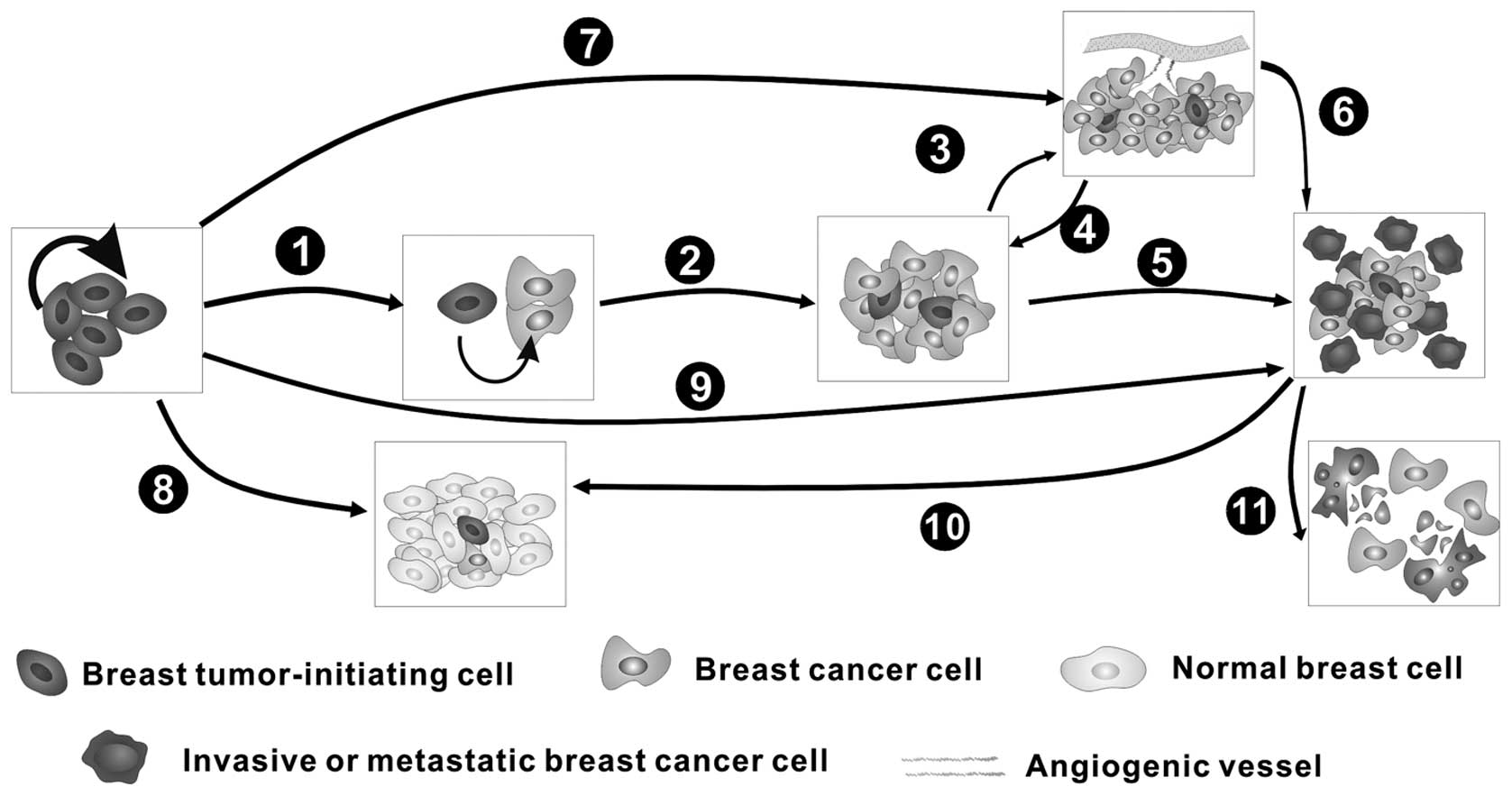
1. Introduction
Breast cancer is one of the most common malignant
cancers in women, and it has become a worldwide public health
issue, with reports of more than 500,000 deaths per year (1). The pathological progression of breast
cancer is multistep and complicated, consisting of oncogenesis,
which is primarily due to the self-renewal and differentiation of
breast tumor-initiating cells (BT-ICs), tumor growth (proliferation
and apoptosis), invasion, metastasis, angiogenesis and possible
post-treatment relapse (Fig. 1).
As breast tissue is steroid responsive, extensive effort has been
directed at identifying steroids and their receptors as well as the
nuclear receptor coactivators (2).
Endocrine therapy using the estrogen receptor antagonist tamoxifen
and aromatase inhibitors for the treatment of breast cancer has
been clinically applied. However, many problems, such as drug
resistance and the adverse effects of these drugs, still exist.
Therefore, novel and effective therapeutic methods for breast
cancer are urgently needed.
MicroRNAs (miRNAs) are non-coding single-stranded
RNAs with a length of approximately 22 nucleotides, and they
function as key post-transcriptional regulators of eukaryotic gene
expression through inhibiting translation or targeting mRNAs for
degradation (3). Recent studies
have uncovered the important roles of miRNAs in diverse
physiological and pathological events such as cell proliferation,
differentiation, embryo development and cancer progression
(4). Accumulating evidence
indicates that the putative functions of miRNAs might have
important clinical significance. For example, they might be
regarded as tumor suppressors and/or promoters (5,6), and
their abnormal expression is highly correlated with the progression
and pathology of breast cancer (7,8),
supporting their diagnostic, prognostic and therapeutic potentials
in breast cancer (4).
2. miRNAs in the pathogenesis and
progression of breast cancer
miRNAs and breast tumor-initiating
cells
Studies have shown that breast tumor-initiating
cells (BT-ICs) are the key contributors to the oncogenesis of
breast cancer; thus, understanding the regulation of BT-ICs is of
great importance. As BT-ICs have gained mutations causing them to
be more tumorigenic, they possess stem-like properties, such as
self-renewal, unlimited replication potential, differentiation and
metastasis.
At present, approximately 40 miRNAs have been found
to be differentially expressed between human BT-ICs and
non-tumorigenic breast cancer cells. Some of them, such as Let-7
(9), miR-16 (10), miR-34c (11), miR-200c (12), miR-183 and miR-203 (13), have been shown to promote the
self-renewal of BT-ICs by targeting different signaling pathways.
Let-7 miRNAs are members of the heterochronic pathway, and they are
necessary for BT-ICs to undergo the correct progression of
stage-specific events at the correct time, including
differentiation (9). In contrast,
other miRNAs such as miR-30 have been shown to inhibit BT-IC
self-renewal by reducing Ubc9 and inducing apoptosis through
silencing ITGB3 (14).
Additionally, three genomic clusters, miR-183-96-182,
miR-200b-200a-429 and miR-200c-141, are downregulated in BT-ICs,
suggesting that they may play key roles in the regulation of
self-renewal (12). Moreover,
miR-145 has been reported to inhibit the differentiation of BT-ICs
via the 3′-UTR of insulin receptor substrate-1 (IRS-1) (15).
Furthermore, miRNAs might also participate in the
metastasis of breast cancers mediated by BT-ICs. The miR-200 family
has been shown to play pivotal roles in the process of the
epithelial-mesenchymal transition (EMT), which is the initial event
of BT-IC-associated metastasis. Two pathways, the p53-miR-200c
pathway (16) and the
ZEB1-miR-200c-BMI1 pathway (17),
regulate both EMT and BT-IC-associated metastasis. Furthermore, the
downregulation of miR-34c (11)
and the overexpression of the miR-106b-25 cluster (18) are also sufficient to induce EMT.
Recently, miR-30a (19) and
miR-30c (20), targeting the
cytoskeleton network genes encoding twinfilin 1 (TWF1) and vimentin
(VIM), have also been shown to be involved in EMT.
miRNAs and breast cancer oncogenesis
Studies have revealed that miRNAs associated with
genomic changes can act as either breast cancer oncogenes
(oncomiRs) or suppressors (5). The
oncomiRs are upregulated in breast cancer and may hinder the
expression of many genes related to tumor suppression, cell cycle
regulation, apoptosis and differentiation. For example, the
overexpression of miR-21 in breast cancers influences several
targets, including the gene expression of tumor suppressor
tropomyosin 1 (TPM1) (21),
programmed cell death 4 (PDCD4) (22), phosphatase and tensin homolog
(PTEN) (23) and TIMP
metallopeptidase inhibitor 3 (TIMP3) (24), in breast cancer. Additionally,
miR-27a, miR-96 and miR-182 can target the 3′-UTR of the mRNA that
encodes the putative tumor suppressor transcription factor FOXO1
(25). Another miRNA, miR-155, has
been reported to act as an oncomiR, targeting caspase-3 as a potent
suppressor of apoptosis (26).
Compared to the oncomiRs, cancer-suppressing miRNAs
have been reported to target and inhibit oncogenes, and their
dysfunction may lead to the development of cancerous cells
(27). Both in vitro (from
MCF-7 cells) and in vivo (from analysis of cyclin
D1-transgenic mice) analyses have shown that the miR17-5p
(miR-91)/miR-20a cluster can inhibit breast cancer cell
proliferation by suppressing the proliferative effect of cyclin D1
(28). miR-27b has been reported
to play a suppressive role in breast cancer cells and to
post-transcriptionally regulate cytochrome P4501B1 (CYP1B1), a key
enzyme in the metabolism of 17β-estradiol, which promotes the
growth of breast cancer (29).
Recently, HER2 and HER3 (erbB2 and erbB3), which significantly
correlate with breast cancer and poor prognosis, have been shown to
be suppressed by miR-125a, miR-125b and miR-205 in breast cancer
cells, leading to a reduction in cell proliferation and migration
and increased apoptosis (30).
Moreover, the suppressive role of miR-205 is mediated through the
direct targeting of oncogenes such as Zeb1 (31). It has been shown that the
overexpression of miR-145 inhibits estrogen receptor-α (ER-α)
protein expression (32) and
reduces RTKN protein expression (33), thus reducing breast cancer cell
growth and inducing apoptosis, indicating its tumor-suppressive
functions. Similarly, miR-22 (34), miR-206 (35) and miR-375 (36) negatively affect the expression of
ER-α.
miRNAs and breast cancer growth
Studies have recently uncovered the roles of miRNAs
in cancer cell proliferation and apoptosis, the dysregulation of
which is the most important mechanism of cancer growth. For
example, oncomiRs, including miR-21 (24), miR-27a, miR-96, miR-182 (25) and miR-155 (37), can target the 3′-UTR of mRNAs that
encode tumor suppressor genes. Moreover, cell growth suppression is
achieved by the overexpression of cancer suppressor miRNAs such as
miR17-5p (miR-91)/miR-20a (28),
miR-27b (29), miR-125a, miR-125b
and miR-205 (30), suggesting
their roles in growth suppression. Furthermore, the upregulation of
ER-α accounts for the abnormal cell proliferation observed in
approximately two-thirds of breast cancer cases. miR-22 (34), miR-145 (32), miR-206 (35) and miR-375 (36) are reported to negatively regulate
ER-α expression, thus inhibiting breast cancer cell
proliferation.
Several studies have addressed the roles that miRNAs
play in the modulation of apoptosis in breast cancer cells. The
overexpression of miR-145 has been demonstrated to induce apoptosis
by directly binding to the 3′-UTR of RTKN (33). miR-26b can also impair the
viability and trigger the apoptosis of human MCF-7 cells by
directly targeting SLC7A11 (38).
Moreover, miR-290 was shown to target Arid4b while simultaneously
enhancing ER signaling and inducing apoptosis, thereby suppressing
breast cancer growth (46). The
overexpression of miR-335 decreases cell viability and increases
apoptosis by simultaneously regulating the known BRCA1 activators
ER-α, IGF1R and SP1 and the repressor ID4 (39).
miRNAs and breast cancer invasion and
metastasis
Invasion and metastasis are the major factors
underlying the poor prognosis or even death of breast cancer
patients. In the past few years, many types of miRNAs have been
identified, and their roles have been explored in cancer invasion
and metastasis; some of these miRNAs function as metastasis
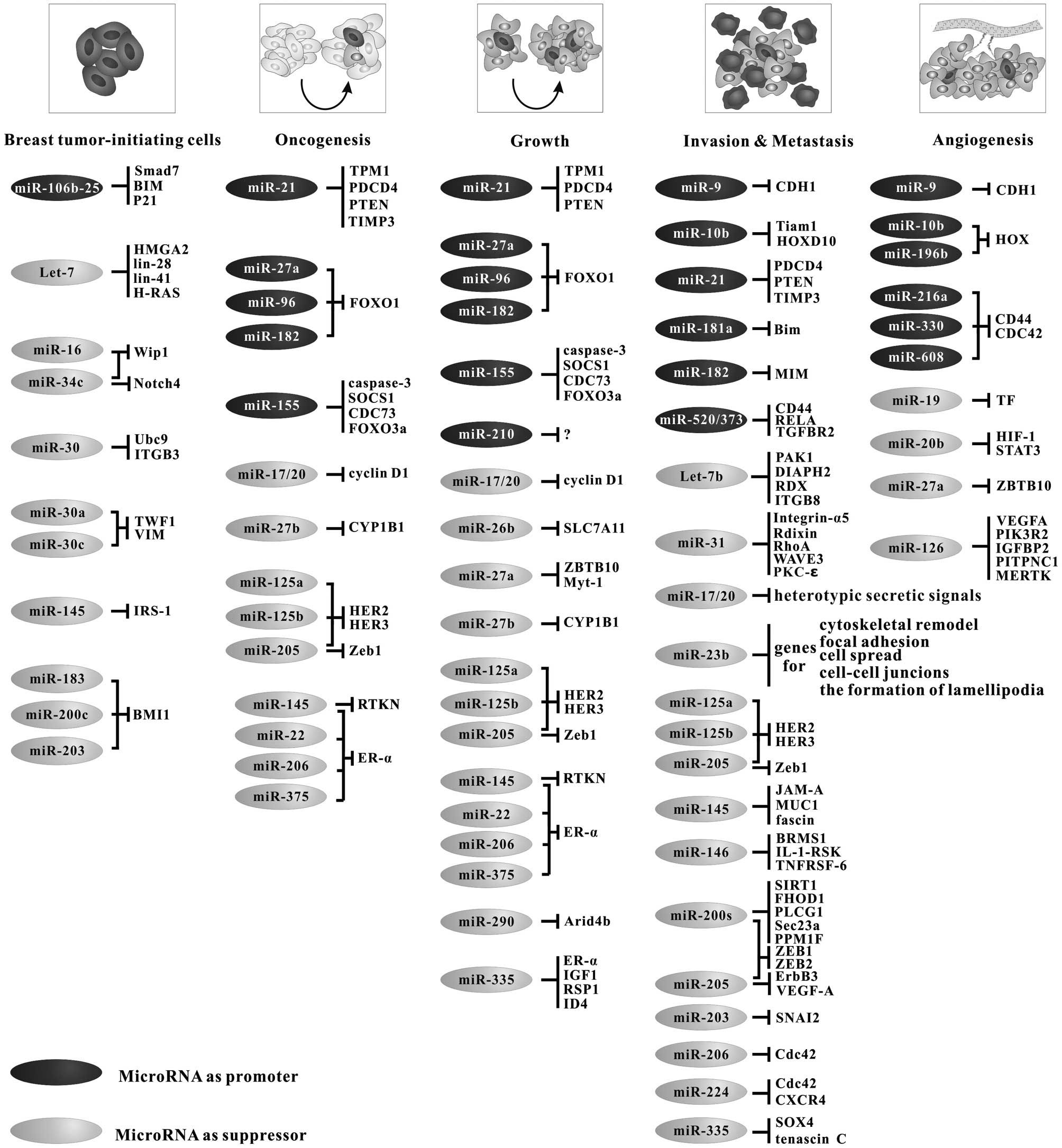
promoters, whereas others function as suppressors (Fig. 2).
miRNAs as suppressors of metastasis
miR-23b has been shown to be a metastatic suppressor
miRNA that directly inhibits a number of genes implicated in
cytoskeletal remodeling, focal adhesion, cell spreading, cell-cell
junction formation and the formation of lamellipodia in breast
cancer cells (40). miR-31 has
also been shown to trigger metastatic regression in the lungs by
eliciting cell cycle arrest and apoptosis, which can be explained
by the miR-31-mediated suppression of integrin-α5, radixin, RhoA
(41), the actin cytoskeleton
remodeling protein WAVE3 (42) and
protein kinase C epsilon (PKC-ɛ) (43). Other cancer-suppressing miRNAs such
as miR-125a, miR-125b and miR-205 significantly inhibit breast
cancer cell invasion by directly targeting HER2 and HER3 (30), whereas miR-205 can target Zeb1
(31). Additionally, miR-145
significantly suppresses cell invasion and lung metastasis by
directly targeting the metastasis gene mucin 1 (MUC1), leading to a
reduction of β-catenin and adherin 11 (44). Moreover, miR-146 may be involved in
the inhibition of the breast cancer metastasis suppressor 1
(BRMS1), a predominant nuclear protein, leading to metastasis
suppression (45); miR-146a/b
functions to negatively regulate NF-κB activity through targeting
IL-1 receptor-associated kinase (IL-1-RSK) and TNF
receptor-associated factor 6 (TNFRSF-6), thus functioning as a
suppressor of breast cancer metastasis (46).
The initiation of metastasis, EMT, has been shown to
be significantly regulated by the miR-200 family (miR-200a,
miR-200b, miR-200c, miR-141 and miR-429), the members of which are
all dramatically downregulated in breast cancer cells that have
undergone the EMT induced by tumor growth factor-β (TGF-β), and the
E-cadherin transcriptional repressors ZEB1 and ZEB2 have been
established as their targets (47). In addition, the miR-200 family also
enhances metastatic colonization, partially through directly
targeting Sec23a, which is involved in the secretion of metastasis
suppressor proteins (48).
Furthermore, the miR-200b/c/429 cluster is a stronger regulator of
EGF-driven invasion than the miR-200a/141 cluster, and
phospholipase C gamma 1 (PLCG1) might be a potential candidate
contributing to these differences (49). Moreover, the overexpression of
miR-200a has been reported to decrease anchorage-independent growth
and migration in breast cancer, targeting the oncogene silent
information regulator 1 (SIRT1) (50); miR-200c greatly impacts the
regulation of several EMT-related processes in breast cancer by
directly targeting the actin cytoskeleton (51).
Studies have reported that in highly metastatic
breast cancer cells, the expression of miR-203 is significantly
downregulated and that its upregulation inhibits tumor cell
invasion and metastatic colonization through the SNAI2 and miR-203
regulatory loop (52). Moreover,
miR-335 has been identified as the first selective metastasis
suppressor in human breast cancer by targeting SOX4 and tenascin-C
(53). Additionally, miR-206
suppresses the invasion and migration of MDA-MB-231 cells in
vitro, partially via regulating the remodeling of the actin
cytoskeleton, with CDC42 as its potential target (54). Another study has shown that the
overexpression of miR-224 inhibits cell division cycle 42 (CDC42)
and chemokine receptor 4 (CXCR4), accounting for the inhibition of
Ubc9-mediated invasion (55).
Furthermore, let-7b and the miR-17-5p/miR-20a cluster are
significantly decreased in lymph node metastases of breast cancer
cells. The forced expression of let-7b significantly inhibits
breast cancer cell migration by targeting four genes that are
correlated with the actin cytoskeleton pathway, including PAK1,
DIAPH2, RDX and ITGB8 (56). The
miR-17-5p/miR-20a cluster plays a suppressive role in metastatic
breast cancer through heterotypic secreted signaling (57).
miRNAs as metastasis promoters
miR-9 is upregulated in breast cancer and increases
cell motility and invasiveness by directly targeting CDH1 and
E-cadherin-encoding mRNAs (58).
miR-10b expression is increased in metastatic breast cancer cells,
leading to cell migration, invasion and metastasis through
indirectly activating the prometastatic gene RHOC by suppressing
the homeobox D10 (HOXD10) tumor suppressor signaling pathway
(59). The regulation of T
lymphoma invasion and metastasis 1 (Tiam1)-mediated Rac activation
in breast cancer cells is a novel target for miR-10b to regulate
the invasion and migration of breast cancer cells (60). miR-21, as an oncogenic miRNA, plays
important roles in breast cancer invasion and metastasis by
inhibiting multiple metastasis suppressor genes. The upregulated
expression of miR-181a in metastatic breast cancer cells has been
shown to enhance breast cancer metastasis by promoting EMT and
migratory and invasive phenotypes, targeting the proapoptotic
molecule Bim, which is involved in metastatic cancer cell anoikis
(61). Similarly, miR-182 promotes
breast cancer metastasis by suppressing the missing in metastasis
(MIM), which activates the Ras homolog family member A (RhoA)
(62). miR-373 and miR-520c can
also promote the invasion and migration of breast cancer cells both
in vitro and in vivo (63) by directly targeting and inhibiting
the expression of RELA, TGFBR2 (63) and CD44 (64).
miRNAs and breast cancer
angiogenesis
Angiogenesis is one of the most important mechanisms
of tumor growth and metastasis (Fig.
1). Much effort has been directed toward the development of
anti-angiogenic tumor therapies. miRNAs appear to have considerable
potential as the gene resource for the gene therapy of breast
cancer. Endothelial cells (ECs) are one of the key components of
the tunica intima, playing vital roles in tumor angiogenesis. It
has been shown that many miRNAs such as miR-216a, miR-330 and
miR-608, which can bind to both the CD44 and CDC42 (CD44 downstream
target mRNA) 3′-UTRs, can modulate EC activities (3). miR-126, which is particularly
expressed in vascular endothelial cells, is reported to play a
vital role in suppressing breast cancer angiogenesis through
regulating the VEGF/PI3K/AKT signaling cascade (65) and targeting IGFBP2, PITPNC1 and
MERTK (66).
Moreover, miR-9 contributes to angiogenesis through
the upregulation of VEGF expression in breast cancer by directly
targeting CDH1 (58). Another
report has demonstrated that VEGF expression in breast cancer cells
is mediated by hypoxia inducible factor 1 (HIF-1) and signal
transducer and activator of transcription 3 (STAT3) in a
miR-20b-dependent manner (67).
The overexpression of miR-10b and miR-196b has been detected in
high-grade breast cancer vasculature and, notably, to be responsive
to VEGF stimulation (68).
Moreover, miR-27a suppresses breast cancer angiogenesis by
inhibiting specificity protein (Sp)-dependent angiogenic gene
expression, including that of survivin, VEGF and VEGFR1, as it
targets the zinc finger ZBTB10 gene (69). The overexpression of miR-19
downregulates tissue factor (TF) expression, which is an important
factor in the regulation of tumor angiogenesis, suggesting the
potential of miR-19 for regulating breast cancer angiogenesis
(70).
Taken together, the involvement of these miRNAs in
the pathogenesis and progression of breast cancer has been
elucidated, with some of them acting as suppressors and others as
tumor promoters, as summarized in Fig.
2. These studies are crucial to evaluating their use as
clinical biomarkers and their therapeutic potential.
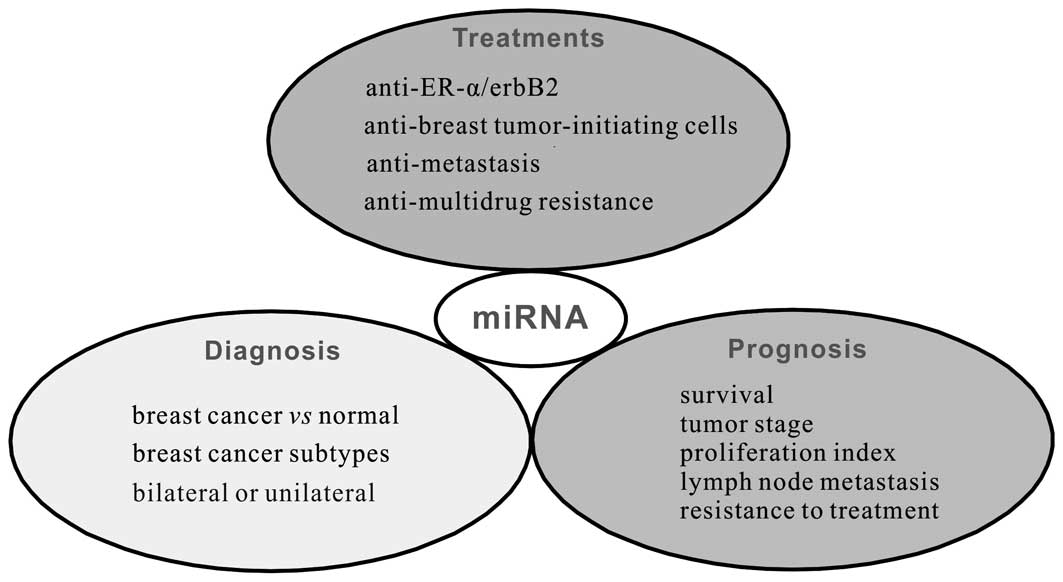
3. Clinical implications of miRNAs in breast
cancer
Many miRNAs have been shown to contribute to the
pathogenesis and progression of breast cancer, which supports the
potential of miRNA-specific strategies for the treatment of breast
cancer (Fig. 3).
Diagnostic and prognostic biomarkers of
breast cancer
The increasing number of reports indicating the
significance of miRNAs in the pathogenesis and progression of
breast cancer has revealed the potential relevance of aberrantly
expressed miRNAs as biomarkers for the detection, diagnosis,
classification and therapy of breast cancer. First, the expression
profile of miRNAs can be used to distinguish breast cancer tissue
from normal tissue. For example, the significant overexpression of
miR-21, miR-106a and miR-155 and the decreased expression of
miR-126, miR-199a and miR-335 have been reported in breast cancer
tissue when compared with normal tissue (71). A study by Iyevleva et al has
reported higher levels of miR-21, miR-10b and miR-31 in bilateral
patients than in unilateral patients (72). Additionally, a number of miRNAs are
differentially expressed in the luminal A, luminal B, basal-like,
HER2+ and normal-like breast cancer subtypes. For
example, the upregulation of miR-17-92 clusters has been shown to
account for the great distinction of triple-negative breast cancer
from other subtypes (6).
The miRNA expression profile has also been
correlated with pathological features such as tumor stage,
proliferation index, lymph node metastasis, resistance to breast
cancer treatment and poor survival, suggesting their prognostic
potential. The most promising prognostic miRNAs for breast cancer
are miR-21 (24), miR-181a
(61) and the miR-221/miR-222
cluster (73) because their
overexpression has been correlated with advanced tumor stage, lymph
node metastasis and poor patient survival. Moreover, a high level
of the miR-106b-25 cluster in breast cancer can significantly
predict a shortened time to relapse (18). Additionally, in ER-positive cases,
high miR-767-3p, miR-128a and/or miR-769-3p expression is
associated with a poor prognosis, whereas high miR-135a expression
is associated with a good prognosis; in contrast, in ER-negative
cases, high levels of miR-27b, miR-144 and/or miR-210 are
associated with a poor prognosis, and high miR-342, miR-150 and/or
miR-30c expression is associated with a good prognosis (74). Moreover, miR-342, miR-27b and
miR-150 are prognostic in triple receptor-negative breast cancers
(74).
Therapeutic miRNAs
The traditional therapies for breast cancer, such as
surgery, chemotherapy and/or radiotherapy, appear to be powerful;
however, regardless of the level of damage directly caused to the
human body, problems such as drug resistance and adverse effects
still exist, making the treatment of breast cancer more difficult
than expected. Therefore, novel, feasible and effective therapeutic
methods for breast cancer are urgently needed. Because of the
distinct roles that miRNAs play in the pathology of breast cancer,
many studies have investigated their therapeutic potential in its
treatment.
Anti-BT-ICs
One of the greatest challenges for breast cancer
research is relapses that occur in patients undergoing chemotherapy
and radiotherapy. Recent studies have shown that targeting BT-ICs
is promising to prevent relapse and provides a new strategy for
breast cancer prevention. Many types of miRNAs are involved in
regulating the self-renewal and differentiation of BT-IC as well as
BT-IC-induced relapse (Fig. 2). As
mentioned above, one attractive candidate for this purpose is let-7
because the administration of let-7 has been found to be effective
in mouse models of breast cancer (75).
Anti-ER-α/HER
As breast cancer is steroid-responsive, much
attention has been given to steroids (e.g., estradiol and
progesterone) and their receptors [e.g., ER-α, ER-β and the
progesterone receptor (PR)] as well as the nuclear receptor
coactivators, particularly steroid receptor coactivator-1 (SRC-1)
and SRC-3 (2). It has been shown
that ER-α upregulation is responsible for the abnormal cell
proliferation observed in approximately two-thirds of breast cancer
cases, and anti-ER-α therapy may be of great value for breast
cancer treatments. Studies have revealed that the levels of miR-22
(34), miR-145 (32), miR-206 (35) and miR-375 (36) are significantly decreased in
ER-α-positive human breast cancer tissues and inversely correlated
with ER-α mRNA expression in breast cancer tissues; thus, they have
been suggested as novel therapeutic agents for anti-multidrug
resistance (MDR) therapies that target only ER-α in breast cancer.
Additionally, the overexpression of HER2 and HER3 (erbB2 and erbB3)
is significantly correlated with breast cancer grade and poor
prognosis, and this overexpression can be suppressed by miR-125a,
miR-125b and miR-205 (30), thus
resulting in the abrogation of HER2- and HER3-mediated resistance
and restoring potent proapoptotic activity.
Anti-metastasis
miR-21 plays a role in invasion and metastasis by
targeting multiple tumor/metastasis suppressor genes such as PDCD4
(22), PTEN (23) and TIMP3 (24), suggesting its potential for
anti-metastasis treatments. miR-31 has also been shown to regulate
a number of metastasis-related genes in breast cancer cells and
tissues, and its expression is inversely correlated with the cell’s
ability to invade and metastasize. Activation of miR-31 in
established metastases elicits metastatic regression and prolongs
patient survival (41). The
overexpression of miR-145 has been shown to suppress breast cancer
cell growth and motility, and a treatment combining Ad-miR-145 with
5-FU has shown significant antitumor effects (76). miR-146 is also involved in
suppressing metastasis, and miR-146a/b-expressing cells show
markedly impaired invasion and migration capacities when compared
with controls (45).
Anti-multidrug resistance (MDR)
Drug resistance and multidrug resistance (MDR) are
the main causes of treatment failure, and these are the most
challenging problems in the treatment of breast cancer. Studies
have demonstrated that some miRNAs might be involved in the
occurrence of MDR, and therefore, targeting these miRNAs might be
of great significance regarding the development of novel strategies
against breast cancer. For example, the expression of miR-155
(77) and miR-663 (78) has been demonstrated to induce
breast cancer cell survival and MDR, whereas their knockdown
renders breast cancer cells susceptible to apoptosis and enhances
their chemosensitivity. miR-326 is downregulated in advanced breast
cancer tissues and is inversely correlated with the expression of
multidrug resistance-associated protein-1 (MRP-1); its upregulation
in VP-16-resistant MCF-7 cell lines can downregulate the expression
of MRP-1, making them sensitive to VP-16 and doxorubicin (DOX)
(79). These findings indicate
that miR-326 may be an efficient and powerful agent for preventing
and reversing MDR in breast cancer cells. Moreover, the
miR-328-mediated downregulation of ABCG2 (a molecular determinant
of the pharmacokinetic properties of many drugs in humans) in
MCF-7/MX100 cells may result in increased mitoxantrone sensitivity
(80). Recently, the multidrug
resistance 1 gene has been identified as a novel target of miR-451,
and transfection of MCF-7/DOX-resistant cells with miR-451 results
in increased sensitivity to DOX, indicating that increased miR-451
expression may have significant implications for overcoming the MDR
of breast cancer cells (81).
The above results are summarized in Fig. 3 and Table I.
 | Table ISelected miRNAs and their clinical
applications in breast cancer. |
Table I
Selected miRNAs and their clinical
applications in breast cancer.
| Application | microRNA | Putative clinical
significance | Refs. |
|---|
| Diagnosis | miR-21↑, miR-106a↑,
miR-155↑, miR-199a↓, miR-126↓, miR-335↓ | Potential
biomarkers for breast cancer diagnosis, grading and prognosis | (70) |
| miR-21↑, miR-10b↑,
miR-31↑ | Biomarkers for the
diagnosis bilateral or unilateral breast cancer | (71) |
| miR-17-92
cluster↑ | Identifies the
subtype of breast cancer: distinction of triple- negative breast
cancer from other subtypes | (72) |
| Biomarker |
| Prognosis | miR-21↑, miR-181a↑,
miR-221/miR 222 cluster↑ | The overexpression
is correlated with advanced tumor stage, lymph node metastasis and
poor survival of the patients | (6,23,60) |
| miR-767-3p,
miR-128a, miR-769-3p, miR-135a | Associated with
prognosis in ER-positive cases: poor, miR-767-3p↑, miR-128a↑,
miR-769-3p↑; good, miR-135a↑ | (73) |
| miR-27b, miR-144,
miR-210
miR-342, miR-150, miR-30c | Associated with
prognosis in ER-negative cases: poor, miR-27b↑, miR-144↑, miR-210↑;
good, miR-342↑, miR-150↑, miR-30c↑ | (73) |
| miR-27b, miR-150,
miR-342 | Prognostic also in
triple receptor-negative breast cancers | (73) |
| miR-106b-25
cluster↑ | Significantly
predicted shortened time to relapse | (17) |
| miR-22↓, miR-145↓,
miR-206↓ and miR-375↓ | Significantly
decreased in ER-α-positive human breast cancer tissues and is
inversely correlated with ER-α mRNA expression in breast cancer
tissues in a dose-and time-dependent manner | (31,33–35) |
| Anti-ER-α/HER | miR-125a↓,
miR-125b↓
miR-205↓ | Directly suppressed
the HER2 and HER3 (erbB2 and erbB3) | (29) |
| Anti-BT-IC | let-7↑ | Effective in
suppressing metastatic breast cancer | (74) |
| Therapy |
|
Anti-metastasis | miR-21↓ | Significantly
reduced invasion and lung metastasis ability | (21–23) |
| miR-31↑ | Elicits the
regression of metastasis and prolongs the survival of patients | (40) |
| miR-145↑ | Ad-miR-145
suppressed breast cancer cell motility and invasiveness; combining
Ad-miR-145 with 5-FU significantly showed anti-tumor effects | (2) |
| miR-146a/b↑ | Inhibited both
invasion and migration, and suppressed experimental lung
metastasis | (44) |
| Anti-MDR | miR-21↑ | miR-21 inhibitor
gene therapy combining with taxol chemotherapy | (81) |
| miR-24-2↓ | Combining with an
anticancer drug such as cisplatin provides a new avenue for
overcoming drug resistance | (86) |
| miR-155↑ | Knockdown of it
renders breast cancer cells to apoptosis and enhance
chemosensitivity | (76) |
| miR-326↑ | Downregulated MRP-1
expression and sensitize cancer cells to VP-16 and doxorubicin | (78) |
| miR-328↑ | Increased
mitoxantrone sensitivity | (79) |
| miR-451↑ | Increased
sensitivity of cancer cells to DOX | (80) |
4. Conclusions
In the past few years, the correlations between
miRNAs and breast cancer have been widely investigated,
contributing greatly to studies of breast cancer pathogenesis and
their clinical implications. These small molecules play significant
roles in the oncogenesis, growth, invasion, metastasis and
angiogenesis of breast cancer; thus, altered expression of miRNAs
can be regarded as a target for the diagnosis and/or treatment of
breast cancer based on the miRNA expression profile. However, many
of the remaining problems urgently require further exploration. For
example, we know little about the upstream, intrinsic factors that
regulate these specific miRNAs; there are tens or hundreds of
miRNAs within a cell, and which is the most prominent indicator for
the diagnosis and treatment as well as the prognosis of cancer
remains to be elucidated. Future work should aim to identify the
limited miRNAs involved in each pathological stage as well as the
therapeutic targets of breast cancers. These under-explored issues
may be the greatest challenges in the future.
Acknowledgements
This study was supported by the National Science
Foundation of China (NSFC, No. 81171035).
References
|
1
|
Zoon CK, Starker EQ, Wilson AM,
Emmert-Buck MR, Libutti SK and Tangrea MA: Current molecular
diagnostics of breast cancer and the potential incorporation of
microRNA. Expert Rev Mol Diagn. 9:455–467. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Johnson AB and O’Malley BW: Steroid
receptor coactivators 1, 2, and 3: critical regulators of nuclear
receptor activity and steroid receptor modulator (SRM)-based cancer
therapy. Mol Cell Endocrinol. 348:430–439. 2012. View Article : Google Scholar
|
|
3
|
Jeyapalan Z, Deng Z, Shatseva T, Fang L,
He C and Yang BB: Expression of CD44 3′-untranslated region
regulates endogenous microRNA functions in tumorigenesis and
angiogenesis. Nucleic Acids Res. 39:3026–3041. 2011.
|
|
4
|
Piva R, Spandidos DA and Gambari R: From
microRNA functions to microRNA therapeutics: novel targets and
novel drugs in breast cancer research and treatment (Review). Int J
Oncol. 43:985–994. 2013.PubMed/NCBI
|
|
5
|
Iorio MV, Ferracin M, Liu CG, et al:
MicroRNA gene expression deregulation in human breast cancer.
Cancer Res. 65:7065–7070. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Farazi TA, Horlings HM, Ten Hoeve JJ, et
al: MicroRNA sequence and expression analysis in breast tumors by
deep sequencing. Cancer Res. 71:4443–4453. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Andorfer CA, Necela BM, Thompson EA and
Perez EA: MicroRNA signatures: clinical biomarkers for the
diagnosis and treatment of breast cancer. Trends Mol Med.
17:313–319. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Melo SA and Esteller M: Dysregulation of
microRNAs in cancer: playing with fire. FEBS Lett. 585:2087–2099.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nimmo RA and Slack FJ: An elegant miRror:
microRNAs in stem cells, developmental timing and cancer.
Chromosoma. 118:405–418. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang X, Wan G, Mlotshwa S, et al:
Oncogenic Wip1 phosphatase is inhibited by miR-16 in the DNA damage
signaling pathway. Cancer Res. 70:7176–7186. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yu F, Jiao Y, Zhu Y, et al: MicroRNA 34c
gene down-regulation via DNA methylation promotes self-renewal and
epithelial-mesenchymal transition in breast tumor-initiating cells.
J Biol Chem. 287:465–473. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shimono Y, Zabala M, Cho RW, et al:
Downregulation of miRNA-200c links breast cancer stem cells with
normal stem cells. Cell. 138:592–603. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Greene SB, Herschkowitz JI and Rosen JM:
Small players with big roles: microRNAs as targets to inhibit
breast cancer progression. Curr Drug Targets. 11:1059–1073. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yu F, Deng H, Yao H, Liu Q, Su F and Song
E: Mir-30 reduction maintains self-renewal and inhibits apoptosis
in breast tumor-initiating cells. Oncogene. 29:4194–4204. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Rubin R, Arzumanyan A, Soliera AR, Ross B,
Peruzzi F and Prisco M: Insulin receptor substrate (IRS)-1
regulates murine embryonic stem (mES) cells self-renewal. J Cell
Physiol. 213:445–453. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chang CJ, Chao CH, Xia W, et al: p53
regulates epithelial-mesenchymal transition and stem cell
properties through modulating miRNAs. Nat Cell Biol. 13:317–323.
2011. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ahmad A, Aboukameel A, Kong D, et al:
Phosphoglucose isomerase/autocrine motility factor mediates
epithelial-mesenchymal transition regulated by miR-200 in breast
cancer cells. Cancer Res. 71:3400–3409. 2011. View Article : Google Scholar
|
|
18
|
Smith AL, Iwanaga R, Drasin DJ, et al: The
miR-106b-25 cluster targets Smad7, activates TGF-beta signaling,
and induces EMT and tumor initiating cell characteristics
downstream of Six1 in human breast cancer. Oncogene. 31:5162–5171.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cheng CW, Wang HW, Chang CW, et al:
MicroRNA-30a inhibits cell migration and invasion by downregulating
vimentin expression and is a potential prognostic marker in breast
cancer. Breast Cancer Res Treat. 134:1081–1093. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bockhorn J, Yee K, Chang YF, et al:
MicroRNA-30c targets cytoskeleton genes involved in breast cancer
cell invasion. Breast Cancer Res Treat. 137:373–382. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhu S, Si ML, Wu H and Mo YY: MicroRNA-21
targets the tumor suppressor gene tropomyosin 1 (TPM1). J Biol
Chem. 282:14328–14336. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Frankel LB, Christoffersen NR, Jacobsen A,
Lindow M, Krogh A and Lund AH: Programmed cell death 4 (PDCD4) is
an important functional target of the microRNA miR-21 in breast
cancer cells. J Biol Chem. 283:1026–1033. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Meng F, Henson R, Wehbe-Janek H, Ghoshal
K, Jacob ST and Patel T: MicroRNA-21 regulates expression of the
PTEN tumor suppressor gene in human hepatocellular cancer.
Gastroenterology. 133:647–658. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li J, Zhang Y, Zhang W, et al: Genetic
heterogeneity of breast cancer metastasis may be related to miR-21
regulation of TIMP-3 in translation. Int J Surg Oncol.
2013:8750782013.PubMed/NCBI
|
|
25
|
Guttilla IK and White BA: Coordinate
regulation of FOXO1 by miR-27a, miR-96, and miR-182 in breast
cancer cells. J Biol Chem. 284:23204–23216. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ovcharenko D, Kelnar K, Johnson C, Leng N
and Brown D: Genome-scale microRNA and small interfering RNA
screens identify small RNA modulators of TRAIL-induced apoptosis
pathway. Cancer Res. 67:10782–10788. 2007. View Article : Google Scholar
|
|
27
|
Zhang B, Pan X, Cobb GP and Anderson TA:
microRNAs as oncogenes and tumor suppressors. Dev Biol. 302:1–12.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yu Z, Wang C, Wang M, et al: A cyclin
D1/microRNA 17/20 regulatory feedback loop in control of breast
cancer cell proliferation. J Cell Biol. 182:509–517. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tsuchiya Y, Nakajima M, Takagi S, Taniya T
and Yokoi T: MicroRNA regulates the expression of human cytochrome
P450 1B1. Cancer Res. 66:9090–9098. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang S, Huang J, Lyu H, et al: Functional
cooperation of miR-125a, miR-125b, and miR-205 in
entinostat-induced downregulation of erbB2/erbB3 and apoptosis in
breast cancer cells. Cell Death Dis. 4:e5562013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wu H and Mo YY: Targeting miR-205 in
breast cancer. Expert Opin Ther Targets. 13:1439–1448. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Spizzo R, Nicoloso MS, Lupini L, et al:
miR-145 participates with TP53 in a death-promoting regulatory loop
and targets estrogen receptor-alpha in human breast cancer cells.
Cell Death Differ. 17:246–254. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang S, Bian C, Yang Z, et al: miR-145
inhibits breast cancer cell growth through RTKN. Int J Oncol.
34:1461–1466. 2009.PubMed/NCBI
|
|
34
|
Xiong J, Yu D, Wei N, et al: An estrogen
receptor alpha suppressor, microRNA-22, is downregulated in
estrogen receptor alpha-positive human breast cancer cell lines and
clinical samples. FEBS J. 277:1684–1694. 2010. View Article : Google Scholar
|
|
35
|
Adams BD, Furneaux H and White BA: The
micro-ribonucleic acid (miRNA) miR-206 targets the human estrogen
receptor-alpha (ERalpha) and represses ERalpha messenger RNA and
protein expression in breast cancer cell lines. Mol Endocrinol.
21:1132–1147. 2007. View Article : Google Scholar
|
|
36
|
de Souza Rocha Simonini P, Breiling A,
Gupta N, et al: Epigenetically deregulated microRNA-375 is involved
in a positive feedback loop with estrogen receptor alpha in breast
cancer cells. Cancer Res. 70:9175–9184. 2010.
|
|
37
|
Rather MI, Nagashri MN, Swamy SS, Gopinath
KS and Kumar A: Oncogenic microRNA-155 down-regulates tumor
suppressor CDC73 and promotes oral squamous cell carcinoma cell
proliferation: implications for cancer therapeutics. J Biol Chem.
288:608–618. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Liu XX, Li XJ, Zhang B, et al:
MicroRNA-26b is underexpressed in human breast cancer and induces
cell apoptosis by targeting SLC7A11. FEBS Lett. 585:1363–1367.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Heyn H, Engelmann M, Schreek S, et al:
MicroRNA miR-335 is crucial for the BRCA1 regulatory cascade in
breast cancer development. Int J Cancer. 129:2797–2806. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Pellegrino L, Stebbing J, Braga VM, et al:
miR-23b regulates cytoskeletal remodeling, motility and metastasis
by directly targeting multiple transcripts. Nucleic Acids Res.
41:5400–5412. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Valastyan S, Chang A, Benaich N, Reinhardt
F and Weinberg RA: Activation of miR-31 function in
already-established metastases elicits metastatic regression. Genes
Dev. 25:646–659. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Sossey-Alaoui K, Downs-Kelly E, Das M,
Izem L, Tubbs R and Plow EF: WAVE3, an actin remodeling protein, is
regulated by the metastasis suppressor microRNA, miR-31, during the
invasion-metastasis cascade. Int J Cancer. 129:1331–1343. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Korner C, Keklikoglou I, Bender C, Worner
A, Munstermann E and Wiemann S: MicroRNA-31 sensitizes human breast
cells to apoptosis by direct targeting of protein kinase C epsilon
(PKCepsilon). J Biol Chem. 288:8750–8761. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Sachdeva M and Mo YY: MicroRNA-145
suppresses cell invasion and metastasis by directly targeting mucin
1. Cancer Res. 70:378–387. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Hurst DR, Edmonds MD, Scott GK, Benz CC,
Vaidya KS and Welch DR: Breast cancer metastasis suppressor 1
up-regulates miR-146, which suppresses breast cancer metastasis.
Cancer Res. 69:1279–1283. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Bhaumik D, Scott GK, Schokrpur S, Patil
CK, Campisi J and Benz CC: Expression of microRNA-146 suppresses
NF-kappaB activity with reduction of metastatic potential in breast
cancer cells. Oncogene. 27:5643–5647. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Gregory PA, Bert AG, Paterson EL, et al:
The miR-200 family and miR-205 regulate epithelial to mesenchymal
transition by targeting ZEB1 and SIP1. Nat Cell Biol. 10:593–601.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Korpal M, Ell BJ, Buffa FM, et al: Direct
targeting of Sec23a by miR-200s influences cancer cell secretome
and promotes metastatic colonization. Nat Med. 17:1101–1108. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Uhlmann S, Zhang JD, Schwäger A, et al:
miR-200bc/429 cluster targets PLCgamma1 and differentially
regulates proliferation and EGF-driven invasion than miR-200a/141
in breast cancer. Oncogene. 29:4297–4306. 2010. View Article : Google Scholar
|
|
50
|
Eades G, Yao Y, Yang M, Zhang Y, Chumsri S
and Zhou Q: miR-200a regulates SIRT1 expression and epithelial to
mesenchymal transition (EMT)-like transformation in mammary
epithelial cells. J Biol Chem. 286:25992–26002. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Jurmeister S, Baumann M, Balwierz A, et
al: MicroRNA-200c represses migration and invasion of breast cancer
cells by targeting actin-regulatory proteins FHOD1 and PPM1F. Mol
Cell Biol. 32:633–651. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Ding X, Park SI, McCauley LK and Wang CY:
Signaling between transforming growth factor beta (TGF-beta) and
transcription factor SNAI2 represses expression of microRNA miR-203
to promote epithelial-mesenchymal transition and tumor metastasis.
J Biol Chem. 288:10241–10253. 2013. View Article : Google Scholar
|
|
53
|
Tavazoie SF, Alarcón C, Oskarsson T, et
al: Endogenous human microRNAs that suppress breast cancer
metastasis. Nature. 451:147–152. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Liu H, Cao YD, Ye WX and Sun YY: Effect of
microRNA-206 on cytoskeleton remodelling by downregulating Cdc42 in
MDA-MB-231 cells. Tumori. 96:751–755. 2010.PubMed/NCBI
|
|
55
|
Zhu S, Sachdeva M, Wu F, Lu Z and Mo YY:
Ubc9 promotes breast cell invasion and metastasis in a
sumoylation-independent manner. Oncogene. 29:1763–1772. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Hu X, Guo J, Zheng L, et al: The
heterochronic microRNA let-7 inhibits cell motility by regulating
the genes in the actin cytoskeleton pathway in breast cancer. Mol
Cancer Res. 11:240–250. 2013. View Article : Google Scholar
|
|
57
|
Yu Z, Willmarth NE, Zhou J, et al:
microRNA 17/20 inhibits cellular invasion and tumor metastasis in
breast cancer by heterotypic signaling. Proc Natl Acad Sci USA.
107:8231–8236. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Ma L, Young J, Prabhala H, et al: miR-9, a
MYC/MYCN-activated microRNA, regulates E-cadherin and cancer
metastasis. Nat Cell Biol. 12:247–256. 2010.PubMed/NCBI
|
|
59
|
Haque I, Banerjee S, Mehta S, et al:
Cysteine-rich 61-connective tissue growth
factor-nephroblastoma-overexpressed 5 (CCN5)/Wnt-1-induced
signaling protein-2 (WISP-2) regulates microRNA-10b via
hypoxia-inducible factor-1alpha-TWIST signaling networks in human
breast cancer cells. J Biol Chem. 286:43475–43485. 2011. View Article : Google Scholar
|
|
60
|
Moriarty CH, Pursell B and Mercurio AM:
miR-10b targets Tiam1: implications for Rac activation and
carcinoma migration. J Biol Chem. 285:20541–20546. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Taylor MA, Sossey-Alaoui K, Thompson CL,
Danielpour D and Schiemann WP: TGF-beta upregulates miR-181a
expression to promote breast cancer metastasis. J Clin Invest.
123:150–163. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Lei R, Tang J, Zhuang X, et al:
Suppression of MIM by microRNA-182 activates RhoA and promotes
breast cancer metastasis. Oncogene. 33:1287–1296. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Huang Q, Gumireddy K, Schrier M, et al:
The microRNAs miR-373 and miR-520c promote tumour invasion and
metastasis. Nat Cell Biol. 10:202–210. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Yu Z, Baserga R, Chen L, Wang C, Lisanti
MP and Pestell RG: microRNA, cell cycle, and human breast cancer.
Am J Pathol. 176:1058–1064. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Zhu N, Zhang D, Xie H, et al:
Endothelial-specific intron-derived miR-126 is down-regulated in
human breast cancer and targets both VEGFA and PIK3R2. Mol Cell
Biochem. 351:157–164. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Png KJ, Halberg N, Yoshida M and Tavazoie
SF: A microRNA regulon that mediates endothelial recruitment and
metastasis by cancer cells. Nature. 481:190–194. 2012. View Article : Google Scholar
|
|
67
|
Cascio S, D’Andrea A, Ferla R, et al:
miR-20b modulates VEGF expression by targeting HIF-1 alpha and
STAT3 in MCF-7 breast cancer cells. J Cell Physiol. 224:242–249.
2010.PubMed/NCBI
|
|
68
|
Plummer PN, Freeman R, Taft RJ, et al:
MicroRNAs regulate tumor angiogenesis modulated by endothelial
progenitor cells. Cancer Res. 73:341–352. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Mertens-Talcott SU, Chintharlapalli S, Li
X and Safe S: The oncogenic microRNA-27a targets genes that
regulate specificity protein transcription factors and the G2-M
checkpoint in MDA-MB-231 breast cancer cells. Cancer Res.
67:11001–11011. 2007. View Article : Google Scholar
|
|
70
|
Zhang X, Yu H, Lou JR, et al: MicroRNA-19
(miR-19) regulates tissue factor expression in breast cancer cells.
J Biol Chem. 286:1429–1435. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Wang F, Zheng Z, Guo J and Ding X:
Correlation and quantitation of microRNA aberrant expression in
tissues and sera from patients with breast tumor. Gynecol Oncol.
119:586–593. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Iyevleva AG, Kuligina E, Mitiushkina NV,
Togo AV, Miki Y and Imyanitov EN: High level of miR-21, miR-10b,
and miR-31 expression in bilateral vs. unilateral breast
carcinomas. Breast Cancer Res Treat. 131:1049–1059. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Chen WX, Hu Q, Qiu MT, et al: miR-221/222:
promising biomarkers for breast cancer. Tumour Biol. 34:1361–1370.
2013. View Article : Google Scholar
|
|
74
|
Buffa FM, Camps C, Winchester L, et al:
microRNA-associated progression pathways and potential therapeutic
targets identified by integrated mRNA and microRNA expression
profiling in breast cancer. Cancer Res. 71:5635–5645. 2011.
View Article : Google Scholar
|
|
75
|
Barh D, Malhotra R, Ravi B and Sindhurani
P: MicroRNA let-7: an emerging next-generation cancer therapeutic.
Curr Oncol. 17:70–80. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Kim SJ, Oh JS, Shin JY, et al: Development
of microRNA-145 for therapeutic application in breast cancer. J
Control Release. 155:427–434. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Kong W, He L, Coppola M, et al:
MicroRNA-155 regulates cell survival, growth, and chemosensitivity
by targeting FOXO3a in breast cancer. J Biol Chem. 285:17869–17879.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Hu H, Li S, Cui X, et al: The
overexpression of hypomethylated miR-663 induces chemotherapy
resistance in human breast cancer cells by targeting heparin
sulfate proteoglycan 2 (HSPG2). J Biol Chem. 288:10973–10985. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Liang Z, Wu H, Xia J, et al: Involvement
of miR-326 in chemotherapy resistance of breast cancer through
modulating expression of multidrug resistance-associated protein 1.
Biochem Pharmacol. 79:817–824. 2010. View Article : Google Scholar
|
|
80
|
Pan YZ, Morris ME and Yu AM: MicroRNA-328
negatively regulates the expression of breast cancer resistance
protein (BCRP/ABCG2) in human cancer cells. Mol Pharmacol.
75:1374–1379. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Kovalchuk O, Filkowski J, Meservy J, et
al: Involvement of microRNA-451 in resistance of the MCF-7 breast
cancer cells to chemotherapeutic drug doxorubicin. Mol Cancer Ther.
7:2152–2159. 2008. View Article : Google Scholar : PubMed/NCBI
|