Introduction
Pancreatic carcinoma (PaCa) is one of the most
lethal solid malignancies and the fourth leading cause of
cancer-related mortality in North America, where over 38,000 cases
are diagnosed annually, with a similar number of patients dying
from the disease (1,2). Pancreatic ductal adenocarcinoma
(PDAC) accounts for more than 95% of PaCa (3). It is characterized by a rapid disease
progression and absence of specific symptoms, largely precluding an
early diagnosis and curative treatment, and is associated with a
very poor prognosis (4). Despite
progress in chemotherapy, radiation therapy, immunotherapy and
surgery over the past several years, the overall 5-year relative
survival rate is less than 5% (5).
Gemcitabine has been the standard systemic therapy for the
palliative treatment of PaCa over the last decade, although the
1-year survival rates ranging at approximately 18% remain
unsatisfactory (6–8). The comparably low rate of
side-effects of gemcitabine treatment makes additional treatment
options feasible (9) and a
substantial number of potential drug combinations have been tested.
However, no convincing results on clinically relevant improvements
in the quality of life and survival have been found. Thus, it is
impartial to state that there are still no evidence-based treatment
options for gemcitabine-refractory advanced PaCa, and effective
additional agents to the first-line gemcitabine treatment have not
yet to be found (10). Therefore,
it is imperative to provide an effective means by which to enhance
the sensitivity of PaCa cells to gemcitabine in order to increase
the curative effect of gemcitabine, and thus improve patient
outcomes.
The concept of using a combination of agents for
cancer chemoprevention has recently received much attention.
Considerable evidence from laboratory studies suggests that
combinations of chemopreventive agents may be more effective for
the prevention of cancer than any single constituent. Recent
studies show that the traditional Chinese medicine, curcumin
(11), and drugs such as
resveratrol (12), thymoquinone
(13) and emodin (14–16)
used in combination with gemcitabine can significantly inhibit
malignant tumor cell proliferation.
Oridonin is a diterpenoid purified from Rabdosia
rubescens and has been reported to exert various
pharmacological and physiological effects, such as
anti-inflammatory, antibacterial, and antitumor effects (17). As regards its antitumor activity,
certain reports have demonstrated that oridonin exhibits remarkable
inhibitory effects on hepatocellular carcinoma, human osteosarcoma
cells, U937 human macrophage-like cells, A431 human epidermoid
carcinoma cells, non-small cell lung cancers, acute promyelocytic
leukemia, as well as glioblastoma multiforme (18–22),
and others have also shown that oridonin has cytotoxic effects on
various cancer cells, such as HeLa human cervical carcinoma, MCF-7
human breast adenocarcinoma, L929 murine fibrosarcoma and
colorectal cancer cells (23–26).
These functions have been identified through the regulation of
tumor cell proliferation and apoptosis-related genes. Apoptosis can
be triggered in many ways, and it is well known that many proteins,
such as cysteine-dependent aspartate-specific proteases (caspase)
family, the Bcl-2 family, the mitogen-activated protein kinase
(MAPK) family, p53 and phosphoinositide 3-kinase (PI3K) signal
transduction pathways play important roles in regulating the
apoptotic process (27–29). MAPKs have been shown to
phosphorylate p53 in response to a variety of stressful stimuli,
leading to p53-mediated cellular responses, causing cell cycle
arrest and apoptosis (30).
Currently, the involvement of p38 and p53 in the attenuation of the
antitumor effect of oridonin in combination with gemcitabine in the
treatment of PaCa has been not reported. Whether oridonin can
potentiate the effect of gemcitabine in PaCa, also remains
unknown.
Therefore, in this study, we investigated the effect
of oridonin on the growth of human PaCa cells in culture and in a
subcutaneous mouse model of PaCa, and examined the effect of
oridonin in combination with gemcitabine both in vitro and
in vivo. We also investigated the role of p38 and p53 in the
induction of apoptosis in the BxPC-3 human PaCa cell line. We
demonstrate that oridonin can potentiate the therapeutic effects of
gemcitabine against PaCa cells in vitro and in vivo.
We show that the upregulation of p38 and p53 by oridonin is one of
the mechanisms of the inhibition of tumor growth of PaCa. Our
results can lead to an improvement in the treatment of PaCa.
Materials and methods
Chemicals and reagents
Oridonin was obtained from the Beijing Institute of
Biological Products (Beijing, China). The purity of oridonin was
measured by high-performance liquid chromatography (HPLC) and
determined to be 99.4%. Oridonin was dissolved in dimethyl
sulfoxide (DMSO) to create a stock solution at a concentration of
10 mmol/l and stored at −20°C. The DMSO concentration was kept
below 0.1% in all the cell cultures and did not exert any
detectable effect on cell growth or cell death. Gemcitabine
(Gemzar) from Eli Lilly was stored at 4°C and dissolved in sterile
phosphate-buffered saline (PBS) at a concentration of 0.2 mmol/l on
the day of use. Fetal bovine serum (FBS), trypsin-EDTA, Roswell
Park Memorial Institute-1640 (RPMI-1640) and the Cell Counting
kit-8 (CCK-8) were obtained from Gibco, the Annexin V-FITC/PI
apoptosis detection kit was from Biological Development Co, Ltd.
Nanjing KGI, the RNA extraction kit was from Life Technologies Co.,
the cDNA first-strand synthesis kit was from Fermentas and the 2X
Taq PCR MasterMix was from Tiangen. Ribonuclease A (RNase A),
propidium iodide (PI) and DMSO were obtained from Sigma. Antibodies
against p53, phosphorylated p53 (P-p53), p38, phosphorylated p38
(P-p38), β-actin and horseradish peroxidase (HRP)-conjugated
secondary antibodies (goat-anti-rabbit and goat-anti-mouse) were
purchased from Sigma.
Cell line and culture
The PaCa cell line, BxPC-3, was obtained from the
American Type Culture Collection (ATCC; Manassas, VA, USA). All
cells were cultured in RPMI-1640 medium supplemented with 10% FBS,
100 U/ml penicillin and 100 μg/ml streptomycin. Cells were
maintained at 37°C in a humidified atmosphere of 5% CO2.
The medium was changed every 2–3 days, and the cells were
subcultured when confluency reached 70–80% by 0.25% trypsin at
37°C.
Cell proliferation assay
Cell survival was determined using CCK-8. Briefly,
the logarithmic phase PaCa cells were plated in 96-well culture
plates (7×103 cells per well). After 24 h of incubation,
the cells were treated with the vehicle alone (0.1% DMSO) and
various concentrations (20, 40, 60, 80 and 100 μM) of
oridonin, followed by 24, 48 and 72 h of cell culture. Each group
had 6 wells. A total of 10 μl CCK-8 was added to each well 1
h before the end of incubation. The absorbance at 450 nm was read
using the Bio-Tek ELx800 Absorbance Microplate Reader. The
experiment was repeated 3 times. The degree of cellular inhibition
by each drug was calculated by the following formula: relative %
inhibition = 1- (dosing absorbance - blank absorbance)/(control
absorbance - blank absorbance) ×100%.
Annexin V apoptosis assay
Apoptosis induction was assessed using the Annexin
V-FITC kit according to the manufacturer’s instructions. BxPC-3
PaCa cells were exposed to the control (DMSO-treated), the desired
concentration of oridonin (40 μM) and gemcitabine (20
μM) alone or in combination with oridonin for 48 h, and the
floating and adherent cells were collected by centrifugation at
1,000 x g for 5 min. Pooled cells were washed with the binding
buffer supplied by the manufacturer. Approximately 5×104
cells were suspended in 200 μl of PBS, and mixed with 5 ml
of Annexin V-FITC and 10 ml of PI. After 15 min of incubation in
the dark, the fluorescence intensities of oridonin of >10,000
viable cells from each cell sample were analyzed by using a Coulter
Epics XL flow cytometer with excitation and emission settings of
488 and 525 nm, respectively. The data were analyzed using
CellQuest software.
Cell cycle distribution by flow
cytometry
The effect of oridonin on cell cycle distribution
was determined by flow cytometry analysis of the DNA content in the
cell nuclei following staining with PI. Asynchronized (70–80%)
confluent cells were treated with 40 μM oridonin for 48 h.
The control cells were treated with 0.1% DMSO only. After
incubation at 37°C for a specified time, the floating and adherent
cells were collected by using 0.05% trypsin, washed twice with cold
PBS and then fixed with ice-cold 70% ethanol overnight at 4°C. The
fixed cells were subsequently centrifuged at 300 x g for 10 min and
the pellets were washed with PBS. The cells were then treated with
80 mg/ml RNase A for 30 min at 37°C. The cells were chilled over
ice for 10 min and stained with PI (50 mg/ml final concentration)
for 1 h in the dark. The stained cells were analyzed using a
Coulter Epics XL flow cytometer. Approximately 20,000 cells were
evaluated for each experiment. In all the determinations, cell
debris and clumps were excluded from the analysis. The cell cycle
data were reanalyzed using ModFit software.
Measurements of p38 and p53 mRNA by
semi-quantitative RT-PCR assay
PaCa cells were treated with the assigned
concentration oridonin (40 μM) and gemcitabine (20
μM) alone or in combination for 48 h. The control cells were
treated with 0.1% DMSO only. Subsequently, total cellular RNAs were
isolated from the cells with TRIzol reagent, and the RNA content
was measured using a UV spectrophotometer under 260 nm. cDNA was
synthesized with 1 μg of total RNA and oligo(dT) primer
according to the manufacturer’s instructions. PCR amplification
conditions were as follows: p38, 94°C 60 sec, 61.8°C 60 sec, 72°C
60 sec, 35 cycles; p53, 94°C 45 sec, 51.9°C 1 min, 72°C 90 sec, 35
cycles; GAPDH, 94°C 45 sec, 58°C 1 min, 72°C 1 min, 35 cycles.
GAPDH was used as the internal control. The primer pairs used for
the amplification are listed in Table
I. The PCR product (5 μl) was analyzed by 1% agarose gel
electrophoresis and the results were photographed.
 | Table IPrimer pairs used in
semi-quantitative PCR analysis. |
Table I
Primer pairs used in
semi-quantitative PCR analysis.
| Genes | Primer pairs
(5′→3′) | Product size
(bp) |
|---|
| p38 | | |
| Sense |
CGGAGTGGCATGAAGCTGTAG | 346 |
| Antisense |
CCCTAGGAAACCAACACAGCA | |
| p53 | | |
| Sense |
TCTGGGACAGCCAAGTCTGT | 435 |
| Antisense |
GGAGTCTTCCAGTGTGATGA | |
| GAPDH | | |
| Sense |
CGCTGCGCTGGTCGTCGACA | 619 |
| Antisense |
GTCACGCACGATTTCCCGCT | |
Western blot analysis in vitro
Briefly, PaCa cells were incubated with the
allocated concentration of oridonin (40 μM) and gemcitabine
(20 μM) alone or in combination for 48 h; the cells in
medium alone were used as the controls. Subsequently, the cells
were harvested, washed twice with ice-cold PBS and then the cell
pellets were resuspended in lysis buffer consisting of 50 mM HEPES
(pH 7.4), 1% Triton-X 100, 2 mM sodium orthovanadate, 100 mM sodium
fluoride, 1 mM edetic acid, 1 mM PMSF, 10 mg/l aprotinin (Sigma),
and 10 mg/l leupeptin (Sigma) and lysed at 4°C for 60 min. After
centrifugation at 13,000 × g for 15 min, the protein content of the
supernatant was determined by the bicinchoninic acid (BCA) assay
kit (Sigma) according to the manufacturer’s instructions. The
protein lysates (20 μg/lane) were separated by
electrophoresis on 12% SDS polyacrylamide gel and blotted onto a
nitrocellulose membrane. Each membrane was blocked with 5% skim
milk and then incubated with the indicated primary antibodies
against p53, P-p53, p38, P-p38 and β-actin overnight at 4°C.
Subsequently, the membrane was incubated with the secondary
antibodies, goat anti-rabbit and goat anti-mouse IgG conjugated
with HRP, for 1 h at room temperature and the formed immunocomplex
was visualized by enhanced chemiluminescence reagent and exposed to
X-ray films. Quantitative data are expressed as the means ± SD of
the relative levels of the objective protein and control β-actin of
each group of cells from 3 independent experiments.
Experimental animals
Female nude mice [4–6 weeks old, BALB/cA-nu (nu/nu),
weight 18–20 g] were purchased from the Shanghai Cancer Institute
for Tumor Implantation and maintained in a specific-pathogen-free
environment at the Wenzhou Medical College Experimental Animal
Center. The animals were kept in a controlled environment with a
temperature of 25±1°C and relative humidity of 40–60%. Ethical
approval for this study was given by the Ethics Committee at
Wenzhou Medical College.
Model establishment and experimental
scheme
BxPC-3 cells in the log-phase were suspended with
serum-free culture medium (5×106 cells in 200
μl), and tumor xenografts were established by a subcutaneous
inoculation of BxPC-3 PaCa cells into the right abdominal flanks of
the nude mice. Tumors were allowed to develop for 4–5 weeks until
they reached a size of 100–150 mm3, and then treatment
was initiated. All the mice were randomly divided into 4 groups and
treated intraperitoneally (IP) with saline, gemcitabine (80 mg/kg)
alone, oridonin (40 mg/kg) alone, or gemcitabine (80 mg/kg) and
oridonin (40 mg/kg) in combination every 3 days for up to 30 days
post-implantation in a volume of 0.2 ml (n=10 per group). Tumor
volume was measured using calipers and estimated according to the
formula: tumor volume (mm3) = L × W2/2, where
L is the length and W is the width. Two weeks after the last
treatment, the mice were sacrificed and the tumors were excised.
Xenograft tumors of the treated and control mice, were harvested
and fixed in 4% formalin, embedded in paraffin, and cut into
suitable sections for western blot and RT-PCR analysis.
Western blot analysis in tumor
tissues
Protein was routinely extracted from tumor tissues
using radioimmunoprecipitation assay (RIPA) buffer. Total protein
concentration was measured using the BCA assay kit (Sigma) with
bovine serum albumin as the standard, according to the
manufacturer’s instructions. Tumor tissue extracts containing 80
μg total protein were subjected to 12% SDS/PAGE, and the
resolved proteins were transferred electrophoretically to
polyvinylidene fluoride (PVDF) membranes. After being blocked with
5% fat-free milk, the membranes were probed with individual primary
antibodies overnight at 4°C and the bound antibodies were detected
with HRP-conjugated goat anti-rabbit IgG for 1 h. The formed
immunocomplex was visualized by enhanced chemiluminescence reagent
(ECL; Pierce).
RT-PCR detection of p38 and p53 mRNA in
tumor tissues
Total RNA was extracted from the tumor tissues using
the TRIzol reagent. The amount and purity of the extracted RNA was
quantified using spectrophotometry. The value of
A260/A280 was measured to evaluate the
quality of the RNA. cDNA was synthesized with 5 μg total RNA
and oligo(dT) primer according to the manufacturer’s instructions.
The PCR amplification reaction was amplified using the GeneAmp PCR
System 9600 (Perkin-Elmer Corp. Norwalk, CT, USA). The
amplification conditions and the primer pairs used for the
amplification were the same as those used for the in vitro
analysis. The PCR products (5 μl) were analyzed by
electrophoresis on 1% agarose gel with ultraviolet (UV)
illumination and the results were photographed.
Statistical analysis of the data
All results were confirmed by at least 3 separate
experiments. The data are expressed as the means ± SD. Statistical
comparisons were performed by one-way ANOVA and the Student’s
t-test for differences between two sample means using the SPSS
v17.0 software. A P-value <0.05 was considered to indicate
statistically significant differences.
Results
The goal of the present study was to determine
whether and how oridonin can be effectively used for the treatment
of PaCa either alone or in combination with gemcitabine and if so,
to clarify the mechanisms behind its therapeutic action. For this
purpose, we selected the PaCa cell line, BxPC-3. In order to
facilitate the imaging of the tumors in the animals, BxPC-3 cells
were also used in the subcutaneous transplant model in mice.
Oridonin suppresses the proliferation and
potentiates the apoptotic effects of gemcitabine in human PaCa
cells in vitro
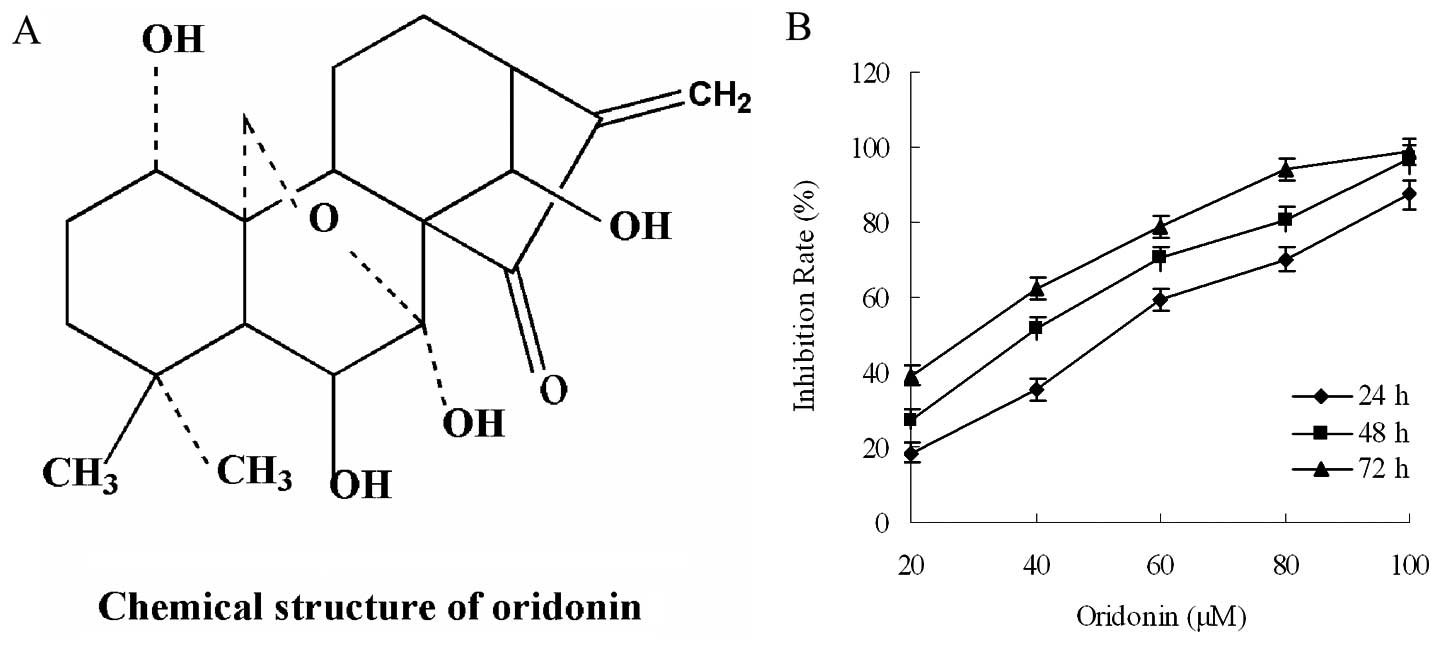
We first investigated the effect of oridonin on the
proliferation of the PaCa cell line, BxPC-3. Oridonin inhibited the
growth of PaCa cells (BxPC-3) in a dose- and time-dependent manner
(Fig. 1). These tumor cells showed
sensitivity to the oridonin treatment. The half maximal inhibitory
concentration (IC50) value of oridonin was determined as
38.86 μM for the BxPC-3 cells following treatment for 48 h.
Oridonin was used at the dose of 40 μM.
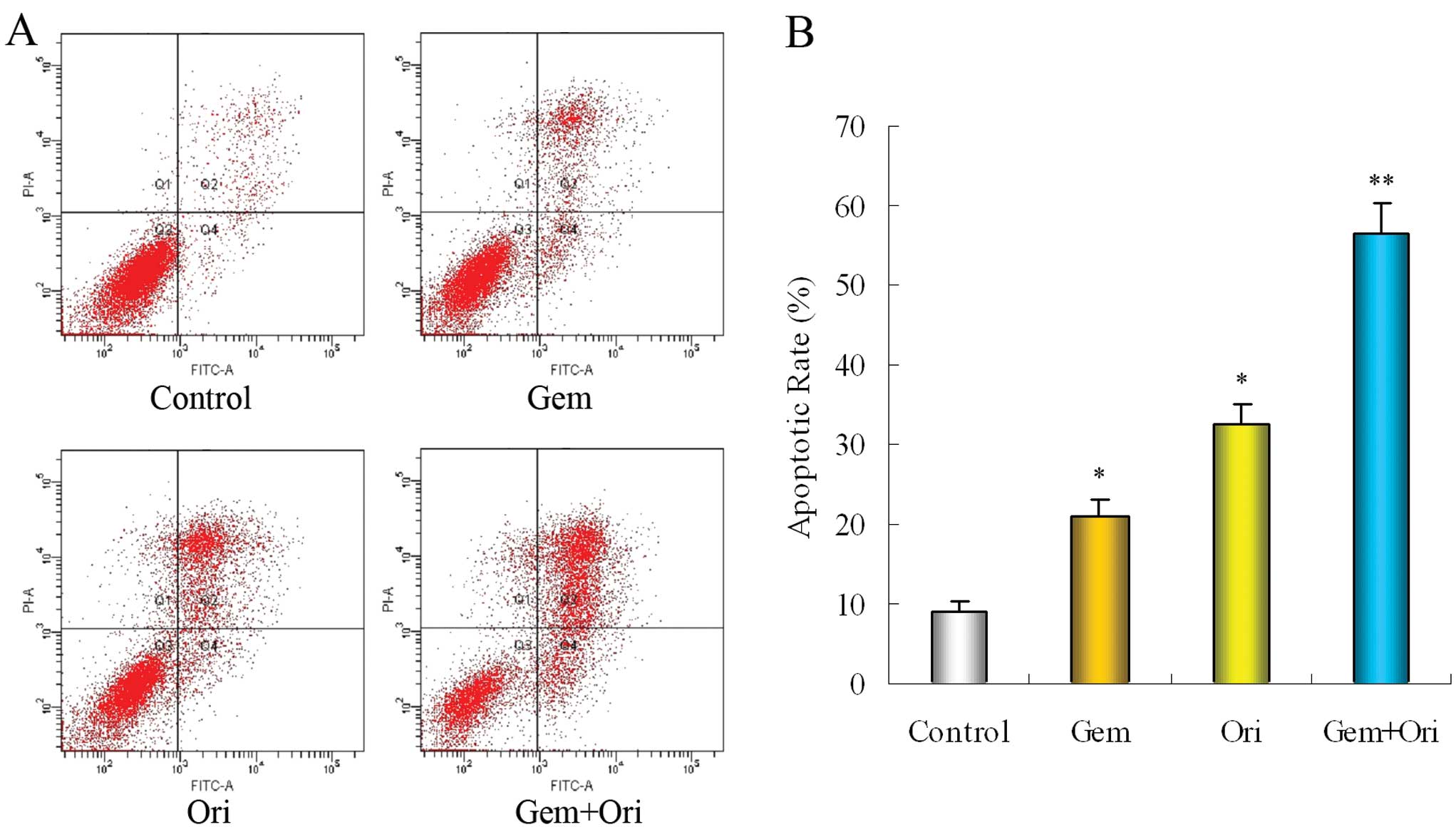
To determine whether oridonin enhances the induction
of apoptosis by gemcitabine, we investigated the occurrence of
apoptosis in the cell lines. Flow cytometry analysis was performed
using Annexin V-FITC/PI-stained PaCa cells. The cells were cultured
with oridonin and gemcitabine, alone or in combination for 48 h.
The results showed that, at a dose at which oridonin and
gemcitabine alone were minimally effective, the combination of both
was highly effective (Fig. 2).
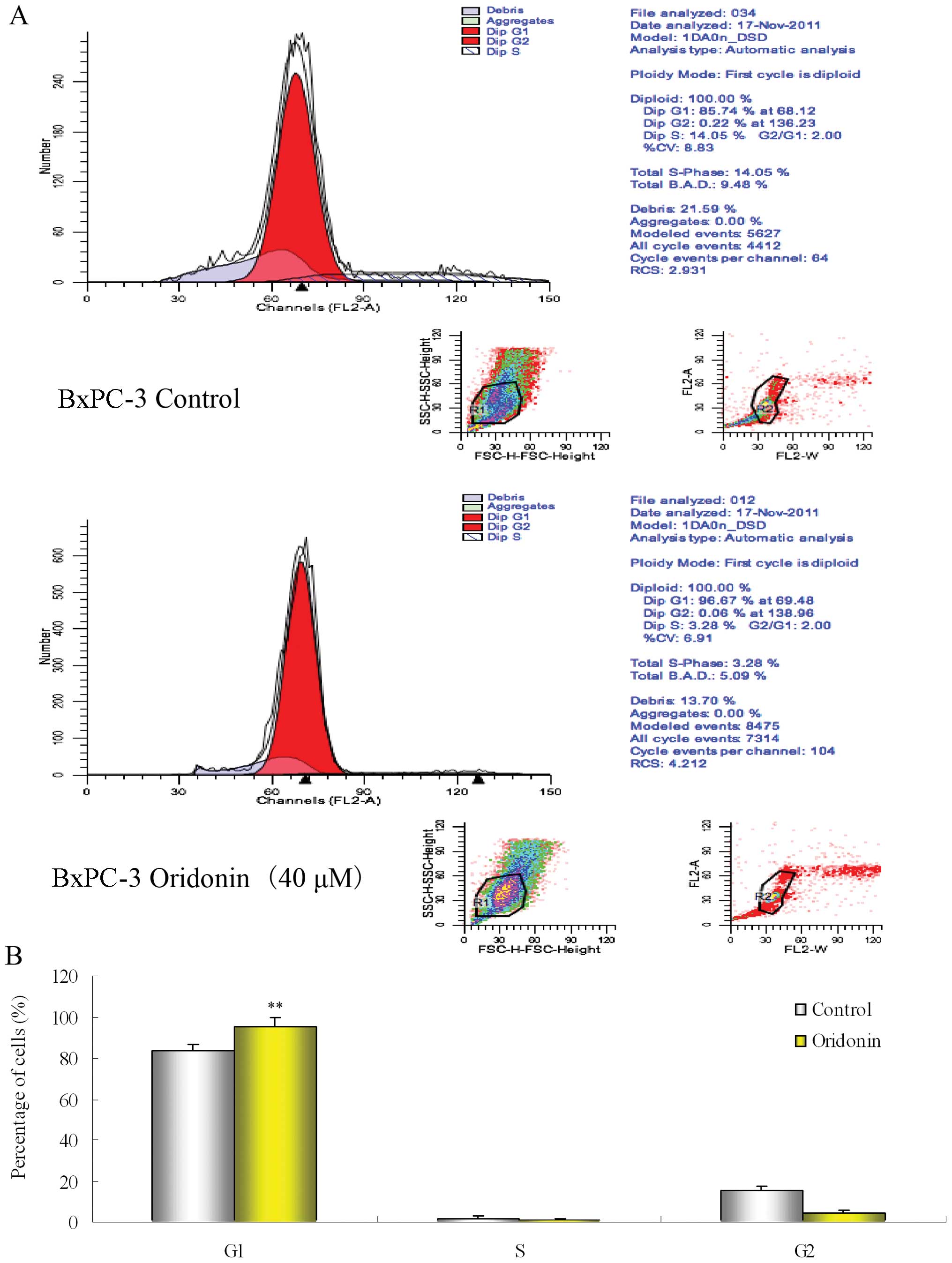
We also investigated the mechanisms by which
oridonin potentiates the effects of gemcitabine in these cells. The
DNA content of the PaCa cells treated for 48 h with 40 μM
oridonin or the vehicle (0.1% DMSO) was analyzed using a flow
cytometer. Fig. 3 shows that there
were marked and consistent changes in the cell cycle at 48 h. The
number of cells in the G1 phase increased significantly
with a concomitant decrease in the number of treated cells in the S
and G2 phase when compared with the control. These
results suggest that oridonin treatment suppresses the growth of
PaCa cells, and synergizes the apoptotic effects of
gemcitabine.
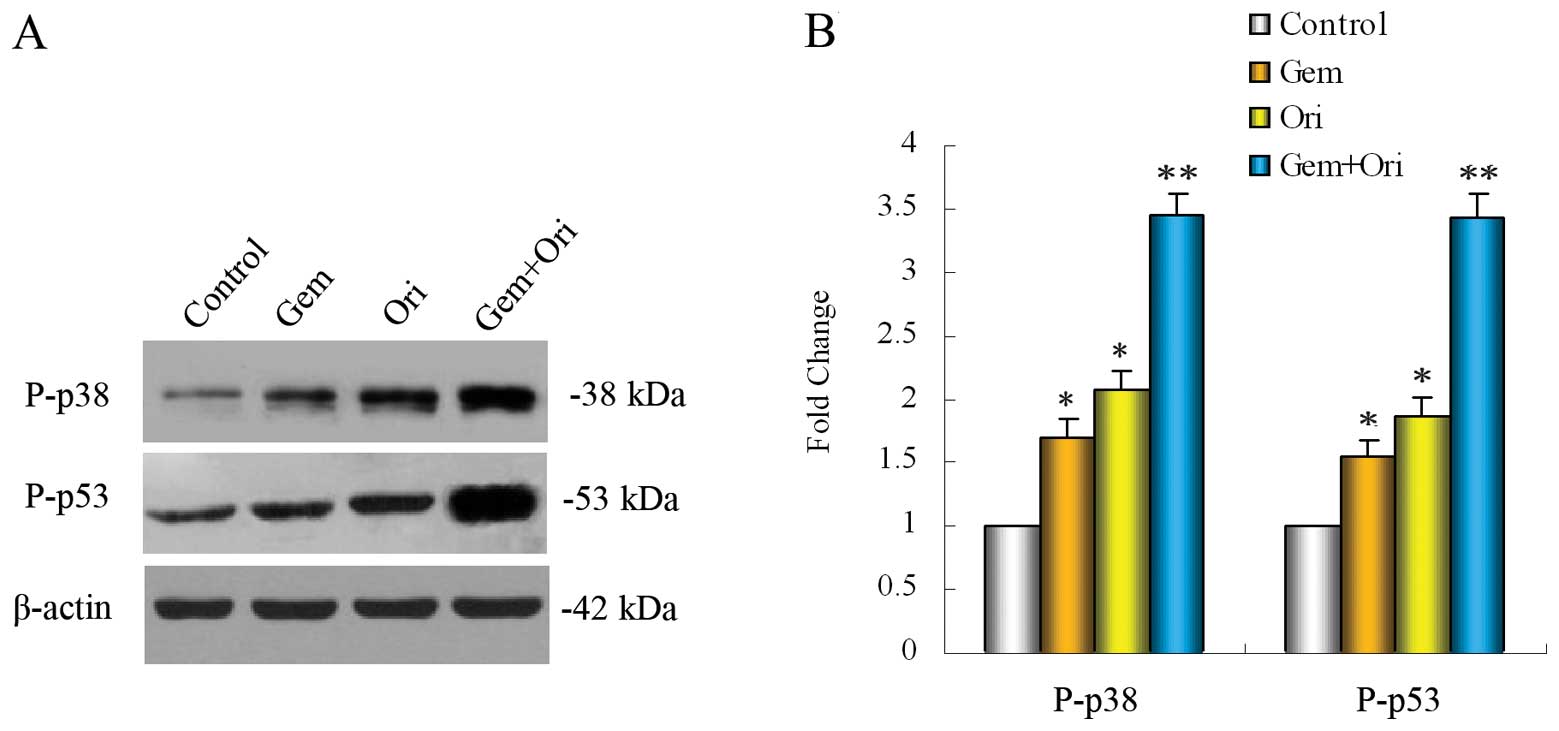
Effect of oridonin on the
gemcitabine-induced expression of P-p38 and P-p53 in vitro
The tumor suppressor gene product, p53, has been
reported to mediate apoptosis in a number of experimental systems.
The MAPK family members, including the extracellular
signal-regulated protein kinase (ERK), c-Jun N-terminal kinase
(JNK) and p38, play important roles in the regulation of apoptosis
(30). Therefore, in order to
confirm whether MAPK-p38 and its downstream p53 protein are
involved in the anti-cancer effects of oridonin, western blot
analysis was carried out. After treatment of the PaCa cells with
oridonin and gemcitabine alone or combination for 48 h, the results
showed that both oridonin (P<0.05) and gemcitabine (P<0.05)
alone significantly increased the level of P-p38 and P-p53
(Fig. 4), as compared with that of
the control cells, and that the combination of both agents was even
more effective (P<0.01). The unphosphorylated forms of p38 and
p53 were not altered by oridonin and gemcitabine alone or
combination treatment (data not shown). These results suggest that
oridonin can improve the p38 activation of gemcitabine in PaCa
cells, which contributes to the further activation of p53. This
could further increase PaCa cell apoptosis.
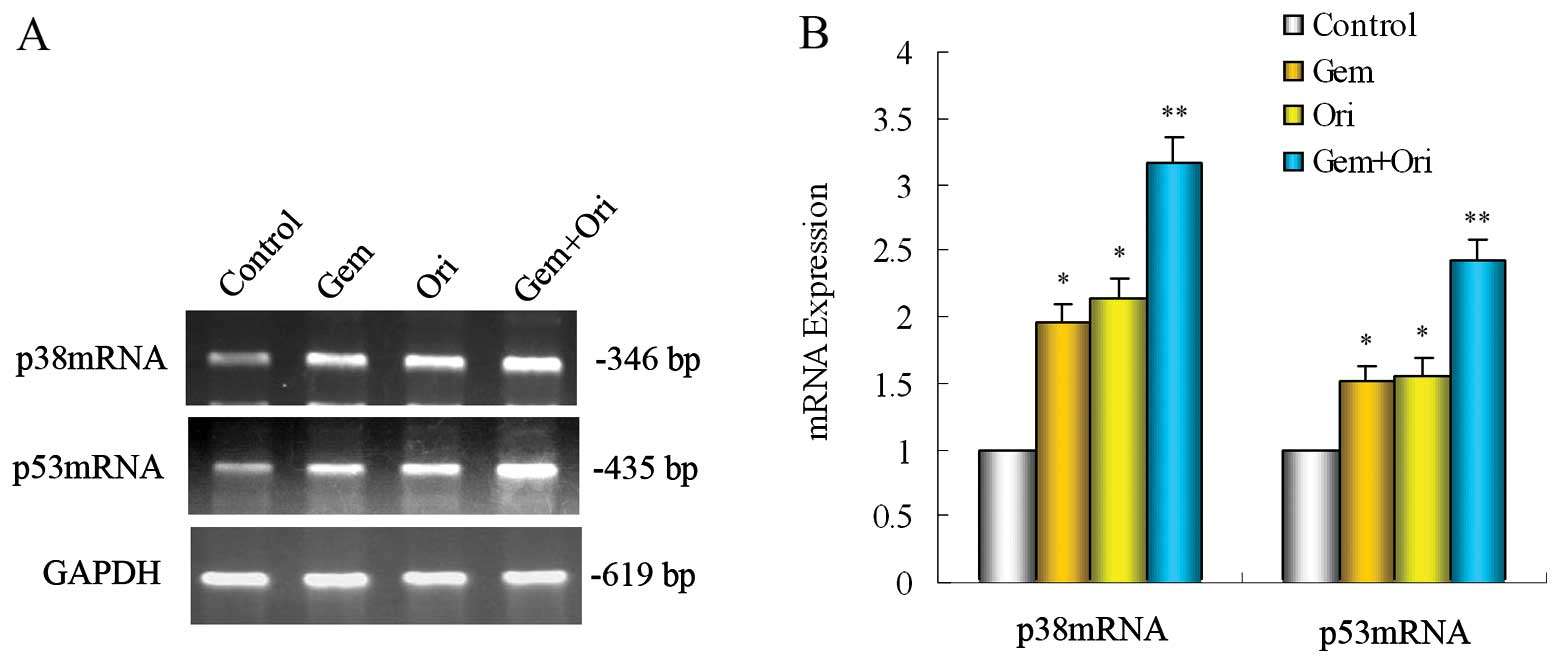
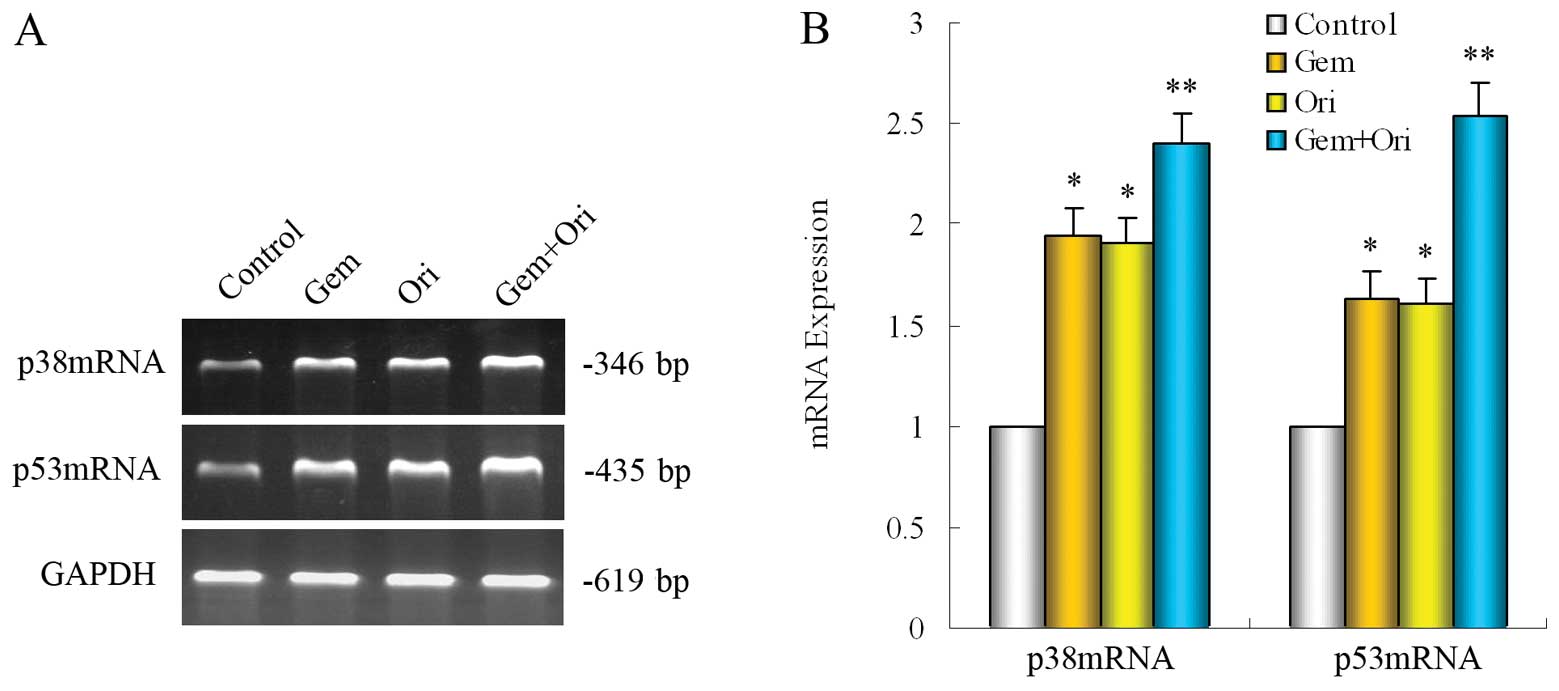
Effects of oridonin and gemcitabine on
p38 and p53 mRNA in PaCa cells
To further clarify whether the possible mechanism is
relevant with the p38 and p53 gene, RT-PCR was carried out to
detect the p38 and p53 mRNA expressions. In the PaCa cell line,
BxPC-3, when compared with the control, gemcitabine alone
significantly upregulated the expression of p38 and p53 mRNA
(P<0.05). Treatment with oridonin alone or in combination with
gemcitabine significantly upregulated the expression of p38 and p53
mRNA (P<0.05). The upregulation induced by the combined
treatment of oridonin and gemcitabine was even more evident than
the other groups (P<0.05) (Fig.
5). These results indicate that p38 and p53 participate in the
apoptosis induced by oridonin and gemcitabine in PaCa cells.
Oridonin potentiates the
anti-proliferative effects of gemcitabine in implanted PaCa cells
in nude mice
Our in vitro data prompted us to examine
whether the effects of oridonin and gemcitabine are equally
demonstrable in vivo. We examined the effects of oridonin
and gemcitabine, alone or in combination, on the growth of
subcutaneously implanted pancreatic tumors in nude mice. The
experimental protocol is depicted in Fig. 6. We found that the oral
administration of oridonin alone at 40 mg/kg significantly
inhibited tumor growth (P<0.05 when compared to the control)
(Fig. 6). Gemcitabine alone was as
effective as oridonin (P<0.05 when compared to the control,
P>0.05 when compared to the oridonin alone group), and the
combination of both agents was even more effective in reducing the
tumor burden. The tumor volume in the combination group was
significantly lower than in the oridonin alone (P<0.05) or
gemcitabine alone group (P<0.05) (Fig. 6).
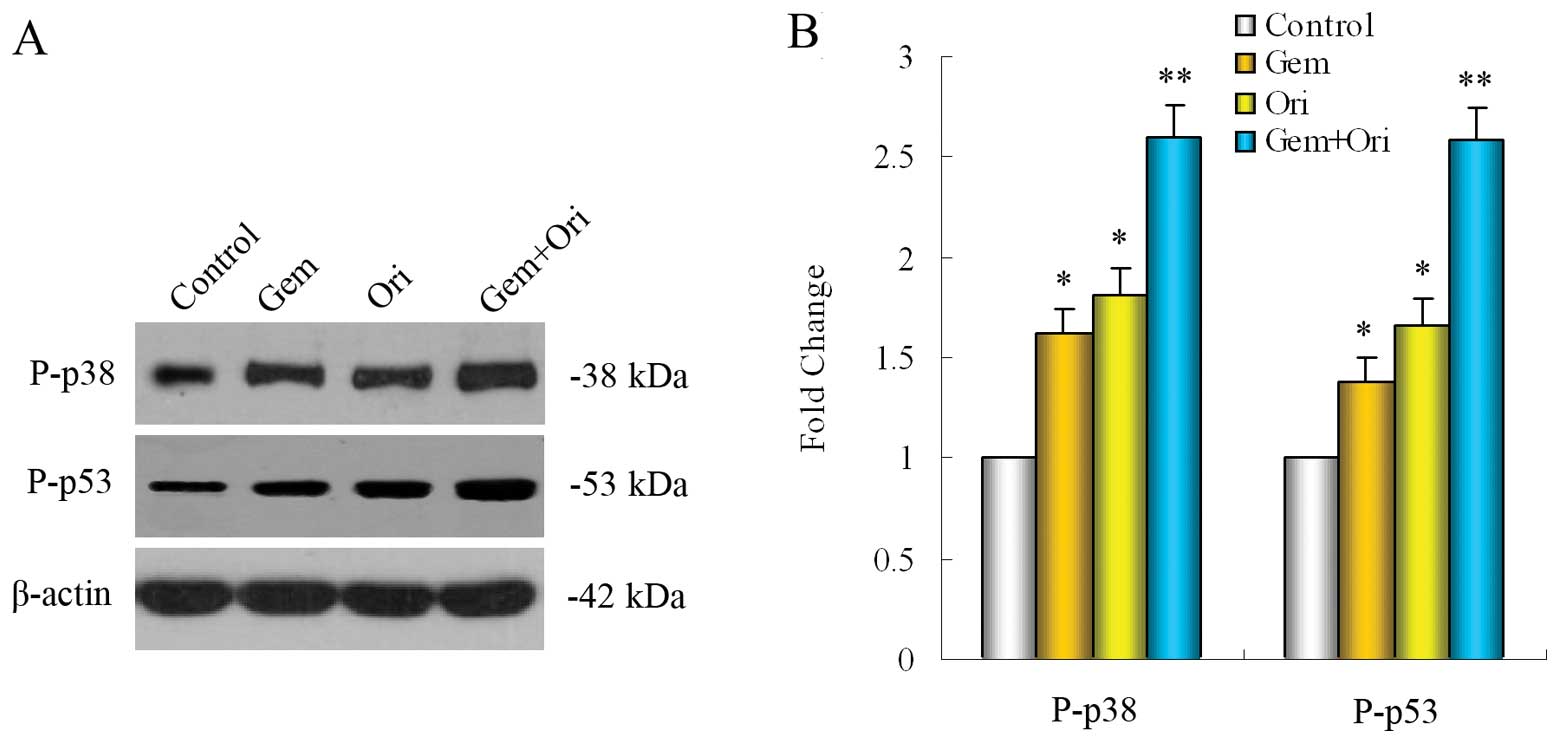
Oridonin enhances the effect of
gemcitabine via upregulating the expression of P-p38 and P-p53 in
vivo
To determine whether the impact on the tumor growth
inhibition of oridonin was related to the p38 and p53 activation,
we evaluated the effects of oridonin and gemcitabine on P-p38 and
P-p53 levels, by western blot analysis in the pancreatic tumor
tissues. The results showed that both oridonin (P<0.05) and
gemcitabine (P<0.05) alone significantly upregulated the
expression of P-p38 and P-p53 in the PaCa tissue as compared to the
control group and that the combination of both was even more
effective (P<0.05 when compared to gemcitabine alone or the
control) (Fig. 7).
Effects of oridonin on expression of p38
and p53 mRNA in vivo
In order to further confirm our conclusion in
pancreatic tumor tissues. we investigated the expression of p38 and
p53 mRNA levels in pancreatic tumor tissues. The results showed
that oridonin either alone or in combination with gemcitabine
(P<0.05 vs. the mice treated with gemcitabine or the control)
was effective in boosting the expression of p38 and p53 mRNA in
PaCa tissue (Fig. 8).
Discussion
The aim of this study was to determine whether
oridonin has potential either alone or in combination with
gemcitabine in the treatment of PaCa, one of the most lethal
cancers. Experiments using cell cultures or an animal model, showed
that oridonin alone inhibited the growth of PaCa and that the
combination of gemcitabine and oridonin significantly enhanced this
inhibition. In the PaCa cell line, BxPC-3, the data indicated that
varying concentrations of oridonin effectively inhibited the
proliferation of the cells at different times, and we found for the
first time that oridonin enhanced the apoptotic effects of
gemcitabine in cultured PaCa cells. Gemcitabine alone had a minimal
effect on apoptosis in BxPC-3 cell line. However, oridonin when
used in combination with gemcitabine, was highly effective in
inducing apoptosis. We also found oridonin that augmented the
apoptotic effects of gemcitabine, which were related to
G1 phase arrest, and the upregulation of p38 and p53.
The activation of p38 and p53 could be one of the mechanisms
invovled in the induction of apoptosis in PaCa cells.
In addition to these in vitro results, in a
subcutaneous nude mice model, we found that oridonin effectively
suppressed the growth of PaCa. In this model, oridonin was found to
be as effective as gemcitabine in inhibiting the tumor volume. When
the 2 agents were used together, maximum abrogation of tumor volume
was observed. Although our study is the first to report the effect
of oridonin alone on PaCa in a mouse model, these results are
consistent with those from previous reports showing the growth
suppressive effects of oridonin alone against colorectal cancer
(26). When we investigated the
mechanisms by which oridonin manifests its effects against PaCa in
an animal model, we found that this effect was also associated with
the activation of p38 and p53.
MAPKs are serine/threonine kinases that mediate
intracellular signaling associated with a variety of cellular
activities, including cell proliferation, differentiation,
survival, death and transformation (31,32).
The 3 main members that integrate the MAPK family in mammalian
cells are stress-activated protein kinase c-Jun NH2-terminal kinase
(JNK), stress-activated protein kinase 2 (SAPK2, p38) and the
extracellular signal-regulated protein kinases (ERK1/2, p44/p42)
(33). MAPK p38 has been shown to
be activated by cellular stress, UV radiation, growth factor
withdrawal and pro-inflammatory cytokines (34). Upon activation, p38 phosphorylates
various transcription factors, as well as the tumor suppressor, p53
(35). A number of studies have
reported the role of MAPK signaling in the regulation of apoptosis
in various cancer cells (18,19,21,36–38).
Previous studies have documented that the activation of the p38
MAPK pathway may lead to p53-induced apoptosis (39). In human osteosarcoma cells and A431
epidermoid carcinoma cells, p38 functions as an upstream kinase of
p53 phosphorylation and subsequently stabilizes and activates p53
transcriptional activity, leading to diverse cellular responses,
such as cell cycle arrest and apoptosis (30). We found that in response to
oridonin treatment, the increased expression of P-p38 was
accompanied by the upregualtion of P-p53, which suggests that the
p38 activation is involved in the G1 cell cycle arrest
and apoptosis induced by oridonin in pancreatic cells.
p53 is one of the most commonly mutated genes in
human cancers and its loss of function is believed to result in
increased genomic instability, with the subsequent acquisition of
additional oncogenic mutations (40). The targeting of p53 is important
for many cancer therapies. p53 has been shown to transactivate a
broad range of pro-apoptotic proteins from the Bcl-2 family (Bax,
the BH3-only proteins Bid, Puma and Noxa), and to downregulate
anti-apoptotic proteins from the Bcl-2 family (Bcl-2 and Bcl-xL),
as well as to induce the upregulation of proteins that localize to
the mitochondria (41). Knowledge
of the p53-dependent pathway in prostate cancer could allow the
development of selective and effective anticancer strategies
involved with the apoptotic response (42). The phosphorylation of p53 can
usually be induced at serine 15 or/and 18. In particular, the
phosphorylation of p53 at serine 15 has been reported to be a key
phosphorylation target during the p53 activation process to
apoptotic cell death (43). These
data indicate that p53 may play a crucial role in oridonin-induced
PaCa cell apoptosis.
Overall, our results suggest that oridonin has
significant potential for the treatment of PaCa and potentiates the
antitumor effects of gemcitabine by upregulating p38 and its
downstream targets, leading to the inhibition of proliferation,
angiogenesis and invasion. However, certain molecular links remain
to be clarified, such as how the p53 signaling is connected to cell
cycle arrest, and how p38 signaling acts on the p53 pathway. The
specific mechanisms involved require further study. The combination
of oridonin with gemcitabine has significant potential as an
effective therapy for PaCa that can enhance the effect of
gemcitabine and overcome chemoresistance. Based on these results,
further clinical studies are warranted to confirm our findings in
patients with PaCa.
Acknowledgements
We are grateful for funding support
from: the Administration of Traditional Chinese Medicine of
Zhengjing Province, China (grant no. 2011ZZ010) and the National
Natural Science Foundation of China (grant no. 81173606). We thank
the entire staff of the Animal Experimental Center of Wenzhou
Medical College and of the Scientific Research Platform of the
Second Affiliated Hospital of Wenzhou Medical College for their
invaluable assistance.
References
|
1.
|
Jemal A, Siegel R, Ward E, Hao Y, Xu J,
Murray T and Thun MJ: Cancer statistics, 2008. CA Cancer J Clin.
58:71–96. 2008. View Article : Google Scholar
|
|
2.
|
Canadian Cancer Society/National Cancer
Institute of Canada: Canadian Cancer Statistics 2008. Toronto: ISSN
0835-2976. 2008
|
|
3.
|
Reske SN: PET and PET-CT of malignant
tumors of the exocrine pancreas. Radiologe. 49:131–136. 2009.(In
German).
|
|
4.
|
Stathis A and Moore MJ: Advanced
pancreatic carcinoma: current treatment and future challenges. Nat
Rev Clin Oncol. 7:163–172. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
American Cancer Society: Cancer Facts and
Figures 2007 [M]. American Cancer Society; New York, NY: pp. 51–52.
2007
|
|
6.
|
Burris HA III, Moore MJ, Andersen J, et
al: Improvements in survival and clinical benefit with gemcitabine
as first-line therapy for patients with advanced pancreas cancer: a
randomized trial. J Clin Oncol. 15:2403–2413. 1997.PubMed/NCBI
|
|
7.
|
O’Reilly EM and Abou-Alfa GK: Cytotoxic
therapy for advanced pancreatic adenocarcinoma. Semin Oncol.
34:347–353. 2007.
|
|
8.
|
O’Reilly EM: Pancreatic adenocarcinoma:
new strategies for success. Gastrointest Cancer Res. 3:S11–S15.
2009.PubMed/NCBI
|
|
9.
|
Tingstedt B, Johansson P, Andersson B and
Andersson R: Predictive factors in pancreatic ductal
adenocarcinoma: role of the inflammatory response. Scand J
Gastroenterol. 42:754–759. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Kang SP and Saif MW: Optimal second-line
treatment options for gemcitabine refractory advanced pancreatic
cancer patients. Can we establish standard of care with available
data? J Oral Pathol. 9:83–90. 2008.
|
|
11.
|
Kunnumakkara AB, Guha S, Krishnan S, et
al: Curcumin potentiates antitumor activity of gemcitabine in an
orthotopic model of pancreatic cancer through suppression of
proliferation, angiogenesis, and inhibition of nuclear
factor-kappaB-regulated gene products. Cancer Res. 67:3853–3861.
2007. View Article : Google Scholar
|
|
12.
|
Harikumar KB, Kunnumakkara AB, Sethi G,
Diagaradjane P, Anand P, Pandey MK, Gelovani J, Krishnan S, Guha S
and Aggarwal BB: Resveratrol, a multitargeted agent, can enhance
antitumor activity of gemcitabine in vitro and in orthotopic mouse
model of human pancreatic cancer. Int J Cancer. 127:257–268.
2010.PubMed/NCBI
|
|
13.
|
Banerjee S, Kaseb AO, Wang Z, Kong D,
Mohammad M, Padhye S, Sarkar FH and Mohammad RM: Antitumor activity
of gemcitabine and oxaliplatin is augmented by thymoquinone in
pancreatic cancer. Cancer Res. 69:5575–5583. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Wang ZH, Chen H, Guo HC, Tong HF, Liu JX,
Wei WT, Tan W, Ni ZL, Liu HB and Lin SZ: Enhanced antitumor
efficacy by the combination of emodin and gemcitabine against human
pancreatic cancer cells via downregulation of the expression of
XIAP in vitro and in vivo. Int J Oncol. 39:1123–1131.
2011.PubMed/NCBI
|
|
15.
|
Chen H, Wei W, Guo Y, Liu A, Tong H, Wang
Z, Tan W, Liu J and Lin S: Enhanced effect of gemcitabine by emodin
against pancreatic cancer in vivo via cytochrome C-regulated
apoptosis. Oncol Rep. 25:1253–1261. 2011.PubMed/NCBI
|
|
16.
|
Wei WI, Chen H, Ni ZL, Liu HI, Tong HF,
Fan L, Liu A, Qiu MU, Liu DL, Guo HC, Wang ZH and Lin SZ: Antitumor
and apoptosis-promoting properties of emodin, an anthraquinone
derivative from Rheum officinale Baill, against pancreatic
cancer in mice via inhibition of Akt activation. Int J Oncol.
39:1381–1390. 2011.PubMed/NCBI
|
|
17.
|
Sun HD, Huang SX and Han QB: Diterpenoids
from isodon species and their biological activities. Nat Prod Rep.
23:673–698. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Huang J, Wu L, Tashiro S, Onodera S and
Ikejima T: Reactive oxygen species mediate oridonin-induced HepG2
apoptosis through p53, MAPK, and mitochondrial signaling pathways.
J Pharmacol Sci. 107:370–379. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Jin S, Shen JN, Wang J, Huang G and Zhou
JG: Oridonin induced apoptosis through AKT and MAPKS signaling
pathways in human osteosarcoma cells. Cancer Biol Ther. 6:261–268.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Liu YQ, Mu ZQ, You S, Tashiro S, Onodera S
and Ikejima T: Fas/FasL signaling allows extracelluar-signal
regulated kinase to regulate cytochrome c release in
oridonin-induced apoptotic U937 cells. Biol Pharm Bull.
29:1873–1879. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Li D, Wu LJ, Tashiro S, Onodera S and
Ikejima T: Oridonin induced A431 cell apoptosis partially through
blockage of the RAS/RAF/ERK signal pathway. J Pharmacol Sci.
103:56–66. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Ikezoe T, Chen SS, Tong XJ, Heber D,
Taguchi H and Koeffler HP: Oridonin induces growth inhibition and
apoptosis of a variety of human cancer cells. Int J Oncol.
23:1187–1193. 2003.PubMed/NCBI
|
|
23.
|
Hu HZ, Yang YB, Xu XD, Shen HW, Shu YM,
Ren Z, Li XM, Shen HM and Zeng HT: Oridonin induces apoptosis via
PI3K/Akt pathway in cervical carcinoma HeLa cell line. Acta
Pharmacol Sin. 28:1819–1826. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24.
|
Hsieh TC, Wijeratne EK, Liang JY,
Gunatilaka AL and Wu JM: Differential control of growth, cell cycle
progression, and expression of NF-kappaB in human breast cancer
cells MCF-7, MCF-10A, and MDA-MB-231 by ponicidin and oridonin,
diterpenoids from the chinese herb Rabdosia rubescens.
Biochem Biophys Res Commun. 337:224–231. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25.
|
Cheng Y, Qiu F, Ye YC, Tashiro S, Onodera
S and Ikejima T: Oridonin induces G2/M arrest and apoptosis via
activating ERK-p53 apoptotic pathway and inhibiting PTK-RAS-RAFJNK
survival pathway in murine fibrosarcoma L929 cells. Arch Biochem
Biophys. 490:70–75. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26.
|
Gao FH, Hu XH, Li W, Liu H, Zhang YJ, Guo
ZY, Xu MH, Wang ST, Jiang B, Liu F, Zhao YZ, Fang Y, Chen FY and Wu
YL: Oridonin induces apoptosis and senescence in colorectal cancer
cells by increasing histone hyperacetylation and regulation of p16,
p21, p27 and c-myc. BMC Cancer. 10:6102010. View Article : Google Scholar : PubMed/NCBI
|
|
27.
|
Rufini A and Melino G: Cell death
pathology: The war against cancer. Biochem Biophys Res Commun.
414:445–450. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28.
|
Kim EK and Choi EJ: Pathological roles of
MAPK signaling pathways in human diseases. Biochim Biophys Acta.
1802:396–405. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29.
|
Martelli AM, Nyåkern M, Tabellini G,
Bortul R, Tazzari PL, Evangelisti C and Cocco L: Phosphoinositide
3-kinase/Akt signaling pathway and its therapeutical implications
for human acute myeloid leukemia. Leukemia. 20:911–928. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
30.
|
Wu GS: The functional interactions between
the p53 and MAPK signaling pathways. Cancer Biol Ther. 3:156–161.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
31.
|
McCubrey JA, Lahair MM and Franklin RA:
Reactive oxygen species-induced activation of the MAP kinase
signaling pathways. Antioxid Redox Signal. 8:1775–1789. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
32.
|
Kholodenko BN and Birtwistle MR:
Four-dimensional dynamics of MAPK information processing systems.
Wiley Interdiscip Rev Syst Biol Med. 1:28–44. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33.
|
Rodríguez-Berriguete G, Fraile B,
Martínez-Onsurbe P, Olmedilla G, Paniagua R and Royuela M: MAP
kinases and prostate cancer. J Signal Transduct.
2012:1691702012.PubMed/NCBI
|
|
34.
|
Wada T and Penninger JM: Mitogen-activated
protein kinases in apoptosis regulation. Oncogene. 23:2838–2849.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
35.
|
Harris SL and Levine AJ: The p53 pathway:
positive and negative feedback loops. Oncogene. 24:2899–2908. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
36.
|
Shi Y, Sahu RP and Srivastava SK: Triphala
inhibits both in vitro and in vivo xenograft growth of pancreatic
tumor cells by inducing apoptosis. BMC Cancer. 8:2942008.
View Article : Google Scholar : PubMed/NCBI
|
|
37.
|
Ma Y, Yu WD, Kong RX, Trump DL and Johnson
CS: Role of nongenomic activation of phosphatidylinositol
3-kinase/Akt and mitogen-activated protein kinase/extracellular
signal regulated kinase kinase/extracellular signal-regulated
kinase 1/2 pathway in 1,25D3-mediated apoptosis in squamous cell
carcinoma cells. Cancer Res. 66:8131–8138. 2006.
|
|
38.
|
Filomeni G, Graziani I, Rotilio G and
Ciriolo MR: trans-Resveratrol induces apoptosis in human breast
cancer cells MCF-7 by the activation of MAP kinases pathways. Genes
Nutr. 2:295–305. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
39.
|
Bulavin DV and Fornace AJ Jr: p38 MAP
kinase’s emerging role as a tumor suppressor. Adv Cancer Res.
92:95–118. 2004.
|
|
40.
|
Vousden KH and Prives C: P53 and
prognosis: new insights and further complexity. Cell. 120:7–10.
2005.PubMed/NCBI
|
|
41.
|
Galluzzi L, Morselli E, Kepp O, Tajeddine
N and Kroemer G: Targeting p53 to mitochondria for cancer therapy.
Cell Cycle. 7:1949–1955. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
42.
|
Califice S, Waltregny D, Castronovo V and
van den Brûle F: Prostate carcinoma cell lines and apoptosis: a
review. Rev Med Liege. 59:704–710. 2004.(In French).
|
|
43.
|
Kim YS, Lee HJ, Jang C, Kim HS and Cho YJ:
Knockdown of RCAN1.4 increases susceptibility to FAS-mediated and
DNA damage-induced apoptosis by upregulation of p53 expression.
Korean J Physiol Pharmacol. 13:483–489. 2009. View Article : Google Scholar : PubMed/NCBI
|