Introduction
Osteoporosis (OP), as a common bone metabolism
disease, is characterized by decreased bone density and destruction
of the bone tissue microstructure. These alterations lead to a
significant increase in bone notable and a substantial rise in the
probability of fractures. As a result, it severely impairs the
patients' quality of life, elevates the mortality rate, and imposes
a heavy economic burden. It has already become a global health
challenge (1). Currently, in
clinical practice, drug-based treatment approaches for OP, such as
bisphosphonates and estrogen replacement therapy, are employed.
However, these methods have certain side effects and limitations,
including issues such as jawbone necrosis, atypical femoral
fractures, and increased risk of cardiovascular disease (2-4).
Therefore, it is particularly important to explore new treatment
methods for OP.
OP is a metabolic disease caused by multiple
factors. Its main causes of onset include endocrine factors,
genetic and immune factors, nutritional factors, sex and age
factors, disease and drug factors, disuse and environmental
factors. Among them, the pathophysiology that has received the
majority of attention mainly emphasizes the endocrine mechanism,
such as estrogen deficiency and aging (5-7).
Changes in these factors lead to the body being in a
microenvironment of chronic inflammation, ultimately resulting in
insufficient osteogenic ability of osteoblasts or enhanced bone
resorption by osteoclasts, thereby disrupting normal bone
metabolism (8,9). According to a previous study, the
levels of interleukin-6 (IL-6), interleukin-1β (IL-1β), and tumor
necrosis factor-α (TNF-α) in the serum of ovariectomized (OVX) rats
are markedly increased compared with the control group (10). As a key molecule that can induce
inflammatory factors IL-6 and TNF-α, nuclear factor-κB (NF-κB) can
markedly inhibit the expression levels of osteocalcin (OCN) and
runt-related transcription factor 2 (RUNX2) in bone marrow
mesenchymal stem cells (BMSCs) when elevated, thereby affecting
their osteogenic differentiation ability (11). In addition, hydrogen peroxide
(H2O2) can activate the NOD-like receptor
pyrin domain-containing 3 (NLRP3) inflammasome mediated by NF-κB,
which in turn leads to inflammatory injury and pyroptosis of
osteoblasts (12). It has been
reported that the expression levels of the NLRP3 inflammasome
increase in the primary BMSCs of OVX mice, and caspase-1 and IL-1β
are activated, which further inhibits osteoblast differentiation
(13). Estrogen deficiency or
oxidative stress may lead to the activation of the NLRP3
inflammasome in osteoblasts, thus triggering the pathological
process of OP.
BA, a natural pentacyclic triterpenoid compound, is
mostly extracted from plants such as birch trees. It has attracted
extensive attention due to its potent pharmacological effects,
including anti-inflammatory, antioxidant and antitumor activities
(14,15). Regarding its anti-inflammatory
properties, BA exerts anti-inflammatory effects by regulating
multiple inflammatory signaling pathways. For example, it can
significantly inhibit the activation of the NF-κB signaling
pathway, the central hub of inflammatory responses, thereby
reducing the transcription and secretion of pro-inflammatory
cytokines such as IL-6, TNF-α and IL-1β in inflammatory cells
(16). Furthermore, studies have
shown that BA can suppress the phosphorylation of p38
mitogen-activated protein kinases (MAPKs), which further blocks
inflammatory reactions in pathological microenvironments (17). In terms of antioxidant activity,
BA can enhance the activity of endogenous antioxidant enzymes in
cells, such as superoxide dismutase and glutathione peroxidase,
while reducing the accumulation of reactive oxygen species (ROS)
(18,19). This regulation helps mitigate
oxidative stress-induced cellular damage and maintain the
structural and functional integrity of cell. Research findings
suggest that part of the mechanisms underlying the actions of BA
can be ascribed to its regulation of autophagy, which verifies that
BA serves as a key regulator in autophagy (20,21). In addition, autophagy plays a
bridging role in inhibiting the activation of the NLRP3
inflammasome (22-24).
Autophagy, an evolutionarily conserved intracellular
degradation process, carries out an indispensable regulatory role
in maintaining bone homeostasis (25). For osteoblasts, moderate
autophagy can protect them against inflammatory and oxidative
stress damage: Under inflammatory conditions, autophagy eliminates
damaged organelles and ROS accumulated in osteoblasts, reducing the
production of pro-inflammatory factors within cells and thereby
preserving the normal proliferation and osteogenic differentiation
capacity of these cells (26).
By contrast, excessive or insufficient autophagy exerts adverse
effects on osteoblasts, persistent inflammation may induce
excessive autophagy in osteoblasts, leading to apoptosis and
further impairing osteogenic function (27); meanwhile, inadequate autophagy
results in the accumulation of cellular waste, which hinders the
synthesis and secretion of osteogenesis-related proteins such as
OCN (28). Additionally,
autophagy activation can promote the differentiation of BMSCs into
mature osteoblasts by upregulating the expression of osteogenic
markers including Runx2 and OCN (29).
Therefore, the present study aimed to explore the
potential of BA in treating bone loss in OVX rats and investigate
the role of osteoblast inflammatory injury in OP and further
confirm whether BA can exert a protective effect on OB by inducing
autophagy and inhibiting NLRP3-induced inflammatory injury.
Materials and methods
Reagents and antibodies
BA, 3-Methyladenin (3-MA), ROS scavenger
N-acetyl-L-cysteine (NAC), NLRP3 inhibitor MCC950 and Dorsomorphin
(synonyms: Compound c, CC) were purchased from MedChemExpress, and
Essential Medium α (α-MEM) and fetal bovine serum (FBS) were
purchased from Gibco, Thermo Fisher Scientific, Inc. The primary
antibodies against NLRP3 (cat. no. BF8029), Asc (cat. no. DF6304),
Caspase-1 (cat. no. AF5418), Cleaved Caspase-1 (cat. no. AF4005),
IL-1β (cat. no. BF8021), AMPK (cat. no. AF6423), phosphorylated
(p-)AMPK (cat. no. AF3423), mTOR and p-mTOR were obtained from
Affinity Biosciences. The primary antibodies for LC3b (cat. no.
ab192890), Beclin-1 (cat. no. ab207612), P62 (cat. no. ab109012),
OPN (cat. no. ab283656) and RUNX2 (cat. no. ab192256) were
purchased from Abcam.
Cell cultures and drug treatments
MC3T3-E1 cells, derived from mouse calvarial
pre-osteoblasts, were purchased from Procell Life Science &
Technology Co., Ltd. (cat. no. CRL-2593). The cells were seeded in
6-well plates containing α-MEM, 10% FBS, 100 IU/ml penicillin and
100 μg/ml streptomycin. After 24 h, the old medium was
discarded and replaced with the osteogenic induction and
differentiation medium (cat. no. CM-0378; Procell Life Science
& Technology Co., Ltd.). The medium was then replaced every 3
days to conduct the induction of osteoblasts. After pretreating
MC3T3-E1 cells with different concentrations of BA, the cells were
treated with 200 μM H2O2 to establish
an inflammatory injury cell model. The cells were cultured in a
humidified incubator with a moist balanced air containing 5% carbon
dioxide at 37°C.
Cell viability assays
MC3T3-E1 cells were seeded into 96-well plates at a
density of 3×103 cells. The cells were then treated with
BA at concentrations of 0, 5, 10, 20, 30 and 40 μM,
respectively, for 24 and 48 h. Subsequently, the cytotoxicity was
evaluated.
The Cell Counting Kit-8 (CCK-8; cat. no. C0037;
Beyotime Institute of Biotechnology) detection kit was used to
determine cell viability. Briefly, 10 μl CCK-8 was added to
each well (100 μl medium), incubated at 37°C for 1 h, and
absorbance was measured. The absorbance value, optical density, was
accurately measured at a wavelength of 450 nm using a microplate
reader (ELx800; Thermo Fisher Scientific, Inc.).
Alkaline phosphatase (ALP) activity and
staining
MC3T3-E1 cells were seeded in 6-well plates. Cells
were pretreated with 0, 10, and 20 μM of BA for 24 h. The
medium was then replaced with osteogenic differentiation medium,
treated with H2O2 for 24 h, and cultured for
7 days, with the medium changed every 3 days. After fixing the cell
samples, they were washed 3-5 times. The BCIP/NBT (cat. no. C3206;
Beyotime Institute of Biotechnology) staining working solution was
then added for 30 min in the dark, the color development reaction
was then terminated. Cell images were obtained using an optical
(bright-field) microscope (Leica Microsystems GmbH).
Alizarin red staining (ARS)
MC3T3-E1 cells were seeded in 6-well plates and
pretreated with 0, 10, and 20 μM of BA for 24 h. The medium
was then replaced with osteogenic differentiation medium containing
H2O2 and cultured for 14 days. The medium was
changed every 3 days. The samples were fixed with 4%
paraformaldehyde (PFA) at room temperature for 20 min. The samples
were washed three times with PBS and the ARS (cat. no. C0148S;
Beyotime Institute of Biotechnology) staining solution was applied
for 30 min at room temperature before terminating the reaction.
Cell images were obtained using an optical microscope (Leica
Microsystems GmbH).
Monodansylcadaverine (MDC) staining
MC3T3-E1 cells were seeded in 96-well plates,
treated per experimental protocols, then culture medium was
aspirated. 100 μl MDC (cat. no. C3018; Beyotime Institute of
Biotechnology) staining solution was added to each well, followed
by 30-min light-protected incubation at 37°C. After discarding the
stain, cells were rinsed thrice with Assay Buffer, and green
fluorescence was visualized via fluorescence microscopy.
Immunofluorescent staining
MC3T3-E1 cells were plated on coverslips and
pretreated with different concentrations of BA. Subsequently, the
culture medium was replaced with fresh α-MEM containing 200
μM H2O2 and incubated under standard
conditions (37°C; 5% CO2) for 24 h. Initial fixation was
performed with 4% PFA for 15 min. Subsequently, the cells were
permeabilized with 0.2% Triton X-100 for 10 min. To minimize
non-specific binding, specimens were incubated with Rapid
Immunofluorescence Blocking Buffer at room temperature for 30 min.
Primary antibody incubation was performed using rabbit anti-LC3b
(1:50) overnight at 4°C, followed by three PBS washes. Secondary
antibody incubation was performed using Cy3-labeled goat
anti-rabbit IgG (1:200; cat. no. A0507; Beyotime Institute of
Biotechnology) at room temperature for 1 h. Nuclear counterstaining
was performed with DAPI (5 μg/ml). Cell images were captured
using an inverted fluorescence microscope (Leica Microsystems
GmbH). Additionally, ImageJ software (v1.53t; National Institutes
of Health) was employed to quantitatively analyze the average
fluorescence intensity.
Intracellular ROS detection
Intracellular ROS generation was assessed using
2',7' dichloro-dihydro-fluorescein diacetate (DCFH-DA; cat. no.
S0034; Beyotime Institute of Biotechnology). MC3T3-E1 cells were
plated in 96-well culture plates at 3×103 cells/well.
Cells were then pretreated with 0, 10, and 20 μM of BA for
24 h. The medium was then replaced with fresh medium containing
H2O2 and culture for 24 h. Prior to analysis,
cells were loaded with 10 μM DCFH-DA in serum-free medium
(37°C; 30 min) and immediately image was captured using an inverted
fluorescence microscope (Leica Microsystems GmbH).
Western blot analysis
After treating the cells under the aforementioned
conditions, the cells were lysed using a buffer containing RIPA
(cat. no. P0013; Beyotime Institute of Biotechnology), PMSF (cat.
no. ST505; Beyotime Institute of Biotechnology) and phosphatase
inhibitors (cat. no. P1081; Beyotime Institute of Biotechnology) to
extract the total cellular proteins. According to the
manufacturer's instructions, a BCA assay kit was used to determine
the protein concentration. Subsequently, the proteins in the
samples were separated using 10% SDS-polyacrylamide gels (loading:
30 μg per lane) and an electrophoresis apparatus. The
proteins then were transferred onto a PVDF membrane. After blocking
the membrane with a rapid protein-free blocking solution at room
temperature for 30 min, the membrane was probed with the primary
antibody overnight at 4°C. After that, the membrane was rinsed
three times with Tris-buffered saline containing 0.1% Tween 20 and
then incubated with the corresponding secondary antibody (1:1,000;
cat. no. A0208 or A0192; Beyotime Institute of Biotechnology) at
room temperature for 2 h. Finally, protein expression was detected
using an enhanced chemiluminescence reagent (cat. no. MI00607B;
Mouse Biotech), and its intensity was analyzed using ImageJ
software.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was isolated from MC3T3-E1 cells using
TRIzol™ Reagent (Thermo Fisher Scientific, Inc.)
following the manufacturer's protocol. For RT, RNA (1 μg)
was combined with oligo (dT) primers (10 μM) and
deoxyribonucleoside triphosphates (dNTPs; 10 mM) in a 20 μl
reaction volume. The reverse transcription reaction was carried out
at 42°C for 60 min, followed by 85°C for 5 min to inactivate the
reverse transcriptase. Primers employed for amplification are shown
in Table I. The obtained
complementary DNA (cDNA) was used as a template to determine the
gene expression levels on the LightCycler® 96 system
using SYBR Green I (Thermo Fisher Scientific, Inc.) as the
fluorophore. The reaction conditions of PCR amplification were as
follows: Denaturation at 95°C for 10 min and followed by 40 cycles
at 95°C for 15 sec, then decreased to 60°C for 15 sec and finally
increased to 72°C for 40 sec. Relative gene expression levels were
calculated using the 2−ΔΔCq method (30).
 | Table IPrimer sequences used for reverse
transcription-quantitative PCR analysis. |
Table I
Primer sequences used for reverse
transcription-quantitative PCR analysis.
| Gene name | Primer sequence
(5'-3') |
|---|
| Runt-related
transcription factor 2 | Forward:
CCGCACGACAACCGCACCAT |
| Reverse:
CGCTCCGGCCCACAAATCTC |
| OPN | Forward:
CGTCCCTACAGTCGATGTCC |
| Reverse:
TGTGGCATCAGGATACTGTTCA |
| β-actin | Forward:
CCTAGGCACCAGGGTGTGAT |
| Reverse:
GGTTGGCCTTAGGGTTCAGG |
Animals and treatment
All animal experimental protocols were examined and
approved by the Laboratory Animal Management Committee of Xi'an
Jiaotong University (approval no. XJTUAE2025-3812). Animals were
housed under controlled conditions (22±2°C, 50±10% humidity,
12/12-h light/dark cycle) with free access to food and water and
were implemented following the Xi'an Jiaotong University Guide for
Animal Experimentation, which conforms to the requirements of the
Guide for the Care and Use of Laboratory Animals issued by the US
National Institutes of Health. A total of 30 Sprague-Dawley female
rats (8-weeks-old; weight, 180±10 g) were acquired from Xi'an
Jiaotong University. Based on the formula provided by
MedChemExpress official guidelines: Animal A (mg/kg)=Animal B
(mg/kg) × (Animal A Km/Animal A Km) and the recommended
intraperitoneal administration volume range for rats, the in
vivo regimen was rationally designed. The rats were divided
into three groups: Sham + Vehicle, OVX + Vehicle, and OVX + BA (15
mg/kg; n=10) randomly. The three groups of rats were acclimated for
1 week. Before all surgical and sampling procedures, the rats were
anesthetized with inhaled isoflurane (4% for induction and 2% for
maintenance) to ensure adequate anesthesia. OVX + BA group rats
were intraperitoneally administered 15 mg/kg/day BA every other day
for 8 weeks. Rats in the sham and OVX + Vehicle groups were
intraperitoneally administered with equal volumes of the excipient.
After 8 weeks of treatment, blood was collected from the heart of
the rats. Then, the rats were euthanized by cervical dislocation.
The femurs were removed and fixed with 4% PFA for subsequent
experiments.
ELISA detection
Cardiac blood samples were obtained via cardiac
puncture and subjected to two-step centrifugation to isolate serum
fractions. Serum concentrations of proinflammatory cytokines were
quantified using ELISA kits: IL-1β (cat. no. GER008), IL-6 (cat.
no. GER011) and TNF-α (cat. no. GER013; all from Elabscience
Biotechnology, Inc.), following the manufacturer's protocols.
Micro-CT analyses
The prepared femur samples were placed in a
high-resolution Micro-CT scanner (Skyscan 1276; Bruker
Corporation). Raw projection data were reconstructed into
three-dimensional models through sequential processing with NRecon
(v1.7.4.2; Bruker-microCT) and Data Viewer (v1.5.6.2,
Bruker-microCT) software packages (Bruker-microCT). Quantitative
morphometric analysis within the metaphyseal region of interest
included bone mineral density (BMD, mg HA/cm3), bone
volume fraction (BV/TV, %), trabecular number (Tb.N,
mm−1) and trabecular separation (Tb.Sp, μm).
H&E, Masson and immunohistochemical
(IHC) staining
Femoral specimens were subjected to decalcification
in 10% EDTA solution, achieving complete decalcification within
8-week duration. Specimens subsequently underwent graded ethanol
dehydration series (70-100%) and paraffin embedding using standard
histology protocols. Subsequently, the specimens were cut into thin
slices with a thickness of 4 μm. Finally, according to the
standard laboratory procedures, the H&E and Masson staining
methods were used respectively to stain the slices.
Paraffin sections underwent xylene dewaxing and
graded ethanol rehydration. After that, they were subjected to
citrate buffer antigen retrieval. Then, 5% bovine serum albumin
(cat. no. HY-D0842; MedChemExpress) was used to block the
non-specific binding at room temperature for 30 min. Incubation
followed successively with the corresponding primary antibody and
secondary antibody. Chromogenic development with
3,3'-diaminobenzidine was monitored microscopically until optimal
signal intensity, followed by hematoxylin counterstaining (30 sec)
and permanent mounting with resinous medium. Brightfield imaging
was performed using a microscope. Quantitative histo-morphometric
analysis of staining intensity was conducted through ImageJ with
IHC Profiler plugin.
mRFP-GFP-LC3 puncta assay
Cells were seeded onto chamber slides and infected
using the 1/2 small-volume method with HBAD-mRFP-GFP-LC3 adenovirus
(cat. no. HB-AP210000; Hanbio Biotechnology Co., Ltd.) at 500
multiplicity of infection at 37°C for 24 h. After transfection, the
cells were subjected to the designated treatments and then examined
under a confocal microscope at 48 h post-infection. Yellow puncta
(RFP+GFP+) were identified as autophagosomes,
while red puncta (RFP+GFP−) represented
autophagolysosomes. Autophagic flux was assessed by manually
counting the number of yellow and red puncta in the merged images.
Successful transfection efficiency was verified, and representative
fluorescence images are shown in Fig. S1.
Statistical analysis
All experimental data were obtained from at least
three independent experiments and were expressed as the mean ± SD.
Statistical analyses were performed using SPSS 24.0 software (IBM
Corp.). Comparisons between two groups were conducted using
unpaired Student's t-test, and Cohen's d was calculated to estimate
effect size. For comparisons among multiple groups, one-way
analysis of variance (ANOVA) followed by Tukey's post hoc test was
applied. When appropriate, 95% confidence intervals (95% CI) were
provided for key outcome measures. P<0.05 was considered to
indicate a statistically significant difference. In addition,
post-hoc power analysis was conducted using G*Power 3.1 based on
key outcome measures (effect size f=0.32, α=0.05, n=10 per group).
P<0.05 was considered to indicate a statistically significant
difference.
Results
Protective effect of BA on OVX-induced
bone loss in rats
To evaluate the in vivo safety of BA, (HC
staining was carried out on major organs (liver, heart, lung and
kidney) of Sham, OVX and BA groups (Fig. S2). Histopathological analysis
revealed intact tissue structures and no obvious abnormalities
(such as inflammatory infiltration, necrosis or structural damage)
in the major organs of the BA groups. No significant histological
differences were observed among the three groups, preliminarily
confirming the favorable in vivo safety of BA at the
experimental dose in OVX rats. To explore the effect of BA on bone
loss in OVX rats, focus was addressed on the rat femurs (Fig. 1A). The micro-CT images and bone
parameters of the rat femurs were analyzed. In addition, the
changes in the microstructure of the rat bone tissue were evaluated
by Masson's trichrome staining and H&E staining. The results of
the CT scan showed that, compared with the OVX group, the BMD,
BV/TV and Tb.N in the BA group were significantly higher, whereas
Tb.Sp was significantly lower. (Fig.
1B-F). The results of Masson's trichrome staining revealed
that, compared with the OVX group, the BA group significantly
increased the generation of bone collagen fibers (Fig. 1G). The results of H&E
staining demonstrated that, compared with the OVX group, the BA
group alleviated the manifestations of trabecular bone reduction
and fracture (Fig. 1G). Post-hoc
power analysis indicated that the current sample size (10 rats per
group) achieved 83.6% power for one-way ANOVA, confirming the
reliability of these primary outcome measures. These findings
indicate that BA can prevent OVX-induced bone loss.
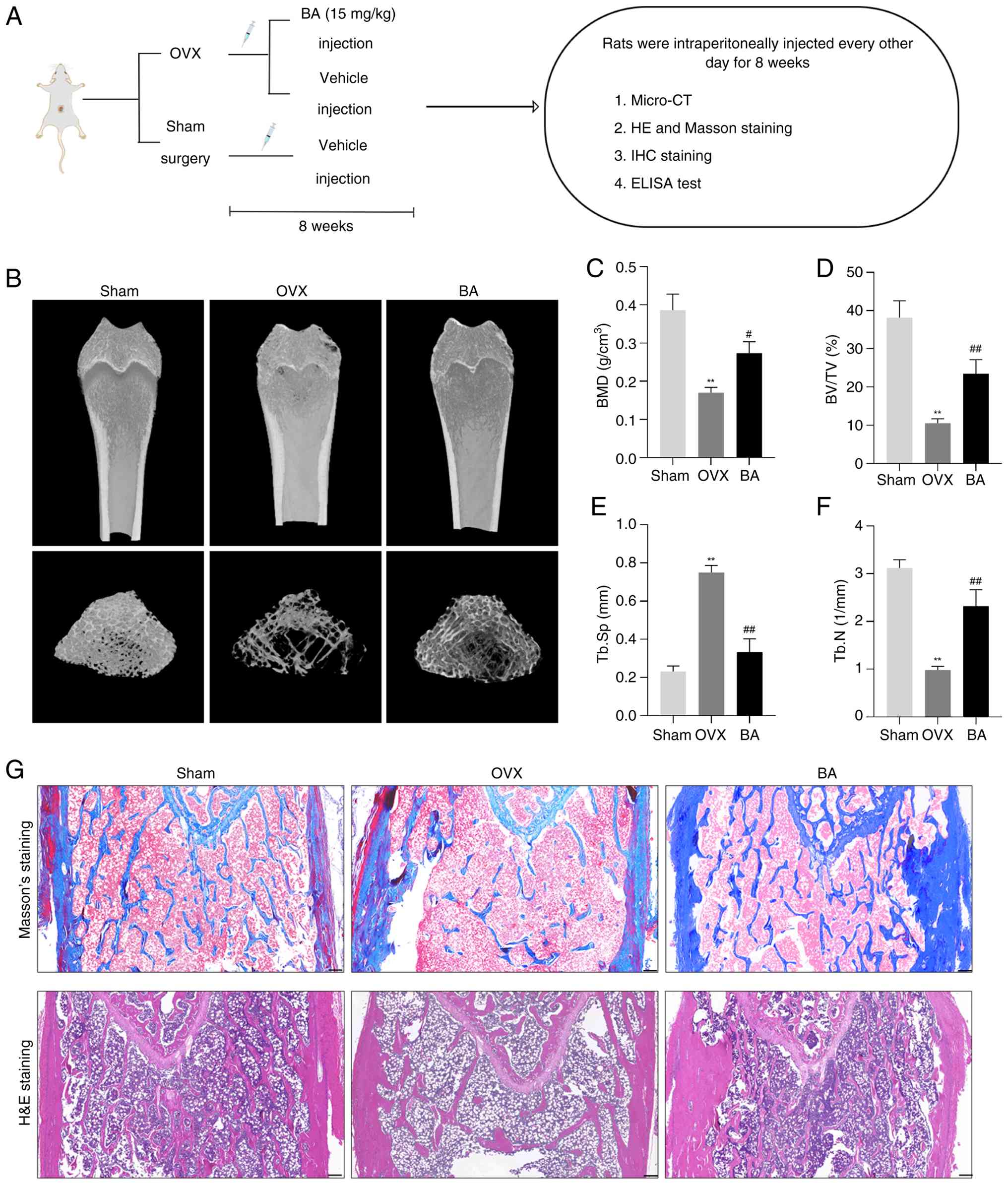 | Figure 1Protective effect of BA on
OVX-induced bone loss in rats. (A) Flow chart representing the
study design protocol to evaluate the therapeutic effect of BA on
OVX rats. (B) Representative images of femurs in each group of rats
detected by micro-CT. (C-F) Quantitative analyses of bone
structural parameters: BMD, BV/TV, Tb.Sp and Tb.N. (G) Masson's
staining and H&E staining of metaphyseal tissue sections of
femurs. Scale bar, 200 μm. Data are expressed as the mean ±
SEM. **P<0.01 vs. sham group; #P<0.05
and ##P<0.01 vs. corresponding OVX group. BA,
betulinic acid; OVX, ovariectomized; BMD, bone mineral density;
BV/TV, bone volume fraction; Tb.N., trabecular number; Tb.Sp,
trabecular separation; IHC, immunohistochemical. |
Protective effect of BA on OVX-induced
inflammation in rats
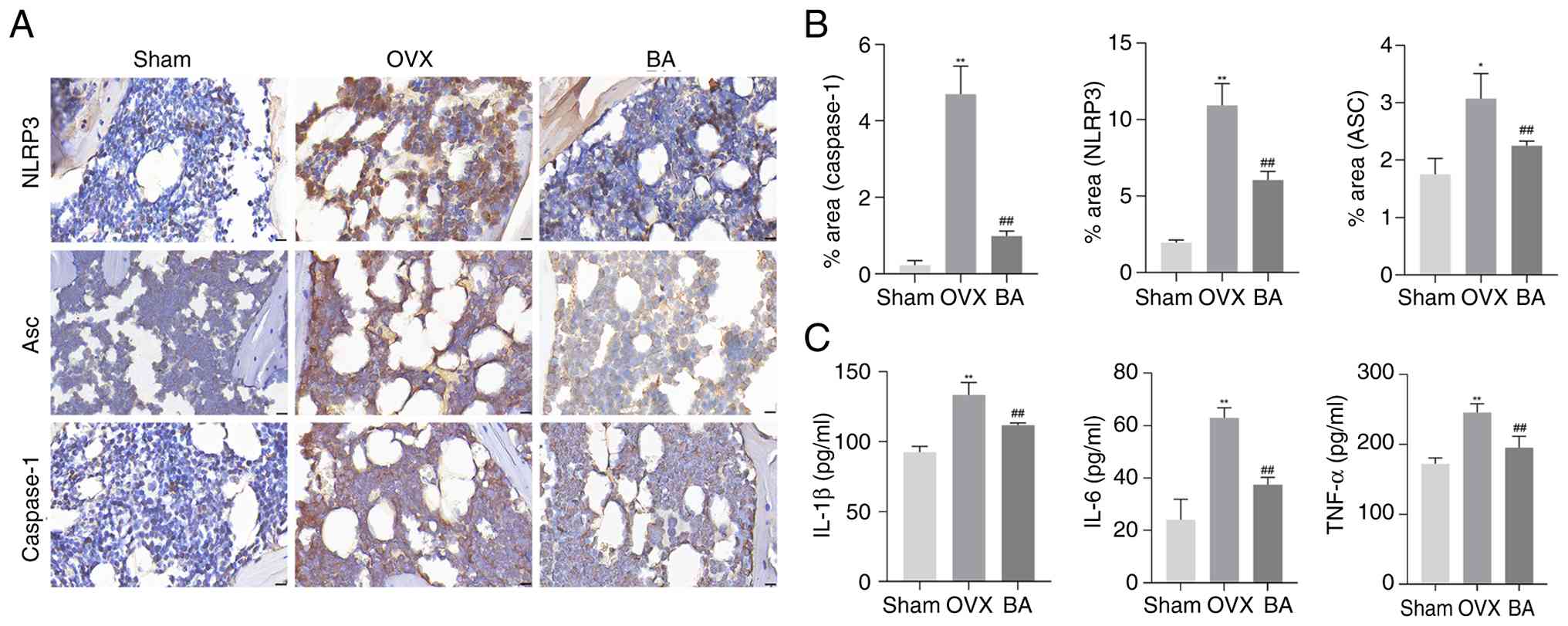
To further investigate the mechanism by which BA
prevents bone loss in OVX rats, the expression levels of NLRP3, Asc
and Caspase-1 protein in the femoral tissues of the rats were
assessed through IHC staining. Additionally, ELISA was used to
detect the inflammatory markers IL-1β, IL-6 and TNF-α in the rat
serum. IHC staining of the femurs of OVX rats identified that,
compared with the sham group, there was a significant increase in
the expression levels of NLRP3, Asc and Caspase-1 protein. However,
administration of BA significantly reduced the expression of these
inflammation-related proteins (Fig.
2A and B). In addition, the results of ELISA showed that BA
could significantly reduce the elevated inflammatory indices of
IL-1β, IL-6 and TNF-α in the serum of OVX rats (Fig. 2C). Therefore, these data suggest
that BA treatment for bone loss in OVX rats may be associated with
its inhibition of the NLRP3 inflammatory pathway in the rat bone
tissue.
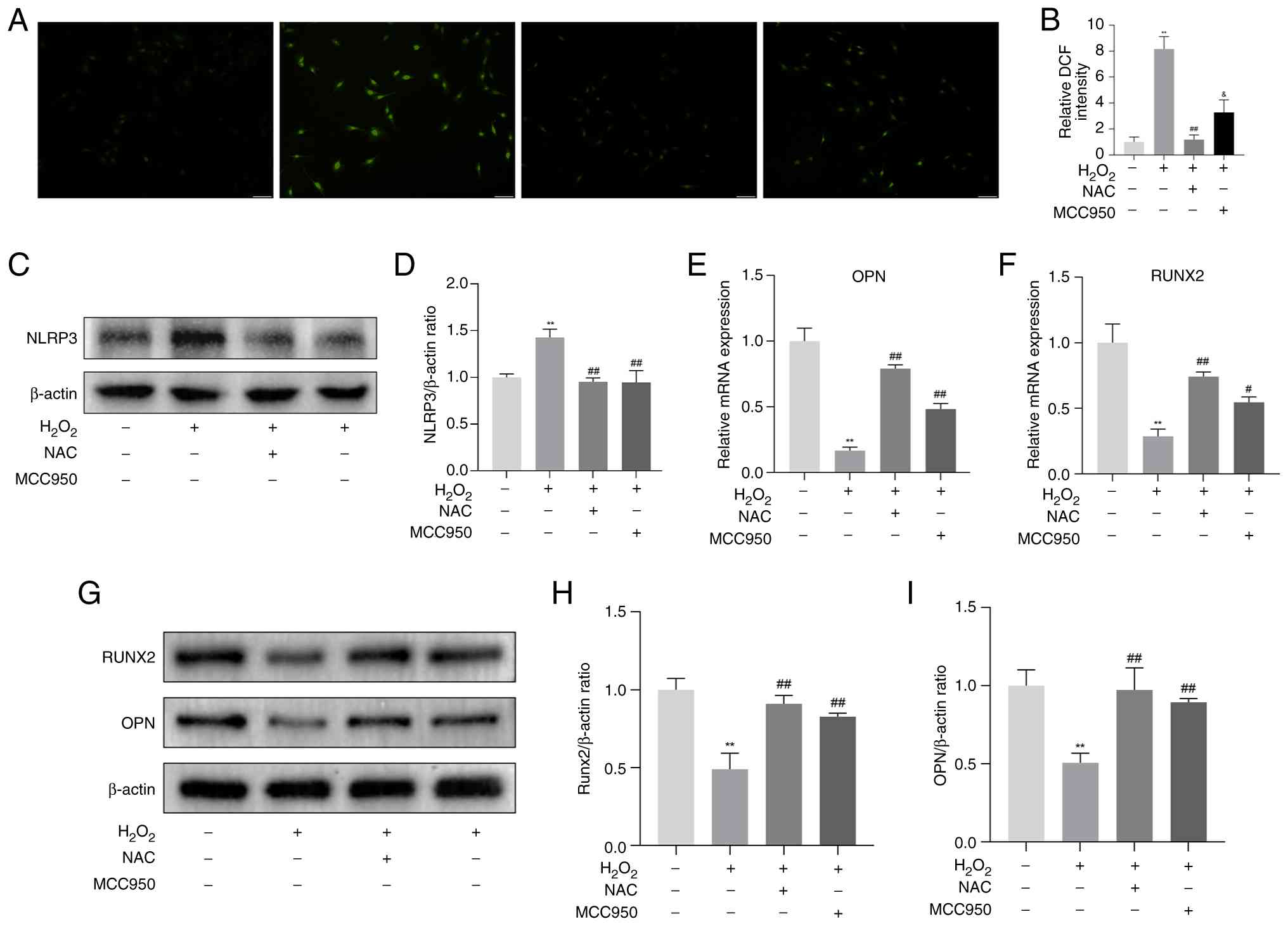
H2O2-induced
osteogenic differentiation decline in MC3T3-E1 cells associated
with ROS/NLRP3
To further explore the potential of BA treatment and
prevention of OP, MC3T3-E1 cells were challenged with
H2O2 (200 μM) and an inflammatory
injury cell model was established in vitro. The experimental
results showed that, compared with the con group,
H2O2 intervention significantly increased ROS
production in MC3T3-E1 cells (Fig.
3A and B), and the protein expression level of the NLRP3
inflammasome increased (Fig. 3C and
D). In addition, as osteogenic differentiation marker factors,
both the mRNA and protein expression levels of RUNX2 and OPN were
inhibited (Fig. 3E-I). When
NLRP3 inhibition experiments were conducted using MCC950
(NLRP3-specific inhibitor) in MC3T3-E1 cells with inflammatory
injury, relative to the H2O2 group, the
expression level of NLRP3 protein decreased significantly (Fig. 3C and D). Moreover, relative to
the H2O2 group, it significantly improved the
expression of RUNX2 and OPN genes and proteins (Fig. 3E-I). Additionally, when NAC
(specific ROS inhibitor) was added, in comparison with the
H2O2 group, the accumulation of ROS was
reduced, the expression of NLRP3 protein in the inflammasome was
inhibited (Fig. 3A-D), and the
inhibition of RUNX2 and OPN gene and protein expression was
alleviated (Fig. 3E-I). The
aforementioned findings indicate that the effect of
H2O2 on the osteogenic differentiation of
MC3T3-E1 is associated with ROS/NLRP3 inflammasome.
BA attenuates NLRP3-induced inflammatory
injury and osteogenic differentiation decline in
H2O2-exposed MC3T3-E1 cells
Before evaluating the effect of BA (Fig. 4A) on the differentiation of
MC3T3-E1 cells treated with H2O2, the CCK-8
activity assay was used to detect whether there were potential
cytotoxic effects on MC3T3-E1 cells within 24 and 48 h. When the
concentration of BA ≤20 μM, there was no significant change
in cell viability (Fig. 4B).
Consistent with previous studies reporting that BA shows no
cytotoxicity to osteoblast-lineage cells within the range of 0-30
μM (31), 10 and 20
μM were therefore selected for subsequent experiments.
MC3T3-E1 cells were treated with BA at different concentrations.
The results showed that BA (20 μM) significantly inhibited
the activation of the NLRP3 inflammasome pathway induced by
H2O2 (Fig.
4C-H). In addition, the results of ARS and ALP staining
revealed that, BA (20 μM) significantly restored the
activity of ALP and the number of mineralized nodules (Fig. 4I). Similarly, by detecting the
mRNA expression of RUNX2 and OPN, it was found that when treated
with BA (20 μM), the mRNA expression of the two was
significantly restored (Fig.
4J-K). These results demonstrate that BA can effectively treat
H2O2-induced inflammatory damage and
significantly alleviate the inhibitory effect of
H2O2 on osteoblast differentiation.
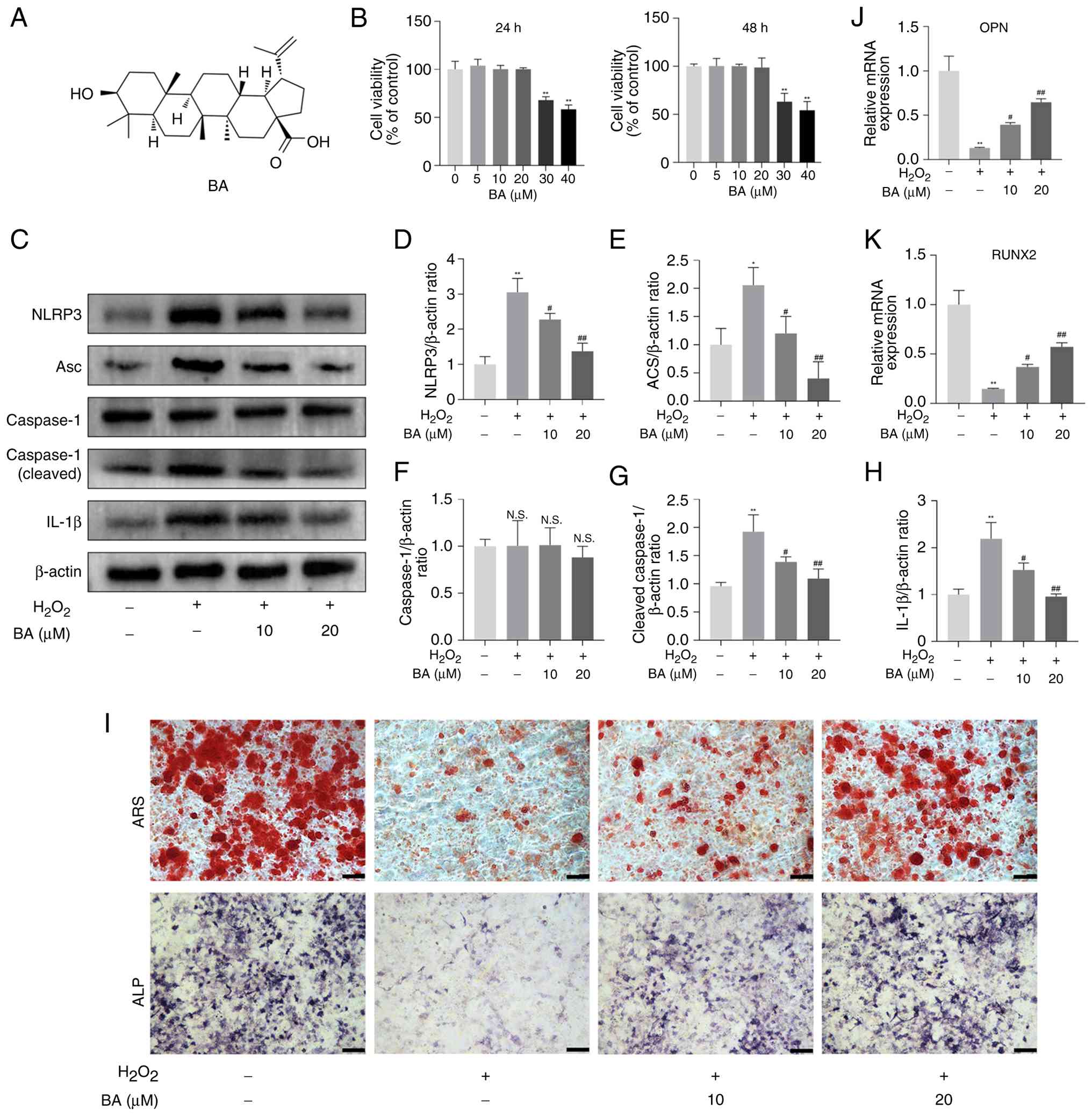 | Figure 4BA attenuates NLRP3-induced
inflammatory injury and osteogenic differentiation decline in
H2O2-exposed MC3T3-E1 cells. (A) The chemical
structure of BA. (B) MC3T3-E1 cells were treated with different
concentrations of BA (0, 5, 10, 20, 30 and 40 μM) for 24 and
48 h. Cell viability was measured by Cell Counting Kit-8 assay. (C)
MC3T3-E1 cells were incubated in the presence of
H2O2 (200 μM) medium with the BA (10
μM) or BA (20 μM), western blot analysis was
performed against NLRP3, Asc, caspase-1, cleaved caspase-1 and
IL-1β. (D-H) Quantification of the results shown in C. (I) ALP and
ARS staining were performed in MC3T3-E1 cells. (J and K)
Quantitative analysis of osteoblast marker gene expression after
treatment with different concentrations of BA using reverse
transcription-quantitative PCR. Data are expressed as the mean ±
SEM. **P<0.01 vs. control group;
#P<0.05 and ##P<0.01 vs. corresponding
H2O2 group. BA, betulinic acid; NLRP3,
NOD-like receptor pyrin domain-containing 3; ALP, alkaline
phosphatase; ARS, alizarin red staining; RUNX2, runt-related
transcription factor 2; N.S., not significant. |
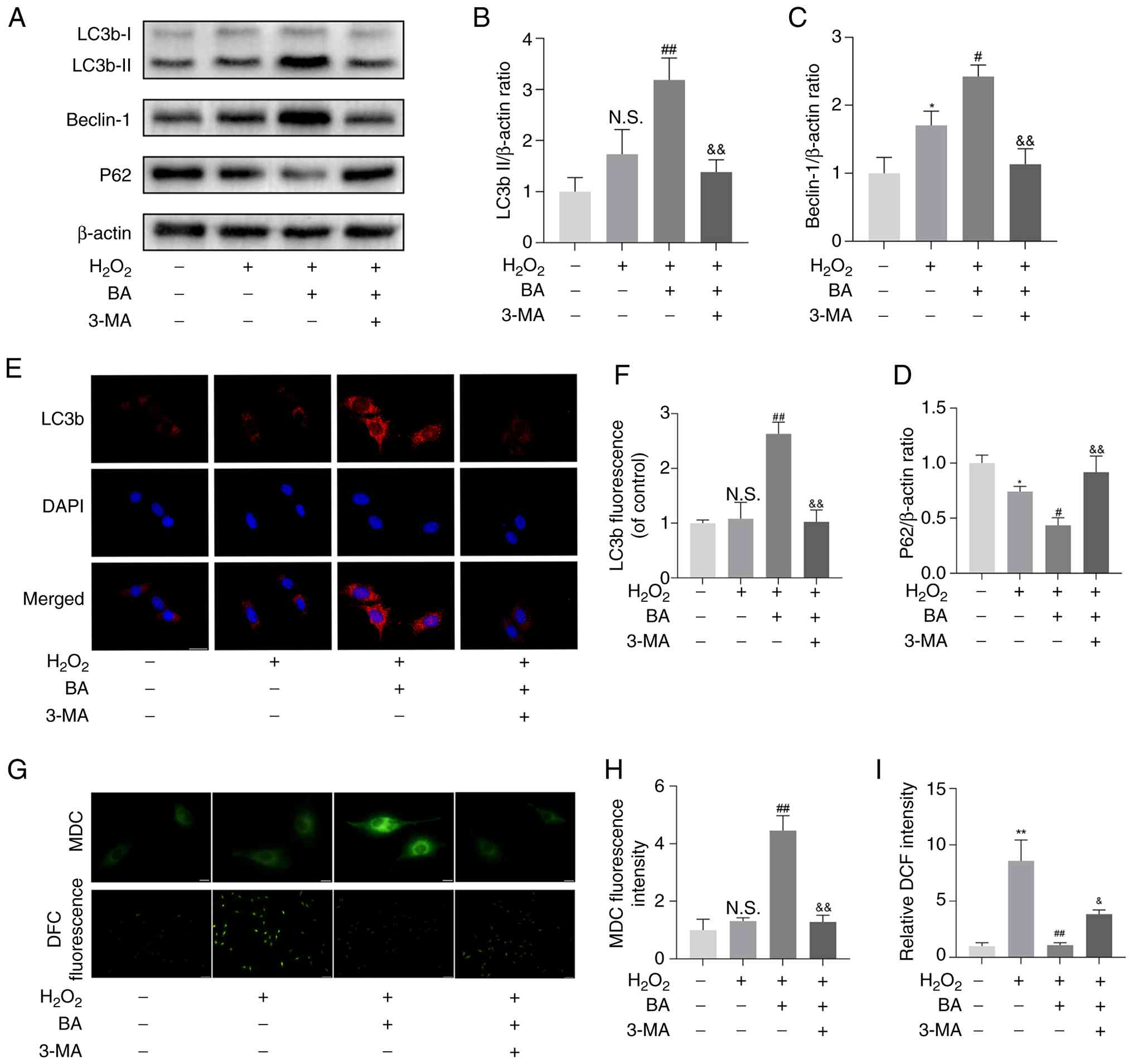
BA enhances autophagy in
H2O2-exposed MC3T3-E1 cells
Based on the previous experimental data, 20
μM was determined as the optimal concentration for BA. To
explore the effect of BA on autophagy in MC3T3-E1 cells treated
with H2O2, the expression of autophagy marker
proteins was detected. The results showed that BA significantly
increased the expression of LC3b II and Beclin-1 in the cells and
inhibited the expression of P62. However, 3-MA could reverse the
effects of BA on the expression of LC3b-II, Beclin-1 and P62
(Fig. 5A-D). Immunofluorescence
results showed that, compared with the control group, the
expression of LC3b in cells increased significantly, and the 3-MA
group reversed this result (Fig. 5E
and F). MDC can specifically label autophagosomes. MDC staining
showed that BA can promote the formation of autophagosomes
(Fig. 5G and H). The fluorescent
probe DCFH-DA was used to detect intracellular ROS. The BA group
significantly inhibited the production of ROS, but the 3-MA group
reversed this result (Fig. 5I).
Additionally, based on the mRFP-GFP-LC3 puncta assay, the
autophagic flux was dynamically evaluated under different
conditions. In the BA treatment group, there was a marked increase
in both yellow puncta (autophagosomes) and red puncta
(autolysosomes), indicating that BA significantly promoted
autophagosome formation and enhanced autophagic flux. By contrast,
when bafilomycin A1 was added together with BA, yellow puncta
remained abundant, but red puncta were markedly reduced. This
suggests that although autophagosome formation was still induced,
the fusion and degradation steps were blocked, leading to
autophagosome accumulation. These findings demonstrate that BA
enhances autophagy by promoting autophagic flux (Fig. S3). This indicates that BA can
enhance autophagy and inhibit ROS production in
H2O2-exposed MC3T3-E1 cells.
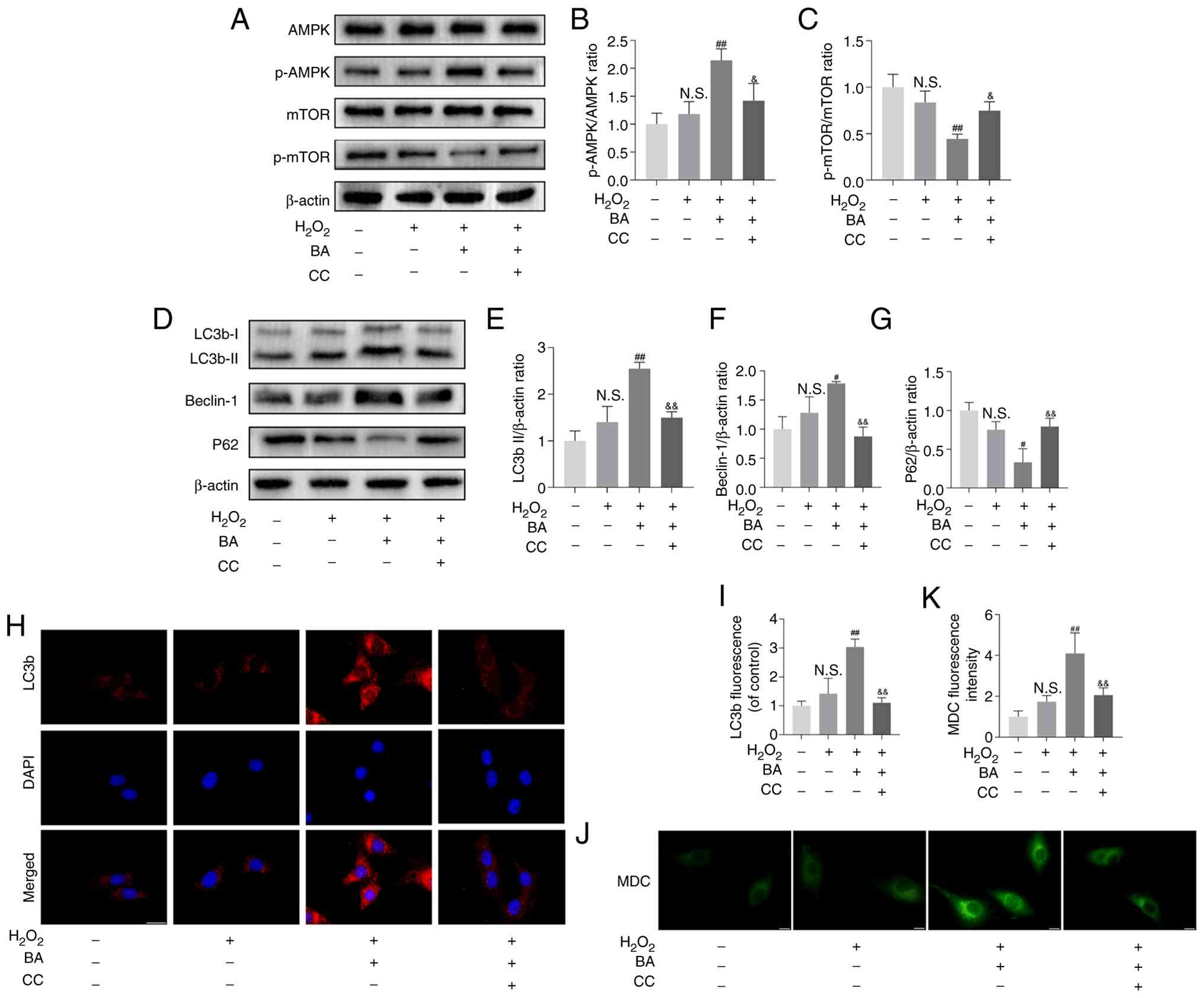
BA activates the AMPK/mTOR signaling
pathway in H2O2-exposed MC3T3-E1 cells
The regulation of autophagy by BA was investigated.
It has been reported that BA can regulate AMPK phosphorylation
(32). Western blotting showed
that BA effectively promoted the phosphorylation of AMPK and
inhibited the phosphorylation of mTOR, while compound C (an AMPK
blocker) significantly reversed this effect and inhibited the
expression of LC3b-II and Beclin-1, promoting the expression of P62
(Fig. 6A-G). The group treated
with compound C also inhibited the expression of LC3b as detected
by immunofluorescence and the formation of autophagosomes as shown
by MDC staining (Fig. 6H-K).
This indicates that BA regulates autophagy in
H2O2-exposed MC3T3-E1 cells through the
AMPK/mTOR pathway.
Discussion
In humans, bones are constantly in a continuous
remodeling cycle. Under physiological conditions,
mesenchymal-derived osteoblasts and hematopoietic-derived
osteoclasts maintain the structural stability of bone tissue
through synergistic actions (33). The former constructs the bone
matrix by synthesizing collagen fibers and inorganic salts, while
the latter decomposes the aged bone tissue by acidifying the
microenvironment. These two populations of cells with antagonistic
functions form a sophisticated functional coupling system under the
regulation of cytokines, jointly determining the quality of bone
tissue and metabolic homeostasis. When there is a dysfunction in
this regulatory network, metabolic imbalance will cause the rate of
bone resorption to exceed the bone formation capacity, thus leading
to a progressive decrease in bone mineral density and
manifestations of OP (34,35).
Studies on the molecular mechanisms of bone
metabolism regulation have shown that fluctuations in endogenous
hormone levels play an important regulatory role in bone
homeostasis (36). Previous
studies have confirmed that a decrease in estrogen levels can
induce a chronic inflammatory response through the amplifying
effect of the NF-κB signaling cascade (37,38). This molecular cascade can
severely disrupt the dynamic balance between bone formation and
bone resorption, and it is a key pathogenic factor for
postmenopausal OP. The regulation of osteogenic differentiation
relies on the synergistic action of core signaling pathways, such
as the Wnt/β-catenin and BMP/Smad pathways. These pathways drive
the differentiation of MSCs into mature osteoblasts and facilitate
bone matrix mineralization by activating key transcription factors,
including RUNX2 and osterix (39-41). Notably, the inflammatory
microenvironment can inhibit osteogenic differentiation by
interfering with the aforementioned pathways, such as suppressing
Wnt signaling and abnormally activating Notch signaling (42,43). The positive feedback loop between
the systemic low-grade inflammatory state and the ROS-NLRP3
inflammasome axis serves as the core pathological mechanism through
which inflammatory damage compromises osteogenic function in
postmenopausal OP (44,45). At the molecular level,
pathological processes such as mitochondrial dysfunction can
activate the NLRP3 inflammasome via ROS-dependent pathways.
Mediated by ASC, this activation triggers Caspase-1 cleavage, which
in turn promotes the maturation and release of IL-1β, ultimately
impairing osteogenic differentiation capacity (46-48). The present study found that
MC3T3-E1 cells intervened with H2O2 showed
increased production of ROS, excessive activation of the NLRP3
pathway, accompanied by a downregulation of the expression levels
of osteogenic markers. This suggests that inflammatory injury is a
key regulatory step in the impairment of osteogenic
differentiation. It is worth noting that both NAC and MCC950 can
inhibit the activation of NLRP3 and effectively restore the
osteogenic differentiation ability. Moreover, compared with MCC950,
NAC can also inhibit the production of ROS. These findings suggest
that targeted regulation of the ROS-NLRP3 signaling axis may
provide a new molecular intervention strategy for the treatment of
OP.
BA, a natural product of pentacyclic triterpenoids,
has become a research hotspot in the field of drug development in
recent years due to its multiple biological activity
characteristics in metabolic diseases, especially its significant
anti-inflammatory and antioxidant abilities (49). However, the protective effect of
this compound on osteoblast function and its potential value in the
treatment of OP have not been elucidated. According to a previous
study, the concentration of pro-inflammatory mediators such as
IL-1β and TNF-α in the peripheral blood of rats with OVX model
markedly increased (50). The
present study revealed that the BA intervention group of OVX rats
not only significantly reduced the levels of serum inflammatory
factors, but also significantly downregulated the protein
expression of NLRP3 inflammasome components (NLRP3, Asc and
Caspase-1) in their femoral tissue. At the same time, bone
microstructural parameters were significantly improved. In
vitro experiments have confirmed that in MC3T3-E1 cells
intervened by H2O2, after pretreatment with
BA, the NLRP3 inflammasome pathway is inhibited. Meanwhile, the ALP
activity of cells is restored, the number of mineralized nodules
significantly rebounds, and the inhibited expression of
osteoblast-specific genes RUNX2 and OPN is effectively improved.
Collectively, these results demonstrate that BA exerts effects in
protecting osteoblasts from inflammatory damage, suppressing
inflammation in rats, and treating OP in OVX rats. These effects
are achieved by inhibition of the activation of the NLRP3
inflammasome by BA.
Autophagy is a key biological activity that responds
to various types of stresses and maintains intracellular
homeostasis. It can assist cells in processes such as clearing
damaged organelles and misfolded proteins, ensuring the stability
of the intracellular environment, thereby enabling cells to
maintain normal physiological functions in various complex internal
and external environmental changes (51). An increasing number of studies
have shown that autophagy plays an important role in scavenging ROS
and inhibiting the activation of the NLRP3 inflammasome (52,53). Moreover, BA can regulate the
autophagy levels of some tissues and cells (18,23,32). Autophagy can be detected by
examining the expression levels of common markers such as
LC3b-II/I, Beclin-1, and P62. Additionally, the formation of
autophagosomes can also be detected by MDC. The present study is
the first to demonstrate that, in MC3T3-E1 cells treated with
H2O2, BA can promote the expression of
LC3b-II and Beclin-1, inhibit the expression of P62, and
significantly increase the formation of autophagosomes. In
addition, BA can inhibit ROS production through autophagy, and the
autophagy inhibitor 3-MA can reverse this effect. These findings
indicate that the protective effect of BA against inflammatory
injury in MC3T3-E1 cells is partly attributed to the enhancement of
autophagy, which reduces ROS production and inhibits the activation
of NLRP3. Among a variety of pathways for regulating autophagy, the
AMPK/mTOR axis is classical and widely known (54). However, it remains unclear
whether BA activates the AMPK/mTOR signaling pathway to regulate
autophagy in MC3T3-E1 cells treated with
H2O2. Findings of the present study revealed
that BA could promote the phosphorylation of AMPK and weaken the
phosphorylation of mTOR, which subsequently led to the activation
of autophagy. Compound C, an AMPK inhibitor, could significantly
reverse the effect of BA in promoting the phosphorylation of AMPK.
These results indicated that the autophagy induced by BA is
mediated through the AMPK/mTOR signaling pathway.
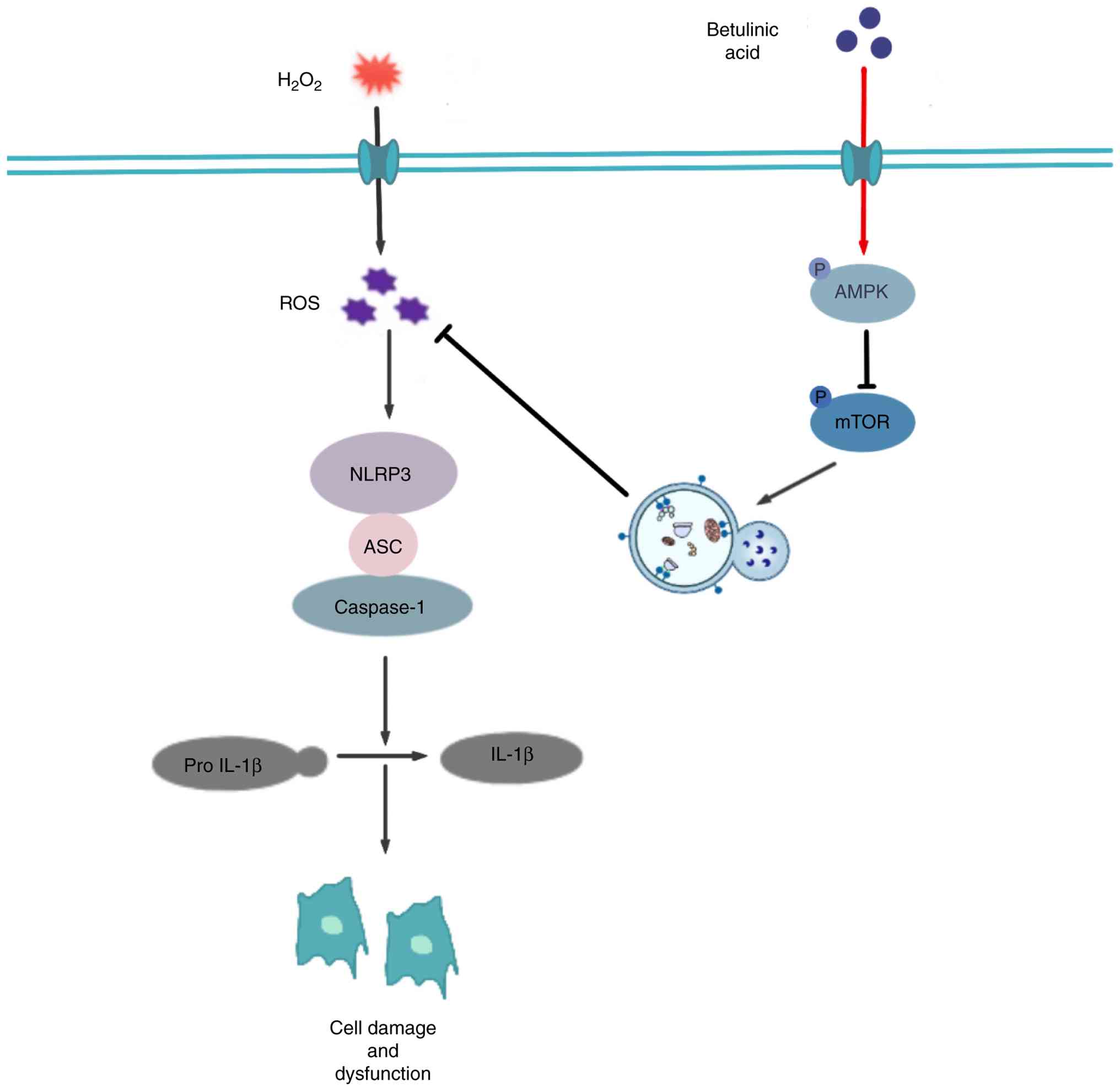
In conclusion, the present study demonstrated for
the first time that BA may alleviate the chronic low-grade
inflammation and OP caused by estrogen deficiency in OVX rats.
In vitro, the potential mechanism by which BA exerts its
effects may be through activating the AMPK/mTOR pathway, promoting
cellular autophagy, reducing the excessive production of ROS,
thereby decreasing the generation and activation of the NLRP3
inflammasome, maintaining the stability of the intracellular
environment, and ensuring the normal differentiation and functions
of osteoblasts (Fig. 7).
However, the present study also has some limitations. Firstly,
pharmacokinetic properties, bioavailability and tissue distribution
of BA were not addressed, and the 15 mg/kg dosing regimen in rats
was based on recommended guidelines rather than detailed
pharmacokinetic modeling. Secondly, previous studies have shown
that BA inhibits the generation and differentiation of osteoclasts
caused by ROS and inflammation (55,56). However, further investigation is
needed to determine whether these regulate autophagy through BA.
Thirdly, further research is needed on the safety of BA in
vivo. Finally, future studies will incorporate positive control
groups, such as rapamycin as an autophagy inducer or MCC950 as an
NLRP3 inhibitor, to directly compare the efficacy of BA with
established therapeutic agents and further clarify its mechanism of
action. In summary, the results of the present study indicate that
BA is a potential drug for treating diseases associated with
osteoblast inflammatory injury and provide relevant evidence for
this.
Supplementary Data
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author.
Authors' contributions
YiZ and ZQ conceptualized and designed the study,
analyzed and interpreted the data. YiZ and LY designed the study,
drafted the manuscript and carried out the experiments. ZQ, YG and
LY revised the manuscript critically for important intellectual
content and provided final approval of the version to be published.
XW, YoZ, LL and BZ analyzed and interpretated the data. YG, LL, YiZ
and BZ carried out the experiments and acquired data. LY acquired
funding. LY and ZQ confirm the authenticity of all the raw data.
All authors read and approved the final version of the
manuscript.
Ethics approval and consent to
participate
All animal care and experimental procedures were
approved (approval no. XJTUAE2025-3812) by Animal Care Committee of
Hong-Hui Hospital, Xi'an Jiaotong University College of Medicine
(Xi'an, China) and conducted strictly following the institutional
guidelines for the care and use of laboratory animals at the
Jiaotong University College of Medicine.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgements
The authors sincerely appreciate the technical
support provided by the Translational Center Laboratory Platform
and Public Technical Service Platform of the Red Cross Hospital
Affiliated to Xi'an Jiaotong University.
Funding
The present study was supported by the National Natural Science
Foundation of China (grant no. 82501067), the Plan for Health and
Medical Research and Innovation Platforms in Shaanxi (grant no.
2025PT-01), the Medical Research Project of Xi'an Science and
Technology Bureau (grant no. 24YXYJ0006) and the Key Project of
Natural Science Basic Research Plan of Shaanxi (grant no.
2025JC-YBQN-1164).
References
|
1
|
Fischer V and Haffner-Luntzer M:
Interaction between bone and immune cells: Implications for
postmenopausal osteoporosis. Semin Cell Dev Biol. 123:14–21. 2022.
View Article : Google Scholar
|
|
2
|
Ott SM: Osteoporosis treatment: Not easy.
Ann Intern Med. 176:278–279. 2023. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Adler RA: Osteoporosis treatment:
Decreased mortality too? J Clin Endocrinol Metab. 108:e48–e49.
2023. View Article : Google Scholar
|
|
4
|
Song S, Guo Y, Yang Y and Fu D: Advances
in pathogenesis and therapeutic strategies for osteoporosis.
Pharmacol Ther. 237:1081682022. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Adami G, Fassio A, Rossini M, Caimmi C,
Giollo A, Orsolini G, Viapiana O and Gatti D: Osteoporosis in
rheumatic diseases. Int J Mol Sci. 20:58672019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Valero C and González Macías J:
Atherosclerosis, vascular calcification and osteoporosis. Med Clin
(Barc). 164:e13–e20. 2025.In English, Spanish. View Article : Google Scholar
|
|
7
|
Weaver CM: Nutrition and bone health. Oral
Dis. 23:412–415. 2017. View Article : Google Scholar
|
|
8
|
Kimball JS, Johnson JP and Carlson DA:
Oxidative stress and osteoporosis. J Bone Joint Surg Am.
103:1451–1461. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yao Y, Cai X, Chen Y, Zhang M and Zheng C:
Estrogen deficiency-mediated osteoimmunity in postmenopausal
osteoporosis. Med Res Rev. 45:561–575. 2025. View Article : Google Scholar
|
|
10
|
Zhang YW, Cao MM, Li YJ, Lu PP, Dai GC,
Zhang M, Wang H and Rui YF: Fecal microbiota transplantation
ameliorates bone loss in mice with ovariectomy-induced osteoporosis
via modulating gut microbiota and metabolic function. J Orthop
Translat. 37:46–60. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang G, Tan J, Huang C, Xu Y, Yang Z and
Huo L: Based on NF-κB and Notch1/Hes1 signaling pathways, the
mechanism of artesunate on inflammation in osteoporosis in
ovariectomized rats was investigated. Front Biosci (Landmark Ed).
29:2662024. View Article : Google Scholar
|
|
12
|
Sun D, Peng Y, Ge S and Fu Q: USP1
inhibits NF-κB/NLRP3 induced pyroptosis through TRAF6 in
osteoblastic MC3T3-E1 cells. J Musculoskelet Neuronal Interact.
22:536–545. 2022.PubMed/NCBI
|
|
13
|
Xu L, Zhang L, Wang Z, Li C, Li S, Li L,
Fan Q and Zheng L: Melatonin suppresses estrogen deficiency-induced
osteoporosis and promotes osteoblastogenesis by inactivating the
NLRP3 inflammasome. Calcif Tissue Int. 103:400–410. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
An T, Zha W and Zi J: Biotechnological
production of betulinic acid and derivatives and their
applications. Appl Microbiol Biotechnol. 104:3339–3348. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lou H, Li H, Zhang S, Lu H and Chen Q: A
review on preparation of betulinic acid and its biological
activities. Molecules. 26:55832021. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zheng X, Cao Z, Wang M, Yuan R, Han Y, Li
A and Wang X: Betulinic acid reduces intestinal inflammation and
enhances intestinal tight junctions by modulating the PPAR-γ/NF-κB
signaling pathway in intestinal cells and organoids. Nutrients.
17:20522025. View Article : Google Scholar
|
|
17
|
Xia G, Shen C, Xiao Y, Wang X, Qiu L, Lei
S and Jiang R: Shenling Baizhu Powder and Betulin attenuate
sepsis-induced intestinal injury by targeting GADD45B/TAOK1/p38
MAPK pathway. J Ethnopharmacol. 353:1202822025. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zheng LY, Zou X, Wang YL, Zou M, Ma F,
Wang N, Li JW, Wang MS, Hung HY and Wang Q: Betulinic
acid-nucleoside hybrid prevents acute alcohol-induced liver damage
by promoting anti-oxidative stress and autophagy. Eur J Pharmacol.
914:1746862022. View Article : Google Scholar
|
|
19
|
Ou Z, Zhu L, Huang C, Ma C, Kong L, Lin X,
Gao X, Huang L, Wen L, Liang Z, et al: Betulinic acid attenuates
cyclophosphamide-induced intestinal mucosa injury by inhibiting the
NF-κB/MAPK signalling pathways and activating the Nrf2 signalling
pathway. Ecotoxicol Environ Saf. 225:1127462021. View Article : Google Scholar
|
|
20
|
Li J, Bao G, ALyafeai E, Ding J, Li S,
Sheng S, Shen Z, Jia Z, Lin C, Zhang C, et al: Betulinic acid
enhances the viability of random-pattern skin flaps by activating
autophagy. Front Pharmacol. 10:10172019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu Y, Bi Y, Mo C, Zeng T, Huang S, Gao L,
Sun X and Lv Z: Betulinic acid attenuates liver fibrosis by
inducing autophagy via the mitogen-activated protein
kinase/extracellular signal-regulated kinase pathway. J Nat Med.
73:179–189. 2019. View Article : Google Scholar
|
|
22
|
Liu B, Wu Y, Liang T, Zhou Y, Chen G, He
J, Ji C, Liu P, Zhang C, Lin J, et al: Betulinic acid attenuates
osteoarthritis via limiting NLRP3 inflammasome activation to
decrease interleukin-1β maturation and secretion. Mediators
Inflamm. 2023:37064212023. View Article : Google Scholar
|
|
23
|
Wu C, Chen H, Zhuang R, Zhang H, Wang Y,
Hu X, Xu Y, Li J, Li Y, Wang X, et al: Betulinic acid inhibits
pyroptosis in spinal cord injury by augmenting autophagy via the
AMPK-mTOR-TFEB signaling pathway. Int J Biol Sci. 17:1138–1152.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu T, Wang L, Liang P, Wang X, Liu Y, Cai
J, She Y, Wang D, Wang Z, Guo Z, et al: USP19 suppresses
inflammation and promotes M2-like macrophage polarization by
manipulating NLRP3 function via autophagy. Cell Mol Immunol.
18:2431–2442. 2021. View Article : Google Scholar
|
|
25
|
Behera J, Ison J, Tyagi A, Mbalaviele G
and Tyagi N: Mechanisms of autophagy and mitophagy in skeletal
development, diseases and therapeutics. Life Sci. 301:1205952022.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang J, Zhang Y, Cao J, Wang Y, Anwar N,
Zhang Z, Zhang D, Ma Y, Xiao Y, Xiao L and Wang X: The role of
autophagy in bone metabolism and clinical significance. Autophagy.
19:2409–2427. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lin Z, Gu Y, Liu Y, Chen Z, Fang S, Wang
Z, Liu Z, Lin Q, Hu Y, Jiang N, et al: Melatonin attenuates
inflammatory bone loss by alleviating mitophagy and lactate
production. Apoptosis. 30:1351–1371. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tang N, Zhao H, Zhang H and Dong Y: Effect
of autophagy gene DRAM on proliferation, cell cycle, apoptosis, and
autophagy of osteoblast in osteoporosis rats. J Cell Physiol.
234:5023–5032. 2019. View Article : Google Scholar
|
|
29
|
Qiao J, Liu A, Sun C and Liu Q: HIF1A
overexpression promotes osteoblast differentiation through
activation of autophagy to alleviate osteoporosis. Sci Rep.
15:303702025. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
31
|
Choi H, Jeong BC, Kook MS and Koh JT:
Betulinic acid synergically enhances BMP2-induced bone formation
via stimulating Smad 1/5/8 and p38 pathways. J Biomed Sci.
23:452016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang Y, He N, Zhou X, Wang F, Cai H,
Huang SH, Chen X, Hu Z and Jin X: Betulinic acid induces
autophagy-dependent apoptosis via Bmi-1/ROS/AMPK-mTOR-ULK1 axis in
human bladder cancer cells. Aging (Albany NY). 13:21251–21267.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang S, Deng Z, Ma Y, Jin J, Qi F, Li S,
Liu C, Lyu FJ and Zheng Q: The role of autophagy and mitophagy in
bone metabolic disorders. Int J Biol Sci. 16:2675–2691. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Du J, Wang Y, Wu C, Zhang X, Zhang X and
Xu X: Targeting bone homeostasis regulation: Potential of
traditional Chinese medicine flavonoids in the treatment of
osteoporosis. Front Pharmacol. 15:13618642024. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kim JM, Lin C, Stavre Z, Greenblatt MB and
Shim JH: Osteoblast-osteoclast communication and bone homeostasis.
Cells. 9:20732020. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li J, Chen X, Lu L and Yu X: The
relationship between bone marrow adipose tissue and bone metabolism
in postmenopausal osteoporosis. Cytokine Growth Factor Rev.
52:88–98. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang C, Li H, Li J, Hu J, Yang K and Tao
L: Oxidative stress: A common pathological state in a high-risk
population for osteoporosis. Biomed Pharmacother. 163:1148342023.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Weitzmann MN and Pacifici R: Estrogen
deficiency and bone loss: An inflammatory tale. J Clin Invest.
116:1186–1194. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Cheng CH, Chen LR and Chen KH:
Osteoporosis due to hormone imbalance: An overview of the effects
of estrogen deficiency and glucocorticoid overuse on bone turnover.
Int J Mol Sci. 23:13762022. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wu M, Chen G and Li YP: TGF-β and BMP
signaling in osteoblast, skeletal development, and bone formation,
homeostasis and disease. Bone Res. 4:160092016. View Article : Google Scholar
|
|
41
|
Zanotti S and Canalis E: Notch signaling
and the skeleton. Endocr Rev. 37:223–253. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Uluçkan Ö, Jimenez M, Karbach S, Jeschke
A, Graña O, Keller J, Busse B, Croxford AL, Finzel S, Koenders M,
et al: Chronic skin inflammation leads to bone loss by
IL-17-mediated inhibition of Wnt signaling in osteoblasts. Sci
Transl Med. 8:330ra3372016. View Article : Google Scholar
|
|
43
|
Zhang H, Hilton MJ, Anolik JH, Welle SL,
Zhao C, Yao Z, Li X, Wang Z, Boyce BF and Xing L: NOTCH inhibits
osteoblast formation in inflammatory arthritis via noncanonical
NF-κB. J Clin Invest. 124:3200–3214. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Livshits G and Kalinkovich A: Targeting
chronic inflammation as a potential adjuvant therapy for
osteoporosis. Life Sci. 306:1208472022. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Iantomasi T, Romagnoli C, Palmini G,
Donati S, Falsetti I, Miglietta F, Aurilia C, Marini F, Giusti F
and Brandi ML: Oxidative stress and inflammation in osteoporosis:
Molecular mechanisms involved and the relationship with microRNAs.
Int J Mol Sci. 24:37722023. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Abais JM, Xia M, Zhang Y, Boini KM and Li
PL: Redox regulation of NLRP3 inflammasomes: ROS as trigger or
effector? Antioxid Redox Signal. 22:1111–1129. 2015. View Article : Google Scholar
|
|
47
|
Que X, Zheng S, Song Q, Pei H and Zhang P:
Fantastic voyage: The journey of NLRP3 inflammasome activation.
Genes Dis. 11:819–829. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Fu J and Wu H: Structural mechanisms of
NLRP3 inflammasome assembly and activation. Annu Rev Immunol.
41:301–316. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Jiang W, Li X, Dong S and Zhou W:
Betulinic acid in the treatment of tumour diseases: Application and
research progress. Biomed Pharmacother. 142:1119902021. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Feng R, Wang Q, Yu T, Hu H, Wu G, Duan X,
Jiang R, Xu Y and Huang Y: Quercetin ameliorates bone loss in OVX
rats by modulating the intestinal flora-SCFAs-inflammatory
signaling axis. Int Immunopharmacol. 136:1123412024. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Liu S, Yao S, Yang H, Liu S and Wang Y:
Autophagy: Regulator of cell death. Cell Death Dis. 14:6482023.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Di Q, Zhao X, Tang H, Li X, Xiao Y, Wu H,
Wu Z, Quan J and Chen W: USP22 suppresses the NLRP3 inflammasome by
degrading NLRP3 via ATG5-dependent autophagy. Autophagy.
19:873–885. 2023. View Article : Google Scholar
|
|
53
|
Lin Q, Li S, Jiang N, Jin H, Shao X, Zhu
X, Wu J, Zhang M, Zhang Z, Shen J, et al: Inhibiting NLRP3
inflammasome attenuates apoptosis in contrast-induced acute kidney
injury through the upregulation of HIF1A and BNIP3-mediated
mitophagy. Autophagy. 17:2975–2990. 2021. View Article : Google Scholar
|
|
54
|
Chen H, Cheng Y, Du H, Zhang C, Zhou Y,
Zhao Z, Li Y, Friedemann T, Mei J, Schröder S, et al: Shufeng Jiedu
capsule ameliorates olfactory dysfunction via the AMPK/mTOR
autophagy pathway in a mouse model of allergic rhinitis.
Phytomedicine. 107:1544262022. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Wei J, Li Y, Liu Q, Lan Y, Wei C, Tian K,
Wu L, Lin C, Xu J, Zhao J and Yang Y: Betulinic acid protects from
bone loss in ovariectomized mice and suppresses RANKL-associated
osteoclastogenesis by inhibiting the MAPK and NFATc1 pathways.
Front Pharmacol. 11:10252020. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Jeong DH, Kwak SC, Lee MS, Yoon KH, Kim JY
and Lee CH: Betulinic acid inhibits RANKL-induced
osteoclastogenesis via attenuating Akt, NF-κB, and
PLCγ2-Ca2+ signaling and prevents inflammatory bone
loss. J Nat Prod. 83:1174–1182. 2020. View Article : Google Scholar : PubMed/NCBI
|