Introduction
Lung cancer is one of the leading causes of
cancer-related mortality worldwide (1), and adenocarcinoma is the most common
histological subtype. In Japan, the mortality rate of lung cancer
is the highest of all types of cancer since 1998 (2). The 5-year survival rate of lung
cancer surgically treated is currently 69.6% in all stages, and
86.8% even in stage IA (3).
Therefore, more effective strategies of therapy are necessary.
NR0B1, also known as dosage-sensitive sex reversal,
adrenal hypoplasia critical region, on X-chromosome gene 1 (DAX-1),
is a member of the orphan nuclear receptor family (4,5), and
its mutations result in adrenal hypoplasia congenita (6). NR0B1 is physiologically expressed in
the adrenal cortex, ovary, Sertoli cells, pituitary gonadotropes,
ventromedial hypothalamic nucleus cells, and others (7), and is correlated with gonadal
development, sex determination, and steroidogenesis (8). NR0B1 is a negative regulator of
steroid production (9–12), which represses the transcription of
other nuclear receptors via heterodimerization, including androgen,
estrogen, and progesterone receptors (13–15).
Such repression is mediated via the N-terminal repeat domain of
NR0B1, containing three LxxLL motifs (4,5,16).
Cancers consist of heterogeneous cell populations
derived from a single clone. It has previously been demonstrated
that cells with tumorigenic potential are limited to a small
population, known as cancer stem cells (CSCs). In flow-cytometric
analysis, CSCs are stained faintly by Hoechst 33342, and are
enriched in a side population, where dimly stained cells are
collected (17–19). NR0B1 is one of the highly expressed
genes in the side population of lung adenocarcinoma (20). NR0B1 expression is detected in a
number of cancers, including endometrial carcinoma, prostate
carcinoma, Ewing’s sarcoma, and lung cancer (21–25).
We previously reported that NR0B1 is correlated with the malignant
potential of lung adenocarcinoma through invasion, colony
formation, and tumorigenic activities (25).
Peroxisome proliferator-activated receptor γ (PPARγ)
is a member of the nuclear receptor superfamily of ligand-activated
transcriptional factors and is implicated in adipogenesis (26,27).
PPARγ binds to the peroxisome proliferator responsive element
(PPRE) (28), and is expressed in
a number of tissues, including type II pneumocytes, in humans
(29–31). PPARγ ligands, such as troglitazone
(TGZ) and 15-deoxy-delta12–14-prostaglandin J2 (15d-PGJ2), inhibit
the growth of human lung adenocarcinoma through ligand-induced
differentiation, growth arrest, and induction of apoptosis
(32–35). PPARγ is reported to interact with
NR0B1 via its DNA binding and hinge domains (36). The domain of NR0B1 which is
essential for the interaction with PPARγ, however, has yet to be
determined. In contrast to NR0B1, PPARγ possesses an antagonistic
function against lung adenocarcinoma. Therefore, there is a
possibility that NR0B1 and PPARγ possess an opposite effect on
tumors. In the present study, we examined the inhibitory effect of
PPARγ on NR0B1 in lung adenocarcinoma.
Materials and methods
Plasmids, cells, and chemicals
The human NR0B1 cDNA was cloned into pIRESpuro and
pTRE3G vectors (Clontech, Palo Alto, CA), and pCMV7.1-3xFLAG vector
(Sigma, St. Louis, MO), respectively. The deletion mutant of NR0B1
(del-NR0B1), lacking the N-terminal region of NR0B1, was
PCR-amplified and cloned into pCMV7.1-3xFLAG vector. The human
PPARγ cDNA was also cloned into pIRESpuro and pHM6 HA (Roche
Diagnostics, Mannheim, Germany) vectors. The luciferase construct
containing three repeats of PPRE (PPRE-Luc) was obtained from
OriGene (Rockville, MD). HEK-293T and human lung adenocarcinoma
cell lines A549 and PC9 were purchased from the American Type
Culture Collection (Rockville, MD). Cells were cultured in
Dulbecco’s Modified Eagle’s Medium (DMEM; Sigma) supplemented with
10% fetal calf serum (FCS; Nippon Bio-Supp. Center, Tokyo, Japan).
GW9662 (Calbiochem, Gibbstown, NJ) was dissolved in
dimethylsulfoxide (DMSO), and used as a PPARγ specific
antagonist.
Luciferase assay
PPRE-Luc was cotransfected with various amounts of
expression plasmid (pIRES-Puro) containing NR0B1, del-NR0B1, and
PPARγ to A549 and PC9 cells, using TransFast Transfection Reagent
(Promega, Madison, WI). Empty vector was added to maintain a
consistent amount of DNA used for transfection. After 48 h, cells
were harvested, and the luciferase activity was measured as
previously reported (37). Cells
transfected with empty vector alone were used as control and the
relative luciferase activity was represented as folds of
control.
Immunoprecipitation assay
The Flag-tagged wt-NR0B1 or del-NR0B1 was
coexpressed with HA-tagged PPARγ in HEK-293T cells. The nuclear
extract was mixed with either anti-FLAG M2 conjugated beads or
anti-HA conjugated beads (Dynabeads, Invitrogen, Carlsbad, CA).
Immunocomplexes were analyzed by immunoblot analysis with anti-Flag
antibody (Sigma).
Induced expression of NR0B1 with
Tet-Express
Inducible expression of NR0B1 was performed with
Tet-Express Inducible Expression Systems (Clontech) according to
the manufacturer’s instructions. Briefly, pTRE3G containing NR0B1
cDNA was transfected into A549 cells with linear puromycin marker
(Clontech). Puromycin-resistant clones were selected as Tet-Express
inducible cells. NR0B1 expression induced by the transfection of
Tet-Express with Xfect transfection reagent (Clontech) was
confirmed by immunoblotting as described below. Cells were
harvested and the nuclear extractions were prepared; they were
isolated on 10% SDS-polyacrylamide gels, transferred into immobilon
(Millipore, Bedford, MA), and incubated with anti-NR0B1 (Abcam Ltd,
Cambridge, UK) or anti-Lamin A/C (Cell Signaling, Beverly, MA)
antibodies. After washing, the blots were incubated with an
appropriate peroxidase-labeled secondary antibody (MBL, Nagoya,
Japan), and then reacted with Renaissance reagents (NEN, Boston,
MA) before exposure.
Quantification of mRNA levels by
real-time reverse-transcription PCR (RT-PCR)
Total RNA was extracted from cells transfected with
Tet-Express or incubated with GW9662 (20 μM overnight) using
an RNeasy kit (Qiagen, Valencia, CA). RNA was reverse-transcribed
into cDNA by Superscript III (Invitrogen). The mRNA levels for
aldehyde dehydrogenase 3A1 (ALDH3A1) and glyceraldehyde-3-phosphate
dehydrogenase (GAPDH) were measured using TaqMan gene expression
assays (Applied Biosystems, Foster City, CA). The amount of ALDH3A1
mRNA was normalized to that of GAPDH mRNA. The mRNA amount of cells
transfected with empty vector alone or supplemented with DMSO were
used as a control. The data were represented as folds of
control.
Aldefluor assay
ALDH activity was detected using the Aldefluor assay
kit (Stemcell Technologies, Vancouver, Canada) as described by the
manufacturer. Briefly, cells were suspended in Aldefluor assay
buffer containing ALDH substrate and BODIPY-aminoacetaldehyde
(BAAA). The BAAA was taken up by living cells and converted by
intracellular ALDH into BODIPY-aminoacetate, which yields bright
fluoresce. The brightly fluorescent ALDH-expressing cells were
detected with FACS Calibur or FACS Aria II (BD Biosciences,
Franklin Lakes, NJ). As a negative control, cells were stained
under identical conditions with the specific ALDH inhibitor,
diethylaminobenzaldehyde (DEAB; Sigma). Data were analyzed by Cell
Quest software (BD Biosciences).
Patients
Fifty-two patients with p-Stage IA lung
adenocarcinoma who had undergone surgery at the Department of
General Thoracic Surgery, Osaka University, between 1995 and 2003,
were included in this study. No patients had chemotherapy or
radiation therapy prior to surgery, and complete resection of
tumors was performed. The clinical characteristics of the patients
are shown in Table I. Survival
data were available for all patients. The mean follow-up duration
after surgery was 6.2±0.3 years. The study was approved by the
ethics review board of the Graduate School of Medicine, Osaka
University.
 | Table IClinical characteristics of 52 cases
of Stage IA lung adenocarcinoma. |
Table I
Clinical characteristics of 52 cases
of Stage IA lung adenocarcinoma.
| Number of
patients |
|---|
| Gender | |
| Male | 20 |
| Female | 32 |
| Histology | |
| Adenocarcinoma
in situ | 27 |
| Minimally invasive
adenocarcinoma | 6 |
| Invasive
adenocarcinoma | 19 |
| Recurrence | |
| Yes | 11 |
| No | 41 |
| Prognosis | |
| Deceased | 6 |
| Alive | 46 |
Immunohistochemical analysis
Histologic specimens were fixed in 10% formalin and
routinely processed for paraffin-embedding. Paraffin-embedded
specimens were stored in the dark room in the Department of
Pathology of Osaka University Hospital at room temperature, and
sectioned at 4 μm thickness at the time of staining. After
antigen retrieval with Pascal pressurized heating chamber (Dako
A/S, Glostrup, Denmark), the sections were incubated with
anti-NR0B1 or anti-PPARγ antibody (Abcam), subjected to the
treatment with ChemMate EnVision kit (Dako). DAB (Dako) was used as
a chromogen. As a negative control, staining was carried out in the
absence of primary antibody. Immunohistochemically stained sections
were evaluated independently by two pathologists (Y.S. and E.M.) in
a blinded manner, without any knowledge of the clinicopathological
parameters or patient outcomes. Cases were categorized into three
groups (score 0, score 1, and score 2) according to the extent of
staining in each section. Cases without any staining were
categorized as score 0, those with <10% positive cells among
tumor were categorized as score 1 and those ≥10% were grouped as
score 2.
Statistical analysis
Statistical analysis for experimental studies was
performed with the Student’s t-test. The values are shown as the
mean ± standard error (SE) of at least three experiments.
Statistical analysis for clinical samples was performed using JMP
ver. 9.0.2 software (SAS Institute Inc., Cary, NC). Five-year
overall survival (OS) and disease-free survival (DFS) were
calculated by the Kaplan-Meier method, and the differences in
survival curves were analyzed by the log-rank test.
Results
Inhibitory effect of NR0B1 on the
transactivation ability of PPARγ in lung adenocarcinoma cell
lines
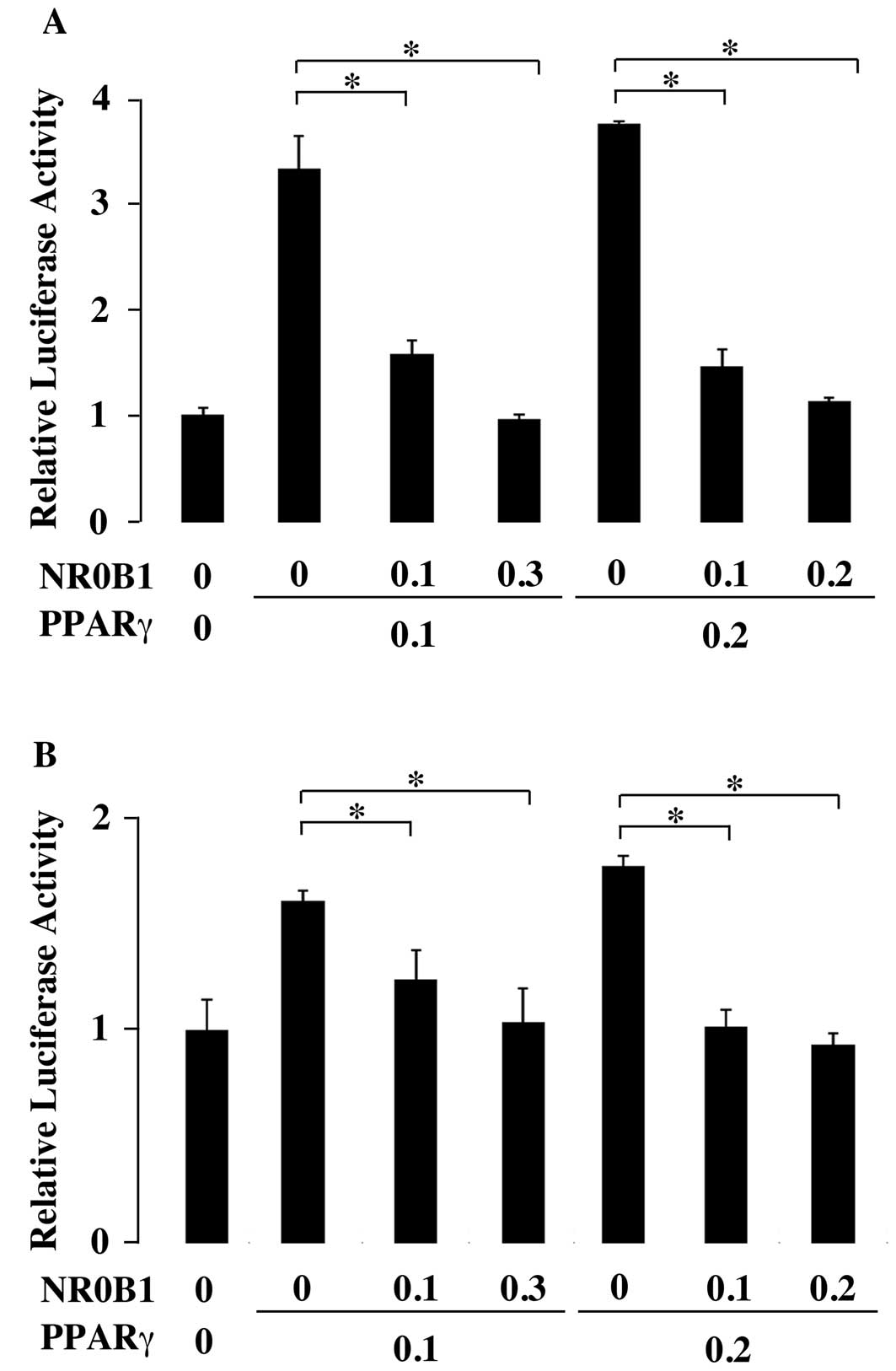
To examine the effect of NR0B1 on the
transactivation ability of PPARγ, the reporter plasmid containing
luciferase gene under the control of PPARγ recognition elements
(PPRE) was transfected to the A549 lung adenocarcinoma cell line.
When the PPARγ expression plasmid was cotransfected, the luciferase
activity increased approximately 3-fold (Fig. 1A). However, the coexpression of
NR0B1 dose-dependently interfered with the increase of luciferase
activity by PPARγ (Fig. 1A). The
comparable results were obtained in PC9, another lung
adenocarcinoma cell line (Fig.
1B).
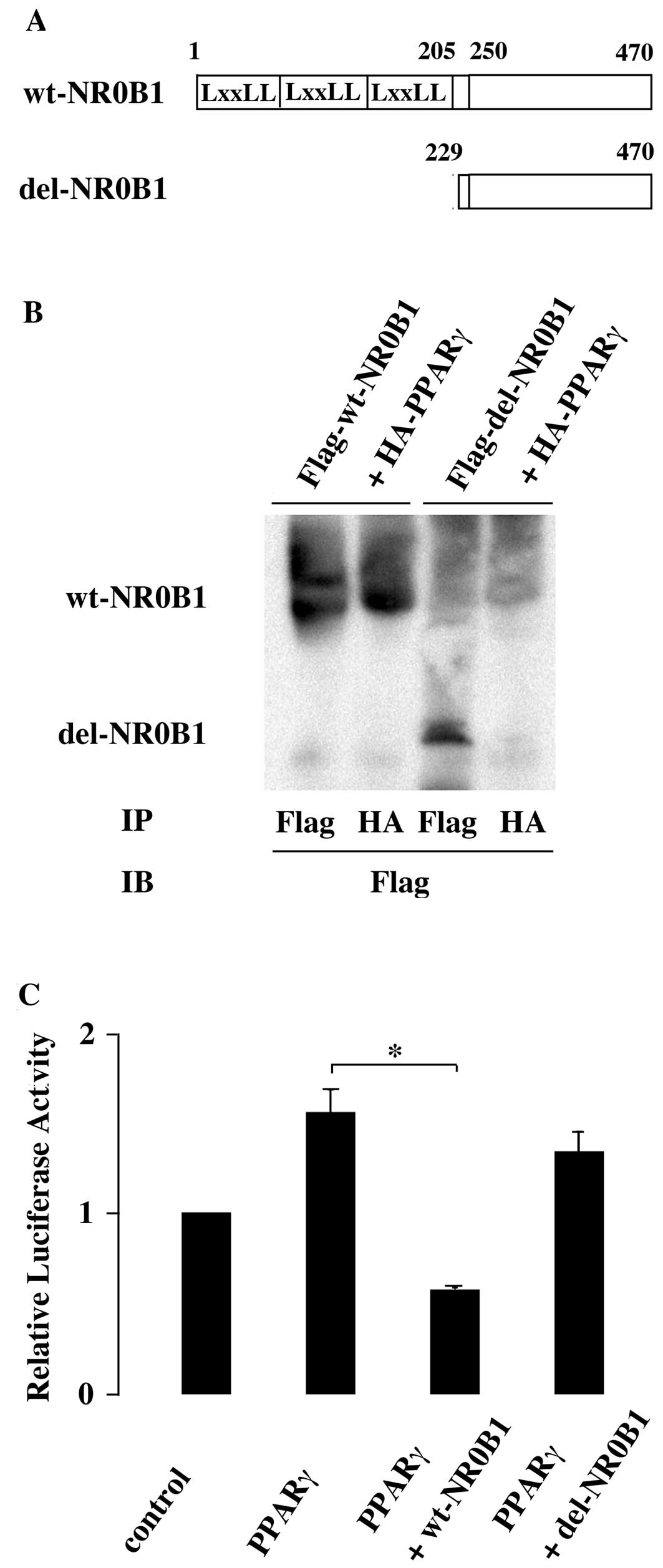
Kim et al (36) demonstrated that PPARγ is physically
bound to NR0B1 via its DNA binding and hinge domains. The domain of
NR0B1 mediating the interaction with PPARγ has yet to be
identified. NR0B1 possesses repeated LxxLL motifs in its N-terminal
half, and interacts with various factors, such as estrogen
receptor, via these LxxLL motifs. Then, the mutant NR0B1 lacking
LxxLL motifs (del-NR0B1) was constructed (Fig. 2A). Flag-tagged wild-type NR0B1 or
del-NR0B1 was coexpressed with HA-tagged PPARγ in HEK293T cells and
the nuclear extract was analyzed. The immunoprecipitated product
with anti-HA antibody contained Flag-tagged wild-type NR0B1, but it
did not contain Flag-tagged del-NR0B1 (Fig. 2B). This indicated that the
interaction of NR0B1 with PPARγ was mediated via its N-terminal
domain containing LxxLL motifs.
Next, the wild-type NR0B1 or del-NR0B1 was
cotransfected with PPARγ. In contrast to wild-type NR0B1, del-NR0B1
did not interfere with the PPARγ transactivation ability,
indicating that the inhibitory effect of NR0B1 was mediated via its
N-terminal domain containing LxxLL motifs (Fig. 2C).
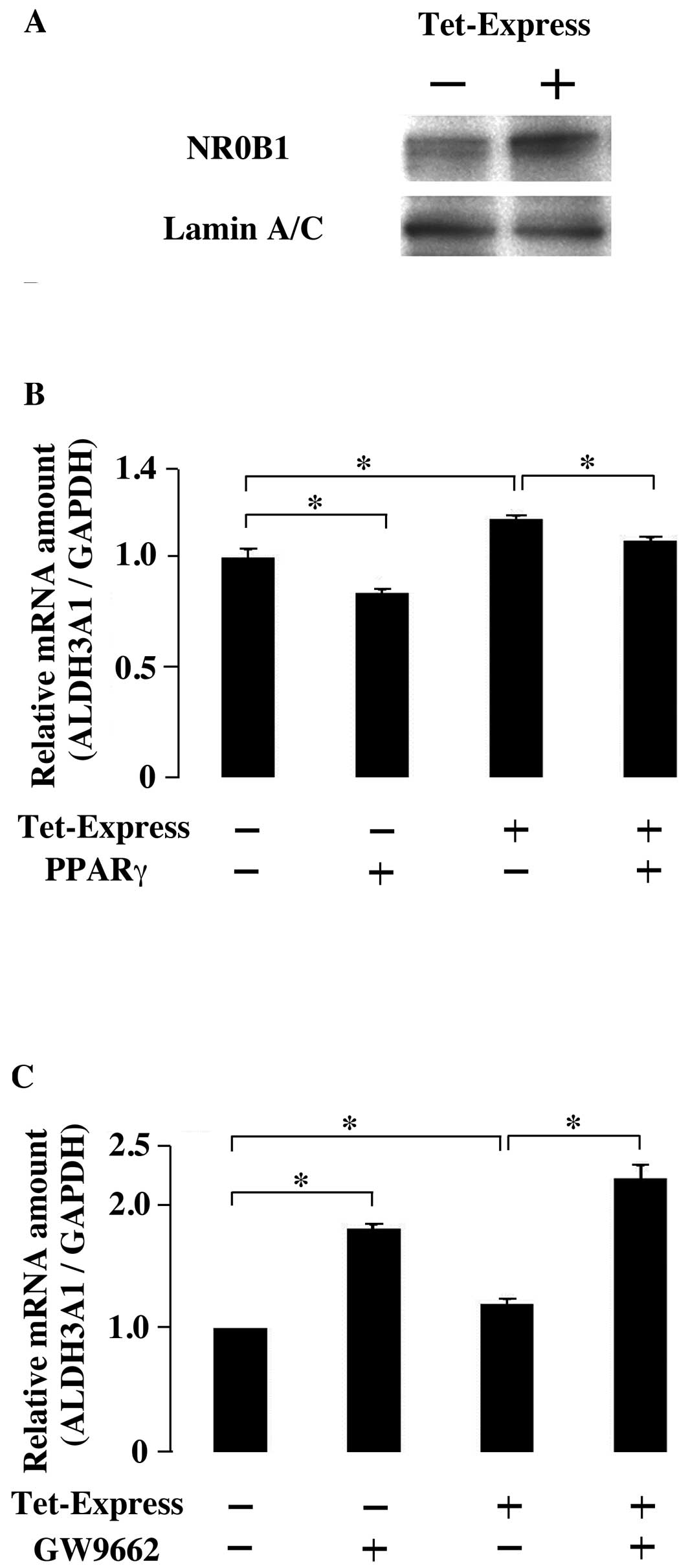
Effect of NR0B1 and PPARγ on the
expression of ALDH3A1
Tumorigenic potential is correlated with ALDH3A1
expression in lung adenocarcinoma (38). Therefore, the effect of NR0B1 and
PPARγ on the expression of ALDH3A1 was examined. Tet-Express
inducible system of NR0B1 was established in A549 cells (Fig. 3A). Without NR0B1, the
overexpression of PPARγ decreased the ALDH3A1 expression level
(Fig. 3B). The induced expression
of NR0B1 increased the ALDH3A1 expression, but this increase
interfered with the overexpression of PPARγ (Fig. 3B). These results indicated that the
effect of NR0B1 on ALDH3A1 expression was inhibited by PPARγ.
Moreover, the functional interaction between NR0B1 and PPARγ was
examined with PPARγ inhibitor GW9662. When GW9662 was added, the
expression level of ALDH3A1 increased. The induced expression of
NR0B1 also increased ALDH3A1 expression. The induced NR0B1
expression and the treatment of GW9662 additively increased ALDH3A1
expression (Fig. 3C).
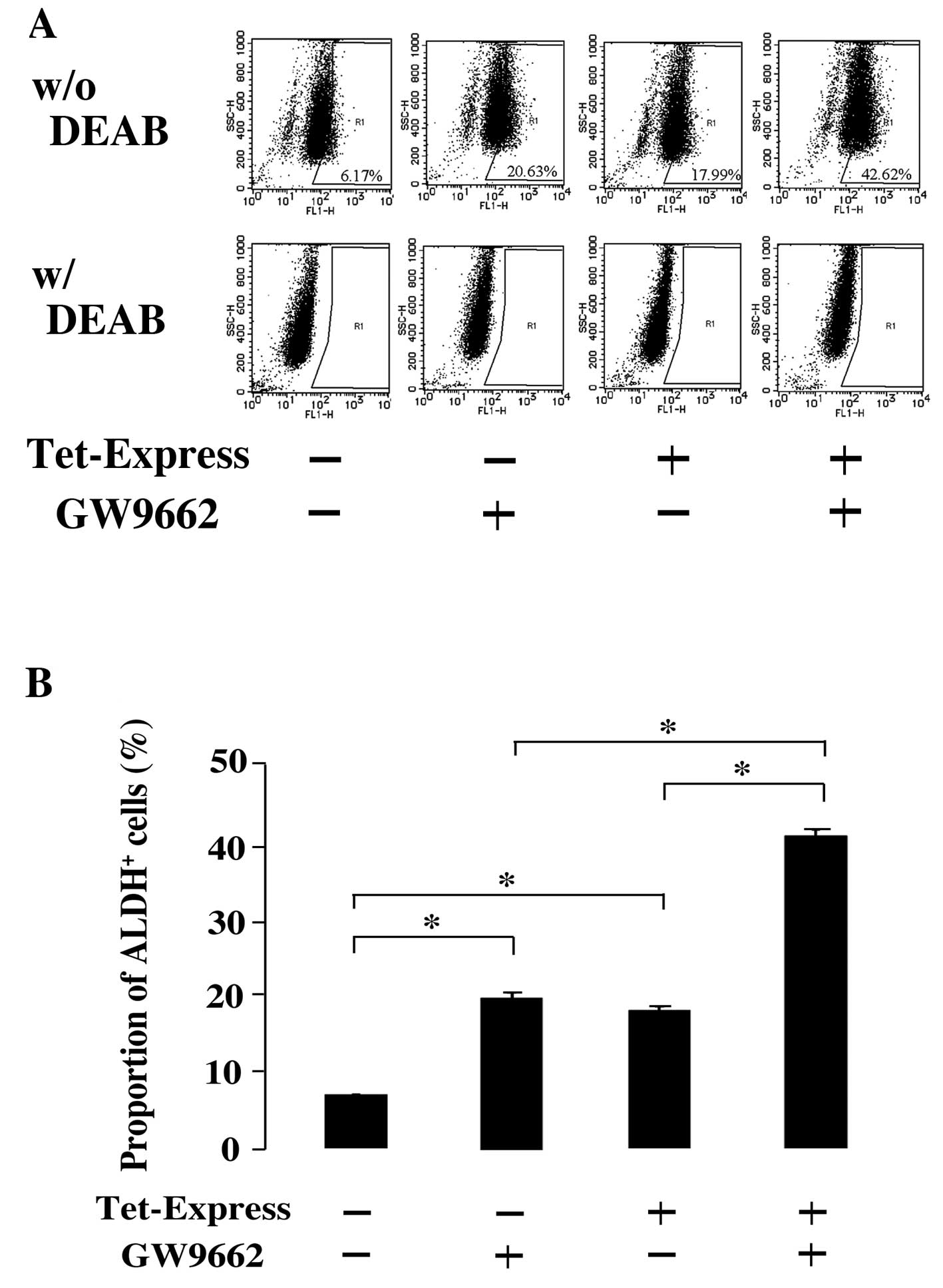
ALDH activity was measured with Aldefluor assay.
ALDH activity was examined in the combination of NR0B1 induction
and GW9662. Consistent with the result of ALDH3A1 expression, the
proportion of ALDH-positive cells increased when NR0B1 was induced
and when GW9662 was treated, respectively (Fig. 4A and B). The simultaneous induction
of NR0B1 and treatment of GW9662 further increased the proportion
of ALDH-positive cells (Fig. 4A and
B).
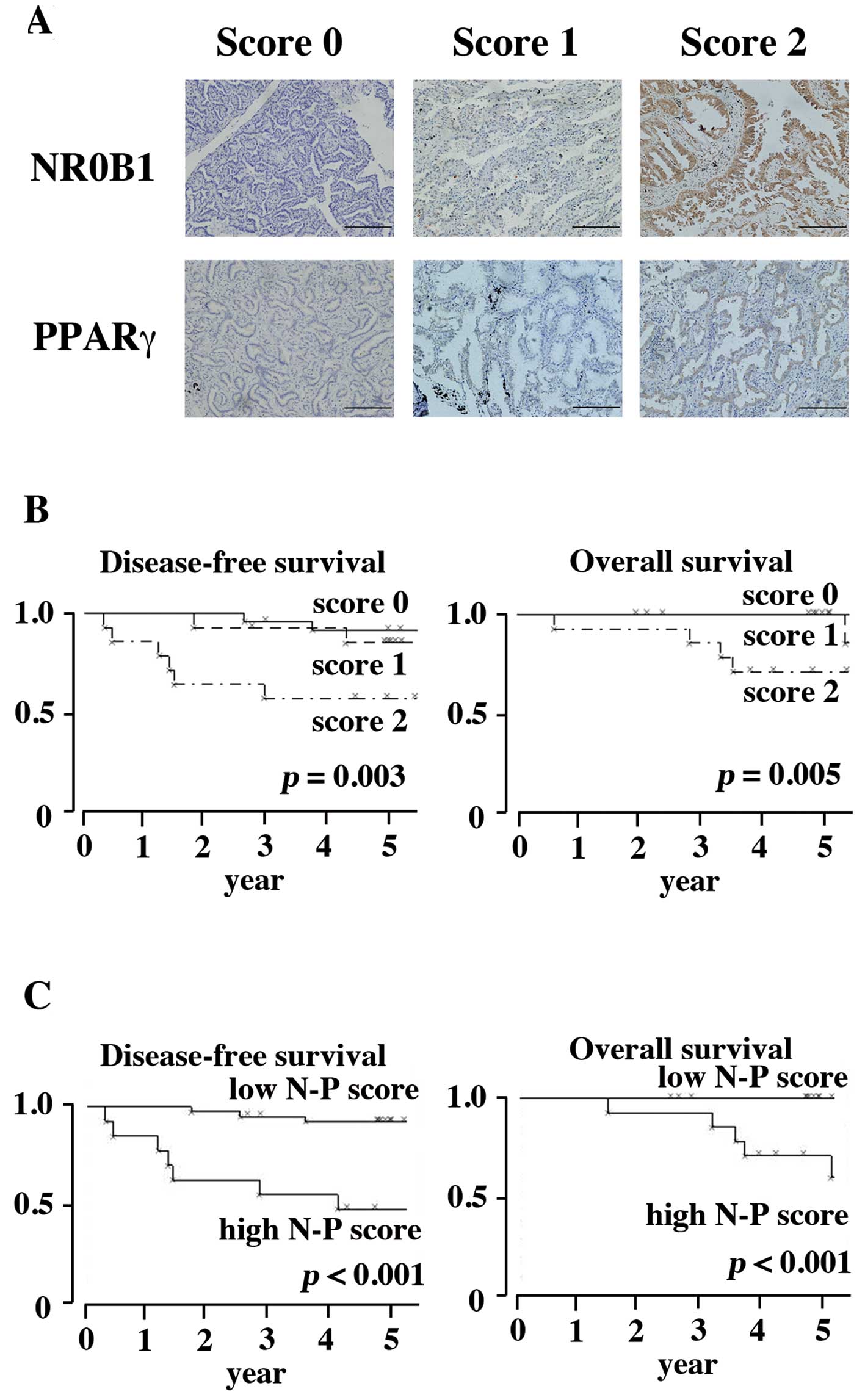
Immunohistochemical findings in clinical
specimens
Expression of NR0B1 and PPARγ was examined
immunohistochemically in 52 clinical cases of lung adenocarcinoma.
Cases were categorized into three groups (score 0, score 1, and
score 2) according to the proportion of positive cells (Fig. 5A). In the NR0B1 score, 24 cases
were categorized as score 0, 14 as score 1, and 14 as score 2. In
the PPARγ score, 21 cases were categorized as score 0, 23 as score
1, and 8 as score 3, respectively. Consistent with our previous
report (25), the high NR0B1 score
was correlated with poor prognosis in both DFS and OS (Fig. 5B, p=0.003 and 0.005, respectively).
To evaluate the functional interaction between NR0B1 and PPARγ, the
PPARγ score was subtracted from the NR0B1 score (NR0B1 score -
PPARγ score), and the resultant value was referred to as ‘N-P
score’. The low ‘N-P score’ corresponds to high PPARγ and low NR0B1
scores, whereas the high ‘N-P score’ to low PPARγ and high NR0B1
scores. Cases were again divided into two categories; cases with ≤0
‘N-P score’ were categorized as ‘low N-P’, and cases with ≥1 ‘N-P
score’ were categorized as ‘high N-P’ (Fig. 5C). The former included 39 cases,
whereas the latter 13 cases. The ‘high N-P’ was correlated with
poor prognosis in both DFS and OS (Fig. 5C).
Eleven out of the 14 cases with NR0B1 score 1 and 4
out of the 14 cases with NR0B1 score 2 were grouped as ‘low N-P’.
The former 11 cases were alive (1 case with recurrence), and the
latter 4 cases were alive without recurrence. These results
indicated that high NR0B1 cases with favorable prognosis could be
categorized as ‘low N-P’ group.
Discussion
NR0B1, an orphan nuclear receptor, is expressed in a
side population of lung adenocarcinoma. We previously reported that
the knockdown expression of NR0B1 reduced tumorigenic and
anti-apoptotic potential in lung adenocarcinoma cell lines, and
that the high expression of NR0B1 was a poor indicator of prognosis
in clinical cases of lung adenocarcinoma. NR0B1 is known to form
heterodimers with various factors, such as estrogen and
progesterone receptors. NR0B1 has previously been reported to
interact with PPARγ in adipose tissue (36). In lung adenocarcinoma, PPARγ
inhibits the growth of tumor cells and induces apoptosis (32–35)
suggesting that NR0B1 and PPARγ might present opposite effects on
tumor cells. In the present study, we examined the interaction
between NR0B1 and PPARγ in lung adenocarcinoma.
The luciferase activity enhanced by PPARγ was
reduced by the coexpression of NR0B1, indicating the inhibitory
effect of NR0B1 on the transactivation ability of PPARγ.
Immunoprecipitated experiments revealed that the N-terminal region
of NR0B1 containing three LxxLL motifs mediated the physical
interaction with PPARγ. Deletion of this region abolished the
inhibitory effect to PPARγ in luciferase assay. The N-terminal
region of NR0B1 appeared to be essential for physical and
functional interaction with PPARγ in lung adenocarcinoma. The
N-terminal region is essential for the interaction of NR0B1 with
various factors, such as estrogen receptors, and this was
applicable to the interaction with PPARγ.
ALDH activity is correlated with tumorigenic
potential in various types of tumors (38). Among 19 distinct isoforms, ALDH3A1
is responsible for ALDH activity of lung adenocarcinoma (38,39).
The level of NR0B1 induced by Tet-Express was correlated with ALDH
activity and ALDH3A1 expression levels. This was consistent with
our previous report that NR0B1 is involved in the malignant
potential of lung adenocarcinoma. In contrast to NR0B1, PPARγ was
inversely correlated with ALDH activity and ALDH3A1 expression
levels, which were reduced with the overexpression of PPARγ and
increased with the addition of PPARγ inhibitor GW9662. The additive
effect of NR0B1 and GW9662 indicated that PPARγ inhibited the
tumorigenic potential of NR0B1 in lung adenocarcinoma. NR0B1 and
PPARγ appeared to be antagonistic in malignant attitude.
We previously reported that the high NR0B1 score was
a negative prognostic indicator, when all stages of lung
adenocarcinoma were included. This was applicable even when only
stage IA cases were included in the present study. Furthermore, we
compared the evaluation using two factors (NR0B1 and PPARγ scores)
with that using a single factor (only NR0B1 score). When evaluated
with two factors, 11 of the 14 NR0B1 score 1 cases and 4 of the 14
NR0B1 score 2 cases were categorized to the group with favorable
prognosis. All of the re-categorized cases were alive, and most of
them were free from recurrence. These results suggested that the
evaluation of the two factors was a more accurate indicator than
that of a single factor. NR0B1 and PPARγ may interact with each
other also in clinical samples of lung adenocarcinoma.
Taken together, NR0B1 and PPARγ were antagonistic
with each other in malignant attitude. The interaction was mediated
through the N-terminal region of NR0B1. The high NR0B1 and low
PPARγ expression was strictly correlated with poor prognosis of
Stage IA lung adenocarcinoma.
Acknowledgements
The authors thank Professor Yuko Ohno
for her helpful discussion on statistics, Ms. Megumi Nihei-Sugano,
Ms. Etsuko Maeno, and Ms. Takako Sawamura for their technical
assistance.
References
|
1
|
Alberg AJ, Ford JG and Samet JM; American
College of Chest Physicians: Epidemiology of lung cancer: ACCP
evidence-based clinical practice guidelines (2nd edition). Chest.
132:29S–55S. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Committee for Scientific Affairs; Sakata
R, Fujii Y and Kuwano H: Thoracic and cardiovascular surgery in
Japan during 2009: Annual report by The Japanese Association for
Thoracic Surgery. Gen Thorac Cardiovasc Surg. 59:636–667. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sawabata N, Miyaoka E, Asamura H, et al:
Japanese lung cancer registry study of 11,663 surgical cases in
2004. J Thorac Oncol. 6:1229–1235. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zanaria E, Muscatelli F, Bardoni B, et al:
An unusual member of the nuclear hormone receptor superfamily
responsible for X-linked adrenal hypoplasia congenita. Nature.
372:635–645. 1994. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Swain A, Zanaria E, Hacker A, Lovell-Badge
R and Camerino G: Mouse Dax1 expression is consistent with a role
in sex determination as well as in adrenal and hypothalamus
function. Nat Genet. 12:404–409. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Burris TP, Guo W and McCabe ERB: The gene
responsible for adrenal hypoplasia congenita, DAX1, encodes a
nuclear hormone receptor that defines a new class within the
superfamily. Recent Prog Horm Res. 51:241–260. 1996.PubMed/NCBI
|
|
7
|
Ikeda Y, Swain A, Weber TJ, et al:
Steroidogenic factor 1 and Dax-1 colocalization in multiple cell
lineages: potential links in endocrine development. Mol Endocrinol.
10:1261–72. 1996.PubMed/NCBI
|
|
8
|
Lessnick SL, Dacwag CS and Golub TR: The
Ewing’s sarcoma oncoprotein EWS/FLI induces a p53-dependent growth
arrest in primary human fibroblasts. Cancer Cell. 1:393–401.
2002.
|
|
9
|
Lalli E, Melner MH, Stocco DM and
Sassone-Corsi P: DAX-1 blocks steroid production at multiple
levels. Endocrinology. 139:4237–4243. 1998.PubMed/NCBI
|
|
10
|
Zazopoulos P, Lalli E, Stocco DM and
Sassone-Corsi P: DNA binding and transcriptional repression by
DAX-1 blocks steroidogenesis. Nature. 390:311–315. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sugawara T, Lin D, Holt JA, et al:
Structure of the human steroidogenic acute regulatory protein
(StAR) gene. Biochemistry. 34:12506–12512. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Stocco DM and Clark BJ: Regulation of
acute production of steroid in steroidgenic tissue. Endocr Rev.
17:221–244. 1996.PubMed/NCBI
|
|
13
|
Park YY, Ahn SW, Kim HJ, et al: An
autoregulatory loop controlling orphan nuclear receptor DAX-1 gene
expression by orphan nuclear receptor ERRγ. Nucleic Acids Res.
33:6756–6768. 2005.PubMed/NCBI
|
|
14
|
Holter E, Kotaja N, Makela S, et al:
Inhibition of androgen receptor(AR) function by the reproductive
orphan nuclear receptor DAX-1. Mol Endocrinol. 16:512–528. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Agoulnik IU, Krause WC, Bingman WE III, et
al: Repressors of androgen and progesterone receptor action. J Biol
Chem. 278:31136–31148. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lalli E, Ohe K, Hindelang C and
Sassone-Corsi P: Orphan receptor DAX-1 is a shuttling RNA binding
protein associated with polyribosomes via mRNA. Mol Cell Biol.
20:4910–4921. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Reya T, Morrison SJ, Clarke MF and
Weissman IL: Stem cells, cancer, and cancer stem cells. Nature.
414:105–111. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dean M, Fojo T and Bates S: Tumor stem
cells and drug resistance. Nat Rev Cancer. 5:275–284. 2005.
View Article : Google Scholar
|
|
19
|
Lou H and Dean M: Targeted therapy for
cancer stem cells: the patched pathway and ABC transporters.
Oncogene. 26:1357–1360. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Seo DC, Sung JM, Cho HJ, et al: Gene
expression profiling of cancer stem cell in human lung
adenocarcinoma A549 cells. Mol Cancer. 6:752007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Saito S, Ito K, Suzuki T, et al: Orphan
nuclear receptor DAX-1 in human endometrium and its disorders.
Cancer Sci. 96:645–652. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nakamura Y, Suzuki T, Arai Y and Sasano H:
Nuclear receptor DAX1 in human prostate cancer: a novel independent
biological modulator. Endocr J. 56:39–44. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mendiola M, Carrillo J, García E, et al:
The orphan nuclear receptor DAX1 is up-regulated by the EWS/FLI1
oncoprotein and is highly expressed in Ewing tumors. Int J Cancer.
118:1381–1389. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kinsey M, Smith R and Lessnick SL: NR0B1
is required for the oncogenic phenotype mediated by EWS/FLI in
Ewing’s sarcoma. Mol Cancer Res. 4:851–859. 2006.
|
|
25
|
Oda T, Tian T, Inoue M, et al: Tumorgenic
role of orphan nuclear receptor NR0B1 in lung adenocarcinoma. Am J
Pathol. 175:1235–1245. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Spiegelman BM: PPAR-γ: Adipogenic
regulator and thiazolidinedine receptor. Diabetes. 47:507–514.
1998.
|
|
27
|
Tontonoz P, Hu E and Spiegelman BM:
Stimulation of adipogenesis in fibroblasts by PPARγ 2, a
lipid-activated transcription factor. Cell. 79:1147–1156. 1994.
|
|
28
|
Gearing KL, Gottlicher M, Teboul M,
Widmark E and Gustafsson JA: Interaction of the
peroxisome-proliferator-activated receptor and retinoid X receptor.
Proc Natl Acad Sci USA. 90:1440–1444. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mueller E, Sarraf P, Tontonoz P, et al:
Terminal differentiation of human breast cancer through PPARγ. Mol
Cell. 1:465–470. 1998.
|
|
30
|
Sarraf P, Mueller E, Jones D, et al:
Differentiation and reversal of malignant changes in colon cancer
through PPARγ. Nat Med. 4:1046–1052. 1998.PubMed/NCBI
|
|
31
|
Lambe KG and Tugwood JD: A human
perioxisome proliferator-activated receptor-γ is activated by
inducers of adipogenesis including thiazolidinedine drugs. Eur J
Biochem. 239:1–7. 1996.
|
|
32
|
Keshamouni VG, Reddy RC, Arenberg DA, et
al: Peroxisome proliferator-activated receptor-γ activation
inhibits tumor progression in non-small-cell lung cancer. Oncogene.
23:100–108. 2004.
|
|
33
|
Satoh T, Toyoda M, Hoshino H, et al:
Activation of peroxisome proliferator-activated receptor-γ
stimulates the growth arrest and DNA-damage inducible 153 gene in
non-small cell lung carcinoma cells. Oncogene. 21:2171–2180.
2002.
|
|
34
|
Li M, Lee TW, Mok TS, Warner TD, Yim AP
and Chen GC: Activation of peroxisome proliferator-activated
receptor-γ by troglitazone (TGZ) inhibits human lung cell growth. J
Cell Biochem. 96:760–774. 2005.
|
|
35
|
Tsubouchi Y, Sano H, Kawahito Y, et al:
Inhibition of human lung cancer cell growth by the peroxisome
proliferator-activated receptor-γ agonists through induction of
apoptosis. Biochem Biophys Res Commun. 270:400–405. 2000.
|
|
36
|
Kim GS, Lee GY, Nedumaran B, et al: The
orphan nuclear receptor DAX-1 acts as a novel transcriptional
corepressor of PPARγ. Biochem Biophys Res Commun. 370:264–268.
2008.PubMed/NCBI
|
|
37
|
Morii E and Oboki K: MITF is necessary for
generation of prostaglandin D2 in mouse mast cells. J Biol Chem.
279:48293–48299. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Patel M, Lu L, Zander DS, Sreerama L, Coco
D and Moreb JS: ALDH1A1 and ALDH3A1 expression in lung cancers:
Correlation with histologic type and potential precursors. Lung
Cancer. 59:340–349. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Muzio G, Maggiora M, Paiuzzi E, Oraldi M
and Canuto RA: Aldehyde dehydrogenase ands and cell proliferation.
Free Radic Biol Med. 52:735–746. 2012. View Article : Google Scholar : PubMed/NCBI
|