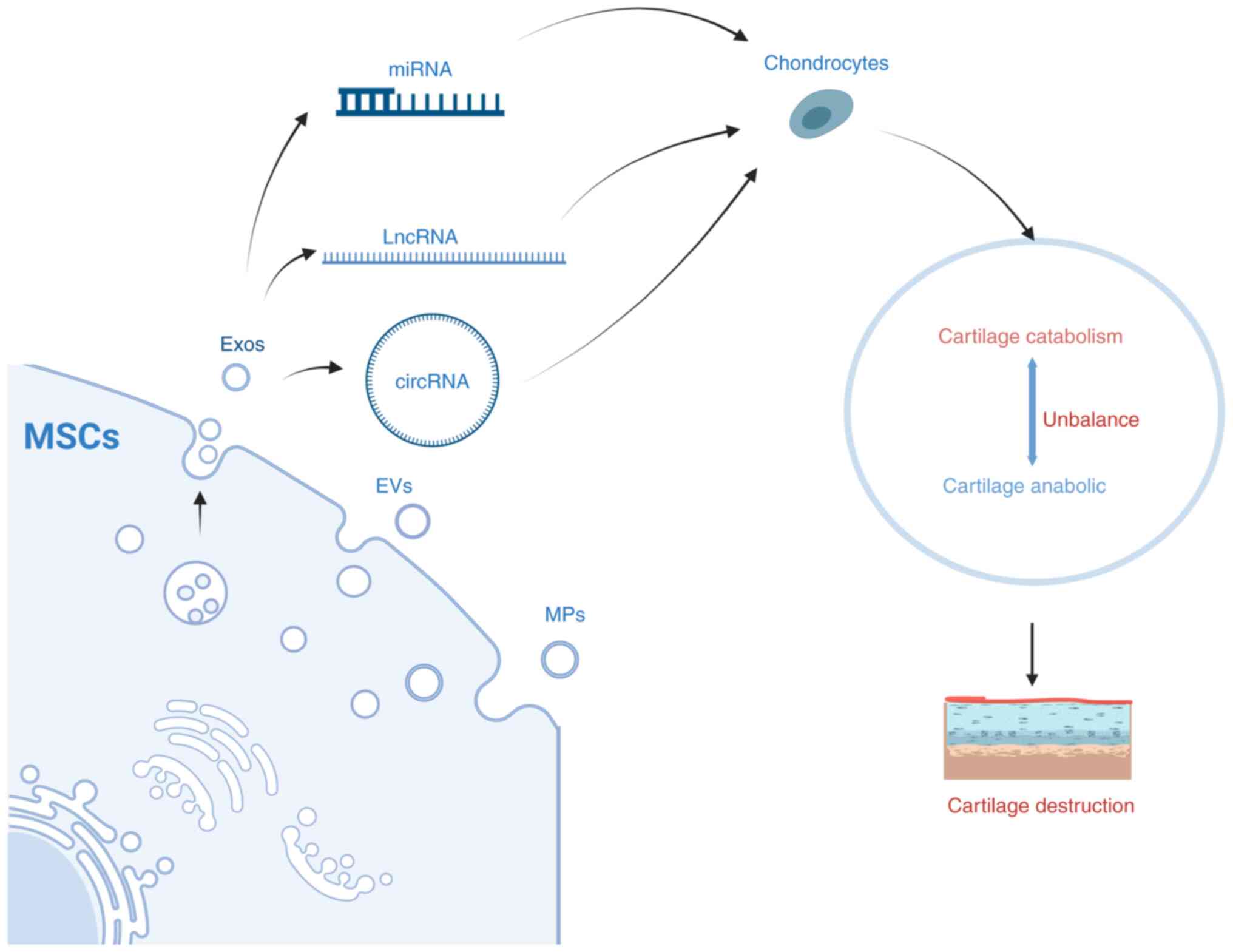
Osteoarthritis (OA) is a chronic joint disease
characterized by articular cartilage degeneration and secondary
bone hyperplasia. The disease affects the articular cartilage or
the entire joint, including the subchondral bone, joint capsule,
synovium and muscles around the joint (1). The cause of primary OA remains not
fully understood, and its occurrence and development is a
long-term, chronic and progressive pathological process. OA is
often considered to be the result of an interaction of multiple
pathogenic factors, including mechanical and biological factors.
Among them, age is the main risk factor, and the other factors
include trauma, obesity, genetics, inflammation and metabolism
(2). The earliest and most
important pathological changes in OA occur in cartilage.
Mesenchymal stem cells (MSCs) can differentiate into chondrocytes
(3). Therefore, MSC aging may be
related to the occurrence and progression of OA. The present review
aimed to describe the association between OA and cellular aging in
terms of genetics, epigenetics, and single-cell sequencing,
focusing on the association between the aging of MSC and OA
(Fig. 1).
The main types of aging include replication, DNA
damage-induced, oncogene-induced, oxidative stress-induced,
chemotherapy-induced, mitochondrial dysfunction-associated,
epigenetically-induced [DNA methylase inhibitors or histone
deacetylase (HDAC) are also known to cause senescence], and
paracrine senescence (senescence-associated secretory
phenotype-induced senescence) produced by primary senescent cells
(4). Cellular senescence refers
to the permanent growth arrest in cells with the ability to
proliferate and respond to various cellular stresses. It promotes
tissue remodeling during development and after an injury, but may
also lead to decreased regenerative potential and function of
tissues and tumorigenesis of inflammatory and senescent organisms
(4). Markers of aging include
genomic instability, telomere depletion, epigenetic alterations,
loss of protein balance, malfunction of nutrient perception,
mitochondrial dysfunction, cellular senescence, stem cell failure
and altered intercellular communication (5). Numerous factors associated with
aging, such as mitochondrial dysfunction, oxidative stress and
cellular senescence, contribute to OA.
Cellular senescence is one of the hallmarks of
senescence, and a key feature of senescent cells is the secretion
of a range of pro-inflammatory cytokines, chemokines and growth
factors such as interleukin (IL)-6, IL-1β, known as the
senescence-associated secretory phenotype (SASP) (6). SASP is observed in numerous
senescent cell types, including fibroblasts and MSCs (7,8).
Phenotypic genes (IL-1β, IL-6, IL-7, IL-8, MMP family) associated
with senescent SASP show increased expression in cartilage and
synovial tissue in patients with OA (9). These SASP factors may be produced
in part by OA chondrocytes (10). In animal experiments, expression
levels of the inflammatory markers matrix metalloproteinase 13
(MMP13) and IL-1β were reduced when senescent cells were removed by
transgenic mouse models or drug intervention (11). Both expressions are
manifestations of OA. Therefore, OA may be associated with the
senescence of chondrocytes.
Stem cells can be roughly divided into embryonic
stem cells and adult stem cells according to the source, and MSCs
belong to adult stem cells, which are stromal cells with a
self-renewal ability and exhibit multidirectional differentiation.
MSCs can be isolated from a variety of tissues, including the
umbilical cord, endometrial polyps, menstrual blood, bone marrow
and adipose tissue (12). MSCs,
with a multidirectional differentiation capacity, can differentiate
into all mesoderm-derived cell types, such as fat cells,
osteoblasts and chondrocytes (3). The function of MSCs declines with
age, a process known as aging. This may be associated with a loss
of tissue homeostasis maintenance, leading to organ failure and
aging diseases (13,14) such as OA. The clinical value of
MSCs comes primarily from their non-stem cell/progenitor cell
properties. That is, MSCs produce extracellular vesicles, including
exosomes (described below), several cytokines and growth factors
involved in regulating tissue metabolism and tissue repair and
regeneration after injury (15),
as in OA.
OA affects joint cartilage or the entire joint, and
cartilage damage is an important part of the development of OA.
Potential mechanisms that contribute to OA include age-related
inflammation, cellular senescence (including SASP), mitochondrial
dysfunction and oxidative stress, energy metabolism dysfunction
(associated with decreased autophagy) and alterations in cell
signaling (16). In recent
years, numerous studies have suggested that chloride ion channels
may also be one of the mechanisms that lead to the development of
OA (17). Cartilage damage,
meaning loss of the extracellular matrix (ECM) of chondrocytes, is
a key early feature of OA, characterized by inadequate ECM
synthesis and degeneration of articular cartilage. Cartilage
consists of a single cell type, the chondrocytes (18), which are surrounded by a large
ECM. The ECM consists of two main components, mainly collagen
(types II, IX and XI) and proteoglycans (mainly aggrecan) (19-21), the gene expression of which is
controlled by transcription factors SRY-related HMG-box (SOX)-5,
SOX-6, and SOX-9 (22).
Aggrecan, a major proteoglycan in the articular cartilage, is
cleaved at a specific 'aggrecanase' site in human osteoarthritic
cartilage; this cleavage is performed by several members of a
disintegrin and metalloproteinase with thrombospondin motifs
(ADAMTS) family of metalloproteases, with ADAMTS-5 playing a major
role (23). Matrix
metalloproteinases (MMPs) are responsible for the breakdown of
chondro-collagen, with MMP-13 playing a central role (24).
OA is characterized by accumulation of senescent
cells and the wearing off of the protective cartilage. A previous
study showed an increase in the number of senescent cells in
articular cartilage, synovial membrane, and fat pad tissues during
OA progression (25). The
development and progression of OA may be caused by the accumulation
of senescent cells within or near the joint (6). With age, the proliferation and
synthesis capacity of articular chondrocytes decreases, the aging
chondrocytes are unable to produce and repair the ECM (26). Characteristic changes in
senescent chondrocytes include increased production of cytokines
(for example, IL-1β, IL-1 and IL-6) and MMP, increased levels of
senescence-associated beta-galactosidase, p53, p21 and p16 and
decreased collagen II (COL2) synthesis and sirtuin 1 (SIRT1)
(10,26). In OA, joint cartilage degradation
triggers an inflammatory response and cytokine production in the
tissues around the joint. These inflammatory molecules stimulate
further ECM catabolism by increasing protease synthesis in
chondrocytes (27). Senescent
fibroblasts and MSCs can also produce a range of pro-inflammatory
cytokines (7,8). In addition, studies have shown that
a decrease in the level of autophagy during aging also increases
the release of pro-inflammatory factors (28). Cartilage destruction begins when
chondrocytes are stimulated by pro-inflammatory cytokines such as
IL-1β, IL-6, IL-8 and tumor necrosis factor (TNF)-α (29,30). Disruption of cartilage stromal
integrity is caused by increased catabolism/apoptosis of
chondrocytes in articular cartilage and decreased chondrocyte
anabolism (31,32). IL-1β and TNF-α are among the key
pro-inflammatory cytokines involved cartilage destruction in OA
(27). Numerous studies have
shown that elevated levels of IL-1β can be observed in the synovial
fluid, synovium, subchondral bone and cartilage in patients with OA
(33). Numerous studies have
confirmed that IL-1β interferes with the synthesis of key
structural proteins such as type II collagen and aggrecan (34,35). IL-1β induced enzymes of the MMP
family, mainly interstitial collagenase (MMP-1),
streptoglobulinase-1 (MMP-3), and collagenase 3 (MMP-13). These
enzymes affect the expression of cartilage collagen (24,36), thereby affecting the balance
between cartilage matrix synthesis and catabolism, leading to
degenerative cartilage diseases such as OA (32). In addition, IL-1β induces
chondrocytes to produce ADAMTS metalloproteinases (36), which affects aggrecan expression
and regulates cartilage metabolism (37). A previous study suggested that
IL-1β-induced chloride channel opening may also be strongly
associated with the development of OA (38). The pro-inflammatory and catabolic
effects of IL-1β and TNF are mediated by the activation of several
signaling pathways, most importantly, the nuclear factor
kappa-light-chain-enhancer of activated B cells (NFκB) signaling
pathway (39). NFκB mediates the
expression of several inflammatory genes, such as those encoding
inducible nitric oxide synthase (iNOS), cyclooxygenase 2 (COX2) and
chemokines, and also helps induce MMP-1, MMP-9, MMP-13 and ADAMTS4
(39,40). IL-1β significantly upregulated
the expression of MMP-13, NFκB, p65, etc., inhibiting the
expression of COL2 and aggrecan (ACAN) (41). In contrast, MSCs enhanced the
expression of COL2 and aggrecan and inhibited the expression of
MMP-13 and NFκB p65 in IL-1β-stimulated rat chondrocytes, partially
through the NFκB signaling pathway (41) (Fig. 2).
Additionally, elevated levels of TNF-α, IL-1, and
IL-6 in synovial fluid, synovium, subchondral bone, and cartilage
in patients with OA induce the production of other cytokines, MMPs,
and prostaglandins, and inhibit the synthesis of proteoglycans and
COL2. Chondrocytes activated by these pro-inflammatory cytokines in
turn produce MMP-1, MMP-3, MMP-13, and aggrecanase 1 and 2
(ADAMTS-4 and ADAMTS-5, respectively) expression (19). This in turn causes cartilage
degradation. Senescent cells also produce SASPs such as IL-1β, IL-6
and TNF-α, which are key pro-inflammatory cytokines associated with
OA (19). Therefore, they play a
key role in cartilage matrix degradation and bone resorption in OA
(42). The increase in
inflammatory factors above in OA reflects the senescence of
chondrocytes (characteristic changes in the senescence of
chondrocytes). Similarly, aging MSCs also produce SASP, which
affects cartilage-specific gene expression and inhibits cartilage
catabolic gene expression, thereby promoting cartilage breakdown
and reducing cartilage synthesis, which is related to the
occurrence and development of OA.
The transcription factor SOX9 is essential for
chondrocyte differentiation and cartilage formation (43,44). Sox9 was the main positive
regulator of chondrogenesis, and its regulation affected cartilage
differentiation by MSCs in a mouse model (45,46). SOX9 controlled the expression of
numerous chondrocyte genes, including its cofactors L-SOX5a and
SOX6, as well as ECM genes, such as type II collagen and aggrecan
(47), involved in regulating
cartilage formation. Chondrogenic RNA modulator (ROCR), a long
non-coding RNA (lncRNA) necessary for successful differentiation of
MSCs into chondrocytes, appeared to aid in the expression of SOX9.
Normalizing SOX9 levels by overexpression reversed the damaged
cartilage phenotype caused by ROCR depletion (47). SOX9, on the other hand, promoted
microRNA (miR)-140 expression (48,49), which in turn promotes collagen II
expression and inhibits MMP-13 and ADAMTS-5 expression (50), thereby blocking OA. However,
studies have shown no significant correlation between SOX9
expression in bone marrow and cellular senescence. Association
between aging MSCs and SOX9 expression needs further study
(51). Nuclear factor kappa-B
ligands (RANKL) are produced by osteoblasts, stromal cells, T cells
and other sources and activate RANK on the surface of osteoclasts
and osteoclast precursors. RANK activation leads to the recruitment
of the adaptor protein TRAF 6 (TNF receptor correlated factor 6),
resulting in NFκB activation (52). This may cause an imbalance in
cartilage metabolism. Studies have shown that RANKL mRNA increases
with age in bone marrow cells, and its expression in OA and
rheumatoid arthritis (RA) bone marrow cells is markedly higher than
normal (51). In animal trials,
transplantation of MSCs can reduce the expression of RANKL in rats
by downregulating the level of IL-22, thereby improving the degree
of RA bone destruction (53).
The expression of RANKL is significantly increased in senescent
MSCs (54). Therefore, senescent
MSCs may affect OA by regulating RANKL expression. Studies have
found that the expression of Runt-associated transcription factor 2
(RUNX2) is elevated in human OA cartilage (55). Runx2 is the most effective
inducer of osteoblast differentiation and a major transcription
factor for chondrocytes hypertrophy. Osteocalcin, Runx2 from
subchondral bone accumulates in the weight-bearing area of the OA
joint. This indicates an upregulation of osteoblast and
pre-osteoblastic activity, possibly due to an increased need for
bone formation due to excessive mechanical load and osteocyte death
(56), thus promoting cartilage
breakdown and aggravating OA. Studies have shown that MSCs undergo
aging and spontaneous osteogenic differentiation after regular
culture amplification, and Runx2 is upregulated (57). Upregulated RUNX2 may help
increase the expression of its downstream catabolic target MMP-13
affecting cartilage catabolism (55). miR-105 bound to and targeted
RUNX2 in chondrocytes, and the expression of miR-105 in human OA
cartilage was downregulated, which was inversely correlated with
the expression of RUNX2, ADAMTS7 and ADAMTS12 (58). The expression of these genes is
upregulated, causing cartilage destruction. Insufficiency of ECM
synthesis and articular cartilage degeneration in patients with OA,
as well as downregulation of miR-140, may be associated with a
decrease in SOX9.
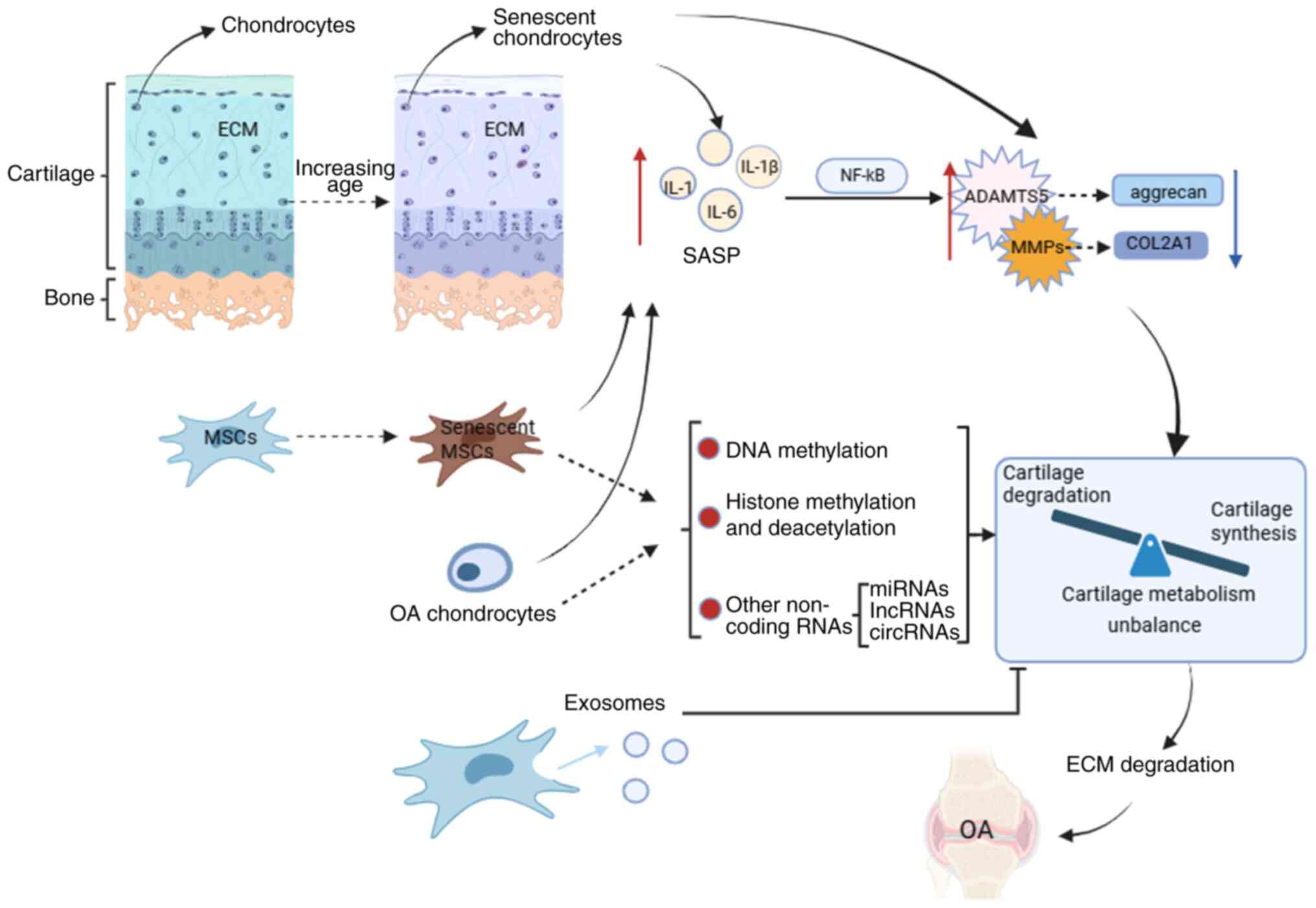
In summary, OA chondrocytes produce a large amount
of SASP, which eventually causes cartilage metabolism imbalance
through different signaling pathways, that is, cartilage catabolism
is greater than cartilage anabolism, resulting in cartilage
destruction, which in turn aggravates OA Senescent MSCs and
chondrocytes can also upregulate the expression of cartilage
catabolic genes by producing SASPs, affecting cartilage metabolism
(Fig. 1).
Over the past decade, OA genetics has changed
through the application of large-scale genome-wide association
scans (GWAS). So far, more than 100 polymorphic DNA variants have
been linked to OA (59).
Interpreting the results of GWAS is biologically challenging,
therefore translating gene discoveries into effective therapies
remains elusive for now. The OA GWAS on the UK Biobank dataset is
the largest to date, surveying over 77,000 OA patients and
identifying 52 new risk loci (60). For example, rs75621460 (hip
and/or knee OA, single variant in the 95% credible set) is an
intergenic variant located downstream of CCDC97 and TGF-β1
(60). SMAD3 (known loci:
rs12901372) encodes a transcriptional regulator, plays a key role
in cartilage differentiation, and regulates TGF-β1 expression
(61). TGF-β1 regulates
articular cartilage metabolism (as aforementioned). In addition,
the TGF-β1/SMAD3 pathway regulates the expression of miR-140 in OA
(62), affecting OA progression.
RUNX2 (known loci: rs2064630) encodes transcription factor
necessary for osteoblast differentiation and chondrocytes
maturation (63), and is
downregulated by TGF-β1 (64).
Senescent MSCs also produce pro-inflammatory cytokines and,
therefore, may have partial crosstalk with genetic locus
variation.
In the future, with the application of large-scale
GWAS, more risk sites will be discovered, which means that more
therapeutic targets will be discovered. At that time, new
treatments for OA will continue to emerge.
Epigenetics is defined as a heritable change in gene
expression caused by external or environmental factors rather than
a change in the DNA sequence (65), mainly including DNA methylation,
histone modification and non-coding RNA (detailed below) (65). Changes in age-related epigenetic
processes may be a potential cause of delayed human diseases such
as OA. In 2017, Simon and Jeffries summarized epigenetic changes in
OA such as low methylation, low HDAC-1 level, increased HDAC-2
level and downregulated miR-140 (66).
DNA methylation is a reversible process catalyzed by
DNA methyltransferases (DNMTs) resulting in the formation of
5-methylcytosine (5 mC), that is, methylation on the fifth carbon
of the cytosine residue at the CpG dinucleotide (67). Overall, DNA methylation in OA is
reduced (66). During MSC
cartilage development, considerable demethylation changes occur in
the epigenetic landscape, especially at sites characterized by
enhancer modifications (68).
Differential DNA methylation of inflammatory factors is associated
with OA in human chondrocytes. For example, IL8 showed 38-fold
higher expression in patients with hip OA than in the control
group, and in vitro DNA methylation was noted to reduce
basal IL8 promoter basal activity. Upregulation of IL-8 reduces the
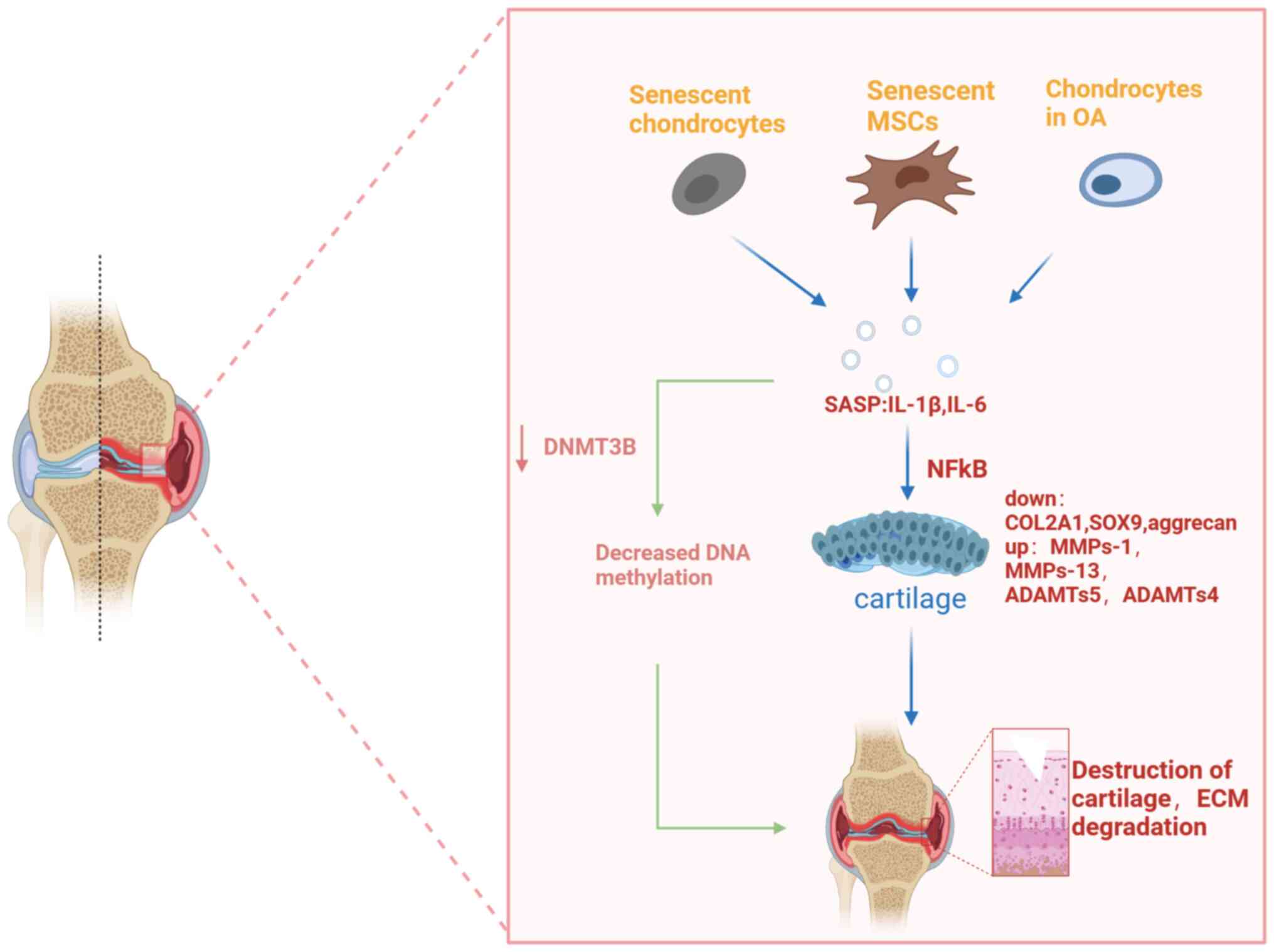
expression of DNMT3B in OA chondrocytes (69), causing decreased DNA methylation.
The pro-inflammatory cytokine IL-1β may downregulate DNMT3B in
mouse and human chondrocytes via the NFκB (70). Long-term repetitive stimulation
of primary generational chondrocytes with inflammatory cytokines
induced demethylation of specific CpG sites in the proximal
promoter of IL-1β, resulting in an increased and sustained IL-1β
expression (71). The expression
of inflammatory factors is upregulated, which in turn leads to an
imbalance in joint cartilage metabolism through the NFκB pathway.
The increase in cartilage degrading enzymes in advanced OA may be
due to epigenetic changes in the methylation state of CpG sites in
the promoter regions of these enzymes (72). In OA, numerous chondrocytes
undergo phenotypic changes and acquire gene expression banks
characterized by abnormal expression of numerous catabolic genes,
including MMPs, aggregation enzymes (ADAMTS-4 and -5), iNOS, IL-1β
and other cytokines (73,74).
These pro-inflammatory cytokines work together with abnormally
expressed cartilage catabolic genes to accelerate degradation of
chondrocytes. Aging MSCs likewise produce numerous pro-inflammatory
factors (SASPs) and may therefore affect OA by influencing DNA
methylation (Fig. 2).
Histones are highly conserved proteins whose
function is to stabilize, organize and concentrate DNA within a
limited range of the nucleus. They consist of duplicate octamers
containing dimers of each protein in four core histones (H2A, H2B,
H3 and H4) and encapsulate the genomic DNA on its outer surface
(66). Acetylation neutralizes
the positive charge on the histone by oxidizing the amine residue
into an amide and reduces the histone's ability to bind to DNA;
this prevents chromatin shrinkage and allows the gene transcription
machinery to enter the underlying DNA and transcribe. Deacetylation
presents a positively charged histone tail, promoting high-affinity
binding between the DNA backbone and histone, resulting in
chromatin condensation, thereby blocking transcription (66).
HDACs constitute a family of enzymes responsible for
histone and non-histone deacetylation, whereas histone
acetyltransferase catalyzes the reverse reaction, that is,
acetylation. HDACs catalyze the removal of the acetyl functional
groups from lysine residues of histones and non-histones. HDAC
enzymes are divided into four classes: Class I Rpd3-like proteins
(HDAC1, HDAC2, HDAC3 and HDAC8); class II Hda1-like proteins
(HDAC4, HDAC5, HDAC6, HDAC7, HDAC9 and HDAC10); class III Sir2-like
proteins (SIRT1, SIRT2, SIRT3, SIRT4, SIRT5, SIRT6 and SIRRT7); and
class IV proteins (HDAC11) (75). Studies have shown elevated levels
of HDAC1 and HDAC2 in chondrocytes and synovial membranes of
patients with OA than in the control group; the new
carboxyl-terminal domain of HDAC1 and HDAC2 worked in tandem with
the transcriptional inhibitor snail 1 to inhibit the expression of
the collagen α1(II) gene (COL2A1) and aggrecan (76,77). Knockdown of human chondrocyte
HDAC3 led to upregulated expression of cartilage oligomeric matrix
protein (COMP), COL2A1, SOX9 and aggrecan, and downregulated
expression of COL10 (78), whose
overexpression may contribute to OA progression. miR-193b promoted
cartilage differentiation of hMSCs by inhibiting HDAC3 expression,
thereby maintaining cartilage-specific gene expression (78). Studies have shown that exogenous
HDAC4 reduces the transcription of RUNX2, MMP1, MMP3, MMP-13,
X-collagen, Indian hedgehog signal and ADAMTS-4 and -5, and
increases the transcription of COL2. Furthermore, overexpression of
HDAC4 not only reduced the expression of IL-1β, COX2 and iNOS and
increased the expression of aggrecan, but also partially blocked
the effect of IL-1β on the catabolic events in human OA
chondrocytes (79); therefore,
it may slow down the progress of OA. Studies have shown that high
cartilage HDAC7 expression levels in human OA may lead to cartilage
degradation by promoting MMP-13 gene expression (80). IL-1β upregulated IL-6 and IL-8
expression in synovial MSCs (SMSCs) through the NFκB pathway, and
HDAC10 overexpression promoted the IL-6, IL-8 and IL-1β-mediated
activation of the NFκB pathway (81). HDAC10 upregulation contributes to
the activation of IL-1β-mediated SMSC inflammation (81), induces cartilage catabolic gene
expression and causes cartilage metabolism imbalance. A decrease in
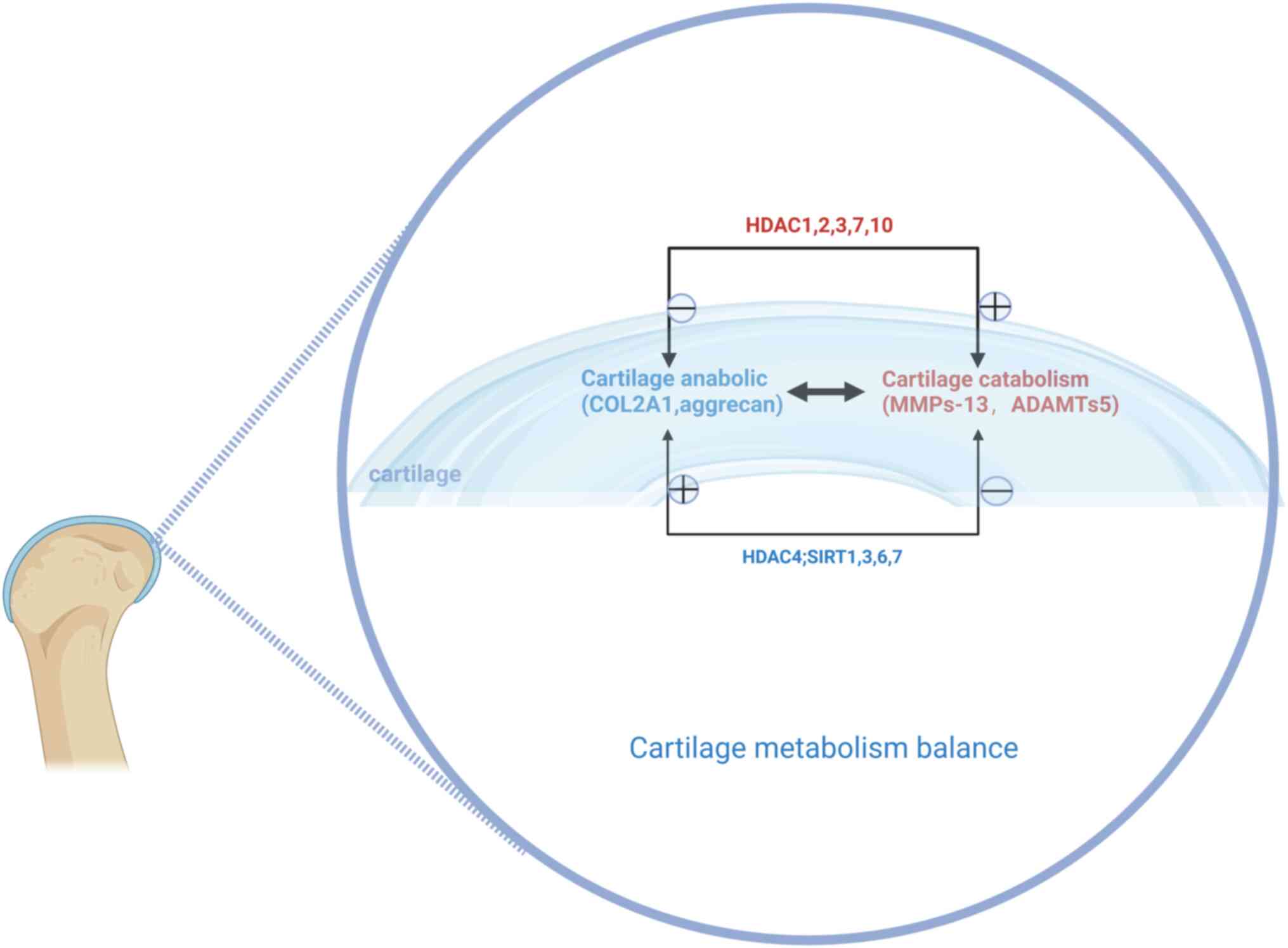
the expression of HDACs can be observed in senescent MSCs (82). These studies suggested that aging
MSCs may affect cartilage catabolism by influencing expression of
HDAC1, 2, 3, 7 and 10, promoting OA progression, and HDAC4 helps
mitigate this process (Fig.
3).
Histone methylation and acetylation are also
important epigenetic regulators during cartilage differentiation
(65,97). Similar to DNA, histones can be
methylated by histone methyltransferase and demethylated by histone
demethylase, both of which can alter gene transcription (66). Decreased expression of HDACs is
observed in senescent MSCs (82). Decreased H3K9 methylation
increased the transcription of MMP-1 and MMP-13 in chondrocytes.
Transcription of the anabolic factors SOX9 and COL2A1, which are
involved in cartilage differentiation, were also reduced by
treatment with histone methyltransferase inhibitor chaetocin
(98). Therefore, histone
methylation contributes to cartilage differentiation. Lysine
demethylase 6B (Kdm6b), also known as Jmjd3, has been identified as
an H3K27 demethylase that catalyzes H3K27me2/3 demethylation;
knockdown of Kdm6b in chondrocytes led to abnormal cartilage
development and accelerated OA progression by inhibiting the
anabolic metabolism of chondrocytes (65). Kdm6b promoted chondroblast
proliferation and hypertrophy during intrachondral osteogenesis in
Kdm6b−/− mice (99).
Depletion of Kdm6A inhibited the expression of SOX9, COL2A1 and
ACAN, resulting in an increase in H3K27me3 and a decrease in
H3K4me3 levels (100).
Similarly, KDM4B is a histone demethylase that mediates
transforming growth factor (TGF)-induced SOX9 activation by
removing H3K9me3 from the SOX9 promoter (101). Both KDM4B and KDM6B promoted
osteogenesis differentiation of human MSCs (102). The pro-inflammatory cytokine
TGF-β1 promotes upregulation of KDM4B and KDM6B (103). Thus, senescent MSCs can affect
articular cartilage metabolism and OA by releasing SASPs, affecting
histone methylation.
The main types of non-coding RNAs include microRNAs
(miRNAs), lncRNAs, and circular RNAs (circRNAs) (104). Non-coding RNA is divided into
small RNA (<200 nucleotides) and long RNA (>200
nucleotides).
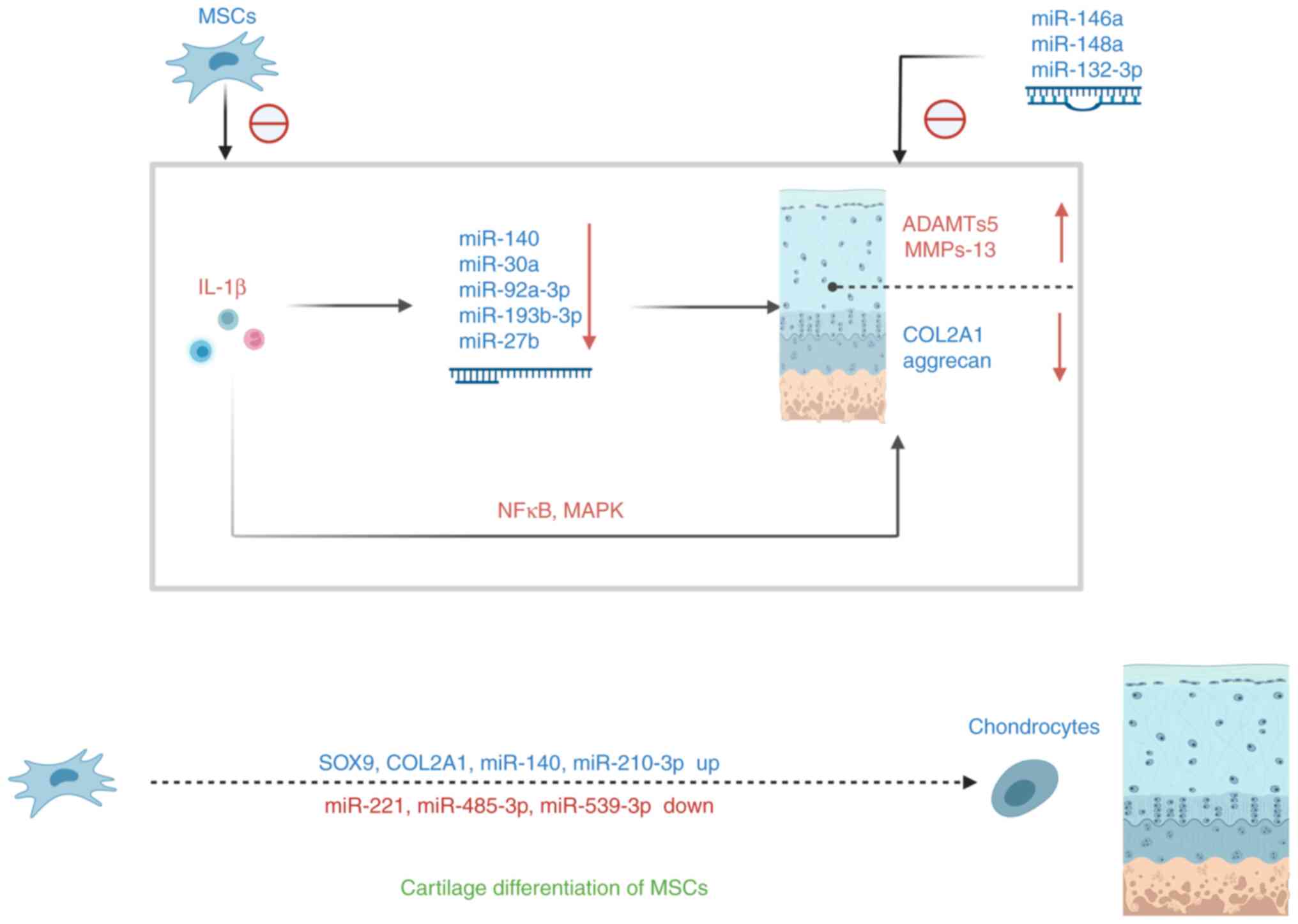
Aging MSCs cause the downregulation of the
corresponding miRNA by producing SASP, thus affecting cartilage
metabolism. Other miRNAs also affect OA through various mechanisms,
such as miR-145, which inhibited cartilage differentiation of
TGF-β3-induced MSCs by directly targeting SOX9 at the
post-transcriptional level, resulting in its downregulation
(114). miR-146a antagonized
the expression of IL-1-induced cartilage degrading enzymes MMP-13
and ADAMTS5 in articular cartilage (115). Overexpression of miR-148a
increased COL2A1 expression and decreased COL10A1, MMP-13 and
ADAMTS5 gene expression (116).
Inhibition of miRNA-495 relieved IL-1β-induced chondroblastic
inflammatory response by salvaging SOX9 expression (117). miR-129-5p decreased in patients
with OA and IL-1β-induced chondrocytes; miR-129-5p in exosomes from
human SMSC (hSMSC-Exo) inhibited IL-1β-mediated OA by inhibiting
the release of high mobility group box protein 1 (HMGB1) (118). Overexpression of miRNA-615-3p
led to increased expression levels of inflammatory cytokines such
as IL-1, IL-6 and IL-α (119).
Therefore, miRNA-615-3p overexpression may also affect articular
cartilage metabolism through the NF-kB pathway. Human bone marrow
MSCs (hBMSCs) exhibited typical MSC differentiation potential.
During cartilage differentiation, the expression of collagen 2 and
10 (COL2 and COL10), SOX9, and RUNX2 are upregulated. Expression
levels of miR-140, miR-143 and miR-181a increased over time,
whereas those of miR-27b, miR-221 and miR-615-3p decreased
(120) during cartilage
differentiation in BMSC. miR-26b expression was significantly
downregulated during the in vitro cartilage differentiation
of rat MSCs, and it played an inhibitory role in the process by
inhibiting Wnt expression (121). miR-485-5p levels were inversely
correlated with the degree of differentiation of BMSCs; miR-485-5p
reduced SOX9 levels, promoted the synthesis of cartilage surface
inflammatory factors, and blocked mouse BMSCs from differentiating
into chondrocytes (122).
miR-539-3p is gradually downregulated during cartilage
differentiation of human adipose-derived MSCs (hADMSCs); its
overexpression inhibited cartilage differentiation of
TGF-β1-induced hADMSCs by reducing gene and protein expression of
cartilage differentiation markers COL2A1 and ACAN, with SOX9 being
a direct target gene for miR-539-3p. During cartilage
differentiation of hADMSCs, the expression of SOX9 is gradually
upregulated over time (123).
miR-210-3p promotes cartilage differentiation of rat BMSCs and
promotes mRNA and protein levels of cartilage expression genes COL2
and SOX9 (124). miR-130b
inhibitors induce chondrocyte differentiation and chondrocyte
growth of BMSCs by targeting SOX9 (125). Long intergenic non-protein
coding RNA, a regulator of reprogramming (Linc-ROR), is
downregulated in the articular cartilage tissue of patients with
OA; it promotes cartilage differentiation and cartilage formation
by BMSCs by acting as competitive endogenous RNAs for miR-138 and
miR-145 and activating SOX9 expression (126). A previous study showed
overexpression of miR-122-5p by indolol 2,3 dioxygenase 1 in
synovial fluid in patients with OA, activating the Wnt1/β-catenin
pathway to impair cartilage differentiation and cartilage
regeneration of MSCs (127).
ThesemiRNA changes induce mesenchymal stem cell cartilage
differentiation through various pathways (Fig. 4). Senescent MSCs release
pro-inflammatory cytokines that mediate cartilage damage and
apoptosis and can partially alleviate cartilage damage by
modulating miRNAs expression.
LncRNAs can be divided into small (200-950 nt),
medium (950-4,800 nt), and large lncRNAs (~4,800 nt) (128), and are rich in MSC-derived
exosomes. Numerous lncRNAs regulate the chondrogenesis of MSCs. For
example, lncRNA ZBED3-AS1 induced chondrogenesis of MSC of human
synovial fluid origin (SFMSCs) (129). LncRNA ROCR contributed to SOX9
expression and cartilage differentiation of hMSC (47). LncRNA GRASLND (originally named
RNF144A-AS1) acted as a regulator of MSC chondrogenesis (130). LncRNA DANCR was involved in
SFMSC proliferation and chondrogenesis. Mechanically, DANCR acted
as a sponge RNA of miR-1275, regulating the expression of the
target gene MMP-13 (131). The
expression of lnc-RNA BLACAT1 in inflammatory BMSCs increased, and
the knockdown of BLACAT1 promoted proliferation and osteogenesis
differentiation of BMSCs targeting miR-142-5p (132). LncRNA XIST silencing promoted
cartilage differentiation of SMSCs. In addition, XIST regulated the
expression of ADAMTS-5 by directly binding to miR-27b-3p (133). TIMP-3 (a natural inhibitor of
MMPs) expression may be downregulated by recruiting DNMT1, DNMT3A
and DNMT3B, thereby increasing the methylation ratio of CpG islands
in the promoter region of TIMP-3, thereby increasing collagen
degradation in OA chondrocytes (134). MSC-derived exosomes
(MSC-exo)-mediated lncRNA KLF3-AS1 inhibited autophagy and
apoptosis of IL-1β-treated chondrocytes via the PI1K/Akt/mTOR
signaling pathway (135).
Exosome lncRNA-KLF3-AS1 from hMSCs is significantly enriched in
MSC-exo, which improves IL-1β-induced cartilage damage; exosome
lnc-KLF3-AS1 inhibited IL-1β-induced chondrocyte apoptosis
(136). Overexpression of
lncRNA CTBP1-AS2 downregulated miR-130a by increasing methylation
levels of the miR-130a gene, ultimately leading to a decrease in
the rate of chondrocyte proliferation in patients with OA (137). These studies have shown that
lncRNAs can regulate chondrogenesis of MSCs, and some lncRNAs can
regulate the expression of cartilage-specific genes and catabolic
genes through various mechanisms. SASP produced by aging MSCs
induces cartilage damage and apoptosis, aggravating OA. Numerous
lncRNAs can delay OA progression by regulating cartilage formation
of MSCs.
Cyclic RNAs (circRNAs) are a new class of discovered
non-coding RNAs with structural stability. For example, Circ_ATRNL1
is significantly higher in cartilage tissues of patients with OA.
Circ_ATRNL1 overexpression enhanced proliferation and
differentiation of hADMSCs into chondrogenesis, promoted the
expression of COL2, ACAN and SOX9, and inhibited the adipose
differentiation of hADMSC and the expression of adipose-related
genes. miR-145-5p is a target miRNA for circ_ATRNL1 and SOX9
(138). A miR-145-5p mimic
inhibited the differentiation of cartilage by hADMSC and the
expression of cartilage-related factors. The miR-145-5p mimic
effectively reversed the regulatory effect of circ_ATRNL1 on
hADMSC. Circ_ATRNL1 upregulated SOX9 expression to promote
cartilage differentiation of hADMSCs mediated by miR-145-5p
(138). MSCs-circHIPK3-EV
[extracellular vesicles (EV) from MSCs overexpressing circHIPK3]
significantly improved IL-1β-induced chondrocyte damage (139). Similar to lncRNA and miRNAs,
circRNAs can also alleviate OA by modulating the expression of
cartilage-specific genes and catabolic genes. Therefore, by
regulating the expression of non-coding RNA, it affects the
expression of cartilage-specific genes (COL2A1 and aggrecan) and
cartilage catabolic genes (MMP-13 and ADAMTS5) in OA.
In summary, epigenetic changes such as decreased DNA
methylation, histone acetylation and methylation, and regulation of
non-coding RNA greatly affect the articular cartilage metabolism in
OA. Epigenetic changes that occur in senescent cells can also
affect joint cartilage metabolism. In the future, the development
of therapies targeting cellular senescence is expected to become a
specific treatment for OA.
Exosomes are tiny membrane-bound vesicles released
from cells. Exosomes from different types of MSCs, including BMSCs,
SMSCs, ADMSCs and embryonic MSCs (EMSCs) regulated cartilage
regeneration and slowed OA progression, whereas exosomes isolated
from the synovial fluid stimulated the release of inflammatory
cytokines and MMPs (140).
MSC-EV promoted the expression of Col2A1, sox9 and
Acan in mouse chondrocytes in OA models, while negatively
regulating the expression of Mmp-13 and Runx2. MSC-circHIPK3-EV
significantly improved IL-1β-induced chondrocyte damage (139), and MSC-KLF3-AS1-exo (exosomes
from KLF3-AS1-overexpressed-MSCs) improved IL-1β-induced
chondrocyte damage by participating in MSC-mediated chondrocyte
proliferation induction and chondrocyte apoptosis inhibition via
the miR-206/G protein-coupled receptor kinase interaction protein-1
axis (141). Expression of
exosome miR-92a-3p in MSC cartilage exosomes is elevated, whereas
that in exosomes secreted by OA chondrocytes is significantly
reduced. MSC-miR-92a-3p-exo promoted cartilage proliferation and
matrix gene expression in MSCs and OA primary human chondrocytes
(PHCs), respectively. Conversely, treatment with
MSC-anti-miR-92a-3p-exo inhibited cartilage differentiation and
reduced cartilage stromal synthesis by enhancing the expression of
WNT5A (142).
Exosomes from hBMSCs overexpressed miR-92a-3p and
directly targeted the WNT5A gene to increase the expression of
chondrocyte markers (for example, COL2 and SOX9) and reduce
catabolic markers (for example, MMP-13 and RUNX2), thereby
increasing the proliferation of chondrocytes and protecting
chondrocytes from apoptosis (142). hBMSC-derived exosome-metastatic
miR-361-5p mitigated chondrocyte damage and inhibited the NFκB
signaling pathway by targeting DDX20. Inhibition of NFκB signaling
reversed the effect of overexpressed DDX20 on IL-1β-induced
chondrocyte damage. In addition, the exosome miR-361-5p reduced OA
damage in vivo (143).
Similarly, BMSC-derived exosomes and microvesicles/microparticles
re-induced the expression of chondrocyte markers (COL2 and ACAN)
while inhibiting catabolism (MMP-13 and ADAMTS5) and inflammatory
(iNOS) markers (144). Thus,
inhibiting IL-1β-induced chondrocytes destruction. BMSC-derived
exosomes are rich in miR-125a-5p, which is endowed with features
that accelerate chondrocyte migration; this is accompanied by a
higher expression of COL2, ACAN and SOX9 and lower expression of
MMP-13 in vitro, and also relieve ECM degradation of
chondrocytes (145). In
clinical samples of traumatic OA cartilage tissue, ELF3 expression
increased and miR-136-5p expression decreased. BM-MSC-exo showed
high levels of miR-136-5p, which was internalized by chondrocytes.
miR-136-5p promoted the migration of chondrocytes, wherein the
expression of COL2, ACAN and SOX9 increased and that of MMP-13
decreased, verifying that miR-136-5p targeted ELF3 and
downregulated its expression (146). Human ADMSC-EV not only promoted
the proliferation and migration of human OA chondrocytes, but also
maintained the chondrocyte matrix in the presence of IL-1β by
increasing COL2 synthesis and reducing the expression of MMP-1,
MMP-3, MMP-13 and ADAMTS-5 (147). Exosomes derived from SMSCs
overexpressing miR-155-5p promoted the proliferation and migration
of OA chondrocytes, inhibited apoptosis and enhanced ECM secretion,
thus effectively blocking OA in mouse models (148). In addition, the SMSC-EV carried
miR-26a-5p into chondrocytes, upregulated miR-26a-5p, and inhibited
phosphatase and tension homologs, thereby inhibiting apoptosis and
inflammation and improving cartilage damage in OA (149). SMSC-EV mitigated chondrocyte
damage during OA by miR-130b-3p-mediated inhibition of the
LRP12/AKT/β-catenin axes (150). miR-129-5p in hSMSC-Exo
alleviated IL-1β-mediated OA by inhibiting the release of HMGB1,
while downregulating the expression of iNOS, COX2, MMP-13, and NFκB
(118). The SMSC-derived
exosome miR-320c enhanced chondrogenesis by targeting ADAM19,
highlighting the potential new mechanism of SMSC in the treatment
of OA (151). Co-culture of
ADMSCs and SMSCs with chondrocytes reduced MMP-13 expression, while
increasing the expression of COL2A1, ACAN and SOX9, and thus,
reversed the IL-1β effect on promoting reactive oxygen species
content and inflammatory factor levels (152). EMSC-exo also had a beneficial
therapeutic effect on OA by increasing the expression of COL2 in
the cartilage matrix and reducing the expression of ADAMTS5
(153). Exosome overexpression
of miR-140-5p from human urine-derived stem cells enhanced
cartilage regeneration and subchondral bone remodeling (154). Exosomes from
miR-95-5p-overexpressing primary chondrocytes (AC-miR-95-5p)
enhanced chondrogenicity and prevented the development of OA by
directly targeting HDAC2/8 (155). The role of exosomes in OA
discussed in this review is shown in (Table I).
The aforementioned studies have shown that
MSCs-derived exosomes improve OA by regulating cartilage
development and cartilage injury repair through mechanisms such as
altering miRNA, lncRNA and circRNA expression (Fig. 5). This has become a research
hotspot in recent years. There is growing evidence that exosomes
can effectively reduce inflammation, promote cartilage regeneration
and reduce pain in OA patients. In the future, the potential of
exosomes in the treatment of OA should be used to a full extent
towards the development of novel specific exosome therapies.
Single-cell RNA sequencing (scRNA seq) assesses
individual cells, identifies new cell populations, reveals
regulatory relationships between genes and tracks the trajectory of
different cell lineages in development and even disease (156). This contributes to a
comprehensive understanding of the pathophysiology of the disease
and facilitates the development of effective treatment options. In
terms of cartilage development, scRNA seq helps identify
self-renewing and pluripotent human bone stem cells that produce
bone, cartilage and stromal progenitor cells (157). Previous studies identified four
chondrocytes subclusters including proliferative chondrocytes
(ProCs), pre-hypertrophic chondrocytes (preHTCs), HTCs and
fibrochondrocytes (FCs). HDAC4 expressed in preHTCs regulates
chondrocytes hypertrophy and endochondral bone formation by
interacting with the transcription factor Runx2, which is necessary
for chondrocytes hypertrophy, and inhibiting its activity (158). Overexpression of HDAC4 in ProCs
in vivo inhibits chondrocytes hypertrophy and
differentiation (158). HDACs
expression decreased in aging MSCs (82). Therefore, aging MSCs may promote
chondrocytes to hypertrophy and aggravate OA. HTCs attract blood
vessels and osteocytes to invade and then undergo apoptosis and
calcium deposition, ultimately triggering the progression of OA in
humans (159). FCs are mainly
present in the late stages of OA and have a high proportion of
genes and angiogenesis ability associated with adverse OA outcomes,
indicating that FCs promote OA progression (159). Previously, a study using scRNA
seq on OA cartilage identified three new subsets of cartilage in
cartilage, effector chondrocytes (ECs), regulatory chondrocytes
(RegCs) and homeostatic chondrocytes (HomCs), and highlighted some
changes associated with OA progression (159). ECs are associated with the
tricarboxylic acid cycle and amino acid metabolism, suggesting that
they provide cellular energy (159). Another study showed that ECs
mainly exert immune function, causing tissue inflammation, and
possibly promoting the development of OA (160). HomCs exhibit high expression of
genes related to cell cycle regulation, metabolic processes and
development in response to external stimuli (159,161). RegCs are involved in numerous
signaling pathways and may play a key role in regulating OA
progression (159). In
addition, Ji et al (159) identified two hypertrophic
chondrocytes subclusters: HTC-A is highly expressed in genes
associated with cartilage development and connective tissue
development; HTC-B is highly expressed in genes associated with ECM
tissue, ossification and mineralization. In 2021, Sebastian et
al (162) identified nine
chondroblast subtypes (Ucmahigh, Cytl1high,
Chil1high, Mef2chigh, Krt16high,
Tnfaip6high, S100a4high, Neat1high
and divC chondrocyte clusters) in the knee cartilage of healthy
mice by analyzing cells from each cluster separately. MMP-3, Inhba,
Sfn, Il11, Ptgs2, Dusp2 and MMP-13 are upregulated among various
chondrocytes subtypes, while Cytl1, Il17b, Fgfr2, Ptch1, Dbp and
Rad are downregulated. The Ucmahigh and
Krt16high clusters have unique ECM signatures with
increased expression of several key ECM proteins, including COL2A1,
COL9A1-a3 and ACAN (162).
Previous studies have found elevated expression of Mmp3, Mmp13,
Ptgs2, Il11 and Inhba in OA (163,164). The results of Sebastian et
al (162) confirmed that
joint chondrocytes are the main source of this elevated expression
in injured joints. SPP1 was found to be highly expressed in OA
cartilage, and Qu et al (165) identified unique chondrocytes
characterized by high SPP1 expression. Gao et al (166) performed scRNA seq on human
articular chondrocytes treated with IL-1β and it found that one of
the initial cell clusters can be transformed into a
pro-inflammatory subpopulation through a pathway called an
inflammatory response, which may be a central target for mitigating
OA progression. Yoshimoto et al (167) demonstrated by analyzing scRNA
seq datasets that cellular senescence signals in chondrocytes are
associated with OA pathogenesis. The emergence of scRNA seq has
broadened our understanding of chondrocytes subsets. scRNA seq can
also play an important role in exploring the association between
MSC aging and OA pathogenesis. In the future, improved
identification of the functional roles of subpopulations and the
description of relevant molecular mechanisms are needed in order to
improve understanding of the pathogenesis of OA, identify new
therapeutic targets and develop specific therapies.
The present review mainly explored the relationship
between mesenchymal stem cell senescence and genetic and epigenetic
changes in the progression of OA from the perspective of
pro-inflammatory cytokines. OA progression is associated with
genetic and epigenetic changes that occur during mesenchymal stem
cell aging.
Non-coding RNA influences OA progression by
modulating cartilage synthesis/catabolism.
The exosomes secreted by various mesenchymal stem
cell sources regulate miRNA expression to affect metabolism in
joint cartilage. Exosome treatment for OA is a promising future
direction.
OA is a disease associated with age and cartilage
destruction. Pro-inflammatory cytokines play a pivotal role in the
development of OA. Cytokines activate white blood cells, which
produce more cytokines. Therefore, even a small pro-inflammatory
stimulus may produce a more systemic chronic inflammatory response.
Spontaneous differentiation and aging of MSCs occur during
expansion, and little is currently known about the molecular
mechanisms involved. Aging MSCs release pro-inflammatory cytokines
that cause an inflammatory storm and aggravate the progression of
OA. More studies are needed to determine the molecular mechanisms
of MSCs aging to identify new specific targets and promote
development of specific OA therapies. With the application of
large-scale GWAS, the role of OA genetics in OA is gradually
emerging. The proportion of known OA risk loci in heritability is
just >20%, and there are still a large number of loci to be
discovered. Interpreting the results of GWAS is biologically
challenging, therefore translating genetic discoveries into
effective treatments remains elusive for now. In the future, as
more GWAS sites are reported, the gap between discovery and utility
will narrow. Epigenetics is a key player in the formation and
maintenance of joints. OA is associated with alterations in
numerous epigenetic markers in the affected tissues, such as DNA
methylation, histones and non-coding RNA. Exosome therapy for OA
has shown exciting promise over the past few years, with
MSC-derived exosomes becoming potential cell-free therapies for OA
because of their anti-inflammatory properties as well as unique
advantages and characteristics. The therapeutic effect of
MSC-derived exosomes is similar to that of MSCs. Numerous studies
have demonstrated that MSC-derived exosomes mediate cartilage
tissue repair, promote cartilage differentiation of MSCs, and
attenuate the development of OA; this suggests that MSC-derived
exosomes have exciting therapeutic potential for OA. However, the
specific signaling molecules that regulate MSC-derived exosomes in
OA are still not well understood. Clarifying the underlying
molecular mechanisms of MSC-derived exosomes in OA pathogenesis
will help identify potential therapeutic targets for OA. Despite
the great potential of exosomes in the treatment of OA, obtaining
consistent and abundant high-quality exosomes remains a technical
challenge. Obtaining high-quality exosomes maximizes their
effectiveness and ensures their safe application. Single-cell
sequencing has shone in recent years in identifying chondrocytes
subsets; however, the understanding of the molecular mechanisms of
orthopedic disease development and homeostasis by single-cell
sequencing is still in infancy. In the future, attention should be
paid to identify subsets or disease-specific subsets, key signaling
pathways and conduct in-depth research on the functions of
subpopulations and their interactions to improve understanding of
OA. Meanwhile, scRNA-seq could be combined with other traditional
and emerging technologies to complement each other to achieve more
detailed and accurate cell classification and expand our
understanding of orthopedic diseases.
Not applicable.
DT and ZH wrote the manuscript. ZZ, ZD and WL
designed and corrected the whole manuscript. DW, XC and JL proposed
the information. DW supervised the process. ZD and WL funded the
project. All authors read and approved the final version of the
manuscript. Data authentication is not applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
Not applicable.
The present study was supported by the National Natural Science
Foundation of China (grant nos. 81800785, 81972085 and 82172465),
the China University Industry-University-Research Innovation Fund
(grant no. 2021JH037), the Natural Science Foundation of Guangdong
(grant nos. 2018A0303100027 and 2021A1515010706), the Guangdong
Provincial Key Clinical Discipline-Orthopedics (grant no. 2000005),
the Sanming Project of Shenzhen Health and Family Planning
Commission (grant no. SZSM201612086), the Shenzhen Science and
Technology Planning (grant nos. JCYJ20180228163401333 and
JCYJ20190806170612680), the Shenzhen Key Medical Discipline
Construction Fund (grant no. SZXK025), the discipline construction
Capacity Improvement project of Shenzhen Municipal Health
Commission (grant no. SZXJ2018065) and the Natural Science
Foundation of Guangdong (grant no. 2023A1515010102).
|
1
|
Loeser RF, Goldring SR, Scanzello CR and
Goldring MB: Osteoarthritis: A disease of the joint as an organ.
Arthritis Rheum. 64:1697–1707. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Abramoff B and Caldera FE: Osteoarthritis:
Pathology, diagnosis, and treatment options. Med Clin North Am.
104:293–311. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lan T, Luo M and Wei X: Mesenchymal
stem/stromal cells in cancer therapy. J Hematol Oncol. 14:1952021.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hernandez-Segura A, Nehme J and Demaria M:
Hallmarks of cellular senescence. Trends Cell Biol. 28:436–453.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
López-Otín C, Blasco MA, Partridge L,
Serrano M and Kroemer G: The hallmarks of aging. Cell.
153:1194–1217. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xu M, Bradley EW, Weivoda MM, Hwang SM,
Pirtskhalava T, Decklever T, Curran GL, Ogrodnik M, Jurk D, Johnson
KO, et al: Transplanted senescent cells induce an
osteoarthritis-like condition in mice. J Gerontol A Biol Sci Med
Sci. 72:780–785. 2017.
|
|
7
|
Coppé JP, Patil CK, Rodier F, Sun Y, Muñoz
DP, Goldstein J, Nelson PS, Desprez PY and Campisi J:
Senescence-associated secretory phenotypes reveal
cell-nonautonomous functions of oncogenic RAS and the p53 tumor
suppressor. PLoS Biol. 6:2853–2868. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Xu M, Tchkonia T, Ding H, Ogrodnik M,
Lubbers ER, Pirtskhalava T, White TA, Johnson KO, Stout MB, Mezera
V, et al: JAK inhibition alleviates the cellular
senescence-associated secretory phenotype and frailty in old age.
Proc Natl Acad Sci USA. 112:E6301–E6310. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Greene MA and Loeser RF: Aging-related
inflammation in osteoarthritis. Osteoarthritis Cartilage.
23:1966–1971. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lotz M and Loeser RF: Effects of aging on
articular cartilage homeostasis. Bone. 51:241–248. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jeon OH, Kim C, Laberge RM, Demaria M,
Rathod S, Vasserot AP, Chung JW, Kim DH, Poon Y, David N, et al:
Local clearance of senescent cells attenuates the development of
post-traumatic osteoarthritis and creates a pro-regenerative
environment. Nat Med. 23:775–781. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ding DC, Shyu WC and Lin SZ: Mesenchymal
stem cells. Cell Transplant. 20:5–14. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li Y, Wu Q, Wang Y, Li L, Bu H and Bao J:
Senescence of mesenchymal stem cells (Review). Int J Mol Med.
39:775–782. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Alt EU, Senst C, Murthy SN, Slakey DP,
Dupin CL, Chaffin AE, Kadowitz PJ and Izadpanah R: Aging alters
tissue resident mesenchymal stem cell properties. Stem Cell Res.
8:215–225. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chew JRJ, Chuah SJ, Teo KYW, Zhang S, Lai
RC, Fu JH, Lim LP, Lim SK and Toh WS: Mesenchymal stem cell
exosomes enhance periodontal ligament cell functions and promote
periodontal regeneration. Acta Biomater. 89:252–264. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Loeser RF, Collins JA and Diekman BO:
Ageing and the pathogenesis of osteoarthritis. Nat Rev Rheumatol.
12:412–420. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lin Z, Deng Z, Liu J, Lin Z, Chen S, Deng
Z and Li W: Chloride channel and inflammation-mediated pathogenesis
of osteoarthritis. J Inflamm Res. 15:953–964. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xu X, Liang Y, Li X, Ouyang K, Wang M, Cao
T, Li W, Liu J, Xiong J, Li B, et al: Exosome-mediated delivery of
kartogenin for chondrogenesis of synovial fluid-derived mesenchymal
stem cells and cartilage regeneration. Biomaterials.
269:1205392021. View Article : Google Scholar
|
|
19
|
McCulloch K, Litherland GJ and Rai TS:
Cellular senescence in osteoarthritis pathology. Aging Cell.
16:210–218. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Duan L, Liang Y, Xu X, Xiao Y and Wang D:
Recent progress on the role of miR-140 in cartilage matrix
remodelling and its implications for osteoarthritis treatment.
Arthritis Res Ther. 22:1942020. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fujii Y, Liu L, Yagasaki L, Inotsume M,
Chiba T and Asahara H: Cartilage homeostasis and osteoarthritis.
Int J Mol Sci. 23:63162022. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu CF and Lefebvre V: The transcription
factors SOX9 and SOX5/SOX6 cooperate genome-wide through
super-enhancers to drive chondrogenesis. Nucleic Acids Res.
43:8183–8203. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Glasson SS, Askew R, Sheppard B, Carito B,
Blanchet T, Ma HL, Flannery CR, Peluso D, Kanki K, Yang Z, et al:
Deletion of active ADAMTS5 prevents cartilage degradation in a
murine model of osteoarthritis. Nature. 434:644–648. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang M, Sampson ER, Jin H, Li J, Ke QH, Im
HJ and Chen D: MMP13 is a critical target gene during the
progression of osteoarthritis. Arthritis Res Ther. 15:R52013.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Philipot D, Guérit D, Platano D, Chuchana
P, Olivotto E, Espinoza F, Dorandeu A, Pers YM, Piette J, Borzi RM,
et al: p16INK4a and its regulator miR-24 link senescence and
chondrocyte terminal differentiation-associated matrix remodeling
in osteoarthritis. Arthritis Res Ther. 16:R582014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Loeser RF: Aging and osteoarthritis: The
role of chondrocyte senescence and aging changes in the cartilage
matrix. Osteoarthritis Cartilage. 17:971–979. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kapoor M, Martel-Pelletier J, Lajeunesse
D, Pelletier JP and Fahmi H: Role of proinflammatory cytokines in
the pathophysiology of osteoarthritis. Nat Rev Rheumatol. 7:33–42.
2011. View Article : Google Scholar
|
|
28
|
Fang H, Deng Z, Liu J, Chen S, Deng Z and
Li W: The mechanism of bone remodeling after bone aging. Clin
Interv Aging. 17:405–415. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bian Q, Wang YJ, Liu SF and Li YP:
Osteoarthritis: Genetic factors, animal models, mechanisms, and
therapies. Front Biosci (Elite Ed). 4:74–100. 2012. View Article : Google Scholar
|
|
30
|
Molnar V, Matišić V, Kodvanj I, Bjelica R,
Jeleč Ž, Hudetz D, Rod E, Čukelj F, Vrdoljak T, Vidović D, et al:
Cytokines and chemokines involved in osteoarthritis pathogenesis.
Int J Mol Sci. 22:92082021. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hwang HS and Kim HA: Chondrocyte apoptosis
in the pathogenesis of osteoarthritis. Int J Mol Sci.
16:26035–2604. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Deng Z, Chen X, Lin Z, Alahdal M, Wang D,
Liu J and Li W: The homeostasis of cartilage matrix remodeling and
the regulation of volume-sensitive ion channel. Aging Dis.
13:787–800. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chen LX, Lin L, Wang HJ, Wei XL, Fu X,
Zhang JY and Yu CL: Suppression of early experimental
osteoarthritis by in vivo delivery of the adenoviral
vector-mediated NF-kappaBp65-specific siRNA. Osteoarthritis
Cartilage. 16:174–184. 2008. View Article : Google Scholar
|
|
34
|
Fei J, Liang B, Jiang C, Ni H and Wang L:
Luteolin inhibits IL-1β-induced inflammation in rat chondrocytes
and attenuates osteoarthritis progression in a rat model. Biomed
Pharmacother. 109:1586–1592. 2019. View Article : Google Scholar
|
|
35
|
Huang X, Xi Y, Pan Q, Mao Z, Zhang R, Ma X
and You H: Caffeic acid protects against IL-1β-induced inflammatory
responses and cartilage degradation in articular chondrocytes.
Biomed Pharmacother. 107:433–439. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Meszaros E and Malemud CJ: Prospects for
treating osteoarthritis: Enzyme-protein interactions regulating
matrix metalloproteinase activity. Ther Adv Chronic Dis. 3:219–229.
2012. View Article : Google Scholar
|
|
37
|
Verma P and Dalal K: ADAMTS-4 and
ADAMTS-5: Key enzymes in osteoarthritis. J Cell Biochem.
112:3507–3514. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Deng Z, Lin Z, Zhong Q, Lu M, Fang H, Liu
J, Duan L, Chen L, Wang L, Wang D and Li W: Interleukin 1
beta-induced chloride currents are important in osteoarthritis
onset: An in vitro study. Acta Biochim Biophys Sin (Shanghai).
53:400–409. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
oman-Blas JA and Jimenez SA: NF-kappaB as
a potential therapeutic target in osteoarthritis and rheumatoid
arthritis. Osteoarthritis Cartilage. 14:839–848. 2006. View Article : Google Scholar
|
|
40
|
Lepetsos P, Papavassiliou KA and
Papavassiliou AG: Redox and NF-κB signaling in osteoarthritis. Free
Radic Biol Med. 132:90–100. 2019. View Article : Google Scholar
|
|
41
|
Tang J, Cui W, Song F, Zhai C, Hu H, Zuo Q
and Fan W: Effects of mesenchymal stem cells on
interleukin-1β-treated chondrocytes and cartilage in a rat
osteoarthritic model. Mol Med Rep. 12:1753–1760. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wang T and He C: Pro-inflammatory
cytokines: The link between obesity and osteoarthritis. Cytokine
Growth Factor Rev. 44:38–50. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Song H and Park KH: Regulation and
function of SOX9 during cartilage development and regeneration.
Semin Cancer Biol. 67:12–23. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lefebvre V and Dvir-Ginzberg M: SOX9 and
the many facets of its regulation in the chondrocyte lineage.
Connect Tissue Res. 58:2–14. 2017. View Article : Google Scholar :
|
|
45
|
Kawakami Y, Tsuda M, Takahashi S,
Taniguchi N, Esteban CR, Zemmyo M, Furumatsu T, Lotz M, Izpisúa
Belmonte JC and Asahara H: Transcriptional coactivator PGC-1alpha
regulates chondrogenesis via association with Sox9. Proc Natl Acad
Sci USA. 102:2414–2419. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Akiyama H, Stadler HS, Martin JF, Ishii
TM, Beachy PA, Nakamura T and de Crombrugghe B: Misexpression of
Sox9 in mouse limb bud mesenchyme induces polydactyly and rescues
hypodactyly mice. Matrix Biol. 26:224–233. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Barter MJ, Gomez R, Hyatt S, Cheung K,
Skelton AJ, Xu Y, Clark IM and Young DA: The long non-coding RNA
ROCR contributes to SOX9 expression and chondrogenic
differentiation of human mesenchymal stem cells. Development.
144:4510–4521. 2017.PubMed/NCBI
|
|
48
|
Nakamura Y, He X, Kato H, Wakitani S,
Kobayashi T, Watanabe S, Iida A, Tahara H, Warman ML, Watanapokasin
R and Postlethwait JH: Sox9 is upstream of microRNA-140 in
cartilage. Appl Biochem Biotechnol. 166:64–71. 2012. View Article : Google Scholar
|
|
49
|
Yang J, Qin S, Yi C, Ma G, Zhu H, Zhou W,
Xiong Y, Zhu X, Wang Y, He L and Guo X: MiR-140 is co-expressed
with Wwp2-C transcript and activated by Sox9 to target Sp1 in
maintaining the chondrocyte proliferation. FEBS Lett.
585:2992–2997. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Si HB, Zeng Y, Liu SY, Zhou ZK, Chen YN,
Cheng JQ, Lu YR and Shen B: Intra-articular injection of
microRNA-140 (miRNA-140) alleviates osteoarthritis (OA) progression
by modulating extracellular matrix (ECM) homeostasis in rats.
Osteoarthritis Cartilage. 25:1698–1707. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Jiang Y, Mishima H, Sakai S, Liu YK,
Ohyabu Y and Uemura T: Gene expression analysis of major
lineage-defining factors in human bone marrow cells: Effect of
aging, gender, and age-related disorders. J Orthop Res. 26:910–917.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Kearns AE, Khosla S and Kostenuik PJ:
Receptor activator of nuclear factor kappaB ligand and
osteoprotegerin regulation of bone remodeling in health and
disease. Endocr Rev. 29:155–192. 2008. View Article : Google Scholar
|
|
53
|
Li F and Li X, Liu G, Gao C and Li X: Bone
marrow mesenchymal stem cells decrease the expression of RANKL in
collagen-induced arthritis rats via reducing the levels of IL-22. J
Immunol Res. 2019:84592812019. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Lin TH, Gibon E, Loi F, Pajarinen J,
Córdova LA, Nabeshima A, Lu L, Yao Z and Goodman SB: Decreased
osteogenesis in mesenchymal stem cells derived from the aged mouse
is associated with enhanced NF-κB activity. J Orthop Res.
35:281–288. 217
|
|
55
|
Wang X, Manner PA, Horner A, Shum L, Tuan
RS and Nuckolls GH: Regulation of MMP-13 expression by RUNX2 and
FGF2 in osteoarthritic cartilage. Osteoarthritis Cartilage.
12:963–973. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Chen D, Kim DJ, Shen J, Zou Z and O'Keefe
RJ: Runx2 plays a central role in osteoarthritis development. J
Orthop Translat. 23:132–139. 2019. View Article : Google Scholar
|
|
57
|
Li Z, Liu C, Xie Z, Song P, Zhao RC, Guo
L, Liu Z and Wu Y: Epigenetic dysregulation in mesenchymal stem
cell aging and spontaneous differentiation. PLoS One. 6:e205262011.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Ji Q, Xu X, Xu Y, Fan Z, Kang L, Li L,
Liang Y, Guo J, Hong T, Li Z, et al: miR-105/Runx2 axis mediates
FGF2-induced ADAMTS expression in osteoarthritis cartilage. J Mol
Med (Berl). 94:681–694. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Aubourg G, Rice SJ, Bruce-Wootton P and
Loughlin J: Genetics of osteoarthritis. Osteoarthritis Cartilage.
30:636–649. 2022. View Article : Google Scholar :
|
|
60
|
Tachmazidou I, Hatzikotoulas K, Southam L,
Esparza-Gordillo J, Haberland V, Zheng J, Johnson T, Koprulu M,
Zengini E, Steinberg J, et al: Identification of new therapeutic
targets for osteoarthritis through genome-wide analyses of UK
Biobank data. Nat Genet. 51:230–236. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Cheung KS, Sposito N, Stumpf PS, Wilson
DI, Sanchez-Elsner T and Oreffo RO: MicroRNA-146a regulates human
foetal femur derived skeletal stem cell differentiation by
down-regulating SMAD2 and SMAD3. PLoS One. 9:e980632014. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Tardif G, Pelletier JP, Fahmi H, Hum D,
Zhang Y, Kapoor M and Martel-Pelletier J: NFAT3 and TGF-β/SMAD3
regulate the expression of miR-140 in osteoarthritis. Arthritis Res
Ther. 15:R1972013. View
Article : Google Scholar
|
|
63
|
Nishimura R, Hata K, Nakamura E, Murakami
T and Takahata Y: Transcriptional network systems in cartilage
development and disease. Histochem Cell Biol. 149:353–363. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Kanaan RA and Kanaan LA: Transforming
growth factor beta1, bone connection. Med Sci Monit.
12:RA164–RA169. 2006.PubMed/NCBI
|
|
65
|
Dai J, Yu D, Wang Y, Chen Y, Sun H, Zhang
X, Zhu S, Pan Z, Heng BC, Zhang S and Ouyang H: Kdm6b regulates
cartilage development and homeostasis through anabolic metabolism.
Ann Rheum Dis. 76:1295–1303. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Simon TC and Jeffries MA: The epigenomic
landscape in osteoarthritis. Curr Rheumatol Rep. 19:302017.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Luo C, Hajkova P and Ecker JR: Dynamic DNA
methylation: In the right place at the right time. Science.
361:1336–1340. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Barter MJ, Bui C, Cheung K, Falk J, Gómez
R, Skelton AJ, Elliott HR, Reynard LN and Young DA: DNA
hypomethylation during MSC chondrogenesis occurs predominantly at
enhancer regions. Sci Rep. 10:11692020. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Takahashi A, de Andrés MC, Hashimoto K,
Itoi E and Oreffo RO: Epigenetic regulation of interleukin-8, an
inflammatory chemokine, in osteoarthritis. Osteoarthritis
Cartilage. 23:1946–1954. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Shen J, Wang C, Li D, Xu T, Myers J,
Ashton JM, Wang T, Zuscik MJ, McAlinden A and O'Keefe RJ: DNA
methyltransferase 3b regulates articular cartilage homeostasis by
altering metabolism. JCI Insight. 2:e936122017. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Hashimoto K, Oreffo RO, Gibson MB,
Goldring MB and Roach HI: DNA demethylation at specific CpG sites
in the IL1B promoter in response to inflammatory cytokines in human
articular chondrocytes. Arthritis Rheum. 60:3303–3313. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Roach HI, Yamada N, Cheung KS, Tilley S,
Clarke NM, Oreffo RO, Kokubun S and Bronner F: Association between
the abnormal expression of matrix-degrading enzymes by human
osteoarthritic chondrocytes and demethylation of specific CpG sites
in the promoter regions. Arthritis Rheum. 52:3110–3124. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Goldring SR and Goldring MB: The role of
cytokines in cartilage matrix degeneration in osteoarthritis. Clin
Orthop Relat Res. 427(427 Suppl): S27–S36. 2004. View Article : Google Scholar
|
|
74
|
Aida Y, Maeno M, Suzuki N, Namba A,
Motohashi M, Matsumoto M, Makimura M and Matsumura H: The effect of
IL-1beta on the expression of inflammatory cytokines and their
receptors in human chondrocytes. Life Sci. 79:764–771. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Seto E and Yoshida M: Erasers of histone
acetylation: The histone deacetylase enzymes. Cold Spring Harb
Perspect Biol. 6:a0187132014. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Hong S, Derfoul A, Pereira-Mouries L and
Hall DJ: A novel domain in histone deacetylase 1 and 2 mediates
repression of cartilage-specific genes in human chondrocytes. FASEB
J. 23:3539–3552. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Huber LC, Brock M, Hemmatazad H, Giger OT,
Moritz F, Trenkmann M, Distler JH, Gay RE, Kolling C, Moch H, et
al: Histone deacetylase/acetylase activity in total synovial tissue
derived from rheumatoid arthritis and osteoarthritis patients.
Arthritis Rheum. 56:1087–1093. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Meng F, Li Z, Zhang Z, Yang Z, Kang Y,
Zhao X, Long D, Hu S, Gu M, He S, et al: MicroRNA-193b-3p regulates
chondrogenesis and chondrocyte metabolism by targeting HDAC3.
Theranostics. 8:2862–2883. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Cao K, Wei L, Zhang Z, Guo L, Zhang C, Li
Y, Sun C, Sun X, Wang S, Li P and Wei X: Decreased histone
deacetylase 4 is associated with human osteoarthritis cartilage
degeneration by releasing histone deacetylase 4 inhibition of
runt-related transcription factor-2 and increasing
osteoarthritis-related genes: A novel mechanism of human
osteoarthritis cartilage degeneration. Arthritis Res Ther.
16:4912014. View Article : Google Scholar
|
|
80
|
Higashiyama R, Miyaki S, Yamashita S,
Yoshitaka T, Lindman G, Ito Y, Sasho T, Takahashi K, Lotz M and
Asahara H: Correlation between MMP-13 and HDAC7 expression in human
knee osteoarthritis. Mod Rheumatol. 20:11–17. 2010. View Article : Google Scholar :
|
|
81
|
Liao W, Sun J, Liu W, Li W, Jia J, Ou F,
Su K, Zheng Y, Zhang Z and Sun Y: HDAC10 upregulation contributes
to interleukin 1β-mediated inflammatory activation of
synovium-derived mesenchymal stem cells in temporomandibular joint.
J Cell Physiol. 234:12646–12662. 2019. View Article : Google Scholar
|
|
82
|
Jung JW, Lee S, Seo MS, Park SB, Kurtz A,
Kang SK and Kang KS: Histone deacetylase controls adult stem cell
aging by balancing the expression of polycomb genes and jumonji
domain containing 3. Cell Mol Life Sci. 67:1165–1176. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Dvir-Ginzberg M, Gagarina V, Lee EJ and
Hall DJ: Regulation of cartilage-specific gene expression in human
chondrocytes by SirT1 and nicotinamide phosphoribosyltransferase. J
Biol Chem. 283:36300–36310. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Tsuda M, Takahashi S, Takahashi Y and
Asahara H: Transcriptional co-activators CREB-binding protein and
p300 regulate chondrocyte-specific gene expression via association
with Sox9. J Biol Chem. 278:27224–27229. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Fujita N, Matsushita T, Ishida K, Kubo S,
Matsumoto T, Takayama K, Kurosaka M and Kuroda R: Potential
involvement of SIRT1 in the pathogenesis of osteoarthritis through
the modulation of chondrocyte gene expressions. J Orthop Res.
29:511–515. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Chen H, Liu X, Zhu W, Chen H, Hu X, Jiang
Z, Xu Y, Wang L, Zhou Y, Chen P, et al: SIRT1 ameliorates
age-related senescence of mesenchymal stem cells via modulating
telomere shelterin. Front Aging Neurosci. 6:1032014. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Diao Z, Ji Q, Wu Z, Zhang W, Cai Y, Wang
Z, Hu J, Liu Z, Wang Q, Bi S, et al: SIRT3 consolidates
heterochromatin and counteracts senescence. Nucleic Acids Res.
49:4203–4219. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Fu Y, Kinter M, Hudson J, Humphries KM,
Lane RS, White JR, Hakim M, Pan Y, Verdin E and Griffin TM: Aging
promotes sirtuin 3-dependent cartilage superoxide dismutase 2
acetylation and osteoarthritis. Arthritis Rheumatol. 68:1887–1898.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Wu Y, Chen L, Wang Y, Li W, Lin Y, Yu D,
Zhang L, Li F and Pan Z: Overexpression of Sirtuin 6 suppresses
cellular senescence and NF-κB mediated inflammatory responses in
osteoarthritis development. Sci Rep. 5:176022015. View Article : Google Scholar
|
|
90
|
Collins JA, Kim CJ, Coleman A, Little A,
Perez MM, Clarke EJ, Diekman B, Peffers MJ, Chubinskaya S,
Tomlinson RE, et al: Cartilage-specific Sirt6 deficiency represses
IGF-1 and enhances osteoarthritis severity in mice. Ann Rheum Dis.
ard-2023-2243852023.Epub ahead of print.
|
|
91
|
Zhai XY, Yan P, Zhang J, Song HF, Yin WJ,
Gong H, Li H, Wu J, Xie J and Li RK: Knockdown of SIRT6 enables
human bone marrow mesenchymal stem cell senescence. Rejuvenation
Res. 19:373–384. 2016. View Article : Google Scholar
|
|
92
|
Ji ML, Jiang H, Li Z, Geng R, Hu JZ, Lin
YC and Lu J: Sirt6 attenuates chondrocyte senescence and
osteoarthritis progression. Nat Commun. 13:76582022. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Wu SY, Du YC and Yue CF: Sirt7 protects
chondrocytes degeneration in osteoarthritis via autophagy
activation. Eur Rev Med Pharmacol Sci. 24:9246–9255.
2020.PubMed/NCBI
|
|
94
|
Mohrin M, Shin J, Liu Y, Brown K, Luo H,
Xi Y, Haynes CM and Chen D: Stem cell aging. A mitochondrial
UPR-mediated metabolic checkpoint regulates hematopoietic stem cell
aging. Science. 347:1374–1377. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Hsu YC, Wu YT, Tsai CL and Wei YH: Current
understanding and future perspectives of the roles of sirtuins in
the reprogramming and differentiation of pluripotent stem cells.
Exp Biol Med (Maywood). 243:563–575. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Bi S, Liu Z, Wu Z, Wang Z, Liu X, Wang S,
Ren J, Yao Y, Zhang W, Song M, et al: SIRT7 antagonizes human stem
cell aging as a heterochromatin stabilizer. Protein Cell.
11:483–504. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Hong S, Cho YW, Yu LR, Yu H, Veenstra TD
and Ge K: Identification of JmjC domain-containing UTX and JMJD3 as
histone H3 lysine 27 demethylases. Proc Natl Acad Sci USA.
104:18439–18444. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Ukita M, Matsushita K, Tamura M and
Yamaguchi T: Histone H3K9 methylation is involved in
temporomandibular joint osteoarthritis. Int J Mol Med. 45:607–614.
2020.PubMed/NCBI
|
|
99
|
Zhang F, Xu L, Xu L, Xu Q, Li D, Yang Y,
Karsenty G and Chen CD: JMJD3 promotes chondrocyte proliferation
and hypertrophy during endochondral bone formation in mice. J Mol
Cell Biol. 7:23–34. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Wang P, Li Y, Meng T, Zhang J, Wei Y, Meng
Z, Lin Y, Liu D and Sui L: KDM6A promotes chondrogenic
differentiation of periodontal ligament stem cells by demethylation
of SOX9. Cell Prolif. 51:e124132018. View Article : Google Scholar
|
|
101
|
Lee HL, Yu B, Deng P, Wang CY and Hong C:
Transforming growth factor-β-induced KDM4B promotes chondrogenic
differentiation of human mesenchymal stem cells. Stem Cells.
34:711–719. 2016. View Article : Google Scholar
|
|
102
|
Ye L, Fan Z, Yu B, Chang J, Al Hezaimi K,
Zhou X, Park NH and Wang CY: Histone demethylases KDM4B and KDM6B
promotes osteogenic differentiation of human MSCs. Cell Stem Cell.
11:50–61. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
He Q, Shi J, Liu W, Zhao W, Wang Z, Liu K,
Zhao D, Wang S, Guo Y, Cheng L and Gao Y: TGF-β1-induced bone
marrow mesenchymal stem cells (BMSCs) migration via histone
demethylase KDM6B mediated inhibition of methylation marker
H3K27me3. Cell Death Discov. 8:3392022. View Article : Google Scholar
|
|
104
|
Duan L, Liang Y, Xu X, Wang J, Li X, Sun
D, Deng Z, Li W and Wang D: Noncoding RNAs in subchondral bone
osteoclast function and their therapeutic potential for
osteoarthritis. Arthritis Res Ther. 22:2792020. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
van Meurs JB, Boer CG, Lopez-Delgado L and
Riancho JA: Role of epigenomics in bone and cartilage disease. J
Bone Miner Res. 34:215–230. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Nakamura Y, Inloes JB, Katagiri T and
Kobayashi T: Chondrocyte-specific microRNA-140 regulates
endochondral bone development and targets Dnpep to modulate bone
morphogenetic protein signaling. Mol Cell Biol. 31:3019–3028. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Miyaki S, Sato T, Inoue A, Otsuki S, Ito
Y, Yokoyama S, Kato Y, Takemoto F, Nakasa T, Yamashita S, et al:
MicroRNA-140 plays dual roles in both cartilage development and
homeostasis. Genes Dev. 24:1173–1185. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Iliopoulos D, Malizos KN, Oikonomou P and
Tsezou A: Integrative microRNA and proteomic approaches identify
novel osteoarthritis genes and their collaborative metabolic and
inflammatory networks. PLoS One. 3:e37402008. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Miyaki S, Nakasa T, Otsuki S, Grogan SP,
Higashiyama R, Inoue A, Kato Y, Sato T, Lotz MK and Asahara H:
MicroRNA-140 is expressed in differentiated human articular
chondrocytes and modulates interleukin-1 responses. Arthritis
Rheum. 60:2723–2730. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Ji Q, Xu X, Zhang Q, Kang L, Xu Y, Zhang
K, Li L, Liang Y, Hong T, Ye Q and Wang Y: The
IL-1β/AP-1/miR-30a/ADAMTS-5 axis regulates cartilage matrix
degradation in human osteoarthritis. J Mol Med (Berl). 94:771–785.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Mao G, Zhang Z, Huang Z, Chen W, Huang G,
Meng F, Zhang Z and Kang Y: MicroRNA-92a-3p regulates the
expression of cartilage-specific genes by directly targeting
histone deacetylase 2 in chondrogenesis and degradation.
Osteoarthritis Cartilage. 25:521–532. 2017. View Article : Google Scholar
|
|
112
|
Mao G, Wu P, Zhang Z, Zhang Z, Liao W, Li
Y and Kang Y: MicroRNA-92a-3p regulates aggrecanase-1 and
aggrecanase-2 expression in chondrogenesis and IL-1β-induced
catabolism in human articular chondrocytes. Cell Physiol Biochem.
44:38–52. 2017. View Article : Google Scholar
|
|
113
|
Akhtar N, Rasheed Z, Ramamurthy S,
Anbazhagan AN, Voss FR and Haqqi TM: MicroRNA-27b regulates the
expression of matrix metalloproteinase 13 in human osteoarthritis
chondrocytes. Arthritis Rheum. 62:1361–1371. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Yang B, Guo H, Zhang Y, Chen L, Ying D and
Dong S: MicroRNA-145 regulates chondrogenic differentiation of
mesenchymal stem cells by targeting Sox9. PLoS One. 6:e216792011.
View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Li X, Gibson G, Kim JS, Kroin J, Xu S, van
Wijnen AJ and Im HJ: MicroRNA-146a is linked to pain-related
pathophysiology of osteoarthritis. Gene. 480:34–41. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Vonk LA, Kragten AH, Dhert WJ, Saris DB
and Creemers LB: Overexpression of hsa-miR-148a promotes cartilage
production and inhibits cartilage degradation by osteoarthritic
chondrocytes. Osteoarthritis Cartilage. 22:145–153. 2014.
View Article : Google Scholar
|
|
117
|
Joung S, Yoon DS, Cho S, Ko EA, Lee KM,
Park KH, Lee JW and Kim SH: Downregulation of MicroRNA-495
alleviates IL-1β responses among chondrocytes by preventing SOX9
reduction. Yonsei Med J. 62:650–659. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Qiu M, Liu D and Fu Q: MiR-129-5p shuttled
by human synovial mesenchymal stem cell-derived exosomes relieves
IL-1β induced osteoarthritis via targeting HMGB1. Life Sci.
269:1189872021. View Article : Google Scholar
|
|
119
|
Zhou JX, Tian ZG, Zhu LF, Wu WD, Zhou SL,
Zhao YT and Huang S: MicroRNA-615-3p promotes the osteoarthritis
progression by inhibiting chondrogenic differentiation of bone
marrow mesenchymal stem cells. Eur Rev Med Pharmacol Sci.
22:6212–6220. 2018.PubMed/NCBI
|
|
120
|
Lv S, Xu J, Chen L, Wu H, Feng W, Zheng Y,
Li P, Zhang H, Zhang L, Chi G and Li Y: MicroRNA-27b targets CBFB
to inhibit differentiation of human bone marrow mesenchymal stem
cells into hypertrophic chondrocytes. Stem Cell Res Ther.
11:3922020. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Huang T, Zhou Y, Wang J, Cao Y and Hang
DH: MiR-26b regulates cartilage differentiation of bone marrow
mesenchymal stem cells in rats through the Wnt/β-catenin signaling
pathway. Eur Rev Med Pharmacol Sci. 23:5084–5092. 2019.PubMed/NCBI
|
|
122
|
Chen HO, Zhang L, Tang ZY and Gong ZM:
MiR-485-5p promotes the development of osteoarthritis by inhibiting
cartilage differentiation in BMSCs. Eur Rev Med Pharmacol Sci.
22:3294–3302. 2018.PubMed/NCBI
|
|
123
|
Qin F, Wang F, Wang XP, Chen J, Zeng FH,
Sun CL, Mao JP and Li CL: MiR-539-3p inhibited chondrogenic
differentiation in human adipose stem cells by targeting Sox9. J
Orthop Surg Res. 17:1682022. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Yang M, Yan X, Yuan FZ, Ye J, Du MZ, Mao
ZM, Xu BB, Chen YR, Song YF, Fan BS and Yu JK: MicroRNA-210-3p
promotes chondrogenic differentiation and inhibits adipogenic
differentiation correlated with HIF-3α signalling in bone marrow
mesenchymal stem cells. Biomed Res Int. 2021:66999102021.
|
|
125
|
Zhang P, Gao G, Zhou Z and He X:
microRNA-130b downregulation potentiates chondrogenic
differentiation of bone marrow mesenchymal stem cells by targeting
SOX9. Braz J Med Biol Res. 54:e103452021. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Feng L, Yang ZM, Li YC, Wang HX, Lo JHT,
Zhang XT and Li G: Linc-ROR promotes mesenchymal stem cells
chondrogenesis and cartilage formation via regulating SOX9
expression. Osteoarthritis Cartilage. 29:568–578. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Alahdal M, Huang R, Duan L, Zhiqin D,
Hongwei O, Li W and Wang D: Indoleamine 2, 3 dioxygenase 1 impairs
chondrogenic differentiation of mesenchymal stem cells in the joint
of osteoarthritis mice model. Front Immunol. 12:7811852021.
View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Ma L, Bajic VB and Zhang Z: On the
classification of long non-coding RNAs. RNA Biol. 10:925–933. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Ou F, Su K, Sun J, Liao W, Yao Y, Zheng Y
and Zhang Z: The LncRNA ZBED3-AS1 induces chondrogenesis of human
synovial fluid mesenchymal stem cells. Biochem Biophys Res Commun.
487:457–463. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Huynh NP, Gloss CC, Lorentz J, Tang R,
Brunger JM, McAlinden A, Zhang B and Guilak F: Long non-coding RNA
GRASLND enhances chondrogenesis via suppression of the interferon
type II signaling pathway. Elife. 9:e495582020. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Fang P, Zhang LX, Hu Y, Zhang L and Zhou
LW: Long non-coding RNA DANCR induces chondrogenesis by regulating
the miR-1275/MMP-13 axis in synovial fluid-derived mesenchymal stem
cells. Eur Rev Med Pharmacol Sci. 23:10459–10469. 2019.PubMed/NCBI
|
|
132
|
Ji Y, Fang QY, Wang SN, Zhang ZW, Hou ZJ,
Li JN and Fu SQ: Lnc-RNA BLACAT1 regulates differentiation of bone
marrow stromal stem cells by targeting miR-142-5p in
osteoarthritis. Eur Rev Med Pharmacol Sci. 24:2893–2901.
2020.PubMed/NCBI
|
|
133
|
Zhu Y, Li R and Wen LM: Long non-coding
RNA XIST regulates chondrogenic differentiation of synovium-derived
mesenchymal stem cells from temporomandibular joint via
miR-27b-3p/ADAMTS-5 axis. Cytokine. 137:1553522021. View Article : Google Scholar
|
|
134
|
Chen H, Yang S and Shao R: Long non-coding
XIST raises methylation of TIMP-3 promoter to regulate collagen
degradation in osteoarthritic chondrocytes after tibial plateau
fracture. Arthritis Res Ther. 21:2712019. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Wen C, Lin L, Zou R, Lin F and Liu Y:
Mesenchymal stem cell-derived exosome mediated long non-coding RNA
KLF3-AS1 represses autophagy and apoptosis of chondrocytes in
osteoarthritis. Cell Cycle. 21:289–303. 2022. View Article : Google Scholar :
|
|
136
|
Liu Y, Zou R, Wang Z, Wen C, Zhang F and
Lin F: Exosomal KLF3-AS1 from hMSCs promoted cartilage repair and
chondrocyte proliferation in osteoarthritis. Biochem J.
475:3629–3638. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Zhang H, Li J, Shao W and Shen N: LncRNA
CTBP1-AS2 is upregulated in osteoarthritis and increases the
methylation of miR-130a gene to inhibit chondrocyte proliferation.
Clin Rheumatol. 39:3473–3478. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Zhu J, Fu Q, Shao J, Peng J, Qian Q, Zhou
Y and Chen Y: Regulating effect of Circ_ATRNL1 on the promotion of
SOX9 expression to promote chondrogenic differentiation of hAMSCs
mediated by MiR-145-5p. J Tissue Eng Regen Med. 15:487–502. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Li S, Liu J, Liu S, Jiao W and Wang X:
Mesenchymal stem cell-derived extracellular vesicles prevent the
development of osteoarthritis via the circHIPK3/miR-124-3p/MYH9
axis. J Nanobiotechnology. 19:1942021. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Bao C and He C: The role and therapeutic
potential of MSC-derived exosomes in osteoarthritis. Arch Biochem
Biophys. 710:1090022021. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Liu Y, Lin L, Zou R, Wen C, Wang Z and Lin
F: MSC-derived exosomes promote proliferation and inhibit apoptosis
of chondrocytes via lncRNA-KLF3-AS1/miR-206/GIT1 axis in
osteoarthritis. Cell Cycle. 17:2411–2422. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Mao G, Zhang Z, Hu S, Zhang Z, Chang Z,
Huang Z, Liao W and Kang Y: Exosomes derived from
miR-92a-3p-overexpressing human mesenchymal stem cells enhance
chondrogenesis and suppress cartilage degradation via targeting
WNT5A. Stem Cell Res Ther. 9:2472018. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Tao Y, Zhou J, Wang Z, Tao H, Bai J, Ge G,
Li W, Zhang W, Hao Y, Yang X and Geng D: Human bone mesenchymal
stem cells-derived exosomal miRNA-361-5p alleviates osteoarthritis
by downregulating DDX20 and inactivating the NF-κB signaling
pathway. Bioorg Chem. 113:1049782021. View Article : Google Scholar
|
|
144
|
Cosenza S, Ruiz M, Toupet K, Jorgensen C
and Noël D: Mesenchymal stem cells derived exosomes and
microparticles protect cartilage and bone from degradation in
osteoarthritis. Sci Rep. 7:162142017. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Xia Q, Wang Q, Lin F and Wang J:
miR-125a-5p-abundant exosomes derived from mesenchymal stem cells
suppress chondrocyte degeneration via targeting E2F2 in traumatic
osteoarthritis. Bioengineered. 12:11225–11238. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Chen X, Shi Y, Xue P, Ma X, Li J and Zhang
J: Mesenchymal stem cell-derived exosomal microRNA-136-5p inhibits
chondrocyte degeneration in traumatic osteoarthritis by targeting
ELF3. Arthritis Res Ther. 22:2562020. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Woo CH, Kim HK, Jung GY, Jung YJ, Lee KS,
Yun YE, Han J, Lee J, Kim WS, Choi JS, et al: Small extracellular
vesicles from human adipose-derived stem cells attenuate cartilage
degeneration. J Extracell Vesicles. 9:17352492020. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Wang Z, Yan K, Ge G, Zhang D, Bai J, Guo
X, Zhou J, Xu T, Xu M, Long X, et al: Exosomes derived from
miR-155-5p-overexpressing synovial mesenchymal stem cells prevent
osteoarthritis via enhancing proliferation and migration,
attenuating apoptosis, and modulating extracellular matrix
secretion in chondrocytes. Cell Biol Toxicol. 37:85–96. 2021.
View Article : Google Scholar
|
|
149
|
Lu L, Wang J, Fan A, Wang P, Chen R, Lu L
and Yin F: Synovial mesenchymal stem cell-derived extracellular
vesicles containing microRN555A-26a-5p ameliorate cartilage damage
of osteoarthritis. J Gene Med. 23:e33792021. View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Zeng Z, Dai Y, Deng S, Zou S, Dou T and
Wei F: Synovial mesenchymal stem cell-derived extracellular
vesicles alleviate chondrocyte damage during osteoarthritis through
microRNA-130b-3p-mediated inhibition of the LRP12/AKT/β-catenin
axis. Immunopharmacol Immunotoxicol. 44:247–260. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Kong R, Gao J, Zhang J, Ji L, Yu Y, Zhang
L and Zhao D: Synovial mesenchymal stem cell-derived exosomal
miR-320c enhances chondrogenesis by targeting ADAM19. Future Med
Chem. 14:81–96. 2022. View Article : Google Scholar
|
|
152
|
Liu X, Liu Y, He H, Xiang W and He C:
Human adipose and synovial mesenchymal stem cells improve
osteoarthritis in rats by reducing chondrocyte reactive oxygen
species and inhibiting inflammatory response. J Clin Lab Anal.
36:e243532022. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Wang Y, Yu D, Liu Z, Zhou F, Dai J, Wu B,
Zhou J, Heng BC, Zou XH, Ouyang H and Liu H: Exosomes from
embryonic mesenchymal stem cells alleviate osteoarthritis through
balancing synthesis and degradation of cartilage extracellular
matrix. Stem Cell Res Ther. 8:1892017. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Liu Y, Zeng Y, Si HB, Tang L, Xie HQ and
Shen B: Exosomes derived from human urine-derived stem cells
overexpressing miR-140-5p alleviate knee osteoarthritis through
downregulation of VEGFA in a rat model. Am J Sports Med.
50:1088–1105. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Mao G, Hu S, Zhang Z, Wu P, Zhao X, Lin R,
Liao W and Kang Y: Exosomal miR-95-5p regulates chondrogenesis and
cartilage degradation via histone deacetylase 2/8. J Cell Mol Med.
22:5354–5366. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Shema E, Bernstein BE and Buenrostro JD:
Single-cell and single-molecule epigenomics to uncover genome
regulation at unprecedented resolution. Nat Genet. 51:19–25. 2019.
View Article : Google Scholar
|
|
157
|
Chan CKF, Gulati GS, Sinha R, Tompkins JV,
Lopez M, Carter AC, Ransom RC, Reinisch A, Wearda T, Murphy M, et
al: Identification of the human skeletal stem cell. Cell.
175:43–56.e21. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Vega RB, Matsuda K, Oh J, Barbosa AC, Yang
X, Meadows E, McAnally J, Pomajzl C, Shelton JM, Richardson JA, et
al: Histone deacetylase 4 controls chondrocyte hypertrophy during
skeletogenesis. Cell. 119:555–566. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Ji Q, Zheng Y, Zhang G, Hu Y, Fan X, Hou
Y, Wen L, Li L, Xu Y, Wang Y and Tang F: Single-cell RNA-seq
analysis reveals the progression of human osteoarthritis. Ann Rheum
Dis. 78:100–110. 2019. View Article : Google Scholar
|
|
160
|
Hu X, Li Z, Ji M, Lin Y, Chen Y and Lu J:
Identification of cellular heterogeneity and immunogenicity of
chondrocytes via single-cell RNA sequencing technique in human
osteoarthritis. Front Pharmacol. 13:10047662022. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Chou CH, Jain V, Gibson J, Attarian DE,
Haraden CA, Yohn CB, Laberge RM, Gregory S and Kraus VB: Synovial
cell cross-talk with cartilage plays a major role in the
pathogenesis of osteoarthritis. Sci Rep. 10:108682020. View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Sebastian A, McCool JL, Hum NR, Murugesh
DK, Wilson SP, Christiansen BA and Loots GG: Single-cell RNA-Seq
reveals transcriptomic heterogeneity and post-traumatic
osteoarthritis-associated early molecular changes in mouse
articular chondrocytes. Cells. 10:14622021. View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Pengas I, Eldridge S, Assiotis A,
McNicholas M, Mendes JE and Laver L: MMP-3 in the peripheral serum
as a biomarker of knee osteoarthritis, 40 years after open total
knee meniscectomy. J Exp Ortho. 5:212018. View Article : Google Scholar
|
|
164
|
Chou CH, Lee MT, Song IW, Lu LS, Shen HC,
Lee CH, Wu JY, Chen YT, Kraus VB and Wu CC: Insights into
osteoarthritis progression revealed by analyses of both knee
tibiofemoral compartments. Osteoarthritis Cartilage. 23:571–580.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Qu Y, Wang Y, Wang S, Yu X, He Y, Lu R,
Chen S, Meng C, Xu H, Pei W, et al: A comprehensive analysis of
single-cell RNA transcriptome reveals unique SPP1+ chondrocytes in
human osteoarthritis. Comput Biol Med. 160:1069262023. View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Gao C, Pu H, Zhou Q, Tao T, Liu H, Sun X,
He X and Xiao J: Two reactive behaviors of chondrocytes in an
IL-1β-induced inflammatory environment revealed by the single-cell
RNA sequencing. Aging (Albany NY). 13:11646–11664. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Yoshimoto M, Sadamori K, Tokumura K,
Tanaka Y, Fukasawa K and Hinoi E: Bioinformatic analysis reveals
potential relationship between chondrocyte senescence and protein
glycosylation in osteoarthritis pathogenesis. Front Endocrinol
(Lausanne). 14:11536892023. View Article : Google Scholar : PubMed/NCBI
|