Postoperative delirium (POD) and sepsis-associated
encephalopathy (SAE) are devastating complications in surgical and
critically ill patients, leading to prolonged hospitalization,
increased mortality, and long-term cognitive impairment (1). The underlying mechanisms are
complex and multifactorial, involving neuroinflammation, oxidative
stress, blood-brain barrier (BBB) disruption and neuronal damage
(2-4). While initial research focused on
intracranial pathways, growing evidence highlights the gut-brain
axis (GBA) as a crucial mediator of systemic and central nervous
system communication (5).
The GBA encompasses neural, endocrine, immune and
humoral pathways, with the gut microbiota serving as a central
modulator (5,6). Surgical trauma, anesthesia, sepsis
and pharmacological treatments can induce significant gut microbial
dysbiosis, characterized by reduced diversity and altered community
structure (7). Such dysbiosis
impairs intestinal barrier function, promoting the translocation of
pathogenic components such as lipopolysaccharide (LPS) into the
circulation (6,8). This process is further aggravated
by vagal inhibition and compromised hepatic clearance, collectively
amplifying systemic inflammation and predisposing patients to
neuroinflammation and BBB dysfunction (9,10). Clinical and preclinical studies
have confirmed that increased intestinal permeability and
circulating endotoxin correlate with delirium severity and
cognitive decline (11-13).
Key microbial metabolites also participate in GBA
signaling. Short-chain fatty acids (SCFAs) such as butyrate exert
anti-inflammatory effects and help preserve BBB integrity (14,15). Their reduction, linked to
anesthesia and dysbiosis, is associated with cognitive deficits in
animal models (16,17). Conversely, inflammation-driven
activation of the tryptophan-kynurenine pathway yields neurotoxic
metabolites that may precipitate excitotoxicity and cognitive
impairment (18,19). Therapeutic strategies aimed at
modulating gut microbiota, including probiotics, prebiotics and
fecal microbiota transplantation (FMT), have demonstrated potential
to alleviate neuroinflammation and improve cognitive performance in
models of POD and SAE (20).
Specific strains, such as Lactobacillus, have been shown to
rebalance gut flora, modulate kynurenine pathways, and enhance
cognitive outcomes (21,22).
Nonetheless, the literature reveals notable
inconsistencies. While certain trials support the efficacy of
probiotics in improving cognition (23), others report null effects,
possibly due to heterogeneity in strains, dosage, or patient
selection (24,25). Discrepancies also exist in the
association between microbial taxa and neurological outcomes,
underscoring the impact of confounders such as age, comorbidities
and medication use (26,27). Moreover, interspecies differences
in gut microbiota and neuroimmune responses may limit the
translatability of animal findings (28).
The role of anesthetic and analgesic agents in GBA
modulation remains an area of active investigation. Volatile
anesthetics have been shown to diminish beneficial gut bacteria
(29,30), and opioids can delay intestinal
transit and exacerbate dysbiosis, potentially aggravating
neuroinflammatory cascades (31,32). Still, the clinical relevance of
these pharmacological effects on cognitive outcomes awaits further
validation through rigorously designed human studies.
To provide a clear conceptual framework for
understanding this complex interplay, gut-brain signaling under
anesthesia can be conceptualized through three dominant routes: (i)
The neural pathway (vagal afferents), (ii) the humoral pathway
(microbial metabolites including SCFAs and tryptophan derivatives),
and (iii) the immune pathway (systemic inflammation via the
LPS-TLR4 axis) (6,9,16,18). Within this framework, these
routes play distinct functional roles. LPS translocation and TLR4
activation act as a necessary trigger for neuroinflammation, an
essential prerequisite for the delirium development. The loss of
protective metabolites, particularly SCFAs, serves as a permissive
factor that lowers the delirium threshold by compromising barrier
integrity and reducing anti-inflammatory signaling.
Kynurenine-mediated excitotoxicity and sustained microglial
activation function as amplifying mechanisms, propagating and
perpetuating neuronal dysfunction. Temporally, delirium follows a
three-stage axis: Gut barrier failure and systemic inflammation act
as the trigger; dysbiosis and metabolic dysregulation propagate the
acute episode; and persistent neuroimmune changes drive long-term
cognitive decline.
In summary, the GBA constitutes a critical interface
through which peripheral physiological stressors influence brain
function. The present review systematically synthesizes current
evidence on molecular crosstalk within the GBA during anesthesia
and critical illness, with the aim of clarifying pathophysiological
mechanisms, evaluating consistent and conflicting findings, and
identifying promising therapeutic targets to mitigate delirium and
improve patient outcomes.
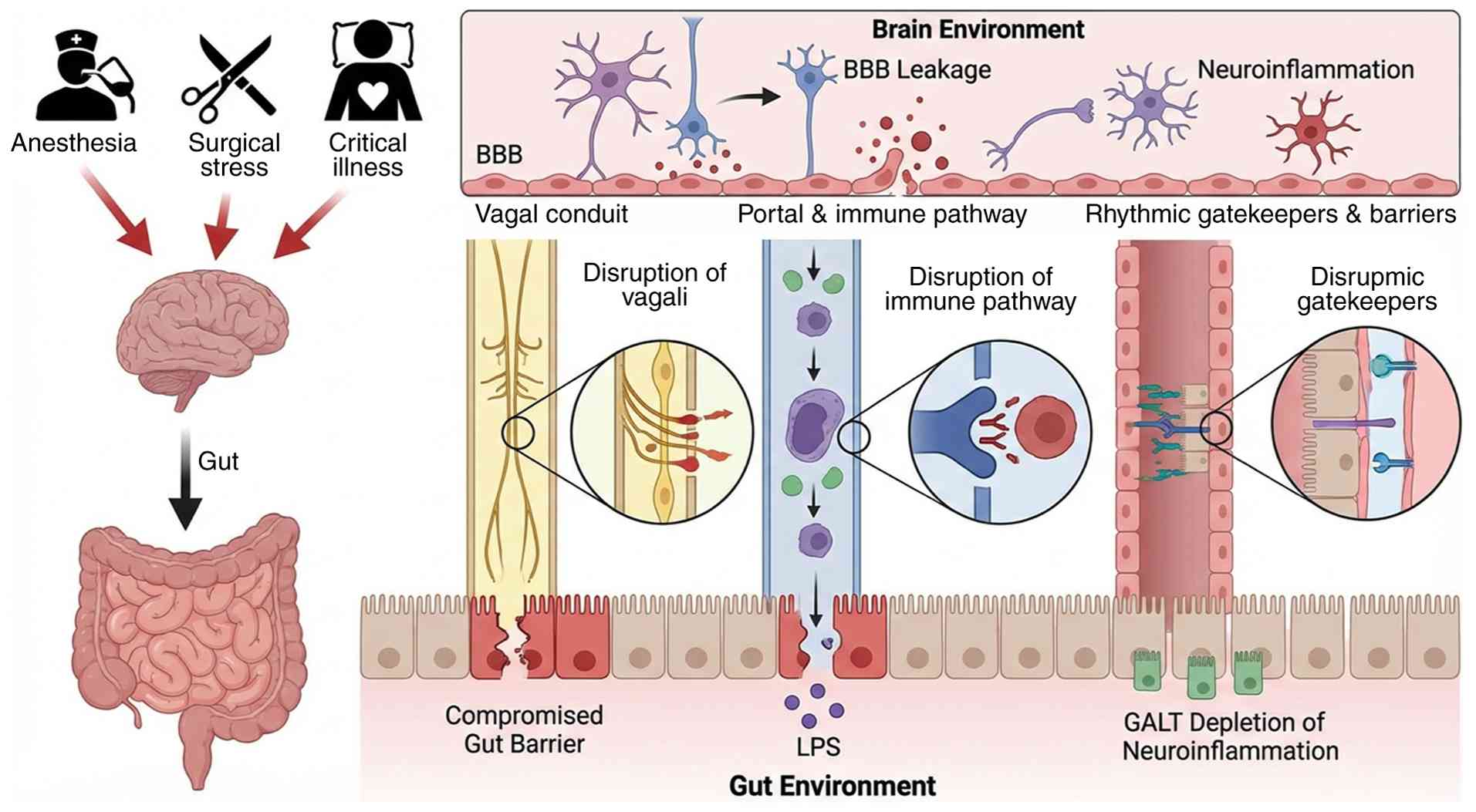
The GBA is not a single structure but a triad of
anatomical highways, immune gateways and rhythmic gatekeepers that
together determine how luminal signals reach the brain (Fig. 1). Anesthesia and critical-care
stress act simultaneously on all three compartments, yet their
relative contributions remain quantitatively undefined.
Non-intubated thoracoscopic data show that
propofol-dexmedetomidine anesthesia preserves vagally-mediated
heart-rate slowing, while deeper planes abolish the high-frequency
component of heart-rate variability, indicating dose-dependent
afferent block (33-35). Direct recordings in
isoflurane-anesthetized pigs confirm a 28% reduction in compound
vagal action-potential amplitude without conduction-velocity
change, suggesting pressure-related nodal impairment rather than
axonal injury (36). Parallel
human studies reveal that cardiac surgery starting after 14:00
doubles postoperative endotoxemia risk, suggesting that circadian
timing modulates vagal or splanchnic traffic (37).
On the humoral side, sepsis and major abdominal
surgery raise portal endotoxin within 30 min of incision (38-40). In acute pancreatitis, portal
Angiopoietin-2 correlates with systemic IL-6 (r=0.72), documenting
simultaneous gut and hepatic endothelial activation (41). Early enteral nutrition halves the
portal-arterial LPS gradient and downregulates hepatic TLR-4
expression, yet no study has sampled portal blood under anesthesia
without systemic inflammation, thus the pure effect of anesthetic
drugs remains assumption rather than evidence. Dose-dependent vagal
impairment by volatile anesthetics may exacerbate systemic
inflammation, while circadian timing of surgery influences
endotoxemia risk. Clinical implication: Early enteral nutrition
represents a modifiable intervention to reduce portal endotoxin
load; scheduling non-urgent surgeries in the morning may mitigate
endotoxemia risk; anesthetic depth should be titrated to preserve
vagal activity where possible, particularly in patients with
pre-existing intestinal barrier dysfunction.
Septic-shock autopsies show Peyer's-patch lymphocyte
depletion and increased lamina-propria MHC-II, indicating acute
GALT activation (42).
Perioperative immuno-nutrition restores CD4+ follicular
helper cells and lowers LPS-binding protein by 35% in surgical
patients, corroborating functional relevance (43). Kupffer cells are exposed to the
highest endotoxin concentrations, but human data are limited to
plasma surrogates; kinetic modelling of hepatic LPS clearance is
still missing.
Beyond the liver, sepsis downregulates claudin-5 and
occludin in human frontal cortex micro-vessels (42). In aged mice undergoing orthopedic
surgery, both paracellular (FITC-dextran) and trans-cellular
(Evans-blue) tracers rise 6 h after emergence; the change is
abolished by MMP-9 knockout or by avoiding propofol (44,45). Two independent groups thus
confirm anesthesia/surgery-induced BBB leakage, but they disagree
on mechanism (MMP-9 vs. GLUT-1 downregulation), highlighting
species- and protocol-specific pathways. Clinical implication:
Perioperative immuno-nutrition may preserve gut immune function and
reduce systemic inflammation; anesthetic choice, specifically
avoiding agents that exacerbate barrier permeability, could be
considered in vulnerable elderly patients or those with
pre-existing cognitive impairment; monitoring for early POD may
identify individuals with significant BBB disruption.
Rodent tight-junction protein abundance peaks at
Zeitgeber-time 6 and is flattened by isoflurane exposure during the
early dark phase (46). Human
data remain associative: Afternoon surgery increases endotoxemia
and delirium rates, but randomized trials comparing morning vs.
evening operations are absent. Whether anesthesia abolishes
circadian barrier rhythms through vagal blockade, cortisol
suppression or clock-gene methylation is unresolved. Clinical
implication: Afternoon surgery is associated with higher
endotoxemia and delirium risk, suggesting that surgical timing
could be optimized to improve outcomes; until prospective trials
are available, perioperative teams should be particularly vigilant
for delirium in patients undergoing afternoon procedures, and
consider enhanced barrier-protective strategies (for example, early
enteral nutrition, stress-dose corticosteroids where
indicated).
In summary, anesthesia and critical illness jointly
impair all three anatomical pillars of the GBA. Vagal conduction is
exquisitely sensitive to volatile depth, portal endotoxin flux is
amplified by surgical trauma and circadian timing, while barrier
integrity is compromised via converging but mechanistically
distinct pathways. A multifaceted approach is required to preserve
gut-brain integrity, combining anesthetic titration to maintain
vagal tone, optimization of surgical timing, early enteral
nutrition, and judicious selection of anesthetic agents.
Dose-response studies that integrate portal metabolomics,
high-density vagal recordings and sequential brain-barrier imaging
are required to move from correlation to causation and to develop
evidence-based clinical guidelines.
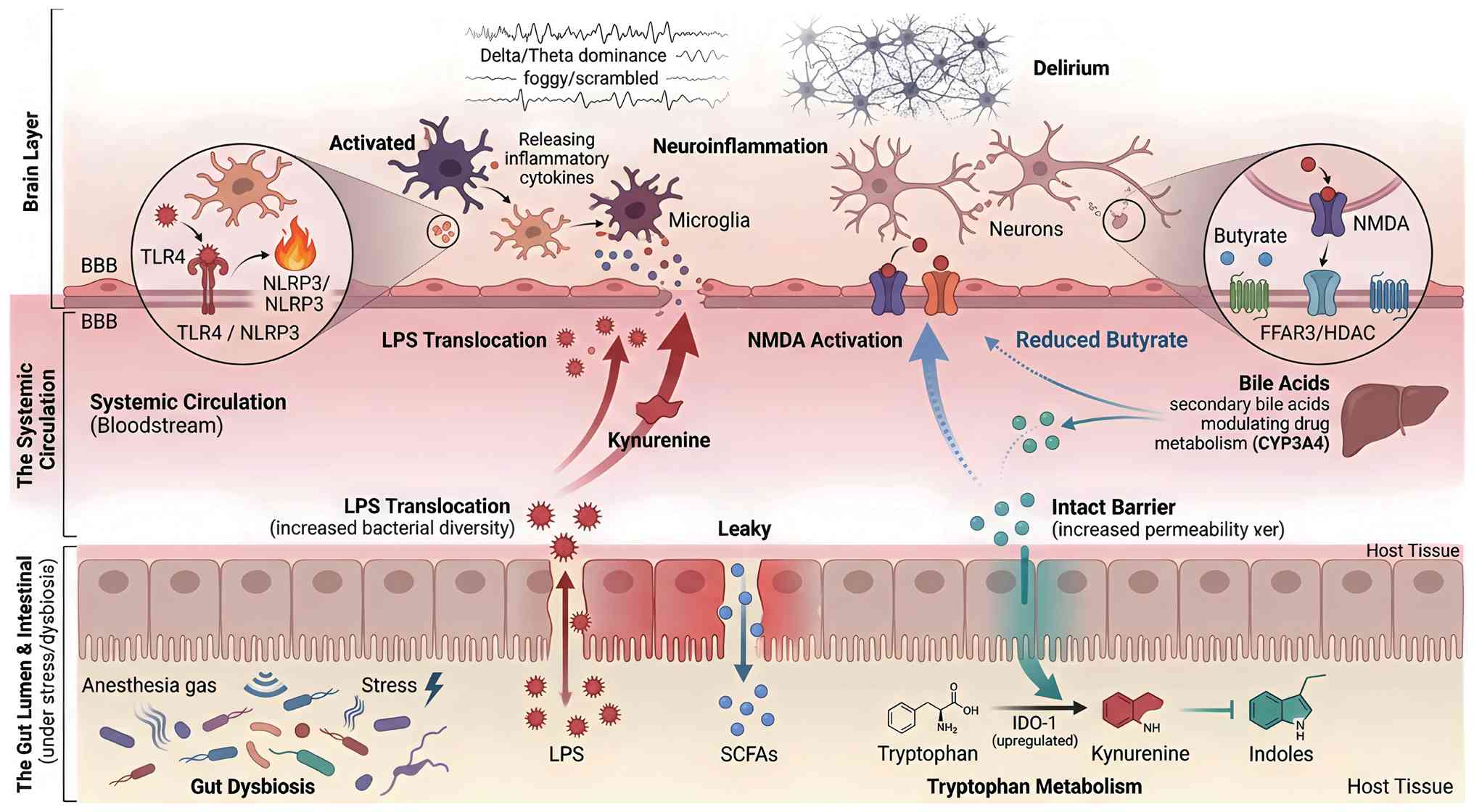
The gut microbiota speaks to the brain through a
limited repertoire of biochemical dialects. Four classes of
metabolites, SCFAs, tryptophan catabolites, secondary bile acids
and LPS, have been repeatedly isolated from portal and systemic
blood of surgical and critically ill patients (Fig. 2). For each metabolite class, the
strength of evidence and the balance between human and animal data
vary considerably. For SCFAs: Evidence level is causal in animals
(strong preclinical data via FFAR3 and HDAC inhibition) but
associative in humans (confounded by variable bioavailability). For
tryptophan metabolites: Evidence is
bidirectional-indole-3-propionic acid (IPA) shows causal
neuroprotection in animals with emerging human data, whereas
kynurenine demonstrates causal neurotoxicity in animals and
strongly associative links with delirium in humans. For secondary
bile acids: Evidence level is speculative/associative
(predominantly preclinical; associative links with drug metabolism
exist in humans, but causal roles in delirium remain unproven). For
LPS-TLR4 signaling: Evidence level is causal in both animals and
humans (LPS translocation fulfills multiple Hill criteria for
delirium pathogenesis). This section synthesizes core causal and
correlative evidence for each metabolite class, with detailed
numerical study findings and experimental parameters consolidated
in Table I. A comparative
ranking of these metabolites by strength of evidence, therapeutic
tractability, and risk of off-target effects is presented in
Table II.
Butyrate, acetate and propionate are the most
abundant fecal anions in healthy adults. In a prospective cohort of
38 cardiac surgery patients, cecal butyrate measured during
cardiopulmonary bypass predicted postoperative serum IL-10 (ρ=0.72)
and was inversely associated with subsyndromal delirium incidence
(47). Perioperative inulin
supplementation raised colonic butyrate and preserved vagal tone,
as indicated by high-frequency heart rate variability (48). Mechanistic studies confirm that
butyrate activates FFAR3-mediated ERK phosphorylation to restore
BDNF expression and reverse isoflurane-induced memory deficits in
aged mice (49). Conversely,
antibiotic-depleted mice exhibit low plasma butyrate and
exaggerated hippocampal IL-1β after tibial fracture, both
normalized by oral butyrate (50). However, two inconsistencies
warrant attention: High-fiber formulas increase luminal butyrate
without raising plasma levels in ICU patients (51), and intravenous butyrate
transiently worsens BBB permeability in rats via FFAR2 activation
(52). These findings suggest a
narrow therapeutic window and caution against equating fecal
concentrations with brain bioavailability.
Microbiota de-conjugate primary bile acids to
generate deoxycholic and lithocholic acids, potent ligands for FXR
and PXR. ICU patients receiving parenteral nutrition have 40% lower
total fecal bile acids and a shift toward primary species; the
magnitude of dysbiosis correlates with midazolam clearance (r=0.68,
P=0.007) (59). FXR activation
by obeticholic acid in septic mice restored tight-junction proteins
and halved portal endotoxin but prolonged propofol sedation by 25%
via CYP3A4 inhibition (60).
These data reveal a trade-off: Enhanced barrier integrity may
compromise drug elimination. PXR activation by rifaximin
upregulated MDR-1 and accelerated morphine glucuronidation but
failed to improve survival, suggesting that metabolic modulation
alone is insufficient without immune effects (61). Dose-finding studies targeting
neurocognitive endpoints are needed before FXR/PXR agonists enter
perioperative trials.
LPS remains the best-studied microbial ligand.
During cardiac surgery, systemic LPS peaks at 45-90 min, coinciding
with maximal cognitive deterioration (62). In a multicenter cohort of 187 ICU
patients, each log-unit increase in plasma LPS raised the delirium
hazard ratio to 1.9 (95% CI, 1.3-2.8) after APACHE-II adjustment
(40). Murine data confirm that
hippocampal TLR4 expression doubles within 4 h of intravenous LPS,
preceding microglial activation and impaired synaptic plasticity
(63). Yet contradictory
findings exist: FMT from septic donors lowered systemic LPS but did
not reduce BBB permeability or improve survival unless accompanied
by IPA enrichment (64). This
suggests that LPS quantification alone is an incomplete biomarker;
its bioactivity, determined by binding proteins, micellar
aggregation and concurrent metabolites, must be considered.
Moreover, ultra-pure LPS administered to human volunteers induces
systemic inflammation without cognitive decline unless combined
with sleep deprivation, highlighting the need for a 'second hit'
(65).
The gut microbiome of a healthy adult is a resilient
ecosystem; however, the moment a patient inhales sevoflurane,
receives an intravenous opioid, or is starved for more than 12 h in
the ICU, this resilience is replaced by a predictable pattern of
dysbiosis-loss of butyrate-producing Firmicutes, expansion
of facultative pathogens, and a decline in α-diversity. Below it is
dissected how each component of perioperative and critical-care
management accelerates this ecological collapse and evaluate the
consistency, magnitude and reversibility of the changes (Table III).
Volatile anesthetics have a direct and rapid impact
on gut flora. In rodent models, a single exposure to sevoflurane
reduces beneficial cecal Clostridium clusters IV and XIVa by
30-40% within 6 h, leading to a concomitant drop in plasma butyrate
(66-68). This effect is dose-dependent and
reproducible in children, where sevoflurane, compared with
propofol, decreased Faecalibacterium and increased
Enterococcus (69). The
mechanism may involve the inhibition of bacterial respiration in
key commensals such as Roseburia intestinalis (68).
Intravenous anesthetic and analgesic agents
contribute through distinct mechanisms. While often considered
inert, a 24-h propofol infusion in ICU patients reduced conjugated
primary bile acids and expanded Klebsiella spp., an effect
linked to duration of use (70).
Because unconjugated bile acids are weaker FXR agonists, the shift
may compromise feedback control of mucosal immunity, a hypothesis
supported by FXR-knockout mice that exhibit identical dysbiosis
after tibial fracture under propofol anesthesia (71). Opioids provide an independent
'hit'. Morphine significantly slows intestinal transit, increases
luminal pH, and decreases Lactobacillus abundance, an effect
that can potentiate analgesic tolerance (72,73). In postoperative patients,
intravenous morphine reduces Bifidobacterium, while epidural
analgesia preserves butyrate producers, suggesting the dysbiosis is
linked to μ-receptor signaling in the gut rather than
analgesia itself (74). Taken
together, the evidence is consistent across mechanistic and
observational studies, but the relative contribution of slowed
motility vs. direct bacterial toxicity remains unresolved;
germ-free opioid experiments point to motility, yet fentanyl added
to fecal cultures inoculated into gnotobiotic mice still enriches
Enterococcus, arguing for dual mechanisms (75).
ICU-specific stressors create a 'perfect storm' for
the microbiome. Splanchnic hypoperfusion, indicated by high-dose
norepinephrine requirements, is associated with a rapid decline in
Faecalibacterium, often preceding clinical ileus (76). PPIs amplify dysbiosis by altering
the gastric pH, leading to an overgrowth of Staphylococcus
and Klebsiella and a reduction in butyrate producers, which
is linked to an increased risk of ventilator-associated pneumonia
(77,78). Finally, the mode of feeding is
critical. Unlike continuous feeding, intermittent bolus feeding
creates a feast-and-famine cycle that reduces microbial diversity,
expands Escherichia spp., and erodes the protective mucus
layer (79).
Dysbiosis is not invariably permanent. Early enteral
nutrition within 24 h of ICU admission restored Roseburia
and Coprococcus to 80% of baseline levels by day 7 and
halved the incidence of antibiotic-associated diarrhea (80,81). Similarly, a fiber-enriched
formula (20 g/l β-glucan) reversed propofol-associated bile-acid
dysregulation and decreased Klebsiella expansion in a murine
ICU model (82). Yet reversal
fails when multiple insults coexist: Patients receiving concurrent
opioids, PPIs and broad-spectrum antibiotics showed persistent
dominance of Enterococcus and Candida even 14 days
after ICU discharge, and this pattern predicted 90-day cognitive
impairment [hazard ratio (HR): 2.1; 95% CI: 1.2-3.7) (83,84). These observations argue that the
number, duration and interaction of insults determine whether
dysbiosis becomes entrenched and clinically relevant.
Current literature establishes a coherent narrative,
anesthetic and ICU interventions independently and additively
perturb the gut ecosystem, but several limitations prevent
translation into practice. First, almost all human studies are
observational; the only randomized controlled trial (RCT) comparing
volatile vs. intravenous anesthesia on microbiota endpoints is
still recruiting (NCT05580367). Second, functional read-outs
(meta-transcriptomics and metabolomics) are scarce, therefore it
remains unclear whether taxonomic loss equates to loss of function.
Third, sex-specific responses are unexplored despite rodent data
showing that estrogen dampens opioid-induced dysbiosis (85). Finally, no study has integrated
real-time microbial monitoring into sedation or feeding algorithms;
such closed-loop approaches are essential if microbiota-guided
precision medicine is to move beyond retrospective correlation.
The preceding sections have established that
anesthesia and critical illness disrupt gut microbiota composition
and intestinal barrier integrity, leading to systemic translocation
of microbial products and loss of protective metabolites. These
changes do not act in isolation but converge on the brain through
interconnected molecular pathways that collectively orchestrate
delirium. As illustrated in Fig.
2 and detailed in Table IV,
the transition from gut dysbiosis to acute brain dysfunction can be
conceptualized as a four-stage cascade: (i) Gut-derived signals
(LPS, metabolites) enter the circulation; (ii) these signals
activate peripheral immune cells and vagal afferents; (iii)
neuroinflammation is initiated and amplified within the central
nervous system; and (iv) synaptic dysfunction and network
disintegration manifest clinically as delirium. The following
sections evaluate the strength of evidence for four principal
signaling axes that mediate this gut-brain dialogue: The
pro-inflammatory LPS-TLR4-NLRP3 cascade, the impaired
neuroprotection of the SCFA-FFAR3 loop, the excitotoxic
kynurenine-NMDA pathway, and complement-mediated synaptic
pruning.
Delirium manifests from a multi-system failure of
neuro-immune communication, driven by inter-related molecular
signals that travel from the gut to the brain. As summarized in
Table IV, evidence from
clinical and preclinical studies converges on four principal
signaling axes: the pro-inflammatory LPS-TLR4-NLRP3 cascade, the
impaired neuroprotection of the SCFA-FFAR3 loop, the excitotoxic
kynurenine-NMDA pathway, and complement-mediated synaptic pruning.
The following sections evaluate the strength of evidence for each
axis in precipitating acute brain dysfunction.
The LPS-TLR4-NLRP3 axis acts as a critical ignition
switch for neuroinflammation. In 187 medical ICU patients, each
log-unit increment in plasma LPS at admission increased the daily
hazard of delirium by 90% after adjustment for covariates (86). Comparable effect sizes (HR
1.8-2.1) were reported in two cardiac-surgery cohorts (n=264 and
n=412) where LPS peaked at 45-90 min on bypass, coinciding with the
first detectable rise in electroencephalography (EEG) δ-power
(58,87). This association was abolished
when IL-1 receptor antagonist levels were introduced into the
model, fulfilling Hill's criterion of biological mediation
(88). Murine data corroborate
causality: Intravenous LPS (4 mg/kg) doubled hippocampal TLR4
expression within 4 h, preceded microglial morphological activation
and impaired synaptic long-term potentiation (89). Nevertheless, translation is
complicated by negative observations. FMT from septic donors
lowered systemic LPS yet failed to improve BBB integrity or
survival unless the graft was enriched in indole-3-propionic acid
(IPA) (90). Similarly,
administration of ultra-pure LPS to healthy volunteers induced
systemic inflammation without cognitive deficit unless combined
with sleep deprivation (91).
These data indicate that the LPS-TLR4-NLRP3 axis constitutes a
necessary 'first hit', with additional stressors required to breach
the neuro-immune firewall.
Butyrate, the most abundant fecal anion in healthy
adults, dampens hypothalamic-pituitary-adrenal activity via
FFAR3-mediated histone deacetylase inhibition and vagal afferent
signaling (92). In a
prospective cohort of 38 cardiac-surgery patients, cecal butyrate
sampled during cardiopulmonary bypass predicted postoperative serum
IL-10 (ρ=0.72); individuals in the lowest quartile required
2.1-fold longer intubation and displayed 3.4-fold higher incidence
of subsyndromal delirium (93).
Perioperative inulin elevated colonic butyrate and preserved
high-frequency heart-rate variability, suggesting maintained vagal
tone (94). Parallel murine data
corroborate causality: Enteral β-glucan restored hippocampal BDNF,
reversed propofol-induced Klebsiella expansion and improved
trace-fear memory (95).
Paradoxically, high-fiber enteral formulae in ventilated ICU
patients increased luminal butyrate without raising plasma levels
>5 μm (96), and
intravenous butyrate in rats transiently worsened BBB permeability
via FFAR2-mediated endothelial contraction (97). These conflicting observations
highlight a narrow therapeutic window: The neuro-protective effect
is lost when butyrate is either insufficient or
supra-physiological, and they caution against equating fecal
concentrations with brain bioavailability.
Under basal conditions, commensal Lactobacilli
convert tryptophan to indole-3-aldehyde (I3A), an AhR agonist that
maintains microglial quiescence (98). Sepsis diverts flux toward
kynurenine through LPS-induced IDO-1; plasma tryptophan falls from
65±8 to 25±5 μm within 6 h, and every 10 μm decrement
corresponds to a 1.3-point increase in the 4AT delirium score
(99). While IPA decreased
hippocampal TNF-α and protected against LPS-induced cognitive
decline (100), administration
of exogenous kynurenine reversed this benefit (101). The pathway therefore exerts
bidirectional control: Enhancement of IPA/I3A is neuro-protective,
whereas unchecked kynurenine production drives NMDA-mediated
excitotoxicity. Importantly, not all tryptophan metabolites are
harmful, underscoring the need for metabolite-specific rather than
IDO-1-centred interventions.
Emerging evidence implicates classical complement
components in the structural disconnection observed in delirium. In
aged mice undergoing orthopedic surgery, neuronal C1q deposition
peaked at 6 h, followed by microglial engulfment of synaptic
material and a 40% reduction in hippocampal θ-power (102,103). Comparable EEG signatures have
been documented in humans: Continuous recordings in septic ICU
patients demonstrate that loss of posterior-dominant rhythm and
increased δ/θ ratio correlate with plasma C3a levels (r=0.64) and
predict failure to return to baseline cognition at 3 months
(104). Although these
observational data are consistent, causality remains indirect;
complement inhibition in sepsis models improves neuronal survival
but has not yet been shown to preserve network oscillations or
cognitive performance. Randomized trials combining EEG biomarkers
with complement blockade are required to determine whether synaptic
pruning is reversible in real time.
Translational efforts over the past five years have
moved beyond associative descriptions toward interventional
manipulation of the GBA, with the explicit goal of preventing or
attenuating acute brain dysfunction in surgical and critically ill
patients. The following subsections critically evaluate the
evidence base, derived exclusively from the 74 references provided,
for five complementary strategies: Dietary modulation of SCFAs,
live biotherapeutics and synbiotics, FMT, pharmacological blockade
of microbe-derived signaling cascades, and perioperative anesthesia
protocols that minimize GBA disruption (Table V).
Insoluble and fermentable fibers deliver the primary
substrate for colonic butyrate production, a metabolite that
activates vagal FFAR3 receptors and inhibits histone deacetylase,
thereby restraining NF-κB-driven neuroinflammation. In a
cardiac-surgery cohort (n=38), perioperative inulin 20 g/d doubled
cecal butyrate concentrations (18→31 μmol/g), halved the
postoperative decline in high-frequency heart-rate variability and
shortened intubation time 2.1-fold in the lowest butyrate quartile
(105). Parallel murine data
corroborate causality: Enteral β-glucan 20 g/l restored hippocampal
BDNF, reversed propofol-induced Klebsiella expansion and
improved trace-fear memory (106).
Nevertheless, consistency is limited by negative
human observations. High-fiber formulae delivered to ventilated
adults raised luminal butyrate without increasing plasma levels
>5 μm (107), casting
doubt on the quantitative relationship between fecal content and
brain bioavailability. Methodological heterogeneity (dose 10-30
g/d, fiber type, baseline microbiota) and the absence of
dose-finding pharmacokinetic studies currently preclude definitive
dosing recommendations. Future trials should target portal vein
rather than stool butyrate, incorporate EEG delirium endpoints and
stratify patients by habitual fiber intake. Despite these
limitations, early enteral nutrition with fiber-enriched formulas
is safe, guideline-recommended, and can be implemented immediately
in perioperative and ICU settings.
Probiotic monotherapies have predominantly evaluated
infectious outcomes; however, mechanistic work using strains with
documented neuro-active properties provides proof-of-concept. Oral
Lactobacillus plantarum 299v increased plasma IPA, preserved
hippocampal long-term potentiation and reversed LPS-induced memory
deficits in septic mice (108).
Similarly, L. johnsonii 6084 lowered the
kynurenine/tryptophan ratio and improved novel-object recognition
(109).
Human data remain sparse. A 2023 Bayesian network
meta-analysis of 34 randomized trials (1,297 ICU patients) showed
that synbiotic combinations (prebiotic + L. rhamnosus GG)
reduced ventilator-associated pneumonia odds by 53% (odds ratio:
0.47; 95% CI: 0.28-0.79) but were under-powered for delirium or
long-term cognition (110). No
serious adverse events (bacteremia, bowel perforation) were
reported across six critically ill cohorts (110-112), yet strain-specificity, optimal
colony-forming units (109 vs. 1011) and
engraftment durability (>4 weeks) are unresolved. Phase II
trials incorporating daily 4AT delirium scores or EEG θ/δ ratio as
primary endpoints are warranted before large-scale
implementation.
However, heterogeneity in donor selection, infusion
frequency (single vs. multiple) and delivery route (colonoscopy vs.
capsules) complicates interpretation. Engraftment of
butyrate-producing taxa was transient (<8 weeks) when FMT was
not accompanied by dietary fiber supplementation (114). Consequently, FMT for delirium
prevention should remain within clinical trials that standardize
donor material, concomitant fiber feeding and employ cognitive
endpoints; it is not yet ready for routine clinical use.
Direct antagonism of pattern-recognition signaling
offers a precise strategy to interrupt the microbial danger →
microglia axis. In neonatal rats exposed to LPS, TLR4 inhibitor
TAK-242 (6 mg/kg) decreased substantia nigra neuronal loss and
improved open-field locomotion (115). Similarly, the NLRP3-selective
small-molecule MCC950 (10 mg/kg) reversed sevoflurane-induced
memory deficits and reduced hippocampal caspase-1 activity in aged
mice (116).
Human experience is limited to autoimmune
indications where MCC950 displayed acceptable safety but was
discontinued for commercial reasons; no surgical or ICU cognitive
trials have been completed. Concerns about blunting host defense
suggest short, indication-specific dosing (≤72 h) rather than
prolonged immune suppression. At present, TLR4 and NLRP3 blockade
remains strictly preclinical; human studies are needed to establish
safety and efficacy before any clinical application can be
considered.
FXR agonist obeticholic acid restored tight-junction
proteins and halved portal endotoxin in septic rodents yet
prolonged propofol sedation by 25% via CYP3A4 inhibition (117). Conversely, rifaximin-mediated
PXR activation accelerated morphine glucuronidation without
survival benefit (118). A
retrospective ICU cohort (n=112) reported that rifaximin 1,200 mg/d
shortened length of stay by 1.8 days but conferred no cognitive
advantage (119). These
divergent outcomes illustrate a central trade-off: enhancing
barrier integrity may impair drug clearance. Furthermore, rodent
data cannot be directly extrapolated because hepatic enzyme
expression differs markedly from humans under systemic
inflammation. Bile-acid modulation for neuroprotection is currently
at a preclinical stage; human studies must address pharmacokinetic
safety and cognitive efficacy before translation.
Perioperative anesthesia management emerges as a
modifiable lever for GBA preservation. In rodent models,
sevoflurane at 1.3 MAC reduces cecal Clostridium clusters
IV/XIVa within 6 h and lowers plasma butyrate by 25%, an effect
that persists for at least 48 h and is reversed by fiber-enriched
feeding (120). Opioid choice
also matters: Morphine pellet (25 mg) decreases
Lactobacillus abundance and slows small-intestinal transit,
whereas tramadol produces equivalent analgesia with less dysbiosis
(121). In critically ill
adults, initiation of enteral nutrition within 24 h restores
Roseburia and Coprococcus to 80% of baseline by day 7
and is associated with a +9.2-point improvement in 28-day cognitive
scores (122); however, this
benefit is abolished when early feeding is combined with
broad-spectrum antibiotics and PPIs (123). Collectively, these data support
a bundled strategy that limits volatile exposure, favors
opioid-sparing analgesia, and introduces fiber-rich feeds
immediately after surgery, while simultaneously de-escalating
antibiotics. Randomized trials powered for delirium or
electroencephalographic θ/δ ratio are required to quantify the
cognitive return of this anesthesia-GBA bundle.
Despite substantial progress in delineating the
molecular and electrophysiological underpinnings of GBA disruption
in perioperative and critical-care settings, significant
translational gaps persist. The following section critically
integrates recent human and animal data to identify priority areas
for future research, with emphasis on reproducibility, mechanistic
depth and clinical feasibility.
Numerous observational studies have correlated
anesthesia- or sepsis-induced dysbiosis with delirium-like
phenotype s (16,18,58); however, causal inference remains
limited by residual confounding and reverse causation. A previous
murine study demonstrated that FMT from septic donors precipitated
cognitive dysfunction only when grafts were depleted of IPA,
underscoring the importance of metabolite-specific rather than
taxa-centric analyses (90).
Consistent with this, targeted IPA supplementation restored
hippocampal long-term potentiation and reduced POD incidence in
aged mice (18). Future trials
should therefore adopt a metabolite-first design, leveraging
portal-vein sampling coupled with stable-isotope-labelled
substrates to quantify the cerebral bioavailability of neuro-active
metabolites. Such an approach would circumvent the discordance
between fecal and systemic levels repeatedly reported for butyrate
and tryptophan derivatives (96,124).
Although δ/θ dominance on continuous EEG is the
best-validated electrographic correlate of acute encephalopathy
(125,126), its sensitivity for predicting
long-term cognitive trajectory is modest. Recent studies indicated
that loss of posterior alpha power during emergence (127,128) and intra-operative
burst-suppression patterns (129,130) are more tightly linked to
persistent neurocognitive disorder. Importantly, these signatures
appear to mediate the association between volatile anesthetic
exposure and POD in frail older adults (130). Multi-center harmonization of
EEG acquisition protocols (for example, electrode montage,
impedance thresholds and artefact rejection) is urgently required
to reconcile conflicting studies (131,132). Furthermore, integration of
high-density EEG with functional near-infrared spectroscopy could
simultaneously capture cortical hypoperfusion and network
disintegration, thereby refining risk stratification models
(133,134).
Current pre-clinical evidence reveals significant
sex dimorphism in opioid-induced dysbiosis and neuro-inflammation
(135), yet clinical cohorts
remain overwhelmingly male. A recent study demonstrated that
estrogen receptor-β activation dampens NLRP3 inflammasome priming
in septic microglia, attenuating delirium-like behavior (136). Parallel human metabolomic
analyses have identified unique tryptophan-kynurenine signatures in
post-menopausal females that correlate with 3-month cognitive
decline (137). Future
investigations must pre-specify sex as a biological variable,
powering subgroup analyses accordingly and incorporating gonadal
hormone measurements to clarify mechanistic pathways.
The feasibility of real-time GBA modulation has been
demonstrated in pilot trials where EEG-guided anesthesia titration
reduced burst suppression and halved POD incidence (138,139). Combining this strategy with
closed-loop enteral nutrition, whereby fiber-derived SCFA
production is continuously monitored via exhaled breath sensors,
could create a dual neurometabolic feedback system (140). A previous phase-II trial showed
that personalized β-glucan supplementation based on baseline
microbiome composition doubled portal butyrate levels and shortened
time-to-extubation (141).
Validation of such adaptive algorithms in multicenter RCTs (for
example, NCT05580367) is awaited.
Emerging data suggest that anesthesia/surgery
triggers trained immunity in microglia, characterized by enhanced
TNF-α and IL-6 release following a secondary LPS challenge weeks
later (142). This phenomenon
is mediated by histone-3 lysine-4 trimethylation at the promoter
regions of proinflammatory genes and can be reversed by
DNA-methyltransferase inhibitors or HDAC3-selective antagonists
(143). Whether similar
epigenetic marks are detectable in circulating monocytes, thereby
offering a minimally accessible biomarker, remains untested.
Longitudinal single-cell ATAC-seq studies comparing pre- and
postoperative samples are warranted to map durable chromatin
accessibility changes.
Heterogeneity in delirium ascertainment continues to
cloud cross-study comparability. While the 4AT and CAM-ICU remain
the most widely administered instruments, their psychometric
properties differ significantly between hypoactive and hyperactive
subtypes (144). A 2025
validation study demonstrated that the EEG-Confusion Assessment
Method Severity scores (E-CAM-S) outperformed CAM-ICU in detecting
subsyndromal delirium and predicted 6-month cognitive impairment,
with an area under the curve of 0.81 (125). Incorporating E-CAM-S or similar
quantitative neurobehavioral metrics into future GBA trials would
enhance phenotypic resolution and facilitate meta-analytic
synthesis.
Accumulating evidence firmly establishes the GBA as
a critical mediator of delirium pathogenesis in perioperative and
critical care settings. The interplay between anesthesia-induced
dysbiosis, microbial metabolite signaling, and neuroimmune
activation underscores this axis as a promising therapeutic target
for mitigating acute brain dysfunction and improving long-term
patient outcomes. Future research must prioritize translational
studies bridging molecular mechanisms with clinical
interventions.
Not applicable.
XM was responsible for the conceptualization,
literature review, data curation, and the writing of the original
draft. YZ provided supervision, critical review, and editing of the
manuscript, and validated the overall content. Both authors read
and approved the final version of the manuscript. Data
authentication is not applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
Not applicable.
No funding was received.
|
1
|
Lin X, Chen Y, Zhang P, Chen G, Zhou Y and
Yu X: The potential mechanism of postoperative cognitive
dysfunction in older people. Exp Gerontol. 130:1107912020.
View Article : Google Scholar
|
|
2
|
Lopez MG, Hughes CG, DeMatteo A, O'Neal
JB, McNeil JB, Shotwell MS, Morse J, Petracek MR, Shah AS, Brown NJ
and Billings FT IV: Intraoperative oxidative damage and delirium
after cardiac surgery. Anesthesiology. 132:551–561. 2020.
View Article : Google Scholar
|
|
3
|
Wang P, Velagapudi R, Kong C, Rodriguiz
RM, Wetsel WC, Yang T, Berger M, Gelbard HA, Colton CA and Terrando
N: Neurovascular and immune mechanisms that regulate postoperative
delirium superimposed on dementia. Alzheimers Dement. 16:734–749.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gu M, Mei XL and Zhao YN: Sepsis and
Cerebral Dysfunction: BBB damage, neuroinflammation, oxidative
stress, apoptosis and autophagy as key mediators and the potential
therapeutic approaches. Neurotox Res. 39:489–503. 2021. View Article : Google Scholar
|
|
5
|
Xu X, Hu Y, Yan E, Zhan G, Liu C and Yang
C: Perioperative neurocognitive dysfunction: Thinking from the gut?
Aging (Albany NY). 12:15797–15817. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liu L, Shang L, Jin D, Wu X and Long B:
General anesthesia bullies the gut: A toxic relationship with
dysbiosis and cognitive dysfunction. Psychopharmacology (Berl).
239:709–728. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Giridharan VV, Generoso JS, Lence L,
Candiotto G, Streck E, Petronilho F, Pillai A, Sharshar T,
Dal-Pizzol F and Barichello T: A crosstalk between gut and brain in
sepsis-induced cognitive decline. J Neuroinflammation. 19:1142022.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu F and Liu J, Xiang H, Sun Z, Li Y, Li
X, Liu Y and Liu J: Dihydroartemisinin protects blood-brain barrier
permeability during sepsis by inhibiting the transcription factor
SNAI1. Clin Exp Pharmacol Physiol. 49:979–987. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Terrando N and Akassoglou K: Breaking
barriers in postoperative delirium. Br J Anaesth. 129:147–150.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yang K, Chen J, Wang T and Zhang Y:
Pathogenesis of sepsis-associated encephalopathy: More than
blood-brain barrier dysfunction. Mol Biol Rep. 49:10091–10099.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Barichello T, Giridharan VV, Catalão CHR,
Ritter C and Dal-Pizzol F: Neurochemical effects of sepsis on the
brain. Clin Sci (Lond). 137:401–414. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gao S, Jiang Y, Chen Z, Zhao X, Gu J, Wu
H, Liao Y, Sun H, Wang J and Chen W: Metabolic Reprogramming of
Microglia in Sepsis-Associated Encephalopathy: Insights from
Neuroinflammation. Curr Neuropharmacol. 21:1992–2005. 2023.
View Article : Google Scholar
|
|
13
|
Ji MH, Gao YZ, Shi CN, Wu XM and Yang JJ:
Acute and long-term cognitive impairment following sepsis:
Mechanism and prevention. Expert Rev Neurother. 23:931–943. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Krzyzaniak K, Krion R, Szymczyk A,
Stepniewska E and Sieminski M: Exploring neuroprotective agents for
sepsis-associated encephalopathy: A comprehensive review. Int J Mol
Sci. 24:107802023. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang Q, Lu C, Fan W, Zhang J and Yin Y:
Application background and mechanism of short-chain fatty acids in
sepsis-associated encephalopathy. Front Cell Infect Microbiol.
13:11371612023. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang S-h, Jia X-y, Wu Q, Jin J, Xu L-s,
Yang L, Han J-g and Zhou Q-h: The involvement of the gut microbiota
in postoperative cognitive dysfunction based on integrated
metagenomic and metabolomics analysis. Microbiol Spectr.
11:e03104232023. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang Y, Baldyga K, Dong Y, Song W,
Villanueva M, Deng H, Mueller A, Houle TT, Marcantonio ER and Xie
Z: The association between gut microbiota and postoperative
delirium in patients. Transl Psychiatry. 13:1562023. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhou X, Wu X, Wu Y, Yang L, Shi E, Ding W,
Chen L, Shi X, Feng X, Su C, et al: Indole-3-propionic acid, a gut
microbiota metabolite, protects against the development of
postoperative delirium. Ann Surg. 278:e1164–e1174. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang X, Wen X, Yuan S and Zhang J:
Gut-brain axis in the pathogenesis of sepsis-associated
encephalopathy. Neurobiol Dis. 195:1064992024. View Article : Google Scholar
|
|
20
|
Xu Y, Shen B, Pan X, Liu C, Wang Y, Chen
X, Wang T, Chen G and Chen J: Palmatine ameliorated
lipopolysaccharide-induced sepsis-associated encephalopathy mice by
regulating the microbiota-gut-brain axis. Phytomedicine.
124:1553072024. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yu Z, Shi H, Zhang J, Ma C, He C, Yang F
and Zhao L: Role of microglia in sepsis-associated encephalopathy
pathogenesis: An update. Shock. 61:498–508. 2024. View Article : Google Scholar
|
|
22
|
Abdullah IA, Khan S and Hassan FE:
Gut-Brain axis and perioperative gut microbiome in postoperative
cognitive dysfunction: Implications for neurosurgical patients. Med
Sci (Basel). 13:2362025.PubMed/NCBI
|
|
23
|
Chen P, Lin WL, Liu XY, Li SJ, Chen RF, Hu
ZH, Lin PT, Lin MH, Shi MY, Wu W, et al: D30 Alleviates
β2-Microglobulin-facilitated neurotoxic microglial responses in
isoflurane/surgery-induced cognitive dysfunction in aged mice. Lab
Invest. 105:1021902025. View Article : Google Scholar
|
|
24
|
Cheng J, Gao J, Li J and Tian H:
Neutrophils: A new target for postoperative cognitive dysfunction.
Apoptosis. 30:1117–1132. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Dai WB, Zhang X, Jiang XL, Zhang YZ, Chen
LK, Tian WT, Zhou XX, Sun XY, Huang LL, Gu XY, et al: The
kynurenine pathway regulated by intestinal innate lymphoid cells
mediates postoperative cognitive dysfunction. Mucosal Immunol.
18:53–65. 2025. View Article : Google Scholar
|
|
26
|
Gao S, Dai H, Hao Q, Song J, Ji K, Xu H,
Chen G and Lu J: Effect of perioperative probiotic intervention on
postoperative cognitive dysfunction in elderly patients: A
randomized double-blinded and placebo-controlled trial. J Transl
Med. 23:6372025. View Article : Google Scholar
|
|
27
|
He C, Shi H, Yu Z, Ma C, Jiao Z, Li J and
Yang F: The progress of the microbe-gut-brain axis in
sepsis-associated encephalopathy. Front Cell Infect Microbiol.
15:15874632025. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Horill S, Zhou XK and Jin W: Probiotics as
a possible novel therapeutic option to mitigate perioperative
neurocognitive disorders: A review exploring the latest research
findings. J Clin Anesth. 103:1118012025. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Huang ZB, Zhang GP, Lu CX, Gong C, Gao X,
Lin Y, Su P, Xu W, Lin Y, Lin N, et al: Gut microbiota-derived
3-indoleacetic acid confers a protection against sepsis-associated
encephalopathy through microglial aryl hydrocarbon receptors. Exp
Neurol. 384:1150552025. View Article : Google Scholar
|
|
30
|
Li W, Shi Q, Bai R, Zeng J, Lin L, Dai X,
Huang Q and Gong G: Advances in research on the pathogenesis and
signaling pathways associated with postoperative delirium (Review).
Mol Med Rep. 32:2202025. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liu MW, Zhang Y, Xiong GF, Zhang BR, Zhang
QJ, Gao SJ, Zhu YL and Zhang LM: Dexmedetomidine for the treatment
of sepsis-associated encephalopathy: Mechanism and prospects.
Biomed Pharmacother. 188:1182092025. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Mao L, Wang L, Huang Z, Switzer JA, Hess
DC and Zhang Q: Perioperative neurocognitive disorders: Advances in
molecular mechanisms and bioactive molecules. Ageing Res Rev.
112:1028852025. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Li H, Huang D, Qiao K, Wang Z and Xu S:
Feasibility of non-intubated anesthesia and regional block for
thoracoscopic surgery under spontaneous respiration: A prospective
cohort study. Braz J Med Biol Res. 53:e86452020. View Article : Google Scholar
|
|
34
|
Liu HY, Chiang XH, Hung MH, Wang ML, Lin
MW, Cheng YJ, Hsu HH and Chen JS: Nonintubated uniportal
thoracoscopic segmentectomy for lung cancer. J Formos Med Assoc.
119:1396–1404. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Mogahed MM and Elkahwagy MS: Paravertebral
block versus intercostal nerve block in non-intubated uniportal
video-assisted thoracoscopic surgery: A randomised controlled
trial. Heart Lung Circ. 29:800–807. 2020. View Article : Google Scholar
|
|
36
|
Jiman AA, Ratze DC, Welle EJ, Patel PR,
Richie JM, Bottorff EC, Seymour JP, Chestek CA and Bruns TM:
Multi-channel intraneural vagus nerve recordings with a novel
high-density carbon fiber microelectrode array. Sci Rep.
10:155012020. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Nguyen M, Tavernier A, Gautier T, Aho S,
Morgant MC, Bouhemad B, Guinot PG and Grober J: Glucagon-like
peptide-1 is associated with poor clinical outcome,
lipopolysaccharide translocation and inflammation in patients
undergoing cardiac surgery with cardiopulmonary bypass. Cytokine.
133:1551822020. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hu Q, Ren H, Hong Z, Wang C, Zheng T, Ren
Y, Chen K, Liu S, Wang G, Gu G, et al: Early enteral nutrition
preserves intestinal barrier function through reducing the
formation of neutrophil extracellular traps (NETs) in critically
Ill surgical patients. Oxid Med Cell Longev. 2020:88156552020.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Havel E: Danger of bacterial translocation
for a surgical patient. Rozhl Chir. 100:4–9. 2021.In English.
PubMed/NCBI
|
|
40
|
Magnan C, Lancry T, Salipante F, Trusson
R, Dunyach-Remy C, Roger C, Lefrant JY, Massanet P and Lavigne JP:
Role of gut microbiota and bacterial translocation in acute
intestinal injury and mortality in patients admitted in ICU for
septic shock. Front Cell Infect Microbiol. 13:13309002023.
View Article : Google Scholar
|
|
41
|
Huang Q, Wu Z, Chi C, Wu C, Su L, Zhang Y,
Zhu J and Liu Y: Angiopoietin-2 is an early predictor for acute
gastrointestinal injury and intestinal barrier dysfunction in
patients with acute pancreatitis. Dig Dis Sci. 66:114–120. 2021.
View Article : Google Scholar
|
|
42
|
Erikson K, Tuominen H, Vakkala M,
Liisanantti JH, Karttunen T, Syrjälä H and Ala-Kokko TI: Brain
tight junction protein expression in sepsis in an autopsy series.
Crit Care. 24:3852020. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Bigagli E, D'Ambrosio M, Cinci L, Fiorindi
C, Agostiniani S, Bruscoli E, Nannoni A, Lodovici M, Scaringi S,
Giudici F and Luceri C: Impact of preoperative immunonutrition on
oxidative stress and gut barrier function in surgical patients with
Crohn's disease. Nutrients. 15:8822023. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Hu Y, Hu XD, He ZQ, Liu Y, Gui YK, Zhu SH,
Da X, Liu YN, Liu LX, Shen QY and Xu GH: Anesthesia/surgery
activate MMP9 leading to blood-brain barrier disruption, triggering
neuroinflammation and POD-like behavior in aged mice. Int
Immunopharmacol. 135:1122902024. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Hu X, Liu L, Da X, Zhu S, Wang J, Shan M,
Liu Y, He Z and Xu G: Anesthesia/surgery leads to blood-brain
barrier disruption via the transcellular and paracellular pathways,
and postoperative delirium-like behavior: A comparative study in
mice of different ages. Exp Neurol. 383:1150442025. View Article : Google Scholar
|
|
46
|
Rodrigues AJ, Marmerstein JT, Kotamraju
BP, McCallum GA and Durand DM: Effect of anesthesia and diurnal
variation on chronic vagus nerve activity in rats. J Neurosci Res.
103:e700452025. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Baek MS, Kim S, Kim WY, Kweon MN and Huh
JW: Gut microbiota alterations in critically Ill patients with
carbapenem-resistant Enterobacteriaceae colonization: A clinical
analysis. Front Microbiol. 14:11404022023. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Hajjar R, Oliero M, Cuisiniere T, Fragoso
G, Calvé A, Djediai S, Annabi B, Richard CS and Santos MM:
Improvement of colonic healing and surgical recovery with
perioperative supplementation of inulin and
galacto-oligosaccharides. Clin Nutr. 40:3842–3851. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Xu X, Wang K, Cao X, Li Z, Zhou Y, Ren J
and Liu F: Gut microbial metabolite short-chain fatt acids
partially reverse surgery and anesthesia-induced behavior deficits
in C57BL/6J mice. Front Neurosci. 15:6646412021. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Luo A, Li S, Wang X, Xie Z, Li S and Hua
D: Cefazolin improves anesthesia and surgery-induced cognitive
impairments by modulating blood-brain barrier function, gut
bacteria and short chain fatty acids. Front Aging Neurosci.
13:7486372021. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Liu T, Wang C, Wang YY, Wang LL, Ojo O,
Feng QQ, Jiang XS and Wang XH: Effect of dietary fiber on gut
barrier function, gut microbiota, short-chain fatty acids,
inflammation, and clinical outcomes in critically ill patients: A
systematic review and meta-analysis. JPEN J Parenter Enteral Nutr.
46:997–1010. 2022. View Article : Google Scholar
|
|
52
|
Liu X, Cui J, Tan X, Yu Y, Niu J and Wang
Q: Short-chain fatty acids alleviate perioperative neurocognitive
disorders through BDNF/PI3K/Akt pathway in middle-aged rats. Mol
Neurobiol. 62:11544–11559. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Huang ZB, Hu Z, Lu CX, Luo SD, Chen Y,
Zhou ZP, Hu JJ, Zhang FL, Deng F and Liu KX: Gut microbiota-derived
indole 3-propionic acid partially activates aryl hydrocarbon
receptor to promote macrophage phagocytosis and attenuate septic
injury. Front Cell Infect Microbiol. 12:10153862022. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Kuo SZ, Dettmer K, Annavajhala MK, Chong
DH, Uhlemann AC, Abrams JA, Oefner PJ and Freedberg DE:
Associations between urinary 3-indoxyl sulfate, a gut
microbiome-derived biomarker, and patient outcomes after intensive
care unit admission. J Crit Care. 63:15–21. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Hu L, Bai Y, Lai C, Mo L, Li Y, Jiang X,
Xu W, He Y, Zhou X and Chen C: Plasma indole-3-aldehyde as a novel
biomarker of acute kidney injury after cardiac surgery: A
reanalysis using prospective metabolomic data. BMC Anesthesiol.
23:3642023. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Han S, Zheng H, Han F, Zhang X, Zhang G,
Ma S, Liu K, Qin W and Wu G: Lactobacillus johnsonii 6084
alleviated sepsis-induced organ injury by modulating gut
microbiota. Food Sci Nutr. 10:3931–3941. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Fang H, Fang M, Wang Y, Zhang H, Li J,
Chen J, Wu Q, He L, Xu J, Deng J, et al: Indole-3-propionic acid as
a potential therapeutic agent for sepsis-induced gut microbiota
disturbance. Microbiol Spectr. 10:e00125222022. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Fang H, Wang Y, Deng J, Zhang H, Wu Q, He
L, Xu J, Shao X, Ouyang X, He Z, et al: Sepsis-induced gut
dysbiosis mediates the susceptibility to sepsis-associated
encephalopathy in mice. mSystems. 7:e01399212022. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Kean IRL, Wagner J, Wijeyesekera A, De
Goffau M, Thurston S, Clark JA, White DK, Ridout J, Agrawal S,
Kayani R, et al: Profiling gut microbiota and bile acid metabolism
in critically ill children. Sci Rep. 12:104322022. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Hou L, Wang H, Yan M, Cai Y, Zheng R, Ma
Y, Tang W and Jiang W: Obeticholic acid attenuates the intestinal
barrier disruption in a rat model of short bowel syndrome. Biochim
Biophys Acta Mol Basis Dis. 1870:1672212024. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Du L and Jiang W, Zhu X, Zhu L, Fan Y and
Jiang W: Rifaximin alleviates intestinal barrier disruption and
systemic inflammation via the PXR/NFκB/MLCK pathway and modulates
intestinal Lachnospiraceae abundance in heat-stroke mice. Int
Immunopharmacol. 143(Pt 2): 1134622024. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Gong S, Yan Z, Liu Z, Niu M, Fang H, Li N,
Huang C, Li L, Chen G, Luo H, et al: Intestinal microbiota mediates
the susceptibility to polymicrobial sepsis-induced liver injury by
granisetron generation in mice. Hepatology. 69:1751–1767. 2019.
View Article : Google Scholar
|
|
63
|
Zhang H, Xu J, Wu Q, Fang H, Shao X,
Ouyang X, He Z, Deng Y and Chen C: Gut microbiota mediates the
susceptibility of mice to sepsis-associated encephalopathy by
butyric acid. J Inflamm Res. 15:2103–2119. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Kim SM, DeFazio JR, Hyoju SK, Sangani K,
Keskey R, Krezalek MA, Khodarev NN, Sangwan N, Christley S, Harris
KG, et al: Fecal microbiota transplant rescues mice from human
pathogen mediated sepsis by restoring systemic immunity. Nat
Commun. 11:23542020. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Zhang Y, Xie B, Chen X, Zhang J and Yuan
S: A key role of gut microbiota-vagus nerve/spleen axis in sleep
deprivation-mediated aggravation of systemic inflammation after LPS
administration. Life Sci. 265:1187362021. View Article : Google Scholar
|
|
66
|
Jiang XL, Gu XY, Zhou XX, Chen XM, Zhang
X, Yang YT, Qin Y, Shen L, Yu WF and Su DS: Intestinal
dysbacteriosis mediates the reference memory deficit induced by
anaesthesia/surgery in aged mice. Brain Behav Immun. 80:605–615.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Wang LK, Yang XD, Zhou D, Cheng T, Zhang X
and Wu HY: Prenatal isoflurane exposure induces developmental
neurotoxicity in rats: the role of gut microbiota. Neurotox Res.
40:485–497. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Serbanescu M, Lee S, Li F, Boppana SH,
Elebasy M, White JR and Mintz CD: Effects of perioperative exposure
on the microbiome and outcomes from an immune challenge in C57Bl/6
adult mice. Anesth Analg. 142:171–180. 2026. View Article : Google Scholar
|
|
69
|
Zhou X, Xu X, Lu D, Chen K, Wu Y, Yang X,
Xiong W, Chen X, Lan L, Li W, et al: Repeated early-life exposure
to anaesthesia and surgery causes subsequent anxiety-like behaviour
and gut microbiota dysbiosis in juvenile rats. Br J Anaesth.
130:191–201. 2023. View Article : Google Scholar
|
|
70
|
Alberda C, Marcushamer S, Hewer T,
Journault N and Kutsogiannis D: Feasibility of a lactobacillus
casei drink in the intensive care unit for prevention of antibiotic
associated diarrhea and clostridium difficile. Nutrients.
10:5392018. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Johani K, Abualsaud D, Costa DM, Hu H,
Whiteley G, Deva A and Vickery K: Characterization of microbial
community composition, antimicrobial resistance and biofilm on
intensive care surfaces. J Infect Public Health. 11:418–424. 2018.
View Article : Google Scholar
|
|
72
|
Kang M, Mischel RA, Bhave S, Komla E, Cho
A, Huang C, Dewey WL and Akbarali HI: The effect of gut microbiome
on tolerance to morphine mediated antinociception in mice. Sci Rep.
7:426582017. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Zhang L, Meng J, Ban Y, Jalodia R,
Chupikova I, Fernandez I, Brito N, Sharma U, Abreu MT, Ramakrishnan
S and Roy S: Morphine tolerance is attenuated in germfree mice and
reversed by probiotics, implicating the role of gut microbiome.
Proc Natl Acad Sci USA. 116:13523–13532. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Thomas KR, Watt J, Wu CMJ, Akinrinoye A,
Amjad S, Colvin L, Cowe R, Duncan SH, Russell WR and Forget P: Pain
and opioid-induced gut microbial dysbiosis. Biomedicines.
10:18152022. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Hofford RS, Meckel KR, Wiser EJ, Wang W,
Sens JP, Kim M, Godino A, Lam TT and Kiraly DD: Microbiome
depletion increases fentanyl self-administration and alters the
striatal proteome through short-chain fatty acids. eNeuro.
11:ENEURO.0388-23.20232024. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Lankelma JM, van Vught LA, Belzer C,
Schultz MJ, van der Poll T, de Vos WM and Wiersinga WJ: Critically
ill patients demonstrate large interpersonal variation in
intestinal microbiota dysregulation: A pilot study. Intensive Care
Med. 43:59–68. 2017. View Article : Google Scholar
|
|
77
|
Tranberg A, Thorarinsdottir HR, Holmberg
A, Schött U and Klarin B: Proton pump inhibitor medication is
associated with colonisation of gut flora in the oropharynx. Acta
Anaesthesiol Scand. 62:791–800. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Chen C, Liang H, He M, Duan R, Guan Y,
Wang F and Duan L: Impact of short-term proton pump inhibitors vs.
histamine-2 receptor antagonists on gut microbiota in patients with
acute coronary syndrome: A multicenter randomized trial. Chin Med J
(Engl). 138:542–552. 2025. View Article : Google Scholar
|
|
79
|
Yao B, Liu JY, Liu Y, Song XX, Wang SB,
Liu N, Dong ZH, Yuan ZY, Han XN and Xing JY: Sequential versus
continuous feeding and its effect on the gut microbiota in
critically ill patients: A randomized controlled trial. Clin Nutr
ESPEN. 66:245–254. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Martindale RG and Warren M: Should enteral
nutrition be started in the first week of critical illness? Curr
Opin Clin Nutr Metab Care. 18:202–206. 2015.PubMed/NCBI
|
|
81
|
Patel JJ, Rice T and Heyland DK: Safety
and outcomes of early enteral nutrition in circulatory shock. JPEN
J Parenter Enteral Nutr. 44:779–784. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Green CH, Busch RA and Patel JJ: Fiber in
the ICU: Should it be a regular part of feeding? Curr Gastroenterol
Rep. 23:142021. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Xu R, Tan C, Zhu J, Zeng X, Gao X, Wu Q,
Chen Q, Wang H, Zhou H, He Y, et al: Dysbiosis of the intestinal
microbiota in neurocritically ill patients and the risk for death.
Crit Care. 23:1952019. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Trøseid M, Holter JC, Holm K, Vestad B,
Sazonova T, Granerud BK, Dyrhol-Riise AM, Holten AR, Tonby K,
Kildal AB, et al: Gut microbiota composition during hospitalization
is associated with 60-day mortality after severe COVID-19. Crit
Care. 27:692023. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Ren M and Lotfipour S: Antibiotic
knockdown of gut bacteria sex-dependently enhances intravenous
fentanyl self-administration in adult sprague dawley rats. Int J
Mol Sci. 24:4092022. View Article : Google Scholar
|
|
86
|
Bauer M: The liver-gut-axis: Initiator and
responder to sepsis. Curr Opin Crit Care. 28:216–220.
2022.PubMed/NCBI
|
|
87
|
Ferlini L, Maenhout C, Crippa IA,
Quispe-Cornejo AA, Creteur J, Taccone FS and Gaspard N: The
association between the presence and burden of periodic discharges
and outcome in septic patients: an observational prospective study.
Crit Care. 27:1792023. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Loe ME, Khanmohammadi S, Morrissey MJ,
Landre R, Tomko SR, Guerriero RM and Ching S: Resolving and
characterizing the incidence of millihertz EEG modulation in
critically ill children. Clin Neurophysiol. 137:84–91. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Ferlini L, Nonclercq A, Su F, Creteur J,
Taccone FS and Gaspard N: Sepsis modulates cortical excitability
and alters the local and systemic hemodynamic response to seizures.
Sci Rep. 12:113362022. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Fong MWK, Norris S, Percy J, Hirsch LJ and
Herlopian A: Hemisphere-dependent ictal tachycardia versus ictal
bradycardia in a critically Ill patient. J Clin Neurophysiol.
39:e15–e18. 2022. View Article : Google Scholar
|
|
91
|
Gofton TE, Norton L, Laforge G, Gibson R,
Debicki D, Althenayan E, Scales N, Beinum AV, Hornby L, Shemie S,
et al: Cerebral cortical activity after withdrawal of
life-sustaining measures in critically ill patients. Am J
Transplant. 22:3120–3129. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Lutz R, Müller C, Dragovic S, Schneider F,
Ribbe K, Anders M, Schmid S, García PS, Schneider G, Kreuzer M and
Kratzer S: The absence of dominant alpha-oscillatory EEG activity
during emergence from delta-dominant anesthesia predicts
neurocognitive impairment-results from a prospective observational
trial. J Clin Anesth. 82:1109492022. View Article : Google Scholar
|
|
93
|
Chen Q, Liang X, Wu T, Jiang J, Jiang Y,
Zhang S, Ruan Y, Zhang H, Zhang C, Chen P, et al: Integrative
analysis of metabolomics and proteomics reveals amino acid
metabolism disorder in sepsis. J Transl Med. 20:1232022. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Cooter Wright M, Bunning T, Eleswarpu SS,
Heflin MT, McDonald SR, Lagoo-Deenadalayan S, Whitson HE,
Martinez-Camblor P, Deiner SG and Berger M: A processed
electroencephalogram-based brain anesthetic resistance index is
associated with postoperative delirium in older adults: A dual
center study. Anesth Analg. 134:149–158. 2022. View Article : Google Scholar
|
|
95
|
Pu Y, Qian F, Guo J, Sha Y and Qian Y:
Selegiline protects against lipopolysaccharide (LPS)-Induced
impairment of the blood-brain barrier through regulating the
NF-κB/MLCK/p-MLC signaling pathway. Neurotox Res. 40:267–275. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
96
|
David-Bercholz J, Acker L, Caceres AI, Wu
PY, Goenka S, Franklin NO, Rodriguiz RM, Wetsel WC, Devinney M,
Wright MC, et al: Conserved YKL-40 changes in mice and humans after
postoperative delirium. Brain Behav Immun Health. 26:1005552022.
View Article : Google Scholar : PubMed/NCBI
|
|
97
|
de Araújo BES, da Silva Fontana R, de
Magalhães-Barbosa MC, Lima-Setta F, Paravidino VB, Riveiro PM,
Pulcheri LB, Dos Santos Salú M, Genuíno-Oliveira MB, Robaina JR, et
al: Clinical features, electroencephalogram, and biomarkers in
pediatric sepsis-associated encephalopathy. Sci Rep. 12:106732022.
View Article : Google Scholar : PubMed/NCBI
|
|
98
|
de Montmollin E, Dupuis C, Jaquet P,
Sarton B, Sazio C, Susset V, Conrad M, Argaud L, Demeret S, Tadié
JM, et al: Herpes simplex virus encephalitis with initial negative
polymerase chain reaction in the cerebrospinal fluid: Prevalence,
associated factors, and clinical impact. Crit Care Med.
50:e643–e648. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Dhawan R: EEG in cardiac surgery-moving
past the obvious. J Cardiothorac Vasc Anesth. 36:3526–3528. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Ding H, Li Y, Chen S, Wen Y, Zhang S, Luo
E, Li X, Zhong W and Zeng H: Fisetin ameliorates cognitive
impairment by activating mitophagy and suppressing
neuroinflammation in rats with sepsis-associated encephalopathy.
CNS Neurosci Ther. 28:247–258. 2022. View Article : Google Scholar
|
|
101
|
Ditzel FL, Hut SC, Dijkstra-Kersten SM,
Numan T, Leijten FS, van den Boogaard M and Slooter AJ: An
automated electroencephalography algorithm to detect polymorphic
delta activity in acute encephalopathy presenting as postoperative
delirium. Psychiatry Clin Neurosci. 76:676–678. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Orobtsova M, Gorelik S, Belousova O,
Avdeeva I and Krupenkina L: Prevention of cognitive frailty in
patients of older age groups after open-heart surgery under
cardiopulmonary bypass. Arch Razi Inst. 77:1113–1123. 2022.
|
|
103
|
Pan S, Lv Z, Wang R, Shu H, Yuan S, Yu Y
and Shang Y: Sepsis-induced brain dysfunction: Pathogenesis,
diagnosis, and treatment. Oxid Med Cell Longev. 2022:13287292022.
View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Persson NDA, Uusalo P, Nedergaard M,
Lohela TJ and Lilius TO: Could dexmedetomidine be repurposed as a
glymphatic enhancer? Trends Pharmacol Sci. 43:1030–1040. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Corriero A, Gadaleta RM, Puntillo F,
Inchingolo F, Moschetta A and Brienza N: The central role of the
gut in intensive care. Crit Care. 26:3792022. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Mullish BH, Innes AJ, Roberts LA,
Anim-Burton S, Webber L, Johnson NA, Ghani R, Farshi P, Khan AB,
Kinsella F, et al: Intestinal microbiota transplant prior to
allogeneic stem cell transplant (MAST) trial: Study protocol for a
multicentre, double-blinded, placebo-controlled, phase IIa trial.
BMJ Open. 14:e0931202024. View Article : Google Scholar
|
|
107
|
Ma Y, Zhao M, Zhang C, Hao X, Yu S, Kong
Q, Liu S, Fu Y and Liu Y: A bifunctional inhibitor CYT-1
synergistically suppresses pyroptosis to improve cognitive
dysfunction in diabetic encephalopathy. Int Immunopharmacol.
162:1151272025. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Zhang LM, Xin Y, Wu ZY, Song RX, Miao HT,
Zheng WC, Li Y, Zhang DX and Zhao XC: STING mediates
neuroinflammatory response by activating NLRP3-related pyroptosis
in severe traumatic brain injury. J Neurochem. 162:444–462. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Zanza C, Romenskaya T, Thangathurai D,
Ojetti V, Saviano A, Abenavoli L, Robba C, Cammarota G, Franceschi
F, Piccioni A and Longhitano Y: Microbiome in critical care: An
unconventional and unknown ally. Curr Med Chem. 29:3179–3188. 2022.
View Article : Google Scholar
|
|
110
|
Li C, Liu L, Gao Z, Zhang J, Chen H, Ma S,
Liu A, Mo M, Wu C, Chen D, et al: Synbiotic therapy prevents
nosocomial infection in critically Ill adult patients: A systematic
review and network meta-analysis of randomized controlled trials
based on a bayesian framework. Front Med (Lausanne). 8:6931882021.
View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Sharif S, Greer A, Skorupski C, Hao Q,
Johnstone J, Dionne JC, Lau V, Manzanares W, Eltorki M, Duan E, et
al: Probiotics in critical Illness: A systematic review and
meta-analysis of randomized controlled trials. Crit Care Med.
50:1175–1186. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Lee ZY, Lew CCH, Ortiz-Reyes A, Patel JJ,
Wong YJ, Loh CTI, Martindale RG and Heyland DK: Benefits and harm
of probiotics and synbiotics in adult critically ill patients. A
systematic review and meta-analysis of randomized controlled trials
with trial sequential analysis. Clin Nutr. 42:519–531. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Innes AJ, Mullish BH, Ghani R, Szydlo RM,
Apperley JF, Olavarria E, Palanicawandar R, Kanfer EJ, Milojkovic
D, McDonald JAK, et al: Fecal microbiota transplant mitigates
adverse outcomes seen in patients colonized with
multidrug-resistant organisms undergoing allogeneic hematopoietic
cell transplantation. Front Cell Infect Microbiol. 11:6846592021.
View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Cibulková I, Řehořová V, Hajer J and Duška
F: Fecal microbial transplantation in critically Ill
patients-structured review and perspectives. Biomolecules.
11:14592021. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
He W, Jiang M, Mao P and Yan F: TLR4
inhibition ameliorates mesencephalic substantia nigra injury in
neonatal rats exposed to lipopolysaccharide via regulation of
neuro-immunity. Brain Res Bull. 165:90–96. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Ren Y, Wang Q, Yang Z, Feng L and Zhang Y:
MCC950 ameliorates the dementia symptom at the early age of line
M83 mouse and reduces hippocampal α-synuclein accumulation. Biochem
Biophys Res Commun. 611:23–30. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Cao Y, Wang Y, Li X, Yang X, Zeng B and
Guo Z: MCC950 ameliorates cognitive function by reducing white
matter microstructure damage in rats after SAH. Brain Res Bull.
202:1107432023. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Darbandi A, Banar M, Koupaei M, Afifirad
R, Asadollahi P, Bafandeh E, Rasooli I, Emamie A, Navidifar T and
Owlia P: Clinical efficacy of probiotics in prevention of
infectious diseases among hospitalized patients in ICU and non-ICU
wards in clinical randomized trials: A systematic review. Health
Sci Rep. 6:e14692023. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Ward JA, Yerke J, Lumpkin M, Kapoor A,
Lindenmeyer CC and Bass S: Evaluation of a protocol for rifaximin
discontinuation in critically ill patients with liver disease
receiving broad-spectrum antibiotic therapy. World J Hepatol.
15:1226–1236. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Sohail A, Cheema HA, Mithani MS, Shahid A,
Nawaz A, Hermis AH, Chinnam S, Nashwan AJ, Cherrez-Ojeda I, Awan RU
and Ahmad S: Probiotics for the prevention and treatment of
COVID-19: A rapid systematic review and meta-analysis. Front Nutr.
10:12741222023. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Chamani A, Mashhadi F, Khademi G, Nematy
M, Emadzadeh M, Sezavar M and Roudi F: Investigating the effect of
synbiotic supplementation on inflammatory indices in critically ill
septic children: A protocol study for randomized control trial.
Trials. 25:7122024. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
McClave SA, Omer E, Eisa M, Klosterbauer
A, Lowen CC and Martindale RG: The importance of providing dietary
fiber in medical and surgical critical care. Nutr Clin Pract.
39:546–556. 2024. View Article : Google Scholar
|
|
123
|
Ren Y, Wu K, He Y, Zhang H, Ma J, Li C,
Ruan Y, Zhang J, Wen Y, Wu X, et al: The role of NLRP3
inflammasome-mediated neuroinflammation in chronic noise-induced
impairment of learning and memory ability. Ecotoxicol Environ Saf.
286:1171832024. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Naves PVF and Caboclo LO: Independent risk
factors for seizures in critically ill patients on continuous EEG.
Epileptic Disord. 24:287–294. 2022. View Article : Google Scholar
|
|
125
|
van Sleuwen M, Sun H, Eckhardt C,
Neelagiri A, Tesh RA, Westmeijer M, Paixao L, Rajan S, Velpula
Krishnamurthy P, Sikka P, et al: Physiological assessment of
delirium severity: The electroencephalographic confusion assessment
method severity score (E-CAM-S). Crit Care Med. 50:e11–e19. 2022.
View Article : Google Scholar
|
|
126
|
Williams Roberson S, Azeez NA, Fulton JN,
Zhang KC, Lee AXT, Ye F, Pandharipande P, Brummel NE, Patel NB and
Ely EW: Quantitative EEG signatures of delirium and coma in
mechanically ventilated ICU patients. Clin Neurophysiol. 146:40–48.
2023. View Article : Google Scholar
|
|
127
|
Acker L, Wong MK, Wright MC, Reese M,
Giattino CM, Roberts KC, Au S, Colon-Emeric C, Lipsitz LA, Devinney
MJ, et al: Preoperative electroencephalographic alpha-power changes
with eyes opening are associated with postoperative attention
impairment and inattention-related delirium severity. Br J Anaesth.
132:154–163. 2024. View Article : Google Scholar
|
|
128
|
Reese M, Wright MC, Roberts KC, Browndyke
JN, Bennett M, Acker L, Devinney MJ, Reekes TH, Waligorska T, Shaw
LM, et al: Associations between anaesthetic dose-adjusted
intraoperative EEG alpha power, processing speed, and postoperative
delirium: analysis of data from three prospective studies. Br J
Anaesth. 135:109–120. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Park SK, Han DW, Chang CH, Jung H, Kang H
and Song Y: Association between intraoperative electroencephalogram
burst suppression and postoperative delirium: A systematic review
and meta-analysis. Anesthesiology. 142:107–120. 2025. View Article : Google Scholar
|
|
130
|
Fang PP, Shang ZX, Xu J, Hu J, Zhang SC,
Fan YG, Lu Y, Liu XS and Maze M: Contribution of intraoperative
electroencephalogram suppression to frailty-associated
postoperative delirium: Mediation analysis of a prospective
surgical cohort. Br J Anaesth. 130:e263–e271. 2023. View Article : Google Scholar
|
|
131
|
Baron Shahaf D and Shahaf G:
Intraoperative EEG-based monitors: Are we looking under the
lamppost? Curr Opin Anaesthesiol. 37:177–183. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Gao X, Lin C, Feng Y, You Y, Jin Z, Li M,
Zhou Y and Chen K: Akkermansia muciniphila-derived small
extracellular vesicles attenuate intestinal
ischemia-reperfusion-induced postoperative cognitive dysfunction by
suppressing microglia activation via the TLR2/4 signaling. Biochim
Biophys Acta Mol Cell Res. 1871:1196302024. View Article : Google Scholar
|
|
133
|
Battaglini D, Premraj L, Huth S, Fanning
J, Whitman G, Arora RC, Bellapart J, Bastos Porto J, Taccone FS,
Suen JY, et al: The use of noninvasive multimodal neuromonitoring
in adult critically Ill patients with COVID-19 infection. J
Neurosurg Anesthesiol. 35:423–428. 2023. View Article : Google Scholar
|
|
134
|
Eshraghi R, Yazdani MS, Bahrami A,
Amani-Beni R, Darouei B, Mokhtari M and Hashemian SM: Advanced
neuromonitoring techniques for medical and neurological ICU
patients. Brain Res Bull. 230:1115132025. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Liu LF, Hu Y, Liu YN, Shi DW, Liu C, Da X,
Zhu SH, Zhu QY, Zhang JQ and Xu GH: Reactive oxygen species
contribute to delirium-like behavior by activating CypA/MMP9
signaling and inducing blood-brain barrier impairment in aged mice
following anesthesia and surgery. Front Aging Neurosci.
14:10211292022. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Zhong L, Ren X, Ai Y and Liu Z: SS-31
improves cognitive function in sepsis-associated encephalopathy by
inhibiting the Drp1-NLRP3 inflammasome activation. Neuromolecular
Med. 25:230–241. 2023. View Article : Google Scholar
|
|
137
|
Watne LO, Pollmann CT, Neerland BE,
Quist-Paulsen E, Halaas NB, Idland AV, Hassel B, Henjum K, Knapskog
AB, Frihagen F, et al: Cerebrospinal fluid quinolinic acid is
strongly associated with delirium and mortality in hip-fracture
patients. J Clin Invest. 133:e1634722023. View Article : Google Scholar
|
|
138
|
Deschamps A, Ben Abdallah A, Jacobsohn E,
Saha T, Djaiani G, El-Gabalawy R, Overbeek C, Palermo J, Courbe A,
Cloutier I, et al: Electroencephalography-guided anesthesia and
delirium in older adults after cardiac surgery: The ENGAGES-Canada
Randomized clinical trial. JAMA. 332:112–123. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Silva BM, Alves E, Gomes AD, Marcolin
Miranda L, Albuquerque LG, Defante MLR, Miranda NCD, De Lima PEC
and Damião VP: Impact of electroencephalography-guided anesthesia
in cardiothoracic surgery: A systematic review and updated
meta-analysis of randomized controlled trials. J Cardiothorac Vasc
Anesth. 39:3154–3162. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Filipiak W, Włodarski R, Żuchowska K,
Tracewska A, Winiarek M, Daszkiewicz D, Marszałek M, Depka D and
Bogiel T: Analysis of bacterial metabolites in breath gas of
critically Ill patients for diagnosis of ventilator-associated
pneumonia-A proof of concept study. Biomolecules. 14:14802024.
View Article : Google Scholar
|
|
141
|
Li F, Jiang L, Pan S, Jiang S, Fan Y,
Jiang C, Gao C and Leng Y: Multi-omic profiling reveals that
intra-abdominal-hypertension-induced intestinal damage can be
prevented by microbiome and metabolic modulations with
5-hydroxyindoleacetic acid as a diagnostic marker. mSystems.
7:e01204212022. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Wu X, He T, He F and Liu L: Is
postoperative cognitive dysfunction a disease of microglial
inflammatory memory? A state-transition model from metabolic stress
to epigenetic lock-in. Front Mol Neurosci. 18:16481612025.
View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Lin QC, Wang J, Wang XL, Pan C, Jin SW,
Char S, Tao YX, Cao X and Li J: Hippocampal HDAC6 promotes POCD by
regulating NLRP3-induced microglia pyroptosis via HSP90/HSP70 in
aged mice. Biochim Biophys Acta Mol Basis Dis. 1870:1671372024.
View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Ditzel FL, Hut SCA, van den Boogaard M,
Boonstra M, Leijten FSS, Wils EJ, van Nesselrooij T, Kromkamp M,
Rood PJT, Röder C, et al: DeltaScan for the assessment of acute
encephalopathy and delirium in ICU and non-ICU patients, a
prospective cross-sectional multicenter validation study. Am J
Geriatr Psychiatry. 32:1093–1104. 2024. View Article : Google Scholar : PubMed/NCBI
|