Chen C, Zhang X, He L
and Tu G: GPR30-mediated HMGB1 upregulation in CAFs induces
autophagy and tamoxifen resistance in ERα-positive breast cancer
cells. Aging. 13:16178–16197. 2021. View Article : Google Scholar
|
|
37
|
Wei Y, Tang X, Ren Y, Yang Y, Song F, Fu
J, Liu S, Yu M, Chen J, Wang S, et al: An RNA-RNA crosstalk network
involving HMGB1 and RICTOR facilitates hepatocellular carcinoma
tumorigenesis by promoting glutamine metabolism and impedes
immunotherapy by PD-L1+ exosomes activity. Signal Transduct Target
Ther. 6:4212021. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chai W, Ye F, Zeng L, Li Y and Yang L:
HMGB1-mediated autophagy regulates sodium/iodide symporter protein
degradation in thyroid cancer cells. J Exp Clin Cancer Res.
38:3252019. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhao M, Zhang Y, Jiang Y, Wang K, Wang X,
Zhou D, Wang Y, Yu R and Zhou X: YAP promotes autophagy and
progression of gliomas via upregulating HMGB1. J Exp Clin Cancer
Res. 40:992021. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hubert P, Roncarati P, Demoulin S, Pilard
C, Ancion M, Reynders C, Lerho T, Bruyere D, Lebeau A, Radermecker
C, et al: Extracellular HMGB1 blockade inhibits tumor growth
through profoundly remodeling immune microenvironment and enhances
checkpoint inhibitor-based immunotherapy. J Immunother Cancer.
9:e0019662021. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zha C, Meng X, Li L, Mi S, Qian D, Li Z,
Wu P, Hu S, Zhao S, Cai J and Liu Y: Neutrophil extracellular traps
mediate the crosstalk between glioma progression and the tumor
microenvironment via the HMGB1/RAGE/IL-8 axis. Cancer Biol Med.
17:154–168. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chou YE, Yang PJ, Lin CY, Chen YY, Chiang
WL, Lin PX, Huang ZY, Huang M, Ho YC and Yang SF: The Impact of
HMGB1 Polymorphisms on Prostate Cancer Progression and
Clinicopathological Characteristics. Int J Environ Res Public
Health. 17:13322020. View Article : Google Scholar
|
|
43
|
Wang JB, Li P, Liu XL, Zheng QL, Ma YB,
Zhao YJ, Xie JW, Lin JX, Lu J, Chen QY, et al: An immune checkpoint
score system for prognostic evaluation and adjuvant chemotherapy
selection in gastric cancer. Nat Commun. 11:63522020. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kam NW, Wu KC, Dai W, Wang Y, Yan LYC,
Shakya R, Khanna R, Qin Y, Law S, Lo AWI, et al: Peritumoral B
cells drive proangiogenic responses in HMGB1-enriched esophageal
squamous cell carcinoma. Angiogenesis. 25:181–203. 2022. View Article : Google Scholar
|
|
45
|
Gong W, Zheng Y, Chao F, Li Y, Xu Z, Huang
G, Gao X, Li S and He F: The anti-inflammatory activity of HMGB1 A
box is enhanced when fused with C-terminal acidic tail. J Biomed
Biotechnol. 2010:9152342010. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wang S and Zhang Y: HMGB1 in inflammation
and cancer. J Hematol Oncol. 13:1162020. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Huebener P, Gwak GY, Pradere JP, Quinzii
CM, Friedman R, Lin CS, Trent CM, Mederacke I, Zhao E, Dapito DH,
et al: High-mobility group box 1 is dispensable for autophagy,
mitochondrial quality control, and organ function in vivo. Cell
Metab. 19:539–547. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Rapoport BL, Steel HC, Theron AJ, Heyman
L, Smit T, Ramdas Y and Anderson R: High mobility group Box 1 in
human cancer. Cells. 9:16642020. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Kang R, Zhang Q, Zeh HJ III, Lotze MT and
Tang D: HMGB1 in cancer: Good, bad, or both? Clin Cancer Res.
19:4046–4057. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Amornsupak K, Thongchot S, Thinyakul C,
Box C, Hedayat S, Thuwajit P, Eccles SA and Thuwajit C: HMGB1
mediates invasion and PD-L1 expression through RAGE-PI3K/AKT
signaling pathway in MDA-MB-231 breast cancer cells. BMC Cancer.
22:5782022. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Jiang J, Wang GZ, Wang Y, Huang HZ, Li WT
and Qu XD: Hypoxia-induced HMGB1 expression of HCC promotes tumor
invasiveness and metastasis via regulating macrophage-derived IL-6.
Exp Cell Res. 367:81–88. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Hernandez C, Huebener P, Pradere JP,
Antoine DJ, Friedman RA and Schwabe RF: HMGB1 links chronic liver
injury to progenitor responses and hepatocarcinogenesis. J Clin
Invest. 128:2436–2451. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Zhang H, Wang J, Li J, Zhou X, Yin L, Wang
Y, Gu Y, Niu X, Yang Y, Ji H and Zhang Q: HMGB1 is a key factor for
tamoxifen resistance and has the potential to predict the efficacy
of CDK4/6 inhibitors in breast cancer. Cancer Sci. 112:1603–1613.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Tan G, Huang C, Chen J and Zhi F: HMGB1
released from GSDME-mediated pyroptotic epithelial cells
participates in the tumorigenesis of colitis-associated colorectal
cancer through the ERK1/2 pathway. J Hematol Oncol. 13:1492020.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Li Pomi F, Borgia F, Custurone P, Vaccaro
M, Pioggia G and Gangemi S: Role of HMGB1 in Cutaneous Melanoma:
State of the Art. Int J Mol Sci. 23:93272022. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Gao Q, Wang S, Chen X, Cheng S, Zhang Z,
Li F, Huang L, Yang Y, Zhou B, Yue D, et al: Cancer-cell-secreted
CXCL11 promoted CD8+ T cells infiltration through
docetaxel-induced-release of HMGB1 in NSCLC. J Immunother Cancer.
7:422019. View Article : Google Scholar
|
|
57
|
Zhang X, Shi H, Yuan X, Jiang P, Qian H
and Xu W: Tumor-derived exosomes induce N2 polarization of
neutrophils to promote gastric cancer cell migration. Mol Cancer.
17:1462018. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Ye F, Chai W, Xie M, Yang M, Yu Y, Cao L
and Yang L: HMGB1 regulates erastin-induced ferroptosis via
RAS-JNK/p38 signaling in HL-60/NRAS(Q61L) cells. Am J Cancer Res.
9:730–739. 2019.PubMed/NCBI
|
|
59
|
Liu Y, Yan W, Tohme S, Chen M, Fu Y, Tian
D, Lotze M, Tang D and Tsung A: Hypoxia induced HMGB1 and
mitochondrial DNA interactions mediate tumor growth in
hepatocellular carcinoma through Toll-like receptor 9. J Hepatol.
63:114–121. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Liang L, Fu J, Wang S, Cen H, Zhang L,
Mandukhail SR, Du L, Wu Q, Zhang P and Yu X: MiR-142-3p enhances
chemosensitivity of breast cancer cells and inhibits autophagy by
targeting HMGB1. Acta Pharm Sin B. 10:1036–1046. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Chen Y, Cai L, Guo X, Li Z, Liao X, Zhang
X, Huang L and He J: HMGB1-activated fibroblasts promote breast
cancer cells metastasis via RAGE/aerobic glycolysis. Neoplasma.
68:71–78. 2021. View Article : Google Scholar
|
|
62
|
Lee HJ, Kim A, Song IH, Park IA, Yu JH,
Ahn JH and Gong G: Cytoplasmic expression of high mobility group B1
(HMGB1) is associated with tumor-infiltrating lymphocytes (TILs) in
breast cancer. Pathol Int. 66:202–209. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Yuan S, Liu Z, Xu Z, Liu J and Zhang J:
High mobility group box 1 (HMGB1): A pivotal regulator of
hematopoietic malignancies. J Hematol Oncol. 13:912020. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Yasinska IM, Gonçalves Silva I, Sakhnevych
SS, Ruegg L, Hussain R, Siligardi G, Fiedler W, Wellbrock J,
Bardelli M, Varani L, et al: High mobility group box 1 (HMGB1) acts
as an 'alarmin' to promote acute myeloid leukaemia progression.
Oncoimmunology. 7:e14381092018. View Article : Google Scholar
|
|
65
|
Liu L, Zhang J, Zhang X, Cheng P, Liu L,
Huang Q, Liu H, Ren S, Wei P, Wang C, et al: HMGB1: An important
regulator of myeloid differentiation and acute myeloid leukemia as
well as a promising therapeutic target. J Mol Med (Berl).
99:107–118. 2021. View Article : Google Scholar
|
|
66
|
Zhao M, Yang M, Yang L, Yu Y, Xie M, Zhu
S, Kang R, Tang D, Jiang Z, Yuan W, et al: HMGB1 regulates
autophagy through increasing transcriptional activities of JNK and
ERK in human myeloid leukemia cells. BMB Rep. 44:601–606. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Ding C, Yu H, Shi C, Shi T, Qin H and Cui
Y: MiR-let-7e inhibits invasion and magration and regulates HMGB1
expression in papillary thyroid carcinoma. Biomed Pharmacother.
110:528–536. 2019. View Article : Google Scholar
|
|
68
|
Wang CQ, Huang BF, Wang Y, Tang CH, Jin
HC, Shao F, Shao JK, Wang Q and Zeng Y: Subcellular localization of
HMGB1 in colorectal cancer impacts on tumor grade and survival
prognosis. Sci Rep. 10:185872020. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Qian F, Xiao J, Gai L and Zhu J:
HMGB1-RAGE signaling facilitates Ras-dependent Yap1 expression to
drive colorectal cancer stemness and development. Mol Carcinog.
58:500–510. 2019. View Article : Google Scholar
|
|
70
|
Zhang JL, Zheng HF, Li K and Zhu YP:
miR-495-3p depresses cell proliferation and migration by
downregulating HMGB1 in colorectal cancer. World J Surg Oncol.
20:1012022. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Wu X, Wang W, Chen Y, Liu X, Wang J, Qin
X, Yuan D, Yu T, Chen G, Mi Y, et al: High mobility group box
Protein 1 serves as a potential prognostic marker of lung cancer
and promotes its invasion and metastasis by matrix
Metalloproteinase-2 in a nuclear Factor-κB-dependent manner. Biomed
Res Int. 2018:34537062018.
|
|
72
|
Zhou Y, Liu SX, Zhou YN, Wang J and Ji R:
Research on the relationship between RAGE and its ligand HMGB1, and
prognosis and pathogenesis of gastric cancer with diabetes
mellitus. Eur Rev Med Pharmacol Sci. 25:1339–1350. 2021.PubMed/NCBI
|
|
73
|
van Beijnum JR, Nowak-Sliwinska P, van den
Boezem E, Hautvast P, Buurman WA and Griffioen AW: Tumor
angiogenesis is enforced by autocrine regulation of high-mobility
group box 1. Oncogene. 32:363–374. 2013. View Article : Google Scholar
|
|
74
|
van Beijnum JR, Buurman WA and Griffioen
AW: Convergence and amplification of toll-like receptor (TLR) and
receptor for advanced glycation end products (RAGE) signaling
pathways via high mobility group B1 (HMGB1). Angiogenesis.
11:91–99. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Zhu L, Li X, Chen Y, Fang J and Ge Z:
High-mobility group box 1: A novel inducer of the
epithelial-mesenchymal transition in colorectal carcinoma. Cancer
Lett. 357:527–534. 2015. View Article : Google Scholar
|
|
76
|
Kuniyasu H, Chihara Y and Kondo H:
Differential effects between amphoterin and advanced glycation end
products on colon cancer cells. Int J Cancer. 104:722–727. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Sharma S, Evans A and Hemers E:
Mesenchymal-epithelial signalling in tumour microenvironment: Role
of high-mobility group Box 1. Cell Tissue Res. 365:357–366. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Gialeli C, Theocharis AD and Karamanos NK:
Roles of matrix metalloproteinases in cancer progression and their
pharmacological targeting. FEBS J. 278:16–27. 2011. View Article : Google Scholar
|
|
79
|
Liu PL, Liu WL, Chang JM, Chen YH, Liu YP,
Kuo HF, Hsieh CC, Ding YS, Chen WW and Chong IW: MicroRNA-200c
inhibits epithelial-mesenchymal transition, invasion, and migration
of lung cancer by targeting HMGB1. PLoS One. 12:e01808442017.
View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Yao S, Zhao T and Jin H: Expression of
MicroRNA-325-3p and its potential functions by targeting HMGB1 in
non-small cell lung cancer. Biomed Pharmacother. 70:72–79. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Zhou J, Chen X, Gilvary DL, Tejera MM,
Eksioglu EA, Wei S and Djeu JY: HMGB1 induction of clusterin
creates a chemoresistant niche in human prostate tumor cells. Sci
Rep. 5:150852015. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Liao M, Qin R, Huang W, Zhu HP, Peng F,
Han B and Liu B: Targeting regulated cell death (RCD) with
small-molecule compounds in triple-negative breast cancer: A
revisited perspective from molecular mechanisms to targeted
therapies. J Hematol Oncol. 15:442022. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Bahar E, Han SY, Kim JY and Yoon H:
Chemotherapy resistance: Role of mitochondrial and autophagic
components. Cancers (Basel). 14:14622022. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Ge R, Liu L, Dai W, Zhang W, Yang Y, Wang
H, Shi Q, Guo S, Yi X, Wang G, et al: Xeroderma pigmentosum Group A
promotes autophagy to facilitate cisplatin resistance in melanoma
cells through the activation of PARP1. J Invest Dermatol.
136:1219–1228. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Ferreira PMP, Sousa RWR, Ferreira JRO,
Militão GCG and Bezerra DP: Chloroquine and hydroxychloroquine in
antitumor therapies based on autophagy-related mechanisms.
Pharmacol Res. 168:1055822021. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Kim JS, Lee JM, Chwae YJ, Kim YH, Lee JH,
Kim K, Lee TH, Kim SJ and Park JH: Cisplatin-induced apoptosis in
Hep3B cells: Mitochondria-dependent and -independent pathways.
Biochem Pharmacol. 67:1459–1468. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Çoku J, Booth DM, Skoda J, Pedrotty MC,
Vogel J, Liu K, Vu A, Carpenter EL, Ye JC, Chen MA, et al: Reduced
ER-mitochondria connectivity promotes neuroblastoma multidrug
resistance. EMBO J. 41:e1082722022. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Yang M, Liu L, Xie M, Sun X, Yu Y, Kang R,
Yang L, Zhu S, Cao L and Tang D: Poly-ADP-ribosylation of HMGB1
regulates TNFSF10/TRAIL resistance through autophagy. Autophagy.
11:214–224. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Pal Singh M, Pal Khaket T, Bajpai VK,
Alfarraj S, Kim SG, Chen L, Huh YS, Han YK and Kang SC: Morin
Hydrate Sensitizes Hepatoma Cells and Xenograft Tumor towards
Cisplatin by Downregulating PARP-1-HMGB1 Mediated Autophagy. Int J
Mol Sci. 21:82532020. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Tang D, Loze MT, Zeh HJ and Kang R: The
redox protein HMGB1 regulates cell death and survival in cancer
treatment. Autophagy. 6:1181–1183. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Tang D, Kang R, Livesey KM, Cheh CW,
Farkas A, Loughran P, Hoppe G, Bianchi ME, Tracey KJ, Zeh HJ III
and Lotze MT: Endogenous HMGB1 regulates autophagy. J Cell Biol.
190:881–892. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Tang D, Kang R, Livesey KM, Cheh CW,
Farkas A, Loughran P, Hoppe G, Bianchi ME, Tracey KJ, Zeh HJ III
and Lotze MT: HMGB1 release and redox regulates autophagy and
apoptosis in cancer cells. Oncogene. 29:5299–5310. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Singh MP, Cho HJ, Kim JT, Baek KE, Lee HG
and Kang SC: Morin hydrate reverses cisplatin resistance by
impairing PARP1/HMGB1-dependent autophagy in hepatocellular
carcinoma. Cancers (Basel). 11:9862019. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Shi Q, Wang Y, Dong W, Song E and Song Y:
Polychlorinated biphenyl quinone-induced signaling transition from
autophagy to apoptosis is regulated by HMGB1 and p53 in human
hepatoma HepG2 cells. Toxicol Lett. 306:25–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Shi Y, Gong W, Lu L, Wang Y and Ren J:
Upregulation of miR-129-5p increases the sensitivity to Taxol
through inhibiting HMGB1-mediated cell autophagy in breast cancer
MCF-7 cells. Braz J Med Biol Res. 52:e86572019. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Zhang Y, Chu X and Wei Q: MiR-451 promotes
cell apoptosis and inhibits autophagy in pediatric acute myeloid
leukemia by targeting HMGB1. J Environ Pathol Toxicol Oncol.
40:45–53. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Run L, Wang L, Nong X, Li N, Huang X and
Xiao Y: Involvement of HMGB1 in vemurafenib resistance in thyroid
cancer cells harboring BRAF (V600E) mutation by regulating
excessive autophagy. Endocrine. 71:418–426. 2021. View Article : Google Scholar
|
|
98
|
Mou K, Liu W, Han D and Li P: HMGB1/RAGE
axis promotes autophagy and protects keratinocytes from ultraviolet
radiation-induced cell death. J Dermatol Sci. 85:162–169. 2017.
View Article : Google Scholar
|
|
99
|
Li H, Li J, Zhang G, Da Q, Chen L, Yu S,
Zhou Q, Weng Z, Xin Z, Shi L, et al: HMGB1-induced p62
overexpression promotes snail-mediated epithelial-mesenchymal
transition in glioblastoma cells via the degradation of GSK-3β.
Theranostics. 9:1909–1922. 2019. View Article : Google Scholar :
|
|
100
|
Liu W, Zhang Z, Zhang Y, Chen X, Guo S,
Lei Y, Xu Y, Ji C, Bi Z and Wang K: HMGB1-mediated autophagy
modulates sensitivity of colorectal cancer cells to oxaliplatin via
MEK/ERK signaling pathway. Cancer Biol Ther. 16:511–517. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Chen SY, Hsu YH, Wang SY, Chen YY, Hong CJ
and Yen GC: Lucidone inhibits autophagy and MDR1 via
HMGB1/RAGE/PI3K/Akt signaling pathway in pancreatic cancer cells.
Phytother Res. 36:1664–1677. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Luo P, Xu Z, Li G, Yan H, Zhu Y, Zhu H, Ma
S, Yang B and He Q: HMGB1 represses the anti-cancer activity of
sunitinib by governing TP53 autophagic degradation via its
nucleus-to-cytoplasm transport. Autophagy. 14:2155–2170. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Dixon SJ, Lemberg KM, Lamprecht MR, Skouta
R, Zaitsev EM, Gleason CE, Patel DN, Bauer AJ, Cantley AM, Yang WS,
et al: Ferroptosis: An iron-dependent form of nonapoptotic cell
death. Cell. 149:1060–1072. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Cao JY and Dixon SJ: Mechanisms of
ferroptosis. Cell Mol Life Sci. 73:2195–2209. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Liang C, Zhang X, Yang M and Dong X:
Recent progress in ferroptosis inducers for cancer therapy. Adv
Mater. 31:e19041972019. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Yan HF, Zou T, Tuo QZ, Xu S, Li H, Belaidi
AA and Lei P: Ferroptosis: Mechanisms and links with diseases.
Signal Transduct Target Ther. 6:492021. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Hassannia B, Vandenabeele P and Vanden
Berghe T: Targeting ferroptosis to Iron out cancer. Cancer Cell.
35:830–849. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Stockwell BR: Ferroptosis turns 10:
Emerging mechanisms, physiological functions, and therapeutic
applications. Cell. 185:2401–2421. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Sosa V, Moliné T, Somoza R, Paciucci R,
Kondoh H and ME LL: Oxidative stress and cancer: An overview.
Ageing Res Rev. 12:376–390. 2013. View Article : Google Scholar
|
|
110
|
Wang D, Tang L, Zhang Y, Ge G, Jiang X, Mo
Y, Wu P, Deng X, Li L, Zuo S, et al: Regulatory pathways and drugs
associated with ferroptosis in tumors. Cell Death Dis. 13:5442022.
View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Chen R, Kang R and Tang D: The mechanism
of HMGB1 secretion and release. Exp Mol Med. 54:91–102. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Wen Q, Liu J, Kang R, Zhou B and Tang D:
The release and activity of HMGB1 in ferroptosis. Biochem Biophys
Res Commun. 510:278–283. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Wiernicki B, Maschalidi S, Pinney J,
Adjemian S, Vanden Berghe T, Ravichandran KS and Vandenabeele P:
Cancer cells dying from ferroptosis impede dendritic cell-mediated
anti-tumor immunity. Nat Commun. 13:36762022. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Efimova I, Catanzaro E, Van der Meeren L,
Turubanova VD, Hammad H, Mishchenko TA, Vedunova MV, Fimognari C,
Bachert C, Coppieters F, et al: Vaccination with early ferroptotic
cancer cells induces efficient antitumor immunity. J Immunother
Cancer. 8:e0013692020. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Vats K, Kruglov O, Mizes A, Samovich SN,
Amoscato AA, Tyurin VA, Tyurina YY, Kagan VE and Bunimovich YL:
Keratinocyte death by ferroptosis initiates skin inflammation after
UVB exposure. Redox Biol. 47:1021432021. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Zhang H, Wang Z, Liu Z, Du K and Lu X:
Protective effects of dexazoxane on rat ferroptosis in
doxorubicin-induced cardiomyopathy through regulating HMGB1. Front
Cardiovasc Med. 8:6854342021. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Friedmann Angeli JP, Krysko DV and Conrad
M: Ferroptosis at the crossroads of cancer-acquired drug resistance
and immune evasion. Nat Rev Cancer. 19:405–414. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Chaudhary N, Choudhary BS, Shah SG,
Khapare N, Dwivedi N, Gaikwad A, Joshi N, Raichanna J, Basu S,
Gurjar M, et al: Lipocalin 2 expression promotes tumor progression
and therapy resistance by inhibiting ferroptosis in colorectal
cancer. Int J Cancer. 149:1495–1511. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
You JH, Lee J and Roh JL: PGRMC1-dependent
lipophagy promotes ferroptosis in paclitaxel-tolerant persister
cancer cells. J Exp Clin Cancer Res. 40:3502021. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Lee J, You JH, Shin D and Roh JL:
Inhibition of Glutaredoxin 5 predisposes Cisplatin-resistant head
and neck cancer cells to ferroptosis. Theranostics. 10:7775–7786.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Wu D, Wang S, Yu G and Chen X: Cell death
mediated by the pyroptosis pathway with the aid of nanotechnology:
Prospects for cancer therapy. Angew Chem Int Ed Engl. 60:8018–8034.
2021. View Article : Google Scholar
|
|
122
|
Qi S, Wang Q, Zhang J, Liu Q and Li C:
Pyroptosis and its role in the modulation of cancer progression and
antitumor immunity. Int J Mol Sci. 23:104942022. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Hage C, Hoves S, Strauss L, Bissinger S,
Prinz Y, Pöschinger T, Kiessling F and Ries CH: Sorafenib induces
pyroptosis in macrophages and triggers natural killer cell-mediated
cytotoxicity against hepatocellular carcinoma. Hepatology.
70:1280–1297. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Wu LS, Liu Y, Wang XW, Xu B, Lin YL, Song
Y, Dong Y, Liu JL, Wang XJ, Liu S, et al: LPS enhances the
chemosensitivity of oxaliplatin in HT29 cells via GSDMD-mediated
pyroptosis. Cancer Manag Res. 12:10397–10409. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Tang Z, Ji L, Han M, Xie J, Zhong F, Zhang
X, Su Q, Yang Z, Liu Z, Gao H and Jiang G: Pyroptosis is involved
in the inhibitory effect of FL118 on growth and metastasis in
colorectal cancer. Life Sci. 257:1180652020. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Yu J, Li S, Qi J, Chen Z, Wu Y, Guo J,
Wang K, Sun X and Zheng J: Cleavage of GSDME by caspase-3
determines lobaplatin-induced pyroptosis in colon cancer cells.
Cell Death Dis. 10:1932019. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Wang F, Liu W, Ning J, Wang J, Lang Y, Jin
X, Zhu K, Wang X, Li X, Yang F, et al: Simvastatin suppresses
proliferation and migration in non-small cell lung cancer via
pyroptosis. Int J Biol Sci. 14:406–417. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Wu L, Lu H, Pan Y, Liu C, Wang J, Chen B
and Wang Y: The role of pyroptosis and its crosstalk with immune
therapy in breast cancer. Front Immunol. 13:9739352022. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Yu P, Zhang X, Liu N, Tang L, Peng C and
Chen X: Pyroptosis: Mechanisms and diseases. Signal Transduct
Target Ther. 6:1282021. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Xia X, Wang X, Cheng Z, Qin W, Lei L,
Jiang J and Hu J: The role of pyroptosis in cancer: Pro-cancer or
pro-'host'? Cell Death Dis. 10:6502019. View Article : Google Scholar
|
|
131
|
Volchuk A, Ye A, Chi L, Steinberg BE and
Goldenberg NM: Indirect regulation of HMGB1 release by gasdermin D.
Nat Commun. 11:45612020. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Deng M, Tang Y, Li W, Wang X, Zhang R,
Zhang X, Zhao X, Liu J, Tang C, Liu Z, et al: The endotoxin
delivery protein HMGB1 mediates Caspase-11-dependent lethality in
sepsis. Immunity. 49:740–753.e7. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Hou W, Wei X, Liang J, Fang P, Ma C, Zhang
Q and Gao Y: HMGB1-induced hepatocyte pyroptosis expanding
inflammatory responses contributes to the pathogenesis of
Acute-on-chronic liver failure (ACLF). J Inflamm Res. 14:7295–7313.
2021. View Article : Google Scholar
|
|
134
|
Peng Z, Wang P, Song W, Yao Q, Li Y, Liu
L, Li Y and Zhou S: GSDME enhances Cisplatin sensitivity to regress
non-small cell lung carcinoma by mediating pyroptosis to trigger
antitumor immunocyte infiltration. Signal Transduct Target Ther.
5:1592020. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Shi F, Zhang L, Liu X and Wang Y:
Knock-down of microRNA miR-556-5p increases cisplatin-sensitivity
in non-small cell lung cancer (NSCLC) via activating NLR family
pyrin domain containing 3 (NLRP3)-mediated pyroptotic cell death.
Bioengineered. 12:6332–6342. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Erkes DA, Cai W, Sanchez IM, Purwin TJ,
Rogers C, Field CO, Berger AC, Hartsough EJ, Rodeck U, Alnemri ES
and Aplin AE: Mutant BRAF and MEK inhibitors regulate the tumor
immune microenvironment via pyroptosis. Cancer Discov. 10:254–269.
2020. View Article : Google Scholar :
|
|
137
|
Chen B, Dragomir MP, Yang C, Li Q, Horst D
and Calin GA: Targeting non-coding RNAs to overcome cancer therapy
resistance. Signal Transduct Target Ther. 7:1212022. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Barth DA, Drula R, Ott L, Fabris L, Slaby
O, Calin GA and Pichler M: Circulating Non-coding RNAs in renal
cell carcinoma-pathogenesis and potential implications as clinical
biomarkers. Front Cell Dev Biol. 8:8282020. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Zeuschner P, Linxweiler J and Junker K:
Non-coding RNAs as biomarkers in liquid biopsies with a special
emphasis on extracellular vesicles in urological malignancies.
Expert Rev Mol Diagn. 20:151–167. 2020. View Article : Google Scholar
|
|
140
|
Barth DA, Slaby O, Klec C, Juracek J,
Drula R, Calin GA and Pichler M: Current concepts of Non-coding
RNAs in the pathogenesis of non-clear cell renal cell carcinoma.
Cancers (Basel). 11:15802019. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Grimson A, Farh KK, Johnston WK,
Garrett-Engele P, Lim LP and Bartel DP: MicroRNA targeting
specificity in mammals: Determinants beyond seed pairing. Mol Cell.
27:91–105. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
He B, Zhao Z, Cai Q, Zhang Y, Zhang P, Shi
S, Xie H, Peng X, Yin W, Tao Y and Wang X: miRNA-based biomarkers,
therapies, and resistance in Cancer. Int J Biol Sci. 16:2628–2647.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Taheri M, Shirvani-Farsani Z,
Ghafouri-Fard S and Omrani MD: Expression profile of microRNAs in
bladder cancer and their application as biomarkers. Biomed
Pharmacother. 131:1107032020. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Deng H, Lv L, Li Y, Zhang C, Meng F, Pu Y,
Xiao J, Qian L, Zhao W, Liu Q, et al: miR-193a-3p regulates the
multi-drug resistance of bladder cancer by targeting the LOXL4 gene
and the oxidative stress pathway. Mol Cancer. 13:2342014.
View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Díaz-Martínez M, Benito-Jardón L, Alonso
L, Koetz-Ploch L, Hernando E and Teixidó J: miR-204-5p and
miR-211-5p Contribute to BRAF inhibitor resistance in melanoma.
Cancer Res. 78:1017–1030. 2018. View Article : Google Scholar :
|
|
146
|
Sur S and Ray RB: Emerging role of lncRNA
ELDR in development and cancer. FEBS J. 289:3011–3023. 2022.
View Article : Google Scholar
|
|
147
|
Zhu L, Zhu Y, Han S, Chen M, Song P, Dai
D, Xu W, Jiang T, Feng L, Shin VY, et al: Impaired autophagic
degradation of lncRNA ARHGAP5-AS1 promotes chemoresistance in
gastric cancer. Cell Death Dis. 10:3832019. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Luo Y, Zheng S, Wu Q, Wu J, Zhou R, Wang
C, Wu Z, Rong X, Huang N, Sun L, et al: Long noncoding RNA (lncRNA)
EIF3J-DT induces chemoresistance of gastric cancer via autophagy
activation. Autophagy. 17:4083–4101. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Lu J, Xu Y, Xie W, Tang Y, Zhang H, Wang
B, Mao J, Rui T, Jiang P and Zhang W: Long noncoding RNA DLGAP1-AS2
facilitates Wnt1 transcription through physically interacting with
Six3 and drives the malignancy of gastric cancer. Cell Death
Discov. 7:2552021. View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Ren C, Han H, Pan J, Chang Q, Wang W, Guo
X and Bian J: DLGAP1-AS2 promotes human colorectal cancer
progression through trans-activation of Myc. Mamm Genome.
33:672–683. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Li Q, Sun H, Luo D, Gan L, Mo S, Dai W,
Liang L, Yang Y, Xu M, Li J, et al: Lnc-RP11-536 K7.3/SOX2/HIF-1α
signaling axis regulates oxaliplatin resistance in patient-derived
colorectal cancer organoids. J Exp Clin Cancer Res. 40:3482021.
View Article : Google Scholar
|
|
152
|
Chioccarelli T, Falco G, Cappetta D, De
Angelis A, Roberto L, Addeo M, Ragusa M, Barbagallo D, Berrino L,
Purrello M, et al: FUS driven circCNOT6L biogenesis in mouse and
human spermatozoa supports zygote development. Cell Mol Life Sci.
79:502021. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Singh V, Uddin MH, Zonder JA, Azmi AS and
Balasubramanian SK: Circular RNAs in acute myeloid leukemia. Mol
Cancer. 20:1492021. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Chen DL, Sheng H, Zhang DS, Jin Y, Zhao
BT, Chen N, Song K and Xu RH: The circular RNA circDLG1 promotes
gastric cancer progression and anti-PD-1 resistance through the
regulation of CXCL12 by sponging miR-141-3p. Mol Cancer.
20:1662021. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Li P, Song R, Yin F, Liu M, Liu H, Ma S,
Jia X, Lu X, Zhong Y, Yu L, et al: circMRPS35 promotes malignant
progression and cisplatin resistance in hepatocellular carcinoma.
Mol Ther. 30:431–447. 2022. View Article : Google Scholar :
|
|
156
|
Zhang Y, Liu Y and Xu X: Upregulation of
miR-142-3p improves drug sensitivity of acute myelogenous leukemia
through reducing P-glycoprotein and repressing autophagy by
targeting HMGB1. Transl Oncol. 10:410–418. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
LeClair K, Bell KJL, Furuya-Kanamori L,
Doi SA, Francis DO and Davies L: Evaluation of gender inequity in
thyroid cancer diagnosis: Differences by Sex in US thyroid cancer
incidence compared with a meta-analysis of subclinical thyroid
cancer rates at autopsy. JAMA Internal Med. 181:1351–1358. 2021.
View Article : Google Scholar
|
|
158
|
Lei T, Huang J, Xie F, Gu J, Cheng Z and
Wang Z: HMGB1-mediated autophagy promotes gefitinib resistance in
human non-small cell lung cancer. Acta Biochim Biophys Sin
(Shanghai). 54:fpage–lpage. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Kishi S, Fujiwara-Tani R, Honoki K, Sasaki
R, Mori S, Ohmori H, Sasaki T, Miyagawa Y, Kawahara I, Kido A, et
al: Oxidized high mobility group B-1 enhances metastability of
colorectal cancer via modification of mesenchymal stem/stromal
cells. Cancer Sci. 113:2904–2915. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Ren Y, Cao L, Wang L, Zheng S, Zhang Q,
Guo X, Li X, Chen M, Wu X, Furlong F, et al: Autophagic secretion
of HMGB1 from cancer-associated fibroblasts promotes metastatic
potential of non-small cell lung cancer cells via NFκB signaling.
Cell Death Dis. 12:8582021. View Article : Google Scholar
|
|
161
|
Ning J, Yang R, Wang H and Cui L: HMGB1
enhances chemotherapy resistance in multiple myeloma cells by
activating the nuclear factor-κB pathway. Exp Ther Med. 22:7052021.
View Article : Google Scholar
|
|
162
|
Zhou P, Zheng ZH, Wan T, Wu J, Liao CW and
Sun XJ: Vitexin inhibits gastric cancer growth and metastasis
through HMGB1-mediated inactivation of the PI3K/AKT/mTOR/HIF-1α
signaling pathway. J Gastric Cancer. 21:439–456. 2021. View Article : Google Scholar
|
|
163
|
Pu Z, Duda DG, Zhu Y, Pei S, Wang X, Huang
Y, Yi P, Huang Z, Peng F, Hu X and Fan X: VCP interaction with
HMGB1 promotes hepatocellular carcinoma progression by activating
the PI3K/AKT/mTOR pathway. J Transl Med. 20:2122022. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Zhang X, Zou N, Deng W, Song C, Yan K,
Shen W and Zhu S: HMGB1 induces radioresistance through
PI3K/AKT/ATM pathway in esophageal squamous cell carcinoma. Mol
Biol Rep. 49:11933–11945. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Xu HZ, Li TF, Wang C, Ma Y, Liu Y, Zheng
MY, Liu ZJ, Chen JB, Li K, Sun SK, et al: Synergy of
nanodiamond-doxorubicin conjugates and PD-L1 blockade effectively
turns tumor-associated macrophages against tumor cells. J
Nanobiotechnol. 19:2682021. View Article : Google Scholar
|
|
166
|
Yang L, Yu Y, Kang R, Yang M, Xie M, Wang
Z, Tang D, Zhao M, Liu L, Zhang H and Cao L: Up-regulated autophagy
by endogenous high mobility group box-1 promotes chemoresistance in
leukemia cells. Leuk Lymphoma. 53:315–322. 2012. View Article : Google Scholar
|
|
167
|
Liu N, Wu Y, Wen X, Li P, Lu F and Shang
H: Chronic stress promotes acute myeloid leukemia progression
through HMGB1/NLRP3/IL-1β signaling pathway. J Mol Med (Berl).
99:403–414. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Wei L, Wang Z, Jing N, Lu Y, Yang J, Xiao
H, Guo H, Sun S, Li M, Zhao D, et al: Frontier progress of the
combination of modern medicine and traditional Chinese medicine in
the treatment of hepatocellular carcinoma. Chin Med. 17:902022.
View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Wei J, Liu Z, He J, Liu Q, Lu Y, He S,
Yuan B, Zhang J and Ding Y: Traditional Chinese medicine reverses
cancer multidrug resistance and its mechanism. Clin Transl Oncol.
24:471–482. 2022. View Article : Google Scholar
|
|
170
|
Xi S, Peng Y, Minuk GY, Shi M, Fu B, Yang
J, Li Q, Gong Y, Yue L, Li L, et al: The combination effects of
Shen-Ling-Bai-Zhu on promoting apoptosis of transplanted H22
hepatocellular carcinoma in mice receiving chemotherapy. J
Ethnopharmacol. 190:1–12. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
171
|
Wang H, Li Y, Lu J, Qiu M, Cheng D, Zhang
X and Yuan W: Shengbai decoction enhances the anti-tumor efficacy
of cyclophosphamide on hepatoma 22-bearing mice. Biomed
Pharmacother. 140:1117752021. View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Xi S, Zhai X, Wang Y, Gong Y, Fu B, Gao C,
Guo X, Li Y, Wang Z, Huang S, et al: The Ciji-Hua'ai-Baosheng II
formula attenuates chemotherapy-induced anorexia in mice with
H22 hepatocellular carcinoma. Front Pharmacol.
12:7158242021. View Article : Google Scholar
|
|
173
|
Li ZJ, Dai HQ, Huang XW, Feng J, Deng JH,
Wang ZX, Yang XM, Liu YJ, Wu Y, Chen PH, et al: Artesunate
synergizes with sorafenib to induce ferroptosis in hepatocellular
carcinoma. Acta Pharmacol Sin. 42:301–310. 2021. View Article : Google Scholar :
|
|
174
|
Ming JX, Wang ZC, Huang Y, Ohishi H, Wu
RJ, Shao Y, Wang H, Qin MY, Wu ZL, Li YY, et al: Fucoxanthin
extracted from Laminaria Japonica inhibits metastasis and enhances
the sensitivity of lung cancer to Gefitinib. J Ethnopharmacol.
265:1133022021. View Article : Google Scholar
|
|
175
|
Li Y, Liu T, Li Y, Han D, Hong J, Yang N,
He J, Peng R, Mi X, Kuang C, et al: Baicalin ameliorates cognitive
impairment and protects microglia from LPS-induced
neuroinflammation via the SIRT1/HMGB1 pathway. Oxid Med Cell
Longev. 2020:47513492020. View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Yan G, Chen L, Wang H, Wu S, Li S and Wang
X: Baicalin inhibits LPS-induced inflammation in RAW264.7 cells
through miR-181b/HMGB1/TRL4/NF-κB pathway. Am J Transl Res.
13:10127–10141. 2021.
|
|
177
|
Chen P, Huang HP, Wang Y, Jin J, Long WG,
Chen K, Zhao XH, Chen CG and Li J: Curcumin overcome primary
gefitinib resistance in non-small-cell lung cancer cells through
inducing autophagy-related cell death. J Exp Clin Cancer Res.
38:2542019. View Article : Google Scholar : PubMed/NCBI
|
|
178
|
Wang Z, Chen W, Li Y, Zhang S, Lou H, Lu X
and Fan X: Reduning injection and its effective constituent
luteoloside protect against sepsis partly via inhibition of
HMGB1/TLR4/NF-κB/MAPKs signaling pathways. J Ethnopharmacol.
270:1137832021. View Article : Google Scholar
|
|
179
|
Wei X, Zhang B, Wei F, Ding M, Luo Z, Han
X and Tan X: Gegen Qinlian pills alleviate carrageenan-induced
thrombosis in mice model by regulating the HMGB1/NF-κB/NLRP3
signaling. Phytomedicine. 100:1540832022. View Article : Google Scholar
|
|
180
|
Roy M, Liang L, Xiao X, Peng Y, Luo Y,
Zhou W, Zhang J, Qiu L, Zhang S, Liu F, et al: Lycorine
downregulates HMGB1 to inhibit autophagy and enhances bortezomib
activity in multiple myeloma. Theranostics. 6:2209–2224. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
181
|
Yu AM, Jian C, Yu AH and Tu MJ: RNA
therapy: Are we using the right molecules? Pharmacol Ther.
196:91–104. 2019. View Article : Google Scholar :
|
|
182
|
Kristensen LS, Andersen MS, Stagsted LVW,
Ebbesen KK, Hansen TB and Kjems J: The biogenesis, biology and
characterization of circular RNAs. Nat Rev Genet. 20:675–691. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
183
|
Ngamcherdtrakul W and Yantasee W: siRNA
therapeutics for breast cancer: Recent efforts in targeting
metastasis, drug resistance, and immune evasion. Transl Res.
214:105–120. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
184
|
Sharma P, Yadav P, Sundaram S, Venkatraman
G, Bera AK and Karunagaran D: HMGB3 inhibition by miR-142-3p/sh-RNA
modulates autophagy and induces apoptosis via ROS accumulation and
mitochondrial dysfunction and reduces the tumorigenic potential of
human breast cancer cells. Life Sci. 304:1207272022. View Article : Google Scholar : PubMed/NCBI
|
|
185
|
Sharma P, Yadav P, Jain RP, Bera AK and
Karunagaran D: miR-142-3p simultaneously targets HMGA1, HMGA2,
HMGB1, and HMGB3 and inhibits tumorigenic properties and in-vivo
metastatic potential of human cervical cancer cells. Life Sci.
291:1202682022. View Article : Google Scholar : PubMed/NCBI
|
|
186
|
Mukherjee A and Vasquez KM: Targeting
chromosomal architectural HMGB proteins could be the next frontier
in cancer therapy. Cancer Res. 80:2075–2082. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
187
|
Wu T, Zhang W, Yang G, Li H, Chen Q, Song
R and Zhao L: HMGB1 overexpression as a prognostic factor for
survival in cancer: A meta-analysis and systematic review.
Oncotarget. 7:50417–50427. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
188
|
Xie J, Yang Y, Gao Y and He J:
Cuproptosis: Mechanisms and links with cancers. Mol Cancer.
22:462023. View Article : Google Scholar : PubMed/NCBI
|
|
189
|
Liu J, Liu Y, Wang Y, Kang R and Tang D:
HMGB1 is a mediator of cuproptosis-related sterile inflammation.
Front Cell Dev Biol. 10:9963072022. View Article : Google Scholar : PubMed/NCBI
|
|
190
|
Hassannia B, Wiernicki B, Ingold I, Qu F,
Van Herck S, Tyurina YY, Bayır H, Abhari BA, Angeli JPF, Choi SM,
et al: Nano-targeted induction of dual ferroptotic mechanisms
eradicates high-risk neuroblastoma. J Clin Invest. 128:3341–3355.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
191
|
Kim KS, Choi B, Choi H, Ko MJ and Kim DH
and Kim DH: Enhanced natural killer cell anti-tumor activity with
nanoparticles mediated ferroptosis and potential therapeutic
application in prostate cancer. J Nanobiotechnol. 20:4282022.
View Article : Google Scholar
|
|
192
|
Zheng X, Zhao Y, Jia Y, Shao D, Zhang F,
Sun M, Dawulieti J, Hu H, Cui L, Pan Y, et al: Biomimetic
co-assembled nanodrug of doxorubicin and berberine suppresses
chemotherapy-exacerbated breast cancer metastasis. Biomaterials.
271:1207162021. View Article : Google Scholar : PubMed/NCBI
|
|
193
|
Meng X, Lou QY, Yang WY, Wang YR, Chen R,
Wang L, Xu T and Zhang L: The role of non-coding RNAs in drug
resistance of oral squamous cell carcinoma and therapeutic
potential. Cancer Commun (Lond). 41:981–1006. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
194
|
Jiang T, Qiao Y, Ruan W, Zhang D, Yang Q,
Wang G, Chen Q, Zhu F, Yin J, Zou Y, et al: Cation-free siRNA
micelles as effective drug delivery platform and potent RNAi
nanomedicines for glioblastoma therapy. Adv Mater. 33:e21047792021.
View Article : Google Scholar : PubMed/NCBI
|
|
195
|
Fu S, Li G, Zang W, Zhou X, Shi K and Zhai
Y: Pure drug nano-assemblies: A facile carrier-free nanoplatform
for efficient cancer therapy. Acta Pharm Sin B. 12:92–106. 2022.
View Article : Google Scholar : PubMed/NCBI
|