Introduction
Recently, bone metastasis has been increasing in
patients with various cancers. This results in bone pain,
pathological fractures, neurological deficits, and/or
hypercalcemia, which are related to a decrease in the quality of
life (QOL) in patients with bone metastasis. Thus, it is important
to understand how these patients should be managed.
Cancer cells are distributed hematogenously to the
bone (1). The process of such
hematogenous spread in soft tissue metastasis includes shedding of
cancer cells from a primary tumor into the circulation,
colonization in a target organ, extravasation into the surrounding
tissue, initiation and maintenance of growth, and vascularization
of the metastatic tumor (2).
However, in bone metastasis, an additional step in which cancer
cells invade and proliferate in the bone matrix is necessary
(3–5). Current evidence suggests that
osteoclasts play a crucial role in this step during
tumor-associated osteolysis, and several osteoclastogenic factors,
i.e., parathyroid hormone-like hormone (PTHLH) (6,7),
secreted matrix metalloproteases (MMPs) (8), interleukin-6 (IL-6) (9), receptor activator of nuclear factor
κB ligand (RANKL) (10),
macrophage inflammatory protein-1α (MIP-1α) (11), interleukin-8 (IL-8) (12,13),
and granulocyte-macrophage colony-stimulating factor (GM-CSF)
(14,15) have been implicated in the
activation of osteoclastic bone resorption during tumor-associated
osteolysis in solid tumors. These factors, released by metastatic
tumor cells in the bone marrow, stimulate differentiation and
activation of osteoclasts either directly or through osteoblasts,
thereby leading to osteolysis and the release of tumor cell growth
factors from the bone matrix. Based on this evidence for the
molecular mechanism of bone metastasis, bisphosphonates (16) and denosumab (17), which inhibit osteoclastic bone
resorption, have been developed for the treatment of bone
metastasis. Although these drugs are widely used, their clinical
benefit is limited. Therefore, the development of a novel drug for
the treatment of bone metastasis is necessary.
It is important to identify factors that are
involved in bone metastasis because such factors could be
attractive therapeutic targets. The present study describes
experimental evidence of the role of CXC chemokine ligand 14
(CXCL14), a member of the CXC chemokine family, in the progression
of bone metastasis from lung cancer.
Materials and methods
Cell culture
HARA cells were obtained from the Health Science
Research Resources Bank (Osaka, Japan). HARA and its bone-seeking
subclones were cultured in RPMI-1640 medium (Invitrogen, Carlsbad,
CA, USA) supplemented with 10% fetal bovine serum (FBS), 50 U/ml of
penicillin, and 50 μg/ml of streptomycin in a humidified
atmosphere under 5% CO2 at 37°C. The murine
pre-osteoclastic/macrophage cell line RAW264.7 and pre-osteoblastic
cell line MC3T3-E1 were cultured in α-modified minimum essential
medium (α-MEM) supplemented with 10% FBS, 50 U/ml of penicillin,
and 50 μg/ml of streptomycin in a humidified atmosphere
under 5% CO2 at 37°C.
In vivo selection of a bone-seeking HARA
subclone
HARA cells (2×105 cells) inoculated into
the left ventricle of the heart in 5-week-old male nude mice (Clea
Japan Inc, Tokyo, Japan) spread mostly to the bone, muscle, and
adrenal glands 4–6 weeks after inoculation, suggesting that batches
of HARA cells contain mixed subpopulations with different
metastatic potentials. To establish a bone-seeking subclone, HARA
cells in bone metastases were isolated by the explant outgrowth
technique, grown in culture (HARA-B), and reinoculated into the
left ventricle of the heart. HARA-B cells that metastasized to bone
were then isolated and grown in culture (HARA-B2). This procedure
was repeated 4 times until no metastases were detected
macroscopically in the adrenal glands (HARA-B4). All animal studies
were carried out according to the protocol approved by the National
Hospital Organization Kyushu Cancer Center Animal Ethics Committee
(permit no. 23-02), in compliance with the Committee
guidelines.
Total RNA preparation and DNA
microarray
Total RNA was extracted from HARA and HARA-B4 cells
using a Qiagen RNeasy kit (Qiagen, Venlo, The Netherlands)
according to the manufacturer’s instructions. RNA samples were
analyzed using a GeneChip Human Genome U133 Plus 2.0 Array
(Affymetrix, Santa Clara, CA, USA). The GeneSpring GX 7.3.1 program
(Agilent Technologies, Santa Clara, CA, USA) was used to filter
gene expression levels. The results were visualized using a Venn
diagram, and the genes known to exhibit expression changes of
≥5-fold with larger raw data values of >100 were listed. The
microarray data described in this report have been deposited in the
NCBI GEO database with the accession no. GSE29367.
Reverse transcription and quantitative
real-time polymerase chain reaction (PCR)
Reverse transcription was performed using the
ThermoScript RT-PCR System (Invitrogen) according to the
manufacturer’s instructions. Quantitative real-time PCR (qRT-PCR)
was performed using the ABI PRISM 7000 Sequence Detection System
(Applied Biosystems, Foster City, CA, USA) and SYBR Premix Ex Taq
(Takara Bio, Otsu, Shiga, Japan). The PCR conditions were as
follows: 95°C (10 sec) followed by 40 cycles at 95°C (5 sec) and
60°C (31 sec). The human gene-specific primers were 5′-CGCTACAGCGAC
GTGAAGAA-3′ and 5′-TTCCAGGCGTTGTACCACTTG-3′ for CXCL14,
5′-CCGTCCGATTTGGGTCTGATG-3′ and 5′-TCTTTTTCTTCCCAGGTGTCTTG-3′ for
PTHLH, 5′-CCGTCGCCCATCATCAAGTT-3′ and 5′-CTGTCTGGG GCAGTCCAAAG-3′
for MMP2, 5′-AAGAAGTGGAATA AGTGGGCTCT-3′ and
5′-GGCCGAATAAGGGTCTGGG-3′ for adrenomedullin (ADM),
5′-AGCTCTGTCTGGACCCC AAG-3′ and 5′-TGGCAACCCTACAACAGACC-3′ for
IL-8, and 5′-CATGTACGTTGCTATCCAGGC-3′ and 5′-CTCC
TTAATGTCACGCACGAT-3′ for β-actin.
RNA interference
Stealth Select RNAi duplexes designed to target
CXCL14 (siCXCL14) and negative controls designed not to
target any known human gene (siNC) were purchased from Invitrogen.
For the silencing assay, we transfected 5×105 cells
(HARA or HARA-B4) with 15 μl of stock Stealth Select RNAi
duplex (20 μM) using Lipofectamine RNAiMAX solution
(Invitrogen) in a 100-mm diameter culture dish. We harvested total
RNA from transfected cells 48 h after transfection to perform gene
expression profiling.
Anchorage-independent proliferation in an
agar matrix
Soft agar experiments were performed using a
CytoSelect 96-well cell transformation assay kit following the
manufacturer’s instructions (Cell Biolabs, San Diego, CA, USA).
Briefly, the bottom layer contained 50 μl of 1X CytoSelect
agar matrix solution and 10% FBS in Dulbecco’s modified Eagle’s
medium (DMEM), and the top layer contained 75 μl of 1X
CytoSelect agar matrix solution, 10% FBS in DMEM, and
5×103 cells. All assays were performed in 96-well
plates. The plates were incubated for 1 week at 37°C with 5%
CO2 to allow cell proliferation in soft agar.
Photographs were taken using an Olympus phase contrast microscope.
Volumes of colonies were calculated according to absorbance values
at 570 nm using the MTT assay after the addition of matrix
solubilization solution.
Resorbing activity of osteoclasts
Osteoclasts were formed in the rat whole bone marrow
cell culture system as described previously (18). Briefly, bone marrow cells were
obtained from the tibia and femur of 4-week-old male Sprague Dawley
rats and cultured in 24-well plates (1×106 cells/well)
in α-MEM containing 15% FBS in the presence of 10−8 M
1α, 25(OH)2D3 and 10% (v/v) htROSCM (19). After 4 days of culture, the cells
were detached from the culture plates with 0.05% trypsin and 0.02%
EDTA. Mature osteoclasts were then cocultured with HARA or HARA-B4
(2×103 cells/well) on Osteologic discs (BD, Franklin
Lake, NJ, USA). After 3 days of culture, attached cells were
removed from the discs using 6% sodium hypochlorite, and von Kossa
staining was performed according to the manufacturer’s
instructions. Images of pits in each well were obtained using
inverted microscopy (BZ-8100, Keyence, Japan). The total area of
resorption pits and the total number of resorption pits per well
were measured using an image analysis system (BZ-II analyzer;
Keyence, Japan).
Transwell cell migration assay
Transwell cell migration was evaluated using a
24-well chemotaxis chamber with 3- and 8-μm pore size
membranes (BD) for MC3T3-E1 and RAW264.7, respectively. HARA and
HARA-B4 (2.5×104 cells/0.8 ml) cells transfected with
various RNAi duplexes were cultured in the lower chamber for 16 h,
and the medium was then changed to serum-free α-MEM (0.8 ml).
Concurrently, MC3T3-E1 and RAW264.7 were cultured in serum-free
α-MEM medium for 12 h and harvested. Cell suspensions of MC3T3-E1
(1.25×104 cells/0.5 ml) and RAW264.7
(1.25×105 cells/0.5 ml) were then added to the upper
chamber and incubated for 4.5 and 6 h, respectively, at 37°C.
Non-migrated cells were wiped off with a cotton swab, the filter
was stained with Diff-Quik stain solution (Siemens, Munch,
Germany), and the remaining cells were counted under the
microscope.
Immunohistochemistry
Lung cancer cells (HARA/siNC, HARA-B4/siNC,
HARA-B4/siCXCL14) were injected into the left cardiac ventricle of
nude mice, which were sacrificed 2 weeks after injection. The legs
of the mice were then fixed in buffered formalin, decalcified in
10% EDTA (pH 7.0), and embedded in paraffin. Human clinical
specimens of pulmonary adenocarcinoma bone metastases (n=7), which
were obtained by computed tomography-guided needle biopsy, were
also subjected to immunohistochemistry for the detection of CXCL14.
Sections (5-μm thick) were treated with target retrieval
solution (Dako, Glostrup, Denmark) for 20 min at 95°C and
subsequently with anti-CXCL14 antibody (1:100, MAB866, R&D
Systems, Minneapolis, MN, USA). CXCL14 was visualized using CSA-II
(Dako) and Autostainer Plus (Dako) according to the manufacturer’s
instructions. Histological sections were also stained with
hematoxylin and eosin. All the human specimens for this study were
collected after obtaining written informed consent from the
patients. This study and the protocols were approved by the
Institutional Ethics Committee of National Hospital Organization
Shikoku Cancer Center, where the patients were treated.
Statistical analysis
The results are presented as mean ± SD. Group
comparisons were performed using either: i) the non-parametric
Mann-Whitney test, or ii) the parametric Student’s t-test and
one-way analysis of variance (ANOVA) followed by Bonferroni’s
multiple comparison test. Differences were considered significant
at p<0.05.
Results
Establishment of a bone-seeking subclone
of HARA and its capacity to metastasize to the bone and adrenal
glands
After 4 passages in vivo and in culture, we
successfully established a bone-seeking subclone (HARA-B4). We then
investigated the metastatic capacity of HARA-B4 compared to the
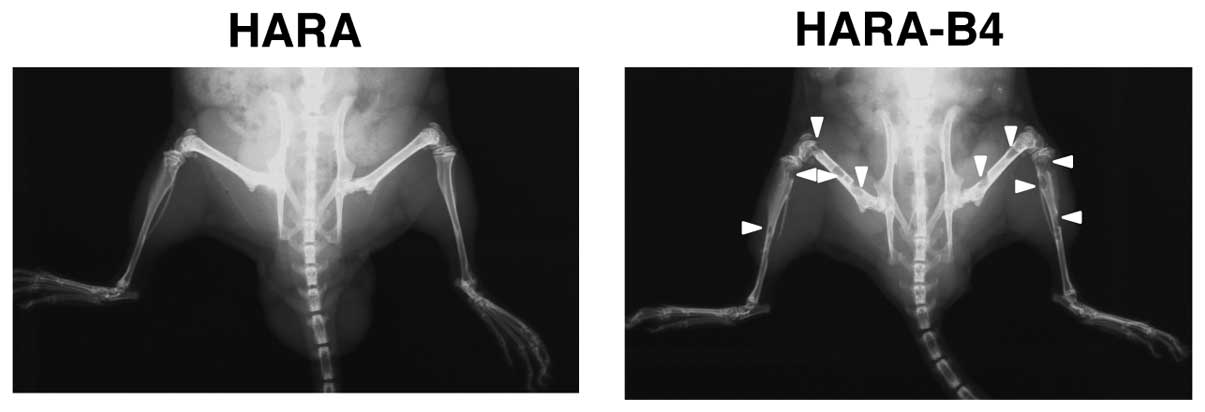
parental line (HARA) in the heart injection model. Radiographs of
hind legs were obtained 4 weeks after intracardiac inoculation to
detect bone metastatic colonies. Few colonies were detected in
HARA-inoculated mice, whereas nearly 15 metastatic colonies were
detected in HARA-B4-inoculated mice (Fig. 1 and Table I). The mice were sacrificed at 7-8
weeks post-inoculation when hind leg paralysis was noted or when
body weight decreased below 20 g. There were no significant
differences in the number of macroscopic bone metastases between
HARA-and HARA-B4-inoculated mice at autopsy, although it tended to
be higher in the HARA-B4-inoculated mice (Table I). All HARA-inoculated mice
developed metastases to the adrenal glands in addition to bone,
while adrenal metastasis was not found in HARA-B4-inoculated mice
(Table I). Metastases to the
muscle were also seen in both HARA- and HARA-B4-inoculated mice
with bone metastasis (data not shown).
 | Table I.Establishment of a bone-seeking
subclone (HARA-B4) from the parental cell line (HARA) that was
established from human squamous cell lung cancer. |
Table I.
Establishment of a bone-seeking
subclone (HARA-B4) from the parental cell line (HARA) that was
established from human squamous cell lung cancer.
| HARA | HARA-B4 |
|---|
| No. of bone
metastases | | |
| 4 weeks | NDa | 15.5±3.7c |
| Autopsy | 26.5±9.4 | 53.5±20.6 |
| Incidence of
adrenal metastases | | |
| Autopsy | 4/4 | 4/0b |
Identification of bone metastasis-related
genes
In order to elucidate the genes involved in bone
metastasis, we compared comprehensive gene expression profiles
between HARA and HARA-B4. Gene expression arrays were performed on
HARA and HARA-B4 cells using Affymetrix GeneChips. The GeneSpring
GX 7.3.1 program (Agilent Technologies) was used to filter gene
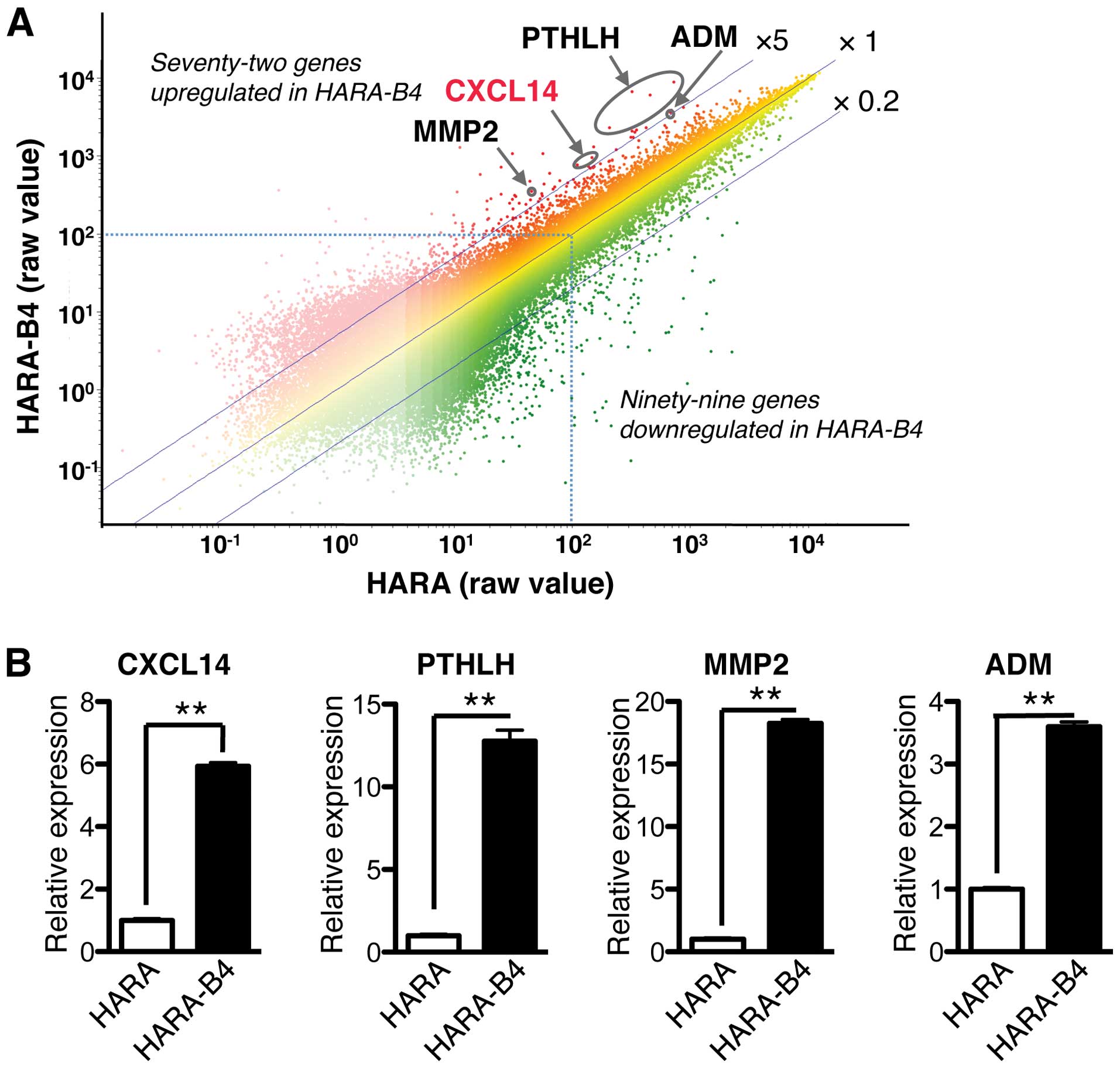
expression levels. The genes with expression changes of ≥5-fold and
raw data values of >100 are selected for further analysis. A
total of 72 genes were upregulated and 99 genes were downregulated
in HARA-B4 compared to HARA (Fig.
2A). To validate the GeneChip data, the expression levels of
four differentially expressed genes (PTHLH, MMP2, ADM and
CXCL14) that may participate in metastasis were assessed by
qRT-PCR, and the results were consistent with the findings obtained
from the microarray (Fig. 2B). Of
these genes, we focused on CXCL14 in the following analyses
because it has not yet been reported in the context of bone
metastasis.
Effects of CXCL14 silencing on bone
metastases in HARA-B4-inoculated nude mice
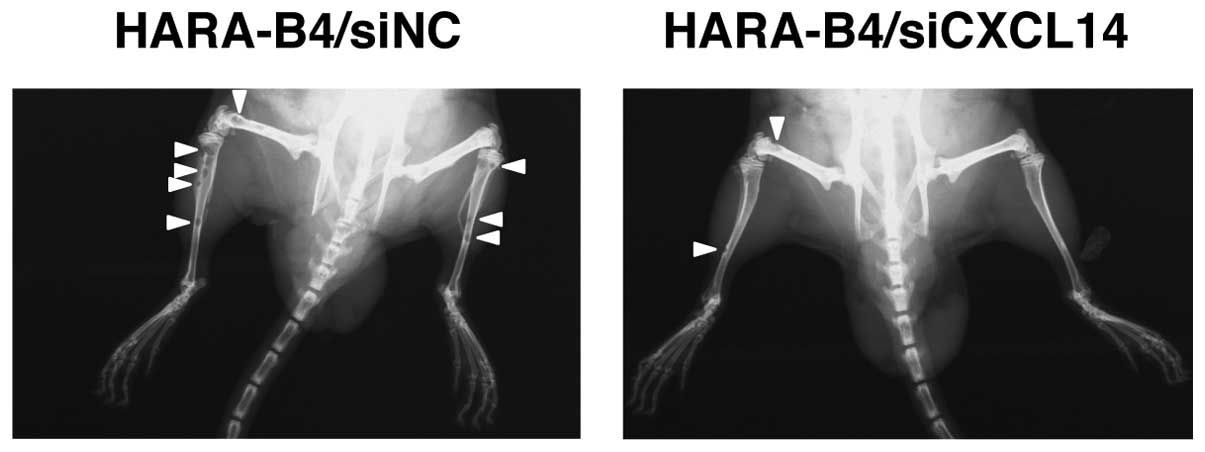
To characterize the role of CXCL14 in bone
metastasis, HARA/siNC (negative control siRNA-transfected HARA),
HARA-B4/siNC, and HARA-B4/siCXCL14 (siRNA that targets
CXCL14-transfected HARA-B4) cells were inoculated into the hearts
of nude mice. Accordingly, 6.3±3.7 and 2.6±1.1 metastatic colonies
were detected in HARA-B4/siNC- and HARA-B4/siCXCL14-inoculated mice
4 weeks after inoculation, respectively (Fig. 3 and Table II). This indicates that the
silencing of CXCL14 significantly reduces bone metastases in
HARA-B4-inoculated nude mice. The number of bone metastases and the
incidence of adrenal metastasis were also examined at autopsy,
which was performed at 7–8 weeks post-inoculation. The number of
bone metastases did not significantly differ between HARA-B4/siNC
and HARA-B4/siCXCL14-inoculated nude mice at autopsy, although it
tended to be lower in the HARA-B4/siCXCL14-inoculated mice
(Table II). Interestingly, none of
the HARA-B4/siNC-inoculated mice exhibited adrenal gland
metastases, whereas all the HARA-B4/siCXCL14-inoculated mice
exhibited metastases to this organ (Table II).
| Table II.Effects of CXCL14 silencing on bone
and adrenal metastases in HARA-B4-inoculated mice. |
Table II.
Effects of CXCL14 silencing on bone
and adrenal metastases in HARA-B4-inoculated mice.
| HARA/siNC |
HARA-B4/siCXCL14 |
|---|
| Number of bone
metastases | | |
| 4 weeks | 6.3±3.7 | 2.6±1.1a |
| Autopsy | 35.1±15.6 | 24.1±13.2 |
| Incidence of
adrenal metastases | | |
| Autopsy | 7/0 | 8/8b |
Effects of CXCL14-silencing on
anchorage-independent proliferation of HARA-B4
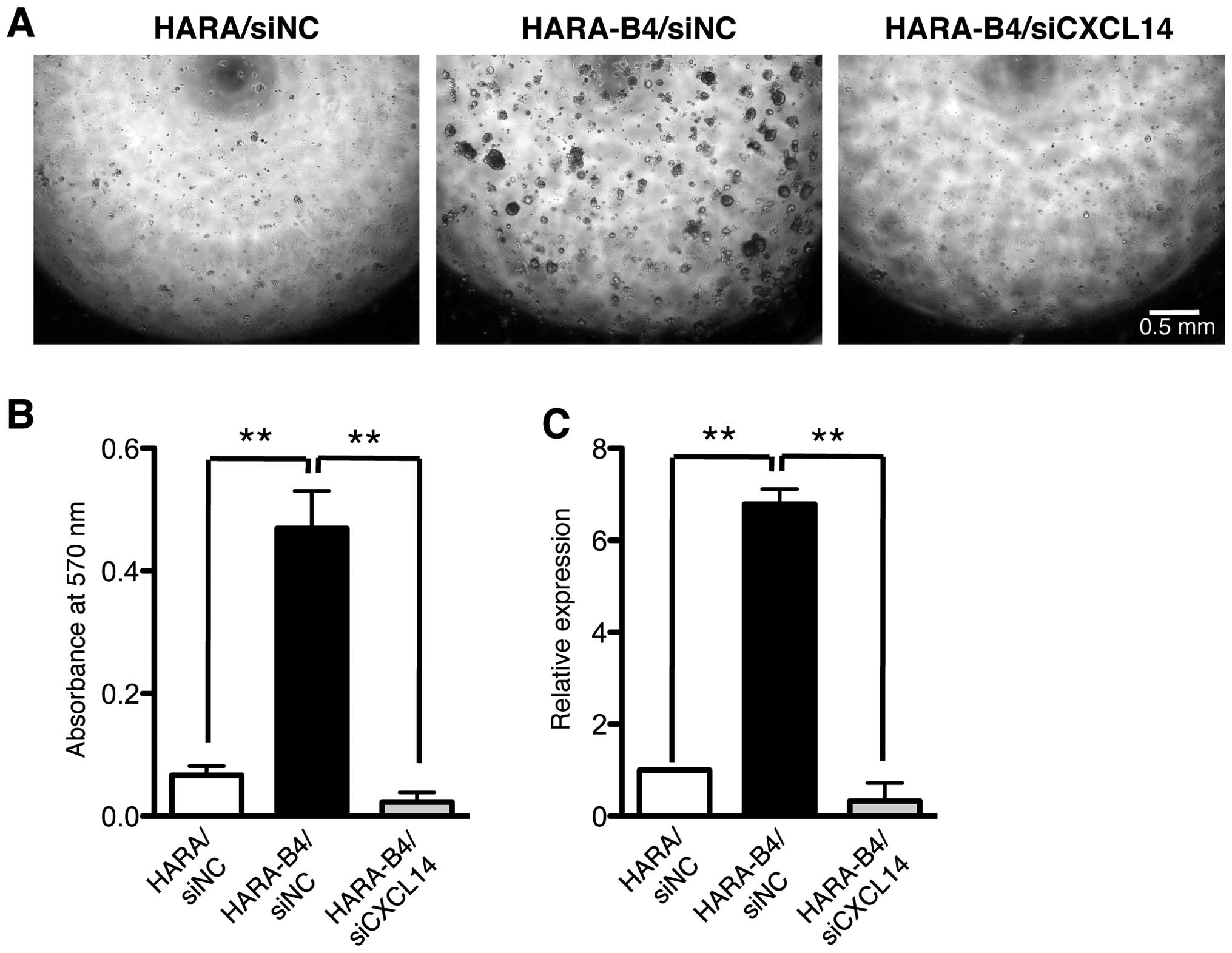
Anchorage-independent proliferation is one of the
hallmarks of malignant tumor cells. Therefore, we assessed the
anchorage-independent proliferation of HARA/siNC and HARA-B4/siNC
using soft agar colony formation assays. One week after plating,
both HARA/siNC and HARA-B4/siNC cells formed colonies in soft agar.
However, the diameters of the HARA-B4/siNC colonies were 2-3 times
larger than those of the HARA/siNC colonies (Fig. 4A). Next, we examined the effect of
siCXCL14-silencing in HARA-B4 on anchorage-independent
proliferation activity. The colony formation activity of HARA-B4
was suppressed by siRNA-induced CXCL14 silencing (Fig. 4A). The total volume of HARA-B4/siNC
colonies was more than 5 times larger than that of HARA/siNC
colonies as quantified by the MTT assay, and the total volume was
decreased more than 10-fold by CXCL14 silencing (Fig. 4B). The efficiency of CXCL14
silencing in HARA-B4 was checked by qRT-PCR 48 h after transfection
(Fig. 4C).
Interaction between cancer cells and bone
marrow cells in the microenvironment of bone metastasis
To investigate the interaction between lung cancer
cells and bone marrow cells in the bone microenvironment, the
effects of HARA and HARA-B4 on the resorbing activity of mature
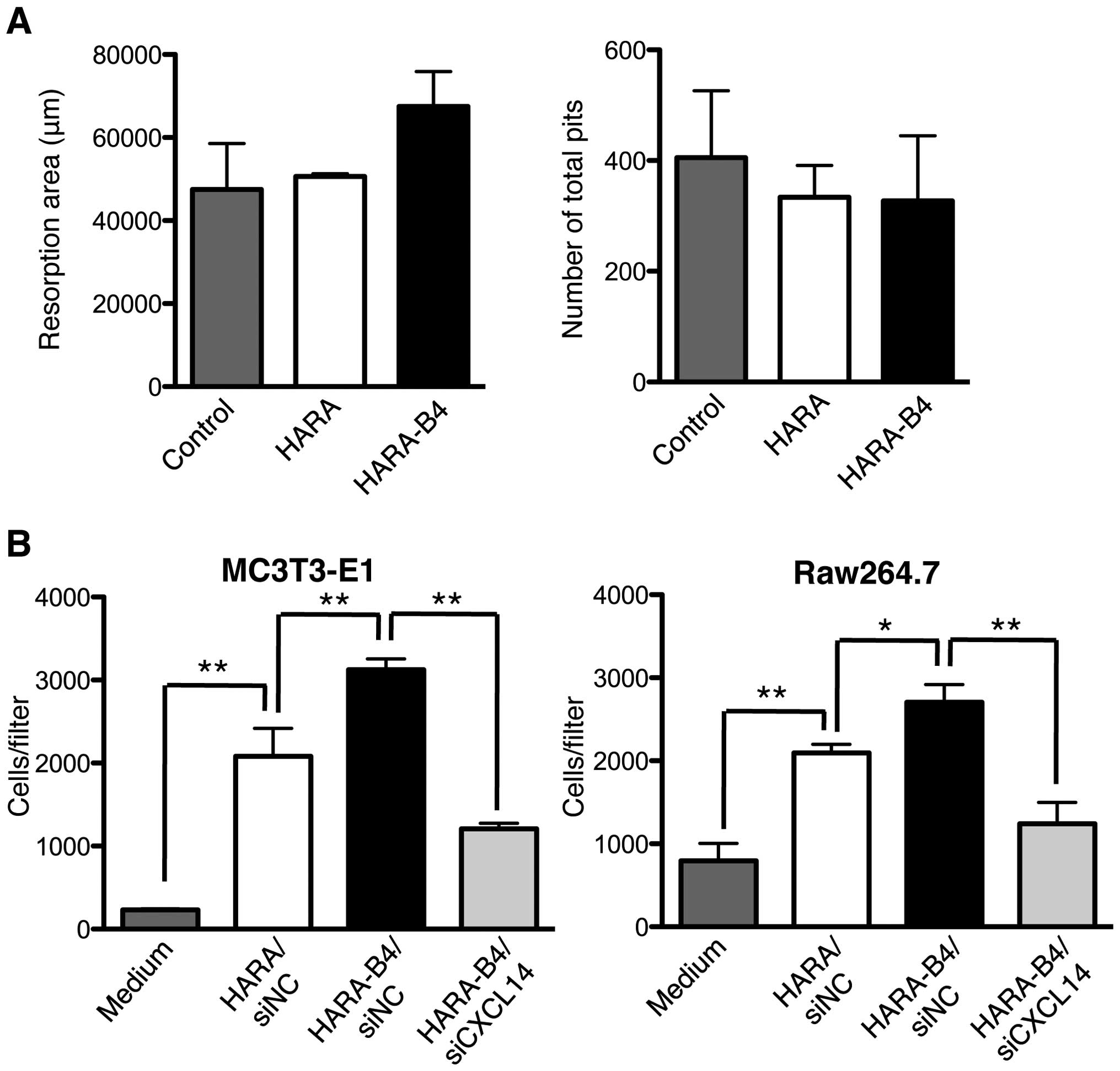
osteoclasts was examined by pit formation. There were no
differences in resorption areas or the number of total pits between
HARA and HARA-B4 (Fig. 5A). Next,
the chemotactic interactions between cancer cells and bone marrow
cells were evaluated using the transwell migration chamber. When we
put the murine pre-osteoblast cell line MC3T3-E1 and the
pre-osteoclast/macrophage cell line RAW264.7 in the upper chamber
with HARA/siNC and HARA-B4/siNC cells in the lower chamber as
stimulants, both MC3T3-E1 and RAW264.7 migrated faster toward the
cultured HARA-B4/siNC cells than toward the HARA/siNC cells
(Fig. 5B). We then silenced
CXCL14 expression in HARA-B4 cells to evaluate the effect of
CXCL14 on the migration of MC3T3-E1 and RAW264.7. Consequently, a
decrease in the migration of both MC3T3-E1 and RAW264.7 was
observed (Fig. 5B).
Histological examination of bone
metastasis
Hematoxylin and eosin staining showed that the bone
marrow cavity included infiltrated tumor cells in the bone
metastatic specimens of the mice inoculated with HARA/siNC and
HARA-B4/siNC (Fig. 6A and C).
Expression of CXCL14 was confirmed immunohistologically in the
tumor cells that infiltrated bone marrow; however, its expression
patterns were patchy for HARA/siNC (Fig. 6B) and abundant and diffuse for
HARA-B4/siNC (Fig. 6D). Expression
of CXCL14 was also noted in stromal cells in the bone metastatic
microenvironment (Fig. 6B and D).
In contrast, in soft tissue metastases of mice inoculated with
HARA-B4/siCXCL14, expression of CXCL14 was not observed in the
infiltrated tumor cells but rather in the stromal cells (Fig. 6E and F). In the clinical specimens
of lung cancer-induced bone metastasis, expression of CXCL14 was
observed in the tumor cells that infiltrated the bone marrow as
well as in the bone stromal cells in all specimens examined
(Fig. 6G and H).
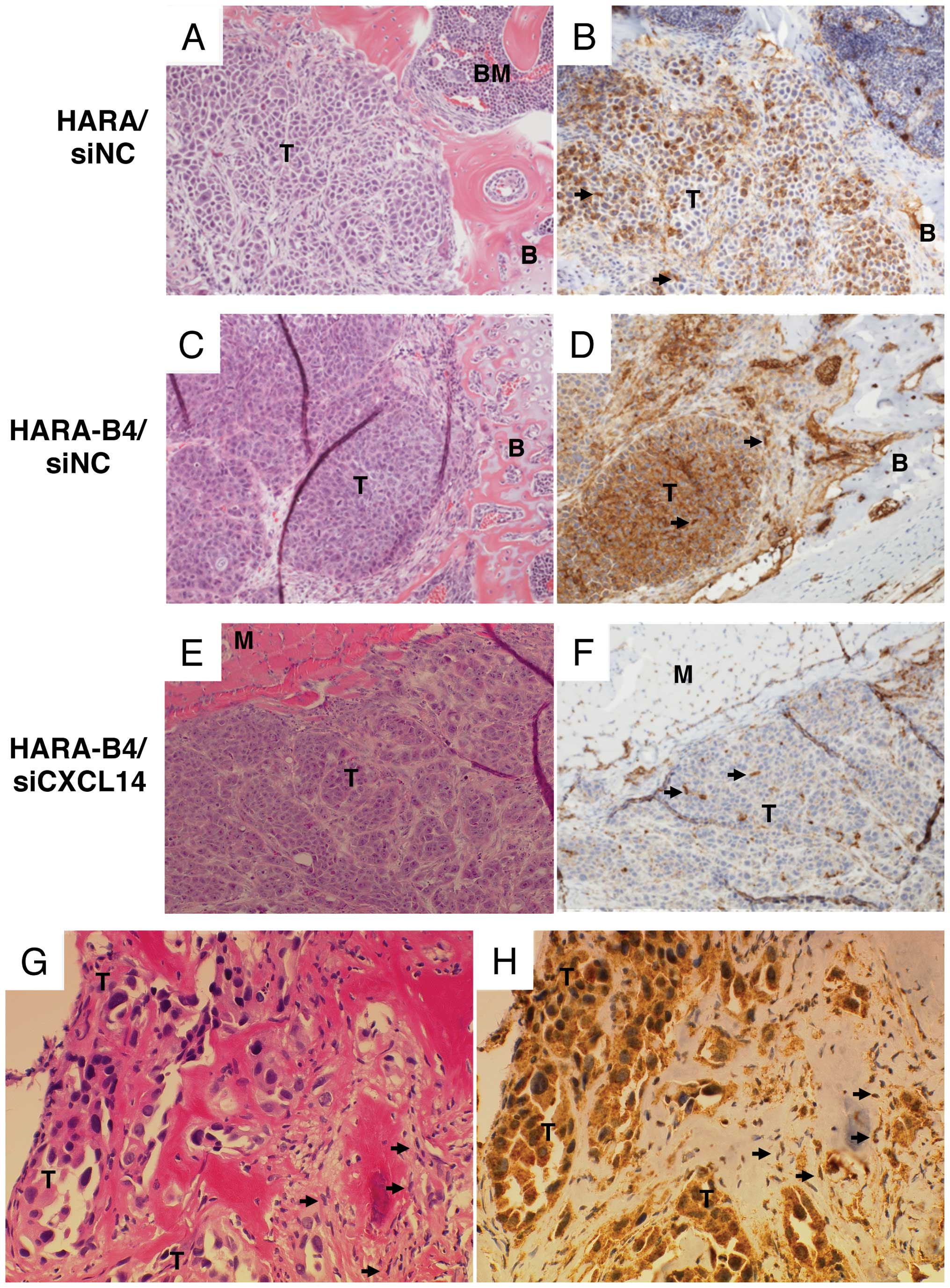 | Figure 6.Histological views and expression of
CXCL14 in lung cancer-induced bone metastases and/or soft tissue
metastases. (A, C, E and G) Hematoxylin and eosin staining,
original magnification (OM) ×200. (B, D, F and H)
Immunohistochemical staining of CXCL14, OM ×200. Specimens of bone
metastases were isolated from nude mice injected with HARA/siNC and
HARA-B4/siNC at 2 weeks after inoculation. Specimens of soft tissue
metastases were isolated from nude mice injected with
HARA-B4/siCXCL14 because bone metastasis was not observed in these
mice. Bone marrow was replaced with infiltrated tumor cells in mice
injected with HARA/siNC (A) and HARA-B4/siNC (C). Expression of
CXCL14 was observed in approximately half of the HARA/siNC cells
(B), whereas it was observed in most HARA-B4/siNC cells (D).
Expression of CXCL14 was also noted in tumor stromal cells and bone
marrow stromal cells in these specimens (B and D). In the soft
tissue metastases of mice injected with HARA-B4/siCXCL14, no CXCL14
expression was noted in tumor cells, and CXCL14 expression was
observed only in tumor stromal cells (arrow) (E and F). (G) Lung
adenocarcinoma metastasized to bone in clinical specimens
(hematoxylin and eosin, OM, ×200). (H) Expression of CXCL14 was
observed in both tumor cells and stromal cells (arrow) in bone
metastases (OM, ×200). T, tumor cells; B, bone; BM, bone marrow; M,
muscle; arrows, stromal cells. |
Discussion
The incidence of bone metastasis has been increasing
in all cancers, given the prolonged survival rate enabled by recent
advances in cancer treatment. Therefore, it is important to
understand how these patients should be managed in order to
maintain their QOL. The molecular mechanism responsible for bone
metastasis formation has been elucidated over the past 20 years,
and osteoclasts have been shown to play pivotal roles during this
process. Therefore, bisphosphonates (16) and denosumab (17), which target osteoclasts, have been
developed for the treatment of bone metastasis. Although these
drugs are widely used in clinical practice, their clinical benefit
is limited. Therefore, the development of a new drug that further
improves the QOL of patients with bone metastasis is necessary.
To identify novel factors related to bone
metastasis, we established a bone-seeking subclone (HARA-B4) from
the parental cell line (HARA) established from human squamous cell
lung cancer. We then compared the gene expression patterns between
HARA and HARA-B4. From these analyses, PTHLH, MMP2, ADM and CXCL14
were identified as factors that are related to bone metastasis
formation based on their increased gene expression in HARA-B4 and
previous reports examining cancer proliferation and metastasis.
Among these factors, PTHLH and MMP2 have been previously reported
to contribute to bone metastasis through the differentiation and
activation of osteoclasts and/or the promotion of angiogenesis
(8,20). ADM was also reported to be involved
in the promotion of angiogenesis (21). Although CXCL14 was identified as a
member of the CXC chemokine family in 1999 (22,23),
a specific receptor for CXCL14 has not yet been uncovered. CXCL14
has been described in the context of cancer cell proliferation and
metastasis; however, its relationship to bone metastasis has not
been elucidated (24–27). Recent studies indicate that the
interaction between a chemokine (CXCL12/SDF-1α) and its receptor
(CXCR4) plays an important role in the establishment of bone
metastasis (28,29). Furthermore, PTHLH induces chemokine
(CCL2) production from osteoblasts and other bone stromal cells,
which in turn stimulates VEGF expression in tumor cells and
enhances angiogenesis (30).
Accordingly, chemokines and/or chemokine receptors have been shown
to play important roles in bone metastasis formation. Thus, we
chose to focus on CXCL14 and analyze its function in relation to
bone metastasis. Indeed, mice inoculated with HARA-B4 in which
CXCL14 expression was silenced exhibited a reduction in bone
metastases and a simultaneous increase in visceral (adrenal)
metastases, suggesting a possible correlation between the
expression of CXCL14 and cancer cell tropism to the bone. HARA-B4
also exhibited an increase in anchorage-independent growth ability,
in the soft agar colony formation assay, which is a hallmark of
malignant cells that form tumors in vivo and is thought to
be associated with highly metastatic cancer cells (31,32).
This anchorage-independent growth ability of HARA-B4 was decreased
by CXCL14 silencing, suggesting a possible correlation
between the expression of CXCL14 and the tumorigenicity of HARA-B4.
Consequently, the HARA-B4 established in the present study had
acquired both tropism to the bone and tumorigenicity in a bone
microenvironment through increased expression of CXCL14, which
resulted in the acquisition and enhancement of bone metastatic
potential.
Current evidence indicates that osteoclasts play a
crucial role in the development of bone metastasis (3). Bone resorption induced by activated
osteoclasts creates spaces for the proliferation of metastatic
cancer cells, and concurrently, growth factors released from the
bone matrix create an attractive microenvironment for the growth of
metastatic cancer cells (4). In
the present study, the effects of HARA and HARA-B4 on the resorbing
activity of mature osteoclasts were examined by pit formation;
however, there were no differences between HARA and HARA-B4. The
interaction between bone marrow cells and metastatic cancer cells
is important for the formation of a bone metastatic niche (5); we therefore evaluated the chemotaxis
of osteoblast-like cells (MC3T3-E1) and osteoclast-like cells
(RAW264.7) toward HARA or HARA-B4 in vitro. The chemotaxis
of both MC3T3-E1 and RAW264.7 toward HARA-B4 was significantly
increased compared with that toward HARA, and furthermore,
CXCL14 silencing in HARA-B4 suppressed the chemotaxis of
these bone marrow cells toward HARA-B4. It is interesting to note
that CXCL14 regulates the chemotaxis of bone marrow cells
(osteoclasts, osteoblasts, etc.) toward metastatic cancer cells.
Indeed, osteoclastogenic factors such as PTHLH and MMP2 are highly
expressed in HARA-B4; therefore, it is readily conceivable that
when bone marrow cells are accumulated around meta-static cancer
cells, differentiation and activation of osteoclasts is facilitated
by the interactions between these cells, and increased bone
resorption aids the formation of metastatic cancer cell colonies.
To determine whether the stimulation of chemotaxis by HARA-B4 was
due to a direct effect of CXCL14, we examined the effect of
recombinant CXCL14 (100 nM) on the chemotaxis of RAW267.4 and
MC3T3-E1 toward HARA or HARA-B4. However, we did not observe any
differences in chemotaxis (data not shown). Regarding the role of
CXCL14 as a chemoattractant, a recent report indicated that
CXCL14-producing mouse fibroblasts, but not recombinant CXCL14,
enhanced the in vitro proliferation and migration of
prostate cancer cells by undefined factors (26). A similar mechanism might be
involved in the chemotaxis of bone marrow cells toward
CXCL14-overproducing cancer cells observed in the present study.
Further experiments will be required to answer this question.
Finally, we performed immunohistochemical staining
to confirm the expression of CXCL14 in the specimens of bone
metastases isolated from nude mice. Expression of CXCL14 was
observed in both HARA and HARA-B4 cells that infiltrated bone
marrow; however, its expression was patchy in HARA cells and
abundant and diffuse in HARA-B4 cells. These results may be
explained by differences in CXCL14 expression in the whole
population: the HARA population is heterogeneous in terms of CXCL14
expression, whereas the HARA-B4 population consists of a
homogeneous population of CXCL14-overexpressing cells enriched
through in vivo selection. Similarly, in 7 clinical
specimens of lung cancer-induced bone metastasis, strong expression
of CXCL14 was observed in tumor cells infiltrated in bone marrow.
It is interesting to note that CXCL14 expression was observed in
the bone marrow stromal cells in addition to the cancer cells that
infiltrated bone marrow in the mouse xenograft model as well as in
the clinical specimens. These results suggest that bone marrow
stromal cells, in cooperation with metastatic cancer cells,
contribute to bone metastasis through a mechanism mediated by
CXCL14.
In conclusion, CXCL14 was able to promote bone
metastasis through enhancement of cancer cell tropism to the bone
and/or recruitment of bone marrow cells (osteoclasts, osteoblasts,
and others) around the metastatic cancer cells. This molecular
mechanism is different from those reported previously regarding
osteoclastogenic factors, which stimulate differentiation and
activation of osteoclasts (3–5).
Although CXCL14 plays an important role during the establishment of
bone metastasis, it remains unclear whether the effects of CXCL14
are mediated directly or indirectly. Thus, further studies will be
required to develop molecular therapy targeting CXCL14.
Acknowledgements
This study was supported in part by
the Grant-in-Aid for Cancer Research (project no. 00054) from the
Ministry of Health, Labour, and Welfare of Japan (to H. Iguchi) and
Grant-in-Aid for Scientific Research (project no. 25462356) from
Japan Society for the Promotion of Science (to S. Takiguchi).
References
|
1.
|
Mundy GR: Metastasis to bone: causes,
consequences and therapeutic opportunities. Nat Rev Cancer.
2:584–593. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
2.
|
Chambers AF, Groom AC and MacDonald IC:
Dissemination and growth of cancer cells in metastatic sites. Nat
Rev Cancer. 2:563–572. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Roodman GD: Mechanisms of bone metastasis.
N Engl J Med. 350:1655–1664. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Weilbaecher KN, Guise TA and McCauley LK:
Cancer to bone: a fatal attraction. Nat Rev Cancer. 11:411–425.
2011. View
Article : Google Scholar
|
|
5.
|
Ell B and Kang Y: SnapShot: bone
metastasis. Cell. 151:690. e12012. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Iguchi H, Tanaka S, Ozawa Y, et al: An
experimental model of bone metastasis by human lung cancer cells:
the role of parathyroid hormone-related protein in bone metastasis.
Cancer Res. 56:4040–4043. 1996.PubMed/NCBI
|
|
7.
|
Guise TA, Yin JJ, Taylor SD, et al:
Evidence for a causal role of parathyroid hormone-related protein
in the pathogenesis of human breast cancer-mediated osteolysis. J
Clin Invest. 98:1544–1549. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
Lynch CC: Matrix metalloproteinases as
master regulators of the vicious cycle of bone metastasis. Bone.
48:44–53. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Ara T and Declerck YA: Interleukin-6 in
bone metastasis and cancer progression. Eur J Cancer. 46:1223–1231.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Croucher PI, Shipman CM, Lippitt J, et al:
Osteoprotegerin inhibits the development of osteolytic bone disease
in multiple myeloma. Blood. 98:3534–3540. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Abe M, Hiura K, Wilde J, et al: Role for
macrophage inflammatory protein (MIP)-1alpha and MIP-1beta in the
development of osteolytic lesions in multiple myeloma. Blood.
100:2195–2202. 2002.PubMed/NCBI
|
|
12.
|
Bendre MS, Gaddy-Kurten D, Mon-Foote T,
Akel NS, Skinner RA, Nicholas RW and Suva LJ: Expression of
interleukin 8 and not parathyroid hormone-related protein by human
breast cancer cells correlates with bone metastasis in vivo. Cancer
Res. 62:5571–5579. 2002.PubMed/NCBI
|
|
13.
|
Bendre MS, Margulies AG, Walser B, et al:
Tumor-derived interleukin-8 stimulates osteolysis independent of
the receptor activator of nuclear factor-kappaB ligand pathway.
Cancer Res. 65:11001–11009. 2005. View Article : Google Scholar
|
|
14.
|
Mancino AT, Klimberg VS, Yamamoto M,
Manolagas SC and Abe E: Breast cancer increases osteoclastogenesis
by secreting M-CSF and upregulating RANKL in stromal cells. J Surg
Res. 100:18–24. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Park BK, Zhang H, Zeng Q, et al: NF-kappaB
in breast cancer cells promotes osteolytic bone metastasis by
inducing osteoclastogenesis via GM-CSF. Nat Med. 13:62–69. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Coleman R, Cook R, Hirsh V, Major P and
Lipton A: Zoledronic acid use in cancer patients: more than just
supportive care? Cancer. 117:11–23. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Lacey DL, Boyle WJ, Simonet WS, et al:
Bench to bedside: elucidation of the OPG-RANK-RANKL pathway and the
development of denosumab. Nat Rev Drug Discov. 11:401–419. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Kukita A, Kukita T, Shin JH and Kohashi O:
Induction of mononuclear precursor cells with osteoclastic
phenotypes in a rat bone marrow culture system depleted of stromal
cells. Biochem Biophys Res Commun. 196:1383–1389. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Kukita A, Kukita T, Hata K, Kurisu K and
Kohashi O: Heat-treated osteoblastic cell (ROS17/2.8)-conditioned
medium induces the formation of osteoclast-like cells. Bone Miner.
23:113–127. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Inoue D and Matsumoto T: Parathyroid
hormone-related peptide and bone: pathological and physiological
aspects. Biomed Pharmacother. 54(Suppl 1): S32–S41. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Nikitenko LL, Fox SB, Kehoe S, Rees MC and
Bicknell R: Adrenomedullin and tumour angiogenesis. Br J Cancer.
94:1–7. 2006. View Article : Google Scholar
|
|
22.
|
Hromas R, Broxmeyer HE, Kim C, Nakshatri
H, Christopherson K II, Azam M and Hou YH: Cloning of BRAK, a novel
divergent CXC chemokine preferentially expressed in normal versus
malignant cells. Biochem Biophys Res Commun. 255:703–706. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Cao X, Zhang W, Wan T, et al: Molecular
cloning and characterization of a novel CXC chemokine macrophage
inflammatory protein-2 gamma chemoattractant for human neutrophils
and dendritic cells. J Immunol. 165:2588–2595. 2000. View Article : Google Scholar
|
|
24.
|
Allinen M, Beroukhim R, Cai L, et al:
Molecular characterization of the tumor microenvironment in breast
cancer. Cancer Cell. 6:17–32. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25.
|
Shellenberger TD, Wang M, Gujrati M, et
al: BRAK/CXCL14 is a potent inhibitor of angiogenesis and a
chemotactic factor for immature dendritic cells. Cancer Res.
64:8262–8270. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26.
|
Augsten M, Hagglof C, Olsson E, et al:
CXCL14 is an autocrine growth factor for fibroblasts and acts as a
multi-modal stimulator of prostate tumor growth. Proc Natl Acad Sci
USA. 106:3414–3419. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27.
|
Pelicano H, Lu W, Zhou Y, Zhang W, Chen Z,
Hu Y and Huang P: Mitochondrial dysfunction and reactive oxygen
species imbalance promote breast cancer cell motility through a
CXCL14-mediated mechanism. Cancer Res. 69:2375–2383. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
28.
|
Muller A, Homey B, Soto H, et al:
Involvement of chemokine receptors in breast cancer metastasis.
Nature. 410:50–56. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
29.
|
Taichman RS, Cooper C, Keller ET, Pienta
KJ, Taichman NS and McCauley LK: Use of the stromal cell-derived
factor-1/CXCR4 pathway in prostate cancer metastasis to bone.
Cancer Res. 62:1832–1837. 2002.PubMed/NCBI
|
|
30.
|
Li X, Loberg R, Liao J, Ying C, Snyder LA,
Pienta KJ and McCauley LK: A destructive cascade mediated by CCL2
facilitates prostate cancer growth in bone. Cancer Res.
69:1685–1692. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31.
|
Chiarugi P and Giannoni E: Anoikis: a
necessary death program for anchorage-dependent cells. Biochem
Pharmacol. 76:1352–1364. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32.
|
Taddei ML, Giannoni E, Fiaschi T and
Chiarugi P: Anoikis: an emerging hallmark in health and diseases. J
Pathol. 226:380–393. 2012. View Article : Google Scholar : PubMed/NCBI
|