Introduction
Osteoporosis (OP) is a prevalent metabolic bone
disorder characterised by reduced bone mass and increased fracture
susceptibility (1,2). With the aging global population, it
poses a significant public health burden worldwide (3,4).
Beyond bone loss itself, accumulating evidence suggests that OP
fundamentally arises from dysregulated cellular dynamics within the
bone marrow niche (5).
Particularly, an imbalance in bone marrow stromal cell (BMSC)
differentiation favouring adipogenesis over osteogenesis has
emerged as a central pathological feature of the disease (6,7).
In addition, inflammatory cytokines, including interleukin-1β
(IL-1β), interleukin-6 (IL-6) and tumour necrosis factor-α, are
well recognised to regulate bone remodelling by promoting
osteoclast differentiation and activity while inhibiting osteoblast
function (8,9), thereby contributing to enhanced
bone resorption and bone loss. These cytokines also participate in
extracellular matrix degradation through upregulation of
proteolytic enzymes such as matrix metalloproteinases and cathepsin
K, which are key mediators of bone matrix degradation in OP
(10,11).
Under physiological conditions, BMSCs maintain
skeletal integrity through tightly coordinated lineage commitment.
However, oestrogen deficiency, aging and chronic oxidative stress
disrupt this equilibrium, resulting in excessive accumulation of
marrow adipocytes and impaired osteoblast formation (12). Reactive oxygen species not only
induce apoptosis of osteoprogenitor cells but also promote
adipogenic differentiation, thereby exacerbating bone loss
(13). Among the key molecular
regulators governing this process, peroxisome
proliferator-activated receptor gamma (PPARγ) serves as a master
transcription factor that drives adipogenesis while simultaneously
suppressing osteoblast differentiation (14,15). Aberrant activation of PPARγ has
been consistently associated with osteoporotic phenotypes,
highlighting it as a critical therapeutic target for restoring
bone-fat balance (16-18).
In parallel with these intrinsic regulatory
mechanisms, the gut microbiota has emerged as an important
extrinsic modulator of skeletal homeostasis through the gut-bone
axis (19,20). Microbial metabolites,
particularly those derived from tryptophan metabolism, have
attracted increasing attention because of their systemic regulatory
functions (21), including the
modulation of immune responses, oxidative stress and tissue
homeostasis (22-24). Tryptophan can be metabolised by
intestinal microbes into indole and its derivatives, including
indole-3-acetic acid (IAA), indole-3-propionic acid (IPA),
indole-3-acrylic acid (IA), indole-3-lactic acid (ILA) and
indole-3-aldehyde (25). Among
these metabolites, IPA, which is predominantly produced by
Clostridium sporogenes (C. sporogenes) (26), has been reported to exert potent
antioxidative (27,28), anti-inflammatory (29) and cytoprotective effects
(30) in multiple disease
contexts (31). Recent studies
have suggested that IPA and related indole derivatives may
influence bone metabolism (32-34), primarily through inhibition of
osteoclastogenesis (35,36) and improvement of gut barrier
function (36). For example, IPA
was reported to improve skeletal quality by suppressing
P65/NLRP3-dependent osteoclast formation in diet-induced obese mice
(32). However, whether IPA
directly regulates BMSC lineage commitment and contributes to the
restoration of osteogenic-adipogenic balance remains largely
unknown.
In the present study, it was demonstrated that
microbiota-derived IPA acts as a critical regulator of bone marrow
lineage fate by suppressing PPARγ signalling. Through integrated
in vitro and in vivo analyses, it was revealed that
IPA restores bone-fat balance, enhances osteogenesis, and
alleviates oestrogen deficiency-induced OP. These findings reveal a
previously unrecognised mechanism linking gut microbial metabolism
to skeletal remodelling and highlight IPA as a promising
therapeutic candidate for OP.
Materials and methods
Reagents and antibodies
IPA (cat. no. SJ-MX4391) was purchased from Shandong
Sparkjade Scientific Instruments Co., Ltd.
H2O2 was purchased from MilliporeSigma.
Anti-GAPDH antibody was obtained from Cell Signalling Technology,
Inc. Antibodies against COL1A1 (cat. no. HA722517), RUNX2 (cat. no.
ET1612-47), SP7 (cat. no. HA722817) and OPN (cat. no. HA723082)
were obtained from HUABIO. Anti-PPARγ antibody (cat. no. 14N93N79)
was purchased from Epizyme, Inc.
Mouse BMSC isolation, culture and
osteogenic differentiation
Mouse BMSCs (mBMSCs) were isolated as previously
described (37,38). Briefly, mBMSCs were obtained from
the femora and tibiae of C57BL/6 mice by flushing the bone marrow
cavity. The C57BL/6 mice were sourced from same mice used in the
animal experiments. The isolated cells were cultured in α-MEM
(Procell Life Science & Technology Co., Ltd.) supplemented with
10% fetal bovine serum (FBS; cat. no. SA101.02; Cellmax; https://www.cellmaxcell.com/) and 1%
penicillin/streptomycin (MedChemExpress). To establish an oxidative
stress microenvironment in vitro, cells were treated with
H2O2 (100 µM) for 2 h (7). For osteogenic differentiation,
mBMSCs were cultured in osteogenic induction medium consisting of
low-glucose Dulbecco's Modified Eagle Medium (Procell Life Science
& Technology Co., Ltd.) supplemented with 10% FBS,
β-glycerophosphate (10 mM), ascorbic acid (200 µM) and
dexamethasone (100 nM) (MedChemExpress).
Alkaline phosphatase (ALP) and alizarin
Red S (ARS) staining
mBMSCs were seeded in 24-well plates and cultured in
osteogenic induction medium. At the indicated time points, ALP
staining was performed using a BCIP/NBT Alkaline Phosphatase
Chromogenic kit (cat. no. C3206; Beyotime Institute of
Biotechnology) according to the manufacturer's instructions. ARS
staining was performed using an Alizarin Red kit (cat. no.
ALIR-10001; Cyagen Biosciences, Inc.) following the manufacturer's
protocol.
Oil Red O staining
mBMSCs were seeded in 24-well plates and cultured in
adipogenic induction medium ((Procell Life Science & Technology
Co., Ltd.) according to the manufacturer's instructions. Cells were
subsequently stained with Oil Red O staining solution (Procell Life
Science & Technology Co., Ltd.).
Cell Counting Kit-8 (CCK-8) assay
To evaluate the effect of IPA on cell proliferation,
2,000 cells were seeded into 96-well plates and treated with
different concentrations of IPA (0, 10, 25, 50, 75 and 100
µM) for the indicated durations. Subsequently, 10 µl
of CCK-8 solution (Beijing Boxbio Science & Technology Co.,
Ltd.) was added to each well and incubated at 37°C for 2 h.
Absorbance at 450 nm was measured using a microplate reader
(BioTek; Agilent Technologies, Inc.).
Mitochondrial membrane potential (MMP)
assay
MMP was assessed using a JC-1 assay kit
(MedChemExpress). Briefly, cells were pretreated with
H2O2 (100 µM) and subsequently exposed
to different concentrations of IPA for 12, 24, or 48 h. After
treatment, cells were incubated with JC-1 working solution for 20
min. Fluorescence intensities were measured at 590 nm (aggregates,
red) and 520 nm (monomers, green). The ratio of red-to-green
fluorescence was calculated to evaluate changes in MMP.
Western blotting
Cells were harvested and lysed in RIPA buffer
containing protease and phosphatase inhibitors (Beijing LABLEAD
Inc.). Lysates were centrifuged at 12,000 × g for 15 min at 4°C,
and the supernatants were collected and mixed with loading buffer.
Total protein concentration was determined using a BCA protein
assay kit (cat. no. P0012; Beyotime Institute of Biotechnology).
Equal amounts of protein (20 µg per lane) were separated by
10% sodium dodecyl sulphate polyacrylamide gel electrophoresis and
transferred onto polyvinylidene fluoride membranes
(MilliporeSigma). Membranes were blocked at room temperature with
5% non-fat milk for 1 h and incubated with primary antibodies
(1:1,000) overnight at 4°C. The membranes were then incubated with
HRP-conjugated secondary antibodies (1:5,000) for 1 h. Protein
bands were visualised using ECL Luminescent Solution (cat. no.
KGC4601; Nanjing KeyGen Biotech Co., Ltd.) using a chemiluminescent
detection system (Thermo Fisher Scientific, Inc.), and band
intensities were analysed using the 'Gel Analysis' function in
ImageJ software (V1.53; National Institutes of Health) as
previously described (39).
GAPDH was used as the internal loading control.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA from BMSCs was extracted using TRIzol
reagent (Shandong Sparkjade Scientific Instruments Co., Ltd.), and
cDNA was synthesised using the SPARKscript II RT Plus Kit (cat. no.
AG0304; Shandong Sparkjade Scientific Instruments Co., Ltd.).
RT-qPCR was performed using SYBR Green Master Mix (cat. no. AH0104;
Shandong Sparkjade Scientific Instruments Co., Ltd.). Thermal
cycling conditions were set according to the manufacturer's
instructions. The mRNA levels of the target genes were normalized
to those of the housekeeping gene GAPDH. The relative gene
expression levels were calculated using the 2−ΔΔCq
method. Primer sequences used in the present study are listed in
Table I.
 | Table ISequences of primers used for reverse
transcription-quantitative PCR. |
Table I
Sequences of primers used for reverse
transcription-quantitative PCR.
| Gene name | Primer sequence
(5′-3′) |
|---|
| Col1a1 | F:
GCCGCAAAGAGTCTACATGT |
| R:
CTTCTTGGCCATGCGTCAG |
| Runx2 | F:
CACCTCGAATGGCAGCACGCTA |
| R:
GCCGCCAAACAGACTCATCCA |
| Sp7 | F:
CCTAAGGGGCACAGCTCGTCT |
| R:
TGCATGTCCCACCAAGGAGTAGG |
| Ocn | F:
CAGTATGGCTTGAAGACCGC |
| R:
GACATCCATACTTGCAGGGC |
| Adiponectin | F:
CTGCAACATTCCGGGACTCT |
| R:
TGAAGAGAACGGCCTTGTCC |
| Srebp-1 | F:
ACAGGAGGACATCTTGCTGC |
| R:
AGATCTCTGCCAGTGTTGCC |
| CEBPα | F:
GACATCAGCGCCTACATCGA |
| R:
CCCGGGTAGTCAAAGTCACC |
| Fabp2 | F:
GCCTGGACCATTGAGGGAAA |
| R:
GCTTGGCCTCAACTCCTTCA |
| PPARγ | F:
AAGCCGTGCAAGAGATCACA |
| R:
TGGTCATGAATCCTTGGCCC |
| GAPDH | F:
GGCAAATTCAACGGCACAGTCAAG |
| R:
TCGCTCCTGGAAGATGGTGATGG |
| *338F_806R | F:
ACTCCTACGGGAGGCAGCAG |
| R:
GGACTACHVGGGTWTCTAAT |
Immunofluorescence (IF)
For IF staining, mBMSCs were fixed with 4%
paraformaldehyde and permeabilised with 0.1% Triton X-100 (Beyotime
Institute of Biotechnology). Cells were then blocked with 5% bovine
serum albumin (BSA; MilliporeSigma) and incubated at 4°C with
primary antibodies [PPARγ, COL1A1, RUNX2 (cat. no. K003506P)
Beijing Solarbio Science & Technology Co., Ltd.] overnight. The
following day, cells were incubated at room temperature with
fluorophore-conjugated secondary antibodies. Nuclei were
counterstained with DAPI with 5 µg/ml (Beyotime Institute of
Biotechnology). Fluorescence signals were detected using a confocal
microscope (Leica Microsystems GmbH) at ×400 magnification.
Fluorescence intensity was analysed using ImageJ software.
Molecular docking
The molecular docking analysis of IPA and PPARγ was
performed using the CB-Dock2 platform (https://cadd.labshare.cn/cb-dock2/index.php), as
previously described (40). The
structure of IPA (PubChem compound ID: 3744) was obtained from
PubChem (https://pubchem.ncbi.nlm.nih.gov/compound/3744),
whereas the crystal structure of PPARγ (8BF1) was downloaded from
the Protein Data Bank (https://www.rcsb.org/structure/8BF1). The binding
sites and binding affinity between IPA and PPARγ were subsequently
predicted.
RNA sequencing
Total RNA from BMSCs was extracted using TRIzol
reagent according to the manufacturer's instructions. RNA quality
and integrity were assessed using 5300 Bioanalyser (Agilent
Technologies, Inc.), and samples with RNA integrity number
OD260/280=1.8~2.2 were used for library construction. Sequencing
libraries were generated using the NEBNext Ultra II RNA Library
Prep Kit (cat. no. E7770; New England Biolabs) according to the
manufacturer's instructions. The final library concentration was
determined using a Qubit fluorometer (Thermo Fisher Scientific,
Inc.) and quantitative PCR, and libraries were loaded at a
concentration of 4 nM for sequencing on the Illumina NovaSeq
platform, yielding ~50 million paired-end reads per sample. Raw
data processing, including adaptor trimming and quality filtering,
was performed using FastQC (v. 0.11.9; https://www.bioinformatics.babraham.ac.uk/projects/fastqc/).
High-quality reads were aligned to the mouse reference genome using
HISAT2 (v. 2.2.1; https://daehwankimlab.github.io/hisat2/). Transcript
abundance was quantified and expressed as transcripts per million.
Gene Ontology (GO) and Kyoto Encyclopaedia of Genes and Genomes
(KEGG) pathway enrichment analyses were performed using the
Majorbio online analysis platform (https://www.majorbio.com/).
16S rRNA gene sequencing
Microbial DNA was extracted from faecal samples
collected from sham and ovariectomy (OVX) mice. The 338F/806R
primers (Table I) were used to
amplify the V3-V4 region of the bacterial 16S rRNA gene, and PCR
products were purified using a PCR Clean-Up Kit (YuHua
Biotechnology). Sequencing libraries were prepared and sequenced
according to the manufacturer's instructions to generate paired-end
300-bp reads. All sequencing and bioinformatics analyses were
performed by Majorbio Bio-Pharm Technology.
IPA concentration measurement
Serum samples were collected from Sham and OVX mice.
Serum IPA concentrations were measured using gas
chromatography-mass spectrometry according to the manufacturer's
instructions.
Animal experiments
All animal procedures were approved by Peking
University Third Hospital (approval no. BCAA0292; Beijing, China)
and conducted in accordance with ARRIVE guidelines. 8-week-old
female C57BL/6 mice were acclimated for 1 week under standard
laboratory conditions. The initial body weight of the mice was
~18-22 g at the start of the experiment. Animals were housed at
22±2°C with 50±10% humidity under a 12 h light/dark cycle. Food and
water were available ad libitum throughout the study. A total of 24
mice were randomly assigned to 4 groups: Sham, OVX, OVX + IPA (10
mg/kg) and OVX + IPA (20 mg/kg). A postmenopausal OP model was
established by bilateral OVX as previously described (41). Briefly, the mice were
anaesthetised via inhalation of 3% isoflurane, with the anaesthesia
maintained via 2% isoflurane inhalation during surgery, and a small
dorsal midline incision was made to expose both ovaries, which were
subsequently ligated and removed. Sham-operated mice underwent the
same surgical procedure without ovary removal. Following surgery,
mice received daily oral administration of IPA or an equal volume
of normal saline for 10 weeks. At the end of the experiment,
animals were euthanized by overdose of pentobarbital sodium
administered via intraperitoneal injection (150 mg/kg). Death was
confirmed by the absence of spontaneous respiration, heartbeat and
pedal withdrawal reflex. The femurs and serum samples were
collected for further analyses. A total of ~600 µl of whole
blood was collected via orbital blood sampling following eyeball
removal, and serum was subsequently isolated by centrifugation at
3,000 × g for 15 min at 4°C.
Micro-CT and histological analysis
Femurs were scanned using a SkyScan micro-CT system
(Bruker Corporation). The region of interest was defined as a
1-mm-high area distal to the growth plate. Three-dimensional
reconstruction images were generated using CTvox software.
Trabecular bone parameters, including bone volume fraction (BV/TV),
trabecular number (Tb.N), trabecular thickness (Tb.Th) and
trabecular separation (Tb.Sp), were analysed using CTAn software
(V2.0; https://www.blue-scientific.com/bruker-micro-ct-software/).
For haematoxylin and eosin (H&E) staining, decalcified bone
tissues were processed and stained according to the manufacturer's
protocols (Wuhan Servicebio Technology Co., Ltd.).
Von Kossa staining
Undecalcified bone samples were subjected to Von
Kossa staining according to the manufacturer's instructions (Wuhan
Servicebio Technology Co., Ltd.). Images were captured using a
light microscope, and mineralised areas were quantified using
ImageJ software.
Immunohistochemistry (IHC)
Decalcified bone tissues were embedded in paraffin.
Tissue sections (5 µm) were deparaffinised in xylene and
rehydrated through a graded ethanol series (100, 95, 85, and 75%
ethanol) before antigen retrieval by trypsin digestion. Sections
were then blocked with 3% hydrogen peroxide and incubated with 5%
BSA for 1 h. Subsequently, the sections were incubated at 4°C with
primary antibodies overnight. The following day, HRP-conjugated
secondary antibodies were applied at room temperature for 1 h.
Signals were developed using a DAB substrate. Images were acquired
under a bright-field microscope, and staining intensity was
quantified using ImageJ software.
Enzyme-linked immunosorbent assay
Serum levels of P1NP and CTX-1 were measured using
ELISA kits (cat. nos. E-EL-M3023 and E-EL-M0233; Elabscience
Biotechnology, Inc.) according to according to the manufacturer.
Absorbance at 450 nm was measured using a microplate reader
(BioTek; Agilent Technologies, Inc.). All samples were analysed in
triplicate.
Statistical analysis
Data were presented as the mean ± SD. All
experiments were independently repeated at least three times.
Comparisons between two groups were performed using unpaired
Student's t-test, whereas comparisons among multiple groups were
analysed using one-way or two-way analysis of variance followed by
Tukey's post hoc test. P<0.05 was considered to indicate a
statistically significant difference. Statistical analyses and data
visualisation were performed using GraphPad Prism (v. 8.0;
Dotmatics).
Results
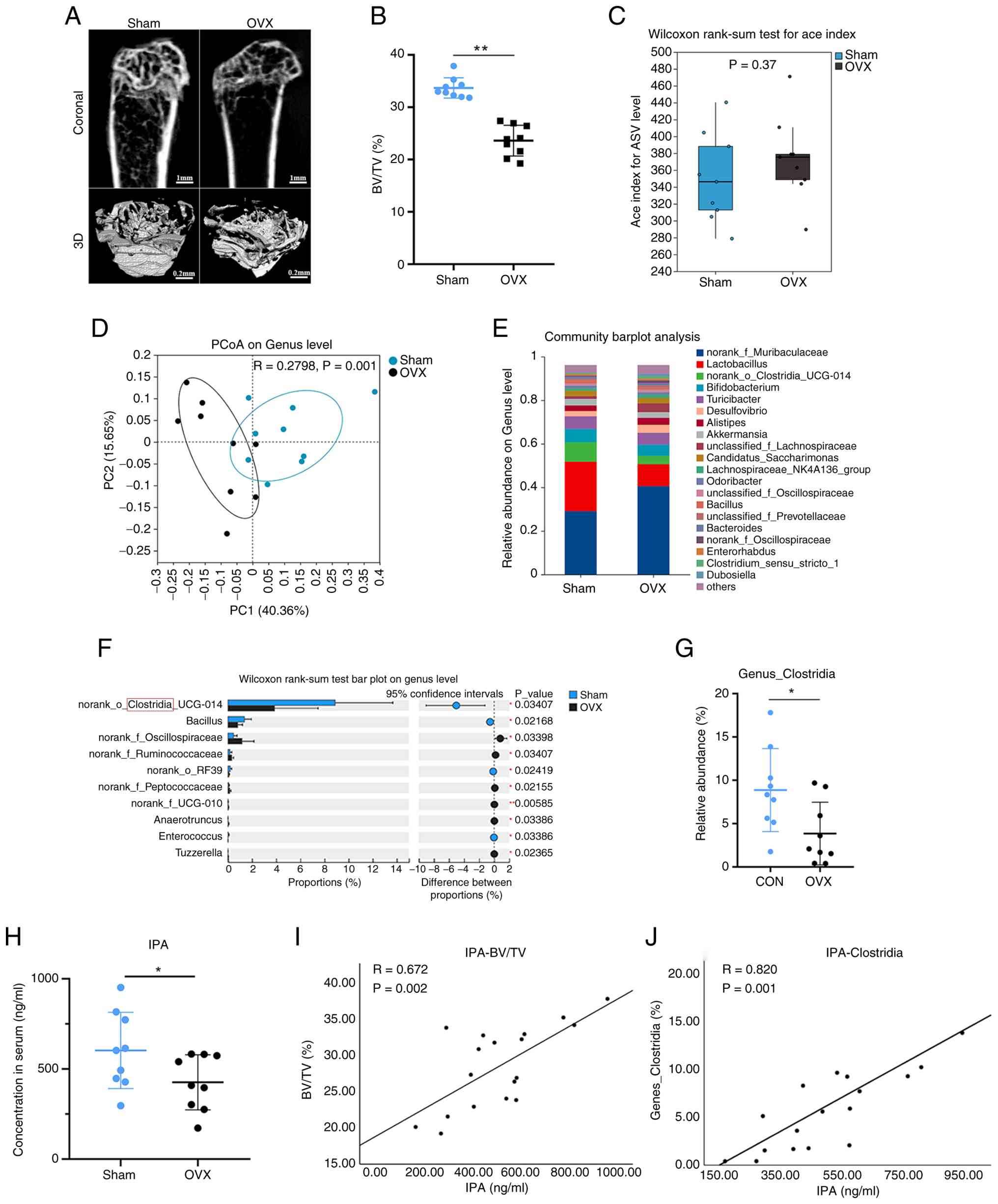
C. sporogenes-derived IPA is depleted in
OP and correlates with bone loss
To investigate whether gut microbiota-derived
metabolites contribute to OP, an OVX-induced mouse model was first
established. The OVX model effectively mimicked postmenopausal OP
(42). Micro-CT analysis
confirmed a marked reduction in trabecular bone mass in OVX mice,
validating the successful establishment of osteoporotic phenotypes
(Fig. 1A and B). 16S rRNA
sequencing was next performed to profile gut microbiota
composition. Although α-diversity did not differ significantly
between groups (Fig. 1C),
β-diversity analysis revealed a clear separation between sham and
OVX mice, indicating substantial alterations in microbial community
structure (Fig. 1D).
Consistently, the gut microbial health index was significantly
reduced in the OVX group (Fig.
S1A), whereas the microbial dysbiosis index was significantly
increased (Fig. S1B).
Taxonomic profiling identified distinct alterations
in bacterial genera between the two groups. OVX mice exhibited
increased abundance of Muribaculaceae and reduced abundance
of the Lactobacillus and Clostridia genera compared
with sham controls (Fig. 1E).
The abundance of Clostridia was significantly decreased in
the OVX group, with statistical significance confirmed by Wilcoxon
analysis (Fig. 1F and G). Given
that Clostridia is a major bacterial genus involved in IPA
production (26), serum IPA
concentrations were next measured. Metabolite analysis demonstrated
a significant reduction in circulating IPA levels in OVX mice
(Fig. 1H). Importantly,
correlation analyses revealed that serum IPA levels were positively
associated with both BV/TV and Clostridium abundance
(Fig. 1I and J). These findings
suggested that depletion of microbiota-derived IPA is a
characteristic feature of oestrogen deficiency-induced OP and may
contribute functionally to bone loss.
IPA protects BMSCs against oxidative
stress-induced apoptosis
It was next investigated whether IPA modulates BMSC
survival under oxidative stress conditions. CCK-8 assays
demonstrated that IPA alone (0-100 µM) neither promoted nor
inhibited mBMSC proliferation (Fig.
2A). Exposure to H2O2 significantly
reduced cell viability, whereas IPA treatment dose-dependently
rescued this effect without altering basal proliferation (Fig. 2B). Live/dead staining further
confirmed that IPA markedly attenuated oxidative stress-induced
cell death (Fig. 2C and D).
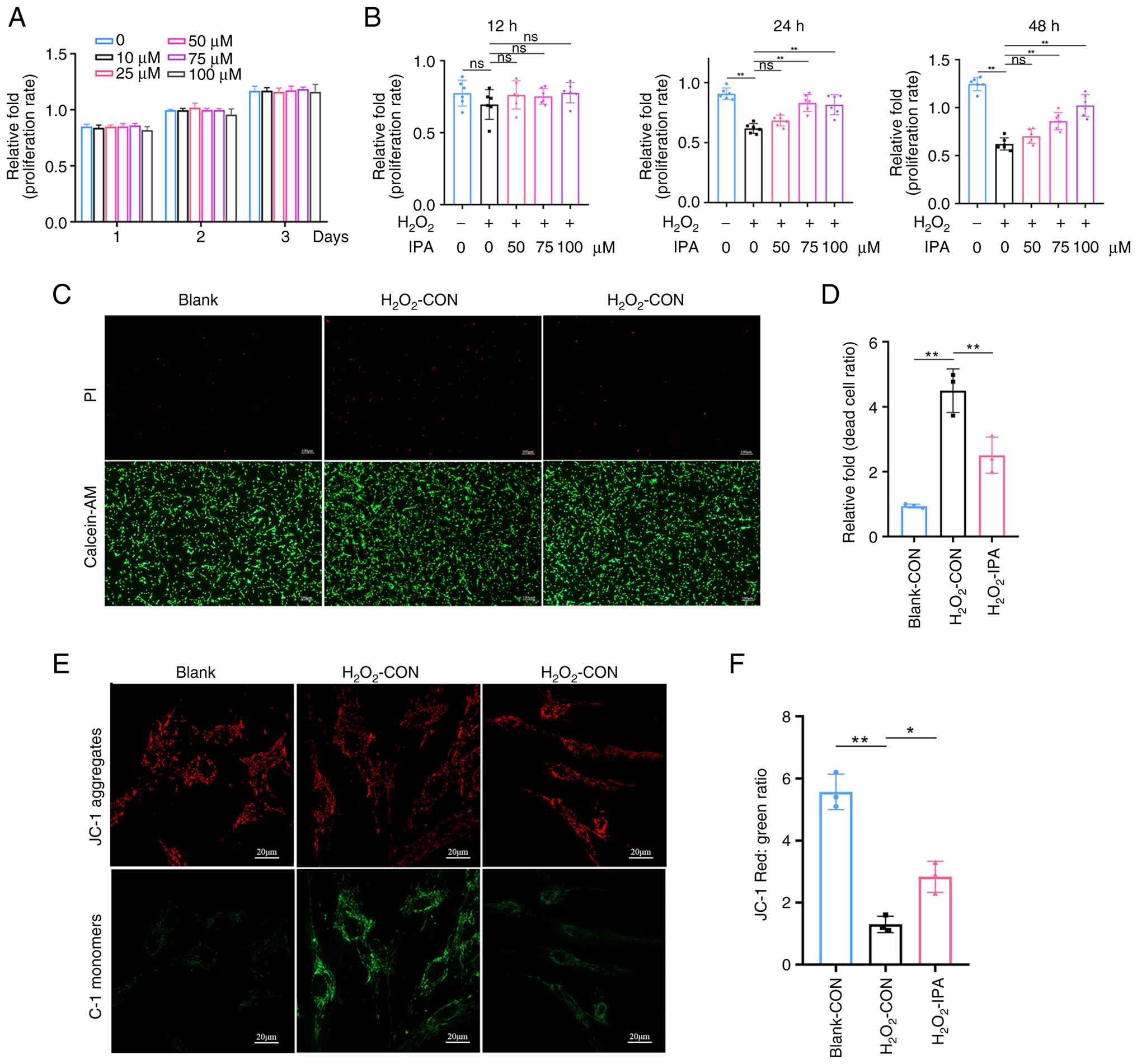 | Figure 2IPA protects mBMSCs against
H2O2-induced apoptotic injury. (A) Effects of
IPA on mBMSC viability over 1, 2, and 3 days, as determined by the
CCK-8 assay. (B) Effects of IPA pretreatment on mBMSC viability
following H2O2 exposure for 12, 24, and 48 h,
as determined by the CCK-8 assay. (C) Live/dead staining of mBMSCs
on day 1 using propidium iodide (dead cells) and Calcein-AM (live
cells). (D) Quantification of the dead-cell ratio based on
live/dead staining. (E) Mitochondrial membrane potential of
H2O2-treated mBMSCs with or without IPA
treatment for 1 day, assessed by JC-1 staining. (F) Quantification
of the red/green fluorescence ratio from JC-1 staining. All
experiments were repeated at least three times. Data are presented
as the mean ± SD. **P<0.01 compared with the control
group. IPA, indole-3-propionic acid; mBMSCs, mouse bone marrow
stromal cells; CCK-8, Cell Counting Kit-8; ns, not significant. |
Because loss of MMP is a hallmark of early apoptosis
(43), JC-1 staining was
performed to evaluate mitochondrial function.
H2O2 treatment induced a significant
reduction in MMP, whereas IPA administration effectively restored
mitochondrial integrity (Fig. 2E and
F). Collectively, these results demonstrated that IPA exerts
cytoprotective effects on BMSCs by preserving mitochondrial
function and attenuating oxidative stress-induced apoptosis.
IPA restores osteogenic capacity and
suppresses adipogenic differentiation under oxidative stress
It was next examined whether IPA directly regulates
BMSC lineage commitment. Under physiological conditions, IPA did
not significantly affect mineralised nodule formation (Fig. S2A and B) or ALP activity
(Fig. S2C and D). Consistently,
western blot analysis showed that IPA had no appreciable effect on
the expression of osteogenic markers, including RUNX2, SP7, and OPN
(Fig. S2E and F). These
findings indicate that IPA does not function as a basal
osteo-inductive factor.
By contrast, under oxidative stress conditions,
H2O2 markedly impaired osteogenic
differentiation, as evidenced by reduced ALP activity and decreased
mineralised nodule formation (Fig.
3A and B). IPA treatment dose-dependently restored osteogenic
capacity (Fig. 3D and E). In
parallel, oxidative stress significantly promoted adipogenic
differentiation (42), as
reflected by increased lipid accumulation and upregulation of
adipogenic markers (Fig. 3C and
F). Further analysis of lineage-specific gene expression
demonstrated that IPA prevented the
H2O2-induced downregulation of osteogenic
genes, including Col1a1, Runx2, Sp7 and Ocn (Fig. S3A), while reversing the
upregulation of adipogenic genes, including Adiponectin,
Cebpα, Fabp2 and Srebp-1 (Fig.
S3B). Western blot analysis further confirmed that IPA rescued
the H2O2-induced downregulation of osteogenic
proteins (Fig. 3G and H).
Similar findings were observed by IF staining (Fig. 3I; Fig. S4A and B), further supporting the
protective effect of IPA on osteogenic differentiation under
oxidative stress conditions. Together, these findings indicated
that IPA functions as a context-dependent regulator that restores
osteogenicadipogenic balance under pathological conditions rather
than altering baseline differentiation.
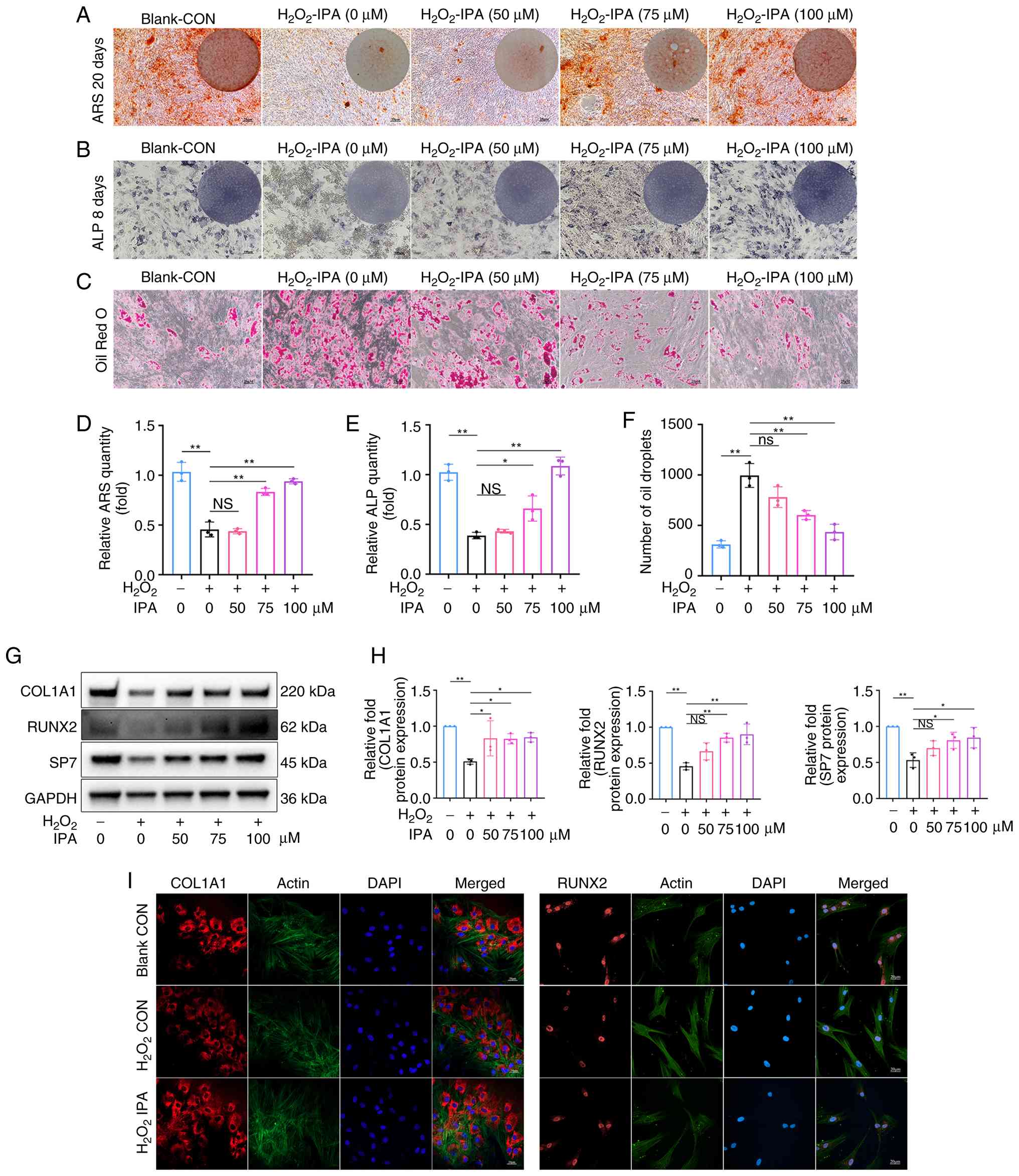 | Figure 3IPA attenuates
H2O2-induced suppression of osteogenic
function in vitro. (A) ARS staining of mBMSCs after 20 days
of osteogenic differentiation. Scale bar, 25 µm. (B) ALP
staining of mBMSCs after 8 days of osteogenic differentiation.
Scale bar, 100 µm. (C) Oil Red O staining of mBMSCs after 8
days of osteogenic differentiation. Scale bar, 25 µm. (D)
Quantification of ARS staining. (E) Quantification of ALP staining.
(F) Quantification of Oil Red O staining. (G) Expression of
osteogenesis-related proteins in mBMSCs after 5 days of osteogenic
differentiation, assessed by western blotting. (H) Quantification
of western blot results. (I) Representative immunofluorescence
images of RUNX2 and COL1A1 expression. Scale bar, 20
µm. All experiments were repeated at least three times. Data
are presented as the mean ± SD. *P<0.05 and
**P<0.01 compared with the control group. IPA,
indole-3-propionic acid; ARS, alizarin Red S; mBMSCs, mouse bone
marrow stromal cells; ALP, alkaline phosphatase; COL1A1, collagen,
type I, alpha 1; RUNX2, Runt-related transcription factor 2; NS,
not significant. |
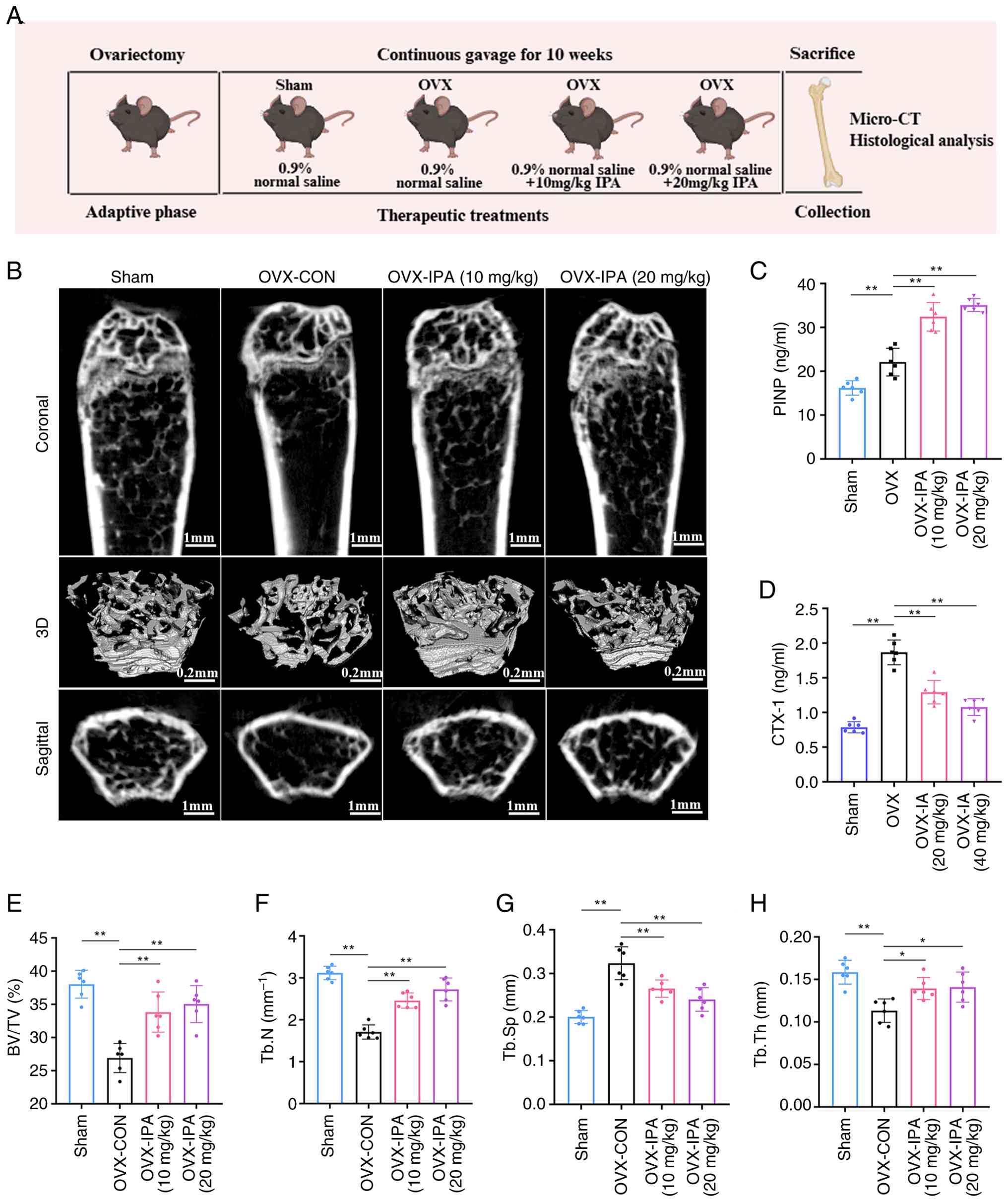
IPA supplementation alleviates bone loss
and enhances bone formation in vivo
To determine whether these protective effects also
occur in vivo, an OVX mouse model was established and IPA
was administered orally for 10 weeks. A schematic overview of the
animal experimental design is presented in Fig. 4A. OVX mice exhibited
significantly increased body weight and reduced uterine weight
compared with the control group (Fig. S5A and B).
Micro-CT analysis revealed that IPA markedly
alleviated OVX-induced bone loss (Fig. 4B). IPA supplementation
significantly improved trabecular bone architecture, as indicated
by increased BV/TV, Tb.N and Tb.Th, along with reduced Tb.Sp
(Fig. 4E-H). Consistently,
biochemical analyses demonstrated elevated levels of the bone
formation marker PINP and decreased levels of the bone resorption
marker CTX-1 following IPA treatment, indicating a shift toward
anabolic bone remodelling (Fig. 4C
and D). These results demonstrated that IPA supplementation
effectively mitigates oestrogen deficiency-induced bone loss and
promotes bone formation in vivo.
IPA corrects the osteogenic-adipogenic
imbalance in the bone marrow of OVX mice
To investigate further the cellular basis of these
effects, BMSCs isolated from experimental mice were analysed. Cells
derived from OVX mice exhibited impaired mineralisation capacity,
as reflected by significantly reduced ALP activity and ARS staining
ability (Fig. 5A and B), along
with increased intracellular lipid accumulation, confirming
disruption of lineage balance (Fig.
5H). Importantly, IPA treatment restored osteogenic capacity
(Fig. 5C and D) while
suppressing adipogenesis (Fig.
5I) in BMSCs isolated from OVX mice. IPA administration
significantly upregulated the expression of osteogenic genes and
proteins (Fig. 5E-G), while
suppressing adipogenic gene expression in OVX mice (Fig. 5J). These findings demonstrated
that IPA alleviates OP by re-establishing bone marrow lineage
equilibrium.
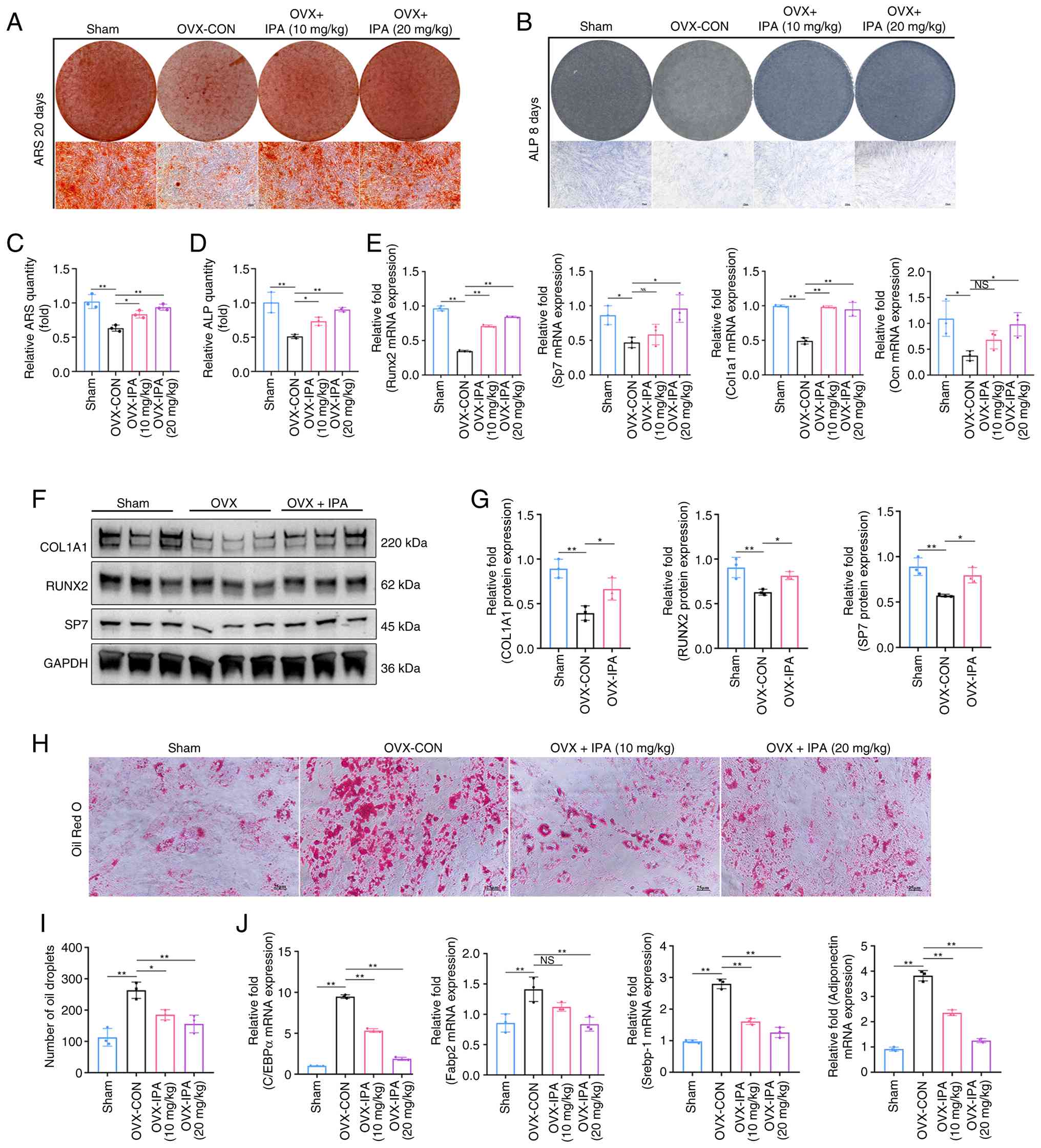 | Figure 5IPA restores OVX-induced bone-fat
imbalance in vivo. (A) ARS staining of mBMSCs isolated from
mice after 20 days of osteogenic differentiation. Scale bar, 25
µm. (B) ALP staining of mBMSCs isolated from mice after 8
days of osteogenic differentiation. Scale bar, 25 µm. (C)
Quantification of ARS staining. (D) Quantification of ALP staining.
(E) Expression of osteogenesis-related genes after 7 days of
osteogenic differentiation, assessed by RT-qPCR. (F) Expression of
osteogenic-related proteins after 7 days of osteogenic
differentiation examined using western blot analysis. (G)
Quantification of results of western blot analysis. (H) Oil Red O
staining of mBMSCs isolated from mice after 8 days of adipogenic
differentiation. Scale bar, 25 µm. (I) Quantification of Oil
Red O staining. (J) Expression of adipogenesis-related genes after
10 days of adipogenic differentiation, assessed by RT-qPCR. All
experiments were repeated at least 3 times. Data are presented as
the mean ± SD. *P<0.05 and **P<0.01
compared with the control group. IPA, indole-3-propionic acid; OVX,
ovariectomy; ARS, alizarin Red S; mBMSCs, mouse bone marrow stromal
cells; ALP, alkaline phosphatase; RT-qPCR, reverse
transcription-quantitative PCR; NS, not significant. |
IPA reprograms BMSC fate through
suppression of PPARγ signaling
To elucidate the underlying mechanisms,
transcriptomic sequencing was performed on mBMSCs treated with
H2O2 in the presence or absence of IPA.
RNA-seq analysis identified a total of 1,254 differentially
expressed genes (DEGs; fold change ≥3.0), including 509 upregulated
and 745 downregulated genes (Figs.
6A and S6A). GO functional
annotation revealed that IPA-regulated genes were enriched in
pathways associated with oxidative stress defence, cell
proliferation and immune regulation (Fig. S6B). GO enrichment analysis
further highlighted biological processes related to osteogenic
differentiation, cell cycle regulation and oxidative
phosphorylation (Fig. 6B). KEGG
pathway analysis demonstrated significant enrichment in PPAR
signalling, cholesterol biosynthesis, adipogenesis-related
pathways, oxidative phosphorylation, and p53-mediated apoptosis
(Fig. 6C). Gene Set Enrichment
Analysis further showed that IPA markedly suppressed several
adipogenesis-related pathways, including unsaturated fatty acid
synthesis, cholesterol biosynthesis, and the PPARγ signalling
pathway, while also inhibiting oxidative phosphorylation (Fig. 6D). These findings suggested that
IPA preserves osteogenic function under oxidative stress, at least
in part, by inhibiting adipogenic differentiation.
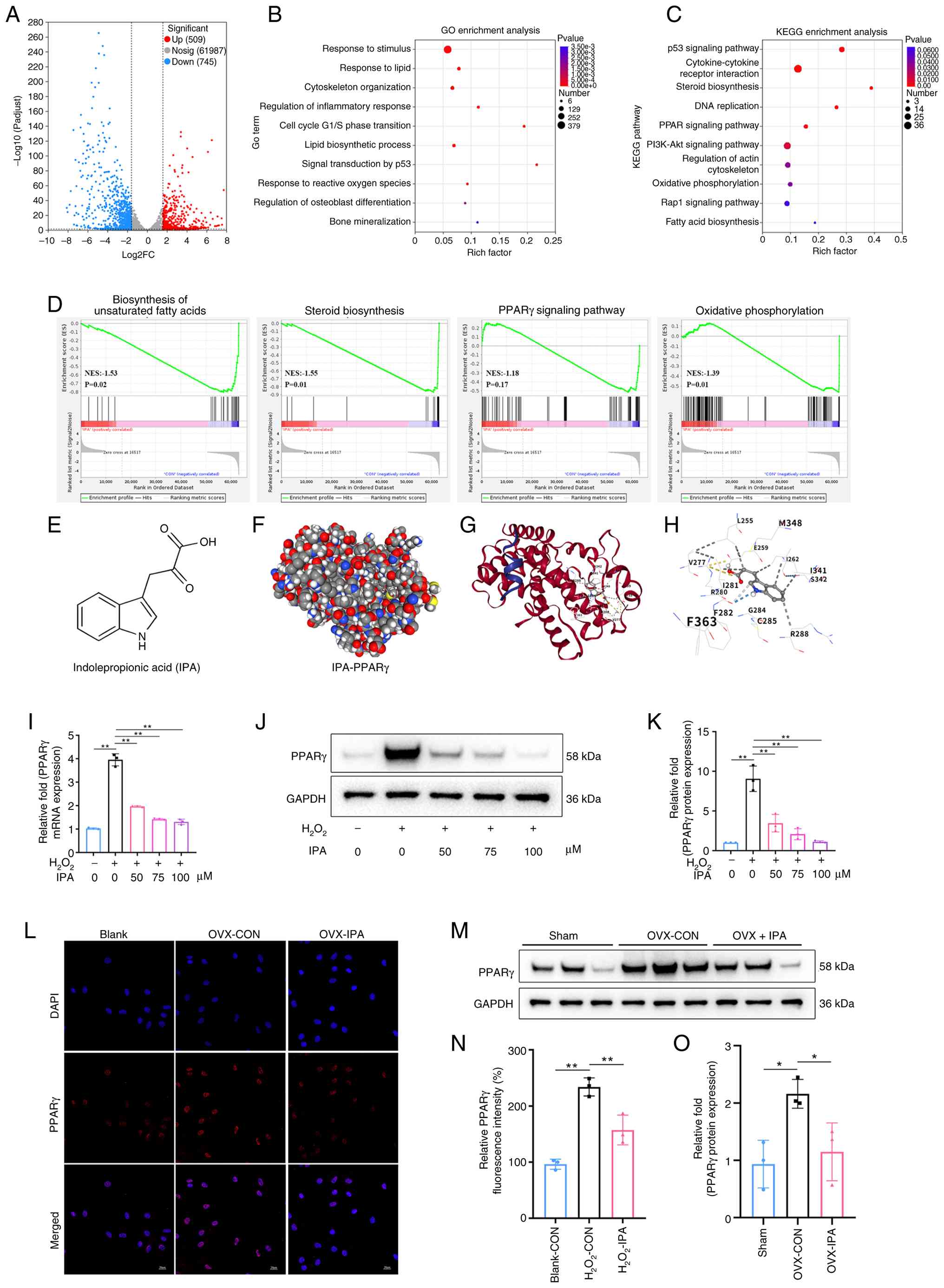 | Figure 6IPA regulates the
osteogenic-adipogenic balance of BMSCs by targeting the PPARγ
signalling pathway. (A) Volcano plots of DEGs between the two
groups (n=3 per group). (B) GO biological process enrichment
analysis of common DEGs. (C) KEGG enrichment analysis showing
significantly altered signalling pathways following IPA treatment.
(D) Gene Set Enrichment Analysis of pathways related to
biosynthesis of unsaturated fatty acids, steroid biosynthesis, the
PPARγ signalling pathway and oxidative phosphorylation. (E)
Chemical structure of IPA. (F) Three-dimensional docking model of
IPA and PPARγ. (G) Two-dimensional docking model of IPA and PPARγ.
(H) Specific binding sites between AKT and including
indole-3-acetic acid. (I) Expression of PPARγ mRNA assessed by
reverse transcription-quantitative PCR. (J) Expression of PPARγ
protein assessed by western blotting. (K) Quantification of western
blot results. (L) Representative IF images of PPARγ expression in
the presence of IPA (100 µM). Scale bar, 20 µm. (M)
Expression of PPARγ protein in mBMSCs isolated from mice in
vivo, assessed by western blotting. (N) Quantification of PPARγ
fluorescence intensity by IF analysis. (O) Quantification of
western blot results. All experiments were repeated at least three
times. Data are presented as the mean ± SD. *P<0.05
and **P<0.01 compared with the control group. IPA,
indole-3-propionic acid; BMSCs, bone marrow stromal cells; PPARγ,
peroxisome proliferator-activated receptor gamma; DEGs,
differentially expressed genes; GO, Gene Ontology; KEGG, Kyoto
Encyclopedia of Genes and Genomes; IF, immunofluorescence. |
The three-dimensional structure of IPA is shown in
Fig. 6E. Molecular docking
analysis revealed a high predicted binding affinity (-6.6 kcal/mol)
between IPA and PPARγ (Fig. 6F and
G). Docking results demonstrated that IPA formed multiple
hydrogen bonds and ionic interactions with key residues, including
Arg280, Ile262 and Ile281, indicating a plausible structural basis
for direct interaction (Fig.
6H). Consistently, H2O2-induced
upregulation of PPARγ expression was significantly attenuated by
IPA at both the mRNA and protein levels (Fig. 6I-K). IF analysis further
confirmed reduced nuclear localisation of PPARγ following IPA
treatment (Fig. 6L and N).
Consistent with the in vitro findings, BMSCs isolated from
OVX mice displayed elevated PPARγ expression, which was
significantly reduced following IPA supplementation in vivo
(Fig. 6M and O). These findings
indicated that IPA regulates BMSC lineage commitment by inhibiting
PPARγ signalling.
IPA supplementation suppresses PPARγ
signalling to promote bone formation in vivo
To validate this mechanism in vivo,
histological and IHC analyses were performed. H&E staining
revealed that IPA supplementation significantly improved BV/TV
compared with the OVX group (Fig.
7A). Consistent with the H&E staining results, Von Kossa
staining demonstrated reduced mineral deposition in OVX mice, which
was effectively restored following IPA administration (Fig. 7B). IHC analysis further confirmed
these observations. BMSCs derived from OVX-induced osteoporotic
mice exhibited lower expression of osteogenic proteins, including
COL1A1 and RUNX2, whereas IPA treatment restored their expression
levels (Fig. 7C and D). By
contrast, PPARγ expression was significantly downregulated by IPA
supplementation compared with the OVX group (Fig. 7E). Furthermore, no obvious
pathological injury was observed in the heart, liver, spleen, lung,
or kidney following oral IPA supplementation (Fig. S7). Collectively, these data
demonstrated that IPA alleviates OP by suppressing PPARγ
signalling, thereby restoring the osteogenic-adipogenic balance and
promoting bone formation.
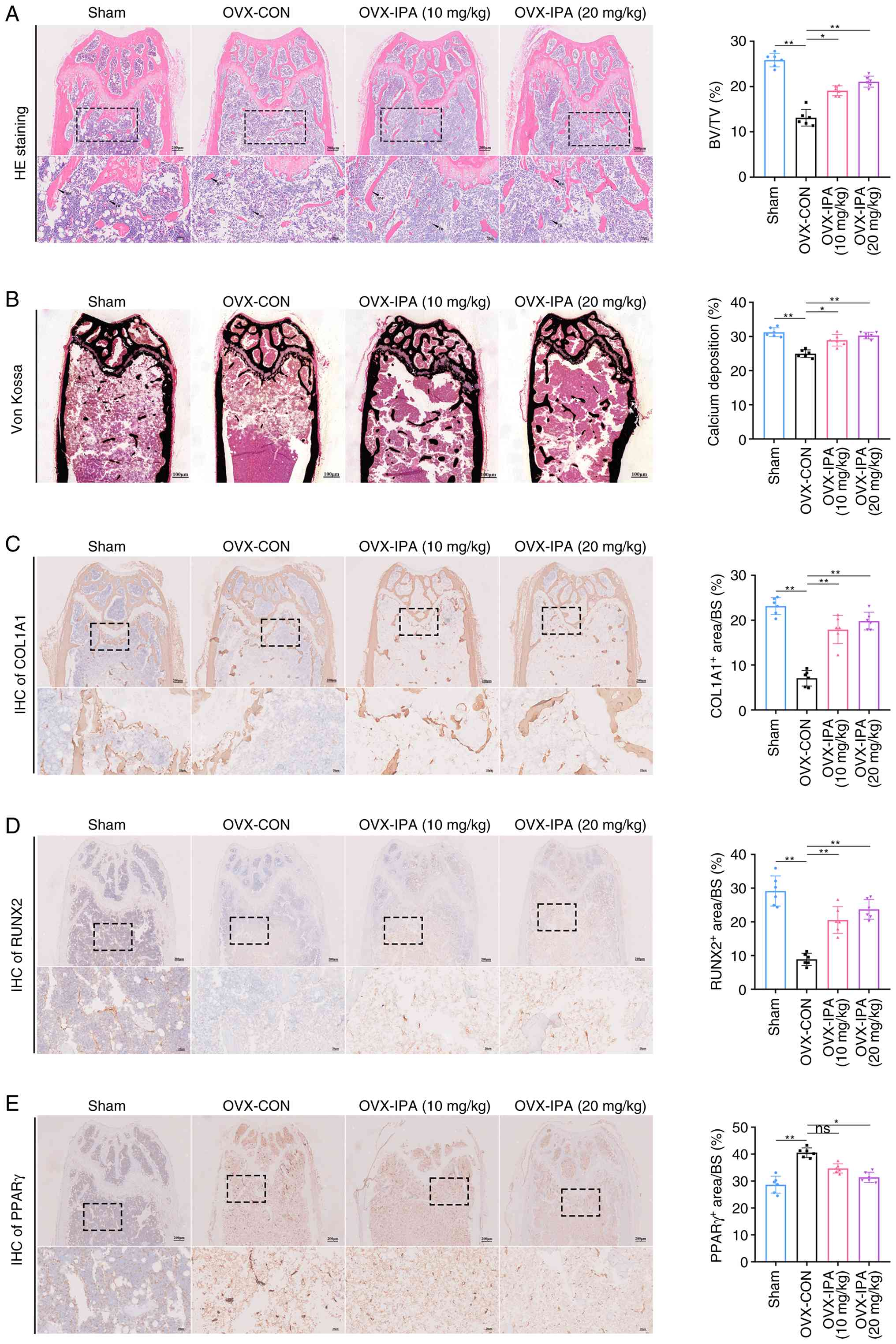 | Figure 7IPA supplementation suppresses PPARγ
signalling to enhance bone formation and attenuate OVX-induced bone
loss. (A) Representative H&E staining images and quantification
of BV/TV in the four experimental groups. Scale bar, 200 µm.
(B) Representative Von Kossa staining images and quantification of
calcium deposition in the four groups. Scale bar, 100 µm.
(C) Histological analysis of femoral tissue stained for COL1A1 and
quantification of the COL1A1-positive area by IHC staining (n=6).
(D) Histological analysis of femoral tissue stained for RUNX2 and
quantification of the RUNX2-positive area by IHC staining (n=6).
(E) Histological analysis of femoral tissue stained for PPARγ and
quantification of the PPARγ-positive area by IHC staining (n=6).
Data are presented as the mean ± SD. *P<0.05 and
**P<0.01 compared with the control group. IPA,
indole-3-propionic acid; PPARγ, peroxisome proliferator-activated
receptor gamma; OVX, ovariectomy; COL1A1, collagen, type I, alpha
1; IHC, immunohistochemistry; RUNX2, Runt-related transcription
factor 2; ns, not significant. |
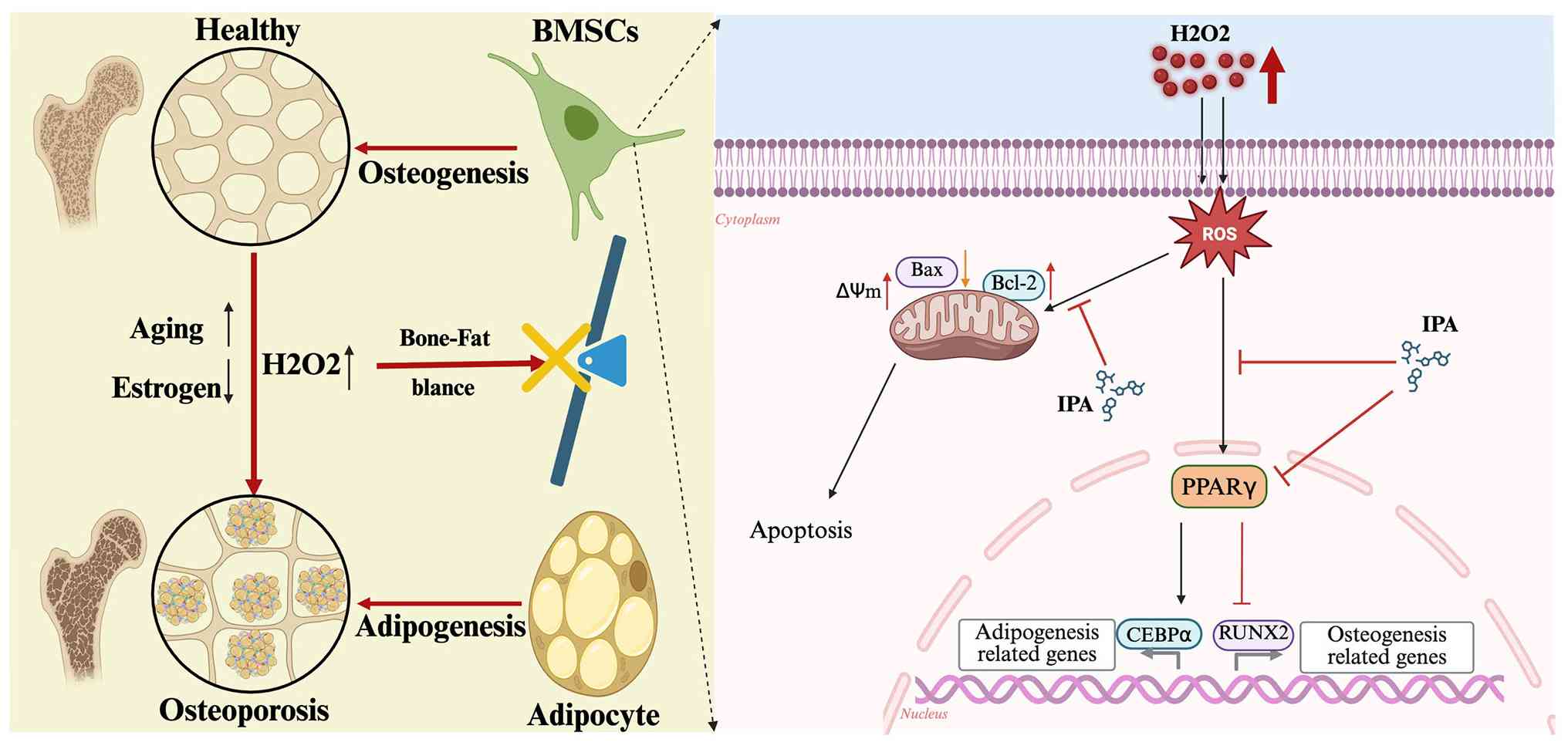
Discussion
In the present study, IPA, a gut microbiota-derived
metabolite, was identified as a critical regulator of bone marrow
lineage commitment and skeletal homeostasis. The present findings
established a previously unrecognised mechanistic link between
microbial metabolism and OP, demonstrating that IPA restores the
osteogenic-adipogenic balance by suppressing PPARγ signalling. A
schematic overview of the study is presented in Fig. 8. A key conceptual advance is the
shift in focus from bone resorption-centred mechanisms toward the
regulation of BMSC fate. Although previous studies have primarily
highlighted the role of microbial metabolites in osteoclast
activity, the present results demonstrated that IPA directly
regulates mesenchymal lineage allocation. This distinction is
important, as impaired osteogenesis, rather than excessive bone
resorption, is increasingly recognised as a major contributor to
age- and oestrogen deficiency-related bone loss.
In recent years, the role and mechanisms of the gut
microbiota and its metabolites in the pathogenesis of OP have
gradually gained recognition (28). It has been demonstrated that gut
microbiota-derived metabolites exert significant regulatory effects
on both osteoblasts and osteoclasts (25). Tryptophan is an essential amino
acid (21). Undigested dietary
tryptophan is metabolised by gut microbiota, including the
Clostridium and Bacteroides genera, to produce indole
and its derivatives. These indole derivatives constitute a distinct
class of tryptophan metabolites produced exclusively through
microbial tryptophan catabolism (44). Among them, IPA, a
tryptophan-derived microbial metabolite predominantly produced by
the Clostridia genus, has attracted increasing attention
because of its involvement in metabolic regulation, inflammation,
and gut barrier protection (26). In the present study, it was found
that both Clostridium abundance and IPA levels were
significantly reduced in OVX mice. Moreover, IPA levels were
positively correlated with the abundance of the Clostridium
genus, suggesting that reduced Clostridium abundance
contributes to decreased IPA production. The observed association
between reduced Clostridium abundance and decreased IPA
levels is correlative. Further studies using microbiota
manipulation or Clostridium-specific interventions are
needed to confirm a direct causal relationship.
The results of the present study further
demonstrated that IPA supplementation alleviated OP-associated bone
loss. It has been shown that IPA derived from Clostridium
inhibits osteoclast formation and that Clostridium abundance
is decreased in OVX-induced OP mouse models (35), which is consistent with the
present findings. Other studies reported that
Lactobacillus-derived IPA and IAA levels were significantly
decreased in OVX mice (34,36). These findings suggest that
variations in intestinal microbiota composition among individuals
may be influenced by host metabolism and environmental factors.
Therefore, further multicentre studies with larger sample sizes are
required to validate gut microbial alterations in OP populations.
In addition, the present study only explored the therapeutic
effects of oral IPA supplementation in OVX mice. The potential
effects of supplementation with IPA-producing bacteria or faecal
microbiota transplantation were not investigated and warrant
further study in the future.
The present data further demonstrated that IPA acts
as a context-dependent regulator that exerts minimal effects under
physiological conditions but becomes highly active under oxidative
stress. This characteristic suggests that IPA may selectively
restore pathological imbalance without disrupting normal tissue
homeostasis, representing a potential advantage over conventional
pharmacological agents. Mechanistically, PPARγ was identified as a
central target of IPA. As a master regulator of adipogenesis, PPARγ
plays a critical role in determining BMSC fate (45). Aberrant activation of PPARγ in OP
promotes adipocyte formation at the expense of osteoblast
differentiation. The current transcriptomic, biochemical and
molecular docking analyses collectively support the notion that IPA
suppresses PPARγ signalling, thereby reprogramming lineage
commitment toward osteogenesis. These findings provide a
mechanistic basis for targeting the bone-fat balance as a
therapeutic strategy.
In addition to its effects on differentiation, IPA
also exhibited cytoprotective properties by mitigating oxidative
stress-induced apoptosis in BMSCs. Given that oxidative stress is a
major driver of skeletal aging and oestrogen deficiency-associated
bone loss (46,47), this dual function further
strengthens the therapeutic potential of IPA. Importantly, the
present in vivo findings demonstrated that oral
administration of IPA significantly improved bone mass and
microarchitecture without detectable systemic toxicity. These
findings highlight the translational potential of
microbiota-derived metabolites as safe and effective therapeutic
agents.
The present study also had certain limitations.
First, although the present findings strongly suggested that IPA
regulates BMSC differentiation through the PPARγ signalling
pathway, based on transcriptomic and molecular docking analyses,
the detailed downstream molecular mechanisms linking PPARγ
suppression to restored osteogenesis were not fully elucidated. In
addition, several key pathways closely associated with osteogenesis
and oxidative stress responses, including the Wnt/β-catenin,
BMP/Smad, AMPK and NRF2 signalling pathways (45), were not investigated. Future
studies employing genetic gain- and loss-of-function models, as
well as pharmacological modulation of specific targets, are needed
to clarify the precise molecular interactions underlying the
effects of IPA. Second, although IPA supplementation improved bone
mass in OVX mice, the broader framework of gut microbial modulation
was not addressed. Because the abundance of Clostridia is
strongly correlated with IPA levels, microbial interventions such
as faecal microbiota transplantation, selective probiotic
colonisation and dietary strategies that enhance IPA-producing
bacteria may provide more sustained and physiological approaches
for correcting the bone-fat imbalance associated with OP. The
stability, colonisation efficiency and host-microbe interactions of
Clostridia and other IPA-producing bacteria should therefore
be further explored in future studies. Additionally, the present
study demonstrated promising therapeutic effects of IPA in
vivo. However, IPA was not compared with standard
anti-osteoporotic drugs, and the study was limited to preclinical
models. Future work should evaluate its comparative efficacy,
safety, and translational potential.
In conclusion, IPA was identified as a key
microbiota-derived regulator of bone-fat balance in OP. IPA
restores skeletal homeostasis by reprogramming BMSC lineage
commitment through suppression of PPARγ signalling and protection
against oxidative stress. These findings uncover a previously
unrecognised gut-bone metabolic axis and highlight IPA as a
promising therapeutic strategy for OP.
Supplementary Data
Availability of data and materials
The data generated in the present study may be found
in the NCBI Sequence Read Archive under accession numbers
PRJNA1327782 and PRJNA1371077 or at the following URL: https://www.ncbi.nlm.nih.gov/bioproject/PRJNA1327782;
https://www.ncbi.nlm.nih.gov/bioproject/PRJNA1371077.
Authors' contributions
JB wrote the original draft, conducted
investigation, developed methodology, curated data, and wrote,
reviewed and edited the manuscript. RW wrote the original draft and
developed methodology. JF conducted investigation, data curation
and visualization. SS conducted formal analysis and data curation.
GS developed methodology and curated data. QY and AS visualized and
curated data. DH validated data and developed methodology. SG and
FZ wrote, reviewed and edited the manuscript, validated data,
conceptualized and supervised the study, and acquired funding. YL
wrote, reviewed and edited the manuscript, validated data,
conceptualized and supervised the study. All authors agree to be
accountable for all aspects of work ensuring integrity and
accuracy. All authors read and approved the final version of the
manuscript. JB and FZ confirm the authenticity of all the raw
data.
Ethics approval and consent to
participate
All animal experiments were approved by the Peking
University Third Hospital (approval no. BCAA0292; Beijing, China)
and conducted in accordance with ARRIVE guidelines.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Abbreviations:
|
ALP
|
alkaline phosphatase
|
|
AST
|
aspartate aminotransferase
|
|
COL1A1
|
collagen, type I, alpha 1
|
|
RUNX2
|
Runt-related transcription factor
2
|
|
PPARγ
|
peroxisome proliferator-activated
receptor gamma
|
|
FBS
|
fetal bovine serum
|
|
CPC
|
cetylpyridinium chloride
|
|
BSA
|
bovine serum albumin
|
|
CTX-1
|
type I collagen C-terminal
telopeptide
|
|
P1NP
|
pro-peptide of type I procollagen
|
|
BV/TV
|
bone volume fraction
|
|
Tb.N
|
trabecular number
|
|
Tb.Th
|
trabecular thickness
|
|
Tb.Sp
|
trabecular separation
|
Acknowledgments
Not applicable.
Funding
The present study was supported by the National Natural Science
Foundation of China (grant nos. 81971160 and 82402806).
References
|
1
|
Wang L, Yu W, Yin X, Cui L, Tang S, Jiang
N, Cui L, Zhao N, Lin Q, Chen L, et al: Prevalence of osteoporosis
and fracture in China: The China osteoporosis prevalence study.
JAMA Netw Open. 4:e21211062021. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Curtis EM, Moon RJ, Dennison EM, Harvey NC
and Cooper C: Recent advances in the pathogenesis and treatment of
osteoporosis. Clin Med (Lond). 16:360–364. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rachner TD, Khosla S and Hofbauer LC:
Osteoporosis: Now and the future. Lancet. 377:1276–1287. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ayers C, Kansagara D, Lazur B, Fu R, Kwon
A and Harrod C: Effectiveness and safety of treatments to prevent
fractures in people with low bone mass or primary osteoporosis: A
living systematic review and network meta-analysis for the american
college of physicians. Ann Intern Med. 176:182–195. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Stegen S and Carmeliet G: Metabolic
regulation of skeletal cell fate and function. Nat Rev Endocrinol.
20:399–413. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang J, Hu W, Zou Z, Li Y, Kang F, Li J
and Dong S: The role of lipid metabolism in osteoporosis: Clinical
implication and cellular mechanism. Genes Dis. 11:1011222024.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zou J, Chen H, Fan X, Qiu Z, Zhang J and
Sun J: Garcinol prevents oxidative stress-induced bone loss and
dysfunction of BMSCs through NRF2-antioxidant signaling. Cell Death
Discov. 10:822024. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Alves CH, Farrell E, Vis M, Colin EM and
Lubberts E: Animal models of bone loss in inflammatory arthritis:
From cytokines in the bench to novel treatments for bone loss in
the Bedside-a comprehensive review. Clin Rev Allergy Immunol.
51:27–47. 2016. View Article : Google Scholar :
|
|
9
|
Zhang W, Wu X, Li W, Zhang H, Wang Y, Xu
J, Li W, Qin Y, Wu Z, Ge G, et al: Pinosylvin inhibits inflammatory
and osteoclastogenesis via NLRP3 inflammasome. Adv Sci (Weinh).
12:e015322025. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Costa AG, Cusano NE, Silva BC, Cremers S
and Bilezikian JP: Cathepsin K: Its skeletal actions and role as a
therapeutic target in osteoporosis. Nat Rev Rheumatol. 7:447–456.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Veis DJ and O'Brien CA: Osteoclasts,
master sculptors of bone. Annu Rev Pathol. 18:257–281. 2023.
View Article : Google Scholar
|
|
12
|
Riegger J, Schoppa A, Ruths L,
Haffner-Luntzer M and Ignatius A: Oxidative stress as a key
modulator of cell fate decision in osteoarthritis and osteoporosis:
A narrative review. Cell Mol Biol Lett. 28:762023. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Iantomasi T, Romagnoli C, Palmini G,
Donati S, Falsetti I, Miglietta F, Aurilia C, Marini F, Giusti F
and Brandi M: Oxidative stress and inflammation in osteoporosis:
Molecular mechanisms involved and the relationship with microRNAs.
Int J Mol Sci. 24:37722023. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Akune T, Ohba S, Kamekura S, Yamaguchi M,
Chung UI, Kubota N, Terauchi Y, Harada Y, Azuma Y, Nakamura K, et
al: PPARgamma insufficiency enhances osteogenesis through
osteoblast formation from bone marrow progenitors. J Clin Invest.
113:846–855. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ahmadian M, Suh JM, Hah N, Liddle C,
Atkins AR, Downes M and Evans RM: PPARgamma signaling and
metabolism: The good, the bad and the future. Nat Med. 19:557–566.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tsai YS and Maeda N: PPARgamma: A critical
determinant of body fat distribution in humans and mice. Trends
Cardiovasc Med. 15:81–85. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tontonoz P and Spiegelman BM: Fat and
beyond: The diverse biology of PPARgamma. Annu Rev Biochem.
77:289–312. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu C, Xiong Q, Li Q, Lin W, Jiang S,
Zhang D, Wang Y, Duan X, Gong P and Kang N: CHD7 regulates bone-fat
balance by suppressing PPAR-gamma signaling. Nat Commun.
13:19892022. View Article : Google Scholar
|
|
19
|
Zhang YW, Song PR, Wang SC, Liu H, Shi ZM
and Su JC: Diets intervene osteoporosis via gut-bone axis. Gut
Microbes. 16:22954322024. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang YW, Wu Y, Liu XF, Chen X and Su JC:
Targeting the gut microbiota-related metabolites for osteoporosis:
The inextricable connection of gut-bone axis. Ageing Res Rev.
94:1021962024. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Al Saedi A, Sharma S, Summers MA, Nurgali
K and Duque G: The multiple faces of tryptophan in bone biology.
Exp Gerontol. 129:1107782020. View Article : Google Scholar
|
|
22
|
Xiang T, Yang C, Xie L, Xiao S, Tang Y,
Huang G, Sun D, Chen Y and Luo F: Aberrant tryptophan metabolism
manipulates osteochondral homeostasis. Research (Wash D C).
8:07282025.PubMed/NCBI
|
|
23
|
Miao H, Zhang SJ, Wu X, Li P and Zhao YY:
Tryptophan metabolism as a target in gut microbiota, ageing and
kidney disease. Int J Biol Sci. 21:4374–4387. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen Y, Yang C, Deng Z, Xiang T, Ni Q, Xu
J, Sun D and Luo F: Gut microbially produced tryptophan metabolite
melatonin ameliorates osteoporosis via modulating SCFA and TMAO
metabolism. J Pineal Res. 76:e129542024. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Su S and Tian L: Association between
dietary tryptophan intake and bone health: A cross-sectional study.
Calcif Tissue Int. 116:62024. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xu H, Luo Y, An Y and Wu X: The mechanism
of action of indole-3-propionic acid on bone metabolism. Food
Funct. 16:406–421. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kim CS, Jung S, Hwang GS and Shin DM: Gut
microbiota indole-3-propionic acid mediates neuroprotective effect
of probiotic consumption in healthy elderly: A randomized,
double-blind, placebo-controlled, multicenter trial and in vitro
study. Clin Nutr. 42:1025–1033. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Anaya JM, Bollag WB, Hamrick MW and Isales
CM: The role of tryptophan metabolites in musculoskeletal stem cell
aging. Int J Mol Sci. 21:66702020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li J, Zhang L, Wu T, Li Y, Zhou X and Ruan
Z: Indole-3-propionic acid improved the intestinal barrier by
enhancing epithelial barrier and mucus barrier. J Agric Food Chem.
69:1487–1495. 2021. View Article : Google Scholar
|
|
30
|
Zeng Y, Guo M, Wu Q, Tan X, Jiang C, Teng
F, Chen J, Zhang F, Ma X, Li X, et al: Gut microbiota-derived
indole-3-propionic acid alleviates diabetic kidney disease through
its mitochondrial protective effect via reducing ubiquitination
mediated-degradation of SIRT1. J Adv Res. 73:607–630. 2025.
View Article : Google Scholar :
|
|
31
|
Zhao ZH, Xin FZ, Xue Y, Hu Z, Han Y, Ma F,
Zhou D, Liu XL, Cui A, Liu Z, et al: Indole-3-propionic acid
inhibits gut dysbiosis and endotoxin leakage to attenuate
steatohepatitis in rats. Exp Mol Med. 51:1–14. 2019.
|
|
32
|
Wu R, Kong Y, Li J, Chen H, Jiao Y, Sun C
and Ju Y: Indole-3 propionate inhibits NF-kappaB/NLRP3-mediated
osteoclastogenesis and improves bone quality in high-fat-diet
induced obese mice. Biochim Biophys Acta Mol Basis Dis.
1871:1679522025. View Article : Google Scholar
|
|
33
|
Bai J, Si G, Wang R, Su S, Fan J, He X, Lv
Y, Gao S and Zhou F: Gut metabolite indoleacrylic acid suppresses
osteoclast formation by AHR mediated NF-κB signaling pathway. Int J
Biol Sci. 22:951–969. 2026. View Article : Google Scholar
|
|
34
|
Bai J, Han G, Fan J, Wang R, Su S, Sun A,
Hu D, Lv Y, Gao S and Zhou F: Gut microbial metabolite alleviates
osteoporosis by attenuating AKT-NFATc1 signaling pathway and ROS
production. Free Radic Biol Med. 243:351–366. 2026. View Article : Google Scholar
|
|
35
|
Peng R, Song C, Gou S, Liu H, Kang H, Dong
Y, Xu Y, Hu P, Cai K, Feng Q, et al: Gut Clostridium
sporogenes-derived indole propionic acid suppresses osteoclast
formation by activating pregnane X receptor. Pharmacol Res.
202:1071212024. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chen C, Cao Z, Lei H, Zhang C, Wu M, Huang
S, Li X, Xie D, Liu M, Zhang L and Chen G: Microbial tryptophan
metabolites ameliorate Ovariectomy-induced bone loss by repairing
intestinal AhR-mediated gut-bone signaling pathway. Adv Sci
(Weinh). 11:e24045452024. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Bai J, Zhang W, Zhou C, Zhao G, Zhong H,
Hang K, Xu J, Zhang W, Chen E, Wu J, et al: MFG-E8 promotes
osteogenic differentiation of human bone marrow mesenchymal stem
cells through GSK3β/β-catenin signaling pathway. FASEB J.
37:e229502023. View Article : Google Scholar
|
|
38
|
Wu X, Wang K, Chen H, Cao B, Wang Y, Wang
Z, Dai C, Yao M, Ji X, Jiang X, et al: Hypoxia-induced
mitochondrial fission regulates the fate of bone marrow mesenchymal
stem cells by maintaining HIF1α stabilization. Free Radic Biol Med.
225:127–144. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jurisic V, Srdic-Rajic T, Konjevic G,
Bogdanovic G and Colic M: TNF-α induced apoptosis is accompanied
with rapid CD30 and slower CD45 shedding from K-562 cells. J Membr
Biol. 239:115–122. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Liu Y, Yang X, Gan J, Chen S, Xiao ZX and
Cao Y: CB-Dock2: Improved protein-ligand blind docking by
integrating cavity detection, docking and homologous template
fitting. Nucleic Acids Res. 50:W159–W164. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Li D, Zhao Z, Zhu L, Feng H, Song J, Fu J,
Li J, Chen Z and Fu H: 7,8-DHF inhibits BMSC oxidative stress via
the TRKB/PI3K/AKT/NRF2 pathway to improve symptoms of
postmenopausal osteoporosis. Free Radic Biol Med. 223:413–429.
2024. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chen M, Liang H, Wu M, Ge H, Ma Y, Shen Y,
Lu S, Shen C, Zhang H, Wang Z and Tang L: Fgf9 regulates bone
marrow mesenchymal stem cell fate and bone-fat balance in
osteoporosis by PI3K/AKT/Hippo and MEK/ERK signaling. Int J Biol
Sci. 20:3461–3479. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Yang L, Liu X, Chen S, Sun J, Tao Y, Ma L,
Zeng Y, Luo K, Tian R and Meng X: Scutellarin ameliorates
mitochondrial dysfunction and apoptosis in OGD/R-insulted HT22
cells through mitophagy induction. Biomed Pharmacother.
179:1173402024. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zhao J, Bai X, Du J, Chen Y, Guo X, Zhang
J, Gan J, Wu P, Chen S, Zhang X, et al: Tryptophan metabolism: From
physiological functions to key roles and therapeutic targets in
cancer (Review). Oncol Rep. 54:862025. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Li Y, Jin D, Xie W, Wen L, Chen W, Xu J,
Ding J and Ren D: PPAR-γ and wnt regulate the differentiation of
MSCs into adipocytes and osteoblasts respectively. Curr Stem Cell
Res Ther. 13:185–192. 2018. View Article : Google Scholar
|
|
46
|
Kim K, Kim JH, Kim I, Seong S, Koh JT and
Kim N: Sestrin2 inhibits RANKL-induced osteoclastogenesis through
AMPK activation and ROS inhibition. Free Radic Biol Med. 211:77–88.
2024. View Article : Google Scholar
|
|
47
|
Ye W, Liao Y, Liu X, Wang Y, Li T, Zhao Y,
He Z, Chen J, Yin M, Sheng Y, et al: Dectin-2 depletion alleviates
osteoclast-induced bone loss in periodontitis via Syk/NOX2/ROS
signaling. Free Radic Biol Med. 229:13–29. 2025. View Article : Google Scholar : PubMed/NCBI
|