Introduction
Smad4 is a key mediator of the transforming growth
factor-β (TGF-β) superfamily that is involved in the control of
cell proliferation, differentiation and apoptosis (1,2).
Smad4 oligomerizes with the R-Smads (Smad2 and 3), phosphorylated
by the type I TGF-β receptor (TβRI), to form transcriptional
complexes Smad2/3-Smad4 that translocate to the nucleus, where they
bind to the promoters of target genes, activating or repressing
their transcription (3,4).
TGF-β is normally expressed and secreted in
epithelial follicular thyroid cells, where it controls the
differentiated phenotype, inhibits iodide trapping (5,6), and
thyroglobulin synthesis (7),
exercising some of these effects through Smad signaling (8,9).
TGF-β is also the negative regulator of thyrocyte proliferation and
is able to antagonize the mitogenic effects of the main growth
factors in thyroid cells of rat (7,10–12),
of porcine (13) and of human
(14), delaying progression during
the mid-late G1 phase (12,15).
Impairment of the TGF-β signaling at the level of
Smad genes is common in human carcinomas. Absent or decreased
expression of Smad4 has been demonstrated in various cancers,
including pancreatic, colorectal, head and neck (16,17),
and, more recently, in papillary thyroid carcinomas (PTCs)
(18), suggesting that the TGF-β
signaling functions as a tumor suppressor. On the other hand, TGF-β
can exhibit tumor-promoting effects as observed in prostate and
skin cancer progression (19,20)
and in papillary thyroid carcinomas (21). We recently demonstrated that a
Smad4 mutation, Smad4 C324Y, isolated from nodal metastases of
papillary thyroid carcinoma, causes an increase of TGF-β signaling
responsible for the acquisition of transformed phenotype and
invasive behaviour in thyroid cells stable expressing this mutation
(22). The TGF-β inhibitory growth
response is also reduced in these cells, this finding is consistent
with the observation that when Smad4 C324Y mutation is expressed in
thyroid cells it exerts a clear pro-oncogenic function. The fine
regulation of thyrocytes growth and differentiation reflects a
critical balance between the promotion and suppression of cell
division. TSH has been shown to stimulate, through the activation
of its receptor, more than one signal transduction pathway, most
notably the adenylcyclase/cAMP (cyclic adenosine monophosphate)
pathway. cAMP seems to account for the mitogenic effects of TSH in
human thyroid cells, mediated by the activation of cAMP-dependent
protein kinase A (PKA) (23,24).
Therefore, the FRTL-5 cell line, that maintains in vitro all
the markers of thyroid cell differentiation, represents an
excellent model to study the mechanism regulating thyroid cell
proliferation, because they require stimulating factors like TSH or
insulin for their growth.
In this study, we demonstrated that the stable
expression of Smad4 C324Y mutation in thyroid cells is responsible
of TSH independent growth ability, without any modulation of
thyroid specific genes, like thyroglobulin (TG). This response is
caused by an increase in oligomerization of Smad4 with Smad3,
responsible, in turn, for an increased phosphorylation of CREB,
necessary for the mitogenic actions of TSH.
Materials and methods
Reagents and constructs
Dulbecco’s modified Eagle’s medium (DMEM), Coon’s
modified Ham’s F-12 medium, PBS, bovine serum (BS), trypsin-EDTA,
L-glutamine 100X (200 mM), the six-hormone mixture (6H) containing:
TSH (10 mU/ml), insulin (10 μg/ml), hydrocortisone
(10−8 M), transferrin (5 μg/ml),
glycyl-L-histidyl-L-lysine acetate (10 ng/ml), and somatostatin (10
μg/ml) were purchased from Sigma-Aldrich (St. Louis, MO,
USA). The human recombinant TGF-β1 isoform was purchased by
PeproTech (Rocky Hill, NJ, USA). Phenilmethylsulfonil fluoride
(PMSF), protease cocktail inhibitor containing 1 mg/ml leupeptin,
2.5 mg/ml aprotinin, 1 mg/ml benzamide hydrochloride and
phosphatase inhibitor cocktail were purchased from Sigma-Aldrich.
Antibodies to Green Fluorescent Protein (GFP FL), cyclin D1 (C20),
cyclin D1 (H-295) and β-actin (C4) from Santa Cruz Biotechnology
(Santa Cruz, CA, USA). Antibodies to the phosphorylated form of
Smad3 (P-Smad3) from Cell Signaling Technology (Beverly, MA, USA),
to P-CREB (phospho S133) and CREB from Abcam (Cambridge, UK).
Horseradish peroxidase conjugated secondary antibodies were
purchased from Sigma-Aldrich. Transfections were performed using
lipofectin kit provided by Lipofectamine Plus Gibco-BRL, Life
Technologies (Rockville, MD, USA). Stable clones were obtained by
selection with geneticine G418 (Invitrogen-Life Technologies,
Carlsbad, CA, USA). GFP-tagged Smad4 constructs were obtained
subcloning the human SMAD4 gene in Clontech (Palo Alto, CA,
USA) pEGFPC3 vector, which allows in frame fusion to the C terminus
of GFP.
Cell cultures
The FRTL-5 (ATCC #8305) were kindly provided by Dr
L.D. Kohn (NHI, Bethesda, MD). This cell line, diploid between
their 5th and 25th passage, maintains the functional
characteristics of iodide uptake, thyroglobulin synthesis and
cyclic nucleotide metabolism over prolonged periods of culture and
doubling time of approximately 36 h (25). These cells were grown as previously
described (8) in W/O supplemented
with 5% calf serum and six-hormone mixture (6H) containing: TSH (10
mU/ml), insulin (10 μg/ml), hydrocortisone (10−8
M), transferrin (5 μg/ml), glycyl-L-histidyl-L-lysyne
acetate (10 ng/ml), and somatostatin (10 μg/ml). Clones
obtained by stable transfection of the FRTL-5 cells with the
expression vector pEGFPC3 containing the human Smad4 cDNA wt or
mutated, tagged with GFP, or with pEGFPC3 empty vector, were grown
in F-12 Coon’s modification medium supplemented with 5% bovine
serum and 6H mixture in presence of 500 μg/ml of G418, as
previously described (22). MDA
MB468 (26), breast cancer cell
lines purchased from American Type Culture Collection (ATCC,
Rockville, MD), were grown in DMEM supplemented with 10% BS. All
cells were maintained in continuous monolayer cultures at 37°C and
5% CO2, expanded up to 70–80% confluent, treated or not
with human recombinant TGF-β1 (10 ng/ml) and then employed for the
experiments as described below.
RNA isolation and analysis
RNA was extracted using Tri Reagent (Sigma-Aldrich),
following the manufacturer’s instructions. Using 1 μg RNA,
cDNAs were synthesized using MuLV Reverse Transcriptase (Applied
Biosystems, NJ, USA) and random primers (Roche, Mannheim, Germany)
according to the manufacturer’s instructions. The primers used in
the amplification of rat TG (TG forward
5′-TGCCCACCCAGAATCAAGGAAC-3′, reverse
5′-TGAAGCCAAAGGTACCCACAACTG-3′) and rat GAPDH, as internal control
(GAPDH forward 5′-TTCACCACCATGGAGAAGGCT-3′, reverse GAPDH
5′-ACAGCCTTGGCAGCACCAGT-3′) were designed to cross intron-exon
junctions. Each experiment was repeated three times using different
total RNA extracts. TG and GAPDH bands were analyzed using Bio-Rad
Laboratories software. Data were collected in terms of average
intensity of bands of TG amplicon per average intensity of
GAPDH.
Proliferation assays
The cell proliferation was assessed by
cell-counting. Briefly, 10×104 cells were seeded into
35-mm plate and left overnight at 37°C in a humidified incubator
with 5% CO2. After a starvation in 4H/0.5% BS medium for
24 h, culture medium was changed in F-12 Coon’s modification medium
supplemented with 0.5% BS and 5H mixture (6H mixture without TSH or
insulin). Growth curves were obtained counting the cells at time
zero (T0), 24, 48, 72 and 96 h. Cells were counted three times by
two independent investigators. Inter-observer variation was below
5%. Values represent mean of triplicate determination ± SD of three
experiments.
Cell cultures and transient transfection
analysis
MDA MB468 (26)
were transiently transfected with pEGFPC3-Smad4 wt or pEGFPC3-Smad4
C324Y using lipofectin technique following the manufacturer’s
instructions. After 24 h incubation, transfected cells were used in
immunoprecipitation experiments and western blot assays.
Immunoblot analysis and
immunoprecipitation
Subconfluent cells, transfected or not, were treated
or untreated with 10 ng/ml of TGF-β1 for 60 min. Protein extracts
were obtained using ice-cold TNE extraction buffer (50 mM Tris-HCl
pH 7.8, 150 mM NaCl, 1 mM EDTA, 1% Triton X-100) supplemented with
1 mM PMSF, protease and phosphatase cocktail inhibitors. Protein
lysates (60 μg) were subjected to immunoblot analysis as
previously described (27) using
primary antibodies to GFP FL (1:500), P-CREB (1:2,000), CREB
(1:1,000), cyclin D1 (1:1,000), P-Smad3 (1:500) and β-actin
(1:5,000). Membranes were, then, incubated with anti-rabbit
(1:50,000) or anti-mouse (1:10,000) horseradish
peroxidase-conjugated secondary antibodies. The western blots were
revealed by chemiluminescence using the Super Signal kit from
Pierce (Rockford, IL, USA) according to the manufacturer’s
instructions and visualized on CL-Xposure Film (Pierce). For
immunoprecipitation experiments, 1 mg of total protein extracted as
previously described, was precleared with protein-A Sepharose CL-4B
(GE Healthcare, Uppsala, Sweden) beads and then immunoprecipitated
with anti-GFP polyclonal antibody (Sigma-Aldrich). Immunocomplexes,
aggregated with 50 μl of protein-A Sepharose CL-4B, were
washed four times with 1 ml of buffer. The pellets were boiled in
Laemmli buffer for 5 min, and the proteins were resolved under
reducing conditions by 8% SDS-PAGE and subjected to immunoblot
analysis as previously described (27) using primary antibodies to P-Smad3
and GFP FL. P-CREB, cyclin D1 bands were analyzed using Bio-Rad
Laboratories software. Data, obtained from three different protein
extracts, were collected in terms of average intensity of bands of
each protein per average intensity of bands of CREB or β-actin.
Immunohistochemistry analysis
Thyroid tissue sections of lymph node metastasis of
PTC from which derives the C324Y mutation of Smad4 and 3 lymph node
metastases of PTC without mutation, collected at the Anatomic
Pathology of Department of Experimental Medicine of Sapienza
University of Rome from 1996 to 2001, were studied using the
catalyzed signal amplification protocol (Dako A/S, Glostrup,
Denmark) (28). Slides were
incubated in a humidified chamber overnight at 41°C with 1:100
dilution of polyclonal serum to cyclin D1 (H-295). The
peroxidase-based LSAB2 Detection kit (Dako A/S), followed by
haematoxylin counterstaining, has been used to visualize the
reactions. Written informed consent was obtained from each patient
according to Helsinki Declaration and approved by the local ethics
committee.
Statistical analysis
All statistical analyses were performed using JMP
Software purchased by Statistical Discovery SAS Institute. Data
were analyzed by Student’s t-test (P<0.05, statistical
significance).
Results
Proliferation and differentiation in
Smad4 C324Y stable clones
The main positive regulators of thyroid growth and
function are TSH and insulin (29). To investigate if Smad4 C324Y
interferes with cell growth mediated by TSH or insulin, growth
curve assays have been performed in FRTL-5 stable transfected with
the expression vector pEGFPC3 containing wt or mutated Smad4 cDNAs,
tagged with GFP, as previously described by D’Inzeo et
al(22). Stable clones C324Y
2M, C324Y cl6, C324Y cl8, V1M, Smad4 wt 6M clones and FRTL-5, were
grown in absence of TSH or insulin. Cells were starved in 4H/0.5%
BS medium for 24 h, plated (10×104 cells/plate) and
counted after 24, 48, 72 and 96 h in medium supplemented with 0.5%
bovine serum and 5H mixture (6H mixture without TSH or insulin).
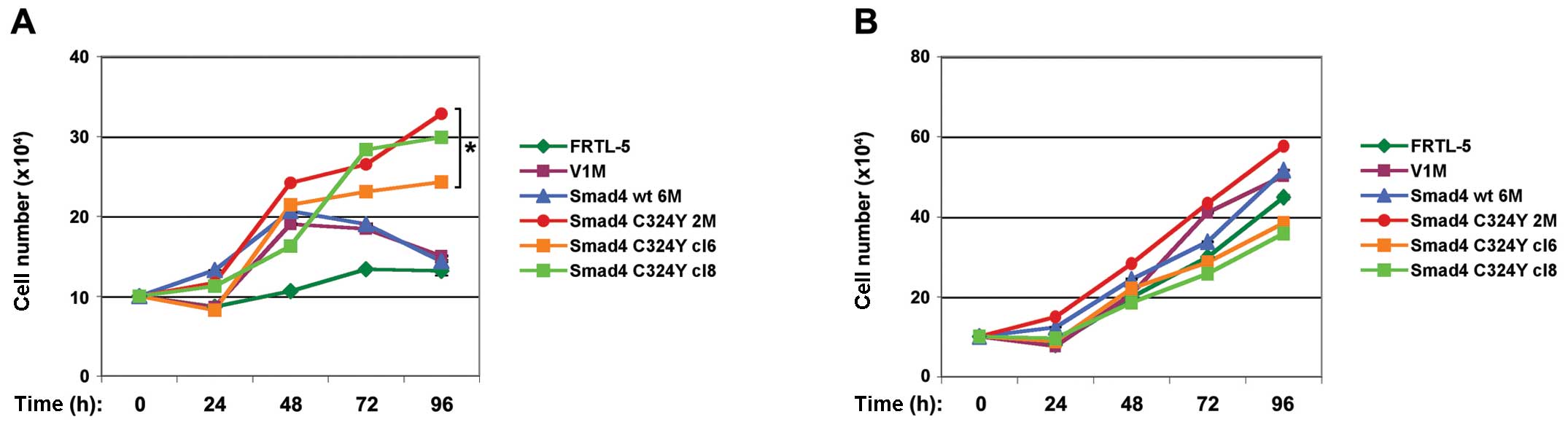
The results shown in Fig. 1A
demonstrated that all Smad4 C324Y clones were able to proliferate
in absence of TSH at all time intervals examined. The growth rate
observed at 96 h was statistically significant (P<0.05) compared
to all control cells (Fig. 1A).
However, no significant differences were observed in proliferation
of C324Y clones grown in absence of insulin, with respect to all
controls, at all times examined (Fig.
1B). These data demonstrated that the expression of Smad4 C324Y
mutation exerted a positive effect on thyroid proliferation in a
TSH-independent manner. Deregulated expression of cyclin D1 is
frequently an early step in neoplastic transformation in various
human cancers including thyroid tumors (30). To evaluate whether the
TSH-independent proliferation observed in Smad4 C324Y cells,
modifies the cyclin D1 expression, we performed western blot
analysis on total lysates obtained from all clones and control
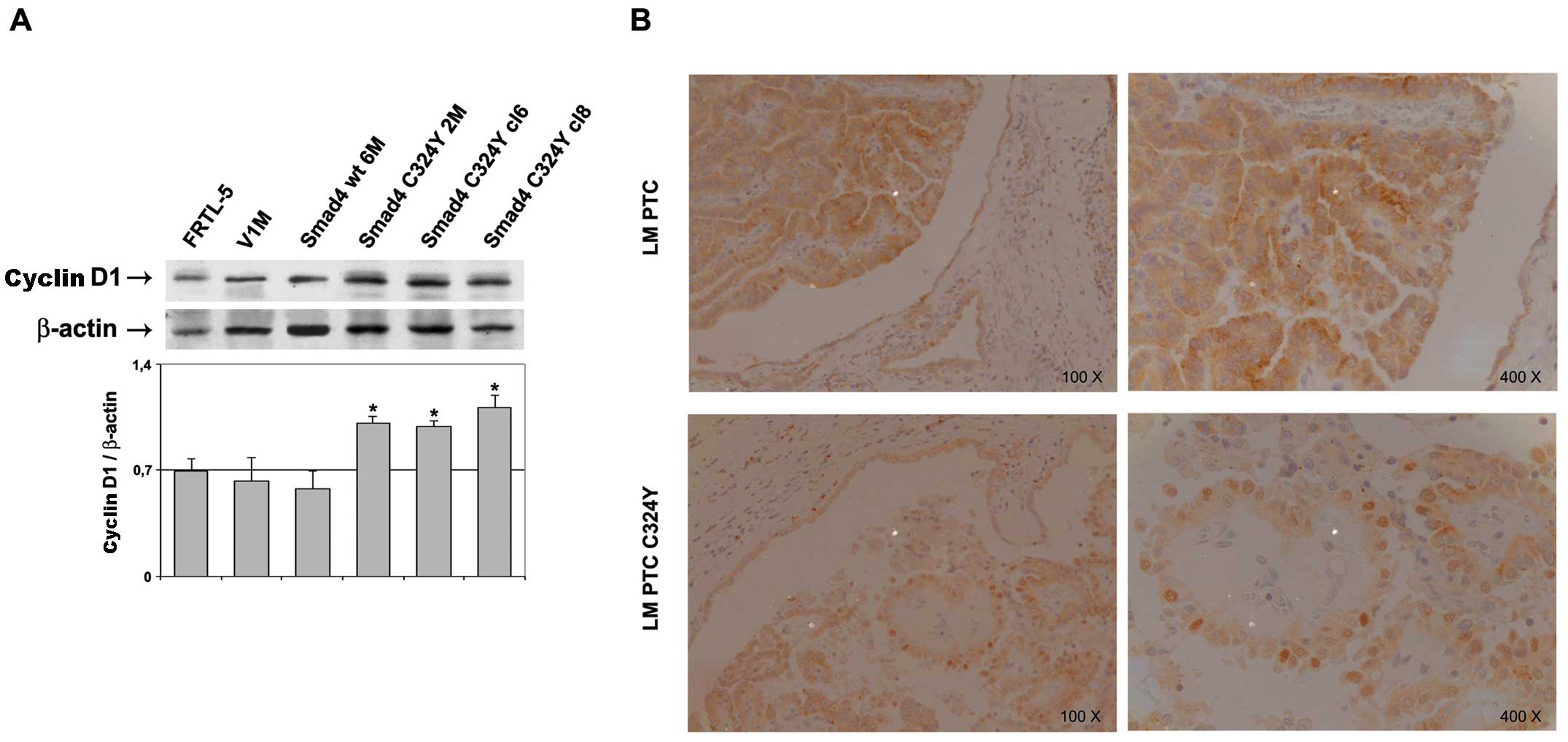
cells, starved in 4H/0.5% BS medium for 24 h. As shown in Fig. 2A, the presence of Smad4 C324Y
mutation determined a statistically significant (P<0.05)
increase of cyclin D1 expression level (about 1.7-fold induction)
with respect to control cells (Fig.
2A). In addition, the lymph nodal metastasis of PTC, from which
the Smad4 C324Y mutation has been isolated, was examined for cyclin
D1 expression by immunohistochemistry and compared to a group of 3
lymph nodal metastases of PTC without mutation. Our results showed
a strong nuclear localization of cyclin D1 in the C324Y lymph nodal
metastases with respect to that of not mutated samples (Fig. 2B). Therefore, these data
demonstrate that the expression of Smad4 C324Y in thyroid cells
might contribute to provide the cells with a growth advantage,
outlining the important role of Smad4 in thyroid
carcinogenesis.
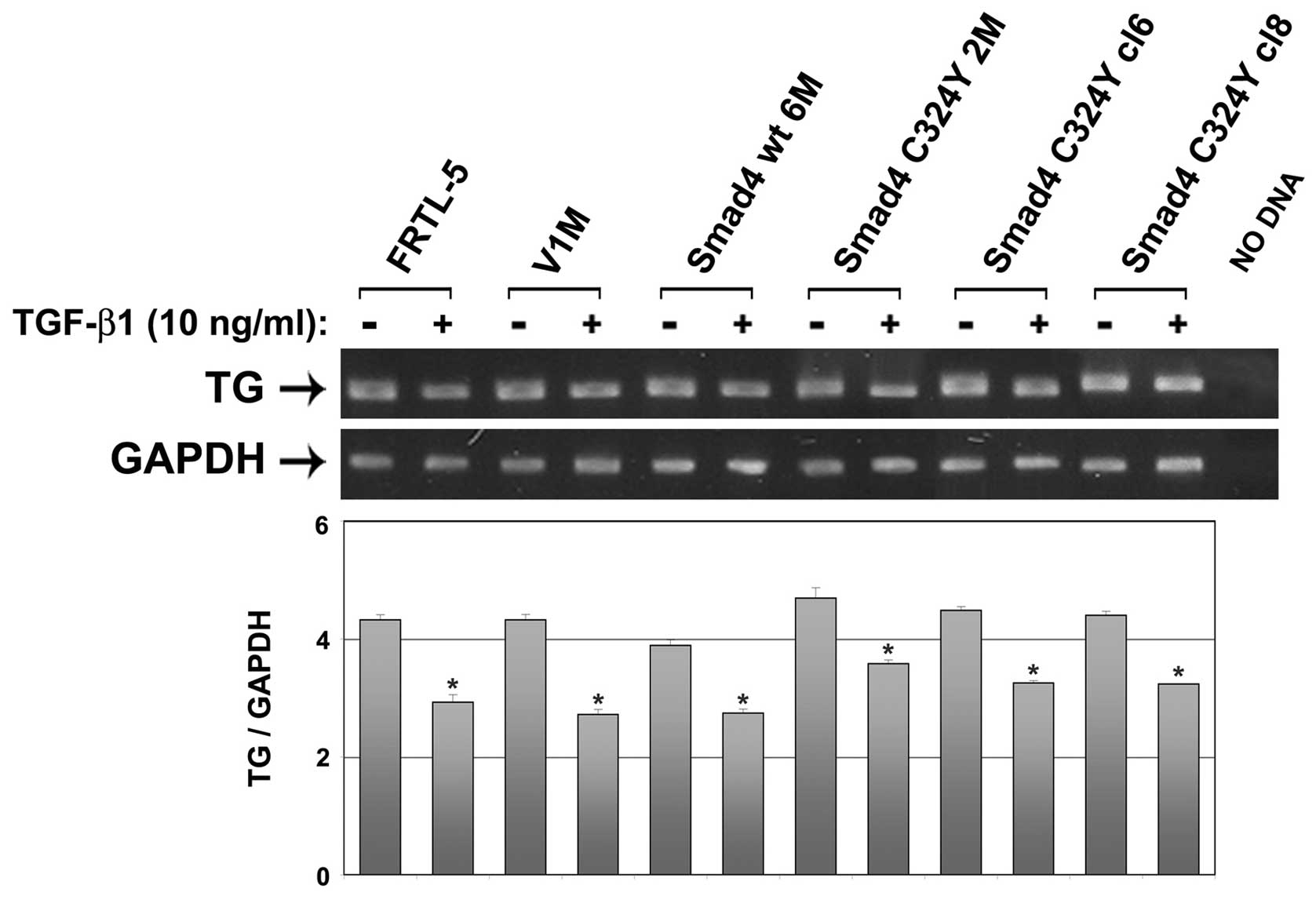
Thyroglobulin represents one of the main
differentiation genes in thyroid cells, whose expression is
modulated by TGF-β in physiological condition (8). By reverse transcription-polymerase
chain reaction (RT-PCR) performed on the same clones (FRTL-5, V1M,
Smad4 wt 6M, Smad4 C324Y clones), treated or not with 10 ng/ml
TGF-β1 for 24 h, we demonstrated that the overexpression of Smad4
C324Y mutation is not associated with significant modification of
TG expression levels in basal condition, neither does it modify the
response to TGF-β treatment (Fig.
3).
Smad3/Smad4 C324Y oligomerization
We already demonstrated that Smad4 C324Y mutation
determines an increase of oligomerization of Smad4 with R-Smad,
Smad2 and a lengthening of nuclear localization, responsible for an
increase of TGF-β signaling (22).
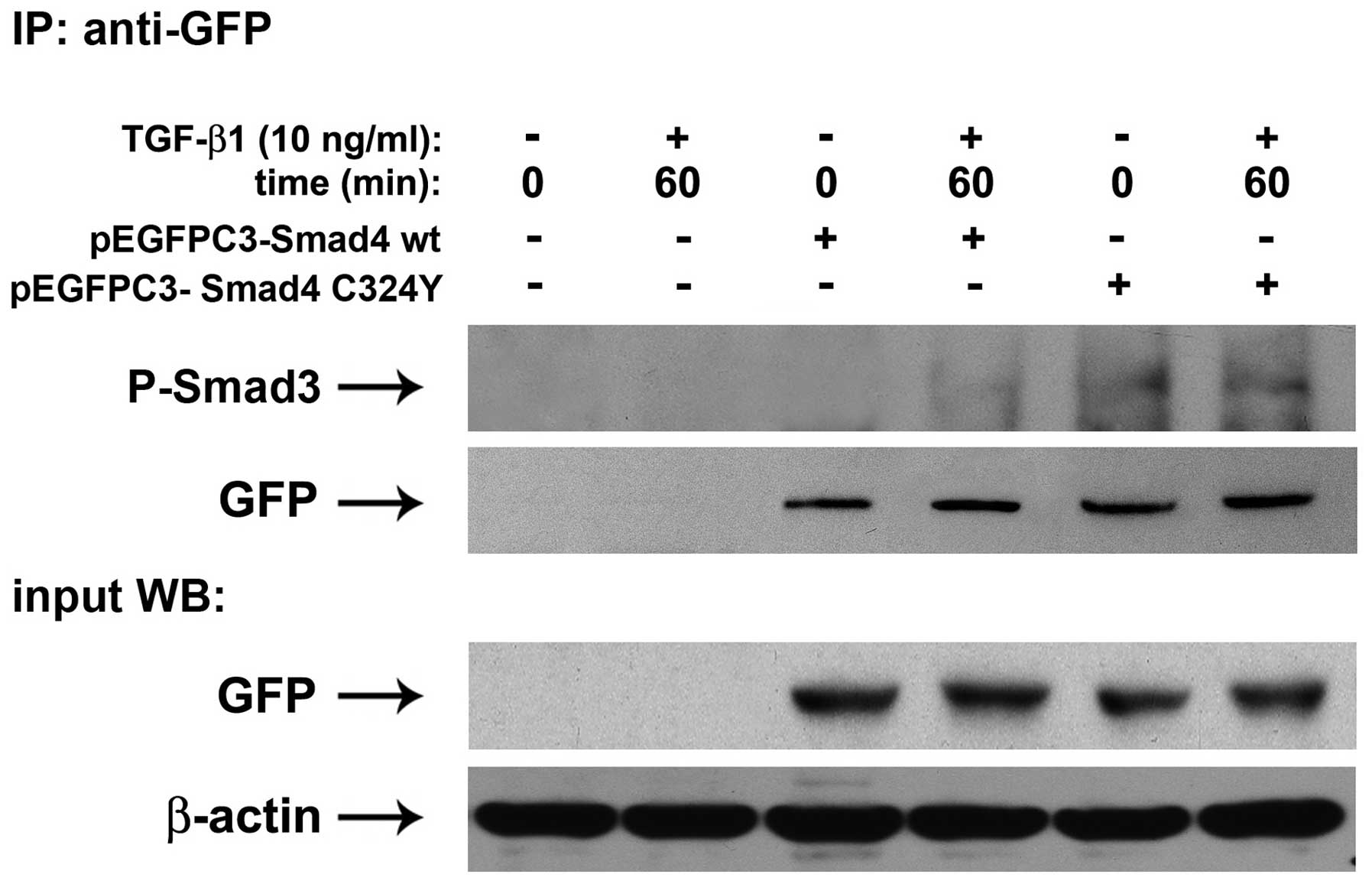
To investigate the effect of Smad4 C324Y in the formation of
complexes with P-Smad3, immunoprecipitation experiments with
anti-GFP antibody were performed on lysates from MDA MB468, cell
line that lacks endogenous SMAD4 because of homozygous deletion of
the SMAD4 gene (26),
untransfected and transiently transfected with pEGFPC3-Smad4 C324Y
or transfected with pEGFPC3-Smad4 wt, untreated or treated with 10
ng/ml TGF-β1 for 60 min. These immunoprecipitates were probed with
an antibody that recognized P-Smad3. The results in Fig. 4 show that in cells overexpressing
Smad4 C324Y, the interaction with P-Smad3 was stronger than that
observed in cells over expressing Smad4 wt, both in basal condition
and after 60 min of treatment (Fig.
4). These results allow us to conclude that the presence of
Smad4 C324Y was responsible for the formation of transcriptional
complexes, able to maintain a longer activation of TGF-β
signaling.
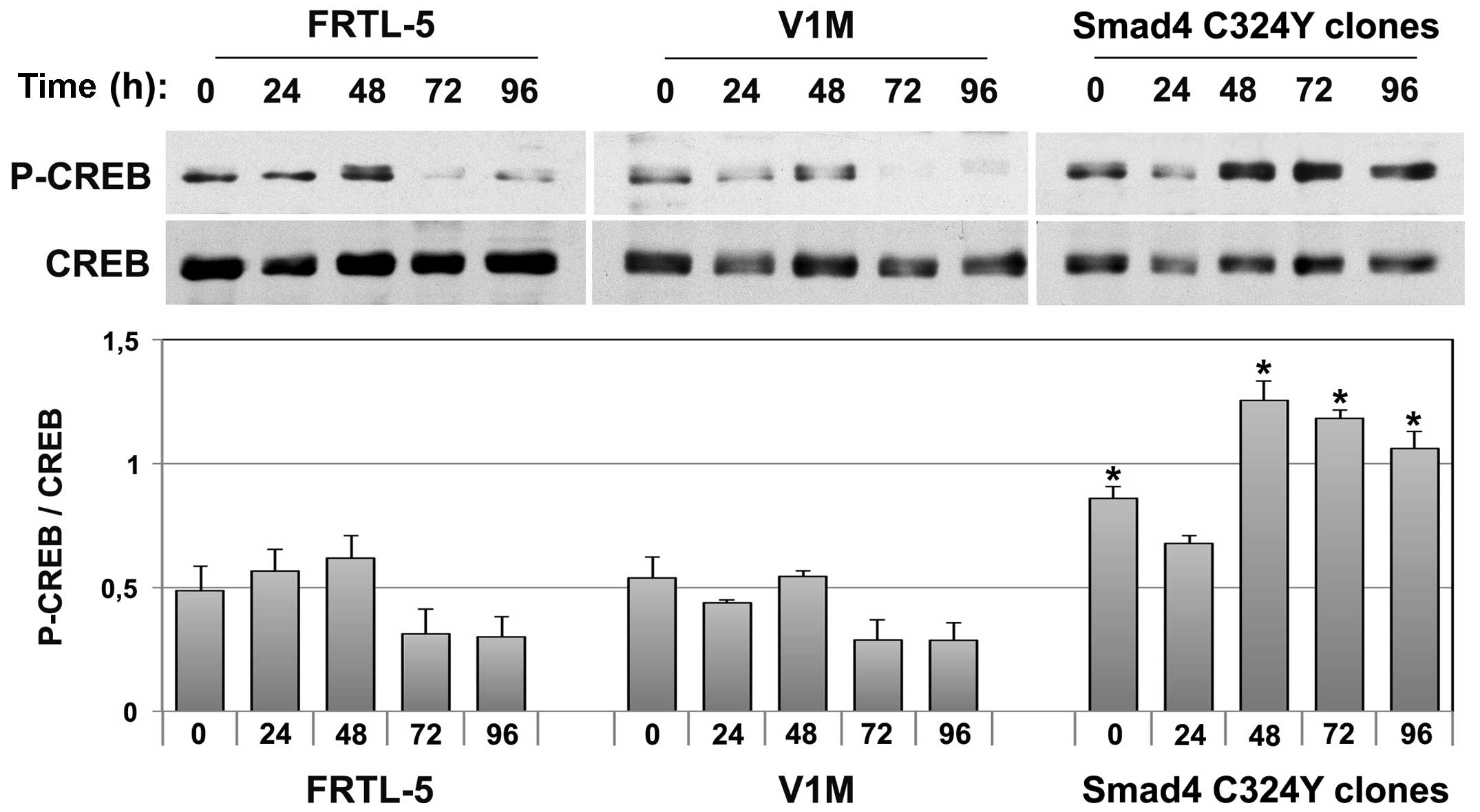
CREB levels in Smad4 C324Y stable
clones
It has been demonstrated that the P-Smad3/Smad4
complex interacts with and activates the PKA, resulting in CREB
phosphorylation and activation of downstream target genes (31). To verify the influence of Smad4
C324Y mutation on CREB phosphorylation, we performed western blot
analysis on total cell lysates obtained from cells starved in
4H/0.5% BS medium for 24 h (T0) and cultured for 24, 48, 72 and 96
h in 5H-TSH/0.5% BS medium. As shown in Fig. 5, P-CREB was significantly
(P<0.05) increased already in basal condition in Smad4 C324Y
clones with respect to FRTL-5 and V1M control cells (1.7- and
1.6-fold induction, respectively); this augmentation was still
evident at 48, 72 and 96 h (P<0.05) (Fig. 5). Given the role of CREB in the
control of proliferation of thyroid cells, an increased level of
its phosphorylation could contribute to the higher proliferative
capacity of the clones overexpressing Smad4 C324Y.
Discussion
TGF-β signaling plays a dichotomous role in tumor
progression and, depending on the cancer type and tumor stage, may
act early as a tumor suppressor and in late-stages as a
pro-metastatic pathway. Smad4 is a key signal transducer of the
TGF-β superfamily that controls a broad range of cellular processes
ranging from proliferation to differentiation and apoptosis
(1,2). Smad4 plays an important role in human
physiology, and its mutations were found with high frequency in a
wide range of human cancers (16,17,32).
We have recently demonstrated that a Smad4 mutation, Smad4 C324Y,
isolated from nodal metastases of papillary thyroid carcinoma
(33), determines a significant
increase of TGF-β signaling, causing the acquisition of transformed
phenotype and invasive behaviour when stably expressed in thyroid
cells (22).
In this study, we focus on the effects of Smad4
C324Y mutation on thyroid cell proliferation and differentiation.
Abnormal thyroid cell proliferation has a very important role in
human diseases. Its deregulation causes goiter, thyroid adenomas,
and carcinomas or primary hypothyroidism resulting from hypoplasia
(34). The main regulators of
thyroid growth and function are TSH and insulin, which recognize
both insulin- and IGF-I receptors. In FRTL-5 cells, that retain
most of the features of differentiated follicular thyroid cells,
these growth factors can produce a dose-dependent increase in DNA
synthesis and cell proliferation (29). Many studies demonstrated that TSH
is the main mitogenic factor in thyroid (35,36),
although others have shown that TSH exerts only a priming effect,
making the cell more competent to progress into G1 phase in
response to insulin/IGF-I alone (37,38).
The view that in FRTL-5 cells, proliferation and DNA synthesis are
synergistically activated by TSH and insulin/IGF-I is almost
unanimously accepted (39,40). In this paper, we demonstrate that
when Smad4 C324Y is expressed, all clones were able to proliferate
in TSH independent manner, while the growth rate of cells growing
without insulin is comparable to that of control cells. These data
let us to state that this Smad4 mutation affects only the
proliferative response to TSH. In FRTL-5, the mitogenic effects of
TSH are mainly or totally mediated by cAMP and require PKA
activity. Binding of TSH to its receptor (TSHR) results in an
increase of cAMP intracellular concentration, which activates PKA
to phosphorylate CREB and other substrates, leading to gene
transcription modification, associated with both proliferation and
differentiation (29). Here we
demonstrated that all Smad4 C324Y clones present a significant
increase of CREB phosphorylation compared to control cell lines
already in basal condition. Given the role of CREB in the control
of proliferation of thyroid cells, an increased level of its
phosphorylation could be responsible of the proliferative behaviour
of the clones overexpressing the mutation of Smad4.
A cyclic AMP-responsive element (CRE), located
upstream of the cyclin D1 mRNA start site, integrates mitogenic
signals that target the CRE-binding factor CREB, which can recruit
the transcriptional coactivator CREB-binding protein (CBP).
Transcriptional activation of the cyclin D1 gene is a key step in
cell proliferation: cyclin D1 is normally expressed during G1 and
regulates the transition from G1 to S phase (30). Deregulated expression of cyclin D1
is frequently an early step in neoplastic transformation in various
human cancers including thyroid tumors (30,41–44).
Interestingly, several studies have also reported correlations
between cyclin D1 overexpression and poor clinical prognostic
characteristics, including advanced tumor stage, tumor recurrence,
and metastatic spread in papillary thyroid (45,46).
Although the copy number of cyclin D1 gene is amplified in a number
of human neoplasms, neither major genetic alterations nor
amplification of this gene has been found in thyroid cancers, but
mainly a strong nuclear cyclin D1 localization (47,48).
This finding allow us to hypothesize that overexpression of cyclin
D1 probably could be a secondary effect, induced by other genetic
alterations in thyroid cancer (30). The expression of cyclin D1 is
increased in all cells overexpressing the Smad4 C324Y mutation
in vitro and it is strongly nuclear in lymph node metastasis
of PTC from which derives the C324Y mutation of Smad4.
An important interaction has been demonstrated
between the TGF-β and PKA signaling pathways, mediating several
physiological responses elicited by TGF-β including CREB
activation. Activated Smad3/Smad4 complexes that bind the
regulatory subunit of PKA, independently of cAMP levels, mediate
this interaction. In this way, the catalytic subunit of PKA is
released, resulting in phosphorylation of CREB and activation of
downstream target genes (31). Our
data show that Smad4 C324Y mutation has a higher capacity to
interact with P-Smad3 than with Smad4 wt, already in basal
condition, thus attesting a strong activity of TGF-β signaling,
responsible also for PKA activation. All together, these data
support the direct involvement of Smad4 C324Y mutation in the
increase of P-CREB levels and clarify the different capacity of
growth observed in these clones in absence of TSH.
Despite hyper-activation of TGF-β signaling causes a
reduction of TG gene expression (8,24,49),
we demonstrated that the overexpression of Smad4 C324Y mutation
does not determine a different behaviour in stable clones either in
basal condition or after TGFβ-treatment. We think that the point
mutation C324Y, localized in the MH2 domain of Smad4 protein, does
not interfere with the control of TG expression. Collectively,
these data demonstrate that Smad4 C324Y mutation, interacting with
PKA pathway, gives the cell the ability to proliferate
independently of TSH. Therefore, we conclude that TGF-β signaling
plays a key role in thyroid carcinogenesis and can be considered as
a new prognostic and therapeutic target for thyroid cancer.
Acknowledgements
The authors would also like to thank
Mr. Daniel Kenton for the careful revisions made. This study is
dedicated to the memory of Dr Davide Lazzereschi.
References
|
1
|
Heldin CH, Miyazono K and ten Dijke P:
TGF-beta signalling from cell membrane to nucleus through SMAD
proteins. Nature. 390:465–471. 1997. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Massagué J: TGF-beta signal transduction.
Annu Rev Biochem. 67:753–791. 1998.
|
|
3
|
Massagué J, Seoane J and Wotton D: Smad
transcription factors. Genes Dev. 9:2783–2810. 2005.PubMed/NCBI
|
|
4
|
Massagué J and Gomis RR: The logic of
TGFbeta signaling. FEBS Lett. 580:2811–2820. 2006.PubMed/NCBI
|
|
5
|
Taton M, Lamy F, Roger PP and Dumont JE:
General inhibition by transforming growth factor beta 1 of
thyrotropin and cAMP responses in human thyroid cells in primary
culture. Mol Cell Endocrinol. 95:13–21. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pekary AE and Hershman JM: Tumor necrosis
factor, ceramide, transforming growth factor-beta1, and aging
reduce Na+/I− symporter messenger ribonucleic
acid levels in FRTL-5 cells. Endocrinology. 139:703–712.
1998.PubMed/NCBI
|
|
7
|
Colletta G, Cirafici AM and Di Carlo A:
Dual effect of transforming growth factor beta on rat thyroid
cells: inhibition of thyrotropin-induced proliferation and
reduction of thyroid-specific differentiation markers. Cancer Res.
49:3457–3462. 1989.
|
|
8
|
Nicolussi A, D’Inzeo S, Santulli M,
Colletta G and Coppa A: TGF-beta control of rat thyroid follicular
cells differentiation. Mol Cell Endocrinol. 207:1–11. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Costamagna E, García B and Santisteban P:
The functional interaction between the paired domain transcription
factor Pax8 and Smad3 is involved in transforming growth
factor-beta repression of the sodium/iodide symporter gene. J Biol
Chem. 279:3439–3446. 2004. View Article : Google Scholar
|
|
10
|
Morris JC 3rd, Ranganathan G, Hay ID,
Nelson RE and Jiang NS: The effects of transforming growth
factor-beta on growth and differentiation of the continuous rat
thyroid follicular cell line, FRTL-5. Endocrinology. 123:1385–1394.
1988. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Coppa A, Mincione G, Mammarella S, Ranieri
A and Colletta G: Epithelial rat thyroid cell clones, escaping from
transforming growth factor beta negative growth control, are still
inhibited by this factor in the ability to trap iodide. Cell Growth
Differ. 6:281–290. 1995.PubMed/NCBI
|
|
12
|
Carneiro C, Alvarez CV, Zalvide J, Vidal A
and Domínguez F: TGF-beta1 actions on FRTL-5 cells provide a model
for the physiological regulation of thyroid growth. Oncogene.
16:1455–1465. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Franzén A, Piek E, Westermark B, ten Dijke
P and Heldin NE: Expression of transforming growth factor-beta1,
activin A, and their receptors in thyroid follicle cells: negative
regulation of thyrocyte growth and function. Endocrinology.
140:4300–4310. 1999.PubMed/NCBI
|
|
14
|
Wyllie FS, Dawson T, Bond JA, Goretzki P,
Game S, Prime S and Wynford-Thomas D: Correlated abnormalities of
transforming growth factor-beta 1 response and p53 expression in
thyroid epithelial cell transformation. Mol Cell Endocrinol.
76:13–21. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Depoortere F, Pirson I, Bartek J, Dumont
JE and Roger PP: Transforming growth factor beta(1) selectively
inhibits the cyclic AMP-dependent proliferation of primary thyroid
epithelial cells by preventing the association of cyclin D3-cdk4
with nuclear p27(kip1). Mol Biol Cell. 11:1061–1076. 2000.
View Article : Google Scholar
|
|
16
|
Bornstein S, White R, Malkoski S, Oka M,
Han G, Cleaver T, Reh D, Andersen P, Gross N, Olson S, Deng C, Lu
SL and Wang XJ: Smad4 loss in mice causes spontaneous head and neck
cancer with increased genomic instability and inflammation. J Clin
Invest. 119:3408–3419. 2009.PubMed/NCBI
|
|
17
|
Ikushima H and Miyazono K: TGFβ
signalling: a complex web in cancer progression. Nat Rev Cancer.
10:415–424. 2010.
|
|
18
|
D’Inzeo S, Nicolussi A, Ricci A, Mancini
P, Porcellini A, Nardi F and Coppa A: Role of reduced expression of
SMAD4 in papillary thyroid carcinoma. J Mol Endocrinol. 45:229–244.
2010.PubMed/NCBI
|
|
19
|
Wikström P, Stattin P, Franck-Lissbrant I,
Damber JE and Bergh A: Transforming growth factor beta1 is
associated with angiogenesis, metastasis, and poor clinical outcome
in prostate cancer. Prostate. 37:19–29. 1998.PubMed/NCBI
|
|
20
|
Padua D and Massagué J: Roles of TGFbeta
in metastasis. Cell Res. 19:89–102. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Riesco-Eizaguirre G, Rodríguez I, De la
Vieja A, Costamagna E, Carrasco N, Nistal M and Santisteban P: The
BRAFV600E oncogene induces transforming growth factor beta
secretion leading to sodium iodide symporter repression and
increased malignancy in thyroid cancer. Cancer Res. 69:8317–8325.
2009. View Article : Google Scholar
|
|
22
|
D’Inzeo S, Nicolussi A, Donini CF, Zani M,
Mancini P, Nardi F and Coppa A: A novel human Smad4 mutation is
involved in papillary thyroid carcinoma progression. Endocr Relat
Cancer. 19:39–55. 2012.PubMed/NCBI
|
|
23
|
Dremier S, Coulonval K, Perpete S,
Vandeput F, Fortemaison N, Van Keymeulen A, Deleu S, Ledent C,
Clément S, Schurmans S, Dumont JE, Lamy F, Roger PP and Maenhaut C:
The role of cyclic AMP and its effect on protein kinase A in the
mitogenic action of thyrotropin on the thyroid cell. Ann NY Acad
Sci. 968:106–1021. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Mincione G, Di Marcantonio MC, Tarantelli
C, D’Inzeo S, Nicolussi A, Nardi F, Donini CF and Coppa A: EGF and
TGF-β1 Effects on Thyroid Function. J Thyroid Res.
2011:4317182011.
|
|
25
|
Ambesi-Impiombato FS, Parks LA and Coon
HG: Culture of hormone-dependent functional epithelial cells from
rat thyroids. Proc Natl Acad Sci USA. 77:3455–3459. 1980.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Schutte M, Hruban RH, Hedrick L, Cho KR,
Nadasdy GM, Weinstein CL, Bova GS, Isaacs WB, Cairns P, Nawroz H,
Sidransky D, Casero RA Jr, Meltzer PS, Hahn SA and Kern SE: DPC4
gene in various tumor types. Cancer Res. 56:2527–2530.
1996.PubMed/NCBI
|
|
27
|
Nicolussi A, D’Inzeo S, Gismondi A and
Coppa A: Reduction of invasive potential in K-ras-transformed
thyroid cells by restoring of TGF-beta pathway. Clin Exp
Metastasis. 23:237–248. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lazzereschi D, Ranieri A, Mincione G,
Taccogna S, Nardi F and Colletta G: Human malignant thyroid tumors
displayed reduced levels of transforming growth factor beta
receptor type II messenger RNA and protein. Cancer Res.
57:2071–2076. 1997.
|
|
29
|
Medina DL and Santisteban P:
Thyrotropin-dependent proliferation of in vitro rat thyroid cell
systems. Eur J Endocrinol. 143:161–178. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kondo T, Ezzat S and Asa SL: Pathogenetic
mechanisms in thyroid follicular-cell neoplasia. Nat Rev Cancer.
6:292–306. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang L, Duan CJ, Binkley C, Li G, Uhler
MD, Logsdon CD and Simeone DM: A transforming growth factor
beta-induced Smad3/Smad4 complex directly activates protein kinase
A. Mol Cell Biol. 24:2169–2180. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Blobe GC, Schiemann WP and Lodish HF: Role
of transforming growth factor beta in human disease. N Engl J Med.
342:1350–1358. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lazzereschi D, Nardi F, Turco A, Ottini L,
D’Amico C, Mariani-Costantini R, Gulino A and Coppa A: A complex
pattern of mutations and abnormal splicing of Smad4 is present in
thyroid tumours. Oncogene. 24:5344–5354. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kimura T, Van Keymeulen A, Golstein J,
Fusco A, Dumont JE and Roger PP: Regulation of thyroid cell
proliferation by TSH and other factors: a critical evaluation of in
vitro models. Endocr Rev. 22:631–656. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ambesi-Impiombato FS, Picone R and
Tramontano D: Influence of hormones and serum on growth and
differentiation of the thyroid cell strain FRTL. Growth of Cells in
Hormonally Defined Media. 9. Sato GH, Pardee AB and Sirbaku DA:
Cold Spring Harbor; New York: pp. 483–492. 1982
|
|
36
|
Ambesi-Impiombato FS and Villone G: The
FRTL-5 thyroid cell strain as a model for studies on thyroid cell
growth. Acta Endocrinol Suppl (Copenh). 281:242–245.
1987.PubMed/NCBI
|
|
37
|
Veneziani BM, Di Marino C, Salvatore P,
Villone G, Perrotti N, Frunzio R and Tramontano D: Transfected
insulin-like growth factor II modulates the mitogenic response of
rat thyrocytes in culture. Mol Cell Endocrinol. 86:11–20. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Takahashi S, Conti M and Van Wyk JJ:
Thyrotropin potentiation of insulin-like growth factor-I dependent
deoxribonucleic acid synthesis in FRTL-5 cells: mediation by an
autocrine amplification factor(s). Endocrinology. 126:736–745.
1990. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Tramontano D, Cushing GW, Moses AC and
Ingbar SH: Insulin-like growth factor-I stimulates the growth of
rat thyroid cells in culture and synergizes the stimulation of DNA
synthesis induced by TSH and Graves’-IgG. Endocrinology.
119:940–942. 1986.PubMed/NCBI
|
|
40
|
Koide T, Ono Y, Ito Y, Akahori M, Nedachi
T, Hakuno F, Takenaka A, Takahashi SI and Noguchi T: Insulin-like
growth factor-I potentiates protein synthesis induced by
thyrotropin in FRTL-5 cells: comparison of induction of protein
synthesis and DNA synthesis. Endocr J. 45:151–163. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Diehl JA: Cycling to cancer with cyclin
D1. Cancer Biol Ther. 1:226–231. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
42
|
Fu M, Wang C, Li Z, Sakamaki T and Pestell
RG: Minireview: Cyclin D1: normal and abnormal functions.
Endocrinology. 145:5439–5447. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Pickett CA, Agoff SN, Widman TJ and
Bronner MP: Altered expression of cyclins and cell cycle inhibitors
in papillary thyroid cancer: prognostic implications. Thyroid.
15:461–473. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Basolo F, Caligo MA, Pinchera A, Fedeli F,
Baldanzi A, Miccoli P, Iacconi P, Fontanini G and Pacini F: Cyclin
D1 overexpression in thyroid carcinomas: relation with
clinico-pathological parameters, retinoblastoma gene product, and
Ki67 labeling index. Thyroid. 10:741–746. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Khoo ML, Beasley NJ, Ezzat S, Freeman JL
and Asa SL: Overexpression of cyclin D1 and underexpression of p27
predict lymph node metastases in papillary thyroid carcinoma. J
Clin Endocrinol Metab. 87:1814–1818. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wang S, Lloyd RV, Hutzler MJ, Safran MS,
Patwardhan NA and Khan A: The role of cell cycle regulatory
protein, cyclin D1, in the progression of thyroid cancer. Mod
Pathol. 13:882–887. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Lazzereschi D, Sambuco L, Carnovale Scalzo
C, Ranieri A, Mincione G, Nardi F and Colletta G: Cyclin D1 and
Cyclin E expression in malignant thyroid cells and in human thyroid
carcinomas. Int J Cancer. 76:806–811. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Khoo ML, Ezzat S, Freeman JL and Asa SL:
Cyclin D1 protein expression predicts metastatic behavior in
thyroid papillary microcarcinomas but is not associated with gene
amplification. J Clin Endocrinol Metab. 87:1810–1813. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Kang HC, Ohmori M, Harii N, Endo T and
Onaya T: Pax-8 is essential for regulation of the thyroglobulin
gene by transforming growth factor-beta1. Endocrinology.
142:267–275. 2001.PubMed/NCBI
|