Introduction
Osteoarthritis (OA), a common chronic degenerative
disease, affects >67 million people worldwide, creating a marked
socioeconomic burden (1).
Characterized by the gradual breakdown of articular cartilage and
abnormal subchondral bone remodeling, OA causes painful joint
symptoms, limited mobility and a notable decrease in the quality of
life of patients (2).
Among the cellular contributors to OA development,
osteoclasts are key mediators, regulating bone resorption by
releasing acids and proteolytic enzymes, such as cathepsin K (Ctsk)
(3). This activity not only
damages the structural stability of the subchondral bone but also
triggers cartilage destruction and joint deterioration. Previous
studies have reported that inhibiting osteoclastogenesis can
effectively alleviate symptoms of OA (4,5).
Despite these promising results, the complex molecular mechanisms
underlying osteoclast-mediated bone remodeling in OA remain
incompletely understood. This presents an obstacle to developing
targeted treatments, underscoring the urgent need for in-depth
research into the regulatory networks that control osteoclast
function in OA.
Macrophage-driven osteoclast differentiation is a
highly coordinated metabolic process characterized by extensive
mitochondrial biogenesis and metabolic reprogramming in response to
RANKL stimulation (6). Studies
have reported a marked increase in key enzymes involved in
glycolysis, the tricarboxylic acid cycle and oxidative
phosphorylation during osteoclastogenesis (7-9).
Inhibition of lactate production or pharmacological inhibition of
lactate dehydrogenase A (LDHA) suppresses osteoclast formation and
alleviates OA progression in preclinical models, suggesting lactate
may serve an important role in osteoclast differentiation (10-13). However, the regulatory mechanism
of lactate in osteoclasts is still unclear.
Epigenetic regulation controls gene expression by
altering chromatin structure, enabling cells to rapidly respond to
metabolic changes. Histone modification serves a key role in this
process. Lactylation, a post-translational modification (PTM)
mediated by lactate, is implicated in regulating cellular
functions, including macrophage polarization and immune responses
(14). However, the role of
lactylation in modulating osteoclastic differentiation and its
potential contribution to OA remains unclear.
The present study aimed to investigate whether
glycolysis and lactylation are involved in the regulation of
osteoclast differentiation and to elucidate the underlying
molecular mechanisms using in vitro and in vivo
experiments, including osteoclast differentiation assays,
measurement of lactate production, detection of histone lactylation
levels, and analyses of gene expression and epigenetic
regulation.
Materials and methods
Animal treatment
In total, 24 9-week-old male C57BL/6 mice (weight,
18-24 g) were obtained from the Animal Center of Southwest Medical
University. Mice were housed under controlled conditions
(temperature: 23±2°C; humidity: 50±10%; 12/12-h light/dark cycle)
with ad libitum access to food and water. Only male mice
were used in this study to eliminate potential confounding effects
of the estrous cycle and associated hormonal fluctuations on bone
metabolism and osteoclast activity (15), as previously described (5). Animal health and behavior were
monitored daily and body weight was recorded weekly. Humane
endpoints were as follows: i) Weight loss >20% of initial body
weight; ii) inability to access food or water; iii) signs of severe
pain or distress (prolonged immobility, hunched posture,
vocalization) unrelieved by analgesia; and iv) any condition
compromising normal murine activities. No animals reached these
endpoints prior to the scheduled experimental endpoint.
OA was induced via anterior cruciate ligament
transection (ACLT) as described previously (5). Briefly, mice were anesthetized with
isoflurane (2-3% induction, 1-2% maintenance). The right knee joint
was exposed and the ACL was transected with microscissors.
Sham-operated controls underwent identical exposure without
ligament transection.
For model validation, 8 mice underwent the bilateral
surgery, with the right knee serving as the OA model and the left
knee as the sham control. A separate cohort of 16 mice all
underwent ACLT surgery to establish the OA model. These mice were
then randomly divided into two groups (n=8/group): OA + Vehicle
group: Received daily intra-articular injections of vehicle for 2
weeks after surgery. OA + oxamic Acid group: Received daily
intra-articular injections of oxamic acid for 2 weeks after
surgery.
All 24 mice survived until the predefined endpoint.
At 4 weeks post-surgery, mice were euthanized by CO2
inhalation at a displacement rate of 50% chamber volume/min,
followed by cervical dislocation. Death was confirmed by the
absence of respiration, heartbeat and pupillary reflex.
CO2 flow was then terminated, and animals were observed
for an additional 2 min prior to tissue harvest. The Animal Care
and Use Committee of Southwest Medical University (Luzhou, China
approved all procedures (approval no. 20241007-008), which were
conducted in adherence to the ARRIVE 2.0 guidelines (16).
Micro-CT analysis
Following sacrifice, bilateral knee specimens were
dissected, thoroughly cleaned of soft tissue using pre-cooled PBS
and fixed in 4% paraformaldehyde at room temperature for 24 h.
Samples were scanned with a high-resolution micro-CT system
(SkyScan1276; Bruker Corporation) at an isotropic voxel size of 18
μm. Three-dimensional reconstruction was performed using
NRecon software (version 1.7.4.2, Bruker Corporation). The
subchondral bone region was analyzed for bone mineral density
(BMD), trabecular thickness, trabecular separation (Tb.Sp) and
bone/tissue volume ratio.
Multiplex immunohistochemistry
Knee joint specimens were fixed in 4%
paraformaldehyde at room temperature for 24 h, decalcified in 10%
EDTA for 14 days at room temperature, embedded in paraffin and
sectioned into 4 μm slices. Sections were washed with
xylene, followed by rehydration through a graded ethanol series.
Antigen retrieval was performed in citrate buffer (pH 6.0) at
95-100°C Endogenous peroxidase activity was blocked with 3%
H2O2 for 10 min, followed by blocking with 5%
BSA at room temperature (Beijing Solarbio Science & Technology
Co., Ltd.; cat. no. SW3015) in TBS for 1 h. Sections were incubated
overnight at 4°C with primary antibodies against Ctsk (1:200;
Proteintech Group, Inc.; cat. no. 11239-1-AP), H3K18la (1:500;
Inc.; cat. no. PTM-1427RM) and L-lactyl lysine (1:200; cat. no.
PTM-1401RM; both Hangzhou Jingjie Biotechnology Co., Ltd.). After
PBS washing, sections were incubated with species-specific
HRP-conjugated secondary antibodies (1:500; Wuhan Servicebio
Technology Co., Ltd.; cat. no. GB23303) for 1 h at room
temperature, followed by tyramide signal amplification using an
Opal fluorophore kit (PerkinElmer, Inc.; cat. no. FP1480). Nuclei
were counterstained with DAPI (Cell Signaling Technology, Inc.;
cat. no. 4083) for 10 min at room temperature and slides were
mounted with ProLong™ Gold Antifade reagent (Thermo Fisher
Scientific, Inc.; cat. no. P36934). Images were captured with a
fluorescence microscope (Olympus VS120 slide scanner, Olympus
Corporation) and analyzed using ImageJ software (version 1.54p,
National Institutes of Health).
Tartrate-resistant acid phosphatase
(TRAP) staining
TRAP staining was performed using a commercial kit
(Beijing Solarbio Science & Technology Co., Ltd.; cat. no.
G1492) following the manufacturer's protocol. Briefly, sections
were fixed in TRAP fixative for 1 min at room temperature, stained
in TRAP substrate solution at 37°C for 50 min and counterstained
with hematoxylin for 3 min at room temperature. TRAP-positive
osteoclasts (red-stained multinucleated cells) were counted using
ImageJ software (version 1.54p, National Institutes of Health) and
the data were presented as the number of osteoclasts/unit of bone
surface area.
Safranin O/Fast green staining
Following deparaffinization and rehydration as
aforementioned, 4 μm sections were stained with 0.1% Fast
Green for 5 min at room temperature to visualize collagen fibers,
followed by 0.1% Safranin O for 5 min at room temperature to
evaluate proteoglycan content (Beijing Solarbio Science &
Technology Co., Ltd.; cat. no. G1371). After dehydration in graded
ethanol and xylene, the sections were mounted with Permount™
(Beijing Solarbio Science & Technology Co., Ltd.; cat. no.
G8593).
Stained sections were observed under a light
microscope (Olympus DP75). The images were acquired using OLYMPUS
OlyVIA software (version 4.2, Olympus Corporation).
Hematoxylin and eosin staining
Following fixation, dehydration, and embedding as
aforementioned, the tissue was sectioned into 4 μm sections.
Tissue sections were first deparaffinized in xylene and rehydrated
through a graded ethanol series at room temperature followed by a
distilled water rinse. Nuclei were stained with Harris hematoxylin
for 3-5 min at room temperature, differentiated briefly in 0.3%
acid alcohol (1-2 sec at room temperature), and blued in running
tap water or 0.5% lithium carbonate solution for 2-3 min at room
temperature. Sections were then stained with 0.5-1% eosin for 1-3
min, dehydrated through 95 and 100% ethanol at room temperature,
cleared in xylene and finally mounted with Neutral Balsam (Beyotime
Biotechnology, cat. no. C0173) under a coverslip. The stained
sections were observed and photographed using a light microscope
(Olympus DP75).
Cell culture and treatment
RAW264.7 macrophages (China Center For Type Culture
Collection; cat. no. GDC0143) were cultured in DMEM (Gibco; Thermo
Fisher Scientific, Inc.) supplemented with 10% FBS (Gibco; Thermo
Fisher Scientific, Inc.; cat. no. A5256701) and 1%
penicillin-streptomycin and maintained at 37°C in a 5%
CO2 incubator. For osteoclastic differentiation, cells
were seeded (1x104 cells/well in 24-well plates) and
treated with recombinant murine RANKL (50 ng/ml; HUABIO, cat. no.
HA210876) for 0, 1, 3 or 5 days at 37°C. To inhibit lactate
production, cells were pre-treated for 24 h at 37°C with oxmacid
(0, 5, 10 or 20 mM; MedChemExpress; cat. no. HY-W013032A) (17,18) before RANKL stimulation.
Lactylation induction was performed by incubating cells with sodium
lactate at 37°C (0, 5, 10 or 20 mM; MedChemExpress; cat. no.
HY-W040233) for 24 h.
Cytotoxicity assay
Cell viability was assessed using the Cell Counting
Kit-8 (CCK-8; APeXBIO Technology LLC; cat. no. K1018). Cells
(1x103/well) were seeded in 96-well plates and treated
with oxmacid (0, 5, 10, or 20 mM) at 37°C for 24 h. Following
incubation, 10 μl CCK-8 solution was added to each well and
absorbance at 450 nm was measured using a microplate reader
(Synergy H1, BioTek Corporation).
Reverse transcription-quantitative
(RT-qPCR)
Total RNA was isolated from cells using the Accurate
RNA Isolation kit (Hunan Accurate Bio-Medical Technology Co., Ltd.;
cat. no. AG21023), followed by RT with the Accurate RT kit (Hunan
Accurate Bio-Medical Technology Co., Ltd.; cat. no. AG11705)
according to the manufacturer's protocols. RT-qPCR was performed on
a CFX96 Touch Real-Time PCR Detection System (Bio-Rad Laboratories,
Inc.; cat. no. 185-5196) using SYBR Green Master Mix (Hunan
Accurate Bio-Medical Technology Co., Ltd.). The thermocycling
conditions were as follows: initial denaturation at 95°C for 30
sec, followed by 40 cycles of 95°C for 5 sec and 60°C for 30 sec.
Primer sequences are listed in Table
I. Relative gene expression was calculated using the
2−ΔΔCq method (19)
and normalized to β-actin.
 | Table IPrimer sequences for reverse
transcription-quantitative PCR. |
Table I
Primer sequences for reverse
transcription-quantitative PCR.
| Gene | Sequence, 5'→3'
|
|---|
| Forward | Reverse |
|---|
| NFATc1 |
CAACGCCCTGACCACCGATAG |
GGCTGCCTTCCGTCTCATAGT |
| Ctsk |
GGGAGAAAAACCTGAAGC |
ATTCTGGGGACTCAGAGC |
| Acp5 |
TGTGGCCATCTTTATGCT |
GTCATTTCTTTGGGGCTT |
| Fosl2 |
GTCACTCCGGGCACCTCGAAC |
TTGGTCCCCGCTGCTACTGCT |
| β-actin |
CTCCATCCTGGCCTCGCTGT |
GCTGTCACCTTCACCGTTCC |
Western blot analysis
Cells were lysed on ice using RIPA buffer (Nanjing
KeyGen Biotech Co.; cat. no. KGB5203-100) supplemented with
protease and phosphatase inhibitors. Protein concentration was
determined using a BCA assay. Equal amounts of protein (30
μg/lane) were separated by SDS-PAGE using either 10 or 12.5%
polyacrylamide gels and transferred to PVDF membranes. Membranes
were blocked with 5% non-fat milk in TBST containing 0.1% Tween-20
for 1 h at room temperature, then incubated overnight at 4°C with
primary antibodies against L-lactyl lysine (1:2,000; PTM BioLab,
Inc.; cat. no. PTM-1427RM), histone H3 (1:2,000; HUABIO; cat. no.
ET1701-64), H3K18la (all 1:2,000; PTM BioLab, Inc; cat. no.
PTM-1401RM) and β-actin (1:10,000; HUABIO; cat. no. HA722023).
After washing, the membranes were incubated with HRP-conjugated
secondary antibodies (1:5,000; goat anti-rabbit IgG, HUABIO; cat.
no. HA1012) for 1 h at room temperature. Protein bands were
visualized using enhanced chemiluminescence (Affinity Biosciences;
cat. no. KF8003) and images were captured using an iBright FL1500
Imaging System (Invitrogen; Thermo Fisher Scientific, Inc.; cat.
no. A44241). Densitometric analysis was performed using ImageJ
software (version 1.54f, National Institutes of Health). β-actin
served as the loading control for non-histone proteins as it is a
stable housekeeping protein with consistent expression across all
experimental conditions. Total histone H3 was used for histone
modifications (L-lactyl lysine, (cat. no. PTM-1427RM) and H3K18la
(both Hangzhou Jingjie Biotechnology Co., Ltd.; cat. no.
PTM-1401RM)) to ensure changes reflect actual levels rather than
variations in total histone loading.
Immunofluorescence staining
Cells were fixed with 4% paraformaldehyde for 15 min
at room temperature, permeabilized with 0.5% Triton X-100 (Beijing
Solarbio Science & Technology Co., Ltd.; cat. no. IT9100) for
10 min and blocked with 5% BSA (Beijing Solarbio Science &
Technology Co., Ltd.; cat. no. SW3015) in PBS for 1 h at room
temperature. The cells were then incubated overnight at 4°C with
L-lactyl lysine and H3K18la (both 1:200) followed by a 1 h
incubation at room temperature with Alexa Fluor-conjugated
secondary antibodies (1:500; Abcam, cat. no. ab150084). F-actin was
stained using Phalloidin-iFluor 488 (Abcam; cat. no. ab176753), and
nuclei were counterstained with DAPI for 10 min at room
temperature. Coverslips were mounted on glass slides using ProLong
Gold Antifade reagent (Thermo Fisher Scientific, Inc.; cat. no.
P36934). Images were acquired using a confocal microscope (Nikon A1
HD25, Nikon Corporation) and analyzed using ImageJ software
(version 1.53t; National Institutes of Health).
Lactate and TRAP assays
Intracellular lactate levels were quantified using a
Lactate Assay kit (Beijing Solarbio Science & Technology Co.,
Ltd.; cat. no. BC2235) according to the manufacturer's
instructions. Briefly, cells were lysed and the supernatant was
mixed with lactate detection reagent. Absorbance at 570 nm was
measured to determine lactate production. TRAP activity was
evaluated using a colorimetric kit (Beijing Solarbio Science &
Technology Co., Ltd.; cat. no. BC5405) and absorbance was measured
at 405 nm to assess osteoclast differentiation.
Cleavage under targets and tagmentation
(CUT&Tag) library construction and sequencing
The CUT&Tag assay was performed using the
Hyperactive Universal CUT&Tag Assay kit (Vazyme Biotech, cat.
no. TD903-01) with modifications. Concanavalin A-coated magnetic
beads (1x106 cells) were used to bind cells, followed by
permeabilization with digitonin at 4°C for 15 min. Cells were then
incubated with anti-H3K18la (1:100) overnight at 4°C. Protein A-Tn5
transposase (100 nM) was then added, and PCR amplification was
conducted with Taq Plus Master Mix (Vazyme, cat. no. P218-01) as
follows: initial denaturation at 95°C for 3 min, 18 cycles of 95°C
for 30 sec, 58°C for 30 sec, 72°C for 30 sec and final extension at
72°C for 5 min. The libraries were size-selected (200-600 bp) with
Ampure XP beads (Beckman Coulter, cat. no. A63880) and quantified
by qPCR using SYBR Green qPCR Master Mix (Vazyme, cat. no. Q711).
The qPCR reaction contained universal primers targeting the
Illumina adapter sequences as follows: forward, 5'-AAT GAT ACG GCG
ACC ACC GAG ATC TAC AC-3') and reverse, 5'-CAA GCA GAA GAC GGC ATA
CGA GAT-3'). The thermocycling conditions were: 95°C for 3 min,
followed by 40 cycles of 95°C for 10 sec and 60°C for 30 sec. For
absolute quantification, a standard curve was generated using a
serially diluted reference library of known concentration. The
concentration of each sample library was determined by comparing
its Cq value to the standard curve.
CUT&Tag data analysis
Raw reads were processed using fastp (v0.23.2;
Shenzhen HaploX Biotechnology Co., Ltd.) for quality trimming and
adapter removal. Clean reads were aligned to the mm10 mouse genome
using Bowtie2 (v2.4.5; Johns Hopkins University). Peaks were
identified using Sparse Enrichment Analysis for CUT&RUN (v1.1;
Epicypher, Inc.) at a false-discovery rate <0.05 and visualized
in Integrative Genomics Viewer (v 2.19.7; igv.org). Motif
analysis was performed using Multiple Em for Motif Elicitation
(version 5.5.3) and Discriminative Regular Expression Motif
Elicitation, version 5.5.3; both from the MEME Suite). Differential
peak analysis was performed using Manorm (v1.1.0;
manorm.readthedocs. io/en/latest/index.html), applying a
significance threshold of P<0.05 and |log2 fold-change
(FC)|>1.
RNA sequencing (RNA-seq) and differential
gene expression analysis
Total RNA was extracted using TRIzol (Thermo Fisher
Scientific, Inc.). RNA integrity was verified on an Agilent 2100
Bioanalyzer (Agilent) and samples with RIN ≥7.0 were used.
Libraries were prepared using the VAHTS Universal V10 RNA-seq kit
(Vazyme; cat. no. NR606). The final library concentration was 10
nM. Libraries were sequenced on an Illumina NovaSeq 6000 platform
using the NovaSeq 6000 S4 Reagent kit (300 cycles) (Illumina, Inc.;
cat. no. 20028312), generating approximately 24 million 150-bp
paired-end reads per sample.
Raw reads were processed using fastp and aligned to
the mm10 genome using HISAT2 (v2.2.1; Johns Hopkins University).
Gene expression was calculated by StringTie (v2.2.1; ccb.jhu.edu/software/stringtie/).
Differential expression analysis was performed using DESeq2
(v1.38.3; Bioconductor). Differentially expressed genes were
defined as P<0.01 and |log2FC| > 1. Principal
component analysis (PCA), volcano plots, WikiPathways, Gene
Ontology (GO; geneontology. org/) and KEGG pathway (https://www.kegg.jp/) enrichment analyses were
performed using R (v4.2.1). The WikiPathways database was accessed
at https://wikipathways.org. Pathways with
adjusted P < 0.05 were considered significantly enriched.
Chromatin immunoprecipitation
(ChIP)-qPCR
ChIP assays were performed using the BeyoChIP™
Enzymatic ChIP Assay kit (Beyotime Biotechnology; cat. no. P2083S)
according to the manufacturer's protocol. Briefly, chromatin from ~
4x106 cells was cross-linked with 1% formaldehyde, lysed
in 100 μl of ChIP buffer, and sonicated to generate
fragments of 200-1,000 bp. For each IP reaction, the lysate was
incubated overnight at 4°C with 4.8 μg of anti-H3K18la
antibody (6 μg per 5x106 cells) or normal rabbit
IgG (Abcam; cat. no. ab133470) as a control. Immune complexes were
isolated by incubation with Protein A/G Magnetic Beads. Beads were
washed sequentially with low-salt, high-salt, LiCl wash buffers,
and TE buffer with centrifugation at 4°C, 12,000 × g for 1 min
between each wash. Following elution and reversal of cross-links,
the co-precipitated DNA was purified. Target DNA enrichment was
quantified by qPCR using primers specific to the Acp5 promoter
region (Table II), as
aforementioned. Enrichment was calculated relative to input DNA and
normalized to the IgG control.
| Table IIPrimer sequences for chromatin
immunoprecipitation-quantitative PCR. |
Table II
Primer sequences for chromatin
immunoprecipitation-quantitative PCR.
| Primer | Amplified region
(relative to the mouse Acp5 gene TSS) | Sequence, 5'→3'
|
|---|
| Forward | Reverse |
|---|
| Acp5-1 | -1,999 to
-1,415 |
CCGCGAGACCCTACACTTAC |
ACCATCGTTCGGGTAGTTGG |
| Acp5-2 | -1,491 to -901 |
TTGGCTGCCTCTCTCTCTCT |
AGTTAGCGTCTCCTGGGTGA |
| Acp5-3 | -624 to -73 |
TGGCAACAGGAACACGCTTA |
GATGGGAGGGGATGCAAACA |
Statistical analysis
All experiments were independently repeated at least
three times and the data are presented as the mean ± standard
deviation. Normality was assessed using the Shapiro-Wilk test, and
homoscedasticity was verified using Levene's test. For data that
followed a normal distribution and had equal variance, comparisons
between two groups used the unpaired Student's t-test. One-way
ANOVA with the Bonferroni post hoc correction was used for >2
group comparisons. Non-normally distributed data were analyzed
using the Kruskal-Wallis test followed by Dunn's multiple
comparisons test. P<0.05 was considered to indicate a
statistically significant difference. All analyses were performed
using GraphPad Prism 9.0 (Dotmatics).
Results
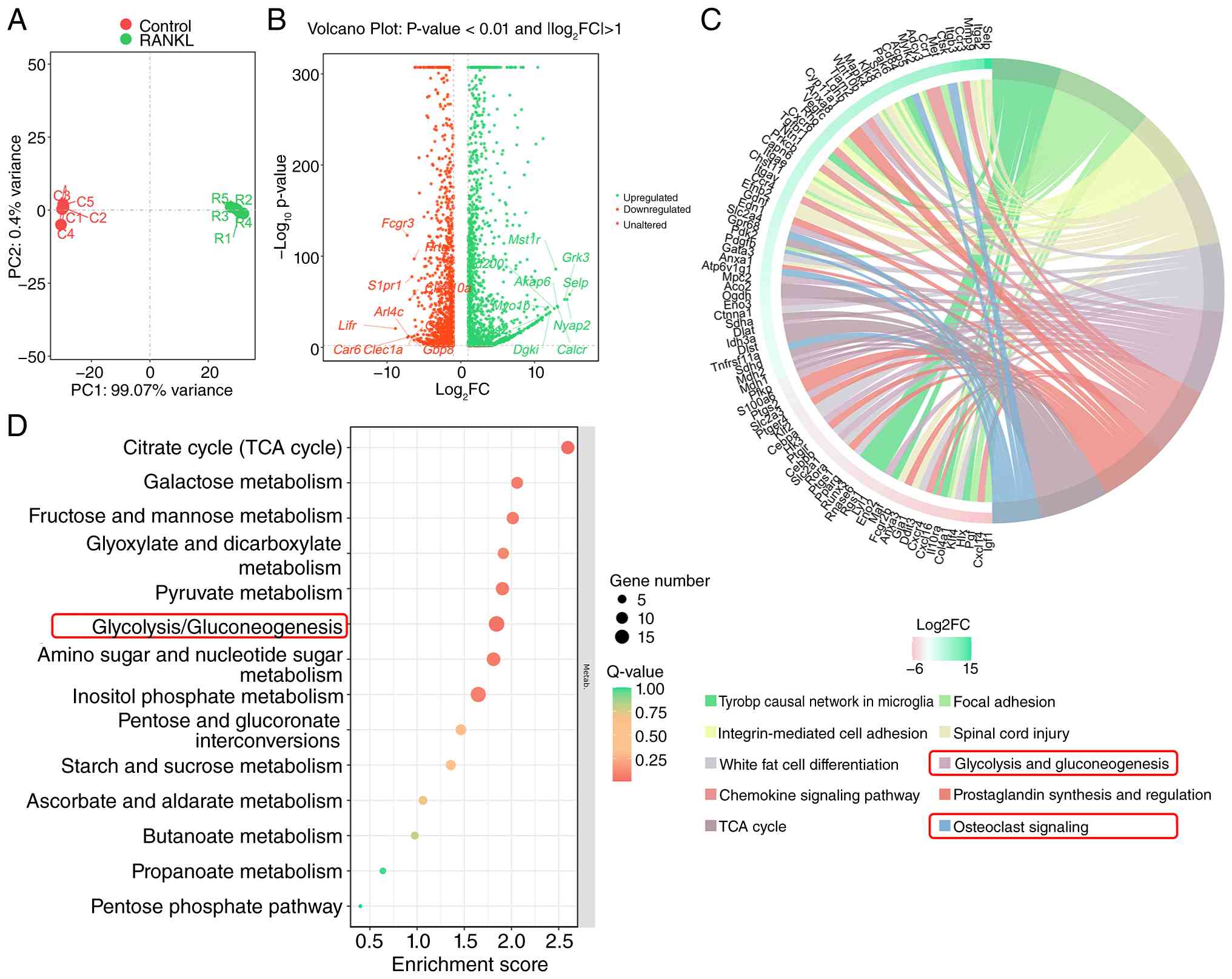
Osteoclast differentiation is accompanied
by changes in carbohydrate metabolism, including the glycolysis
process
To assess metabolic reprogramming during osteoclast
differentiation, RNA-seq was performed on RAW264.7 macrophages with
or without RANKL stimulation. PCA demonstrated clear separation
between the control and RANKL-treated groups, indicating
substantial transcriptomic changes during osteoclastogenesis
(Fig. 1A). Differential
expression analysis identified 1,496 up- and 1,321 downregulated
genes (Fig. 1B).
To explore the functional implications of these
transcriptional changes, KEGG pathway enrichment analysis was
performed. Notably, 'glycolysis/gluconeogenesis' was among the most
significantly enriched pathways, alongside 'osteoclast signaling'
(Fig. 1C and D). A chord diagram
illustrated the direct association between osteoclast-associated
and glycolysis pathway genes (Fig.
1C). These findings suggested that glycolytic metabolism is
coupled with osteoclast differentiation, providing the rationale
for investigating lactate, the end product of glycolysis, in this
process.
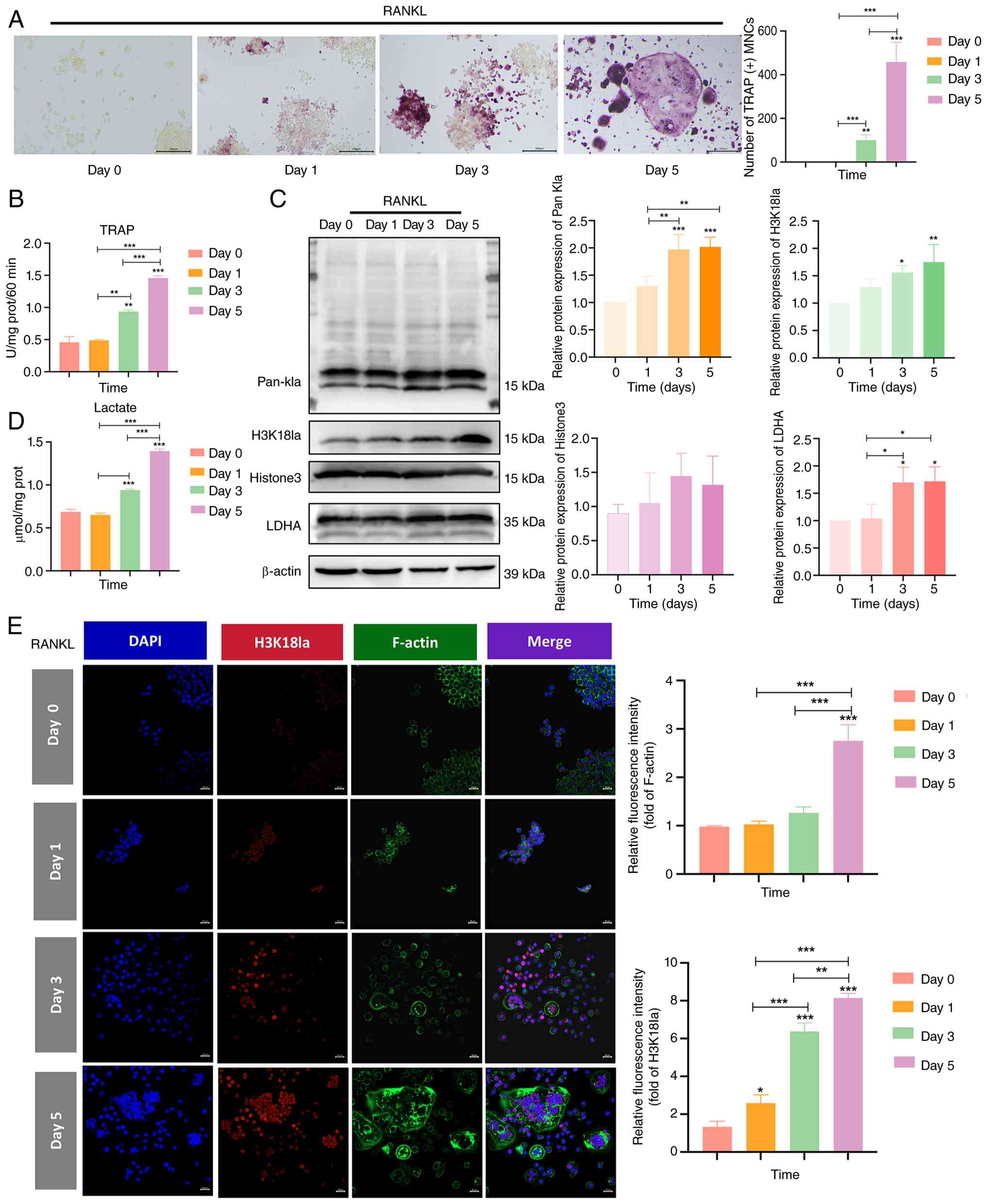
H3K18la increases during osteoclastic
differentiation
Given the enrichment of 'glycolysis/gluconeogenesis'
during osteoclastogenesis, the present study evaluated whether
lactate accumulation leads to histone lactylation. RAW264.7
macrophages were treated with RANKL for 0, 1, 3 or 5 days. TRAP
staining revealed a progressive increase in osteoclast formation,
with mature multinucleated cells appearing by day 5 (Fig. 2A and B). Concurrently,
intracellular lactate levels increased in a time-dependent manner
(Fig. 2D), suggesting enhanced
glycolytic flux during differentiation.
Protein lactylation levels were assessed using
western blotting. Pan-Kla and H3K18la signals gradually increased
from day 1 to 5, paralleling the kinetics of lactate production and
osteoclast maturation (Fig. 2C).
LDHA, a key glycolytic enzyme, also demonstrated elevated
expression. Total H3 levels remained stable across all time points,
indicating that the increased H3K18la signal reflected enhanced
lactylation rather than changes in histone abundance.
Immunofluorescence staining revealed the nuclear accumulation of
H3K18la in mature osteoclasts, with notable co-localization with
F-actin rings (Fig. 2E). These
findings suggested that H3K18la is dynamically upregulated during
osteoclast differentiation in a lactate-dependent manner.
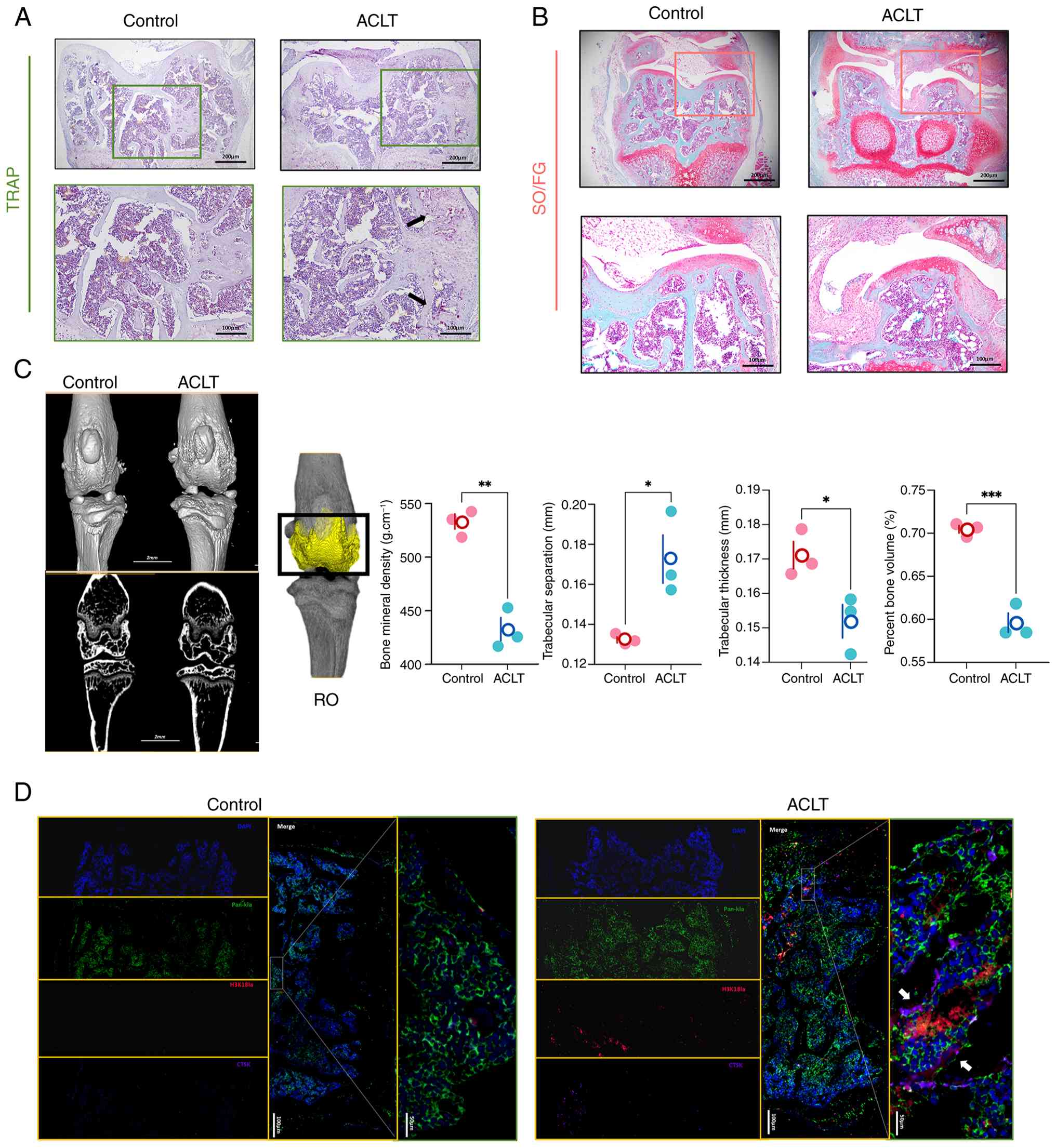
H3K18la is elevated in osteoclasts of OA
model mice
To determine whether H3K18la levels are altered
during OA pathogenesis in vivo, the present study
established an ACLT-induced OA mouse model. Micro-CT analysis at 2
weeks post-surgery revealed notable subchondral bone loss in ACLT
mice compared with that in sham-operated controls, as demonstrated
by decreased BMD and increased Tb.Sp (Fig. 3A-C). Safranin O/Fast Green
staining revealed cartilage degeneration and subchondral bone
erosion in ACLT mice, while TRAP staining confirmed increased
osteoclast numbers at the bone surface (Fig. 3A and B).
Multiplex immunofluorescence staining revealed that
H3K18la signal was markedly elevated in TRAP-positive osteoclasts
of ACLT mice, with a 2.3-fold increase compared with that in sham
controls (Fig. 3D). Pan-Kla and
Ctsk, an osteoclast marker, also revealed increased expression and
co-localization. These findings indicated that H3K18la is
specifically upregulated in osteoclasts under OA pathological
conditions, suggesting its involvement in disease-associated bone
resorption.
Inhibition of H3K18la suppresses
osteoclastic differentiation
To establish the association between lactate
production, H3K18la and osteoclastogenesis, RAW264.7 macrophages
were treated with oxmacid, a glycolysis inhibitor that blocks
lactate production (Fig. 4A).
The CCK-8 assay demonstrated that oxmacid at concentrations <10
mM had no notable cytotoxicity, while 10 and 20 mM significantly
decreased cell viability compared with 0 mM (Fig. 4B).
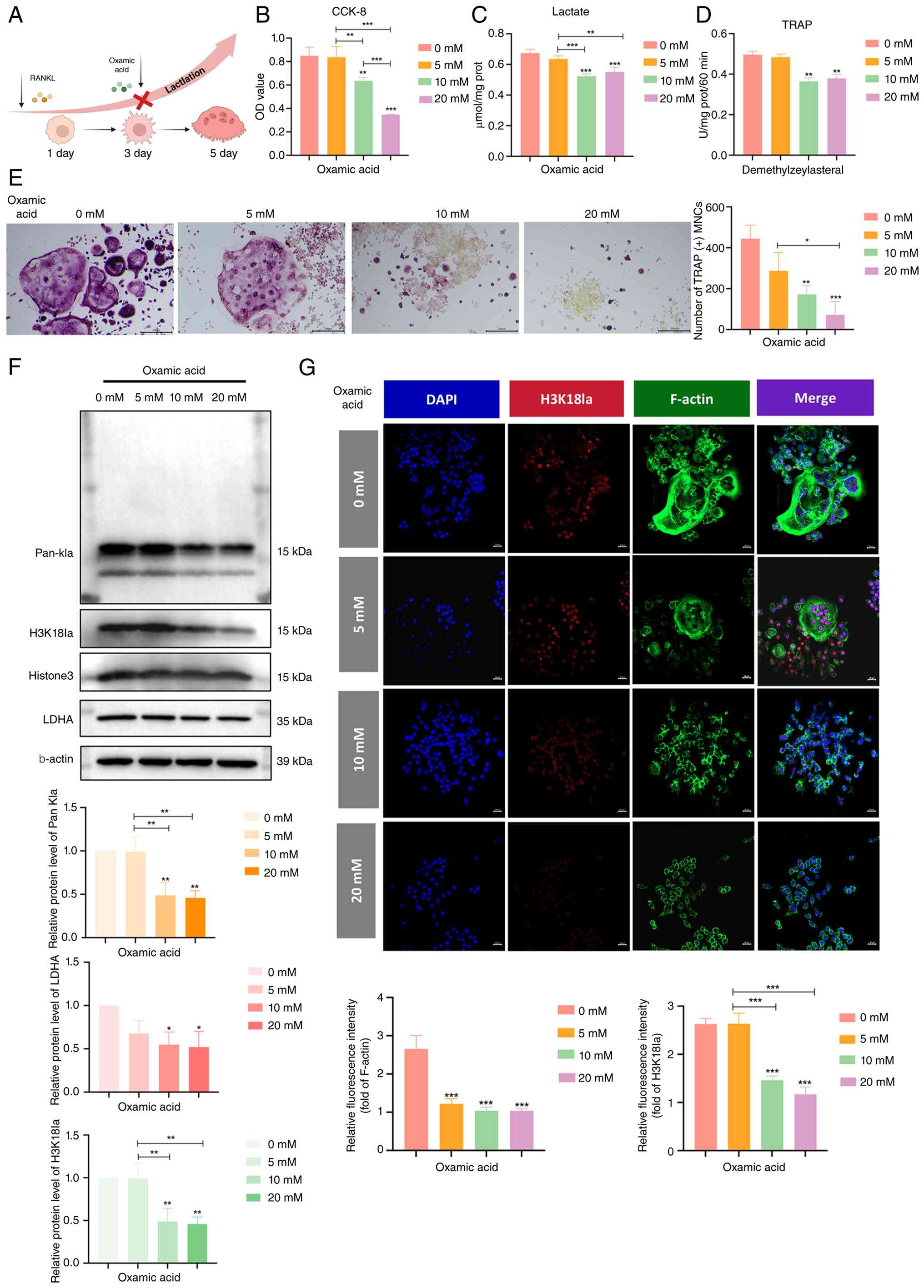 | Figure 4Inhibition of H3K18la suppresses
osteoclastic differentiation. (A) Experimental inhibition of
macrophage osteoclast differentiation. (B) Toxicity, (C) lactate
levels and (D) TRAP-positive staining of macrophages treated with
lactate inhibitors (0, 5, 10 and 20 mM). (E) TRAP staining, (F)
relative protein expression of Pan-Kla, H3K18la and LDHA and (G)
immunofluorescence staining of H3K18la and F-actin protein
following treatment with lactate inhibitors (0, 5, 10 and 20 mM).
n=3. *P<0.05; **P<0.01;
***P<0.001 vs. 0 mM. H3K18la, histone H3 lactylation
at lysine 18; TRAP, tartrate-resistant acid phosphatase; Ctsk,
Cathepsin K; LDHA, lactate dehydrogenase A; prot, protein; CCK-8,
Cell Counting Kit-8; OD, optical density; Pan-Kla, Pan-lysine
lactylation; MNC, multinucleated cells. |
Oxmacid treatment dose-dependently decreased
intracellular lactate levels (Fig.
4C) and suppressed TRAP-positive osteoclast formation (Fig. 4D and E). Moreover, western blot
analysis revealed that oxmacid decreased Pan-Kla and H3K18la levels
in a dose-dependent manner, while total H3 remained unchanged
(Fig. 4F). LDHA expression was
also reduced, consistent with glycolysis inhibition. Therefore,
subsequent experiments used 10 mM oxmacid. Furthermore,
immunofluorescence staining demonstrated decreased H3K18la nuclear
signals in the oxmacid-treated cells (Fig. 4G). These results indicated that
blocking lactate production attenuated H3K18la and inhibited
osteoclast differentiation, supporting a functional role for
H3K18la in this process.
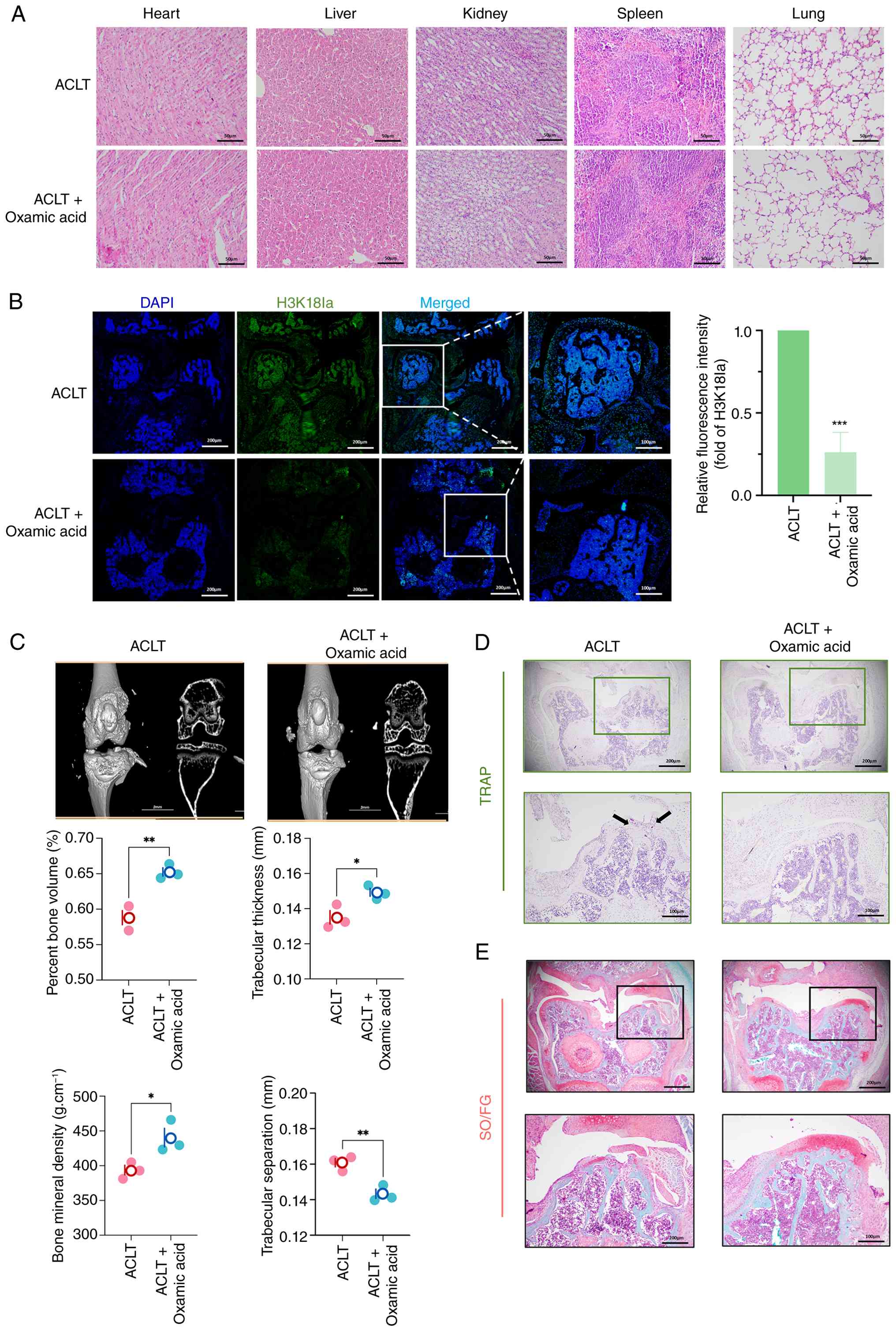
Inhibition of histone lactylation
alleviates OA pathology
Based on the aforementioned in vitro
findings, the present study evaluated the therapeutic potential of
targeting lactylation in vivo. ACLT-operated mice received
intra-articular injections of oxmacid or vehicle for 2 weeks.
Hematoxylin and eosin staining of major organs showed no obvious
toxicity, indicating that local administration was well-tolerated
(Fig. 5A).
Immunofluorescence staining revealed that oxmacid
treatment significantly reduced H3K18la levels in knee joints
compared with the vehicle controls (Fig. 5B). Micro-CT analysis demonstrated
that oxmacid-treated mice exhibited preserved subchondral bone
architecture, with higher BMD and decreased bone resorption
compared with that in vehicle-treated ACLT mice (Fig. 5C). Moreover, TRAP staining
revealed fewer osteoclasts in oxmacid-treated joints (Fig. 5D) and Safranin O/Fast Green
staining showed attenuated cartilage degeneration compared with
ACLT group (Fig. 5E). These data
indicate that pharmacological inhibition of lactate production
alleviated OA pathology by suppressing H3K18la-mediated osteoclast
activity.
Identification of H3K18la targets during
osteoclastic differentiation
To identify direct transcriptional targets of
H3K18la, the present study performed CUT&Tag sequencing using
H3K18la-specific antibodies in RAW264.7 macrophages in the presence
or absence of RANKL stimulation. Compared with the control, H3K18la
peaks were notably enriched near transcription start sites (TSS) in
RANKL-treated cells, consistent with its role in transcriptional
regulation (Fig. 6A). Moreover,
the distribution of peaks indicated predominant localization in
promoter regions (Fig. 6C).
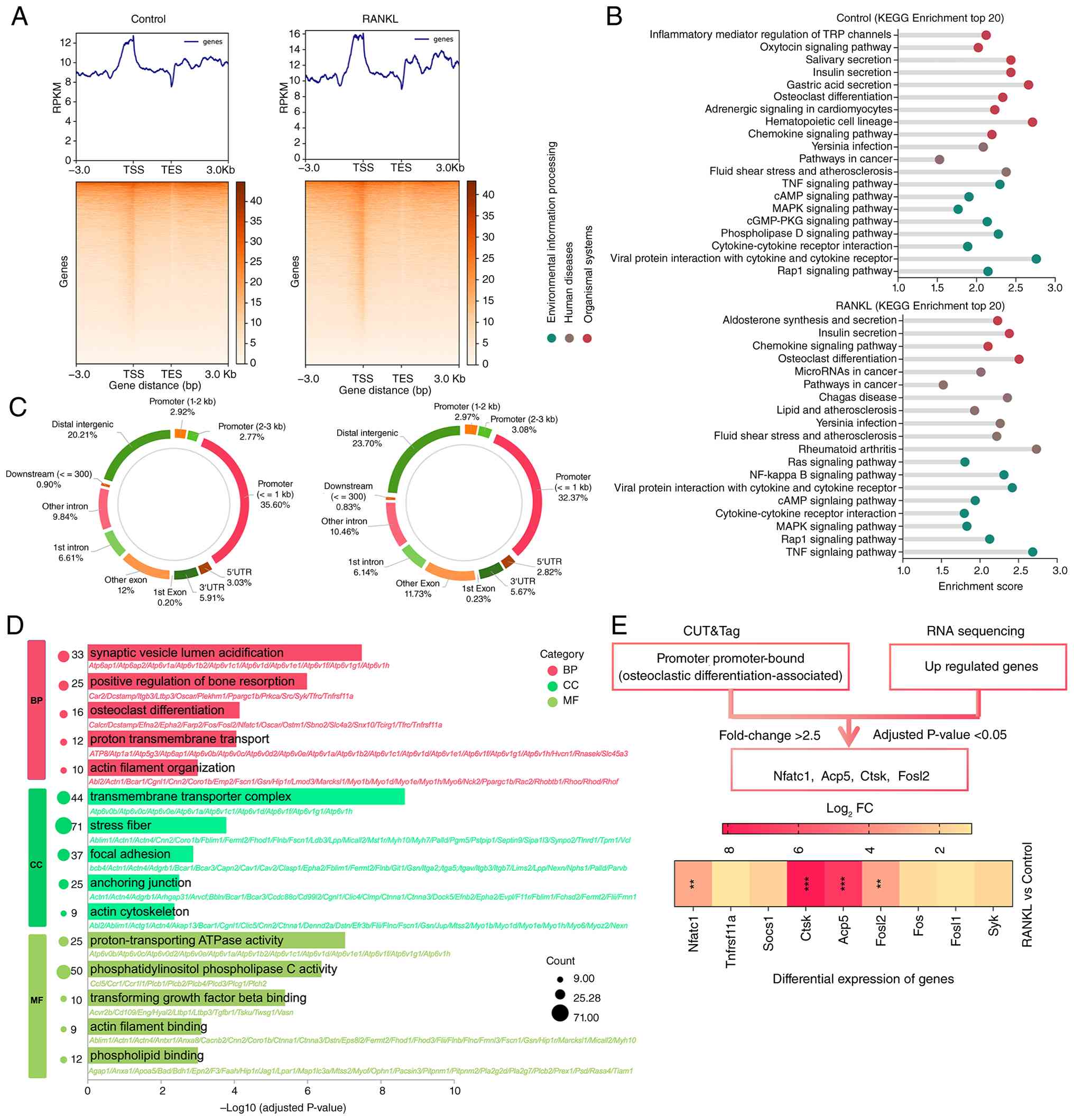 | Figure 6Identification of H3K18la targets
during osteoclastic differentiation. (A) Binding density of H3K18la
in macrophages with and without RANKL treatment. (B) KEGG
enrichment analysis of genes with differential H3K18la binding
(increased in RANKL-treated vs. control) (n=1). (C) Genome-wide
distribution of H3K18la-binding peaks. (D) Gene Ontology enrichment
analysis of the differentially expressed genes (P<0.01 and
|log2FC| >1) between the two groups with RNA-seq
(P<0.05; n=5). (E) CUT&Tag and scRNA-seq analyses identified
upregulated targets of H3K18la (P<0.05, |log2FC|
>2.5). **P<0.01; ***P<0.001.
H3K18la, histone H3 lactylation at lysine 18; KEGG, Kyoto
Encyclopedia of Genes and Genomes; FC, fold-change; scRNA-seq,
single cell RNA sequencing; BP, biological process; CC, cellular
component; MF, molecular function; Nfatc1, nuclear factor of
activated T cells 1; Acp5, acid phosphatase 5; Ctsk, cathepsin K;
RPKM, reads per kilobase of transcript per million mapped reads;
Fosl2, FOS-like antigen 2. |
KEGG enrichment analysis of genes associated with
increased H3K18la binding identified 'osteoclast differentiation'
as one of the top pathways (Fig.
6B). To refine candidate targets, the CUT&Tag data were
integrated with RNA-seq data from macrophages with or without RANKL
stimulation. GO enrichment analysis also showed 'osteoclast
differentiation' as one of the top pathways. This combined analysis
identified four osteoclast-associated genes as potential direct
targets of H3K18la: Nuclear factor of activated T cells 1 (Nfatc1),
Acp5, Ctsk and FOS-like antigen 2 (Fosl2; Fig. 6D and E). Among these, Acp5
demonstrated the most robust enrichment. Therefore, the present
study focused on this gene for subsequent validation.
H3K18la activates Acp5 transcription
To evaluate Acp5 as a direct target of H3K18la,
enriched peaks were assessed at the Acp5 locus. A clear H3K18la
peak was observed at the Acp5 promoter region in the RANKL-treated
cells, while a minimal signal was detected in controls (Fig. 7A). ChIP-qPCR was performed using
three primer pairs spanning the Acp5 promoter: P1 (-1,753 to
-1,177), P2 (-1,196 to -1,042) and P3 (-285 to -51). Significant
H3K18la enrichment was detected at the P1 and P2 regions near the
TSS, confirming direct binding at these sites (Fig. 7E).
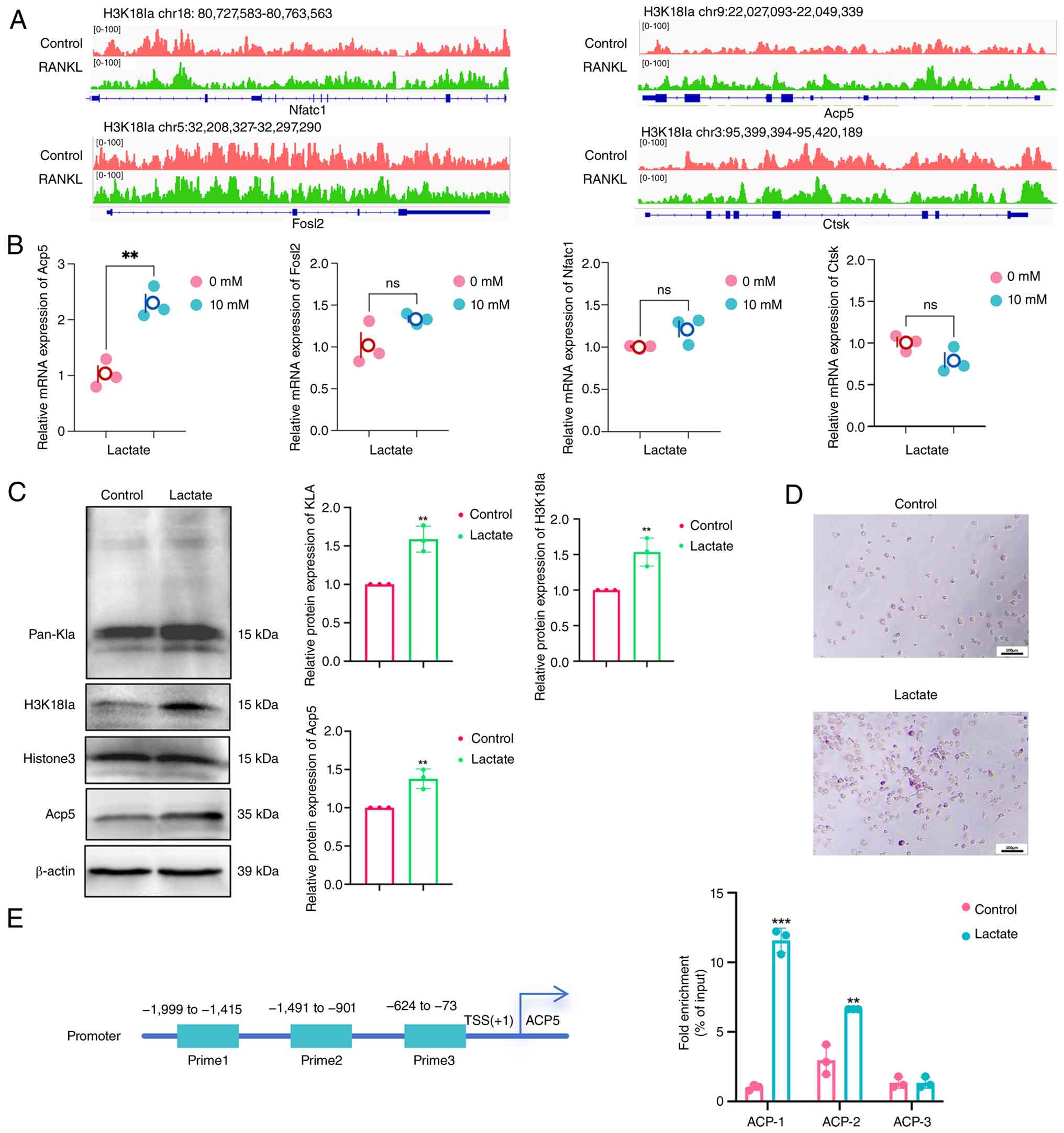 | Figure 7H3K18la activates Acp5 transcription.
(A) Genome browser tracks of the CUT&Tag signal at NFATc1,
Acp5, Ctsk and Fosl2. Relative (B) mRNA expression of Nfatc1, Acp5,
Ctsk and Fosl2 and (C) protein expression of Pan-Kla, H3K18la and
Acp5 in macrophages with or without lactate (10 mM) treatment. (D)
TRAP staining of macrophages with or without lactate (10 mM)
treatment. (E) H3K18la binding at the Acp5 promoter assessed by
ChIP-qPCR. **P<0.01; ***P<0.001 vs.
controls. H3K18la, histone H3 lactylation at lysine 18; Acp5,
tartrate-resistant acid phosphatase type 5; NFATc1, nuclear factor
of activated T cells 1; Ctsk, Cathepsin K; Fosl2, FOS-like antigen
2; TRAP, tartrate-resistant acid phosphatase; CHIP-q, chromatin
immunoprecipitation-quantitative; Pan-Kla, Pan-lysine Lactylation;
TSS, transcription start site; chr, chromosome; ns, not
significant. |
To assess the functional consequence of this
binding, macrophages were treated with exogenous lactate (10 mM) to
enhance H3K18la levels. RT-qPCR revealed that compared with
untreated controls, 10 mM lactate-treated significantly upregulated
Acp5 mRNA expression, whilst other osteoclast markers (Nfatc1, Ctsk
and Fosl2) showed no changes (Fig.
7B). Western blotting demonstrated that compared with untreated
controls, 10 mM lactate increased Acp5 protein expression and
elevated H3K18la (Fig. 7C). TRAP
staining also revealed enhanced osteoclast formation in 10 mM
lactate-treated cells, compared with untreated controls (Fig. 7D), consistent with Acp5
upregulation.
Discussion
Lactate is an important indicator of OA progression,
and the gradual increase in lactate promotes the progression of OA
by enhancing inflammation and degrading the cartilage matrix
(17). To the best of our
knowledge, however, its exact role in controlling subchondral bone
destruction, a key feature of advanced OA, has not been clearly
explored. The present study provided in vivo and in
vitro evidence indicating that pharmacological inhibition of
lactate production with oxmacid significantly decreased subchondral
bone loss in an ACLT-induced mouse model of OA. This not only
strengthens the association between lactate metabolism and OA
progression but also reveals a previously unknown epigenetic
mechanism through which lactate influences osteoclast
differentiation, offering novel insights into how metabolism
controls bone remodeling in OA.
Histone lactylation, a recently identified PTM, is a
key regulator of gene expression in several biological processes,
such as metabolic adaptation, immune response, cell death and
cytoskeletal remodeling (18,20). To the best of our knowledge, the
present study is the first to demonstrate a significant increase of
H3K18la in osteoclasts from OA joints, which was associated with
heightened osteoclast activity and faster subchondral bone
resorption. This supports the glycolytic phenotype of osteoclasts,
as inhibiting LDHA hampers osteoclastogenesis by disrupting energy
metabolism (21-24). As lactate is the primary
substrate for lactylation, it may serve as a metabolic sensor,
modifying H3K18, a lysine residue located within a histone tail
region that is highly responsive to metabolic changes (25,26). Using a comprehensive multi-omics
approach that combined CUT&Tag sequencing, ChIP-qPCR and
functional experiments, the present study identified Acp5 as a
direct downstream target of H3K18la. Acp5, a key enzyme that
hydrolyzes phosphate esters in the bone matrix, is essential for
osteoclast-driven bone resorption (27,28). The present results demonstrated a
novel metabolic-epigenetic axis whereby lactate-derived lactylation
of H3K18 activated Acp5 transcription, promoting osteoclast
differentiation and accelerating OA progression. While the present
study demonstrated the direct role for H3K18la in activating Acp5
expression, the complex interplay between histone lactylation and
other PTMs, such as ubiquitination, methylation and acetylation,
remains largely unexplored (25,29). These modifications typically work
together to regulate gene expression during cell differentiation
(30,31). H3K18la may cooperate or oppose
other PTMs to regulate osteoclastogenesis and future research using
advanced proteomic and epigenomic techniques is key to elucidate
this complex regulatory network. Translating the findings of the
present study to clinical settings requires validation in larger
animal models and human patients. Although the preclinical data
demonstrated the potential of targeting H3K18la in OA, the safety
and effectiveness of lactate-lowering or lactylation-inhibiting
drugs in humans need to be confirmed. Additionally, as lactate has
diverse roles in different cell types within the joint environment,
including chondrocytes, synovial fibroblasts and immune cells
(32-35), consideration is necessary
regarding potential off-target effects of such treatment.
From a broader perspective, the present study has
implications for understanding the metabolic reprogramming of
osteoclasts in OA. Research has highlighted the key role of cell
metabolism in regulating osteoclast differentiation and function
(8,36). For example, mitochondrial
respiration is reported to support osteoclast maturation, while
glycolysis supplies key precursors for macromolecule synthesis
during this energy-intensive process (37,38). The H3K18la-Acp5 axis adds another
layer of complexity to this metabolic regulation, indicating that
lactate serves not only as an energy source but also as a signaling
molecule to influence gene expression in osteoclasts. This may lead
to new approaches for developing targeted therapies that address
both metabolic dysregulation and epigenetic abnormalities in OA.
However, further validation should be performed in larger animal
models to avoid off-target effects caused by the pleiotropic
functions of lactate. Additionally, the results of the present
study have potential implications for other types of metabolic bone
disease characterized by abnormal osteoclast activity, including
osteoporosis and rheumatoid arthritis. Given the conserved nature
of histone lactylation across different cell types and species
(26), the H3K18la-Acp5 pathway
may also serve a role in these diseases.
However, the present study had certain limitations.
Due to the lack of specific inhibitors targeting H3K18la,
inhibiting H3K18la may impact cell activities. Furthermore, in the
CUT&Tag sequencing, there was only one sample/group, which may
compromise the reliability. Thus, it was combined with ChIP
experiments for validation to ensure the reliability of the
results. Moreover, the present study only investigated H3K18la as a
direct factor in osteoclastogenesis, without exploring other
factors involved in the overall process of H3K18la-mediated
macrophage osteoclast differentiation, such as transcription
factors, metabolic regulation and epigenetic modifications. These
aspects require further investigation in future studies.
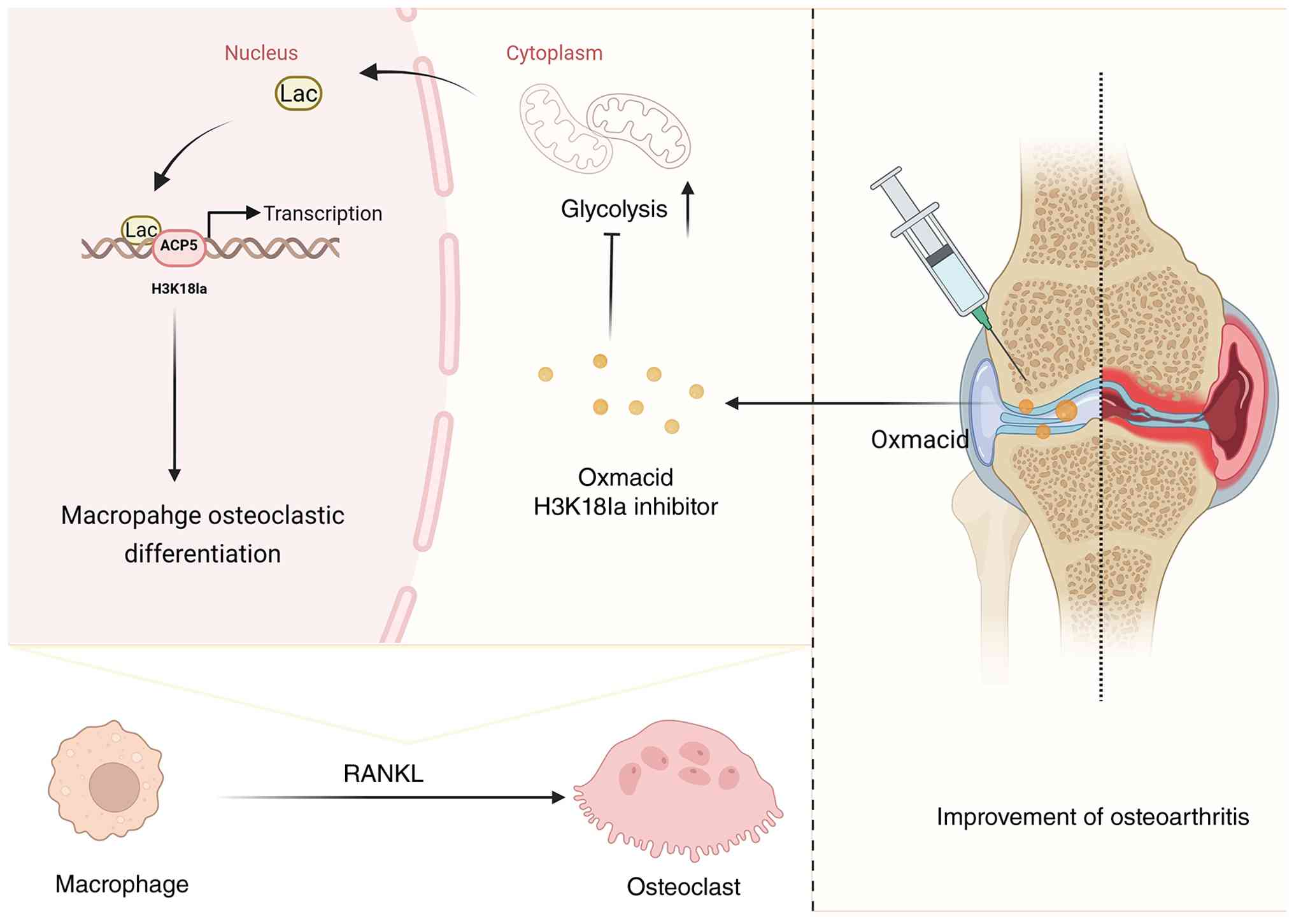
In conclusion, the present study identified H3K18la
as a crucial metabolic-epigenetic switch that connects glycolysis
to osteoclast differentiation in OA (Fig. 8). By elucidating the molecular
mechanisms underlying this new regulatory pathway, the findings
offer a conceptual framework for understanding the complex
association between metabolism, epigenetics and bone remodeling in
OA. Although there are challenges in translating these findings
into clinical practice, the present results demonstrate a promising
novel target for developing disease-modifying therapy for OA and
other metabolic bone disorders. Future research on the detailed
regulatory networks controlling H3K18la and its downstream
effectors may enhance the understanding of OA development and
support the development of more effective treatment strategies.
Availability of data and materials
The sequencing data generated in the present study
may be found in the National Center for Biotechnology Information
under accession number PRJNA1374074 or at the following URL:
ncbi.nlm.nih.gov/bioproject/?term=%20PRJNA1374074.
Authors' contributions
ZBF and SHL confirm the authenticity of all the raw
data. ZBF, SHL and XMX conceived and designed the study and wrote
the manuscript. ZBF, QYF, TS, YTZ, JWJ, SSY and YFM performed the
experiments and analyzed data. ZBF, QYF, TS, JWJ, YTZ, YFM, SHL and
XMX reviewed and edited the manuscript. All authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
The animal study was reviewed and approved by the
Committee of Medical Experimental Animal Center of Southwest
Medical University (Luzhou, China; approval no. 20241007-008).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Collaborative Project of
Luzhou Municipal People's Government and Southwest (grant no.
2024LZXNYDJ041) and the Project of 'Youth Scientific Research
Climbing Program' of the Stomatological Hospital Affiliated to
Southwest Medical University (grant no. 2024KQ05).
References
|
1
|
Zhen G, Wen C, Jia X, Li Y, Crane JL,
Mears SC, Askin FB, Frassica FJ, Chang W, Yao J, et al: Inhibition
of TGF-β signaling in mesenchymal stem cells of subchondral bone
attenuates osteoarthritis. Nat Med. 19:704–712. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Qiu L, Alhaskawi A and Moqbel SAA:
Osteoarthritis: Multitissue pathology, molecular mechanisms,
clinical management, and emerging precision and regenerative
therapies. Front Pharmacol. 16:16971922026. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jiang W, Jin Y, Zhang S, Ding Y, Huo K,
Yang J, Zhao L, Nian B, Zhong TP, Lu W, et al: PGE2 activates EP4
in subchondral bone osteoclasts to regulate osteoarthritis. Bone
Res. 10:272022. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ledesma-Colunga MG, Passin V, Lademann F,
Hofbauer LC and Rauner M: Novel insights into osteoclast energy
metabolism. Curr Osteoporos Rep. 21:660–669. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhu S, Zhu J, Zhen G, Hu Y, An S, Li Y,
Zheng Q, Chen Z, Yang Y, Wan M, et al: Subchondral bone osteoclasts
induce sensory innervation and osteoarthritis pain. J Clin
Investigat. 129:1076–1093. 2019. View Article : Google Scholar
|
|
6
|
Takegahara N, Kim H and Choi Y: Unraveling
the intricacies of osteoclast differentiation and maturation:
Insight into novel therapeutic strategies for bone-destructive
diseases. Exp Mol Med. 56:264–272. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhang D, Tang Z, Huang H, Zhou G, Cui C,
Weng Y, Liu W, Kim S, Lee S, Perez-Neut M, et al: Metabolic
regulation of gene expression by histone lactylation. Nature.
574:575–580. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Park-Min KH: Metabolic reprogramming in
osteoclasts. Semin Immunopathol. 41:565–572. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Karner CM and Long F: Glucose metabolism
in bone. Bone. 115:2–7. 2018. View Article : Google Scholar :
|
|
10
|
Xin Q, Wang H, Li Q, Liu S, Qu K, Liu C
and Zhang J: Lactylation: A passing fad or the future of
posttranslational modification. Inflammation. 45:1419–1429. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang D, Gao J, Zhu Z, Mao Q, Xu Z, Singh
PK, Rimayi CC, Moreno-Yruela C, Xu S, Li G, et al: Lysine
l-lactylation is the dominant lactylation isomer induced by
glycolysis. Nat Chem Biol. 21:91–99. 2025. View Article : Google Scholar
|
|
12
|
Wang J, Yang P, Yu T, Gao M, Liu D, Zhang
J, Lu C, Chen X, Zhang X and Liu Y: Lactylation of PKM2 suppresses
inflammatory metabolic adaptation in Pro-inflammatory macrophages.
Int J Biol Sci. 18:6210–6225. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Du S, Zhang X, Jia Y, Peng P, Kong Q,
Jiang S, Li Y, Li C, Ding Z and Liu L: Hepatocyte HSPA12A inhibits
macrophage chemotaxis and activation to attenuate liver
ischemia/reperfusion injury via suppressing glycolysis-mediated
HMGB1 lactylation and secretion of hepatocytes. Theranostics.
13:3856–3871. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang H, Wang L, Cui J, Wang S, Han Y,
Shao H, Wang C, Hu Y, Li X, Zhou Q, et al: Maintaining hypoxia
environment of subchondral bone alleviates osteoarthritis
progression. Sci Adv. 9:eabo78682023. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lorenzo J: Sexual dimorphism in
osteoclasts. Cells. 9:20862020. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Percie Du Sert N, Ahluwalia A, Alam S,
Avey MT, Baker M, Browne WJ, Clark A, Cuthill IC, Dirnagl U,
Emerson M, et al: Reporting animal research: Explanation and
elaboration for the ARRIVE guidelines 2.0. PLoS Biol.
18:e30004112020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yang J, Li W, Lin X and Liang W: A lactate
metabolism-related gene signature to diagnose osteoarthritis based
on machine learning combined with experimental validation. Aging
(Albany NY). 16:13076–13103. 2024.PubMed/NCBI
|
|
18
|
Xia J, Qiao Z, Hao X and Zhang Y:
LDHA-induced histone lactylation mediates the development of
osteoarthritis through regulating the transcription activity of
TPI1 gene. Autoimmunity. 57:23848892024. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
20
|
Huang YF, Wang G, Ding L, Bai ZR, Leng Y,
Tian JW, Zhang JZ, Li YQ, Ahmad, Qin YH, et al: Lactate-upregulated
NADPH-dependent NOX4 expression via HCAR1/PI3K pathway contributes
to ROS-induced osteoarthritis chondrocyte damage. Redox Biology.
67:1028672023. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang J, Guan H, Liu H, Lei Z, Kang H, Guo
Q, Dong Y, Liu H, Sun Y, Fang Z and Li F: Inhibition of PFKFB3
suppresses osteoclastogenesis and prevents ovariectomy-induced bone
loss. J Cell Mol Med. 24:2294–2307. 2020. View Article : Google Scholar :
|
|
22
|
Hu W, Chen Y, Dou C and Dong S:
Microenvironment in subchondral bone: Predominant regulator for the
treatment of osteoarthritis. Ann Rheum Dis. 80:413–422. 2021.
View Article : Google Scholar :
|
|
23
|
Li B, Lee W, Song C, Ye L, Abel ED and
Long F: Both aerobic glycolysis and mitochondrial respiration are
required for osteoclast differentiation. FASEB J. 34:11058–11067.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nishioku T, Anzai R, Hiramatsu S, Terazono
A, Nakao M and Moriyama M: Lactate dehydrogenase A inhibition
prevents RANKL-induced osteoclastogenesis by reducing enhanced
glycolysis. J Pharmacol Sci. 153:197–207. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Peng X and Du J: Histone and non-histone
lactylation: Molecular mechanisms, biological functions, diseases,
and therapeutic targets. Mol Biomed. 6:382025. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Galle E, Wong CW, Ghosh A, Desgeorges T,
Melrose K, Hinte LC, Castellano-Castillo D, Engl M, de Sousa JA,
Ruiz-Ojeda FJ, et al: H3K18 lactylation marks tissue-specific
active enhancers. Genome Biol. 23:2072022. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Rathod B, Desai S, Samvelyan HJ, Bock L,
Wu J, Ohlsson C, Palmquist A, Alm JJ, Newton PT, Andersson G and
Windahl SH: Tartrate-resistant acid phosphatase (TRAP/ACP5)
promotes bone length, regulates cortical and trabecular bone mass,
and maintains growth plate architecture and width in a sex- and
site-specific manner in mice. Bone. 188:1172232024. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ivanova MM, Dao J, Loynab N, Noor S,
Kasaci N, Friedman A and Goker-Alpan O: The expression and
secretion profile of TRAP5 isoforms in gaucher disease. Cells.
13:7162024. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Shen Y, Mao Z, Chen H, Zhu W, Guan Q, Yang
Y, Liu J and Li L: Exercise-specific post-translational
modification signatures: Unveiling precise regulatory mechanisms of
molecular exercise language and cellular adaptation. Front Sports
Act Living. 8:17651702026. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Balsalobre A and Drouin J: Pioneer factors
as master regulators of the epigenome and cell fate. Nat Rev Mol
Cell Biol. 23:449–464. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chisolm DA and Weinmann AS: Connections
between metabolism and epigenetics in programming cellular
differentiation. Annu Rev Immunol. 36:221–246. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zheng J, Lu Y, Lin Y, Si S, Guo B, Zhao X
and Cui L: Epitranscriptomic modifications in mesenchymal stem cell
differentiation: Advances, mechanistic insights, and beyond. Cell
Death Differ. 31:9–27. 2024. View Article : Google Scholar
|
|
33
|
Cheung KC, Ma J, Wang L, Chen X, Fanti S,
Li M, Azevedo LR, Gosselet F, Shen H, Zheng X, et al: CD31
orchestrates metabolic regulation in autophagy pathways of
rheumatoid arthritis. Pharmacol Res. 207:1073462024. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhu Z, Chen Y, Zou J, Gao S, Wu D, Li X,
Hu N, Zhao J, Huang W and Chen H: Lactate mediates the bone
anabolic effect of High-intensity interval training by inducing
osteoblast differentiation. J Bone Joint Surg Am. 105:369–379.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Pucino V, Nefla M, Gauthier V, Alsaleh G,
Clayton SA, Marshall J, Filer A, Clark AR, Raza K and Buckley CD:
Differential effect of lactate on synovial fibroblast and
macrophage effector functions. Front Immunol. 14:11838252023.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Da W, Tao L and Zhu Y: The role of
osteoclast energy metabolism in the occurrence and development of
osteoporosis. Front Endocrinol (Lausanne). 12:6753852021.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Estell E, Ichikawa T, Giffault P, Bonewald
L, Spiegelman B and Rosen C: Irisin enhances mitochondrial function
in osteoclast progenitors during differentiation. Biomedicines.
11:33112023. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chen L, Su Y, Wang C, Huang Q, Chen W, Hai
N, Wang J, Lian H, Zhao J, Xu J and Liu O: Rc3h1 negatively
regulates osteoclastogenesis by limiting energy metabolism.
Theranostics. 14:7554–7568. 2024. View Article : Google Scholar : PubMed/NCBI
|