Introduction
Neutral endopeptidase (NEP, CD10) is a 90–110 kDa
zinc dependent metallopeptidase that cleaves peptide bonds on the
amino side of hydrophobic amino acids and inactivates a variety of
bioactive peptides including atrial natriuretic factor, substance
P, bradykinin, oxytocin, Leu- and Met-enkephalins, neurotensin,
bombesin, endothelin-1, bombesin-like peptides and amyloid-β
(1,2). Numerous tissues normally express NEP,
including epithelial cells of the prostate, kidney, intestine,
adrenal glands and lung. NEP expression is decreased or absent in
nearly 50% of primary and metastatic prostate cancers (3–5).
This loss can result from methylation of the NEP promoter or may
occur after androgen withdrawal because NEP transcription is
regulated by androgen (3,5,6).
Replacement of NEP using either exogenous recombinant NEP or
overexpression of NEP at the cell-surface using an NEP expression
vector inhibits PC cell growth, cell migration and tumorigenicity
(7,8). NEP actions have been demonstrated to
result from catalytic inactivation of NEP substrates such as
bombesin, endothelin-1 and basic fibroblast growth factor (bFGF),
and via protein-protein interaction of NEP’s cytoplasmic domain
with a variety of proteins, including ezrin/radixin/moesin
proteins, Lyn kinase, and the phosphatase and tensin homolog (PTEN)
protein (7,9,10).
NEP recruits endogenous PTEN to the cell membrane, leading to
prolonged PTEN protein stability and increased PTEN phosphatase
activity, resulting in constitutive downregulation of Akt activity.
NEP also indirectly inhibits Akt phosphorylation by catalytically
inactivating peptides that normally activate the insulin growth
factor-1 receptor (IGF-1R) leading to Akt phosphorylation. Thus,
NEP regulates Akt via both catalytic-dependent and -independent
pathways. In this regard, loss of NEP together with increased Akt
phosphorylation in primary prostate cancers predicts a
significantly shorter time to biochemical relapse in patients who
have undergone radical prostatectomy (11).
We previously reported that recombinant NEP augments
chemosensitivity of prostate cancer cells in vitro by
promoting PKC δ-mediated mitochondrial apoptosis as determined by
cytochrome-c release and caspase-9 activation (12). Taxanes, such as paclitaxel and
taxotere, are active anticancer drugs that are currently used to
treat advanced prostate cancer (13). The predominant mode of action of
paclitaxel is the binding to β-tubulin, stabilizing the
microtubule, and preventing its depolymerization (14). Recent studies suggest that
activated Akt contributes to paclitaxel-induced resistance and thus
inhibition of Akt may synergistically increase paclitaxel
sensitivity (15,16). In the current study, we examined
the combined antitumor effects on DU145 castration resistant
prostate cancer cells of treatment with an adenovirus expressing
NEP that inhibits Akt activation with paclitaxel.
Materials and methods
Cells and reagents
DU145 human PC cells (ATCC, Manassas, VA) were grown
in RPMI-1640 medium supplemented with 2 mM glutamine, 1%
non-essential amino acids, 100 units/ml streptomycin and
penicillin, and 10% fetal calf serum (FCS). Antibodies used include
anti-NEP antibody (NCL-CD10-270, Novocastra Laboratories Ltd.,
Newcastle upon Tyne, UK), anti-phospho-Akt antibody (Ser473, Cell
Signaling Technology, Inc., Beverly, MA), anti-Akt antibody (Cell
Signaling Technology, Inc.), anti-PTEN antibody (A2B1, Santa Cruz
Biotechnology, Inc.), anti-phospho-BAD antibody (Ser136, Cell
Signaling Technology, Inc.), anti-BAD antibody (Cell Signaling
Technology, Inc.), caspase-3 antibody (Cell Signaling Technology,
Inc.), PARP antibody (Cell Signaling Technology, Inc.), and
anti-β-actin antibody (Sigma-Aldrich, St. Louis, MO).
Adenovirus vector production and
transduction
cDNA containing the entire full-length human NEP was
used to construct and produce recombinant AdNEP plasmid (AdEasy™
Adenoviral Vector System, Stratagene). Adenovirus vectors were
amplified in 293 cells and purified by cesium chloride density
gradient ultracentrifugation. The AdLacZ vector was used as a
negative control.
NEP enzyme activity assay
NEP-specific enzyme activities was assessed as
described using Suc-Ala-Ala-Phe-pNA (Bachem Bioscience Inc.,
Philadelphia, PA, USA) as substrate (3). Specific activities were expressed as
picomoles per microgram of protein per minute and represent an
average of two separate measurements performed in duplicate on
separate occasions.
Cell counts and cell viability
assays
Following incubation overnight in T25 flask, cells
were infected with AdNEP or AdLacZ at the indicated concentrations.
Total cell numbers in three independent flasks in each group were
counted using a hemocytometer, and the mean value of four fields
was recorded. Cell viability was assessed by trypan blue, which was
added to cell cultures at a ratio 1:1 and left for 10 min, and
cells counted using a hemocytometer. The ratios of viable and dead
cells were determined. For all experiments, data presented are
representative of experiments performed at least three times in
triplicate or quadruplicate.
TUNEL assay
Apoptosis was determined in adherent DU145 cells
cultured in a chamber slide or deparaffinized DU145 tumor tissue
sections by Terminal deoxynucleotidyl transferase (TdT) mediated
d-Uridine Tri Phosphate nick end labeling (TUNEL) technique using
the In Situ Cell Death Detection kit, POD (Roche, Germany)
according to the recommendations of the manufacturer. Quantitative
evaluation of apoptotic cells was done by counting the TUNEL
positive cells among those cells under light microscopy (×1000).
The apoptotic index was expressed as TUNEL-positive cells per 100
cells.
Immunoblotting
Total cell lysate preparation and immuno-blotting
were performed as described (7,10).
Proteins were visualized using enhanced chemiluminescence (Amersham
Biosciences).
Xenograft model
Under an IACUC approved protocol, DU145 cells
(5×106 cells) were inoculated subcutaneously into the
flanks of 60 athymic male mice (Taconic, Hudson, NY, USA). When
tumors reached a size of 4–6 mm in diameter, mice were randomly
divided into 5 groups and treated with intratumor injection into
the center and periphery of each mass on days 1 and 15, as follows:
Group 1, 100 μl of saline (control); Groups 2 and 4, 100 μl of
saline containing 1×108 pfu of Ad-LacZ; Groups 3 and 5,
100 μl of saline containing 1×108 pfu of Ad-NEP. Groups
4 and 5 also received paclitaxel administered i.p. at dose 10
mg/kg, daily from days 2 to 4 and from days 16 to 18 (17). Tumor volume was calculated as [0.52
× (W × W × L)] where W, minor axis and L, major axis and was
measured every 3 days until day 28 post-adenoviral infection. Two
animals were sacrificed on day 6 and the remainder on day 28 and
the tumors dissected, weighed and collected for further
analysis.
Immunohistochemisty
Paraffin-embedded tissue sections were stained with
anti-NEP antibody (NCL) at 1:80 dilution, anti-PTEN antibody at
1:100 dilution, anti-phospho-Akt antibody at 1:100 dilution using
the Dako Envision + System HRP (Dako Cytomation, Carpinteria, CA)
according to the manufacturer’s instructions.
Statistical analysis
All analyses were done using Statview 5.0 (SAS
Institute, Inc., Cary, NC). Results were expressed as the mean ± SE
for three independent measurements. Analysis of variance was used
for analysis of continuous data followed by Fisher’s protected
least significant difference for post hoc analyses. Differences
with a P<0.05 were determined as statistically significant.
Results
Adenoviral NEP expression in DU145
cells
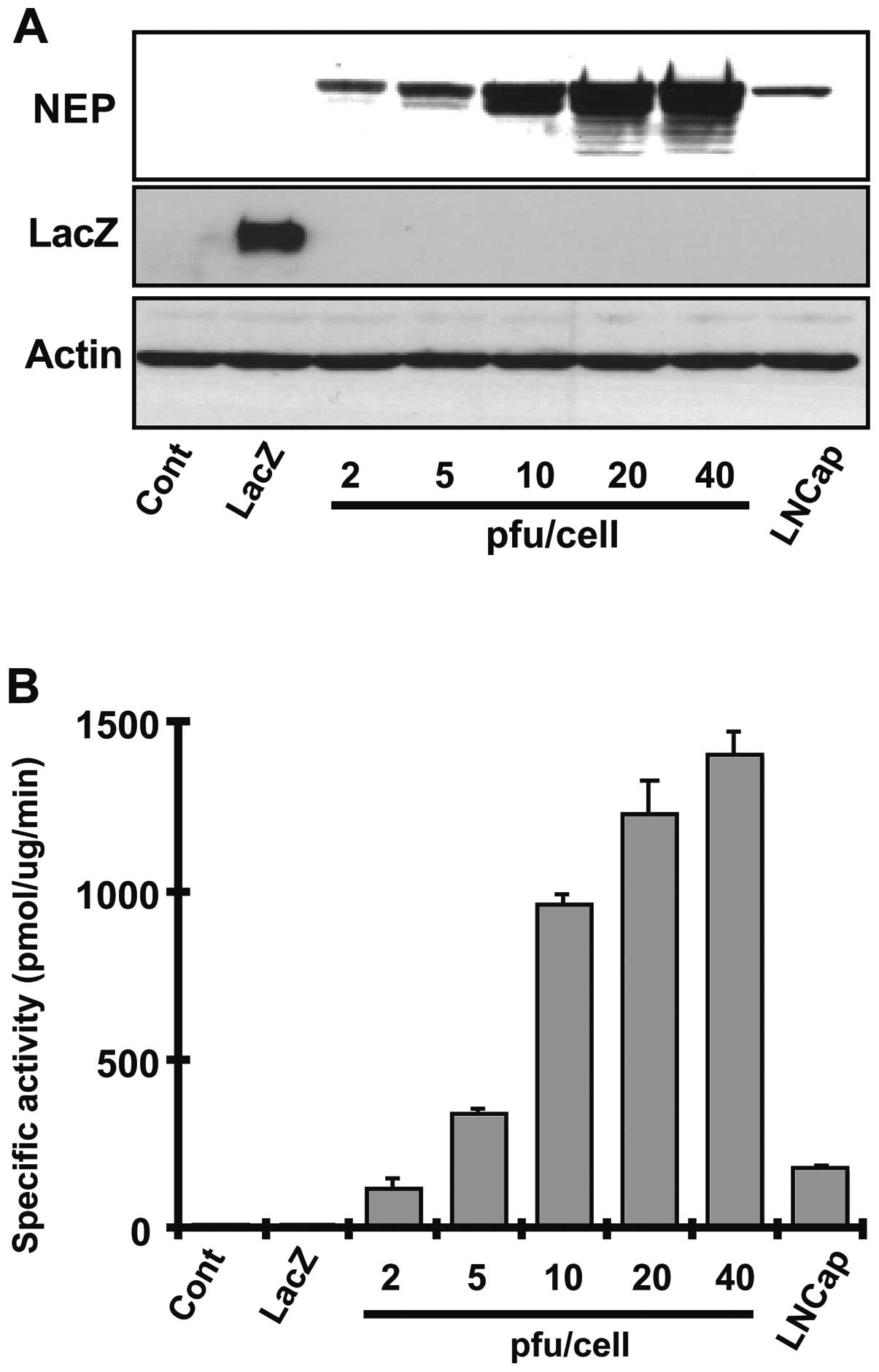
Adenoviral vectors expressing NEP or LacZ protein as
control were introduced into DU145 cells at a pfu/cell
concentration ranging from 2 to 40 and NEP protein levels and
enzymatic activity were measured after 72 h. NEP protein expression
and NEP specific enzyme activity increased in a pfu/cell
dose-dependent manner (Fig. 1A and
B). NEP activity in DU145 cells infected with AdNEP at 10
pfu/cell achieved an enzyme activity of 962.36±24.5 pmol/mg/min, a
10-fold higher specific activity than that achieved in DU145
infected lentiviral vector encoding NEP (96.57±10.2 pmol/mg/min)
(18). NEP activity in uninfected
DU145 cells or in DU145 cells infected with AdLacZ was 10.06±2.8
pmol/mg/min and 8.05±2.2 pmol/mg/min, respectively. We next
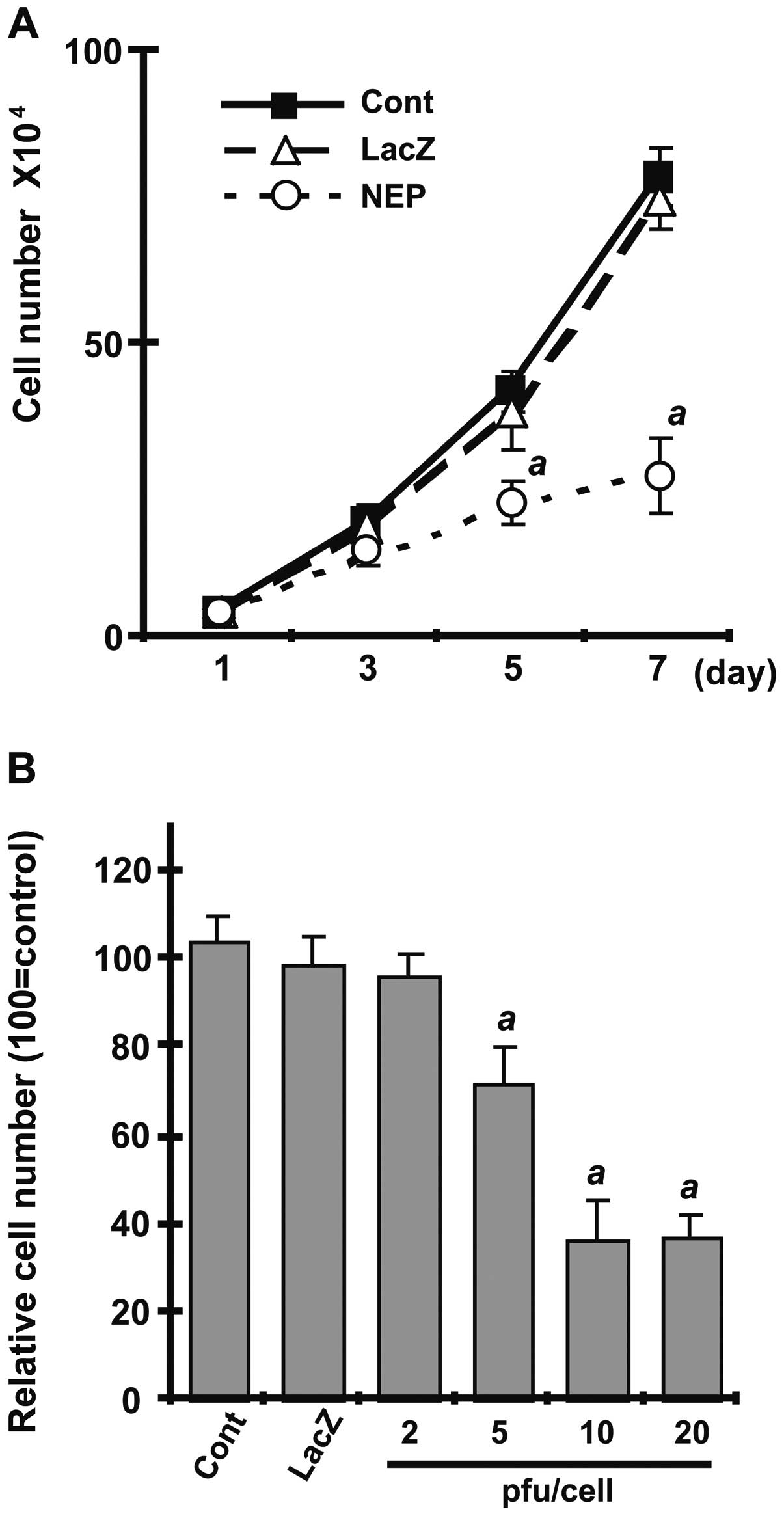
examined the effect of NEP expression on cell growth, comparing
uninfected DU145 cells with cells infected with AdNEP or AdLacZ at
10 pfu/cell. As shown in Fig. 2A,
AdNEP significantly inhibited DU145 cell growth (P<0.01)
compared with cells infected with AdLacZ or uninfected DU145 cells.
Growth inhibition was dose-dependent (Fig. 2B). Together, these results show
that adenoviral vectors can transduce catalytically active NEP
protein into DU145 cells resulting in high levels of NEP catalytic
activity and inhibition of cell growth.
Adenoviral NEP infection in combination
with paclitaxel
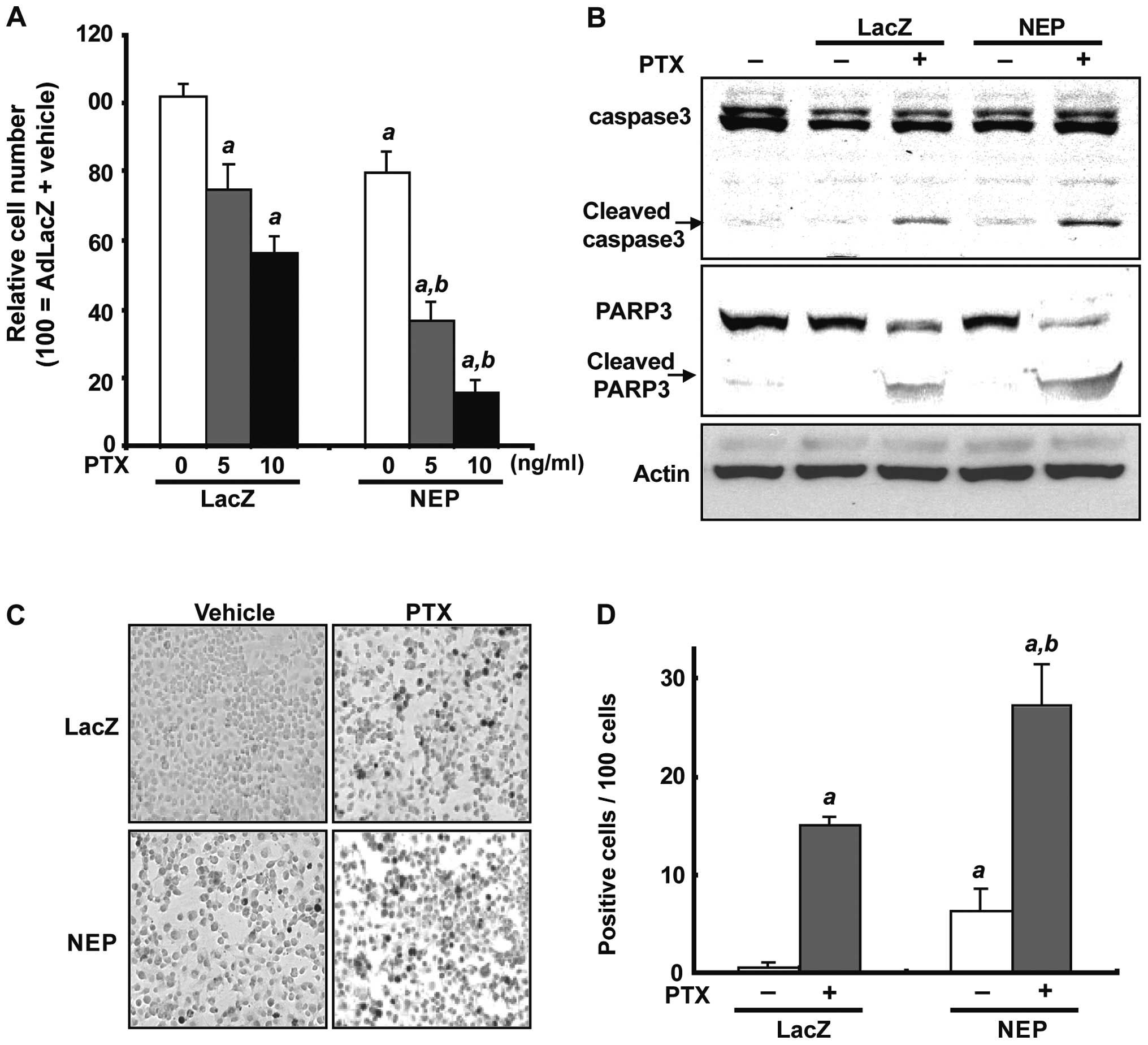
To assess the effect of NEP overexpression alone and
in combination with paclitaxel, DU145 cells were infected with
AdNEP or AdLacZ at 10 pfu/cell for 72 h and treated with paclitaxel
at concentrations of 5 or 10 ng/ml for 48 h. As expected,
paclitaxel significantly inhibited cell viability in cells infected
with AdLacZ compared to control cells treated with vehicle with a
relative viable cell number of 75.0±7.8% (5 ng/ml paclitaxel) or
56.4±4.8% (10 ng/ml paclitaxel) (p<0.01; Fig. 3A). In DU145 cells infected with
AdNEP, paclitaxel treatment at 5 and 10 ng/ml resulted in decreased
cell viability with a relative cell viability ratio of 36.7±5.3%
and 15.5±3.8%, respectively, which was statistically significant
compared to either control cells or cells infected with AdLacZ
(p<0.01; Fig. 3A). Measurement
of the AdNEP effect on paclitaxel-induced apoptosis was assessed by
examining cleavage of caspase-3 and Poly-ADP-ribose polymerase 1
(PARP-1) and counting the number of cells undergoing apoptosis
using a TUNEL assay. As shown in Fig.
3B, proteolytic fragments of caspase-3 and PARP-1 were evident
at 48 h following paclitaxel treatment in DU145 cells infected with
AdLacZ, which were increased in cells infected with AdNEP. The
number of TUNEL-positive cells per 100 cells was significantly
higher in DU145 cells treated with AdNEP plus paclitaxel
(28.3±4.16) compared with cells treated with AdLacZ plus paclitaxel
(16.3±1.52) or AdNEP plus vehicle (6.3±2.30) (P<0.01, Fig. 3C and D). These data demonstrate
that NEP overexpression using AdNEP significantly accelerates
paclitaxel-induced apoptosis in DU145 cells.
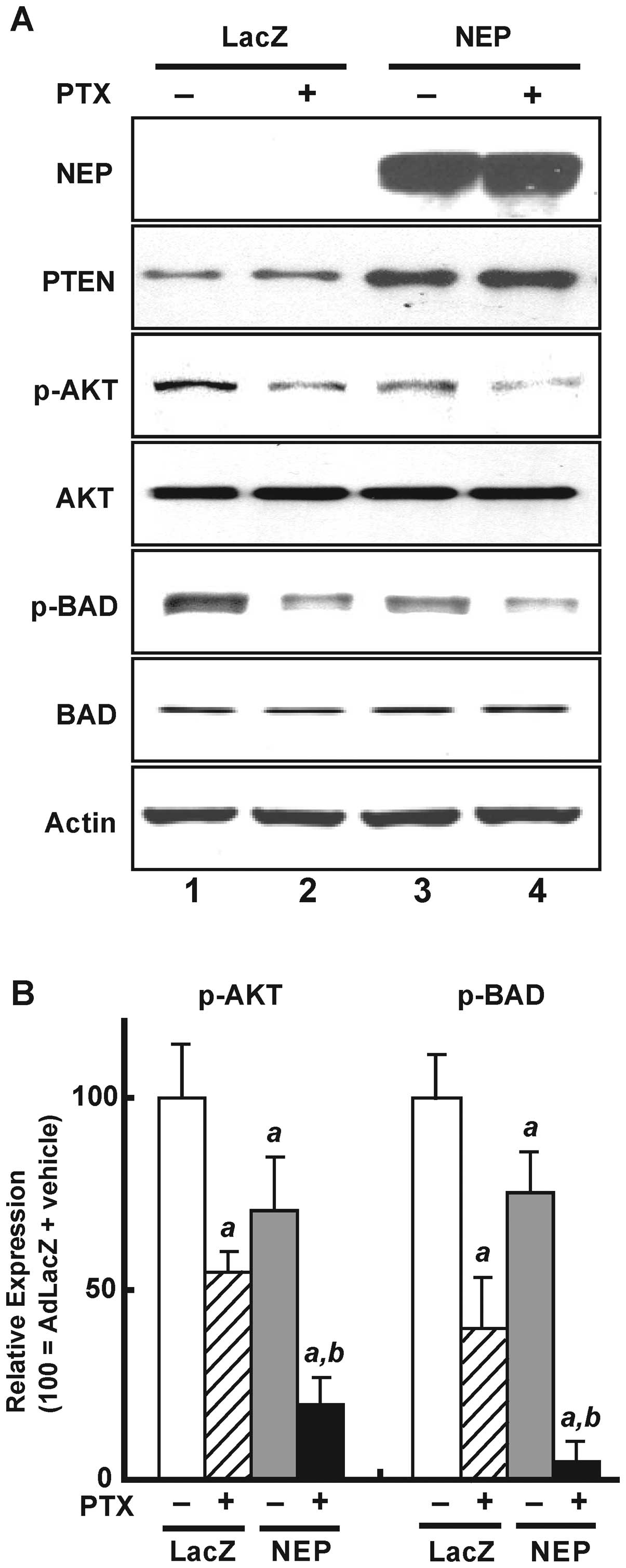
Effects of AdNEP on Akt signaling and BAD
expression
NEP stabilizes PTEN protein resulting in decreased
phosphor-Akt expression (9). As
shown in Fig. 4A, AdNEP infection
(rows 3 and 4) leading to NEP expression (first panel) resulted in
increased PTEN protein (panel 2, row 3) and decreased Akt
phosphorylation (panel 3, row 3) compared with cells infected with
AdLacZ (row 1). The addition of paclitaxel to cells infected with
AdLacZ (row 2) decreased Akt phosphorylation but did not affect
PTEN protein expression. The addition of paclitaxel to cells
infected with AdNEP (row 4) significantly further decreased Akt
phosphorylation (illustrated in Fig.
4B, left panel) (P<0.02).
We also examined expression of BAD, a pro-apoptotic
member of the Bcl-2 family and a substrate of Akt (19). As shown in Fig. 4A (panel 5) and B (right panel),
compared with DU145 cells infected with AdLacZ and treated with
vehicle (control), BAD phosphorylation significantly decreased in
cells infected with AdLacZ treated with paclitaxel (40.4±14.9% of
control; P<0.01), in cells infected with AdNEP treated with
vehicle (73.8±10.9% of control; P<0.01), and most dramatically
in cells infected with AdNEP and treated with paclitaxel (10.2±5.0%
of control; P<0.01). Together these results demonstrate that NEP
overexpression from adenoviral NEP infection significantly
potentiates paclitaxel-induced inactivation of the Akt/Bad
signaling pathway.
AdNEP inhibits DU145 xenograft growth in
vivo
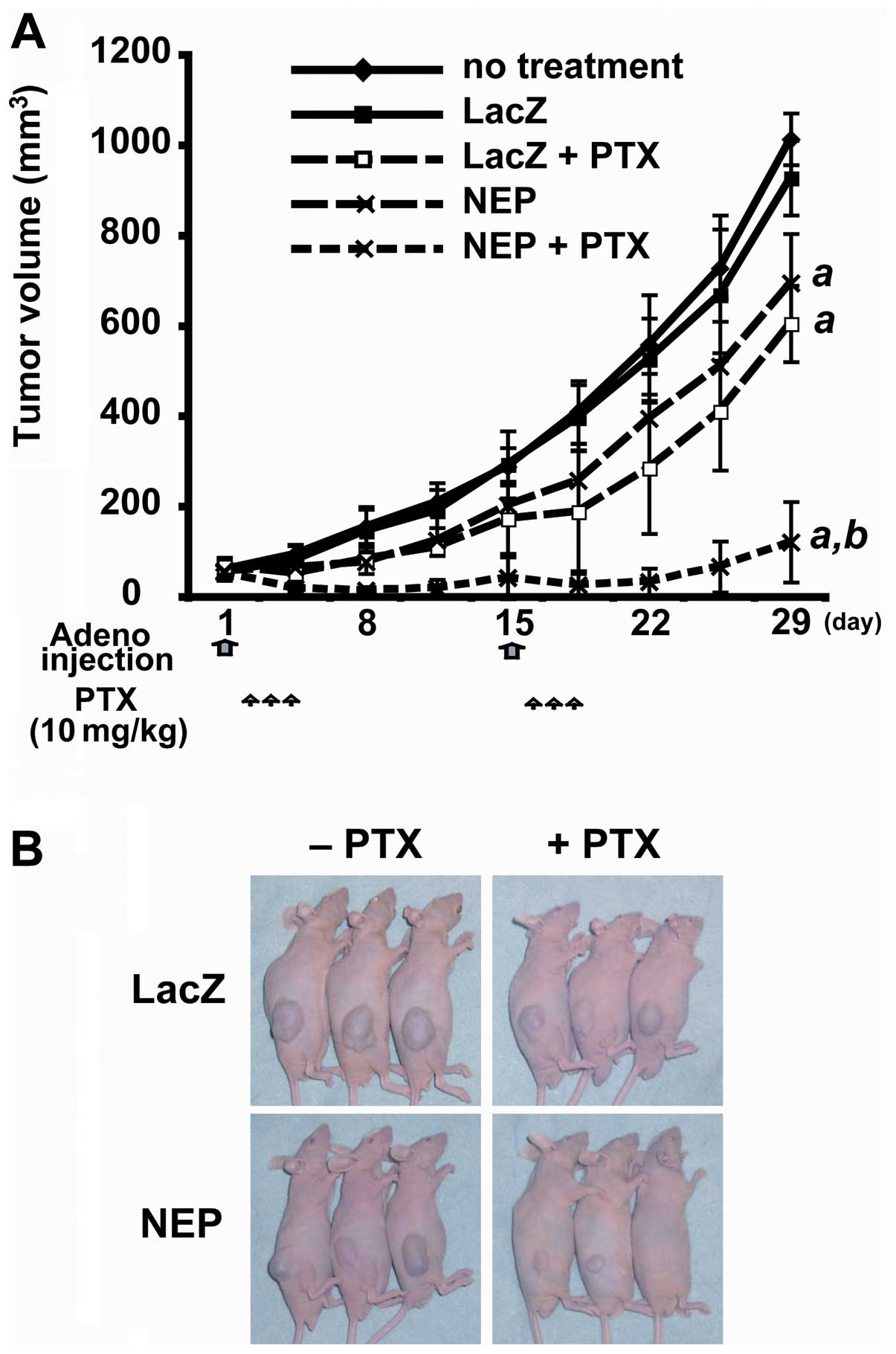
To evaluate the combined therapeutic efficacy of
AdNEP plus paclitaxel in vivo, we injected AdNEP or AdLacZ
intratumorally on days 1 and 15 into established DU145 xenografts
followed by intraperitoneal injection of paclitaxel or saline from
days 2 to 4 and from days 16 to 18. At day 28, tumor measurements
were performed and the animals sacrificed and the tumors were
dissected and weighed. There was no difference between the mean
tumor volume of the untreated control cohort (1022.6±255.5
mm3) and the AdLacZ plus saline cohort (920.2±238.2
mm3). However, the mean tumor volume of the AdNEP plus
saline cohort (653.9±230.3 mm3, P<0.01) and AdLacZ
plus paclitaxel cohort (575.9±176.6 mm3, P<0.01) were
significantly less (Fig. 5A and
B). Furthermore, the tumor volume of the cohort treated with
AdNEP plus paclitaxel (122.85±89.5 mm3) was markedly
less than cohorts treated with AdNEP plus saline or paclitaxel.
Similarly, the mean tumor weight of AdNEP plus paclitaxel cohort
was significantly less at 28 days than other treatment groups (data
not shown). Immunohistochemical staining and for the degree of
apoptosis by TUNEL assay of tumors resected at day 6 demonstrated a
significant increase in tumors injected with AdNEP and receiving
paclitaxel (data not shown). Tumor cells infected with AdNEP showed
an increase in NEP protein, an increase in PTEN protein expression
and a decrease in p-Akt (data not shown). Phospho-Akt was most
diminished in tumors injected with AdNEP and receiving paclitaxel.
Together, these results show that in an established prostate cancer
tumor xenograft, expression of NEP by injection with AdNEP together
with systemic paclitaxel results in an increase in PTEN protein, a
decrease in activated Akt, an increase in cell death and inhibition
of tumor growth.
Discussion
The current study was aimed at investigating the
potential as therapy for castration resistant prostate cancer of
adenoviral delivered NEP in combination with taxane chemotherapy.
We have previously demonstrated that recombinant NEP inhibits
prostate cancer cell growth in vitro(3); overexpression of NEP using a
tetracycline repressible system inhibits growth within the mouse
prostate in an orthotopic prostate cancer model (8); and injection of Lentiviral-NEP into
established xenograft tumors of the CWR22R castration-resistant
prostate cancer subline 22RV1 significantly inhibited tumor growth
(20). Lentiviral delivered NEP
was not effective in inhibiting growth of DU145 cells, although we
could demonstrate inhibition of angiogenesis resulting from a
decrease in FGF-2 levels as a consequence of NEP catalytic
inactivation (20). This lack of
effect in DU145 cells presumably resulted in part from the lower
level of NEP expression and enzyme activity in lentiviral infected
DU145 cells (96.57±10.2 pmol/mg/min) compared to lentiviral
infected 22RV1 cells (1171.27 pmol/mg/min). In contrast to
lentivirus, infection of DU145 cells with AdNEP in the current
study resulted in high levels of NEP protein expression and enzyme
activity (962.36±24.5 pmol/mg/min at an infection ratio of 10
pfu/cell). Based on our prior studies of combining recombinant NEP
with chemotherapeutic agents, we elected to study the effects of
adenoviral delivered NEP in combination with paclitaxel since
taxanes are effective in inhibiting prostate cancer growth. We
demonstrate that the combination of NEP and paclitaxel
significantly and dramatically inhibits Akt activation, DU145 cell
growth and tumorigenicity in a mouse model.
There are many potential pathways in which the
anti-tumor effects of taxanes and NEP may intersect and augment
each other. Akt signaling plays an important role in prostate
cancer cell survival and proliferation, as well as chemoresistance
(21). NEP negatively effects Akt
through multiple mechanisms, including directly interacting with
and stabilizing the PTEN protein (22) and catalytic inactivation of
neuropeptides, each leading to suppression of Akt
phosphorylation.
Studies have also shown that paclitaxel can also
inhibit Akt activation and synergize with other agents to induce
apoptosis and inhibit cell growth (15,23).
We similarly show that paclitaxel treatment results in lower levels
of activated Akt which are decreased further by NEP. This is
substantiated by increased cleavage of caspase-3 and PARP and
inhibition of BAD phosphorylation, resulting in a significant
increase in the number of cells undergoing apoptosis. AdNEP
augmented the effect of paclitaxel-induced apoptosis through
modulation of PTEN/Akt/Bad signaling both in DU145 cells in
vitro and in vivo in xenograft tumors. Our results are
similar to a previous report showing that siRNA silencing of Akt
enhances the antitumor effects of paclitaxel (24).
Microtubule stabilization through binding of taxane
to β-tubulin is the most widely accepted mechanism of taxane’s
anti-neoplastic action (25). Once
bound by taxanes, microtubules cannot be disassembled and this
static polymerization disrupts the normal mitotic process, arrest
cells in the G2M cycle phase ultimately leading to apoptosis.
Recent studies demonstrate that the cytoskeletal regulatory family
of ezrin-radixin-moesin (ERM) can also stabilize microtubule
networks (26,27). We and others have shown that the
N-terminal domain of NEP directly binds to ERM proteins (10,28,29),
and that cells expressing wild-type NEP demonstrate decreased
adhesion to hyaluronic acid and cell migration (10). Thus, it is conceivable that via its
interaction with ERM proteins, NEP also enhances taxane effects on
microtubule stabilization. Finally, the added effect of NEP and
paclitaxel may also result from a combined inhibition of protein
kinase C (PKC) signaling. PKC isoform PKCδ activity is required for
mitochondrial apoptosis in response to etoposide (30) and paclitaxel (31) in various cell types including
prostate cancer cells (32,33).
We previously reported that NEP stabilizes PKCδ protein expression
in prostate cancer cells by inhibiting neuropeptide-induced Src
signaling, which in the absence of NEP results in PKCδ protein
degradation (34). NEP may augment
paclitaxel-induced mitochondrial-dependent apoptosis through
increasing PKCδ expression and kinase activity.
The clinical feasibility of adenovirus gene delivery
has been demonstrated through intraprostatic injection of
adenovirus (35,36). The current study supports the
concept of local therapy involving AdNEP injected into recurrent
tumor in combination with systemic taxane chemotherapy. The
combination of NEP overexpression plus paclitaxel was very
effective in inhibiting DU145 tumor growth. This represents a novel
therapeutic strategy to target NEP-deficient prostate cancer cells,
and would likely be most effective in tumors that have retained
PTEN expression. Histone deacetylase inhibition can also increase
NEP expression in prostate cancer cells (37), suggesting that HDAC inhibitors may
represent another strategy to induce NEP expression. As prostate
cancer therapy evolves towards personalized treatments, the
approach of identifying NEP deficient tumors for AdNEP replacement
therapy in combination with systemic paclitaxel represents a novel
approach for treatment.
Acknowledgements
This study was supported by NIH Grants
CA80240 (RZ, RS, DMN) and DOD PC040758 (DMN), and the Robert H.
McCooey Memorial Cancer Research Fund.
References
|
1
|
Turner AJ: Exploring the structure and
function of zinc metallopeptidases: old enzymes and new
discoveries. Biochem Soc Trans. 31:723–727. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Maguer-Satta V, Besancon R and
Bachelard-Cascales E: Concise review: neutral endopeptidase (CD10):
a multifaceted environment actor in stem cells, physiological
mechanisms, and cancer. Stem Cells. 29:389–396. 2011. View Article : Google Scholar
|
|
3
|
Papandreou CN, Usmani B, Geng YP,
Bogenrieder T, Freeman RH, Wilk S, Finstad CL, Reuter VE, Powell
CT, Scheinberg D, Magill C, Scher HI, Albino AP and Nanus DM:
Neutral endopeptidase 24.11 loss in metastatic human prostate
cancer contributes to androgen-independent progression. Nat Med.
4:50–57. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Freedland SJ, Seligson DB, Liu AY, Pantuck
AJ, Paik SH, Horvath S, Wieder JA, Zisman A, Nguyen D, Tso CL,
Palotie AV and Belldegrun AS: Loss of CD10 (neutral endopeptidase)
is a frequent and early event in human prostate cancer. Prostate.
55:71–80. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Osman I, Yee H, Taneja SS, Levinson B,
Zeleniuch-Jacquotte A, Chang C, Nobert C and Nanus DM: Neutral
endopeptidase protein expression and prognosis in localized
prostate cancer. Clin Cancer Res. 10:4096–4100. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Usmani BA, Shen R, Janeczko M, Papandreou
CN, Lee WH, Nelson WG, Nelson JB and Nanus DM: Methylation of the
neutral endopeptidase gene promoter in human prostate cancers. Clin
Cancer Res. 6:1664–1670. 2000.PubMed/NCBI
|
|
7
|
Sumitomo M, Shen R, Walburg M, Dai J, Geng
Y, Navarro D, Boileau G, Papandreou CN, Giancotti FG, Knudsen B and
Nanus DM: Neutral endopeptidase inhibits prostate cancer cell
migration by blocking focal adhesion kinase signaling. J Clin
Invest. 106:1399–1407. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dai J, Shen R, Sumitomo M, Goldberg JS,
Geng Y, Navarro D, Xu S, Koutcher JA, Garzotto M, Powell CT and
Nanus DM: Tumor suppressive effects of neutral endopeptidase in
androgen-independent prostate cancer cells. Clin Cancer Res.
7:1370–1377. 2001.PubMed/NCBI
|
|
9
|
Sumitomo M, Iwase A, Zheng R, Navarro D,
Kaminetzky D, Shen R, Georgescu MM and Nanus DM: Synergy in tumor
suppression by direct interaction of neutral endopeptidase with
PTEN. Cancer Cell. 5:67–78. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Iwase A, Shen R, Navarro D and Nanus DM:
Direct binding of neutral endopeptidase 24.11 to
ezrin/radixin/moesin (ERM) proteins competes with the interaction
of CD44 with ERM proteins. J Biol Chem. 279:11898–11905. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Osman I, Dai J, Mikhail M, Navarro D,
Taneja SS, Lee P, Christos P, Shen R and Nanus DM: Loss of neutral
endopeptidase and activation of protein kinase B (Akt) is
associated with prostate cancer progression. Cancer. 107:2628–2636.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sumitomo M, Asano T, Asakuma J, Asano T,
Nanus DM and Hayakawa M: Chemosensitization of androgen-independent
prostate cancer with neutral endopeptidase. Clin Cancer Res.
10:260–266. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mancuso A, Oudard S and Sternberg CN:
Effective chemotherapy for hormone-refractory prostate cancer
(HRPC): present status and perspectives with taxane-based
treatments. Crit Rev Oncol Hematol. 61:176–185. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Orr GA, Verdier-Pinard P, McDaid H and
Horwitz SB: Mechanisms of Taxol resistance related to microtubules.
Oncogene. 22:7280–7295. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kim SH, Juhnn YS and Song YS: Akt
involvement in paclitaxel chemoresistance of human ovarian cancer
cells. Ann N Y Acad Sci. 1095:82–89. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Weng D, Song X, Xing H, Ma X, Xia X, Weng
Y, Zhou J, Xu G, Meng L, Zhu T, Wang S and Ma D: Implication of the
Akt2/survivin pathway as a critical target in paclitaxel treatment
in human ovarian cancer cells. Cancer Lett. 273:257–265. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang H, Yu D, Agrawal S and Zhang R:
Experimental therapy of human prostate cancer by inhibiting MDM2
expression with novel mixed-backbone antisense oligonucleotides: in
vitro and in vivo activities and mechanisms. Prostate. 54:194–205.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Horiguchi A, Zheng R, Goodman OB Jr, Shen
R, Guan H, Hersh LB and Nanus DM: Lentiviral vector neutral
endopeptidase gene transfer suppresses prostate cancer tumor
growth. Cancer Gene Ther. 14:583–589. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mitsiades CS, Mitsiades N and Koutsilieris
M: The Akt pathway: molecular targets for anti-cancer drug
development. Curr Cancer Drug Targets. 4:235–256. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Horiguchi A, Chen DY, Goodman OB Jr, Zheng
R, Shen R, Guan H, Hersh LB and Nanus DM: Neutral endopeptidase
inhibits prostate cancer tumorigenesis by reducing FGF-2-mediated
angiogenesis. Prostate Cancer Prostatic Dis. 11:79–87. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
De Souza PL, Russell PJ and Kearsley J:
Role of the Akt pathway in prostate cancer. Curr Cancer Drug
Targets. 9:163–175. 2009.PubMed/NCBI
|
|
22
|
Siepmann M, Kumar S, Mayer G and Walter J:
Casein kinase 2 dependent phosphorylation of neprilysin regulates
receptor tyrosine kinase signaling to Akt. PLoS One. 5:e131342010.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sumitomo M, Asano T, Asakuma J, Asano T,
Horiguchi A and Hayakawa M: ZD1839 modulates paclitaxel response in
renal cancer by blocking paclitaxel-induced activation of the
epidermal growth factor receptor-extracellular signal-regulated
kinase pathway. Clin Cancer Res. 10:794–801. 2004. View Article : Google Scholar
|
|
24
|
Priulla M, Calastretti A, Bruno P,
Azzariti A, Paradiso A, Canti G and Nicolin A: Preferential
chemosensitization of PTEN-mutated prostate cells by silencing the
Akt kinase. Prostate. 67:782–789. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
McGrogan BT, Gilmartin B, Carney DN and
McCann A: Taxanes, microtubules and chemoresistant breast cancer.
Biochim Biophys Acta. 1785:96–132. 2008.PubMed/NCBI
|
|
26
|
Naghavi MH, Valente S, Hatziioannou T, de
Los SK, Wen Y, Mott C, Gundersen GG and Goff SP: Moesin regulates
stable microtubule formation and limits retroviral infection in
cultured cells. EMBO J. 26:41–52. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Haedicke J, de Los SK, Goff SP and Naghavi
MH: The Ezrinradixin-moesin family member ezrin regulates stable
microtubule formation and retroviral infection. J Virol.
82:4665–4670. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Niv MY, Iida K, Zheng R, Horiguchi A, Shen
R and Nanus DM: Rational redesign of neutral endopeptidase binding
to merlin and moesin proteins. Protein Sci. 18:1042–1050. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Terawaki S, Kitano K and Hakoshima T:
Structural basis for type II membrane protein binding by ERM
proteins revealed by the radixin-neutral endopeptidase 24.11 (NEP)
complex. J Biol Chem. 282:19854–19862. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Reyland ME, Anderson SM, Matassa AA,
Barzen KA and Quissell DO: Protein kinase C delta is essential for
etoposide-induced apoptosis in salivary gland acinar cells. J Biol
Chem. 274:19115–19123. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Matassa AA, Carpenter L, Biden TJ,
Humphries MJ and Reyland ME: PKCdelta is required for
mitochondrial-dependent apoptosis in salivary epithelial cells. J
Biol Chem. 276:29719–29728. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sumitomo M, Ohba M, Asakuma J, Asano T,
Kuroki T, Asano T and Hayakawa M: Protein kinase Cdelta amplifies
ceramide formation via mitochondrial signaling in prostate cancer
cells. J Clin Invest. 109:827–836. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Majumder PK, Pandey P, Sun X, Cheng K,
Datta R, Saxena S, Kharbanda S and Kufe D: Mitochondrial
translocation of protein kinase C delta in phorbol ester-induced
cytochrome c release and apoptosis. J Biol Chem. 275:21793–21796.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sumitomo M, Shen R, Goldberg JS, Dai J,
Navarro D and Nanus DM: Neutral endopeptidase promotes phorbol
ester-induced apoptosis in prostate cancer cells by inhibiting
neuropeptide-induced protein kinase C delta degradation. Cancer
Res. 60:6590–6596. 2000.
|
|
35
|
Freytag SO, Stricker H, Peabody J, Pegg J,
Paielli D, Movsas B, Barton KN, Brown SL, Lu M and Kim JH:
Five-year follow-up of trial of replication-competent
adenovirus-mediated suicide gene therapy for treatment of prostate
cancer. Mol Ther. 15:636–642. 2007.PubMed/NCBI
|
|
36
|
Patel P, Young JG, Mautner V, Ashdown D,
Bonney S, Pineda RG, Collins SI, Searle PF, Hull D, Peers E,
Chester J, Wallace DM, Doherty A, Leung H, Young LS and James ND: A
phase I/II clinical trial in localized prostate cancer of an
adenovirus expressing nitroreductase with CB1984. Mol Ther.
17:1292–1299. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hong Y, Beckett C, Belyaev ND and Turner
AJ: The impact of amyloid precursor protein signalling and histone
deacetylase inhibition on neprilysin expression in human prostate
cells. Int J Cancer. 130:775–786. 2012. View Article : Google Scholar : PubMed/NCBI
|