Introduction
Human cervical carcinoma is one of the leading
malignancies in women worldwide (1). Approximately 80% of human cervical
carcinoma is squamous cell carcinoma (SCC), while only 20% of
remaining cervical carcinoma is adenocarcinoma (2). Epidemiological and laboratory data
have shown that human cervical carcinoma is mainly associated with
human papillomavirus (HPV) infections (3,4). The
HPV infection and replication in cervical squamous epithelial cells
resulted in increasing the risk of developing cervical carcinoma
(5). However, accumulated data
indicate that only a small number of women infected with HPV
finally develop a highly malignant cervical carcinoma, suggesting
that other factors are necessary for development of human cervical
carcinoma (6).
Interleukin-6 (IL-6) is a central pro-inflammatory
cytokine, which has been described as an important factor for the
pathogenesis of human cervical carcinoma (7). In addition, IL-6 is highly expressed
in invasive cervical carcinoma and is associated to the
pathogenesis of HPV-related cervical carcinoma (8). These data support the hypothesis that
IL-6 signaling may participate in the development of human cervical
carcinoma. Classic IL-6 signaling requires binding of IL-6 to
interleukin-6 receptor (IL-6R) and subsequent activation of signal
transducer and activator of transcription 3 (Stat3), a downstream
protein of IL-6R (9). Stat3 is
typically located in the cytoplasm when Stat3 is inactive, then
Stat3 forms homodimers and enters the nucleus binding to
DNA-response elements after phosphorylation of Stat3 (10). Stat3 has been demonstrated to play
an important role in human cervical carcinoma and its activation by
IL-6 stimulation is connected with enhanced cancer cell growth,
survival and immune evasion (11–13).
Epithelial-mesenchymal transition (EMT) was
originally described as an essential process for specific
developmental stages and embryonic epithelial cells migration in
early embryogenesis (14). Studies
have also shown that EMT is an important process in many types of
human epithelial carcinoma including cervical carcinoma (15–19).
During EMT, cancer cells lose their epithelial properties and gain
mesenchymal characteristics (20).
As an epithelial cell marker, E-Cadherin is a cell adhesion
molecule and is associated with suppression of cancer cell motility
and invasion (21), whereas, the
expression of mesenchymal cell marker Vimentin in cancer cells is
believed to enhance migration and invasiveness, and is regulated by
several transcription factors including Slug in cancer progression
(22,23). Several factors such as EGF are
known to regulate EMT in human cervical carcinoma (19). However, whether IL-6 plays an
important role in EMT induction and the mechanisms underlying the
IL-6-induced EMT remain to be determined.
In this study, we investigated the role of IL-6 and
its downstream protein Stat3 in regulating the EMT program in human
cervical carcinoma. We show that IL-6R and Stat3 were highly
expressed in human cervical squamous cell carcinoma (CSCC) tissues,
and the expression of EMT markers was reversed in
well-differentiated and poorly-differentiated human CSCC.
Furthermore, IL-6 exposure of cervical carcinoma cell lines induced
IL-6R and Stat3 expression, markedly promoting cell growth, and
altered cell morphology. The treatment of cervical carcinoma cells
with IL-6 resulted in significant upregulation of Vimentin
expression and downregulation of E-Cadherin expression. Knockdown
of Stat3 in cervical carcinoma cells significantly reversed the
IL-6-induced EMT program, suggesting that Stat3 is necessary for
IL-6-induced EMT in progression of human cervical carcinoma.
Moreover, we show a correlation between Stat3 and EMT regulatory
factor Slug in cervical carcinoma cells. Our findings, therefore,
identify a novel mechanism of IL-6-induced EMT program and a
potential therapeutic target for human cervical carcinoma.
Materials and methods
Primary antibodies and reagents
Recombinant human IL-6, human IL-6 receptor α and β
antibodies, IL-6 receptor inhibitor and β-actin were obtained from
Sigma. Stat3 and p-Stat3 antibodies were obtained from Cell
Signaling Technology. E-Cadherin and p-Slug antibodies were
obtained from BD Transduction Laboratories. Ki67 antibody was
obtained from Dako. CK8, CK14, CDK4, Cyclin D1 and p63 antibodies
were obtained from Santa Cruz Biotechnology, Inc. Vimentin antibody
was obtained from Chemicon.
Human normal and cervical squamous cell
carcinoma tissues
Human normal and CSCC tissues were obtained from the
Department of Pathology and Pathophysiology, School of Medicine,
Jianghan University in Wuhan, China, and from Department of
Gynecologic Oncology, Beijing Obstetrics and Gynecology Hospital,
Capital Medical University in Beijing, China, respectively. The
ethics were reviewed and approved by the board of the hospital at
Jianghan University and by the board of the hospital at Capital
Medical University. Patients with previous radiotherapy or
chemotherapy were excluded from this study. All human normal and
CSCC tissues were fixed in 10% formalin until analyzed.
Cell culture
The HeLa cervical cancer cell line (HPV-positive)
and the C33A cervical cancer cell line (HPV-negative) were obtained
from American Type Culture Collection. Both cell lines were grown
in Dulbecco’s Modified Eagle’s medium (DMEM) (HyClone) supplemented
with 10% fetal bovine serum (FBS) (Gemini Bio-Products) and 1%
penicillin/streptomycin solution (Thermo Fisher Scientific). Cells
were incubated at 37°C in a humidified incubator with 5%
CO2 until cells reach 80% confluence, then cultured in
serum-free medium for 24 h. Triplicate wells of cells were
incubated for up to 48 h in the presence of 0–200 ng/ml IL-6. In
some experiments, cells were also treated with 10 μM
curcumin (an inhibitor of IL-6 receptor). Cells were evaluated for
mRNA and/or protein levels of IL-6 receptor, E-Cadherin, Vimentin,
p-Slug, p-Stat3 and Stat3.
Tissue immunofluorescence staining
Human normal and CSCC tissues were fixed in 10%
formalin and embedded in paraffin. After deparaffinization,
sections were boiled in an antigen unmasking solution for 15 min
and blocked in 5% non-fat milk in PBS for 30 min at room
temperature. Sections were incubated with primary antibodies (Ki67,
1:200; CDK4, 1:100; Cyclin D1, 1:100; CK8, 1:100; CK14, 1:200; p63,
1:50; E-Cadherin, 1:100; Vimentin, 1:200; IL-6Rα, 1:100; IL-6Rβ,
Stat3, 1:100; and p-Stat3 1:100) overnight at 4°C, followed by
incubation with fluorescent-conjugated secondary antibodies
(Molecular Probes) for 1 h at room temperature in the dark.
Mounting medium (Vector Laboratories) was used with
4′,6′-diamino-2-phenylindole (DAPI) at 1 μg/ml for 30 min,
and the sections were analyzed by fluorescence microscopy with a
Carl Zeiss Axioscope (Carl Zeiss).
Immunocytochemical staining
Following treatment with IL-6 (50 ng/ml) or IL-6 +
curcumin (10 μM) for 48 h, HeLa and C33A cells were plated
on 8-well coverslips and fixed with ice cold methanol for 5 min at
−20°C. Cells were washed 3 times with ice-cold PBS, exposed to 0.1%
Triton X-100 in PBS for 5 min, and then washed with 0.1% Triton-X
100 in PBS. Cells were incubated with primary antibodies diluted in
PBS with 3% bovine serum albumin (BSA) followed by a 1-h incubation
at room temperature in 0.1% Nonidet P-40 in PBS. Coverslips were
incubated with a solution containing fluorescent
isothiocyanate-conjugated goat anti-mouse IgG (green) (Wako Pure
Chemical Industries) and Rhodamine-conjugated goat anti-rabbit IgG
(red) (Sigma) for 1 h at room temperature. Finally, the coverslips
were mounted with 4′,6′-diamino-2-phenylindole (DAPI) at 1
μg/ml for 30 min and analyzed by fluorescence microscopy
with a Carl Zeiss Axioscope (Carl Zeiss, Inc.).
RNA extraction and real-time polymerase
chain reaction (PCR)
Total RNA was extracted using RNA STAT-60 (Tel-Test)
according to the manufacturer’s protocol. First-strand cDNA was
synthesized using 1 μg of total RNA in a 20-μl final
volume by reverse transcription utilizing SuperScript II Reverse
Transcriptase (Invitrogen) with oligo-dT(18)-primers (Invitrogen). The real-time
PCR reactions were performed using SYBR-Green Master Mix kit
according to the manufacturer’s instructions (Applied Biosystems).
RNA for IL-6, E-Cadherin, Vimentin, and Stat3 was amplified using
ABI Prism 7000 Sequence Detection System (Applied Biosystems). The
primers for this study (Invitrogen) are shown in Table I. For all real-time PCR studies,
negative controls were a non-reverse transcriptase reaction and a
non-sample reaction (data not shown). Glyceraldehyde-3-phosphate
dehydrogenase (GAPDH) was amplified as an internal standard.
 | Table I.Primer sequences for amplification of
human Interleukin-6, Vimentin, E-Cadherin, Stat3 or GAPDH. |
Table I.
Primer sequences for amplification of
human Interleukin-6, Vimentin, E-Cadherin, Stat3 or GAPDH.
| Gene (human) | GeneBank accession
no. | Primers
| Product size
(bp) |
|---|
| Sense | Anti-sense |
|---|
| Interleukin-6 | NM_000600 |
5′-TCTCGAGAGCCCAGCTATGAACTC-3′ |
5′-ATAGCGGCCGCTTACTACATTTGCCGAAGA-3′ | 648 |
| Vimentin | NM_003380 |
5′-GACAATGCGTCTCTGGCACGTCTT-3′ |
5′-TCCTCCGCCTCCTGCAGGTTCTT-3′ | 229 |
| E-Cadherin | NM_044560 |
5′-CCCATCAGCTGCCCAGAAAATGAA-3′ |
5′-CTGTCACCTTCAGCCATCCTGTTT-3′ | 174 |
| Stat3 | NM_213662 |
5′-TTGCCAGTTGTGGTGATC-3′ |
5′-AGAACCCAGAAGGAGAAGC-3′ | 313 |
| GAPDH | NM_002046 |
5′-TGCACCACCAACTGCTTAGC-3′ |
5′-GGCATGGACTGTGGTCATGAG-3′ | 66 |
Western blotting
HeLa and C33A cell lysates (1 mg/ml) were run on 12%
sodium dodecyl sulfate/polyacrylamide gels under non-reducing
conditions and transferred to nitrocellulose. After blocking the
membrane with 3% non-fat milk, the membrane was incubated with
monoclonal or polyclonal anti-IL-6 (1:500), anti-IL-6 receptor
(1:1000), p-Stat3 (1:1000), Stat3 (1:2000), E-Cadherin (1:500),
Vimentin (1:1000), p-Slug (1: 500) and β-actin (1:100) overnight at
4°C. The membrane was then incubated for 1 h with HRP-linked
anti-mouse or anti-rabbit IgG (1:5000) diluted in TBST (10 mmol/l
Tris-HCl, pH 8.0, containing 150 mmol/l NaCl and 0.1% Tween-20) and
washed extensively with TBST and detected using the enhanced
chemiluminescence system. The membranes were exposed to X-ray film
(Amersham Biosciences). Intensities of immunoreactive bands were
quantified using image analysis software (Scion).
Transient transfection of small
interfering RNA
HeLa and C33A cells were transfected with small
interfering RNA (siRNA) of Stat3 (Invitrogen) and scrambled
oligonucleotide siRNA as a negative control using lipofectamine
RNAiMAX (Invitrogen) according to the manufacturer’s protocols.
Transfection efficiency was confirmed by western blot analysis.
Statistical analysis
All data were assessed using the Student’s t-test.
Levels of statistical significance were set at P<0.05.
Results
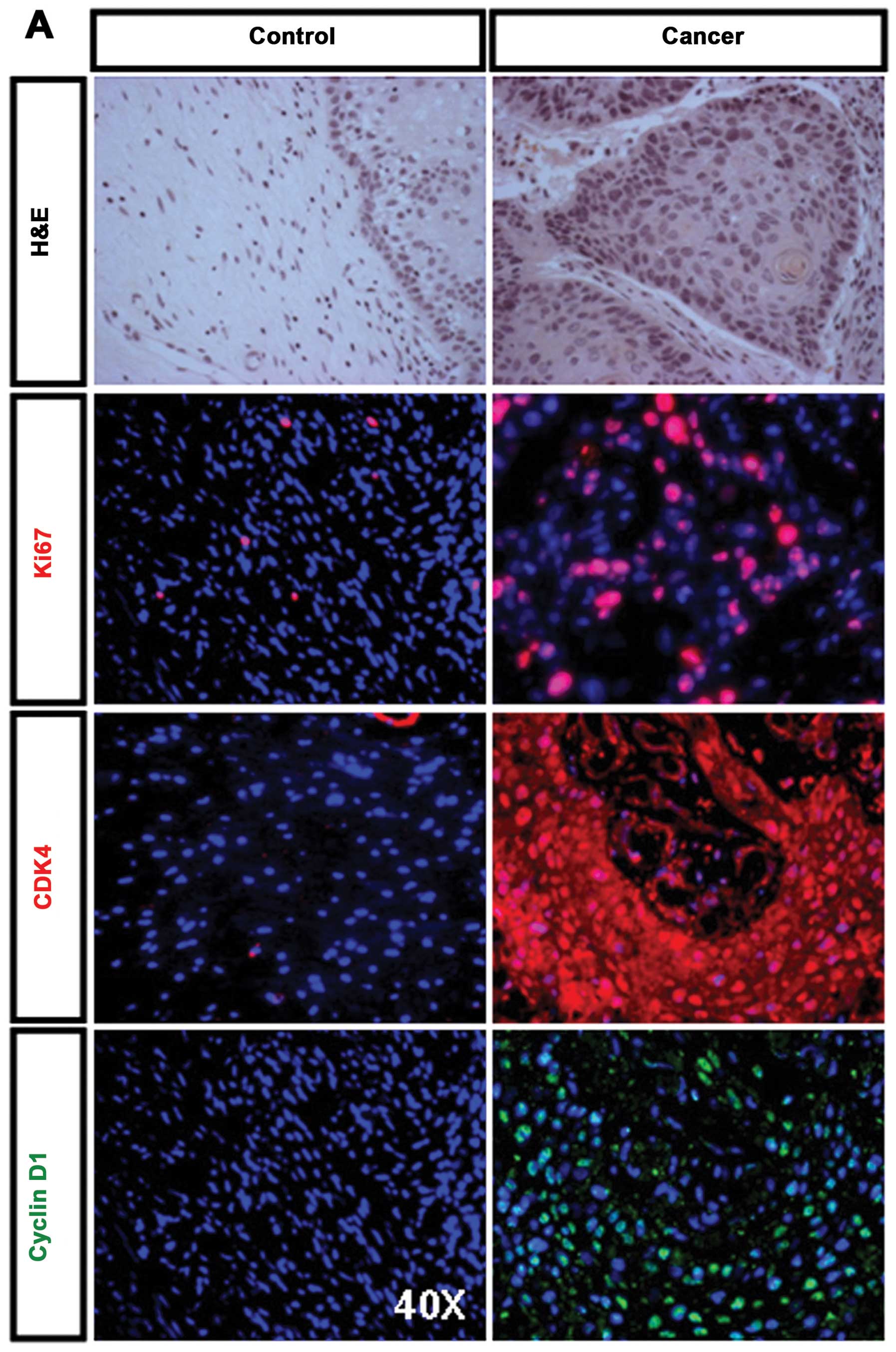
Immunohistochemical features of human
CSCC tissues
To examine immunohistochemical characteristics of
human CSCC, 10 normal human cervical epithelia and 16 human CSCC
tissues were randomly selected for this study. Well-differentiated
and poorly-differentiated human CSCC were diagnosed by two
pathologists. H&E staining showed histological features of
well-differentiated CSCC as compared to normal cervical epithelium
(Fig. 1A, upper panel).
Immunofluorescence staining for Ki67 in well-differentiated CSCC
tissues showed that the majority of Ki67-positive cells (indicated
by red nuclear staining) are restricted to the basal and parabasal
layers (Fig. 1A, right middle
panel). Immunofluorescence staining for CDK4 showed a uniform
cytoplasmic and nuclear staining in well-differentiated CSCC
tissues (Fig. 1A, right middle
panel). Strong nuclear staining for Cyclin D1 was observed in
well-differentiated CSCC tissues (Fig.
1A, right bottom panel). The normal cervical epithelia were
either completely non-immunoreactive or exhibited weak
immunoreactivity (Fig. 1A, left
panels). Cytokeratin (CK) is one of intermediate filament proteins
and has been used as epithelial-developed cancer marker (24). To examine expression patterns of
cytokeratin in human CSCC, well-differentiated and
poorly-differentiated CSCC tissues were selected for staining with
anti-CK8 and anti-CK14 antibodies. The significant positive
staining for CK8 and CK14 was observed in all well-differentiated
(Fig. 1B, bottom panel) and a few
poorly-differentiated (Fig. 1B,
upper panel) CSCC tissues. Both CK8 and CK14 showed more extensive
and intense expression in well-differentiated CSCC tissues
(Fig. 1B, bottom panel) as
compared to poorly-differentiated CSCC tissues (Fig. 1B, upper panel). The p63 is a member
of p53 family. Studies have shown that p63 plays an important role
in the development and differentiation of various epithelial cells
(25). To determine the expression
of p63 in human CSCC, well-differentiated CSCC tissues were
selected for immunofluorescence staining with anti-p63 antibody.
The p63 was expressed robustly in well-differentiated CSCC tissues
(Fig. 1C, upper panel), and
colocalization of p63 and Ki67 in cancer cells was demonstrated to
significantly induce proliferation potential (Fig. 1C, bottom panel). Quantitative
analysis shows that 60% of p63 positive cancer cells were
colocalized with Ki67 in well-differentiated CSCC tissues (Fig. 1D). These data indicate that cell
cycle-associated proteins, cytokeratins and p63 as
immunohistochemical markers are highly expressed in
well-differentiated human CSCC tissues.
Expression of IL-6R, Stat3, E-Cadherin
and Vimentin in human CSCC tissues
IL-6 has been reported to play an important role in
tumor progression and metastasis in a variety of human cancers as a
central proinflammatory factor. To examine the expression of IL-6R
and its downstream protein Stat3 in human CSCC tissues, IL-6R and
Stat3 expression were analyzed using immunofluorescence staining.
Both IL-6Rα and IL-6Rβ staining were observed in cancer lesions in
15 out of 16 (93.8%) of human CSCC tissues (Fig. 2A, right upper panels), whereas the
negative control showed lack of staining for IL-6Rα and IL-6Rβ in
normal cervical tissues and the positive staining for IL-6Rα and
IL-6Rβ was restricted to the most basal and parabasal cells of
normal squamous epithelium (Fig.
2A, left upper panels). Similarly, staining of Stat3 and
p-Stat3 was observed in cancer lesions in 14 out of 16 (87.5%) of
the human CSCC tissues (Fig. 2A,
right bottom panels), whereas the negative control showed lack of
staining for Stat3 and p-Stat3 in normal cervical tissues (Fig. 2A, left bottom panels). EMT is
characterized by dramatic phenotypic changes in human cancers. To
evaluate expression patterns of EMT in human CSCC tissues, both
well-differentiated and poorly-differentiated CSCC tissues were
selected for immunofluorescence staining with epithelial marker
E-Cadherin and mesenchymal marker Vimentin. E-Cadherin was absent
in poorly-differentiated CSCC tissues (Fig. 2B, bottom panel), but was markedly
observed in well-differentiated CSCC tissues (Fig. 2B, upper panel). In contrast,
Vimentin was absent in well-differentiated CSCC tissues (Fig.2-B, upper panel), but was clearly
expressed in poorly-differentiated CSCC tissues (Fig. 2B, bottom panel). These data
indicate that IL-6R and p-Stat3 are positively expressed in human
CSCC tissues, and well-differentiated CSCC expresses high levels of
E-Cadherin and low levels of Vimentin, whereas
poorly-differentiated CSCC expresses high levels of Vimentin and
low levels of E-Cadherin.
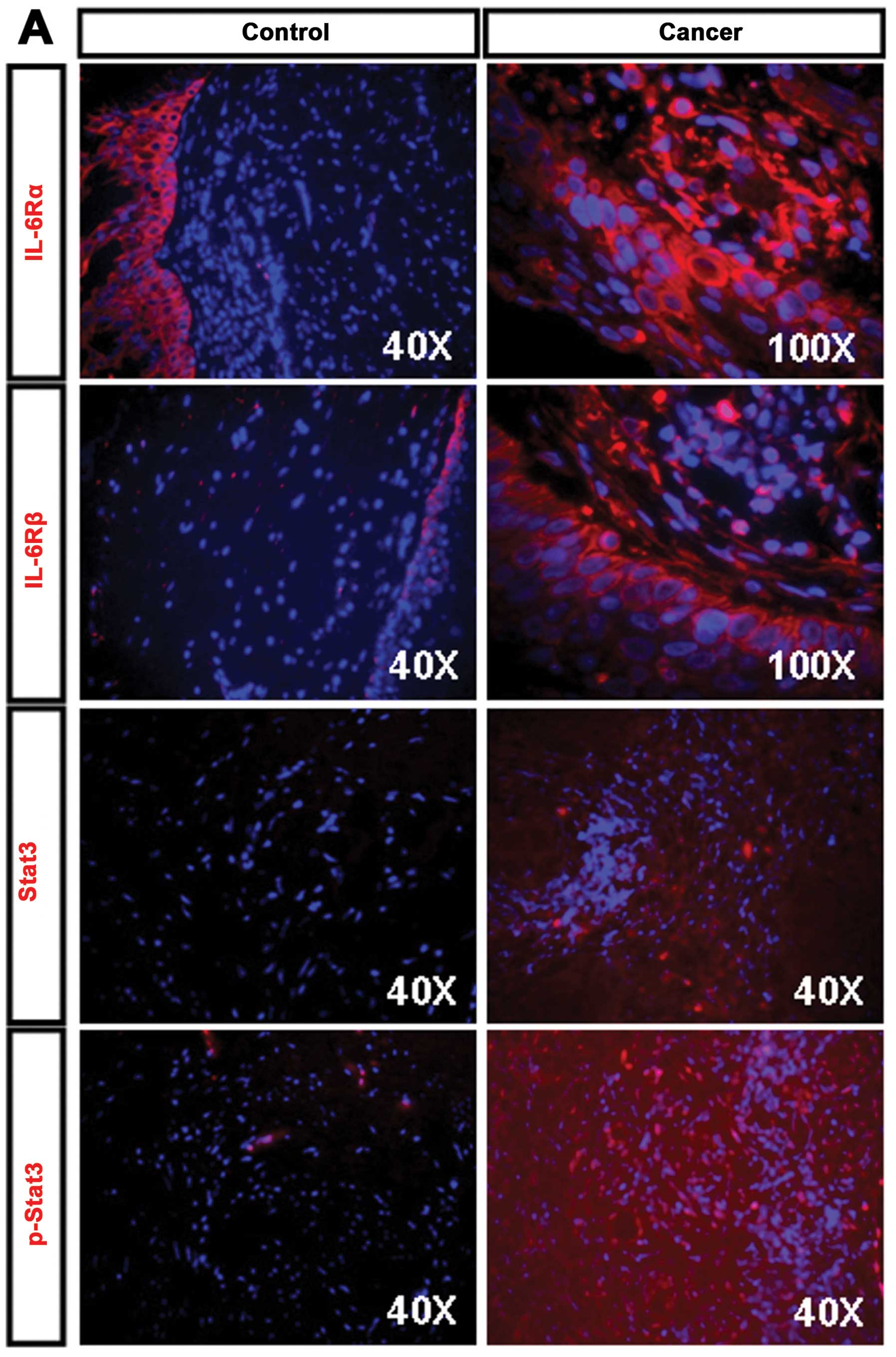 | Figure 2.Expression of IL-6R, Stat3, E-Cadherin
and Vimentin in human CSCC tissues. (A) Human normal cervical and
well-differentiated CSCC tissues were stained using anti-IL-6Rα,
IL-6Rβ Stat3 and p-Stat3 antibodies. Strong staining of IL-6Rα and
IL-6Rβ was detected on the cancer cell membrane in CSCC tissues
(right upper and middle panels), whereas the positive staining of
IL-6Rα and IL-6Rβ was only restricted in squamous epithelium in
normal cervical tissues (left upper and middle panels). The
expression of Stat3 was weakly detected in CSCC tissues (right
middle panel), whereas p-Stat3 showed strong nuclear expression in
CSCC tissues (right bottom panel) as comparing to normal cervical
tissues (left middle and bottom panels). Nuclei were stained with
DAPI (blue) (original magnification, ×400 and ×1000). (B)
E-Cadherin was weakly expressed in poorly-differentiated human CSCC
tissues (bottom panel), but it was markedly observed in
well-differentiated human CSCC tissues (upper panel). In contrast,
Vimentin was lost in well-differentiated human CSCC tissues (upper
panel), but it was clearly expressed in poorly-differentiated human
CSCC tissues (bottom panel). Nuclei were stained with DAPI
(original magnification, ×400). |
IL-6 stimulation induces IL-6 receptor
expression in HeLa and C33A cells
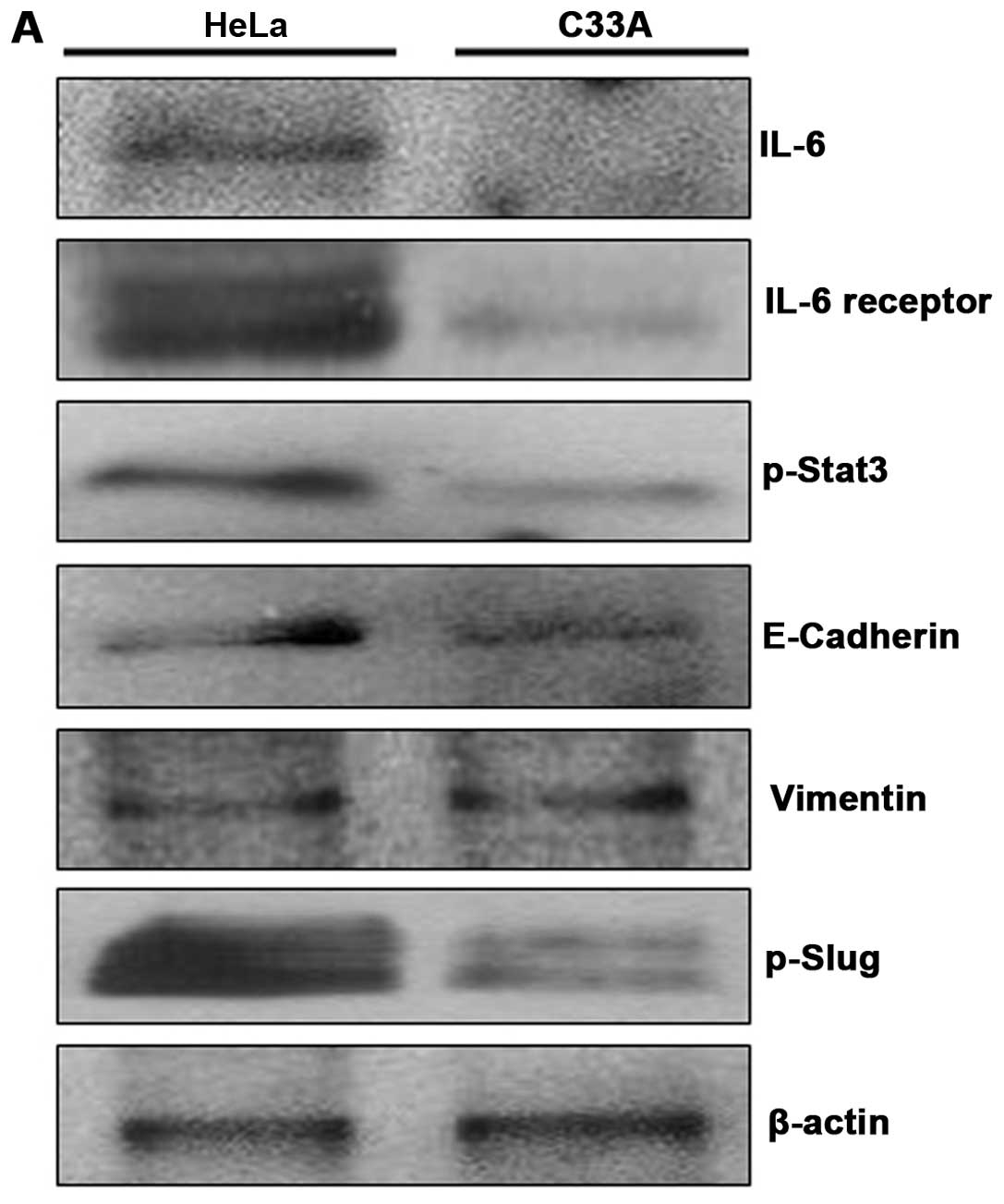
To examine expression level of IL-6 receptor in HeLa
and C33A cells. We first investigated protein expression levels of
endogenous IL-6, IL-6 receptor, p-Stat3, E-Cadherin, Vimentin and
p-Slug using western blot analysis. The expression of IL-6, IL-6R
and p-Stat3 proteins was significantly weaker in C33A cells as
compared to HeLa cells (Fig. 3A).
Similarly, a low level in endogenous p-Slug protein was observed in
C33A cells as compared to HeLa cells (Fig. 3A). However, protein levels of
endogenous E-Cadherin and Vimentin were not significantly different
in both HeLa and C33A cells (Fig.
3A). We next examined expression levels of IL-6R protein and
mRNA in HeLa and C33A cells in response to IL-6 as compared to
control and IL-6 + curcumin for 48 h using western blot analysis
and real-time RT-PCR. The expression levels of IL-6R protein and
mRNA were significantly increased in IL-6-treated HeLa (Fig. 3B and C, quantitative results) and
C33A (Fig. 3D and E, quantitative
results) cells as comparing to control and IL-6 + curcumin. These
data indicate that expression level of IL-6R is similar in response
to IL-6 stimulation in HeLa and C33A cells, while endogenous IL-6R
and p-Stat3 are weakly expressed in C33A cells.
Treatment of IL-6 mediates cell
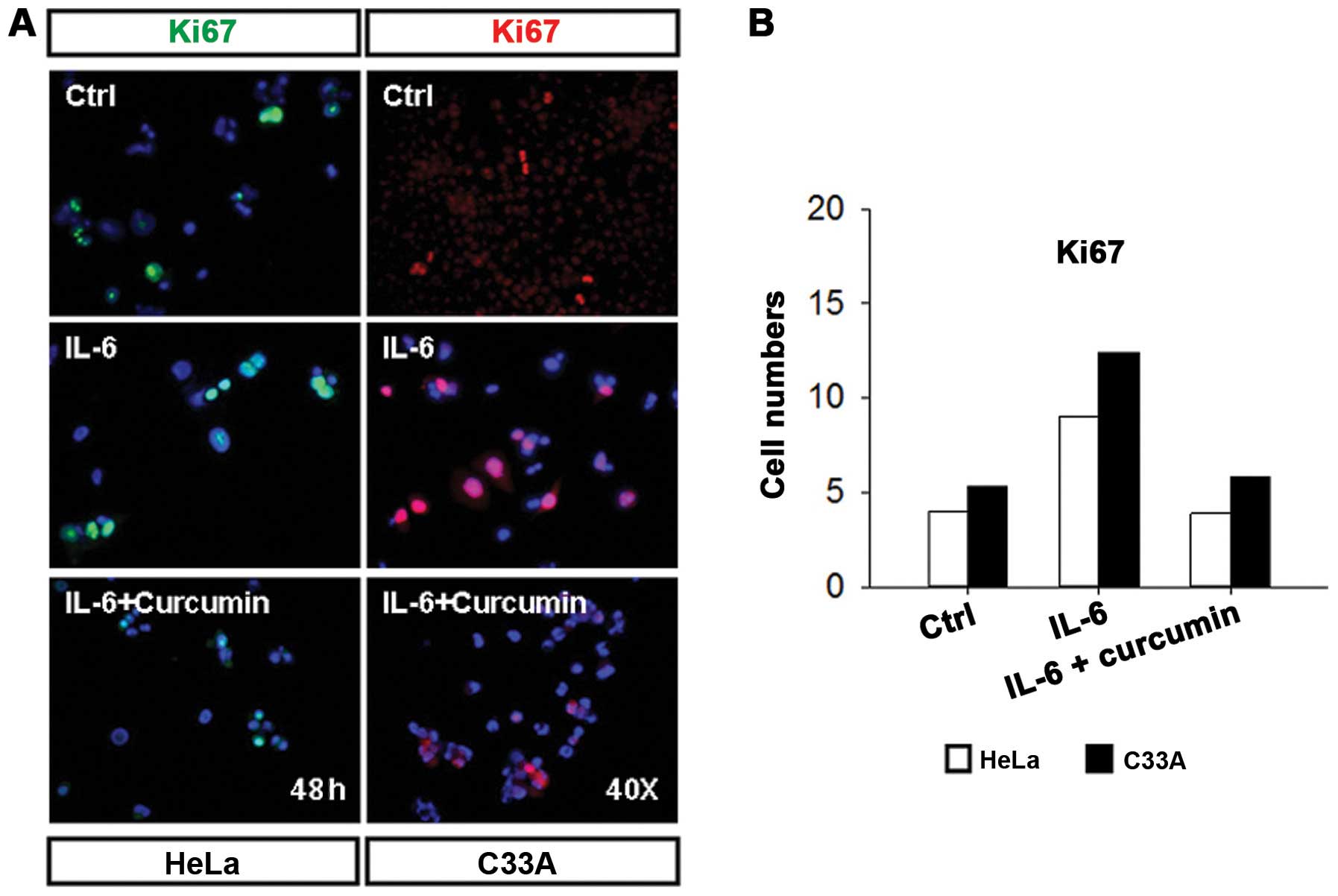
proliferation and alters cell morphology
There is increasing evidence to show that IL-6 is a
regulator of cancer cell proliferation (26). To examine whether IL-6 mediates
growth of human cervical carcinoma cells, HeLa and C33A cells were
treated with IL-6 or IL-6 + curcumin for 48 h. Cells were stained
with anti-Ki67 antibody (Fig. 4A).
The cell proliferation was significantly mediated by IL-6
stimulation in IL-6-treated HeLa and C33A cells as compared to
control and IL-6 + curcumin treatment groups (Fig. 4A and B, quantitative results). To
examine whether IL-6 mediates changes in cell morphology, we
treated HeLa cells with IL-6 for 48 h. In the absence of IL-6
treatment, HeLa cells exhibited well-organized cell-cell
association and islet-like structure, and characteristics of
squamous cell carcinoma (Fig. 4C,
upper panel). Interestingly, IL-6 treatment induced cell elongation
and increased scattering, resulting in a similar morphology to
mesenchymal cells (Fig. 4C, bottom
panel). These data indicate that IL-6 is able to mediate cell
proliferation and alter cell morphology in HeLa cells.
IL-6 stimulation induces the EMT program
in HeLa and C33A cells
EMT is a process by which epithelial cells lose
their cell polarity and cell-cell adhesion, and gain migratory and
invasive properties to become mesenchymal cells. Many factors have
been reported to induce EMT program in cancer progression and
metastasis (27). To evaluate
whether EMT program caused by IL-6 exposure in human cervical
carcinoma, we first examined the expression of EMT markers in HeLa
cells by immunofluorescence staining. Cells treated with IL-6 for
48 h decreased E-Cadherin expression, but significantly increased
Vimentin expression as comparing to the cells treated with IL-6 +
curcumin (Fig. 5A). Similar
effects of E-Cadherin and Vimentin were observed in C33A cell line
(Fig. 5D). Interestingly, a
transition to spindle-shaped morphology was also observed in HeLa
cells (Fig. 5A), but not in C33A
cells (Fig. 5D). We next
determined the expression levels of E-Cadherin and Vimentin in HeLa
cells in response to IL-6 treatment for 48 h by western blot
analysis and real-time RT-PCR. The protein and mRNA levels of
E-Cadherin were significantly decreased in HeLa cells treated with
IL-6 as comparing to HeLa cells treated with DMSO and IL-6 +
curcumin (Fig. 5B). In contrast,
the protein and mRNA levels of Vimentin markedly increased in HeLa
cells in response to IL-6 treatment as comparing to HeLa cells
treated with DMSO and IL-6 + curcumin (Fig. 5C). Similar effects of E-Cadherin
and Vimentin were observed in C33A cells (Fig. 5E and F). These results indicate
that IL-6 exhibits an important role on the EMT induction in human
cervical carcinoma cells by downregulation of epithelial markers
and upregulation of mesenchymal markers.
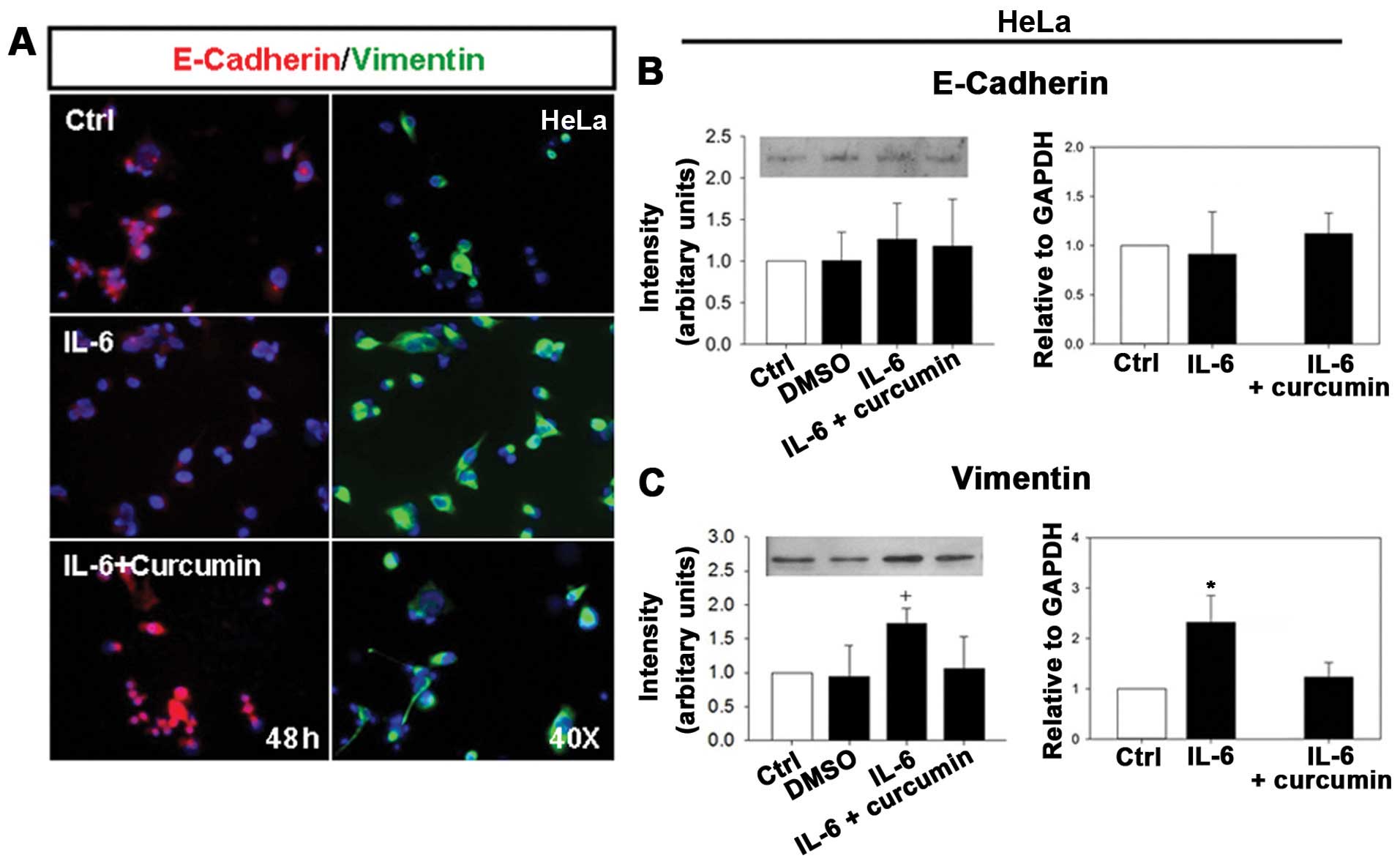 | Figure 5.IL-6 treatment induces EMT in HeLa and
C33A cells. (A) HeLa cells were treated without IL-6 (as a control)
and with IL-6 (50 ng/ml), IL-6 + curcumin (10 μM) for 48 h.
Cells were stained with anti-E-Cadherin antibody and
rhodamine-conjugated secondary antibody (red) or stained with
anti-Vimentin antibody and fluorescein isothiocyanate-conjugated
secondary antibody (green). Nuclei were stained with DAPI (blue)
(original magnification, ×400). (B and C) HeLa cells were treated
without IL-6 (as a control) and with DMSO, IL-6 (50 ng/ml) and IL-6
+ curcumin (10 μM) for 48 h. Total protein was extracted
from control, DMSO and treated cells. The protein levels of
E-Cadherin (B, left) and Vimentin (C, left) were analyzed using
western blot analysis. Similarly, total RNA was extracted from
control and treated cells. The mRNA levels of E-Cadherin (B, right)
and Vimentin (C, right) were determined using real-time RT-PCR.
Real-time PCR for GADPH served as an internal control. (D) C33A
cells were treated without IL-6 (as a control) and with IL-6 (50
ng/ml), IL-6 + curcumin (10 μM) for 48 h. Cells were stained
with anti-E-Cadherin antibody and rhodamine-conjugated secondary
antibody (red) or stained with anti-Vimentin antibody and
fluorescein isothiocyanate-conjugated secondary antibody (green).
Nuclei were stained with DAPI (blue) (original magnification,
×400). (E and F) C33A cells were treated without IL-6 (as a
control) and with DMSO, IL-6 (50 ng/ml) and IL-6 + curcumin (10
μM) for 48 h. Total protein was extracted from control, DMSO
and treated C33A cells. The protein levels of E-Cadherin (E, left)
and Vimentin (F, left) were analyzed using western blot analysis.
Similarly, total RNA was extracted from control and treated cells.
The mRNA levels of E-Cadherin (E, right) and Vimentin (F, right)
were determined using real-time RT-PCR. Real-time PCR for GADPH
served as an internal control. The intensity of bands was
quantified using ImageJ software and normalized to β-actin. |
Treatment of HeLa and C33A cells with
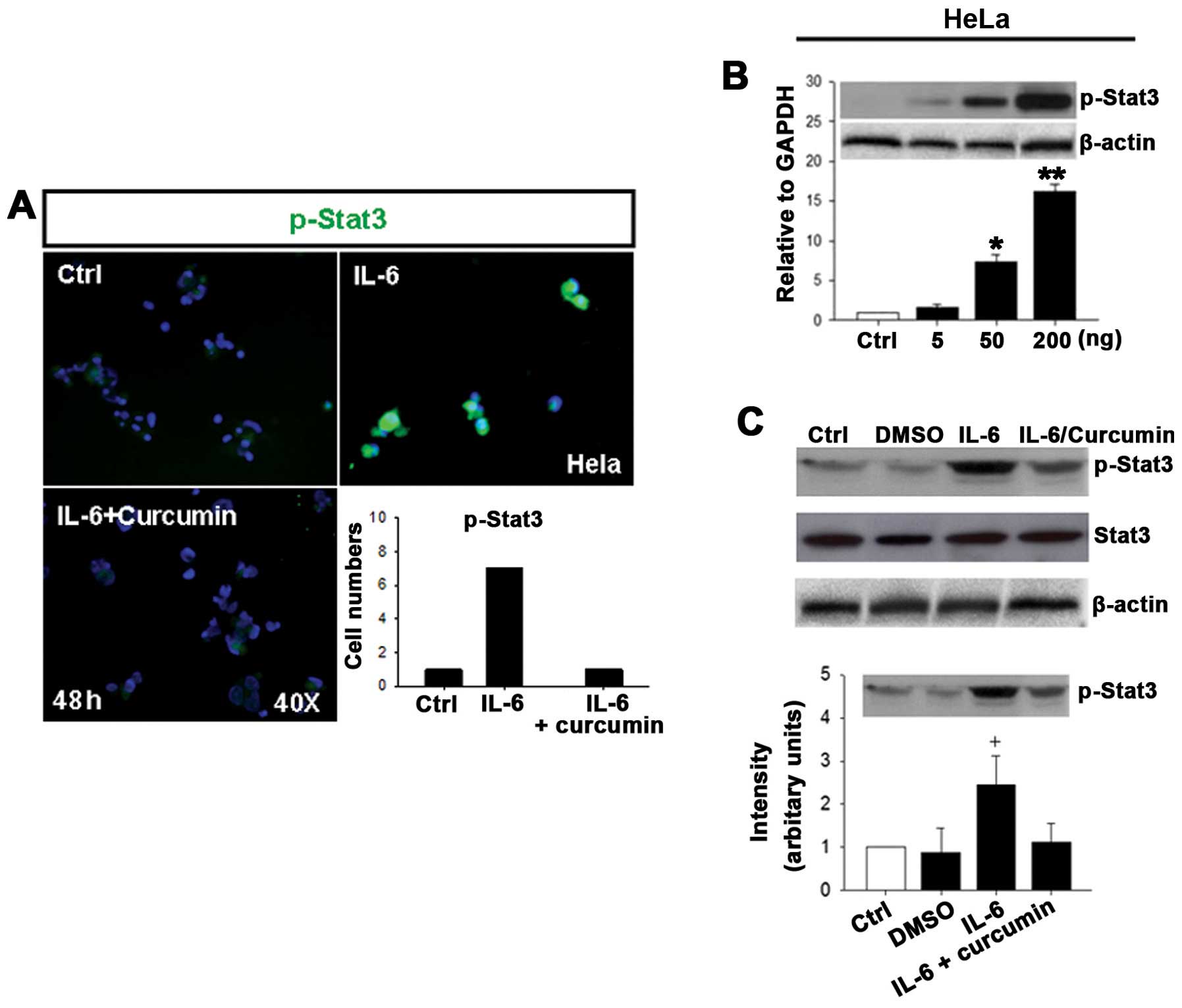
IL-6 increases phosphorylation of Stat3
Stat3 is a downstream protein of IL-6R and plays a
potential role in cervical cancer development. To investigate
whether IL-6 directly activates Stat3 in human cervical carcinoma,
we examined the expression of p-Stat3 in HeLa cells after treatment
with IL-6 for 48 h. IL-6 treatment significantly increased the
expression of p-Stat3 as compared to control and IL-6 + curcumin
groups as demonstrated by immunofluorescence (Fig. 6A, quantitative results). Also, an
increase in p-Stat3 protein levels was observed in HeLa cells
treated with various concentrations of IL-6 for 48 h and was
significant difference at concentrations of 50 and 200 ng/ml
(Fig. 6B, quantitative results).
Similar effect of p-Stat3 expression by IL-6 stimulation was
observed by real-time RT-PCR in HeLa cells (data not shown). Stat3
and p-Stat3 protein levels were then measured by western blot
analysis following treatment of HeLa cells with IL-6 and IL-6 +
curcumin for 48 h. No changes in total Stat3 levels were noted at
48 h in treated cells as comparing to control, DMSO and IL-6 +
curcumin groups (Fig. 6C). In
contrast, IL-6 stimulation significantly increased expression of
p-Stat3 protein as comparing to control, DMSO and IL-6 + curcumin
groups (Fig. 6C, quantitative
results).
We next determined the p-Stat3 expression in C33A
cells in response to IL-6 treatment for 48 h by immunofluorescence
and western blot analysis. IL-6 treatment significantly increased
the expression of p-Stat3 in C33A (Fig. 6D). In addition, the p-Stat3
expression was increased in C33A cells treated with various
concentrations of IL-6 for 48 h (Fig.
6E, quantitative results). The protein levels of p-Stat3 were
significantly increased in C33A cells treated with IL-6 as
comparing to C33A cells treated with DMSO and IL-6 + curcumin
(Fig. 6F, quantitative results).
These results indicate that IL-6 can directly induce
phosphorylation of Stat3 in human cervical carcinoma cells, HeLa
and C33A.
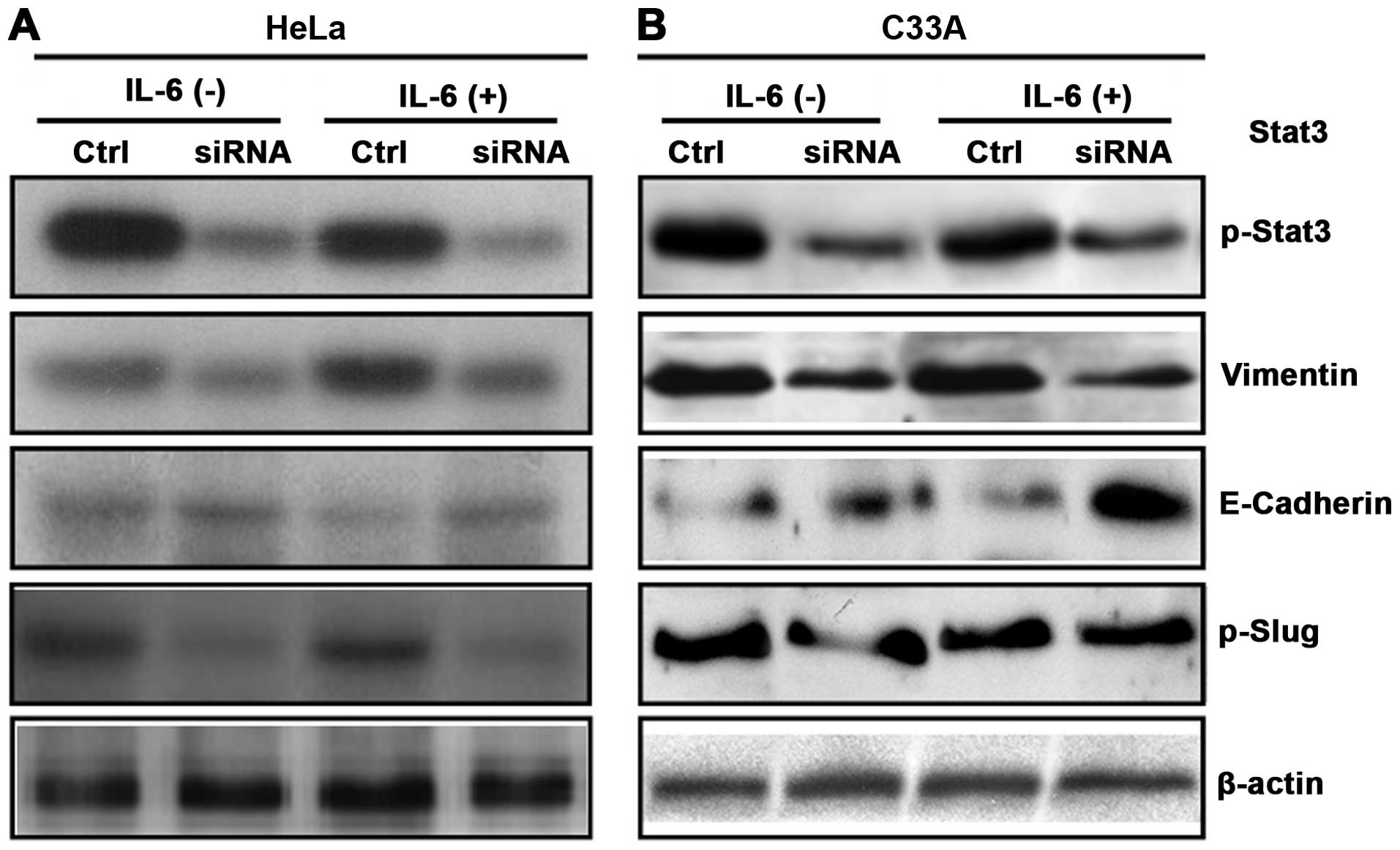
IL-6-induces the EMT program via Stat3 in
HeLa and C33A cells
To evaluate the role of Stat3 in IL-6-induced EMT
program, we examined expression of p-Stat3, E-Cadherin, Vimentin
and p-Slug in HeLa and C33A cells treated with and without IL-6 for
48 h after knockdown of Stat3. The expression of p-Stat3 was
markedly decreased in HeLa cells treated with and without IL-6 as
compared to control by western blot analysis (Fig. 7A). Similar effects were observed in
C33A cell line (Fig. 7B). The
Vimentin and E-Cadherin levels were then measured by western blot
analysis following treatment of HeLa cells with and without IL-6
for 48 h. The protein levels of E-Cadherin were significantly
increased in HeLa cells treated with IL-6 as compared to HeLa cells
treated with control, but no difference for E-Cadherin was observed
in HeLa cells untreated with IL-6 in control and siRNA groups
(Fig. 7A). In contrast, the
protein levels of Vimentin markedly decreased in HeLa cells in
response to IL-6 treatment as compared to HeLa cells treated with
control, but Vimentin levels were significantly decreased in HeLa
cells untreated with IL-6 in siRNA group as comparing to control
(Fig. 7A). Similar effects were
observed in C33A cell line (Fig.
7B). IL-6 treatment in HeLa cells significantly decreased
expression of p-Slug protein in siRNA group as compared to control.
In contrast, p-Slug levels were also decreased in HeLa cells
untreated with IL-6 in siRNA group as compared to control (Fig. 7A). No changes in p-Slug protein
levels were noted in C33A cells treated and untreated with IL-6 in
control and siRNA groups (Fig.
7B). These results indicate that IL-6-induced E-Cadherin
downregulation and Vimentin upregulation were dramatically blocked
by knockdown of Stat3, suggesting that Stat3 may participate in
IL-6-induced EMT program in HeLa and C33A cells.
Discussion
EMT is an important process in many types of human
epithelial carcinoma including cervical carcinoma (19). Despite HPV infection has been
reported to be a major risk for developing cervical carcinoma,
however cervical carcinoma is also closely associated with
inflammatory conditions (3,4).
Studies have shown that many Inflammation mediators can trigger EMT
program in progression of human cancers. IL-6 is one of
pro-inflammatory cytokines that activates its downstream protein
Stat3 within the tumor microenvironment and plays an important role
in the pathogenesis of human cervical carcinoma (28,29).
In this study, we investigated the role of IL-6 as an inducer of
EMT through its downstream signaling molecule Stat3 in human
cervical carcinoma.
In human CSCC tissues, we observed overexpression of
IL-6R signaling molecules and EMT markers, suggesting that these
molecules and markers are necessary for development of human CSCC.
Interestingly, the expression of epithelial cell marker E-Cadherin
was more dominant than mesenchymal cell marker Vimentin in
well-differentiated CSCC tissues, whereas Vimentin was more
strongly expressed in poorly-differentiated CSCC tissues than
E-Cadherin. This is consistent with data obtained from Cheng et
al (30). Further evidence
from cytokeratins (CK8 and CK14) and p63 staining support that the
expression of epithelial cell markers was the major phenotype in
well-differentiated human CSCC tissues. Our data show that 60% of
p63 positive cancer cells were colocalized with Ki67 in
well-differentiated CSCC tissues, suggesting that the
colocalization of p63 and Ki67 in cancer cells was demonstrated to
significantly induce proliferation potential. We also observed that
the expression of N-Cadherin and β-Catenin (data not shown) was
significantly increased in well-differentiated human CSCC tissues.
E-Cadherin and N-Cadherin are known to anchor and sequester
β-Catenin on the cell membrane and preventing its activation; the
activation of β-Catenin may result from the upregulation of
E-Cadherin and the downregulation of N-Cadherin (31). Moreover, we observed a loss of
Vimentin in well-differentiated CSCC lesion and a strong expression
of Vimentin in the connective tissues surrounding the CSCC lesion.
However, we do not know whether EMT can be triggered directly by
IL-6 in human CSCC tissue, because we were unable to examine the
relationship between IL-6 and EMT program using human tumor
tissues.
To identify whether IL-6 is involved in EMT
induction and development of cervical carcinoma, HeLa, an
HPV-positive cervical cancer cell line and C33A, an HPV-negative
cervical cancer cell line were chosen for in vitro
experiment. Interestingly, both HeLa and C33A cells treatment with
IL-6 promoted IL-6R expression, cell proliferation, and morphology
alteration, despite the expression levels of endogenous IL-6, IL-6R
and p-Stat3 were weaker in C33A cells as comparing to HeLa cells.
One possibility we consider is that the activation of IL-6R
signaling may not be dependent on HPV infection. We found that IL-6
treatment in HeLa and C33A cells decreased the expression of
epithelial cell markers such as E-Cadherin and β-Catenin (data not
shown), but increased the expression of mesenchymal cell markers
such as Vimentin and N-Cadherin (data not shown). Interestingly, a
transition to spindle-shaped morphology was detectable in HeLa
cells, but not in C33A cells. We also found that Stat3 was
upregulated upon IL-6 stimulation in both HeLa and C33A cells.
Furthermore, induction of Stat3 by IL-6 was successfully inhibited
by the IL-6R inhibitor curcumin, indicating that IL-6-mediated
Stat3 induction and subsequent EMT are dependent on IL-6R
activation. Although Sullivan et al (32) have reported that IL-6 can induce
EMT change in human breast cancer cells through Stat3 activation,
whether Stat3 plays a role in EMT induction in human cervical
carcinoma still remains to be determined. In our present study, we
found that knockdown of Stat3 reversed IL-6-induced EMT program in
cervical carcinoma cells, suggesting that Stat3 may be involved in
EMT change and may be one of the mechanisms responsible for the
regulation of IL-6R expression. Slug has been reported as a major
transcriptional repressor to downregulate E-Cadherin expression
resulted in initiating events in EMT (33). In our study, we further found that
p-Slug expression was significantly decreased by knockdown of Stat3
in HeLa cells, but not in C33A cells, suggesting that the
activation of Stat3 is important for Slug expression in human
cervical carcinoma. However, we do not know why p-Slug expression
in HeLa cells and C33A cells is different after knockdown of Stat3.
Therefore, our data indicated that IL-6 is a novel inducer of EMT
via Stat3 in human cervical carcinoma cell lines, HeLa and
C33A.
Our study provides new insight into the regulation
of EMT by IL-6 via activation of Stat3 in human cervical carcinoma.
First, our data demonstrate that epithelial cell markers, CK8, CK14
and p63, showed more extensive and intense expression in
well-differentiated CSCC lesion with epithelial morphology,
suggesting that these markers, like E-Cadherin, may play important
roles in the development of human cervical carcinoma. Second, our
data indicate that knockdown of Stat3 resulted in upregulation of
E-Cadherin and downregulation of Vimentin in cervical carcinoma
cells, suggesting that IL-6 may participate in Stat3-induced Slug
expression in HeLa cells, and may mediate Stat3-induced
downregulation of E-Cadherin.
The present study also has some limitations.
Well-differentiated and poorly-differentiated human CSCC tissues
were surgically collected from patients rather than radiotherapy or
chemotherapy. However, the problem with this is that by only
studying tissues from a few well-differentiated and
poorly-differentiated human CSCC, determining the role that IL-6
may be playing throughout disease progression is limited.
Therefore, further studies using larger patient populations are
necessary to explore the role of IL-6 for different types of
cervical carcinoma in order to obtain more reliable results.
In conclusion, our results reveal that IL-6 induced
the EMT program in human cervical carcinoma cells via the
activation of Stat3, and Stat3 knockdown significantly reversed
IL-6-induced EMT program. These results provide a mechanistic
explanation for Stat3 as an important player in IL-6-driven EMT
program, it is possible that blockage of Stat3 may be a good
approach to treat human cervical carcinoma that are driven by
IL-6/IL-6R signaling. Overall, our findings indicate that Stat3
plays an important role in IL-6-induced EMT and may serve as an
attractive therapeutic target for human cervical carcinoma driven
by IL-6/IL-6R signaling.
Acknowledgements
This study was supported by a grant
from The Scientific Research Foundation for Returned Overseas
Chinese Scholars, Bureau of Human Resources and Social Security of
Beijing, P.R. China to J.-W.M. (grant no. 2008002).
References
|
1.
|
Thun MJ, Oliver DeLancey J, Center MM,
Jemal A and Ward EM: The global burden of cancer: priorities for
prevention. Carcinogenesis. 31:100–110. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2.
|
Lee MY and Shen MR: Epithelial-mesenchymal
transition in cervical carcinoma. Am J Transl Res. 4:1–13.
2012.PubMed/NCBI
|
|
3.
|
Castellsague X: Natural history and
epidemiology of HPV infection and cervical cancer. Gynecol Oncol.
110:S4–S7. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Burd EM: Human papillomavirus and cervical
cancer. Clin Microbiol Rev. 16:1–17. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
zur Hausen H: Papillomaviruses and cancer:
from basic studies to clinical application. Nat Rev Cancer.
2:342–350. 2002.PubMed/NCBI
|
|
6.
|
Au WW, Abdou-Salama S, Sierra-Torres CH
and Al-Hendy A: Environmental risk factors for prevention and
molecular intervention of cervical cancer. Int J Hyg Environ
Health. 210:671–678. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Wei LH, Kuo ML, Chen CA, Cheng WF, Cheng
SP, Hsieh FJ and Hsieh CY: Interleukin-6 in cervical cancer: the
relationship with vascular endothelial growth factor. Gynecol
Oncol. 82:49–56. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
Wei LH, Kuo ML, Chen CA, Chou CH, Cheng
WF, Chang MC, Su JL and Hsieh CY: The anti-apoptotic role of
interleukin-6 in human cervical cancer is mediated by up-regulation
of Mcl-1 through a PI3-K/Akt pathway. Oncogene. 20:5799–5809. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Rose-John S: IL-6 trans-signaling via the
soluble IL-6 receptor: importance for the pro-inflammatory
activities of IL-6. Int J Biol Sci. 8:1237–1247. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Braunstein J, Brutsaert S, Olson R and
Schindler C: Stats dimerize in the absence of phosphorylation. J
Biol Chem. 278:34133–34140. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Shukla S, Shishodia G, Mahata S, et al:
Aberrant expression and constitutive activation of Stat3 in
cervical carcinogenesis: implications in high-risk human
papillomavirus infection. Mol Cancer. 9:2822010. View Article : Google Scholar
|
|
12.
|
Chen CL, Hsieh FC, Lieblein JC, et al:
Stat3 activation in human endometrial and cervical cancers. Br J
Cancer. 96:591–599. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Aggarwal BB, Kunnumakkara AB, Harikumar
KB, et al: Signal transducer and activator of transcription-3,
inflammation, and cancer: how intimate is the relationship? Ann N Y
Acad Sci. 1171:59–76. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Lee JM, Dedhar S, Kalluri R and Thompson
EW: The epithelial-mesenchymal transition: new insights in
signaling, development, and disease. J Cell Biol. 172:973–981.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Trimboli AJ, Fukino K, Bruin A, et al:
Direct evidence for epithelial-mesenchymal transitions in breast
cancer. Cancer Res. 68:937–945. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Vergara D, Merlot B, Lucot JP, Collinet P,
Vinatier D, Fournier I and Salzet M: Epithelial-mesenchymal
transition in ovarian cancer. Cancer Lett. 291:59–66. 2009.
View Article : Google Scholar
|
|
17.
|
Brabletz T, Hlubek F, Spaderna S,
Schmalhofer O, Hiendlmeyer E, Jung A and Kirchner T: Invasion and
metastasis in colorectal cancer: epithelial-mesenchymal transition,
mesenchymal-epithelial transition, stem cells and beta-catenin.
Cells Tissues Organs. 179:56–65. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Usami Y, Satake S, Nakayama F, et al:
Snail-associated epithelial-mesenchymal transition promotes
oesophageal squamous cell carcinoma motility and progression. J
Pathol. 215:330–339. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Lee MY, Chou CY, Tang MJ and Shen MR:
Epithelial-mesenchymal transition in cervical cancer: correlation
with tumor progression, epidermal growth factor receptor
overexpression, and snail up-regulation. Clin Cancer Res.
14:4743–4750. 2008. View Article : Google Scholar
|
|
20.
|
Cervantes-Arias A, Pang LY and Argyle DJ:
Epithelial-mesenchymal transition as a fundamental mechanism
underlying the cancer phenotype. Vet Comp Oncol. 11:169–184. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Chen H, Paradies NE, Fedor-Chaiken M and
Brackenbury R: E-cadherin mediates adhesion and suppresses cell
motility via distinct mechanisms. J Cell Sci. 110:345–356.
1997.PubMed/NCBI
|
|
22.
|
McInroy L and Määttä A: Down-regulation of
vimentin expression inhibits carcinoma cell migration and adhesion.
Biochem Biophys Res Commun. 360:109–114. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Vuoriluoto K, Haugen H, Kiviluoto S, et
al: Vimentin regulates EMT induction by Slug and oncogenic H-Ras
and migration by governing Axl expression in breast cancer.
Oncogene. 30:1436–1448. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24.
|
Karantza V: Keratins in health and cancer:
more than mere epithelial cell markers. Oncogene. 30:127–138. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
25.
|
Di Como CJ, Urist MJ, Babayan I, Drobnjak
M, Hedvat CV, Teruya-Feldstein J, Pohar K, Hoos A and Cordon-Cardo
C: p63 expression profiles in human normal and tumor tissues. Clin
Cancer Res. 8:494–501. 2002.PubMed/NCBI
|
|
26.
|
Lin DL, Whitney MC, Yao Z and Keller ET:
Interleukin-6 induces androgen responsiveness in prostate cancer
cells through up-regulation of androgen receptor expression. Clin
Cancer Res. 7:1773–17781. 2001.PubMed/NCBI
|
|
27.
|
Xiao D and He J: Epithelial mesenchymal
transition and lung cancer. J Thorac Dis. 2:154–159.
2010.PubMed/NCBI
|
|
28.
|
Castrilli G, Tatone D, Diodoro MG, Rosini
S, Piantelli M and Musiani P: Interleukin 1alpha and interleukin 6
promote the in vitro growth of both normal and neoplastic human
cervical epithelial cells. Br J Cancer. 75:855–859. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
29.
|
Tjiong MY, van der Vange N, ten Kate FJ,
Tjong-A-Hung SP, ter Schegget J, Burger MP and Out TA: Increased
IL-6 and IL-8 levels in cervicovaginal secretions of patients with
cervical cancer. Gynecol Oncol. 73:285–291. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
30.
|
Cheng Y, Zhou Y, Jiang W, et al:
Significance of E-cadherin, β-catenin, and vimentin expression as
postoperative prognosis indicators in cervical squamous cell
carcinoma. Human Pathol. 43:1213–1220. 2012.
|
|
31.
|
Scanlon CS, Van Tubergen EA, Inglehart RC
and D’Silva NJ: Biomarkers of epithelial-mesenchymal transition in
squamous cell carcinoma. J Dent Res. 92:114–121. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32.
|
Sullivan NJ, Sasser AK, Axel AE, Vesuna F,
Raman V, Ramirez N, Oberyszyn TM and Hall BM: Interleukin-6 induces
an epithelial-mesenchymal transition phenotype in human breast
cancer cells. Oncogene. 28:2940–2947. 2009. View Article : Google Scholar
|
|
33.
|
Bolós V, Peinado H, Pérez-Moreno MA, Fraga
MF, Esteller M and Cano A: The transcription factor Slug represses
E-cadherin expression and induces epithelial to mesenchymal
transitions: a comparison with Snail and E47 repressors. J Cell
Sci. 116:499–511. 2003.PubMed/NCBI
|