Introduction
Tuberous sclerosis complex (TSC) is a genetic
disorder characterized by multisystem involvement and wide
phenotypic variability. This condition results in the development
of non-cancerous tumors in various organs and most frequently
affects the brain, skin, kidney, lung, heart, and retina. TSC
manifestations in the central nervous system include cortical
tubers, subependymal nodules, subependymal giant cell astrocytomas,
and scattered abnormal cells throughout the brain (1). A majority of patients with TSC reveal
neurological and/or psychiatric symptoms, including epilepsy,
intellectual disability, autism spectrum disorder (ASD), attention
deficit, depression, and anxiety disorder, which range from mild to
severe and may impair their ability to live an independent
life.
Mutation of either TSC1 or TSC2 causes
TSC (2,3). Protein products of TSC1
(hamartin) and TSC2 (tuberin) form a complex that inhibits
the Ras homologue enriched in the brain (Rheb), a small G protein
that activates mammalian target of rapamycin complex 1 (mTORC1).
Defects of TSC1 or TSC2 cause excessive mTORC1
activation, which in turn provokes abnormal regulation of important
cellular processes such as cellular growth and proliferation
(4,5). The Knudson’s ‘two-hit’ model
(6) has been the working molecular
model for tumor development in TSC for several years. In fact, loss
of heterozygosity (LOH) of TSC1 or TSC2 has been
demonstrated in renal angiomyolipomas (7–9) and
in subependymal giant cell astrocytomas (10). However, evidence for LOH in TSC
cortical tubers is limited (11).
On the other hand, haploinsufficiency of these genes is also
speculated to be involved in TSC pathogenesis. To reveal Tsc
mutation-related mechanisms of the pathogenesis, rodents harboring
a defect of the Tsc1 or Tsc2 gene have been
extensively investigated (12–15).
For instance, Tsc1+/− and
Tsc2+/− mouse models exhibit learning and memory
deficits (16,17). Eker rats are heterozygous for a
mutation of Tsc2 and develop hereditary kidney cancer by the
age of 1 year (18–20). Although kidney cancer is rare in
human patients with TSC, it is the only cancer known to occur at an
increased incidence in TSC. The embryonic lethality of
Tsc2−/− Eker rat embryos is characterized by
disrupted neuroepithelial growth (21). Although cortical tubers are rare
(22), 63% of Eker rats develop
brain lesions comprising a mixture of large and elongated cells in
both subependymal and subcortical regions (23,24).
In contrast, among Tsc1 and Tsc2 knock-out mouse
models, only conditional ablation in the brain can induce such
lesions (25). Consequently, with
regard to brain lesions, the Eker rat model is more similar to the
human patients compared with other mouse models.
We observed that the tumorigenicity of
Tsc2−/− cells derived from mice was effectively
inhibited by rapamycin treatment (26). Other groups reported a similar
effect when Eker rats or knock-out mice were treated with
rapamycin, although some residual tumors were detected (27,28).
These findings have provided the rationale for therapy with
rapalogues to treat TSC lesions such as lymphangioleiomyomatosis,
SEGAs, and angiomyolipomas, directed at the abnormal activation of
mTORC1 (29–31). Although decreased tumor volume has
been documented, complete cure was not achieved in most cases. In
addition, there are several problems associated with long-term use
of rapalogues, including various undesirable side-effects.
Consequently additional therapeutic molecular targets are required.
The pathogenesis of TSC is assumed to be related to abnormal
differentiation as a result of TSC1/2 deficiency. For
instance, abnormal giant cells that appear in brain lesions of
patients with TSC express both neuronal and glial lineage markers
(32). In recent years, a number
of articles have revealed differentiation- and cell type-specific
abnormalities using in vitro differentiation protocols to
investigate differentiation of embryonic stem cells (ESCs) and
induced pluripotent stem cells (iPSCs). To evaluate roles of
Tsc2 from the viewpoint of differentiation and
tissue-specific pathogenesis as well as to compare and combine
in vivo and in vitro models, we established ESCs from
Eker rats.
In 2008, authentic rat ESCs were established for the
first time (33,34), lagging behind the establishment of
mouse (35,36) and human ESCs (37). Using methods described by Buehr
et al (33), we generated
ESCs from blastocysts of Eker rats to establish an in vivo
experimental system to explore the role of Tsc2 in TSC
pathogenesis. Although several reports have indicated the necessity
of Tsc1 and Tsc2 regulation to maintain ESCs
(38) and somatic stem cells
(39) or to establish iPS cells
(40), we were able to establish
not only Tsc2+/+ and Tsc2+/−
ESCs but also Tsc2−/− ESCs. To our knowledge,
this is the first report describing the generation of
Tsc2-deficient ESCs.
Materials and methods
Ethics statement
All animal experiments were conducted in strict
accordance with the institutional guidelines of Juntendo University
for animal experiments. The protocol was approved by the Animal
Experimentation Committee of Juntendo University (Tokyo, Japan)
(approval no. 250105). All surgical procedures were performed under
isoflurane anesthesia, and all efforts were made to minimize animal
suffering.
Animals
Genetic homogeneity of Eker rats was maintained in
our laboratory by brother-sister mating. Wistar rats, Brown Norway
rats, C57BL/6J mice, and nude mice were purchased from Charles
River Laboratories Japan, Inc. (Kanagawa, Japan). All animals were
housed under specific pathogen-free conditions.
Mouse embryonic fibroblasts (MEFs)
MEFs were derived from embryonic day 14.5 C57BL6/J
mouse embryos. MEFs were cultured in Knockout DMEM supplemented
with 10% fetal bovine serum, 1% L-glutamine, and penicillin
streptomycin (all from Gibco Life technologies, Carlsbad, CA, USA)
on gelatin-coated dishes. MEFs were treated with mitomycin C
(Sigma-Aldrich, St. Louis, MO, USA) for use as feeder cells.
Culture of ESCs
We generated ESCs from Eker rats according to the
method reported by Buehr et al (33). After double heterozygous mating of
Eker rats, E4.5 blastocysts were gently flushed out from uteri
using the N2B27 medium (StemCells, Inc., Newark, CA, USA). After
removal of zonae pellucidae with acid Tyrode’s solution, whole
blastocysts were plated and cultured on mitomycin C-treated MEFs in
N2B27 medium supplemented with 3 μM of CHIR99021, 1 μM of PD0325901
(both from Axon Medchem BV, Groningen, The Netherlands), 1,000 U/ml
rat leukemia inhibitory factor (LIF) (ESGRO®; Millipore,
Bedford, MA, USA) [two inhibitors (2i) + LIF condition]. After 5–7
days, blastocyst outgrowths were cut into pieces and replated in
the same 2i + LIF medium. Thereafter, emerging ESC colonies were
dissociated using Accutase (Innovative Cell Technologies, Inc., San
Diego, CA, USA) and passaged every 2–4 days.
Alkaline phosphatase staining
Alkaline phosphatase staining was performed with an
alkaline phosphatase kit (85L3R; Sigma-Aldrich) according to the
manufacturer’s instructions.
Chromosomal analysis
A standard chromosome preparation method using
colchicine treatment was employed. Chromosome preparations were
analyzed after Giemsa staining. At least 30 metaphase chromosome
sets were analyzed for each line.
Genotyping polymerase chain reaction
(PCR)
Genotyping of ESCs was conducted using PCR on ESC
DNA. To discriminate Tsc2 mutant or wild-type alleles, the
following primers were used: 5MFJ (5′-ACC ATC AGG ATG CTG CTG
AA-3′), 3MFJ2 (5′-CTA TGG CCA CAT GTG ACC AA-3′), and TSR27 (5′-GCG
CCA GAT TCA CCT CAT TA-3′) (41).
PCR was used to identify the gender of ESCs by amplification of the
rat Y chromosome-specific Sry gene using the primer pair
Sry-F (5′-CAT CGA AGG GTT AAA GTG CCA-3′) and Sry-R (5′-ATA GTG TGT
AGG TTG TTG TCC-3′) (33).
Reverse transcription (RT)-PCR
Total RNA was obtained using a
NucleoSpin® RNA II kit (Macherey-Nagel GmbH & Co.
KG, Düren, Germany) according to the manufacturer’s instructions.
Complementary DNA was synthesized using a SuperScript III
First-Strand Synthesis SuperMix kit (Invitrogen Life Technologies,
Carlsbad, CA, USA) and an oligo-dT primer, according to the
manufacturer’s instructions. PCR was performed in a thermal cycler
(Hybaid MBS 0.2G Thermal Cycler; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). The following primer pairs were used: Oct4-F
(5′-GGG ATG GCA TAC TGT GGA C-3′), Oct4-R (5′-CTT CCT CCA CCC ACT
TCT C-3′), Sox2-F (5′-GGC GGC AAC CAG AAG AAC AG-3′), Sox2-R
(5′-GTT GCT CCA GCC GTT CAT GTG-3′), rat Nanog-F (5′-GCC CTG AGA
AGA AAG AAG AG-3′), rat Nanog-R (5′-CGT ACT GCC CCA TAC TGG AA-3′)
(33), rat nestin-F (5′-AGC CAT
TGT GGT CTA CTG A-3′), rat nestin-R (5′-TGC AAC TCT GCC TTA
TCC-3′), Sox17-F (5′-AGG AGA GGT GGT GGC GAG TAG-3′), and Sox17-R
(5′-GTT GGG ATG GTC CTG CAT GTG-3′) (34).
Western blotting
Cells were harvested and lysed in sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) sample buffer
(50 mM Tris-HCl, pH 6.8, 2% SDS, and 10% glycerol). Proteins were
separated by SDS-PAGE and transferred onto a polyvinylidene
fluoride (PVDF) membrane (Millipore). The membrane was blocked with
1% skimmed milk in Tris-buffered saline containing 0.05% Tween-20
and probed with appropriate antibodies using the EnVision System
(DakoCytomation, Glostrup, Denmark). Antibody signals were
developed using ECL reagents and Hyperfilm ECL film (both from GE
Healthcare, Little Chalfont, UK), which were then scanned using
CEPROS SV (Fujifilm, Tokyo, Japan). The following primary
antibodies were used: anti-Tsc2 antibody (C20; 1:200; Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA), anti-Tsc1 primary
antibody (c-Tsc1, 1:500), anti-phospho-S6 ribosomal protein
(Ser235/236) rabbit polyclonal antibody (1:1,000, no. 2211),
anti-S6 ribosomal protein rabbit monoclonal antibody (1:1,000, no.
2217) (both from Cell Signaling Technology, Inc., Danvers, MA,
USA), and anti-β-actin mouse monoclonal antibody (1:1,000;
Sigma-Aldrich).
Embryoid body (EB) formation
ESCs were plated into low-adhesion 96-well dishes
(MS-9096; Sumitomo Bakelite Co., Ltd., Tokyo, Japan). After 10 days
of suspension culture, EBs were plated onto Matrigel-coated dishes
in GMEM/10% fetal bovine serum medium (both from Gibco Life
Technologies).
Immunocytochemistry
Cells were fixed and permeabilized with 4%
paraformaldehyde and 0.25% Triton X-100 (both from Wako Pure
Chemical Industries, Ltd., Osaka, Japan) in PBS for 30 min at 4°C
and then washed (3×5 min) with PBS/0.1% bovine serum albumin (BSA)
(Iwai Kagaku Co., Tokyo, Japan). Cells were incubated with a
primary antibody in PBS with 1% BSA for 1 h at room temperature.
Thereafter, cells were washed and incubated with
fluorophore-conjugated secondary antibodies and
4′,6-diamidino-2-phenylindole (DAPI) for 1 h at room temperature.
Immunofluorescent images were captured using a Leica TCS SP5 v2.0
system (Leica, Heidelberg, Germany). The following primary
antibodies were used: anti-Oct3/4 mouse monoclonal antibody (1:50,
C-10; Santa Cruz Biotechnology, Inc.), anti-Sox2 rabbit polyclonal
antibody (1:100, poly6309; BioLegend, San Diego, CA, USA),
anti-β-III tubulin mouse monoclonal antibody (1:500, Tuj-1; Covance
Laboratories, Princeton, NJ, USA), anti-myosin heavy chain mouse
monoclonal antibody (1:50, MF20; R&D Systems, Minneapolis, MN,
USA), and anti-Gata4 mouse monoclonal antibody (1:50; Santa Cruz
Biotechnology, Inc.). Alexa Fluor (488 or 568)-conjugated goat
anti-mouse or goat anti-rabbit secondary antibodies (Invitrogen
Life Technologies) were used at 1:1,000 dilutions.
Teratoma formation
Approximately 5×105 cells were injected
under kidney capsules of nude mice. Tumors were dissected after 4–5
weeks and fixed in 10% buffered formalin. Tumor tissues were
embedded in paraffin wax, sectioned, and examined after hematoxylin
and eosin staining.
Blastocyst injection and generation of
chimeric rats
Because collection of many blastocysts from Brown
Norway rats is inefficient, we attempted the chimera formation
assay using Brown Norway as well as Wistar rats. Rat blastocysts at
E4.5 days were collected on the day of injection and cultured for
2–3 h to ensure cavitation. ESCs were disaggregated using Accutase,
and 10–12 cells were injected into blastocyst cavities. Injected
embryos were transferred into uteri of pseudopregnant rats.
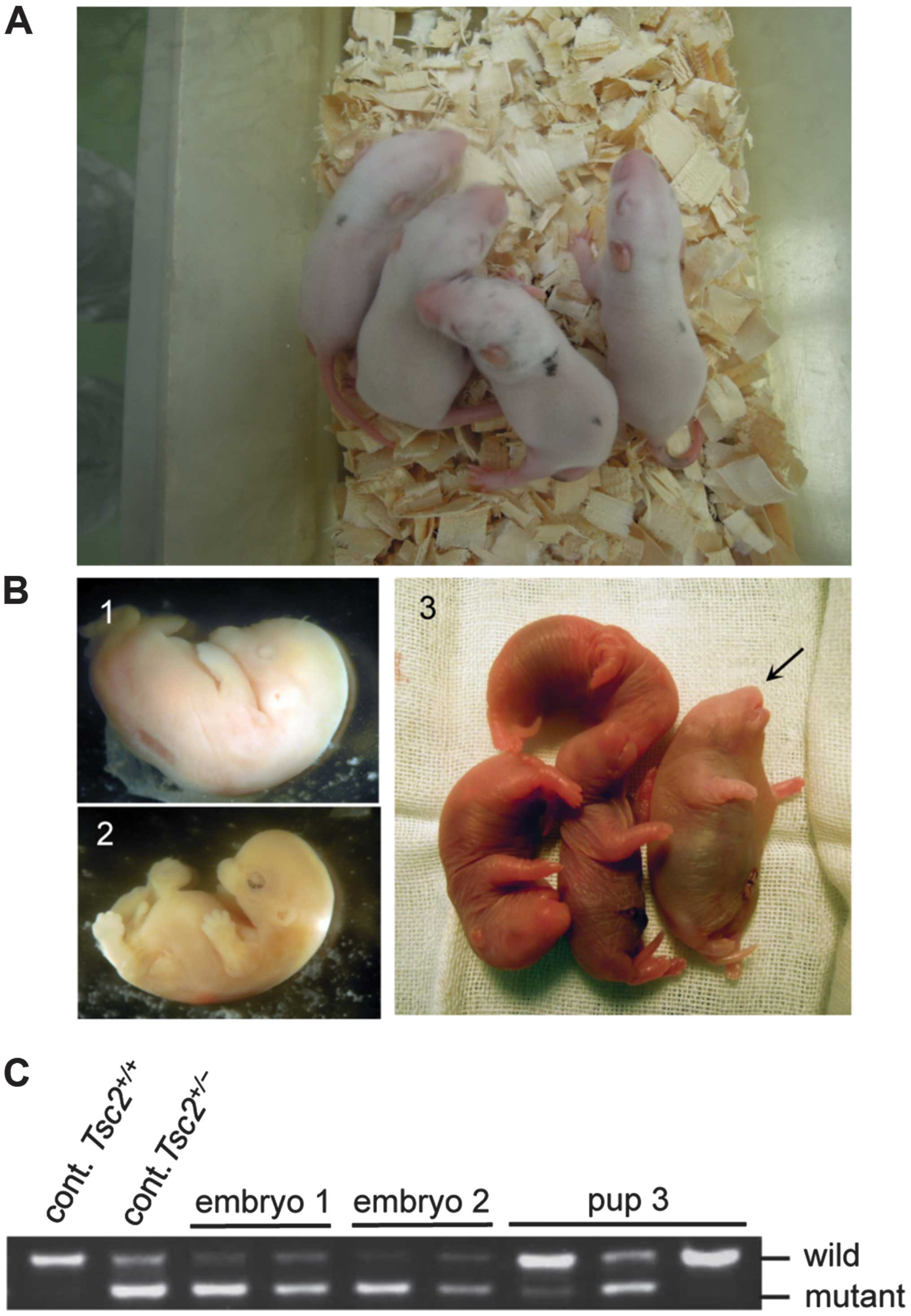
Dead embryos were collected from uteri by cesarean
section. For Tsc2 genotyping PCR, genomic DNA was obtained
from several parts of each embryo or pup.
Gene expression microarray analysis
The Rat Affymetrix GeneChip Gene 1.0 ST Array
(Affymetrix, Inc., Santa Clara, CA, USA) was used for microarray
analysis. Amplification and labeling of probes and hybridization
were performed according to the manufacturer’s instructions.
Hierarchical clustering analysis was performed using GeneSpring
software version 12.1 (Agilent Technologies, Inc., Santa Clara, CA,
USA).
Results
Establishment of Tsc2-deficient stem
cells from Eker rat embryos
After mating of double heterozygous Eker rats, a
total of 34 blastocysts were collected. Zonae pellucidae were
removed, and most blastocysts were successfully cultured on feeder
cells, revealing outgrowths from embryonic fibroblasts (MEFs) in
N2B27 medium supplemented with 2i + LIF (33). After several passages, a total of
26 cell lines were established. We routinely passaged these cells
every 2–4 days by dissociating them into single cells and replating
onto new feeder cells. They grew as dome-shaped or spherical
colonies and were maintained for >25 passages without losing
their morphology (Fig. 1A). A
majority of colonies expressed alkaline phosphatase, an indicator
of stem cell character (Fig. 1A).
Thereafter, we checked Tsc2 genotypes of established cell
lines by PCR. Surprisingly, we identified that not only
Tsc2+/+ and Tsc2+/− cell lines
but also Tsc2−/− cell lines had been established
(Fig. 1B). Considering that
previous reports had indicated that Tsc2 is necessary for
the maintenance of stem cell characteristics, this result was
unexpected. Both male and female cell lines were established for
each genotype (Fig. 1C).
Chromosome analysis revealed that most cell lines had normal ploidy
(n=42, data not shown). RT-PCR analysis revealed that
Tsc2−/− cells expressed the pluripotency markers
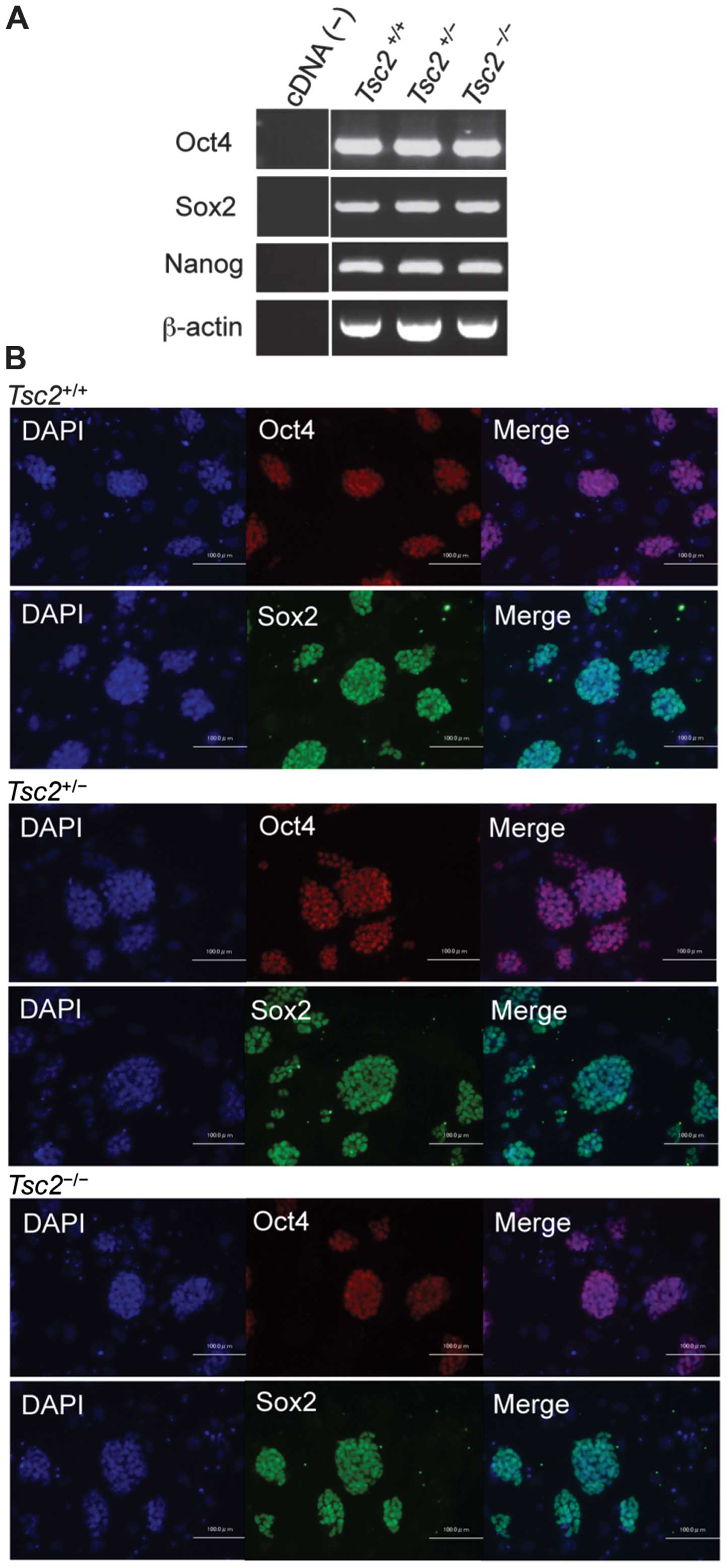
Oct4, Sox2, and Nanog (Fig. 2A). Oct4 and Sox2 expressions were
confirmed by immunofluorescence microscopy (Fig. 2B). These results indicate that
Tsc2-deficient stem cells could be established from Eker rat
embryos. We performed further experiments using two independent
cell lines of each genotype.
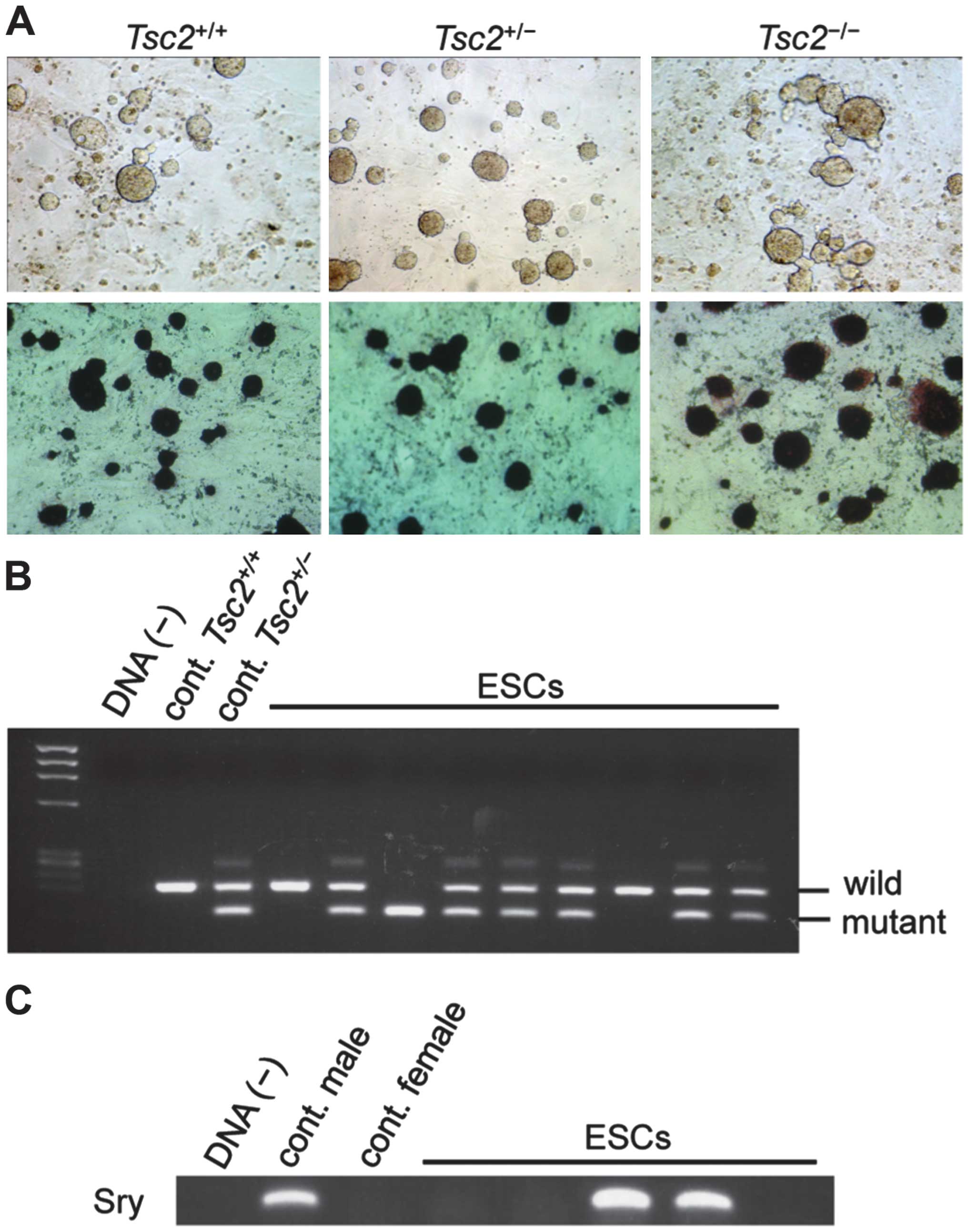 | Figure 1Establishment of
Tsc2-deficient cell lines from blastocysts of Eker rats. (A)
Colonies of established cell lines. Tsc2+/+,
Tsc2+/−, and Tsc2−/− represent
wild-type, Tsc2 heterozygous mutant, and Tsc2
homozygous mutant, respectively. Morphology of colonies established
from blastocysts of Eker rats cultured on mouse embryonic
fibroblasts (MEFs) in two inhibitors (2i) with leukemia inhibitory
factor (LIF) (upper panels). Alkaline phosphatase staining of
colonies (lower panels). Representative colonies are presented. (B)
Polymerase chain reaction (PCR) genotyping for the Tsc2
gene. Upper and lower bands represent wild-type and mutant
Tsc2 alleles, respectively. DNA(−), negative control; cont.
Tsc2+/+, wild-type rat; cont.
Tsc2+/−, Tsc2 heterozygous mutant rat.
Results of representative lines [embryonic stem cells (ESCs)] are
presented. (C) PCR of the Sry gene for gender determination
of established cell lines. DNA(−), negative control; cont. male,
male control; and cont. female, female control. Results of
representative lines (ESCs) are presented. |
Activation of the mTORC1 pathway in
Tsc2−/− ESCs
To evaluate the mTORC1 activation status, we
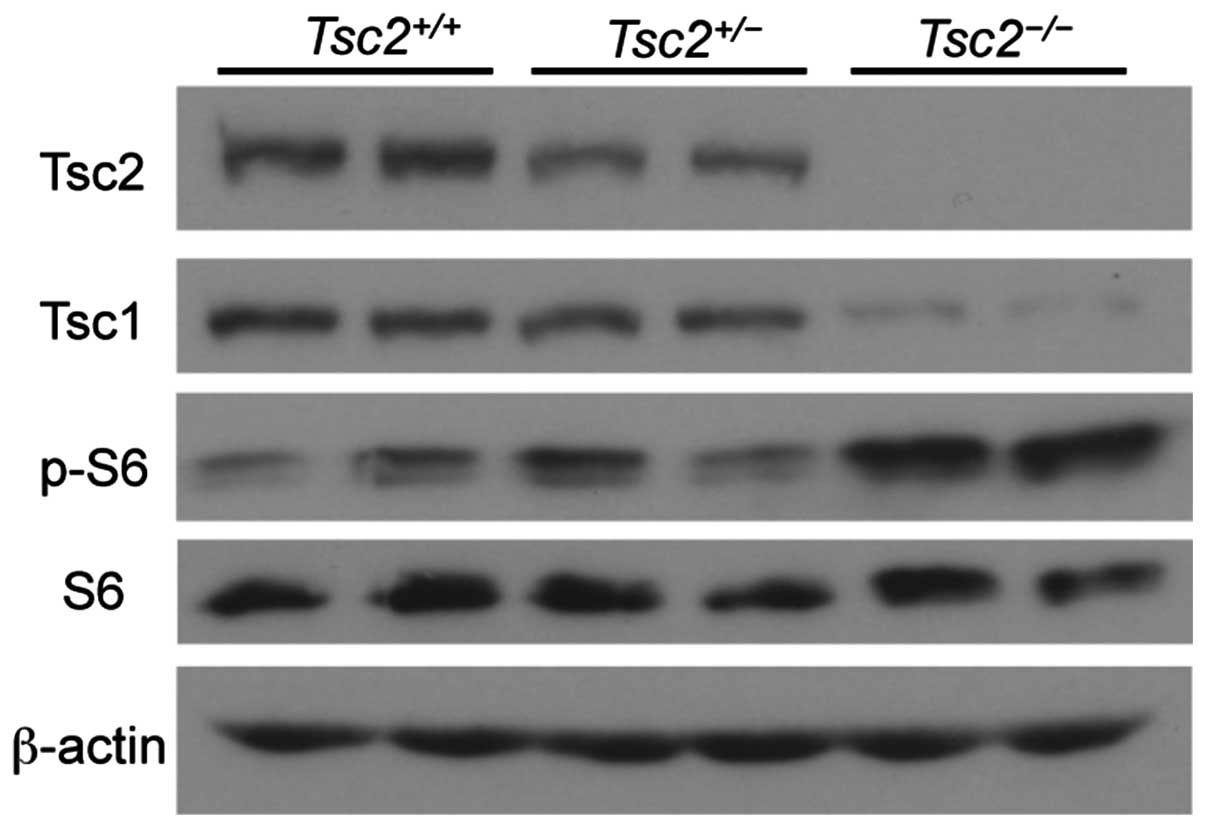
analyzed ESCs by western blotting. Tsc2 protein was detected in
Tsc2+/+ and Tsc2+/− cells but
not in Tsc2−/− cells, thereby confirming results
of the genotype analysis. Tsc1 protein levels were decreased in
Tsc2−/− ESCs, thereby reflecting the reciprocal
stabilization between Tsc1 and Tsc2 proteins (42). As expected, an increase in S6
phosphorylation was detected in Tsc2−/− cells
compared with that in Tsc2+/+ and
Tsc2+/− cells, which indicates abnormal
activation of the mTORC1 pathway in Tsc2−/− ESCs
(Fig. 3). These results indicate
that despite abnormal activation of the mTORC1 pathway,
Tsc2−/− ESCs can be established.
In vitro differentiation of
Tsc2−/− ESCs into three germ layers
Using the EB formation assay, we evaluated the
differentiation potential of the established cell lines. We
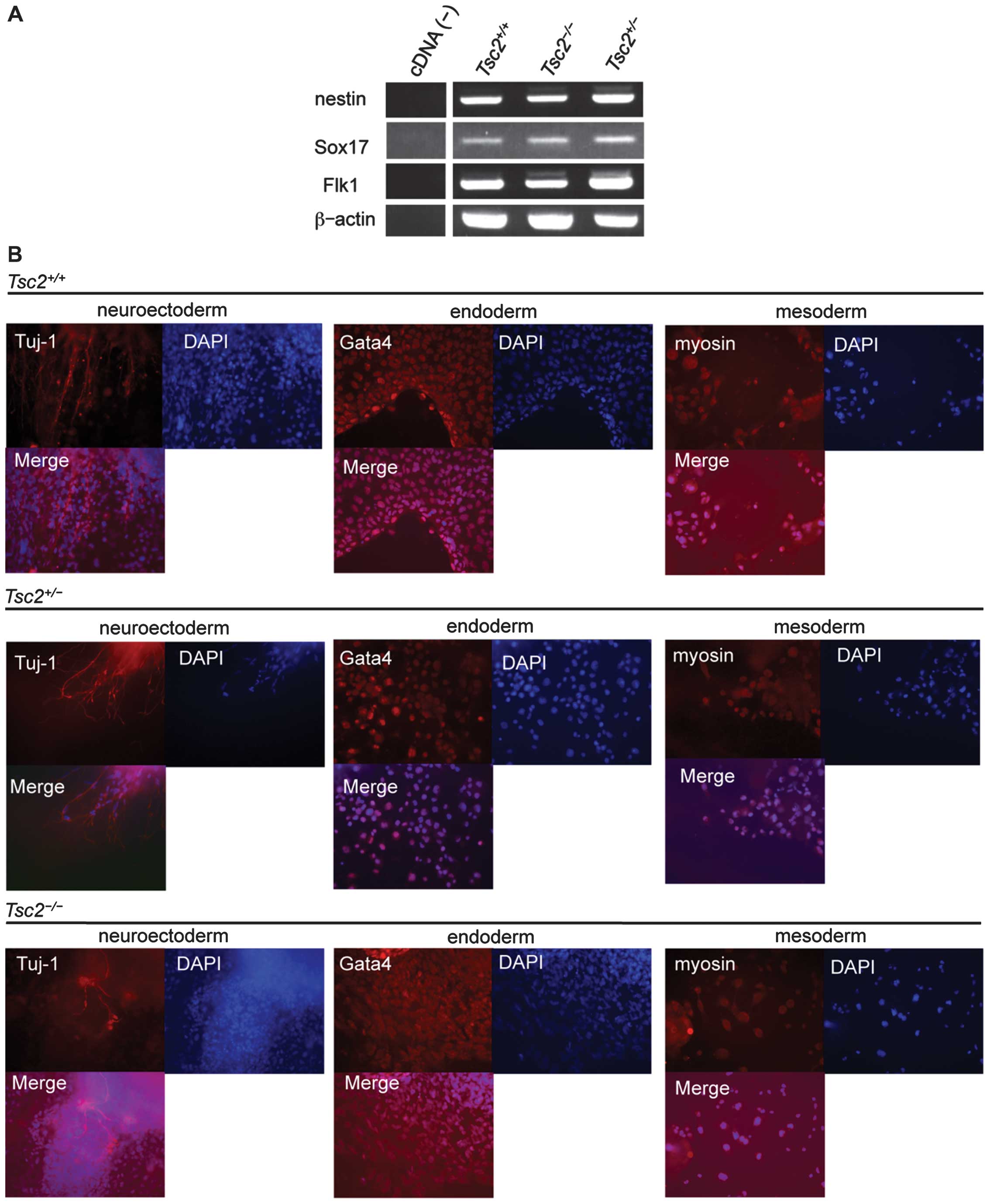
assessed the expression of differentiation markers by RT-PCR.
Expression of markers for ectoderm (Nestin), endoderm
(Sox17), and mesoderm (Flk1) were all observed in EBs
(Fig. 4A). We plated EBs onto
Matrigel-coated dishes and assessed their differentiation status by
immunofluorescent staining for β-III tubulin (neuroectoderm),
myosin (mesoderm), and Gata4 (endoderm) (Fig. 4B). Not only
Tsc2+/+ and Tsc2+/− cells but
also Tsc2−/− cells demonstrated the potential to
differentiate into all three germ layers. In addition, we observed
spontaneously beating areas in EBs of all Tsc2 genotypes
(data not shown). These results suggest that most differentiation
processes of ESCs were not blocked by Tsc2 deficiency.
Differentiation of Tsc2−/−
ESCs into multiple lineages in teratomas
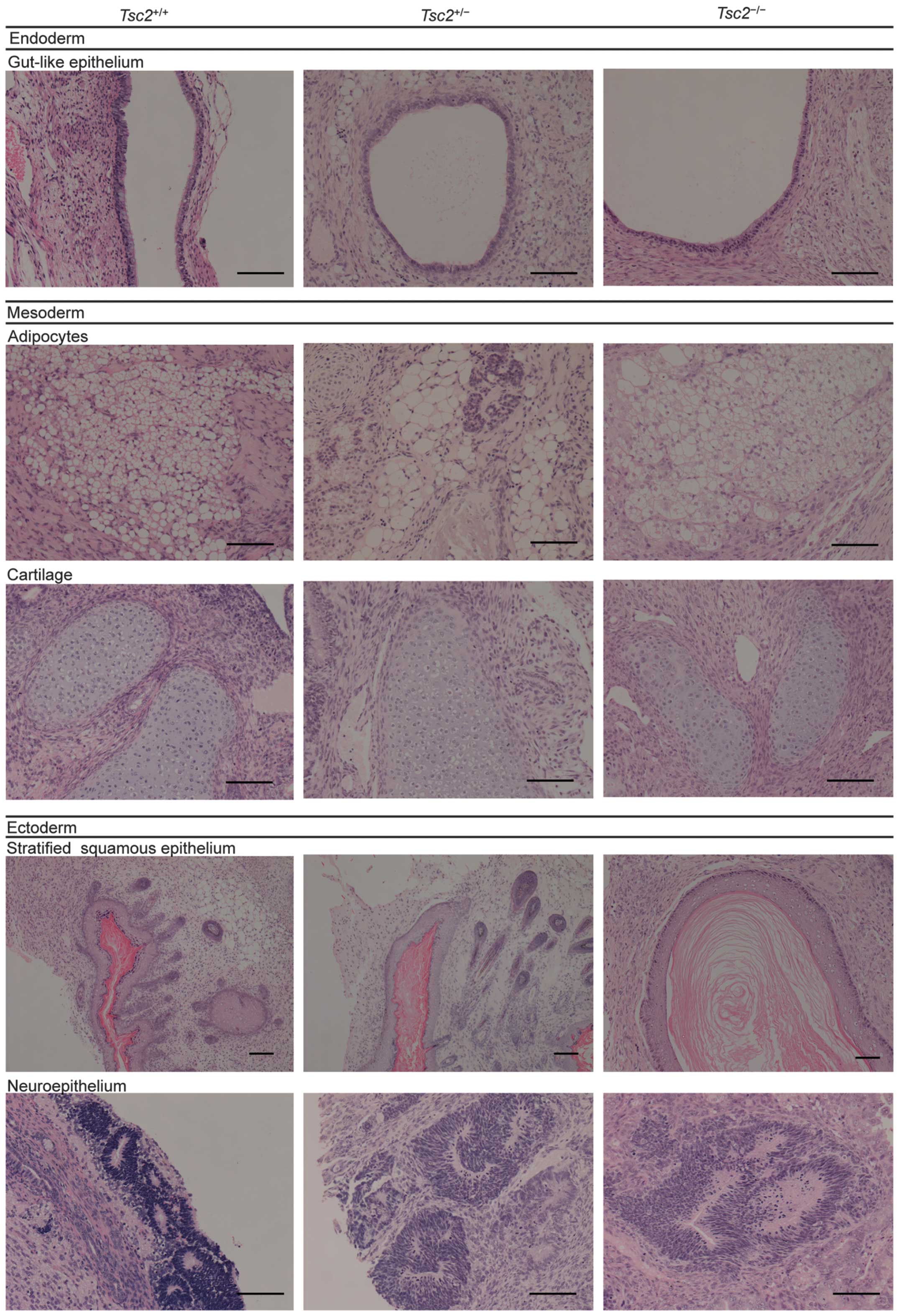
When Tsc2−/− ESCs were
transplanted under the kidney capsule of nude mice, they
differentiated into tissues derived from all three germ layers,
including gut-like epithelium (endoderm), cartilage and adipocytes
(mesoderm), stratified squamous epithelium, and neuroepithelium
(ectoderm) (Fig. 5). These results
indicate that Tsc2−/− ESCs are multipotent,
although detailed characterization of each of the differentiated
tissues remains to be elucidated. Interestingly, we observed that
abnormal ductal structures appeared in Tsc2−/−
teratomas (Kawano H, et al, unpublished data). Further
characterization of these abnormal structures is described in
another report (Kawano H, et al, unpublished data).
Contribution of Tsc2+/+ ESCs
in chimeras
Next, to determine the ability of established ESCs
to form chimeras, we injected Tsc2+/+ and
Tsc2−/− ESCs into blastocysts of Wistar rats or
Brown Norway rats (Materials and methods). Although ratios were
low, four chimeras with black coat color were born from Wistar
blastocysts injected with Tsc2+/+ ESCs,
indicating the contribution of ESCs from the Eker rat strain
(Fig. 6A). In contrast, we were
unable to obtain pups demonstrating chimeric coat color in repeated
trials using two independent Tsc2−/− ESCs.
However, in these trials, we detected dead embryos in the uterus of
recipient mother rats at term (Fig.
6B1 and 2). The appearance of dead embryos suggested
developmental retardation. In addition, one live pup was delivered
by cesarean section but died shortly after birth (Fig. 6B3). This pup revealed various
morphological abnormalities such as an enlarged trunk. PCR
genotyping of dead embryos and the pup indicated the contribution
of Tsc2−/− ESCs in their tissues (Fig. 6C). On the basis of the band pattern
of two dead embryos, we concluded that they had a greater
contribution of Tsc2−/− ESCs compared with the
live pup. These results suggest that a greater contribution of
Tsc2−/− ESCs in the chimera results in embryonic
lethality. Although germline transmission has not been confirmed
yet, the contribution in chimeras suggests that ESCs established in
this study possessed characteristics of multipotent stem cells.
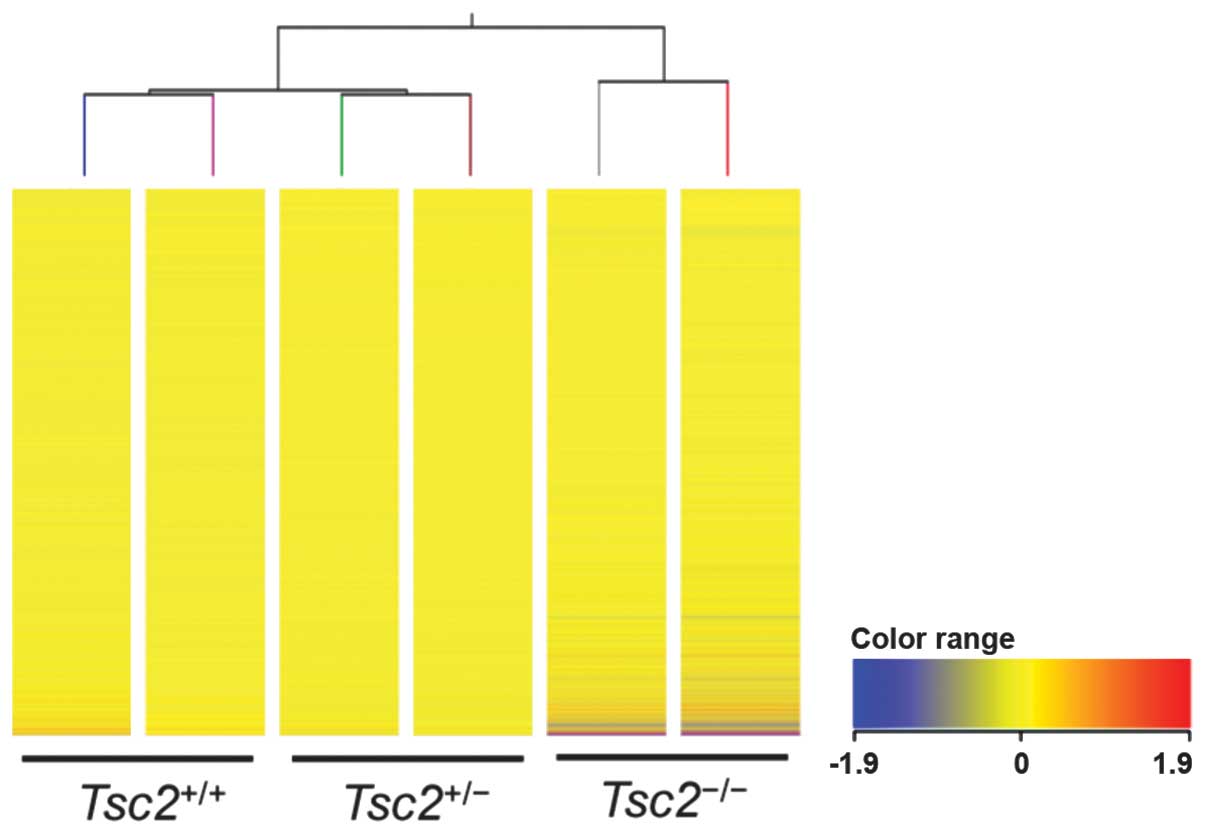
Distinct gene expression pattern in
Tsc2−/− ESCs on microarray analysis
To compare gene expression profiles of established
ESCs, we employed microarray analysis (Fig. 7). Similar expression levels of
pluripotency-related genes were identified in all these cells.
Moreover, hierarchical clustering analysis revealed that gene
expression profiles of Tsc2+/+ and
Tsc2+/− ESCs resembled each other, but those of
Tsc2−/− ESCs revealed an apparently distinct
pattern. These results suggest that the homozygous Tsc2
mutation causes extensive gene expression changes in rat ESCs.
Discussion
In this study, we successfully established
Tsc2−/− ESCs from Eker rats. These cells
possessed characteristic features of ESCs, including expression of
pluripotency markers, long-term self-renewal, and the capacity to
differentiate into derivatives of all three germ layers. Although
detailed mechanisms are still not clear, there have been several
reports indicating the importance of the Tsc2-mTOR pathway in stem
cell maintenance and differentiation (38,39,43).
Gan et al reported that Tsc1 is a critical regulator
of self-renewal, mobilization, and multilineage development in
hematopoietic stem cells and that it executes these phenomena via
both mTORC1-dependent and -independent pathways (39). Further, it was reported that the
activation of S6K by expression of the constitutively active S6K1
or siRNA-mediated knockdown of TSC2 and RICTOR
induced differentiation of human ESCs (38). Recently, Betschinger et al
reported that siRNA-mediated knockdown of Tsc2 or
Flcn inhibits differentiation of ESCs (43). In contrast to results in these
reports, we successfully established Tsc2−/− ESCs
possessing multipotent differentiation capacity despite the
presence of the activated mTORC1 pathway. There are several
possible reasons why the derivation of Tsc2−/−
ESCs was possible in this study. Previous studies utilized siRNA-
or shRNA-mediated knockdown of Tsc2 in already established
ESCs or conditional knockout of Tsc1 in somatic stem cells.
Such ‘acute’ downregulation of Tsc1/Tsc2 may cause
some aberrant gene regulation that restrains the maintenance of the
multipotent nature and differentiation capacity of stem cells.
Microarray analysis revealed a distinct gene expression pattern in
Tsc2−/− ESCs compared with their
Tsc2+/+ and Tsc2+/−
counterparts. Our system enables comparison of gene expression
profiles between Tsc2−/− ESCs and
Tsc2+/− ESCs with Tsc2 knockdown. Such
analysis is of interest to further explore Tsc2
mutation-related pathogenesis.
We were unable to obtain pups demonstrating
chimeric coat color using Tsc2−/− ESCs. Results
of dead embryos suggested that higher contribution of
Tsc2−/− cells in chimeras induced embryonic
lethality. Further, it has been reported that when human
TSC2-deficient fibroblast-like cells were grafted into mice,
differentiated tissues revealed features of TSC skin tumors and
that TSC2-deficient cells directly or indirectly induce
abnormal follicular neogenesis and epidermal proliferation
(44). Because
Tsc2−/− ESCs may cause abnormal differentiation
of hair in chimeras, it may not be appropriate to determine the
contribution of ESCs on the basis of hair color of chimeras.
He et al reported that reprogramming of
somatic cells derived from Tsc2−/− mouse embryos
to iPSCs was not possible (40).
In this study, we provide evidence that the derivation of
Tsc2−/− ESCs from Eker rat embryos is possible.
In somatic cells, some epigenetic abnormalities caused by
Tsc2 deficiency may not be corrected even under
reprogramming conditions. Conversely, during early embryogenesis,
epigenetic abnormalities in Tsc2−/− cells may be
tuned to maintain the stemness. With reprogramming experiments
using Eker rat-derived embryonic fibroblasts, ESCs established in
this study will serve as useful tools to compare effects of
Tsc2 deficiency on epigenetic status in reprogramming and
ESC derivation.
In recent years, various patient-derived iPSCs have
been used for in vitro differentiation experiments to mimic
the pathogenesis of human diseases (45,46).
Moreover, such cellular models are useful to research novel drug
target molecules by high-throughput screening (47). With regard to tumorigenesis, tissue
specificity and abnormal differentiation are relevant to its
molecular basis. Lineage-specific in vitro differentiation
of tumor suppressor-deficient ESCs will provide valuable
experimental models to explore the mechanism of pathogenesis.
However, in humans, establishment of tumor suppressor-deficient
(i.e., homozygously inactivated) ESCs or iPSCs has been technically
difficult. In rodents, homozygous mutant ESCs for tumor
suppressors, including Rb, Tp53, and Apc, have
been established (48–50). To date, none of those ESCs have
been extensively used for in vitro differentiation
experiments. For example, Apc-deficient ESCs failed to
differentiate into multiple lineages in the teratoma formation
assay, suggesting that the induction of various cell types was not
applicable to these ESCs (50). In
contrast, Tsc2−/− ESCs exhibited the potential to
differentiate into all germ layers and multiple cell lineages, both
in vitro and in vivo. We already observed development
of abnormal ductal structures in Tsc2−/−
teratomas, suggesting that cell type-specific effects of
Tsc2 deficiency could be reproduced in differentiation of
ESCs (Kawano H, et al, unpublished data). Combined with
in vivo experiments, in vitro differentiation models
using ESCs established in this study will facilitate understanding
of Tsc2 mutation-related pathogenesis as well as aid in the
search for therapeutic target pathways.
Acknowledgements
We thank Takako Ikegami, and Tomomi Ikeda,
Laboratory of Molecular and Biochemical Research, Research Support
Center, Juntendo University Graduate School of Medicine (Tokyo,
Japan) for technical assistance. The authors would like to thank
Enago (www.enago.jp) for the English language review.
This study was supported in part by the following grants:
Grants-in-Aid for Scientific Research from the Ministry of
Education, Culture, Sports, Science and Technology (MEXT) (Japan);
MEXT-Supported Program for the Strategic Research Foundation at
Private Universities; Grants-in-Aid for Scientific Research from
the Japan Society for the Promotion of Science (Japan); and
Grants-in-Aid for Scientific Research from the Ministry of Health,
Labour and Welfare (Japan). This study was also supported by the
Research Institute for Diseases of Old Age, Juntendo University
Graduate School of Medicine.
References
|
1
|
Kwiatkowski DJ, Whittemore VH and Thiele
EA: Tuberous Sclerosis Complex: Genes, Clinical Features, and
Therapeutics. Wiley-VCH Verlag GmbH & Co. KGaA; Weinheim: 2010,
View Article : Google Scholar
|
|
2
|
European Chromosome 16 Tuberous Sclerosis
Consortium. Identification and characterization of the tuberous
sclerosis gene on chromosome 16. Cell. 75:1305–1315. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
van Slegtenhorst M, de Hoogt R, Hermans C,
et al: Identification of the tuberous sclerosis gene TSC1 on
chromosome 9q34. Science. 277:805–808. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Curatolo P, Bombardieri R and Jozwiak S:
Tuberous sclerosis. Lancet. 372:657–668. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Laplante M and Sabatini DM: mTOR signaling
in growth control and disease. Cell. 149:274–293. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Knudson AG Jr: Mutation and cancer:
Statistical study of retino-blastoma. Proc Natl Acad Sci USA.
68:820–823. 1971. View Article : Google Scholar
|
|
7
|
Henske EP, Scheithauer BW, Short MP,
Wollmann R, Nahmias J, Hornigold N, van Slegtenhorst M, Welsh CT
and Kwiatkowski DJ: Allelic loss is frequent in tuberous sclerosis
kidney lesions but rare in brain lesions. Am J Hum Genet.
59:400–406. 1996.PubMed/NCBI
|
|
8
|
Au KS, Hebert AA, Roach ES and Northrup H:
Complete inactivation of the TSC2 gene leads to formation of
hamartomas. Am J Hum Genet. 65:1790–1795. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tucker T and Friedman JM: Pathogenesis of
hereditary tumors: Beyond the ‘two-hit’ hypothesis. Clin Genet.
62:345–357. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chan JA, Zhang H, Roberts PS, Jozwiak S,
Wieslawa G, Lewin-Kowalik J, Kotulska K and Kwiatkowski DJ:
Pathogenesis of tuberous sclerosis subependymal giant cell
astrocytomas: Biallelic inactivation of TSC1 or TSC2 leads to mTOR
activation. J Neuropathol Exp Neurol. 63:1236–1242. 2004.PubMed/NCBI
|
|
11
|
Niida Y, Stemmer-Rachamimov AO, Logrip M,
Tapon D, Perez R, Kwiatkowski DJ, Sims K, MacCollin M, Louis DN and
Ramesh V: Survey of somatic mutations in tuberous sclerosis complex
(TSC) hamartomas suggests different genetic mechanisms for
pathogenesis of TSC lesions. Am J Hum Genet. 69:493–503. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Onda H, Lueck A, Marks PW, Warren HB and
Kwiatkowski DJ: Tsc2(+/−) mice develop tumors in multiple sites
that express gelsolin and are influenced by genetic background. J
Clin Invest. 104:687–695. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kobayashi T, Minowa O, Kuno J, Mitani H,
Hino O and Noda T: Renal carcinogenesis, hepatic hemangiomatosis,
and embryonic lethality caused by a germ-line Tsc2 mutation in
mice. Cancer Res. 59:1206–1211. 1999.PubMed/NCBI
|
|
14
|
Kobayashi T, Minowa O, Sugitani Y, Takai
S, Mitani H, Kobayashi E, Noda T and Hino O: A germ-line Tsc1
mutation causes tumor development and embryonic lethality that are
similar, but not identical to, those caused by Tsc2 mutation in
mice. Proc Natl Acad Sci USA. 98:8762–8767. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Meikle L, Pollizzi K, Egnor A, Kramvis I,
Lane H, Sahin M and Kwiatkowski DJ: Response of a neuronal model of
tuberous sclerosis to mammalian target of rapamycin (mTOR)
inhibitors: Effects on mTORC1 and Akt signaling lead to improved
survival and function. J Neurosci. 28:5422–5432. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Goorden SMI, van Woerden GM, van der Weerd
L, Cheadle JP and Elgersma Y: Cognitive deficits in
Tsc1+/− mice in the absence of cerebral lesions and
seizures. Ann Neurol. 62:648–655. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ehninger D, Han S, Shilyansky C, Zhou Y,
Li W, Kwiatkowski DJ, Ramesh V and Silva AJ: Reversal of learning
deficits in a Tsc2+/− mouse model of tuberous sclerosis.
Nat Med. 14:843–848. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Eker R and Mossige J: A dominant gene for
renal adenomas in the rat. Nature. 189:858–859. 1961. View Article : Google Scholar
|
|
19
|
Hino O, Mitani H and Knudson AG: Genetic
predisposition to transplacentally induced renal cell carcinomas in
the Eker rat. Cancer Res. 53:5856–5858. 1993.PubMed/NCBI
|
|
20
|
Kobayashi T, Hirayama Y, Kobayashi E, Kubo
Y and Hino O: A germline insertion in the tuberous sclerosis (Tsc2)
gene gives rise to the Eker rat model of dominantly inherited
cancer. Nat Genet. 9:70–74. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Rennebeck G, Kleymenova EV, Anderson R,
Yeung RS, Artzt K and Walker CL: Loss of function of the tuberous
sclerosis 2 tumor suppressor gene results in embryonic lethality
characterized by disrupted neuroepithelial growth and development.
Proc Natl Acad Sci USA. 95:15629–15634. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mizuguchi M, Takashima S, Yamanouchi H,
Nakazato Y, Mitani H and Hino O: Novel cerebral lesions in the Eker
rat model of tuberous sclerosis: Cortical tuber and anaplastic
ganglioglioma. J Neuropathol Exp Neurol. 59:188–196.
2000.PubMed/NCBI
|
|
23
|
Yeung RS, Katsetos CD and Klein-Szanto A:
Subependymal astrocytic hamartomas in the Eker rat model of
tuberous sclerosis. Am J Pathol. 151:1477–1486. 1997.PubMed/NCBI
|
|
24
|
Takahashi DK, Dinday MT, Barbaro NM and
Baraban SC: Abnormal cortical cells and astrocytomas in the Eker
rat model of tuberous sclerosis complex. Epilepsia. 45:1525–1530.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Prabhakar S, Goto J, Zhang X, et al:
Stochastic model of Tsc1 lesions in mouse brain. PLoS One.
8:e642242013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kobayashi T, Adachi H, Mitani H, Hirayama
Y and Hino O: Toward chemotherapy for Tsc2-mutant renal tumor. Proc
Jpn Acad. 79:22–25. 2003. View Article : Google Scholar
|
|
27
|
Kenerson HL, Aicher LD, True LD and Yeung
RS: Activated mammalian target of rapamycin pathway in the
pathogenesis of tuberous sclerosis complex renal tumors. Cancer
Res. 62:5645–5650. 2002.PubMed/NCBI
|
|
28
|
Lee L, Sudentas P, Donohue B, et al:
Efficacy of a rapamycin analog (CCI-779) and IFN-γ in tuberous
sclerosis mouse models. Genes Chromosomes Cancer. 42:213–227. 2005.
View Article : Google Scholar
|
|
29
|
McCormack FX, Inoue Y, Moss J, et al:
National Institutes of Health Rare Lung Diseases Consortium; MILES
Trial Group: Efficacy and safety of sirolimus in
lymphangioleiomyomatosis. N Engl J Med. 364:1595–1606. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Franz DN, Belousova E, Sparagana S, et al:
Efficacy and safety of everolimus for subependymal giant cell
astrocytomas associated with tuberous sclerosis complex (EXIST-1):
A multicentre, randomised, placebo-controlled phase 3 trial.
Lancet. 381:125–132. 2013. View Article : Google Scholar
|
|
31
|
Bissler JJ, Kingswood JC, Radzikowska E,
et al: Everolimus for angiomyolipoma associated with tuberous
sclerosis complex or sporadic lymphangioleiomyomatosis (EXIST-2): A
multicentre, randomised, double-blind, placebo-controlled trial.
Lancet. 381:817–824. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Mizuguchi M: Abnormal giant cells in the
cerebral lesions of tuberous sclerosis complex. Congenit Anom
(Kyoto). 47:2–8. 2007. View Article : Google Scholar
|
|
33
|
Buehr M, Meek S, Blair K, Yang J, Ure J,
Silva J, McLay R, Hall J, Ying QL and Smith A: Capture of authentic
embryonic stem cells from rat blastocysts. Cell. 135:1287–1298.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li P, Tong C, Mehrian-Shai R, et al:
Germline competent embryonic stem cells derived from rat
blastocysts. Cell. 135:1299–1310. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Evans MJ and Kaufman MH: Establishment in
culture of pluripotential cells from mouse embryos. Nature.
292:154–156. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Martin GR: Isolation of a pluripotent cell
line from early mouse embryos cultured in medium conditioned by
teratocarcinoma stem cells. Proc Natl Acad Sci USA. 78:7634–7638.
1981. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Thomson JA, Itskovitz-Eldor J, Shapiro SS,
Waknitz MA, Swiergiel JJ, Marshall VS and Jones JM: Embryonic stem
cell lines derived from human blastocysts. Science. 282:1145–1147.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Easley CA IV, Ben-Yehudah A, Redinger CJ,
Oliver SL, Varum ST, Eisinger VM, Carlisle DL, Donovan PJ and
Schatten GP: mTOR-mediated activation of p70 S6K induces
differentiation of pluripotent human embryonic stem cells. Cell
Reprogram. 12:263–273. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Gan B, Sahin E, Jiang S, Sanchez-Aguilera
A, Scott KL, Chin L, Williams DA, Kwiatkowski DJ and DePinho RA:
mTORC1-dependent and -independent regulation of stem cell renewal,
differentiation, and mobilization. Proc Natl Acad Sci USA.
105:19384–19389. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
He J, Kang L, Wu T, et al: An elaborate
regulation of Mammalian target of rapamycin activity is required
for somatic cell reprogramming induced by defined transcription
factors. Stem Cells Dev. 21:2630–2641. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Shiono M, Kobayashi T, Takahashi R, et al:
The G1556S-type tuberin variant suppresses tumor formation in
tuberous sclerosis 2 mutant (Eker) rats despite its deficiency in
mTOR inhibition. Oncogene. 27:6690–6697. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Benvenuto G, Li S, Brown SJ, Braverman R,
Vass WC, Cheadle JP, Halley DJ, Sampson JR, Wienecke R and DeClue
JE: The tuberous sclerosis-1 (TSC1) gene product hamartin
suppresses cell growth and augments the expression of the TSC2
product tuberin by inhibiting its ubiquitination. Oncogene.
19:6306–6316. 2000. View Article : Google Scholar
|
|
43
|
Betschinger J, Nichols J, Dietmann S,
Corrin PD, Paddison PJ and Smith A: Exit from pluripotency is gated
by intracellular redistribution of the bHLH transcription factor
Tfe3. Cell. 153:335–347. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Li S, Thangapazham RL, Wang JA, Rajesh S,
Kao TC, Sperling L, Moss J and Darling TN: Human TSC2-null
fibroblast-like cells induce hair follicle neogenesis and hamartoma
morphogenesis. Nat Commun. 2:2352011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ebert AD, Yu J, Rose FF Jr, Mattis VB,
Lorson CL, Thomson JA and Svendsen CN: Induced pluripotent stem
cells from a spinal muscular atrophy patient. Nature. 457:277–280.
2009. View Article : Google Scholar :
|
|
46
|
Lee G, Papapetrou EP, Kim H, et al:
Modelling pathogenesis and treatment of familial dysautonomia using
patient-specific iPSCs. Nature. 461:402–406. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yang YM, Gupta SK, Kim KJ, et al: A small
molecule screen in stem-cell-derived motor neurons identifies a
kinase inhibitor as a candidate therapeutic for ALS. Cell Stem
Cell. 12:713–726. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Williams BO, Schmitt EM, Remington L,
Bronson RT, Albert DM, Weinberg RA and Jacks T: Extensive
contribution of Rb-deficient cells to adult chimeric mice with
limited histopathological consequences. EMBO J. 13:4251–4259.
1994.PubMed/NCBI
|
|
49
|
Kawamata M and Ochiya T: Two distinct
knockout approaches highlight a critical role for p53 in rat
development. Sci Rep. 2:9452012. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Kielman MF, Rindapää M, Gaspar C, van
Poppel N, Breukel C, van Leeuwen S, Taketo MM, Roberts S, Smits R
and Fodde R: Apc modulates embryonic stem-cell differentiation by
controlling the dosage of beta-catenin signaling. Nat Genet.
32:594–605. 2002. View
Article : Google Scholar : PubMed/NCBI
|