Introduction
The uncontrolled growth of cancer cells is
frequently driven by the acquisition of activating mutations in
growth-promoting genes (1,2), inactivating mutations in
growth-inhibitory genes and epigenetic abnormalities that lead to
the altered expression of growth-modulating proteins (1,3).
These genetic or/and epigenetic changes can result in the
constitutive activation of signaling pathways that lead to
uncontrolled proliferation, metastasis and resistance to anticancer
therapy.
The epidermal growth factor receptor (EGFR)/Ras
pathway is a key signaling cascade involved in cell proliferation,
stress response, migration and cell cycle progression (4,5). The
constitutive activation of the EGFR/Ras pathway may result from
activating mutations in Ras (KRAS, HRAS, NRAS) and
RAF and via the mutation or overexpression of EGFR (6–9).
Cells harboring a constitutively activated EGFR/Ras pathway via
mutant Ras or Raf are known to be more sensitive to
MAPK/extracellular signal-regulated kinase (ERK) (MEK) inhibitors
(10–12); however, sensitivity in Ras and Raf
mutant cells is not uniform and even Ras and Raf mutant cells may
exhibit resistance to MEK inhibition (10,12,13).
Collectively, these data suggest that sensitivity to these agents
may be complex and dependent on a host of mutations, gene
expression patterns, or the inherent dependence of the cell to
aberrant signaling through the Ras/mitogen-activated protein kinase
(Ras/MAPK) pathway (14).
We previously reported that the treatment of 3 human
cancer lines with the MEK1/2 inhibitor, selumetinib (AZD6244,
ARRY-142886), sensitized the cells to ionizing radiation (IR)
(15). We observed an increase in
mitotic catastrophe in the cell lines exposed to selumetinib prior
to IR compared to those that were treated with IR alone. To further
evaluate the mechanisms by which the inhibition of the Ras/MAPK
pathway results in radiosensitization, we focused on autocrine
growth factors that signal through EGFR following radiation.
Ligands of EGFR have been shown to be secreted by different types
of cancer cells following IR, resulting in the autocrine activation
of the EGFR/Ras/MEK/MAPK pathways which can protect irradiated
cells from IR-induced death (16–18).
In the present study, we investigated the role of
secreted transforming growth factor-α (TGF-α), an EGFR ligand, on
the radiosensitization mediated by selumetinib in A549 cells (KRAS
mutant), in DU145 transfectants expressing wild-type Ras, and in
DU145 transfectants harboring a KRAS mutation. We hypothesized that
the interruption of MAPK signaling with selumetinib in
KRAS-transformed tumor cells would decrease the production of TGF-α
and prevent the secondary activation of the EGFR downstream
signaling pathways, a known resistance mechanism following the
inhibition of mutant Ras (19).
Our data support the notion that inhibiting MEK provides a means to
sensitize cells to IR through the interference of Ras/MAPK and
TGF-α signaling via EGFR.
Materials and methods
Cell lines and treatment
The A549 non-small cell lung cancer (NSCLC) and
DU145 (prostate cancer) cell lines were obtained from American Type
Culture Collection (ATCC; Manassas, VA). All cell lines were
verified by DNA fingerprinting and confirmed to be mycoplasma-free
by ATCC. Cells were cultured in RPMI-1640 medium (ATCC),
supplemented with 5% fetal bovine serum (Hyclone, Logan, UT). Cells
were maintained at 37°C, 5% CO2. Selumetinib, provided by
AstraZeneca (Macclesfield, UK), was reconstituted in DMSO and
stored at −20°C. Recombinant human TGF-α and anti-TGF-α antibodies
were purchased from R&D Systems (Minneapolis, MN) and EMD
Chemicals (Gibbstown, NJ), respectively. Cultures were irradiated
using a Pantak (Solon, OH) X-ray source at a dose rate of 1.55
Gy/min.
Plasmid and transfection
DU145 cells were transfected with an empty vector or
a plasmid expressing a hemagglutinin (HA)-tagged KRAS2A G12V
(Biomyx Technology, San Diego, CA) using the Nucleofector
transfection system (Amaxa Inc., Gaithersburg, MD) according to the
manufacturer’s instructions. Transfectants were placed under
selection with G418 (Invitrogen, Carlsbad, CA) and pooled stable
cell lines (DU145 vec, DU145 mut) were established. Transgene
expression was confirmed by western blot analysis using a HA
antibody.
Clonogenic assays
Cell cultures were trypsinized to generate a single
cell suspension and a specified number of cells were seeded into
6-well tissue culture plates. After allowing 6 h for attachment,
the cells were incubated with 250 nM selumetinib or DMSO (vehicle
control) for 16 h prior to IR. Anti-TGF-α antibody (final
concentration, 1 μg/ml) was added 30 min prior to IR to
neutralize endogenous TGF-α in the culture medium and recombinant
TGF-α (10 pg/ml) was added to the cultures immediately before IR.
Twelve to 14 days after seeding, colonies were stained with crystal
violet, the number of colonies containing at least 50 cells was
determined, and the surviving fractions were calculated. Survival
curves were generated after normalizing for cytotoxicity generated
by treatment alone for each independent experiment. Data presented
are the means ± SEM from at least 3 independent experiments. The
dose enhancement factor (DEF) was determined by taking the ratio of
the dose leading to a surviving fraction of 0.1 for untreated
versus treated cells (for each condition: neutralizing antibody,
selumetinib, or selumetinib + TGF-α).
Mitotic catastrophe
The presence of fragmented nuclei was used as the
criteria for defining cells undergoing mitotic catastrophe
(15,20). To visualize nuclear fragmentation,
cells were fixed with methanol for 15 min at −20°C and stained with
anti-α-tubulin monoclonal antibody (T6199; Sigma-Aldrich, St.
Louis, MO) followed by staining with FITC-conjugated secondary
antibody (Jackson ImmunuoResearch Laboratories Inc., West Grove,
PA). Nuclei were counterstained with DAPI. A total of 150 randomly
selected cells were analyzed from each treatment group and
photographed under an epifluorescence microscope. Nuclear
fragmentation was defined as the presence of >2 distinct nuclear
lobes within a single cell.
Western blot analysis
Cell extracts were prepared using RIPA buffer
(Pierce, Rockford, IL) containing protease inhibitors (Roche
Applied Science, Indianapolis, IN) and phosphatase inhibitors
(Sigma-Aldrich), followed by measurement of protein concentrations
by the Bradford method (Bio-Rad, Hercules, CA). Equal amounts of
protein were subjected to western blot analysis, and were probed
with the primary antibody indicated. ImageQuant software (GE
Healthcare, Pittsburgh, PA) was used to evaluate the relative
expression of each target protein normalized to actin.
ELISA
Culture supernatants were obtained from the cells
(1×104/100 μl) pre-treated with/without
selumetinib at various time-points after IR as indicated. In order
to obtain lysates from A549 tumors, tissue pieces were collected
from mice treated as indicated, and then homogenized in RIPA buffer
containing protease inhibitors. Soluble proteins were collected by
centrifugation (10,000g × 10 min) and followed by the measurement
of protein concentrations by the Bradford method (Bio-Rad). The
levels of soluble TGF-α, amphiregulin and heregulin in the culture
supernatants or lysates from the tumor tissues were assessed using
the human Quantikine ELISA kit for TGF-α and the human DuoSet ELISA
kits for amphiregulin and heregulin (R&D Systems) according to
the manufacturer’s instructions.
Mouse xenograft model
Animal experiments were conducted in accordance with
the principles and procedures outlined in the National Institutes
of Health (NIH) Guide for the Care and Use of Laboratory Animals.
Nude mice, 4-6 weeks old [National Cancer Institute (NCI),
Frederick, MD], were injected subcutaneously with A549 cells
(1×106/100 μl PBS/mouse) on the lateral aspect of
the rear leg. When tumors reached 250 mm3 mice were
treated with selumetinib (50 mg/kg) or the vehicle control by oral
gavage. Restrained mice were irradiated to the hind leg 4 h after
selumetinib administration using a Pantak irradiator. Tumor tissue
was excised at the indicated time-points. For ELISA, tumors were
homogenized in RIPA buffer containing protease inhibitors to
extract soluble proteins. For immunohistochemistry, tumors were
fixed with 10% neutral-buffered formalin and embedded in
paraffin.
Immunohistochemistry
Sections (6-μm-thick) mounted on
poly-L-lysine coated glass slides were deparaffinized, rehydrated,
incubated in 3% H2O2 for 5 min, and boiled
for 30 min in 10 mM sodium citrate buffer (pH 6.0; Vector
Laboratories, Burlingame, CA). TGF-α expression was assayed with an
indirect immunoperoxidase method (ImmPRESS, Vector Laboratories)
using anti-TGF-α polyclonal antibody (1:50 dilution; Abcam,
Cambridge, MA). Following treatment with 3,3-diaminobenzidine
(Roche) sections were counterstained in hematoxylin, dehydrated
through graded alcohols, cleared in xylenes, and mounted in
Permount (Sigma-Aldrich).
Statistical analysis
In vitro experiments were repeated thrice,
and statistical analysis was carried out using a Student’s t-test.
Data are presented as the means ± SD. A probability level of
P<0.05 was considered to indicate a statistically significant
difference.
Results
Exposure to selumetinib alters the
activation of EGFR after radiation
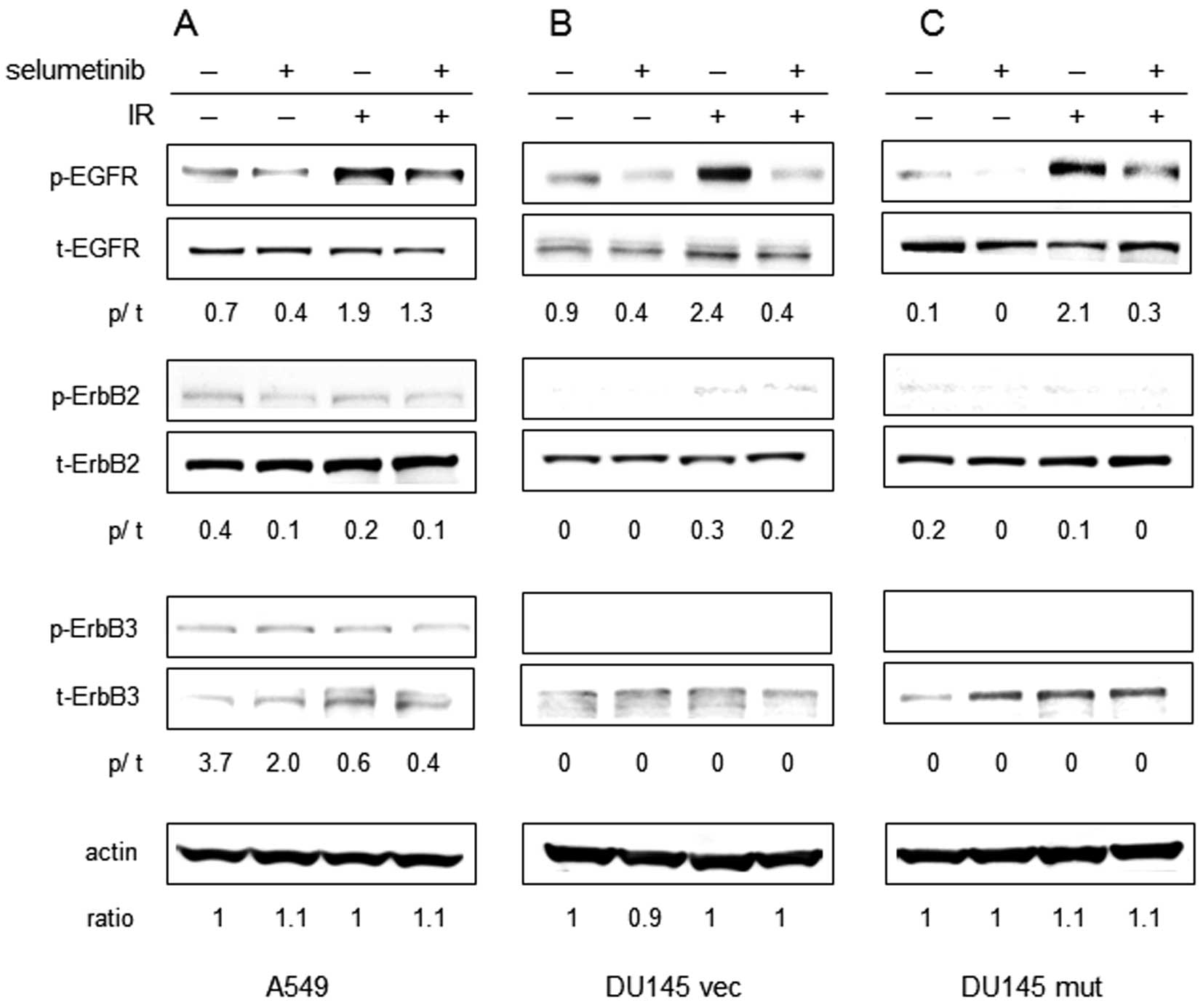
EGFR, ErbB2 and ErbB3 are members of the ErbB
receptor family of tyrosine kinases expressed on the cell surface.
The heterodimerazation or homodimerization of these receptors plays
an important role in the association of EGFRs with ligands and
downstream signaling pathways. To investigate whether the exposure
to selumetinib alters the magnitude of ErbB receptor activation in
response to radiation in our cell lines, the level of
phosphorylation of each receptor was examined at 24 h following
radiation in the A549, DU145 vec and DU145 mut cells (Fig. 1). As expected, irradiation resulted
in the increased phosphorylation of EGFR (Tyr845) in all 3 cell
lines. There was no evidence of the altered phosphorylation of
ErbB2 (Tyr1221/1222) and ErbB3 (Tyr1197) following irradiation. The
phosphorylation of EGFR decreased significantly following treatment
with selumetinib in the presence or absence of IR in all 3 cell
lines. Treatment with selumetinib moderately reduced the
phosphorylation of ErbB2 in the A549 and DU145 mut cells (both Ras
mutants) with or without IR. ErbB3 phosphorylation appeared
minimally affected by selumetinib treatment in A549 cells and was
not detectable in the DU145 vec or DU145 mut cells.
Selumetinib inhibits EGFR ligand
secretion through the downregulation of metalloproteinase tumor
necrosis factor (TNF)-α converting enzyme (TACE) activation
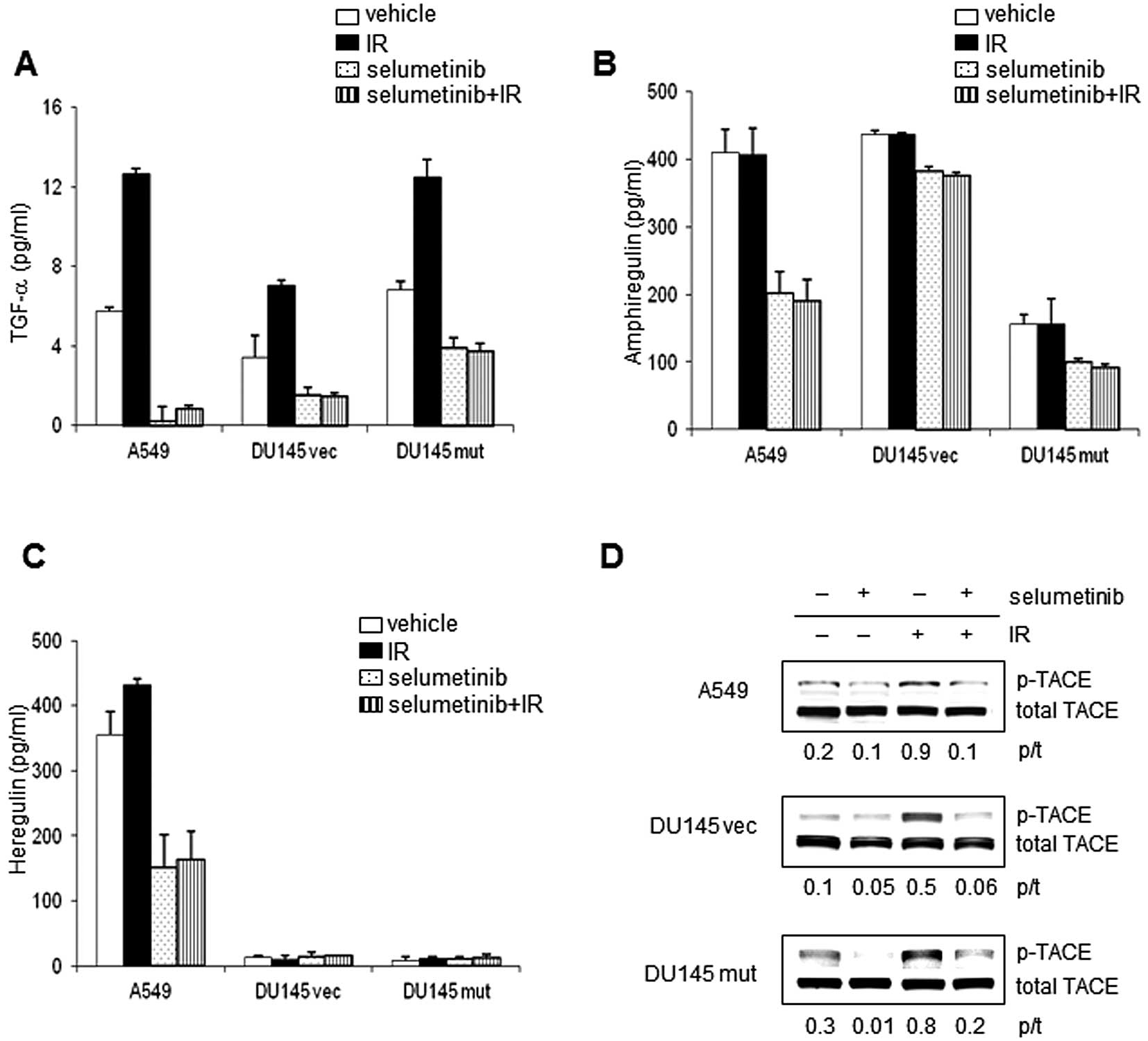
TGF-α, amphiregulin and heregulin are soluble
factors which have been linked to radiation resistance in
Ras-transformed cells (17,21).
To investigate whether the inhibition of MEK can alter the
elaboration EGFR ligands, levels of soluble TGF-α, heregulin and
amphiregulin were assessed by ELISA in the A549, DU145 vec and
DU145 mut cells treated with IR (4 Gy) and/or selumetinib (Fig. 2). TGF-α secretion was induced by IR
in all 3 cell lines. DU145 mut cells secreted significantly higher
levels of TGF-α than DU145 vec cells, at a level similar to the
A549 cell line. MEK inhibition reduced TGF-α secretion in all 3
cell lines, under irradiated and unirradiated conditions (Fig. 2A). In all the cell lines, treatment
with selumetinib reduced TGF-α secretion after IR to levels lower
than those observed under untreated conditions. Amphiregulin
secretion was not induced by radiation in the 3 cell lines tested;
however, basal levels of amphiregulin secretion were inhibited by
MEK inhibition (Fig. 2B). Although
the induction of heregulin secretion in response to IR was
statistically significant compared to the control (p<0.008), the
relative increase was minimal. Furthermore, the increase in
phosphorylation of ErbB3 in irradiated A549 cells compared to the
unirradiated controls was minimal. Basal and radiation-induced
levels of heregulin were markedly inhibited by selumetinib
(Fig. 2C).
The secretion of soluble EGFR ligands is known to be
regulated by TACE, also known as ADAM-17 (22,23).
The activation (phosphorylation) of TACE occurred after IR in all 3
cell lines (Fig. 2D). Treatment
with selumetinib was sufficient to inhibit the phosphorylation of
TACE in the presence or absence of IR in all 3 cell lines. The
activation of TACE has been reported to require an association with
phosphorylated ERK1/2 (24,25).
The association between TACE and phosphorylated ERK1/2 was
increased by 1.8-fold, 4 h after IR in the A549 cells compared to
the unirradiated cells (data not shown). Treatment with selumetinib
decreased this association following IR to levels lower than those
observed in the controls, possibly due to a reduction in the amount
of phosphorylated ERK.
TGF-α autocrine signal is required for
cancer cell survival and xenograft tumor growth following
radiation
To investigate the importance of radiation-induced
TGF-α for clonogenic survival in our cell lines, a neutralizing
antibody against TGF-α was added to the cultures 30 min prior to
IR. As shown in Fig. 3, the
neutralization of endogenous TGF-α decreased clonogenic survival in
the A549, DU145 vec and DU145 mut cells, suggesting that TGF-α
increases the survival of irradiated tumor cells. The dependency on
TGF-α in the post-irradiation setting was greatest in the
KRAS mutant cells with TGF-α neutralization providing a DEF
of 1.4 for the A549, 1.3 for the DU145 mut, and 1.12 for the DU145
vec cells.
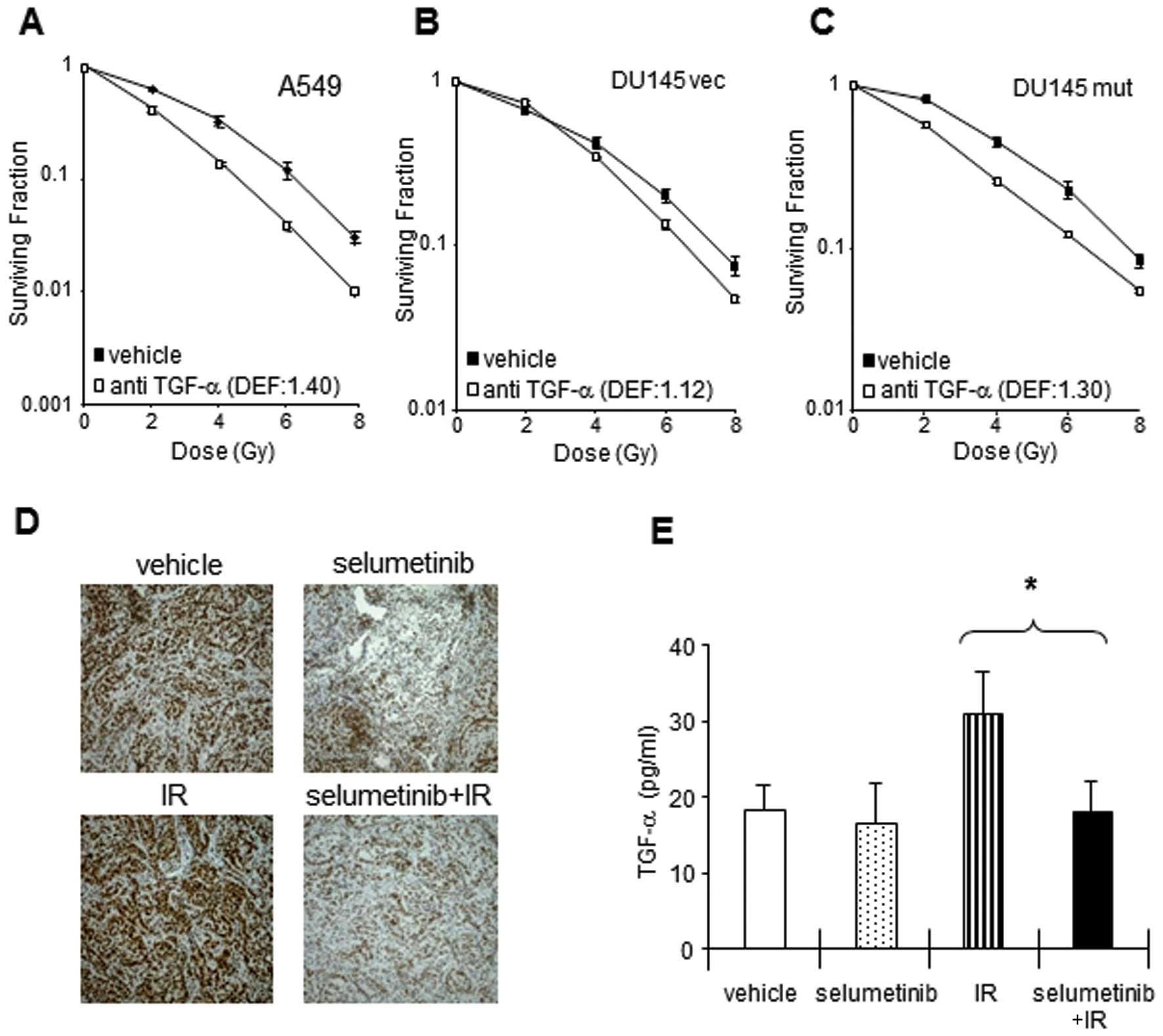 | Figure 3Increased TGF-α expression in
response to IR is required for A549 cell survival in in
vitro cultures and in vivo tumors. (A–C) Neutralizing
anti-TGF-α antibody decreased the clonogenic survival in tumor
cells exposed to IR. (A) A549, (B) DU145 vec and (C) DU145 mut
cells were exposed to neutralizing TGF-α antibody (final
concentration, 1 μg/ml), followed 30 min later by IR, and
incubated at 37°C with 5% CO2. Colony-forming efficiency
was determined 10 to 14 days later and survival curves were
generated after normalizing for cell killing by anti-TGF-α alone.
Clonogenic survival after IR was inhibited by the elimination of
soluble TGF-α in A549, DU145 vec and DU145 mut cells. The data
represent the means of 3 independent experiments. PE, plating
efficiency with selumetinib; DEF, dose enhancement factor. Points,
mean; bars, + SE. (D–E) Effects of selumetinib on TGF-α induction
in response to IR in A549 xenograft tumors. When A549 tumors
reached 250 mm3 in size, the mice were randomized into 4
groups: vehicle, selumetinib, IR (3 Gy), or selumetinib plus IR.
Selumetinib was administered by mouth (oral gavage) in a single
dose of 50 mg/kg. IR (3 Gy) was delivered 4 h after selumetinib
treatment. Tumors were harvested at 24 h after IR and subjected to
TGF-α IHC (D) or ELISA (E). The levels of endogenous TGF-α were
increased 24 h after IR in A549 xenografts. Selumetinib treatment
decreased the level of endogenous TGF-α with/without IR in A549
tumors. Columns, mean; bars, ± SE. |
We previously reported an enhancement in
radiosensitivity with MEK inhibition in vivo in an A549
xenograft model (15). To
determine whether MEK inhibition is capable of reducing TGF-α
elaboration in vivo, the levels of TGF-α following treatment
with IR and/or selumetinib were assessed by immunohistochemistry
and ELISA in A549 xenografts (Fig.
3E). The total expression of TGF-α was lower in the xenografts
from mice treated with selumetinib alone or selumetinib in
combination with IR compared to basal levels. Given the
heterogeneity of the expression of TGF-α observed after
immunohistochemical assay (Fig.
3E), further confirmation of a reduction in TGF-α expression
was achieved with the more quantitative approach of ELISA. TGF-α
expression in the xenograft tumors was increased 24 h after IR.
Pre-treatment with selumetinib 4 h prior to IR resulted in
decreased TGF-α expression to a level similar to that achieved with
selumetinib alone.
TGF-α partially rescues tumor cells from
selumetinib-mediated radiation sensitization
The results presented above suggest that the
radiation-induced secretion of TGF-α may act as a survival factor,
and that MEK inhibition may block the elaboration of basal and
radiation-induced TGF-α levels. To confirm that TGF-α remains an
important survival factor following IR in the setting of MEK
inhibition, clonogenic assays were performed with selumetinib with
or without the addition of TGF-α. Radiosensitization with
selumetinib was evident to a greater extent in KRAS mutant cell
lines with a DEF of 1.9 in the A549 cell line and 1.5 in DU145 mut
(DEF of 1.5) compared to 1.13 in the DU145 vec line. The addition
of exogenous TGF-α rescued all the cell lines from
selumetinib-enhanced radiation-induced cytotoxicity (Fig. 4A–C) with almost complete rescue in
the DU145 vec and DU145 mut lines and partial rescue in the A549
cell line.
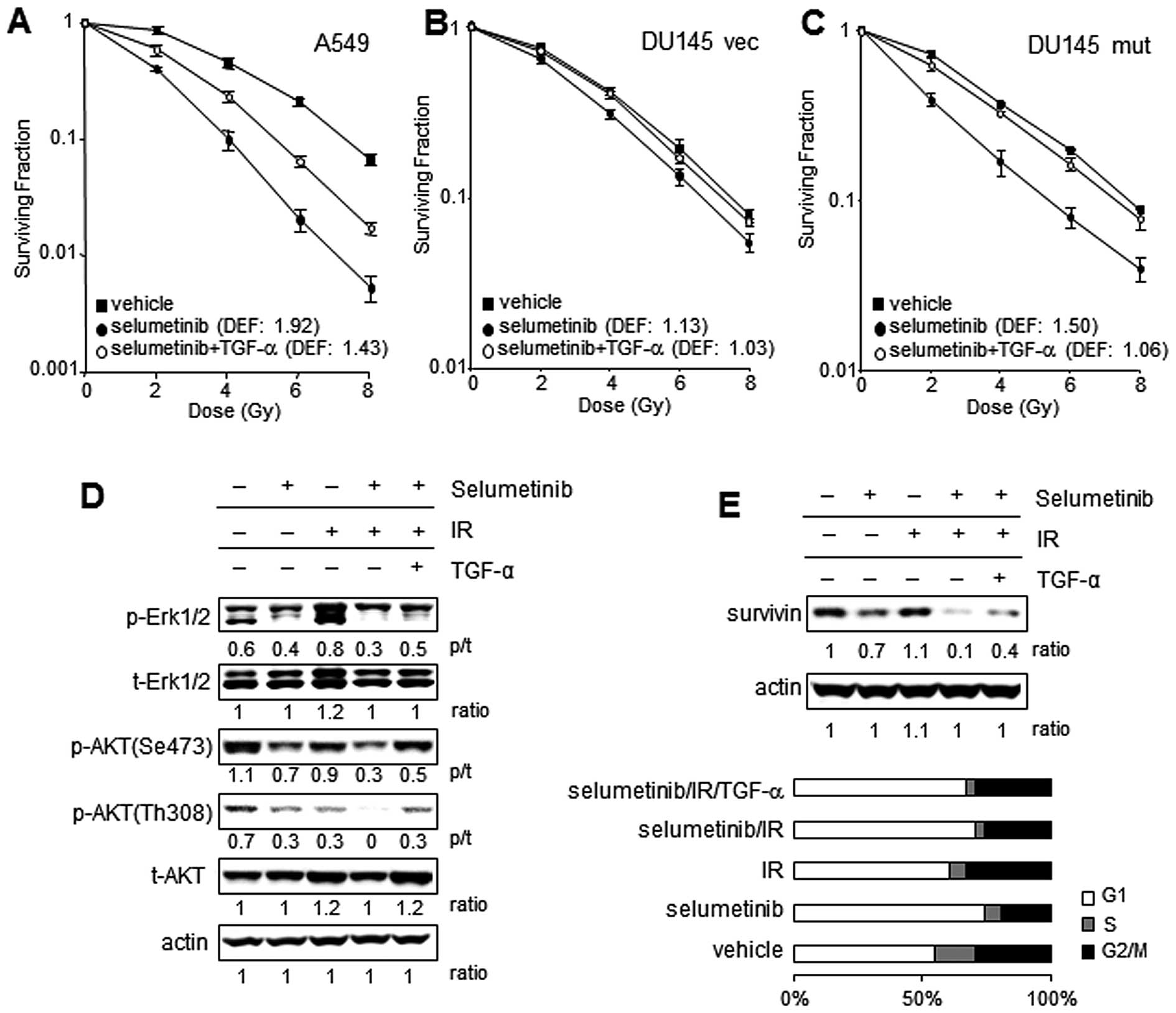 | Figure 4Exogenous TGF-α supplementation
restores EGFR downstream signaling after selumetinib-mediated
inhibition in irradiated tumor cells. (A–C) Clonogenic assays:
Cells were exposed to 250 nM selumetinib or the vehicle control for
16 h, irradiated with graded doses of X-rays and supplemented with
recombinant human TGF-α (rhTGF-α) (10 pg/ml) or PBS immediately
after IR. Colony-forming efficiency was determined 10 to 14 days
later and survival curves were generated after normalizing for cell
killing by selumetinib alone. The data represent the means of 3
independent experiments. Significant sensitizations to IR with
selumetinib were observed in (A) A549 and (C) DU145 mut cells
compared to (B) DU145 vec cells. Exogenous TGF-α partially rescued
the A549 cells and the DU145 transfectant cells almost completely
from selumetinib-induced radiosensitization. DEF, dose enhancement
factor; points, mean ± SE. (D and E) Restoration of EGFR downstream
signals by exogenous TGF-α. A549 cells were exposed to 250 nM
selumetinib or the vehicle control for 16 h, irradiated and
harvested 24 h following IR (4 Gy) for immunoblotting. To evaluate
the downstream signaling after EGFR activation by TGF-α binding,
the levels of phosphorylated AKT and ERK1/2 were assessed in
lysates obtained from the cells treated with various combinations
of IR, TGF-α and selumetinib. (D) The phosphorylation of ERK1/2 was
increased by IR, while the phosphorylation of AKT was slightly
decreased by IR. The effects of the inhibition by selumetinib were
assessed in the cells treated with or without IR. The addition of
TGF-α to the selumetinib-treated cells partially restored the
phosphorylation of AKT and ERK1/2. The levels of survivin, and
EGFR/MAPK downstream target molecule were also investigated. (E)
Survivin expression was partially decreased by selumetinib, and
significantly by the combination treatment with IR. Exogenous TGF-α
reversed the inhibitory effects on survivin expression in A549
cells treated with selumetinib and IR. As survivin expression is
related to the cell cycle, cell cycle profiles of cells treated
with IR, selumetinib and selumetinib/IR were investigated 24 h
after IR. The expression levels of survivin were not a result of
the number of cells in each phase of the cell cycle between the
cells treated with selumetinib alone and selumetinib/IR. |
To further evaluate the molecular events underlying
the ability of TGF-α to rescue cells from radiation sensitization
by MEK inhibition, the A549 cell line was investigated. Our primary
hypothesis was that TGF-α is depleted by MEK inhibition and
recovery to post-irradiation levels activates the EGFR pro-survival
signaling pathway which permits the rescue of irradiated cells. To
examine whether the addition of exogenous TGF-α restores the EGFR
signaling altered by selumetinib in irradiated A549 cells, the
phosphorylation of EGFR and the downstream molecules, ERK1/2 and
AKT, and the expression levels of survivin were assessed by
immunoblotting (Fig. 4D and E).
The exposure to radiation increased phosphorylated ERK1/2, but
decreased the phosphorylation of AKT at serine 473 and threonine
308 in A549 cells at 24 h. Treatment with selumetinib decreased the
levels of ERK1/2 phosphorylation and AKT phosphorylation in the
presence or absence of IR. The addition of TGF-α to the cells
treated with selumetinib and IR partially restored the
phosphorylation of ERK1/2, although it completely recovered AKT
phosphorylation inhibited by selumetinib in irradiated A549 cells.
This suggests that ERK1/2 was inhibited continuously after the
addition of TGF-α due to selumetinib remaining in the culture.
Survivin is known to be a prosurvival molecule, a known downstream
target of the MAPK/ERK pathway and is involved in the progression
of mitosis. As shown in Fig. 4E,
survivin expression was markedly inhibited by the combination
treatment with selumetinib and IR compared to that observed after
single treatment with IR or selumetinib. The addition of TGF-α
partially restored the expression of survivin in the A549 cells
exposed to selumetinib and IR. The expression of survivin is
related to the cell cycle, with reported dominant expression in the
G2/M phase (26). Cell cycle
analysis confirmed that there was no marked alteration in the
percentage of cells in G2/M following treatment with selumetinib
and/or TGF-α in irradiated A549 cells, suggesting that the
increased survivin expression was not a result of cell cycle
changes (Fig. 4E).
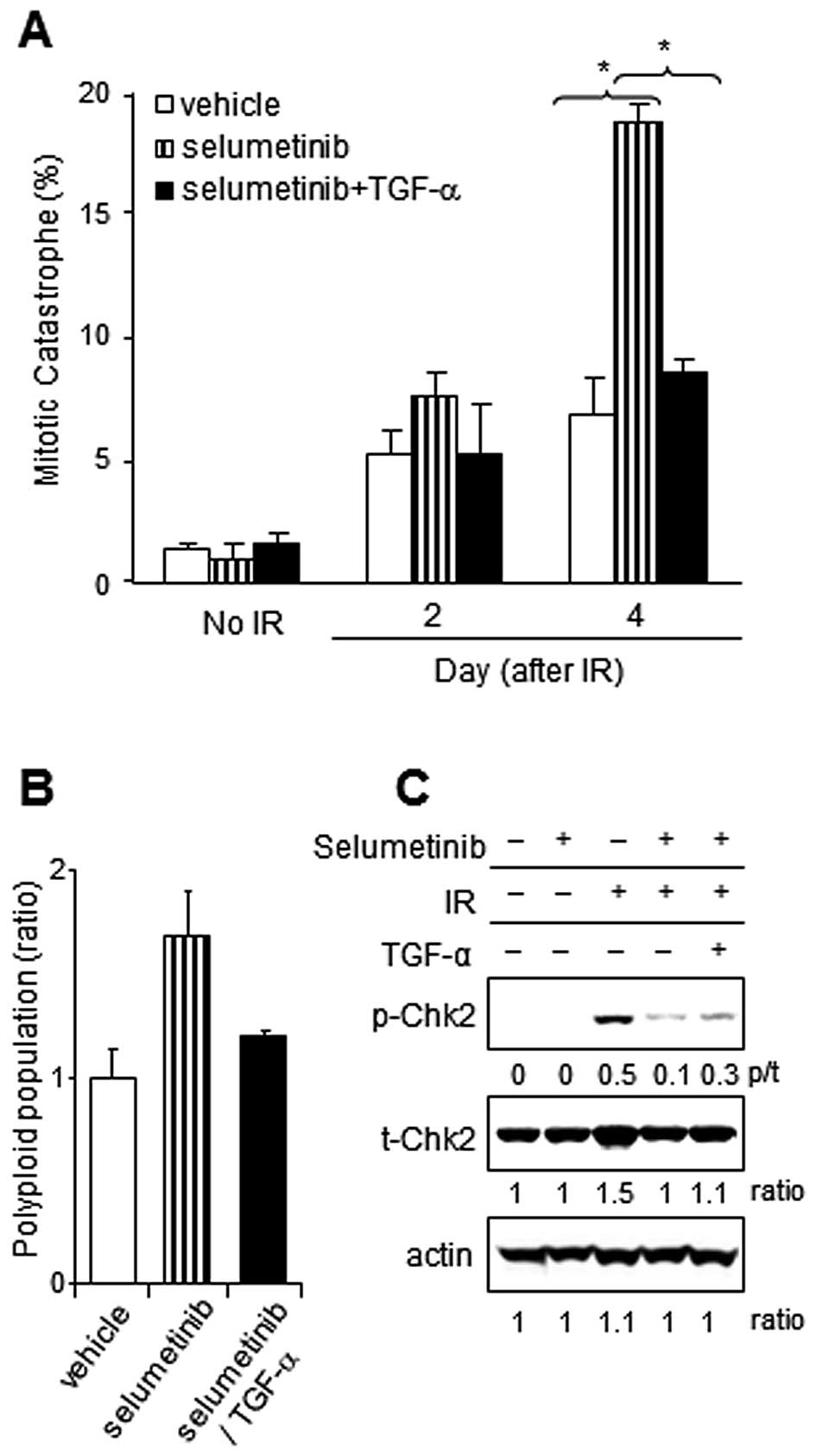
TGF-α supplementation reduces mitotic
catastrophe after IR in selumetinib-treated tumor cells
In our previous study, an increase in the number of
cells undergoing mitotic catastrophe was defined as an important
mechanism of cell death after the combined treatment with
selumetinib and IR compared to either treatment alone (15). In the present study, the mitotic
catastrophe induced by the combination of selumetinib and IR was
inhibited significantly by TGF-α supplementation in A549 cells
(Fig. 5A). The increase in the
polyploid population with selumetinib supplementation was confirmed
at 24 h after IR exposure in A549; however, it was reduced by the
addition of TGF-α (Fig. 5B). To
evaluate the mechanism by which TGF-α protects cancer cells from
mitotic catastrophe, we examined the expression and phosphorylation
of checkpoint kinase 2 (Chk2), which is known as both a regulator
of mitotic catastrophe (27) and
as a kinase that phosphorylates survivin in response to DNA damage
(34). As observed in Fig. 5C, the phosphorylation of Chk2 was
detected in irradiated A549 cells, but not in unirradiated cells.
The IR-induced Chk2 phosphorylation was inhibited by selumetinib
treatment and was partially restored with the addition of
TGF-α.
Discussion
The acute effects of IR-induced DNA damage have been
well documented. Since DNA double-strand breaks (DSBs) are
considered to be a lethal event following IR (28,29),
much emphasis has been placed on the evaluation of DNA repair and
events occurring early after IR, when novel radiation modifiers are
evaluated. We previously reported the radiosensitizing effects of
selumetinib in human cancer cell lines of 3 different histologies
(15). We observed enhanced
sensitization to radiation with selumetinib treatment in KRAS
mutant cell lines in this, as well as our previous study. We also
observed that prolonged post-IR exposure to selumetinib increased
the degree of sensitization in all 3 cell lines (data not shown).
These findings suggest that constitutively active KRAS and
prolonged MEK/ERK1/2 activation enhances survival at later
time-points after IR (>24 h) at a time when DNA damage repair is
likely to be complete. These data suggest that a mechanism other
than DNA repair is responsible for the radiosensitizing effect of
selumetinib treatment, consistent with our prior findings (15). In our previous report, we presented
data from 3 cell lines with varying levels of sensitization to IR
with selumetinib. These data suggest that the presence of a KRAS
mutation may increase the efficacy of radiation sensitization
observed with selumetinib. To explore the hypothesis that the
efficacy of selumetinib as a radiation sensitizer is greater in
cells harboring mutant KRAS, we generated a DU145 cell line
harboring an activating KRAS mutant. As we expected, the
radiosensitizing effects observed with selumetinib were greater in
DU145 cells harboring mutant KRAS compared to Ras wild-type cells.
However, since we observed a degree of sensitization in the Ras
wild-type cells, these data also suggest that the inhibition of the
activation of downstream effectors of Ras after IR can sensitize
even Ras wild-type cell lines, albeit to a lesser degree.
TGF-α has been well described as a factor that
promotes cell proliferation, survival, transformation and protects
against radiation-induced damage by activating EGFR downstream
intermediates, such as AKT and ERK1/2 (18,21,30).
Of note, the transformation of human mammary epithelial cells by
the c-Ha-Ras gene has been shown to enhance TGF-α expression
(31), and the presence of mutant
KRAS also promotes TGF-α secretion through TACE activation.
Treatment with exogenous TGF-α or conditioned medium collected from
cells with oncogenic KRAS has been shown to reverse the
radiosensitizing effect of KRAS inhibition (21). Collectively, these former findings
suggest that ErbB ligands produced downstream of Ras/MEK/ERK1/2
signaling play an important role in the radiation sensitization
obtained with selumetinib in Ras-transformed cells.
The radiation-induced phosphorylation of EGFR and
TGF-α secretion coupled with the finding that treatment with a
neutralizing TGF-α antibody resulted in radiosensitization. This
confirms the importance of TGF-α as a resistance factor to IR,
particularly mutant KRAS. With the knowledge that TGF-α is a
resistance factor after IR in our cell lines, we investigated the
secretion of TGF-α after IR in the setting of treatment with
selumetinib. We confirmed that selumetinib reduced TGF-α secretion
when delivered alone or in combination with radiation. This
suggests that selumetinib may have particular efficacy in tumor
cells that rely on basal or inducible TGF-α autocrine signaling.
The ability of selumetinib to inhibit TGF-α secretion was confirmed
in A549 xenografts, which we have previously shown to be sensitive
to selumetinib-induced radiation sensitization (15). Selumetinib treatment diminished the
basal levels of TGF-α expression and abrogated an increase
following IR. Collectively, these data also suggest that TGF-α
expression may be a useful biomarker of drug effects in the setting
of radiation sensitization, particularly in KRAS mutant cell
lines.
Our data indicate that a possible underlying
mechanism for the reduction in TGF-α secretion with selumetinib
treatment is the inhibition of the activation of TACE, likely due
to a reduction in the association between TACE and phosphorylated
ERK1/2. Phosphorylated, active TACE was increased by radiation and
decreased by selumetinib treatment in all 3 cell lines. We
therefore suggest that the inhibition of pro-TGF-α shedding in
irradiated tumor cells treated with MEK inhibition results in the
reduction in soluble TGF-α, in turn resulting in the downregulation
of the PI3K/AKT pathway. The addition of TGF-α to
selumetinib-treated tumor cells following IR restored AKT
phosphorylation and partially overcame MEK1/2 inhibition induced by
radiation sensitization.
The addition of TGF-α reduced the radiosensitizing
effects of MEK1/2 inhibition in all cell lines evaluated.
Collectively, these findings suggest that TGF-α is a critical
survival factor in KRAS mutant cells and that the radiosensitizing
effects of MEK1/2 inhibition are partially related to the
inhibition of TGF-α. The rescue of selumetinib and
radiation-treated cells after the addition of TGF-α was not
complete, suggesting that ERK1/2 activation downstream of TGF-α is
partly responsible for its pro-survival effect. Alternatively, it
is possible that other unknown pathways and molecules targeted by
selumetinib are involved in the radiation sensitizing effect.
Additionally, ERK phosphorylation is known to be important in a
variety of cellular functions, including cell cycle progression and
assembly of the mitotic spindle (32) that are known to be important in the
recovery after IR.
In our previous study (15), radiosensitization with selumetinib
treatment resulted from mitotic catastrophe rather than apoptosis.
We therefore investigated the effects of TGF-α supplementation on 2
known regulators of mitotic catastrophe, Chk2 and survivin. Chk2
phosphorylated on threonine 68 accumulates in BRCA1 nuclear bodies
during cell cycle arrest induced by DNA damage (27). Phosphorylated Chk2 facilitated by
ATM/ATR in response to radiation-induced DNA damage mediates G2/M
cell cycle arrest (33). Of note,
the conflict provided by Chk2 inhibition between cell cycle
progression and DNA damage could lead to mitotic catastrophe
(27). Chk2 has also been shown to
correlate with survivin expression and regulate its localization by
mediating phosphorylation in response to DNA damage (34–35).
Selumetinib treatment diminished most of the radiation-induced Chk2
phosphorylation (Fig. 5C) and
enhanced mitotic catastrophe (Fig.
5A) in A549 cells. Survivin is also known to protect against
apoptosis and mitotic catastrophe (36,37).
When survivin levels are high, cells are protected against drugs
that induce apoptosis and mitotic catastrophe. In this study, the
level of survivin protein was reduced with selumetinib treatment in
irradiated A549 cells and restored by TGF-α. Consistent with these
results, the addition of TGF-α to A549 cells treated with
selumetinib and radiation, reduced mitotic catastrophe to a level
similar to that observed in the untreated irradiated cells.
Taken together, these findings suggest that
selumetinib has greater efficacy in KRAS mutant compared to Ras
wild-type cells and that this effect may be due to a relatively
greater dependence of KRAS mutant cells on TGF-α autocrine
signaling following IR. Based on these observations, TGF-α appears
to act as a survival factor following radiation, preventing mitotic
catastrophe at later time-points via the activation of EGFR. Our
results raise the possibility that the radiosensitizing effects of
selumetinib may be predicted by determining the dependence of
cancer cells on TGF-α after IR.
Abbreviations:
|
TGF-α
|
transforming growth factor-α
|
|
ERK1/2
|
extracellular signal-regulated kinases
1/2
|
|
EGFR
|
epidermal growth factor receptor
|
|
MAPK
|
mitogen-activated protein kinase
|
|
MEK1/2
|
MAPK/extracellular signal-regulated
kinase 1/2
|
|
TACE
|
tumor necrosis factor-α converting
enzyme
|
|
Chk2
|
checkpoint kinase 2
|
|
IR
|
radiation
|
|
TNF-α
|
tumor necrosis factor-α
|
|
NIH
|
National Institutes of Health
|
|
NCI
|
National Cancer Institute
|
Acknowledgements
This study was supported by the
Intramural Research Program of the National Cancer Institute,
National Institutes of Health.
References
|
1
|
Adamson ED: Oncogenes in development.
Development. 99:449–471. 1987.
|
|
2
|
Grander D: How do mutated oncogenes and
tumor suppressor genes cause cancer? Med Oncol. 15:20–26. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Thomas RK, Baker AC, Debiasi RM, Winckler
W, Laframboise T, Lin WM, Wang M, Feng W, Zander T, MacConaill L,
et al: High-throughput oncogene mutation profiling in human cancer.
Nat Genet. 39:347–351. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yarden Y: The EGFR family and its ligands
in human cancer. signalling mechanisms and therapeutic
opportunities. Eur J Cancer. 37:S3–S8. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Singh AB and Harris RC: Autocrine,
paracrine and juxtacrine signaling by EGFR ligands. Cell Signal.
17:1183–1193. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Oliveira C, Velho S, Moutinho C, Ferreira
A, Preto A, Domingo E, Capelinha AF, Duval A, Hamelin R, Machado
JC, et al: KRAS and BRAF oncogenic mutations in MSS colorectal
carcinoma progression. Oncogene. 26:158–163. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kumar R, Angelini S and Hemminki K:
Activating BRAF and N-Ras mutations in sporadic primary melanomas:
an inverse association with allelic loss on chromosome 9. Oncogene.
22:9217–9224. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Davies H, Bignell GR, Cox C, Stephens P,
Edkins S, Clegg S, Teague J, Woffendin H, Garnett MJ, Bottomley W,
et al: Mutations of the BRAF gene in human cancer. Nature.
417:949–954. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shigematsu H and Gazdar AF: Somatic
mutations of epidermal growth factor receptor signaling pathway in
lung cancers. Int J Cancer. 118:257–262. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Solit DB, Garraway LA, Pratilas CA, Sawai
A, Getz G, Basso A, Ye Q, Lobo JM, She Y, Osman I, et al: BRAF
mutation predicts sensitivity to MEK inhibition. Nature.
439:358–362. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Leboeuf R, Baumgartner JE, Benezra M,
Malaguarnera R, Solit D, Pratilas CA, Rosen N, Knauf JA and Fagin
JA: BRAFV600E mutation is associated with preferential sensitivity
to mitogen-activated protein kinase kinase inhibition in thyroid
cancer cell lines. J Clin Endocrinol Metab. 93:2194–2201. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Davies BR, Logie A, McKay JS, Martin P,
Steele S, Jenkins R, Cockerill M, Cartlidge S and Smith PD: AZD6244
(ARRY-142886), a potent inhibitor of mitogen-activated protein
kinase/extracellular signal-regulated kinase kinase 1/2 kinases:
mechanism of action in vivo, pharmacokinetic/pharmacodynamic
relationship, and potential for combination in preclinical models.
Mol Cancer Ther. 6:2209–2219. 2007.
|
|
13
|
Pohl G, Ho CL, Kurman RJ, Bristow R, Wang
TL and Shih IeM: Inactivation of the mitogen-activated protein
kinase pathway as a potential target-based therapy in ovarian
serous tumors with KRAS or BRAF mutations. Cancer Res.
65:1994–2000. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dry JR, Pavey S, Pratilas CA, Harbron C,
Runswick S, Hodgson D, Chresta C, McCormack R, Byrne N, Cockerill
M, et al: Transcriptional pathway signatures predict MEK addiction
and response to selumetinib (AZD6244). Cancer Res. 70:2264–2273.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chung EJ, Brown AP, Asano H, Mandler M,
Burgan WE, Carter D, Camphausen K and Citrin D: In vitro and in
vivo radiosensitization with AZD6244 (ARRY-142886), an inhibitor of
mitogen-activated protein kinase/extracellular signal-regulated
kinase 1/2 kinase. Clin Cancer Res. 15:3050–3057. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hagan M, Yacoub A and Dent P: Ionizing
radiation causes a dose-dependent release of transforming growth
factor alpha in vitro from irradiated xenografts and during
palliative treatment of hormone-refractory prostate carcinoma. Clin
Cancer Res. 10:5724–5731. 2004. View Article : Google Scholar
|
|
17
|
Dent P, Reardon DB, Park JS, Bowers G,
Logsdon C, Valerie K and Schmidt-Ullrich R: Radiation-induced
release of transforming growth factor alpha activates the epidermal
growth factor receptor and mitogen-activated protein kinase pathway
in carcinoma cells, leading to increased proliferation and
protection from radiation-induced cell death. Mol Biol Cell.
10:2493–2506. 1999.
|
|
18
|
Dent P, Yacoub A, Fisher PB, Hagan MP and
Grant S: MAPK pathways in radiation responses. Oncogene.
22:5885–5896. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cengel KA, Voong KR, Chandrasekaran S,
Maggiorella L, Brunner TB, Stanbridge E, Kao GD, McKenna WG and
Bernhard EJ: Oncogenic K-Ras signals through epidermal growth
factor receptor and wild-type H-Ras to promote radiation survival
in pancreatic and colorectal carcinoma cells. Neoplasia. 9:341–348.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Castedo M, Perfettini JL, Roumier T,
Andreau K, Medema R and Kroemer G: Cell death by mitotic
catastrophe: a molecular definition. Oncogene. 23:2825–2837. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Grana TM, Sartor CI and Cox AD: Epidermal
growth factor receptor autocrine signaling in RIE-1 cells
transformed by the Ras oncogene enhances radiation resistance.
Cancer Res. 63:7807–7814. 2003.PubMed/NCBI
|
|
22
|
Borrell-Pages M, Rojo F, Albanell J,
Baselga J and Arribas J: TACE is required for the activation of the
EGFR by TGF-alpha in tumors. EMBO J. 22:1114–1124. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lee DC, Sunnarborg SW, Hinkle CL, Myers
TJ, Stevenson MY, Russell WE, Castner BJ, Gerhart MJ, Paxton RJ,
Black RA, et al: TACE/ADAM17 processing of EGFR ligands indicates a
role as a physiological convertase. Ann N Y Acad Sci. 995:22–38.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rousseau S, Papoutsopoulou M, Symons A,
Cook D, Lucocq JM, Prescott AR, O’Garra A, Ley SC and Cohen P:
TPL2-mediated activation of ERK1 and ERK2 regulates the processing
of pre-TNF alpha in LPS-stimulated macrophages. J Cell Sci.
121:149–154. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gomez MI, Seaghdha MO and Prince AS:
Staphylococcus aureus protein A activates TACE through
EGFR-dependent signaling. EMBO J. 26:701–709. 2007. View Article : Google Scholar
|
|
26
|
Otaki M, Hatano M, Kobayashi K, Ogasawara
T, Kuriyama T and Tokuhisa T: Cell cycle-dependent regulation of
TIAP/msurvivin expression. Biochim Biophys Acta. 1493:188–194.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Castedo M, Perfettini JL, Roumier T,
Yakushijin K, Horne D, Medema R and Kroemer G: The cell cycle
checkpoint kinase Chk2 is a negative regulator of mitotic
catastrophe. Oncogene. 23:4353–4361. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Valerie K and Povirk LF: Regulation and
mechanisms of mammalian double-strand break repair. Oncogene.
22:5792–5812. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ward JF, Joner EI and Blakely WF: Effects
of inhibitors of DNA strand break repair on HeLa cell
radiosensitivity. Cancer Res. 44:59–63. 1984.PubMed/NCBI
|
|
30
|
Toulany M, Dittmann K, Kruger M, Baumann M
and Rodemann HP: Radioresistance of K-Ras mutated human tumor cells
is mediated through EGFR-dependent activation of PI3K-AKT pathway.
Radiother Oncol. 76:143–150. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Martinez-Lacaci I, Kannan S, De Santis M,
Bianco C, Kim N, Wallace-Jones B, Ebert AD, Wechselberger C and
Salomon DS: RAS transformation causes sustained activation of
epidermal growth factor receptor and elevation of mitogen-activated
protein kinase in human mammary epithelial cells. Int J Cancer.
88:44–52. 2000. View Article : Google Scholar
|
|
32
|
Horne MM and Guadagno TM: A requirement
for MAP kinase in the assembly and maintenance of the mitotic
spindle. J Cell Biol. 161:1021–1028. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Rainey MD, Black EJ, Zachos G and
Gillespie DA: Chk2 is required for optimal mitotic delay in
response to irradiation-induced DNA damage incurred in G2 phase.
Oncogene. 27:896–906. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lopergolo A, Tavecchio M, Lisanti S, Ghosh
JC, Dohi T, Faversani A, Vaira V, Bosari S, Tanigawa N, Delia D,
Kossenkov AV, Showe LC and Altieri DC: Chk2 phosphorylation of
survivin-DeltaEx3 contributes to a DNA damage-sensing checkpoint in
cancer. Cancer Res. 72:3251–3259. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ghosh JC, Dohi T, Raskett CM, Kowalik TF
and Altieri DC: Activated checkpoint kinase 2 provides a survival
signal for tumor cells. Cancer Res. 66:11576–11579. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Shi X, Wang D, Ding K, Lu Z, Jin Y, Zhang
J and Pan J: GDP366, a novel small molecule dual inhibitor of
survivin and Op18, induces cell growth inhibition, cellular
senescence and mitotic catastrophe in human cancer cells. Cancer
Biol Ther. 9:640–650. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Asanuma H, Torigoe T, Kamiguchi K,
Hirohashi Y, Ohmura T, Hirata K, Sato M and Sato N: Survivin
expression is regulated by coexpression of human epidermal growth
factor receptor 2 and epidermal growth factor receptor via
phosphatidylinositol 3-kinase/AKT signaling pathway in breast
cancer cells. Cancer Res. 65:11018–11025. 2005. View Article : Google Scholar
|