Introduction
B cell lymphoma (BCL) includes Mantle cell lymphoma
(MCL), primary effusion lymphoma (PEL), Burkitt’s lymphoma (BL) and
diffuse large B-cell lymphoma (DLBCL) (1); both MCL and BL are highly aggressive
non-Hodgkin lymphomas (NHLs). MCL represents ~4% of all lymphomas
in the US and 7–9% in Europe, with a median overall survival (OS)
of 4–5 years and poor susceptibility to standard chemotherapy
strategy (2,3). BL constitutes ~1–2% of all adult NHL
cases and 30–50% of all childhood lymphomas in Western countries
(4). The current chemotherapy
regimens for MCL and BL are not very effective and a number of
patients cannot afford the dose-intensive side effects, although
dose-intensive induction regimens result in the improvement of
median OS (5). Therefore, finding
new therapy options is urgent to improve patients’ quality of
life.
Green tea contains multiple polyphenolic compounds
such as (−)-epigallocatechin-3-gallate (EGCG), (−)-epigallocatechin
(EGC), (−)-epicatechin-3-gallate (ECG) and (−)-epicatechin (EC)
(6). Among these, EGCG is the
major active component and the most abundant catechin (7). EGCG has been intensively studied in
the past years, mainly because of its diversified effects in
chemoprophylaxis and disease treatment. Indeed, EGCG has been shown
to possess anti-oxidative (8),
antimicrobia (9), and
anticarcinogenic (10) effects, in
addition to reducing the risk for cardiovascular diseases (11), diabetes (12), neurodegenerative diseases (13) and cancer. The anticancer effects of
EGCG on different cancer events have been assessed: EGCG reversed
the progression of colon carcinomas by decreasing methylation
(14), reduced mRNA expression and
showed anti-proliferative potential in cervical cancer cells
(15), induced apoptosis in skin
cancer cells (16), and inhibited
motility and angiogenesis of lung and breast cancer cells (17–19).
In addition, the effect of EGCG on suppressing division and
inducing apoptosis of cancer cells is mediated by several signaling
pathways. EGCG has been shown to activate killer caspases and
suppress NF-κB activation in epidermoid carcinoma A431 cells
(20). It has also been suggested
that EGCG induces apoptosis by activating caspases and MAPK
pathways in lung adenocarcinoma and esophageal carcinoma cells
(21). The PI3K/AKT/mTOR pathway
is involved in EGCG-induced apoptosis of pancreatic carcinoma cells
(22). Therefore, we hypothesized
that EGCG may also induce BCL cells apoptosis.
Apoptosis is an essential process, which eliminates
anomalous cells and balances the internal environment whose
disturbance can bring about cancer (23). Intrinsic (mitochondrial) and
extrinsic (death receptor) pathways are the major apoptosis routes.
Apoptosis is controlled by a crossed pathway network which contains
some targets for activity such as the crucial executor caspases
(cysteine-dependent aspartate-specific proteases). The extrinsic
pathway is triggered by tumour necrosis factor receptor (TNFR), Fas
and TNF-related apoptosis-inducing ligand (TRAIL), and activates
the initiator caspase-8. Subsequently, the signaling starts cascade
reactions that activate caspase-3 and -7. The intrinsic pathway
occurs when cytochrome c released from mitochondria to the
cytosol acts with Apaf-1 to promote the activation of caspase-9 and
the downstream members caspase-3 and -7 (24). Furthermore, B cell lymphoma-2
(Bcl-2) family members, including both pro-apoptotic (such as Bax
and Bid) and anti-apoptotic (such as Bcl-2) molecules, modulate the
mitochondrial pathway by mainly regulating the release of apoptosis
factors such as cytochrome c (25).
Although EGCG was shown to be effective in multiple
cancers, few studies have discussed the relationship between EGCG
and lymphoma, especially MCL and BL. In this study, we assessed the
effects of EGCG on Jeko-1 and Raji cells. We demonstrated that EGCG
markedly suppresses cell growth. In addition, our results revealed
that the EGCG-mediated growth inhibition occurs through increased
activation of caspases and Bcl-2 family proteins that induce
apoptosis. Consequently, EGCG may serve as emerging targeted
therapy option for B cell lymphoma.
Materials and methods
Cell culture
The MCL Jeko-1 and BL Raji cell lines were kindly
provided by Professor Hong Cen (Affiliated Tumour Hospital of
Guangxi Medical University, Nanning, China). Cells were grown in
RPMI-1640 medium supplemented with 10% fetal bovine serum (FBS, all
from Hyclone, USA) for Raji cells and 20% FBS for Jeko-1 cells, in
a humidified atmosphere containing 5% CO2 at 37°C. Cells
were subcultured every 2–3 days.
Cell proliferation assay
Cell growth inhibition effect was determined using
Cell Counting Kit-8 (CCK-8) (Dojindo Co., Japan). Jeko-1 and Raji
cells were plated at 4×104 or 5×105
cells/well in 96-well plates and cultured for 1–2 h. Then, cells
were treated with different concentrations of EGCG (0–120 μg/ml)
(Sigma-Aldrich, USA) for 12, 24 and 36 h. Cell culture medium of
the same volume was used as control. After incubation, 10 μl CCK-8
reagent were added to each well and further incubated at 37°C for 2
h. Optical density (OD) values were measured at 450 nm with a
microplate reader (Thermo Multiskan MK3, USA). Finally, growth
inhibition rate (IR) was calculated as follows: IR (%) =
(ODcontrol−ODexperiment)/(ODcontrol−ODbackgroud)
×100%.
Flow cytometry analysis of apoptosis
Apoptosis was measured with Annexin V: PE apoptosis
detection kit (BD Biosciences). First, Jeko-1 cells
(5×105 cells/well) or Raji cells (1×106
cells/well) were plated in 6-well plates, respectively, and treated
with different concentrations of EGCG (0, 20, 40 and 60 μg/ml) for
12, 24 and 36 h. Then, cells were washed with cold PBS and
resuspended in 100 μl 1X binding buffer, followed by addition of 5
μl Annexin V-PE and 5 μl 7-AAD. The cells were incubated for 15 min
at room temperature in the dark. Finally, 400 μl 1X binding buffer
were added to the cells, which were analyzed by flow cytometry (BD
Caliber, USA).
RNA extraction and reverse transcription
polymerase chain reaction (RT-PCR)
Jeko-1 and Raji cells were treated with EGCG (0, 20,
40 and 60 μg/ml) for 24 h. Total RNA was extracted using TRIzol
reagent (Invitrogen, USA) according to the manufacturer’s
instructions and quantified by NanoDrop2000 (Thermo Scientific,
USA). Equal amounts of RNA (maximum 1 μg) were reverse-transcribed
into cDNA using the ReverTra Ace® qPCR RT kit (Toyobo,
Japan). First, total RNA samples were incubated at 65°C for 5 min
and kept on ice. Then, the denatured RNA was added to the reaction
solution containing nuclease-free water, 5X RT buffer, RT enzyme
mix and primer mix, and subsequently incubated at 37°C for 15 min
and 98°C for 5 min. The resulting cDNA samples were stored at
−20°C.
Quantitative real-time PCR
Quantitative RT-PCR was performed in 20 μl reaction
containing cDNA, nuclease-free water, primers and SYBR®
Green Real-time PCR Master Mix (Toyobo, Japan) according to the
manufacturer’s instructions. Primers used were as follows: GAPDH,
5′-GTCAAGGCTGA GAACGGGAA-3′ (forward) and 5′-AAATGAGCCCCAGCCT
TCTC-3′(reverse);Bcl-2,5′-AACATCGCCCTGTGGATGAC-3′ (forward) and
5′-AGAGTCTTCAGAGACAGCCAGGAG-3′ (reverse); Bax,
5′-AGATGTGGTCTATAATGCGTTTTCC-3′ (forward) and
5′-CAGAAGGCACTAATCAAGTCAAGGT-3′ (reverse); Fas,
5′-TCTGGTTCTTACGTCTGTTGC-3′ (forward) and
5′-CTGTGCAGTCCCTAGCTTTCC-3′ (reverse). The PCR was carried out for
30 sec at 95°C (pre-denaturation) followed by 35 cycles of 5 sec at
95°C (denaturation), 10 sec at 60°C (annealing) and 15 sec at 72°C
(extension). GAPDH served as internal control to normalize the
data. All treatments were performed in triplicate and gene
expression was assessed by the 2−ΔΔCt method.
Analysis of caspase activity
Caspase-8 activation was assessed with the caspase
colorimetric assay kit (KeyGen, China). This assay is based on
spectrophotometric detection of the chromophore p-nitroanilide
(pNA) released following the cleavage of the substrate caspase.
Briefly, Jeko-1 or Raji cells were harvested after treatment with
EGCG (0, 20, 40 and 60 μg/ml) for 24 h, and lysed on ice in 100 μl
lysis buffer (containing 1 μl DTT) for 30 min. Cells lysates were
centrifuged for 1 min at 10,000 × g and the supernatants assessed
for protein content. Subsequently, supernatant samples containing
200 μg proteins were incubated with 50 μl reaction buffer
(containing 0.5 μl DTT) and 5 μl caspase-8 substrate at 37°C for 4
h in the dark. The OD values were measured at 405 nm with a
microplate reader. The activation level of caspase-8 was calculated
by ODexperiment/ODcontrol.
Western blot analysis
Jeko-1 or Raji cells were harvested after treatment
with EGCG (0, 20, 40 and 60 μg/ml) for 24 h. To inhibit apoptosis,
Z-VAD-FMK (Merck Millipore, Germany) was used to pretreat cells for
1 h before EGCG addition. Cell proteins were obtained after lysis
on ice with RIPA lysis buffer [20 mM Tris (pH 7.5), 150 mM NaCl, 1
mM Na2EDTA, 1 mM EGTA, 1% Triton X-100, 2.5 mM sodium
pyrophosphate, 1 mM β-glycerophosphate, 1 mM
Na3VO4, 1 μg/ml leupeptin] (Cell Signaling
Technology, USA) containing 1 mM PMSF. The lysates were clarified
by centrifugation at 4°C for 10 min at 14,000 × g; protein content
was measured by the BCA kit (Beyotime, China) according to the
manufacturer’s instructions. Equal amounts of protein were mixed
with loading buffer and preheated 5 min at 100°C. Proteins were
electrophoretically separated on 10–15% sodium dodecyl sulfate
(SDS)-polyacrylamide gels and transferred onto PVDF membranes on
ice for 1–2 h. After blocking with 5% non-fat milk in TBST for 1 h,
membranes were incubated with primary antibodies against β-actin,
PARP, caspase-3, -7 and -9, cleaved proteins (PARP, caspase-3, -7
and -9), Bcl-2 and Bax overnight at 4°C. Then, the membranes were
washed 3 times for 5 min with TBST and subsequently incubated with
anti-rabbit IgG HRP-secondary antibody for 2 h and washed 3 times
for 10 min with TBST. All antibodies were purchased from Cell
Signaling Technology. Finally, immunoreactive proteins were
detected with the ECL kit (Boster, China) and analyzed by GelDoc XR
system (Bio-Rad, USA).
Statistical analysis
All experiments were performed at least three times
separately. The data are presented as mean ± SD. One-way ANOVA was
used for comparisons among multiple groups. Statistical
significance was established at p<0.05.
Results
EGCG inhibits proliferation of both
Jeko-1 and Raji cells
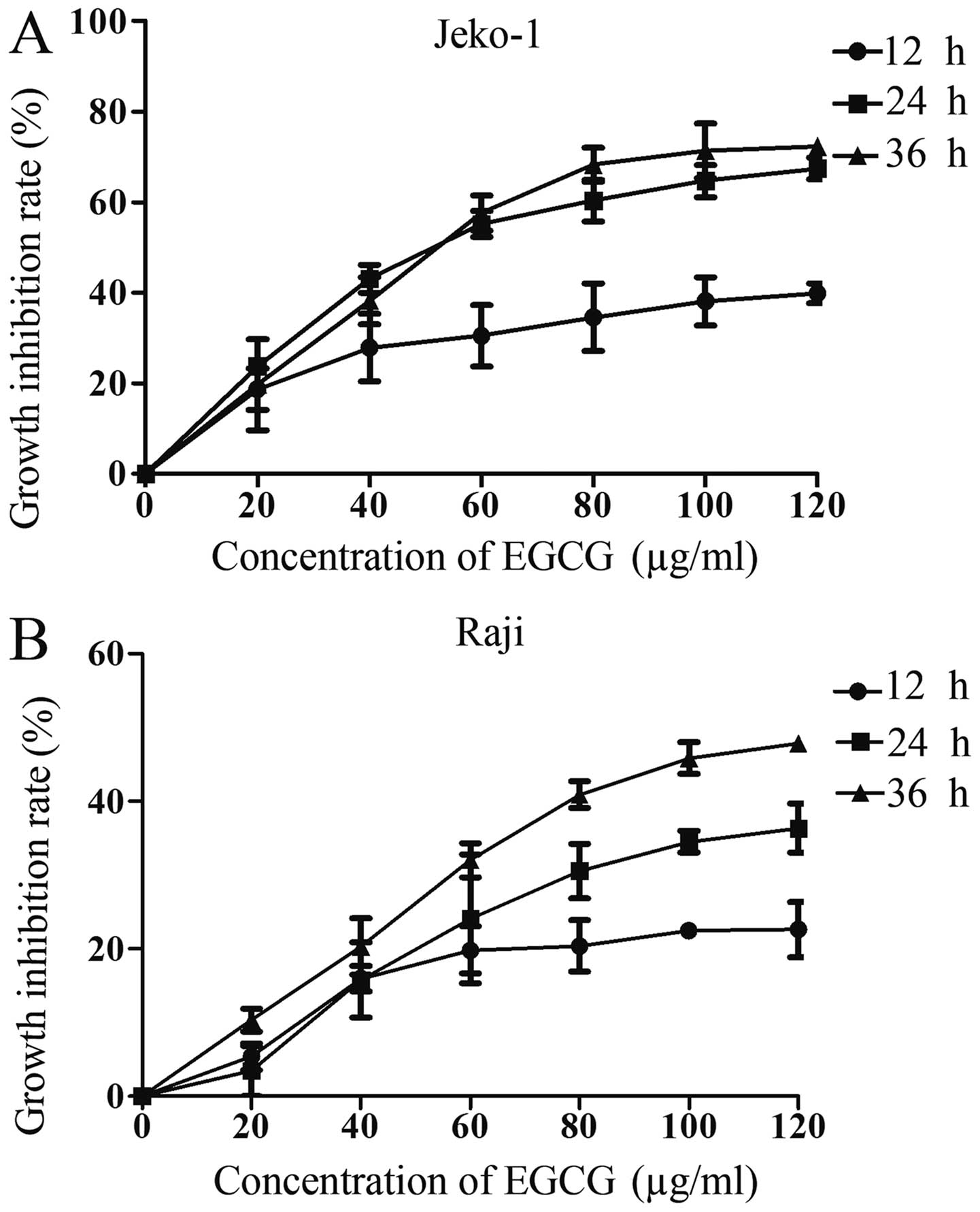
To determine the effect of EGCG on cancer cells,
Jeko-1 (Fig. 1A) or Raji (Fig. 1B) cells were treated with different
EGCG concentrations and various time-points. The growth inhibition
rate was assessed by the CCK-8 assay. As demonstrated in Fig. 1, cell inhibition rate at various
time-points increased with EGCG concentration in both cell lines.
The half-maximal inhibitory concentration (IC50) of EGCG
at 36 h for Jeko-1 and Raji was 57.98 and 61.24 μg/ml,
respectively. Therefore, 20, 40 and 60 μg/ml were selected for
subsequent experiments. These data indicated a time- and
dose-dependent effect of EGCG on growth inhibition of both Jeko-1
and Raji cells.
EGCG induces apoptosis in Jeko-1 and Raji
cells
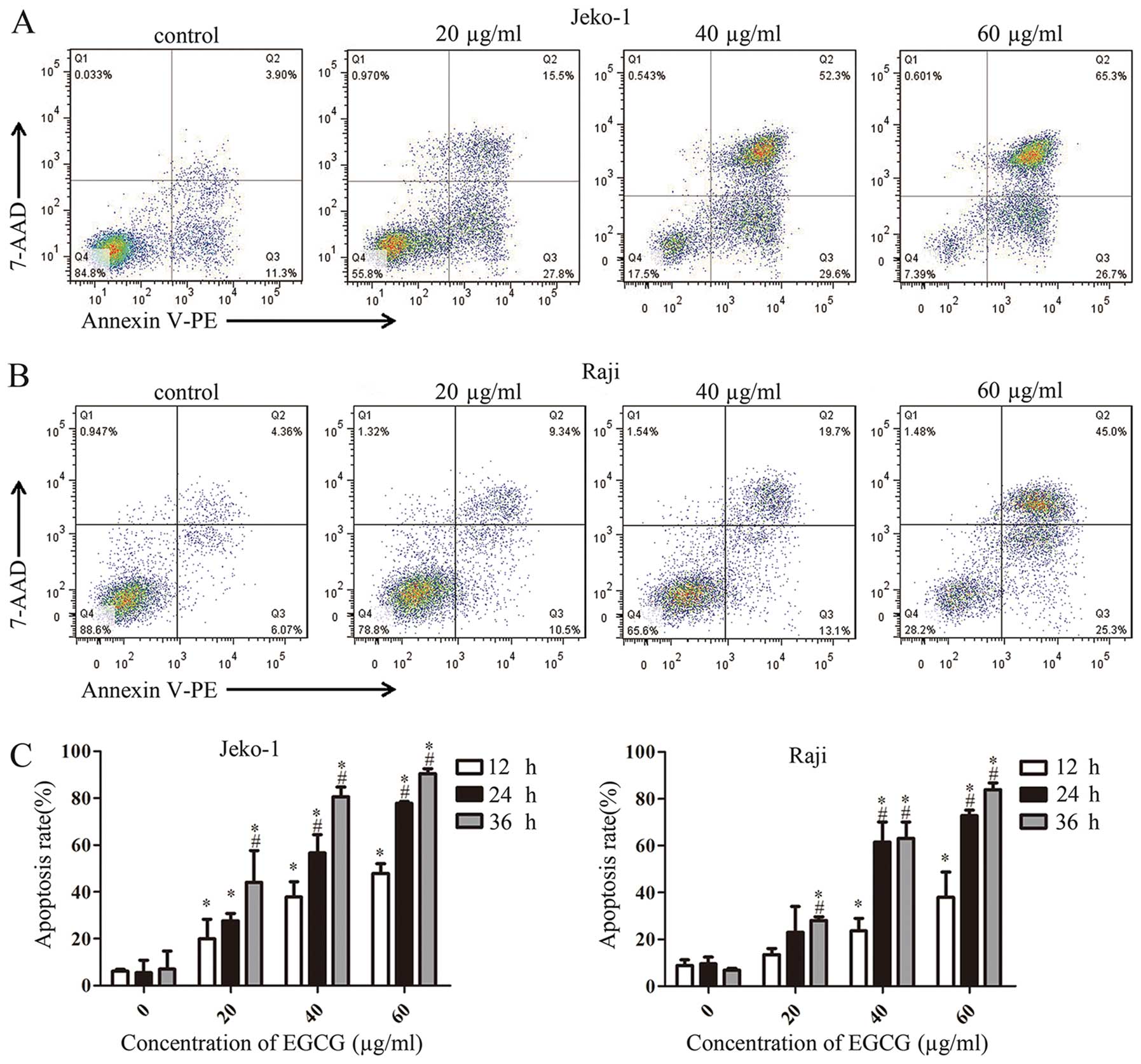
To assess whether EGCG-associated growth inhibition
resulted from induced apoptosis in Jeko-1 and Raji cells, Annexin
V-PE and 7-AAD were used to stain cells treated with different EGCG
concentrations. Annexin V is a protein with high affinity for the
phospholipid phosphatidylserine (PS) exposed from inner layer to
external environment in apoptotic cells; thus, Annexin V is used to
determine undergoing apoptosis. 7-Amino-actinomycin (7-AAD) is a
probe used to distinguish viable cells from dead and damaged ones.
As shown in Fig. 2, apoptotic
Jeko-1 (Fig. 2A) and Raji
(Fig. 2B) cells in Q2 (early
apoptosis) and Q3 (late apoptosis) increased gradually with EGCG
concentrations. Fig. 2C shows that
apoptosis rates in Jeko-1 and Raji cells were time- and
dose-dependent, in agreement with CCK-8 assay data.
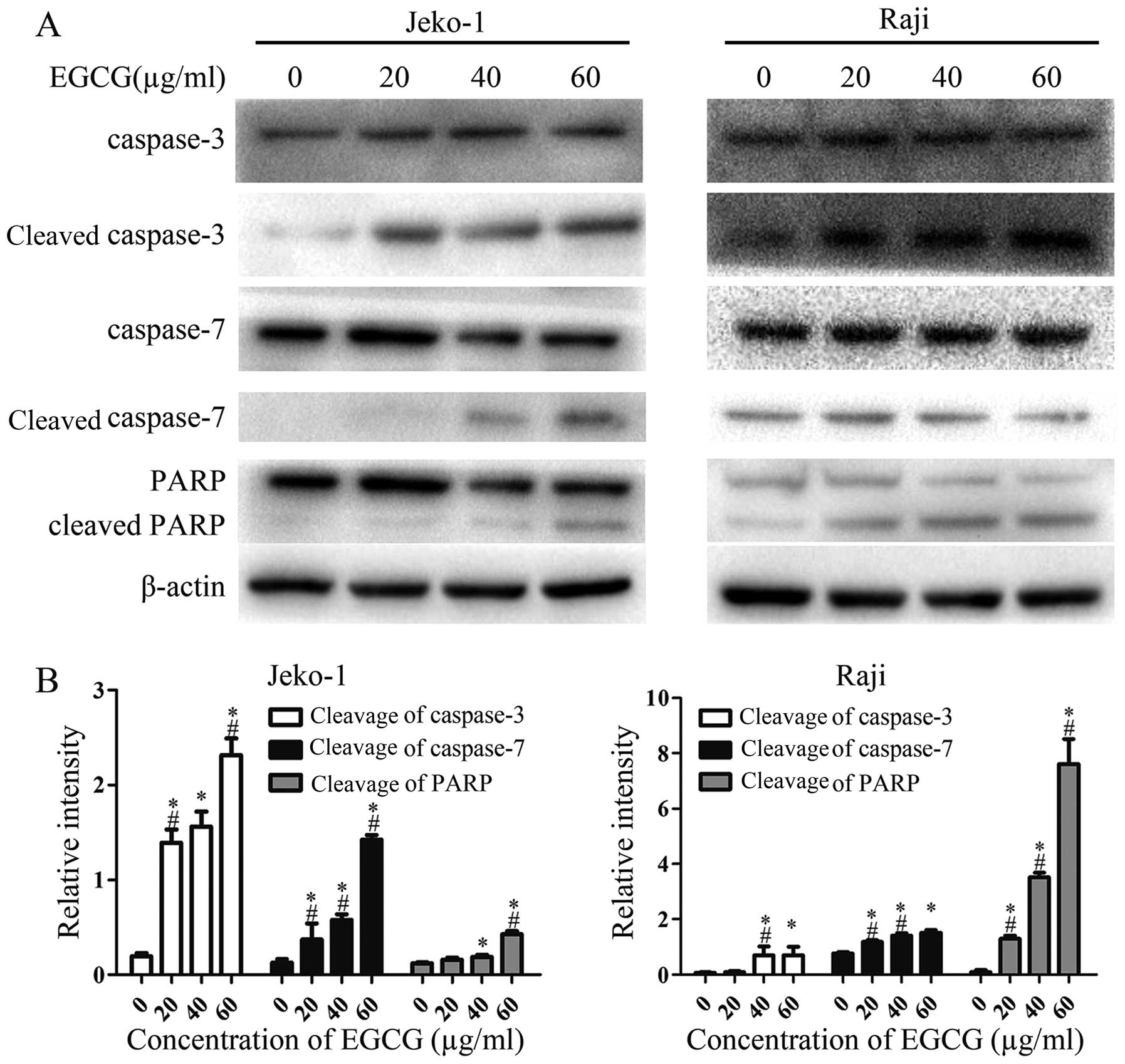
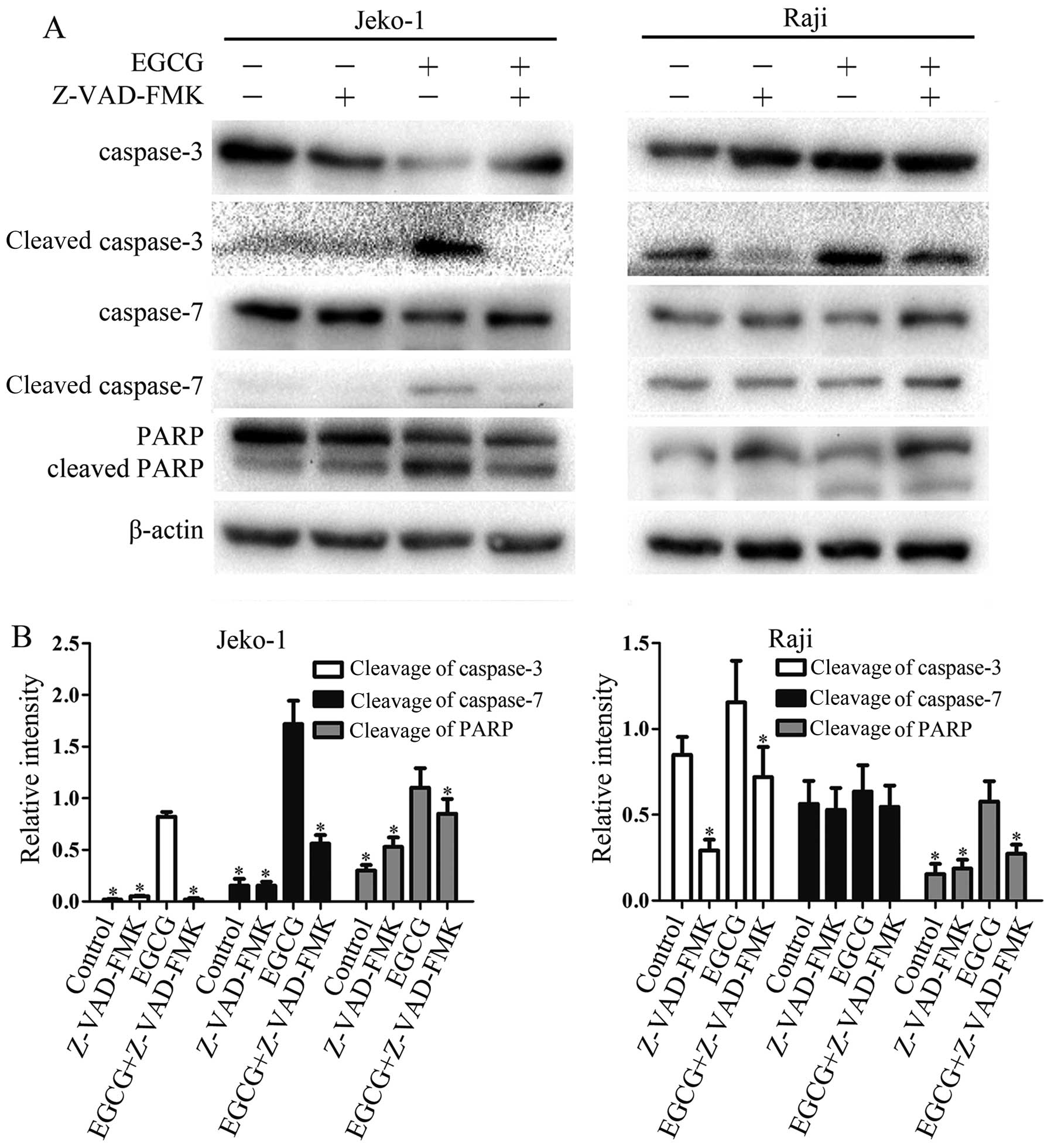
EGCG increases the activities of
caspase-3, -7 and PARP
To further investigate the mechanisms involved in
EGCG-induced apoptosis, the activities of caspase-3, -7 and PARP
were determined by western blot analysis. As shown in Fig. 3, EGCG significantly increased
caspase-3, -7 and PARP activities, and activation levels increased
in a dose-dependent manner. This suggests a relationship between
caspase activity and the EGCG-induced apoptosis in Jeko-1 and Raji
cells.
Effect of the general caspase inhibitor
Z-VAD-FMK on EGCG-induced activity of caspase-3, -7 and PARP
To determine the role of caspases in EGCG-induced
apoptosis, cells were pretreated with Z-VAD-FMK. As expected,
Z-VAD-FMK sufficiently inhibited the EGCG-induced activities of
caspase-3, -7 and PARP. Taken together, the above data strongly
suggested that the EGCG-induced apoptosis rely on caspase-dependent
pathways in Jeko-1 and Raji cells.
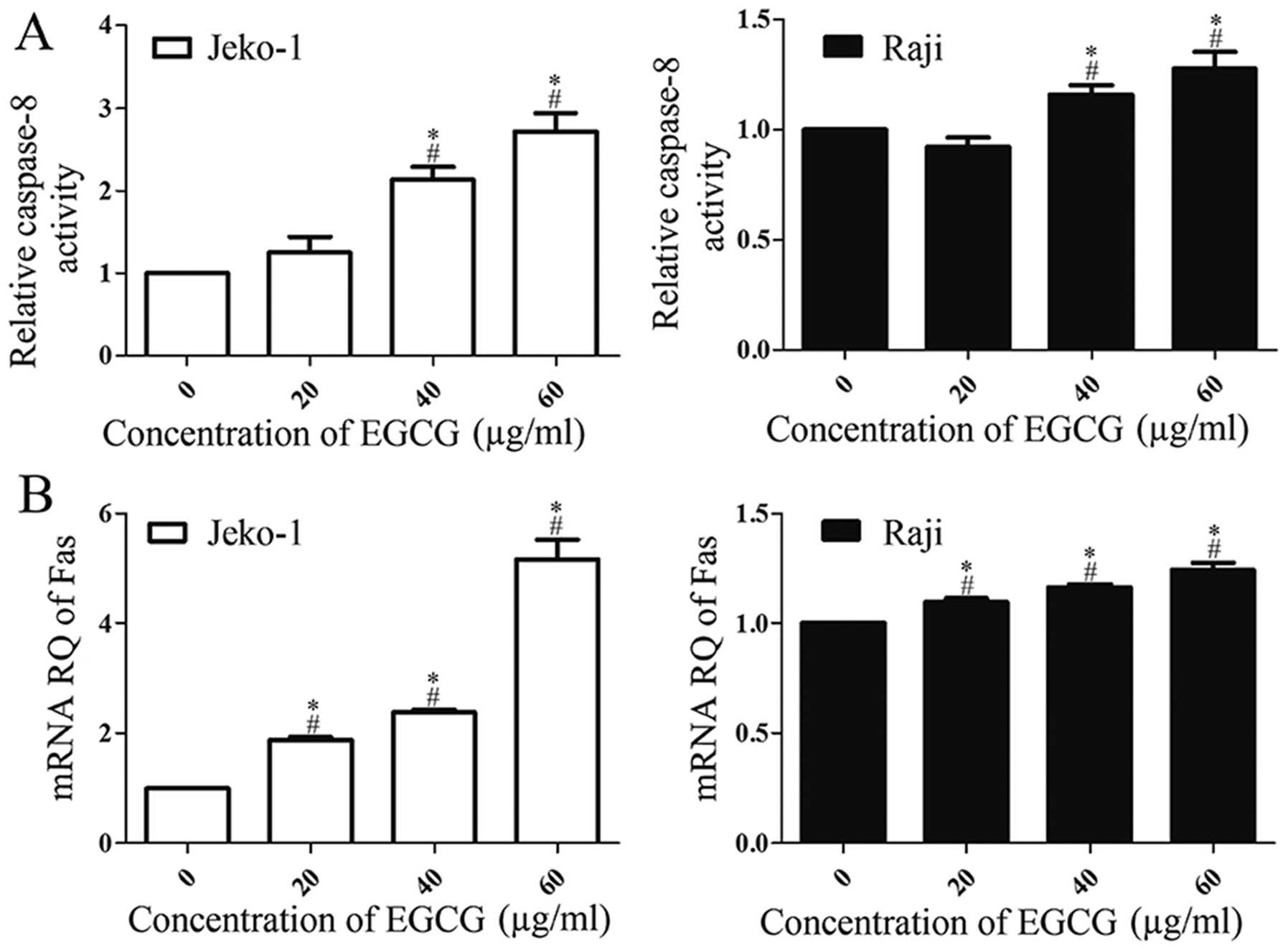
EGCG induces apoptosis through the death
receptor pathway: increases caspase-8 activity and Fas
upregulation
To explore how the upstream molecules triggered the
apoptosis executors caspase-3 and -7, and their substrate PARP, we
first determined caspase-8 activity using the caspase colorimetric
assay kit. As shown in Fig. 5A
EGCG-treated cells showed a dose-dependent increase of caspase-8
activity. Then, to explore the upstream effector that actives
caspase-8, the expression of Fas mRNA was assessed by RT-PCR.
Fig. 5B showed that Fas mRNA
expression levels increased in a dose-dependent manner in
EGCG-treated cells. Of note, the maximum caspase-8 activation and
Fas mRNA expression levels in Jeko-1 cells were at least doubled
compared with controls and significantly higher than in Raji cells.
These results suggested that EGCG-induced apoptosis was associated
with the death receptor pathway.
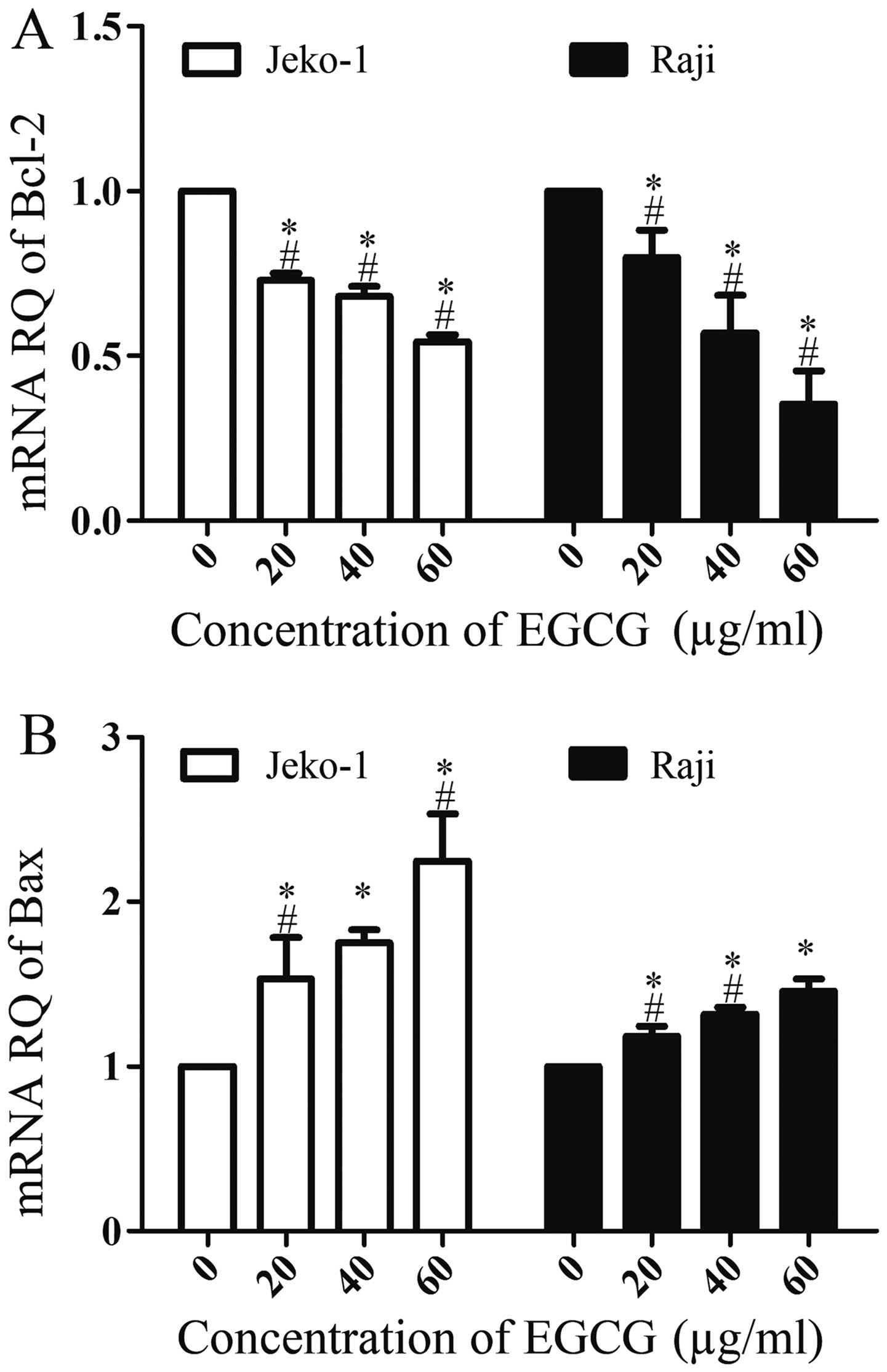
EGCG affects Bcl-2 and Bax mRNA
expression
In order to investigate whether other molecular
events were involved in EGCG-induced apoptosis in Jeko-1 and Raji
cells, we examined the expression levels of Bcl-2 and Bax mRNA by
RT-PCR. As shown in Fig. 6, EGCG
upregulated Bax mRNA expression in a dose-dependent manner; in
contrast, Bcl-2 was downregulated, also in a dose-dependent manner.
These results indicated that Bcl-2 and Bax were also involved in
EGCG-induced apoptosis.
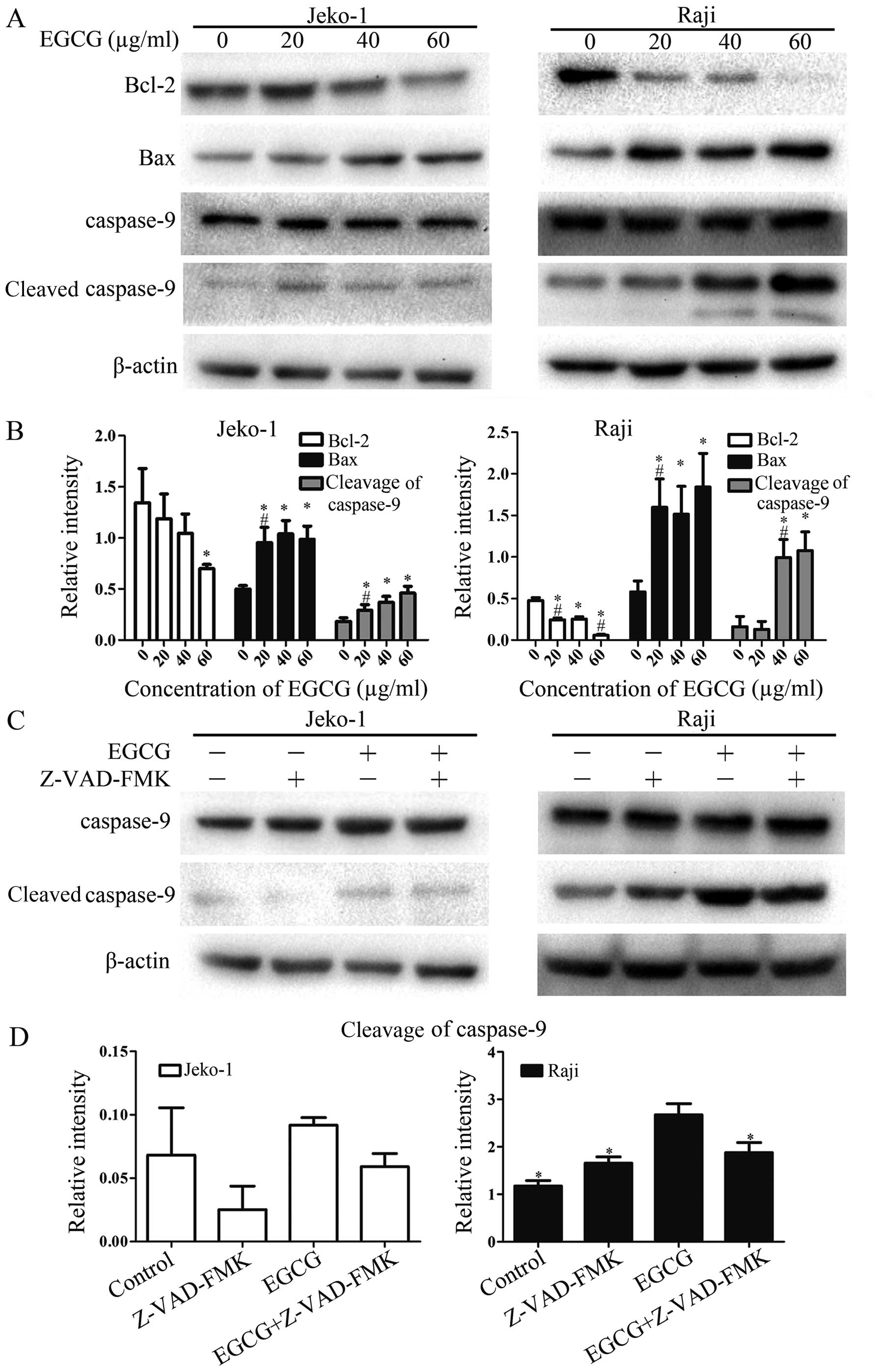
EGCG induces apoptosis through the
mitochondrial pathway: increases caspase-9 activity and regulation
of Bcl-2 and Bax
In order to further confirm the involvement of the
mitochondrial pathway, the most important and common apoptosis
pathway, in EGCG-induced apoptosis in Jeko-1 and Raji cells,
caspase-9 activity and expression levels of Bcl-2 family proteins
Bcl-2 and Bax were assessed. Treatment of both cell lines with EGCG
resulted in increased activity of caspase-9 (Fig. 7A and B). The expression of the
anti-apoptotic protein Bcl-2 was downregulated significantly,
whereas that of the pro-apoptotic protein Bax was upregulated.
Importantly, these changes were also EGCG dose-dependent. When
cells were pretreated with the general caspase inhibitor Z-VAD-FMK,
caspase-9 activity was decreased compared with the EGCG-treated
group as indicated in Fig. 7C and
D. These results suggested that EGCG triggers the mitochondrial
pathway, regulating Bcl-2 family proteins to induce apoptosis in
Jeko-1 and Raji cells.
Discussion
Natural substances are increasingly used around the
world for both chemoprophylaxis and treatment of various diseases.
For instance, EGCG has attracted growing attention due to its low
toxicity in normal cells and significant inhibitory effect on
cancer cells (26,27). Our previous research demonstrated
that EGCG significantly inhibits the growth of hepatocellular
carcinoma cells SMMC7721 at 48 h (28). In this study, we assessed the
apoptotic effects of EGCG in B lymphoma cells and investigated the
possible molecular mechanisms. We showed that EGCG significantly
inhibits Jeko-1 and Raji cell growth in a dose- and time-dependent
manner (Fig. 1), in accordance
with previous studies assessing other cancer cells (16,26,27).
Apoptosis is an essential process and fundamental
cellular activity, which eliminates anomalous cells. Inducing
cancer cell apoptosis is the key to cancer treatment, and recent
studies have shown that many natural substances induce apoptosis as
their primary anticancer mechanism (29–31).
Therefore, apoptosis rate is considered an indicator of anticancer
activity. As shown above (Fig. 2),
the apoptotic cell rate increased significantly with time and EGCG
dose. In agreement, Hazawa et al suggested that EGCG causes
time-dependent apoptosis in Raji cells (32). These findings indicated that EGCG
inhibits cell growth through induction of apoptosis.
Previous studies have proposed that potential novel
agents should mainly inhibit growth and induce apoptosis in
different lymphoma cells. Activation of caspases is a central
process of the 2 major apoptosis pathways, and activated caspases
provide a link between cell signaling and apoptotic execution
(24). It has been reported that
green tea polyphenols induce cell death by caspase-3 activation
(33). We investigated the
molecular mechanisms of EGCG induced Jeko-1 and Raji cells growth
inhibition, and found a dose-dependent increase of caspase-3 and -7
activation (Fig. 3). PARP is
associated with DNA damage in apoptosis and is the major downstream
substrate of caspase-3 (34). PARP
activation increased overtly with EGCG concentration as shown in
Fig. 3. Furthermore, the
activation of caspase-3, -7 and PARP was inhibited by the general
caspase inhibitor Z-VAD-FMK (Fig.
4). These results indicated that EGCG-induced apoptosis is
caspase-dependent in Jeko-1 and Raji cells. This finding was
consistent with a previous study demonstrating that EGCG suppresses
VEGF-R phosphorylation and induces apoptosis by increasing the
activity of caspase-3 and PARP in chronic lymphocytic leukemia B
cells (35).
Next, we focused on upstream molecules to figure out
which pathway triggers the EGCG-induced apoptosis. The death
receptor pathway triggered by stimulation of the death receptor
CD95 (APO-1/Fas) results in receptor aggregation, and FADD and
caspase-8 recruitment; subsequently, caspase-8 becomes activated
and directly leads to a cascade that activates the apoptosis
executor caspase-3 (36). The role
of EGCG and a radiolytic product in triggering the
Fas-caspase-8-medicated pathway in lymphoma U937 cells has been
demonstrated by others (37,38).
The enhanced expression of Fas mRNA and cleavage of caspase-8 with
increasing EGCG concentrations observed in this study demonstrated
that EGCG initiates the death receptor pathway to induce apoptosis
(Fig. 5), in accordance with the
above previous studies.
The mitochondrial pathway has been shown by multiple
studies to play a major role in malignant cell apoptosis induced by
many natural agents, including EGCG (25–27).
The Bcl-2 family plays a pivotal role in regulating cell life and
death; many of these apoptosis-related proteins reside in the outer
mitochondrial membrane. Once the balance of Bcl-2 (preventing cell
survival from various chemotherapeutic agents) and Bax (a
pro-apoptosis protein) is broken, the proteins move towards the
cytosol and modulate mitochondria to promote cytochrome c
release, then combine with caspase-9 to form a complex that
triggers caspase-3 activation (39,40).
Our results also indicated that EGCG increases the protein and mRNA
expression of Bax as well as caspase-9 activation while decreasing
the expression of Bcl-2, all in a dose-dependent manner (Figs. 6 and 7). In other words, EGCG initiated the
mitochondrial pathway at the same time to induce apoptosis in
Jeko-1 and Raji cells. However, it has been demonstrated that EGCG
induces apoptosis in Hep2 cells, relying on p53-mediated and
AIF-dependent mitochondrial pathway, which is not a
caspase-dependent pathway (41).
In addition, EGCG was shown to induce apoptosis in
imatinib-resistance chronic myelogenous leukemia K562 cells via a
caspase-independent mechanism (42). Whether this phenomenon is
idiosyncratic requires further studies in other cell lines.
Previous studies have revealed the means by which EGCG or other
natural agents cause the mitochondria to release cytochrome
c: one direct means is mediated by Bcl-2 family proteins;
the other is through Fas aggregated caspase-8, not directly
activating the caspase-3 cascade but followed by cleaving of Bid to
tBid and subsequent cytochrome c release that activates
caspase-9 and caspase-3 (37,43).
Although our results suggested that EGCG induces apoptosis through
both the extrinsic (death receptor) and intrinsic (mitochondrial)
pathways, it is unclear whether the two pathways are linked by Bid
cleavage in Jeko-1 and Raji cells. To our knowledge, the present
data demonstrate for the first time that EGCG induces cytotoxicity
in MCL Jeko-1 cells. The cytotoxic effect of EGCG on BL Raji cells
corroborates with a previous study (32). We further unveiled the molecular
mechanisms by which EGCG activates the caspase pathways to cause
apoptosis of MCL and BL cells. Our results indicate that
EGCG-induced apoptosis mainly involves both the death receptor and
mitochondrial pathways, which depend on the activation of
caspase-3, -7, -8, -9 and PARP as well as the mediation of Bcl-2
family proteins. Overall, EGCG may be a potential novel therapeutic
agent against B cell lymphoma.
Acknowledgements
This study was supported by grants from the Youth
Science Foundation of Guangxi Medical University (no. 02602211011),
Guangxi Natural Science Foundation (no. 2013GXNSFBA019186) and
Guangxi Science and Technology Development Program (no.
1298003-2-9).
References
|
1
|
Said JW: Aggressive B-cell lymphomas: How
many categories do we need. Mod Pathol. 26(Suppl 1): S42–S56. 2013.
View Article : Google Scholar
|
|
2
|
Vose JM: Mantle cell lymphoma: 2012 update
on diagnosis, risk-stratification, and clinical management. Am J
Hematol. 87:604–609. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bosch F, López-Guillermo A, Campo E,
Ribera JM, Conde E, Piris MA, Vallespí T, Woessner S and Montserrat
E: Mantle cell lymphoma: Presenting features, response to therapy,
and prognostic factors. Cancer. 82:567–575. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li ZJ, Yao C, Liu SF, Chen L, Xi YM, Zhang
W and Zhang GS: Cytotoxic effect of icaritin and its mechanisms in
inducing apoptosis in human burkitt lymphoma cell line. Biomed Res
Int. 2014:3915122014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chang JE and Kahl BS: Current status of
targeted therapies for Mantle cell lymphoma. Drugs. 71:2307–2326.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen D, Milacic V, Chen MS, et al: Tea
polyphenols, their biological effects and potential molecular
targets. Histol Histopathol. 23:487–496. 2008.PubMed/NCBI
|
|
7
|
Khan N, Afaq F, Saleem M, Ahmad N and
Mukhtar H: Targeting multiple signaling pathways by green tea
polyphenol (−)-epigallocatechin-3-gallate. Cancer Res.
66:2500–2505. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Łuczaj W, Waszkiewicz E, Skrzydlewska E
and Roszkowska-Jakimiec W: Green tea protection against
age-dependent ethanol-induced oxidative stress. J Toxicol Environ
Health A. 67:595–606. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Betts JW and Wareham DW: In vitro activity
of curcumin in combination with epigallocatechin gallate (EGCG)
versus multidrug-resistant Acinetobacter baumannii. BMC Microbiol.
14:1722014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xu X, Zhou XD and Wu CD: The tea catechin
epigallocatechin gallate suppresses cariogenic virulence factors of
Streptococcus mutans. Antimicrob Agents Chemother. 55:1229–1236.
2011. View Article : Google Scholar :
|
|
11
|
Kuriyama S, Shimazu T, Ohmori K, Kikuchi
N, Nakaya N, Nishino Y, Tsubono Y and Tsuji I: Green tea
consumption and mortality due to cardiovascular disease, cancer,
and all causes in Japan: The Ohsaki study. JAMA. 296:1255–1265.
2006. View Article : Google Scholar
|
|
12
|
Song Y, Manson JE, Buring JE, Sesso HD and
Liu S: Associations of dietary flavonoids with risk of type 2
diabetes, and markers of insulin resistance and systemic
inflammation in women: A prospective study and cross-sectional
analysis. J Am Coll Nutr. 24:376–384. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hu G, Bidel S, Jousilahti P, Antikainen R
and Tuomilehto J: Coffee and tea consumption and the risk of
Parkinson’s disease. Mov Disord. 22:2242–2248. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Moseley VR, Morris J, Knackstedt RW and
Wargovich MJ: Green tea polyphenol epigallocatechin 3-gallate,
contributes to the degradation of DNMT3A and HDAC3 in HCT 116 human
colon cancer cells. Anticancer Res. 33:5325–5333. 2013.PubMed/NCBI
|
|
15
|
Muthusami S, Prabakaran DS, An Z, Yu JR
and Park WY: EGCG suppresses Fused Toes Homolog protein through p53
in cervical cancer cells. Mol Biol Rep. 40:5587–5596. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Singh T and Katiyar SK: Green tea
polyphenol, (−)-epigallocatechin-3-gallate, induces toxicity in
human skin cancer cells by targeting β-catenin signaling. Toxicol
Appl Pharmacol. 273:418–424. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Takahashi A, Watanabe T, Mondal A, Suzuki
K, Kurusu-Kanno M, Li Z, Yamazaki T, Fujiki H and Suganuma M:
Mechanism-based inhibition of cancer metastasis with
(−)-epigallocatechin gallate. Biochem Biophys Res Commun. 443:1–6.
2014. View Article : Google Scholar
|
|
18
|
Sakamoto Y, Terashita N, Muraguchi T,
Fukusato T and Kubota S: Effects of epigallocatechin-3-gallate
(EGCG) on A549 lung cancer tumor growth and angiogenesis. Biosci
Biotechnol Biochem. 77:1799–1803. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Braicu C, Gherman CD, Irimie A and
Berindan-Neagoe I: Epigallocatechin-3-Gallate (EGCG) inhibits cell
proliferation and migratory behaviour of triple negative breast
cancer cells. J Nanosci Nanotechnol. 13:632–637. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gupta S, Hastak K, Afaq F, Ahmad N and
Mukhtar H: Essential role of caspases in
epigallocatechin-3-gallate-mediated inhibition of nuclear factor
kappa B and induction of apoptosis. Oncogene. 23:2507–2522. 2004.
View Article : Google Scholar
|
|
21
|
Gao Y, Li W, Jia L, Li B, Chen YC and Tu
Y: Enhancement of (−)-epigallocatechin-3-gallate and
theaflavin-3-3′-digallate induced apoptosis by ascorbic acid in
human lung adenocarcinoma SPC-A-1 cells and esophageal carcinoma
Eca-109 cells via MAPK pathways. Biochem Biophys Res Commun.
438:370–374. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu S, Wang XJ, Liu Y and Cui YF:
PI3K/AKT/mTOR signaling is involved in
(−)-epigallocatechin-3-gallate-induced apoptosis of human
pancreatic carcinoma cells. Am J Chin Med. 41:629–642. 2013.
View Article : Google Scholar
|
|
23
|
Cotter TG: Apoptosis and cancer: The
genesis of a research field. Nat Rev Cancer. 9:501–507. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kumar S: Caspase function in programmed
cell death. Cell Death Differ. 14:32–43. 2007. View Article : Google Scholar
|
|
25
|
Tsujimoto Y: Bcl-2 family of proteins:
Life-or-death switch in mitochondria. Biosci Rep. 22:47–58. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li W, Nie S, Yu Q and Xie M:
(−)-Epigallocatechin-3-gallate induces apoptosis of human hepatoma
cells by mitochondrial pathways related to reactive oxygen species.
J Agric Food Chem. 57:6685–6691. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wu PP, Kuo SC, Huang WW, Yang JS, Lai KC,
Chen HJ, Lin KL, Chiu YJ, Huang LJ and Chung JG:
(−)-Epigallocatechin gallate induced apoptosis in human adrenal
cancer NCI-H295 cells through caspase-dependent and
caspase-independent pathway. Anticancer Res. 29:1435–1442.
2009.PubMed/NCBI
|
|
28
|
Shen X, Zhang Y, Feng Y, Zhang L, Li J,
Xie YA and Luo X: Epigallocatechin-3-gallate inhibits cell growth,
induces apoptosis and causes S phase arrest in hepatocellular
carcinoma by suppressing the AKT pathway. Int J Oncol. 44:791–796.
2014.PubMed/NCBI
|
|
29
|
Ryu DS, Kim SH, Kwon JH and Lee DS:
Orostachys japonicus induces apoptosis and cell cycle arrest
through the mitochondria-dependent apoptotic pathway in AGS human
gastric cancer cells. Int J Oncol. 45:459–469. 2014.PubMed/NCBI
|
|
30
|
Li J, Zhang F and Wang S: A polysaccharide
from pomegranate peels induces the apoptosis of human osteosarcoma
cells via the mitochondrial apoptotic pathway. Tumour Biol.
35:7475–7482. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Benarba B, Meddah B and Aoues A: Bryonia
dioica aqueous extract induces apoptosis through mitochondrial
intrinsic pathway in BL41 Burkitt’s lymphoma cells. J
Ethnopharmacol. 141:510–516. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hazawa M, Takahashi K, Sugata S and
Kashiwakura I: (−)-Epigallocatechin-3-O-gallate induces
nonapoptotic cell death in leukemia cells independent of the 67 kDa
laminin receptor. J Nat Prod. 74:695–700. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Islam S, Islam N, Kermode T, Johnstone B,
Mukhtar H, Moskowitz RW, Goldberg VM, Malemud CJ and Haqqi TM:
Involvement of caspase-3 in epigallocatechin-3-gallate-mediated
apoptosis of human chondrosarcoma cells. Biochem Biophys Res
Commun. 270:793–797. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Talanian RV, Quinlan C, Trautz S, Hackett
MC, Mankovich JA, Banach D, Ghayur T, Brady KD and Wong WW:
Substrate specificities of caspase family proteases. J Biol Chem.
272:9677–9682. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lee YK, Bone ND, Strege AK, Shanafelt TD,
Jelinek DF and Kay NE: VEGF receptor phosphorylation status and
apoptosis is modulated by a green tea component,
epigallocatechin-3-gallate (EGCG), in B-cell chronic lymphocytic
leukemia. Blood. 104:788–794. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Debatin KM and Krammer PH: Death receptors
in chemotherapy and cancer. Oncogene. 23:2950–2966. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yu DY, Zhao QL, Furuta M, Todoriki S,
Izumi K, Yamakage K, Matsumoto K, Nomura T and Kondo T: Molecular
mechanisms of apoptosis induction by 2-dodecylcyclobutanone, a
radiolytic product of palmitic acid, in human lymphoma U937 cells.
Apoptosis. 17:636–645. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hayakawa S, Saeki K, Sazuka M, Suzuki Y,
Shoji Y, Ohta T, Kaji K, Yuo A and Isemura M: Apoptosis induction
by epigallocatechin gallate involves its binding to Fas. Biochem
Biophys Res Commun. 285:1102–1106. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ola MS, Nawaz M and Ahsan H: Role of Bcl-2
family proteins and caspases in the regulation of apoptosis. Mol
Cell Biochem. 351:41–58. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Reed JC, Jurgensmeier JM and Matsuyama S:
Bcl-2 family proteins and mitochondria. Biochim Biophys Acta.
1366:127–137. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Lee JH, Jeong YJ, Lee SW, Kim D, Oh SJ,
Lim HS, Oh HK, Kim SH, Kim WJ and Jung JY: EGCG induces apoptosis
in human laryngeal epidermoid carcinoma Hep2 cells via mitochondria
with the release of apoptosis-inducing factor and endonuclease G.
Cancer Lett. 290:68–75. 2010. View Article : Google Scholar
|
|
42
|
Iwasaki R, Ito K, Ishida T, Hamanoue M,
Adachi S, Watanabe T and Sato Y: Catechin, green tea component,
causes caspase-independent necrosis-like cell death in chronic
myelogenous leukemia. Cancer Sci. 100:349–356. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Das A, Banik NL and Ray SK: Mechanism of
apoptosis with the involvement of calpain and caspase cascades in
human malignant neuroblastoma SH-SY5Y cells exposed to flavonoids.
Int J Cancer. 119:2575–2585. 2006. View Article : Google Scholar : PubMed/NCBI
|