Introduction
Neuroblastoma (NB) is a common childhood malignant
tumor of neural crest origin, generated in the paravertebral
sympathetic ganglia and the adrenal medulla. NB is also the most
common extracranial solid tumor in children (>7% of tumors in
patients younger than 15 years) and a major cause of neoplastic
death in infancy (15% of deaths in pediatric oncology) (1). Current surgery and radiotherapy in
conjunction with chemotherapy has greatly improved survival rates
for the patients with low-risk and intermediate-risk NB. However,
high-risk patients still have an overall survival rate of <40%
despite intensive therapy (2).
Thus, it is urgent to develop new drugs for the treatment of
high-risk NB.
Phenanthroindolizidine and phenanthroquinolizine
alkaloids are family of plant-derived alkaloids composed of more
than sixty compounds (3). The
biological properties of these alkaloids include cancer cell growth
inhibition (in vitro and in vivo), anti-inflammatory
activity, anti-ameobicidal and anti-viral activity (4–6). Our
previous studies show that (+)-13a-(S)-Deoxytylophorinidine (CAT)
(Fig. 1A) had potent antitumor
activity both in vivo and in vitro (7,8).
Furthermore, 3-O-Desmethyl-13a-(S)-deoxytylophorinine (PF403)
(Fig. 1B), one of CAT in
vivo metabolites, showed stronger proliferation inhibitory
activity than CAT on several tumor cell lines (9).
In the present study, we investigated the
anti-neuroblastoma potential of PF403 in human NB cell line and in
an in vivo NB mouse model. Since our previous studies on
pharmacokinetic had proved poor bioavailability of PF403, the
pro-drug of PF403, CAT3, was chosen to study
anti-neuroblastoma activity in vivo (8). Both MRI and pathology analysis
verified the anti-neuroblastoma ability of PF403 in situ.
Then mechanisms of PF403 on anti-neuroblastoma were studied.
Collectively, our results for the first time addressed multiple
mechanisms associated with PF403-mediated neuroblastoma cell death
and suggested the possible application of PF403 for neuroblastoma
therapy.
Materials and methods
Cells and chemicals
SH-SY5Y cell line was obtained from the cell center
of CAMS&PUMC. The cells were maintained in Dulbecco’s modified
Eagle’s medium (DMEM; Invitrogen, Carlsbad, CA, USA) supplemented
with 10% fetal bovine serum (FBS; Gibco, Waltham, MA, USA), 100
IU/ml penicillin and 100 μg/ml streptomycin (Invitrogen) in a
humidified incubator containing 5% CO2 at 37°C. PF403
and CAT3 were obtained from the research group of
Professor Shi-shan Yu (Institute of Materia Medica, China).
Cell viability assay
SH-SY5Y cells were seeded in a 96-well plate. After
24 h, the cells were treated with different concentrations of
PF403. After 48, 72 and 96 h of incubation, cells were incubated
with 1 mg/ml of MTT (Sigma, St. Louis, MO, USA) for 4 h at 37°C in
a CO2 incubator. Mitochondrial reduction of MTT to
formazan was determined. Amount of formazan was measured by
absorbance at 570 nm with reference wavelength at 450 nm.
IC50 values were calculated and expressed as mean ±
standard deviation of three independent experiments.
Colony formation assay
The cells were seeded at a density of 200 cells/ml
into 6-well culture plates for 24 h, then washed with
phosphate-buffered saline (PBS) and cultured by DMEM with a series
concentration of PF403 added. Colonies were allowed to grow for 14
days. After removing the medium, each well was carefully washed
twice with PBS. The cells were fixed in methanol for 15 min and
then stained with crystal violet for 20 min. Finally, positive
colony formation (>50 cells per colony) was counted.
Flow cytometry assay
Cells were treated with a different of
concentrations of PF403. After 72 h of treatment, cells were fixed
with 70% ethanol, stained with propidium iodide (PI), and were
analyzed by flow cytometry. The data were analyzed with CellQuest
Pro software (BD Biosciences, San Jose, CA, USA). The experiments
were repeated at least three times.
Invasion assay
Matrigel (10 μl) (BD Biosciences), a reconstituted
basement membrane, was pre-coated on the upper compartment of
Millicell (Millipore, Billerica, MA, USA) containing 8-μm pores.
Approximately 600 μl of DMDM with 10% FBS was added to the lower
compartment; 200 μl of the completed medium containing
2×105 SH-SY5Y cells were seeded to the upper
compartment. PF403 was added to the upper compartment at a series
of concentrations. After culturing for 24 h at 37°C in a humidified
5% CO2, cells migrated to the lower compartment were
stained and counted under the microscope.
Pharmacokinetic analyses
Animals (Vital River Laboratories, Beijing, China)
were randomly assigned to ten groups (n=3/group) that were given a
single dose of CAT3 (10 mg/kg) orally. Mice were
decapitated following 5, 15, 30 min, 1, 2, 4, 6, 8, 12 and 24 h.
Brain structures were dissected on dry ice. Brain tissue homogenate
with PF403 (5, 10, 25, 100, 250, 500, 1,000 and 2,500 ng/ml) of
blank group was used as standard solution. High performance liquid
chromatography (HPLC) system Agilent 1100 Series (Agilent
Technologies, Santa Clara, CA, USA), equipped with a binary pump,
an autosampler and degasser. Separation was performed using a
Zorbax C18 column (2.1×100 mm, 3.5 μm). The column was thermostated
at 25°C. The optimized mobile phase solvents were: 0.1% formic
acid/water =35/65. CAT3 and its metabolites were
separated at a flow rate of 0.2 ml/min with a linear gradient. Mass
spectral data were obtained in positive electrospray mode. High
purity nitrogen used as a sheath gas, was generated with a nitrogen
generator. The m/z of PF403 was 350.1-281. Levels of
CAT3 and its metabolites were calculated using the
calibration standard curves, constructed by linear regression
analysis of peak area vs. concentration curves. CAT3 and
PF403 (2 μl) incubated with hepatic microsome, plasma, artificial
intestinal or artificial gastric juice at 37°C for 15 or 20 min.
LC/MS analyzed concentration of CAT3 and PF403.
Animals and surgical technique
Five-week-old female Balb/c nude (nu/nu) mice (Vital
River Laboratories) were housed in standard facilities and given
free access to water and rodent food. All animal experiments in the
present study were conducted according to the guidelines of the Law
for The Care and Welfare of Animals in China and approved by the
Animal Experiment Committee of Institute of Materia Medica. For
intracranial injection, mice were anesthetized with pentobarbital
sodium (i.p) in a dosage of 50 mg/kg, placed in a stereotaxic
restrain, and a small surgical incision was made in the skin
covering the skull 2 mm to the right of the bregma. Cell
suspensions (5 μl) containing ~5×105 SH-SY5Y cells were
slowly (~30 sec) injected intracranially at 2.5 mm below the skull
surface using a 26-gauge needle. Immediately thereafter the
incision was closed with surgical adhesive and the animals were
monitored until conscious and returned to their cages. Animals were
randomly assigned to control, TMZ treated group (120 mg/kg/d),
CAT3 low-dose group (6 mg/kg/d) and CAT3
high-dose group (12 mg/kg/d) (n=3). From the 15th day of the
operation, mice in CAT3 low-dose groups and
CAT3 high-dose groups were treated of CAT3
orally; and mice in TMZ group were treated of TMZ orally.
Imaging analysis
All mice experienced high resolution MR (Bruker,
Bremen, Germany) scanning weekly after operation. The MR images
(Siemens, Munich, Germany) were acquired using a 1.5 tesla clinical
unit. The slice thickness for coronal and transverse scan was 0.5
mm. The mice were anesthetized with intraperitoneal injection of
pentobarbital sodium solution in a dosage of 50 mg/kg. After
coronal and transverse T2 weighted images had been obtained,
coronal plane enhanced T1 weighted images were acquired by an
intravenous injection of dimeglumine gadopentetate (Magnevist,
Bayer Schering Parma, Germany) at a dose of 2 ml/kg body weight via
caudal vein.
Histopathological analysis
Paraffin-embedded mouse whole brain. The thickness
of sections was 5 μm. Serial sections were also stained with
H&E. The stained images of each section were acquired using a
microscope (Nikon, Tokyo, Japan). H&E staining was used in
diagnosis, and analysis of the pathology changes.
Western blot analysis
For western blotting, samples transferred to a
nitrocellulose membrane by semi-wet electrophoresis were subjected
to SDS-PAGE and incubated with primary antibody rabbit anti-FAK,
rabbit anti-phosphorylated FAK, rabbit anti-MMP-2, rabbit
anti-MMP-9, rabbit anti-PI3K p85, rabbit anti-phosphorylated PI3K
p85, rabbit anti-AKT, rabbit anti-phosphorylated AKT, rabbit
anti-mTOR, rabbit anti-phosphorylated mTOR, rabbit anti-c-Raf,
rabbit anti-phosphorylated c-Raf, rabbit anti-MEK, rabbit
anti-phosphorylated MEK, rabbit anti-ERK, rabbit
anti-phosphorylated ERK1/2, rabbit anti-Mcl1, rabbit anti-Bcl-2,
rabbit anti-BAX, mouse anti-Bad, mouse anti-Bid, mouse anti-Bim,
rabbit anticaspase-9, rabbit anti-cleave caspase-9, rabbit
anti-caspase-3, rabbit anti-cleave caspase-3, rabbit anti-PARP and
rabbit anti-cleave PARP antibodies (Cell Signaling Technology,
Danvers, MA, USA) overnight at 4°C. The samples were detected with
horseradish peroxidase-conjugated anti-rabbit or anti-mouse IgG
(Cell Signaling Technology) and developed using an ECL. Membranes
were tested for equal loading by probing for actin.
Statistical analysis
The results were expressed as means ± SD for
repeated measures, and t-test was used for comparison of
differences between groups. Differences were considered
statistically significant when P≤0.05.
Results
PF403 induces cytotoxicity in SH-SY5Y
neuroblastoma cells in a dose- and time-dependent manner
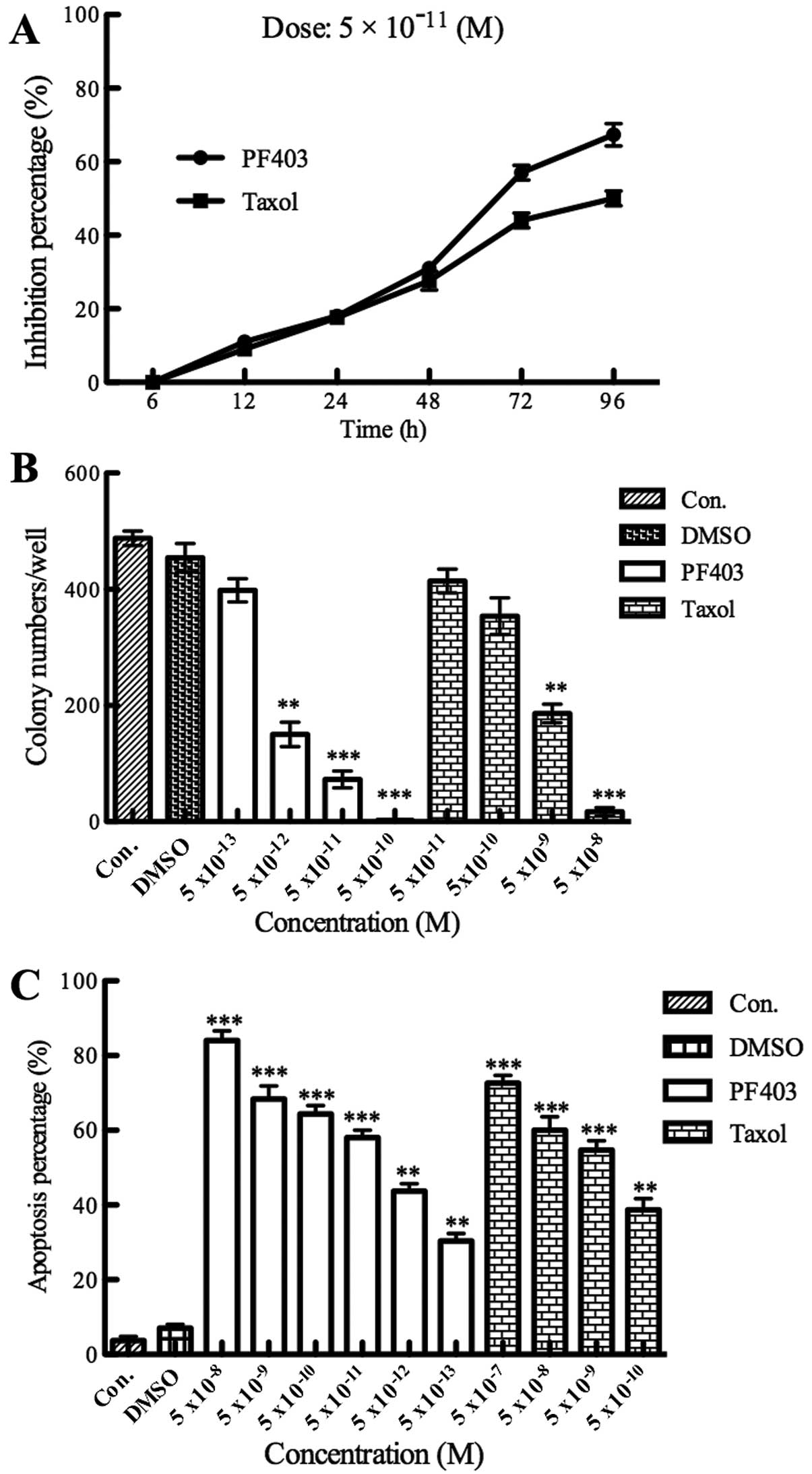
The MTT assay, colony forming assay and flow
cytometry assay were chosen to test the cytotoxicity of PF403 in
NB. Administration with different concentrations of PF403
(5×10−5–5×10−13 M) for 48 h, 72 and 96 h,
cell viability was determined using MTT assay. The viability of the
untreated cells was regarded as 100%. The results revealed that
PF403 could inhibit SH-SY5Y cell proliferation significantly from
48 to 96 h, and its potency was higher than taxol (Table I). The time-dependent curve of MTT
assay demonstrated the concentration of PF403 at 5×10−11
M, its proliferation inhibitory ability in SH-SY5Y cells reached
stable stage after 72 h, and PF403 had stronger antitumor
proliferation ability than taxol after 48 h (Fig. 2A). The results of PF403 on
proliferation were confirmed by the colony formation assay, the
IC50 of PF403 was 3.81±1.29×10−13 M, and the
IC50 of taxol was 4.77±1.29×10−9 M (Fig. 2B). The results of flow cytometry
displayed that the apoptotic cells were increased dramatically in
PF403 treated groups compared with control and DMSO groups, the
IC50 was at 10−11 M (Fig. 2C). These results preliminarily
demonstrated that PF403 had a strong anti-NB activity in
vitro.
 | Table IAnti-proliferative activity of PF403
on SHSY-5Y cells by MTT assay. |
Table I
Anti-proliferative activity of PF403
on SHSY-5Y cells by MTT assay.
| IC50
(M) |
|---|
|
|
|---|
| Time (h) | PF403 | Taxol |
|---|
| 48 |
5.25±1.41×10−6 |
1.57±0.42×10−4 |
| 72 |
4.21±1.23×10−11 |
5.16±2.24×10−8 |
| 96 |
2.84±1.13×10−12 |
2.61±1.43×10−8 |
PF403 inhibits invasion of SH-SY5Y
cells
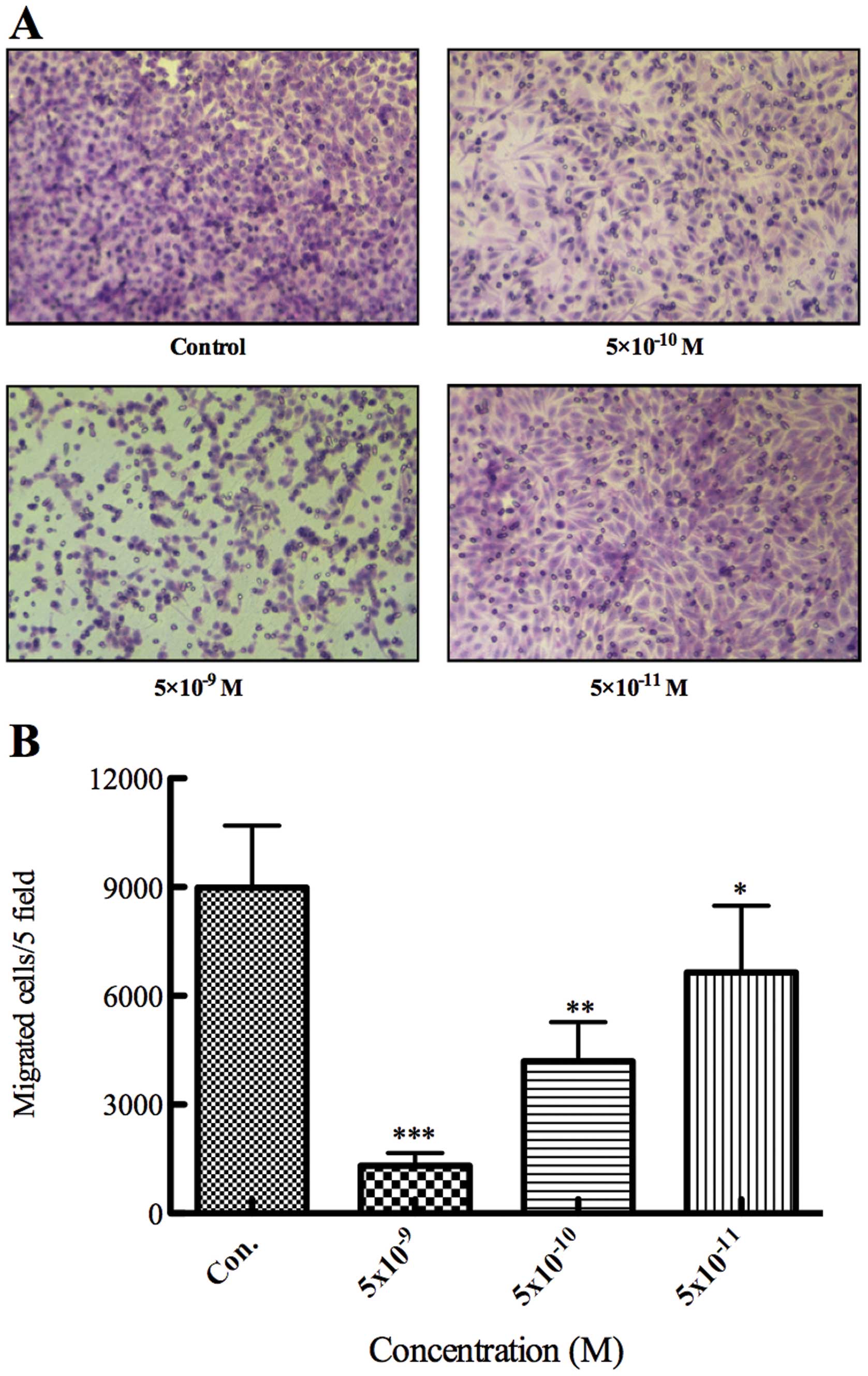
To investigate the effect of PF403 on invasion in
SH-SY5Y cells, we treated the SH-SY5Y NB cancer cells with various
concentrations of PF403 for 24 h and observed cell invasion by
Transwell assay. The results indicated that the dose required for
inhibition of SH-SY5Y cell invasion was much less than the dose
needed to cause apoptosis (IC50 value at 10−3
M, at 24 h) (Table II and
Fig. 3). These results suggested
that PF403 was able to affect invasion ability of SH-SY5Y
neuroblastoma cells at a low concentration.
| Table IIPF403 affected the invasion capacity
in SH-SY5Y cells. |
Table II
PF403 affected the invasion capacity
in SH-SY5Y cells.
| Groups | Concentration
(M) | Cell count | Inhibition rate
(%) |
|---|
| Control | - | 8981±1710 | - |
| PF403 |
5×10−9 | 1313±355a | 83.36 |
|
5×10−10 | 4192±1087a | 53.34 |
|
5×10−11 | 6648±1836b | 26.06 |
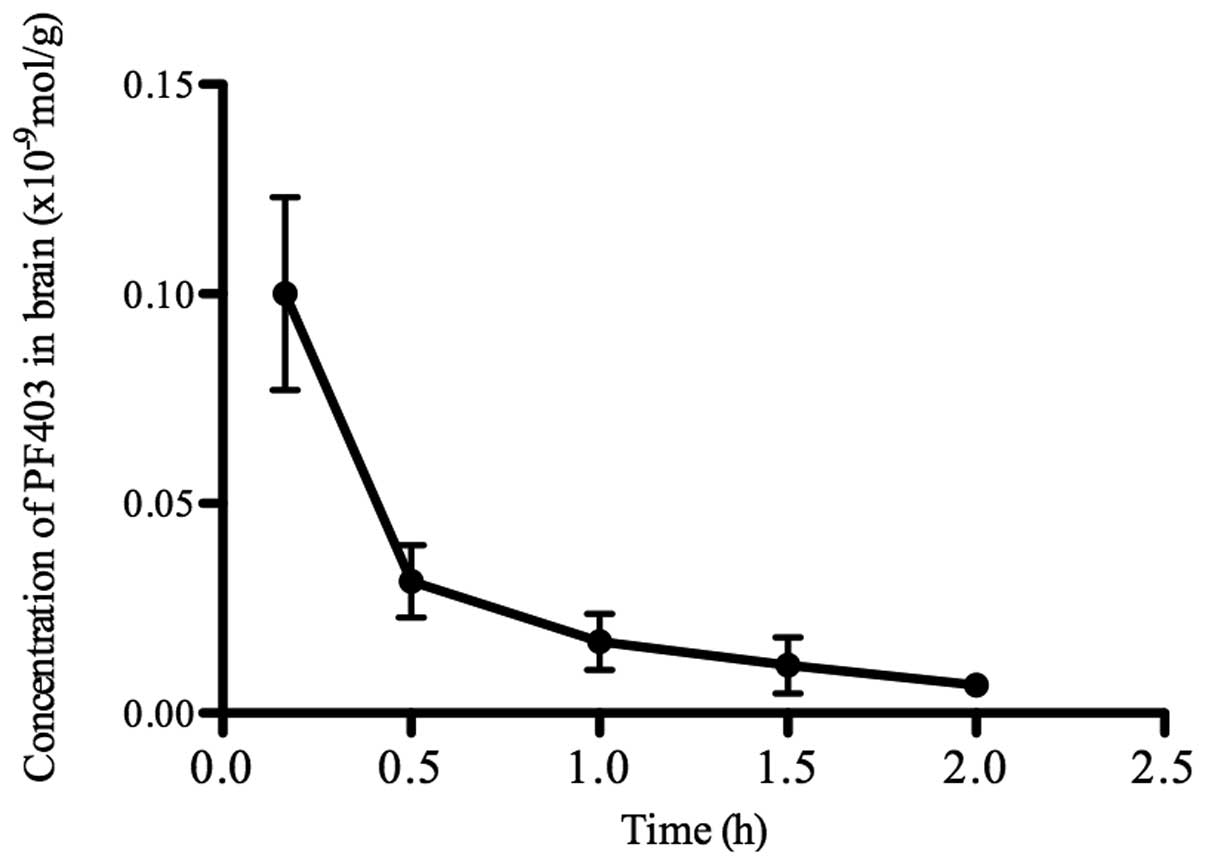
CAT3 metabolizes into PF403 in
vivo
There was no significant change in the concentration
of CAT3 and PF403 incubated with artificial gastric
juice for 15 min. However, the concentrations of CAT3
were reduced, and PF403 could be detected while incubated with
hepatic microsome, plasma or artificial intestinal fluid for 15 or
20 min. CAT3 (59%) metabolized into PF403 after
incubated with artificial intestinal fluid for 15 min. PF403 was
detected in brain tissue after CAT3 oral administration
for 5 min (Fig. 4). These results
demonstrated that CAT3 metabolized into PF403 in
vivo, and PF403 was proved to penetrate the blood-brain
barrier.
The pro-drug of PF403 and
CAT3, has a potent anti-neuroblastoma activity in
situ
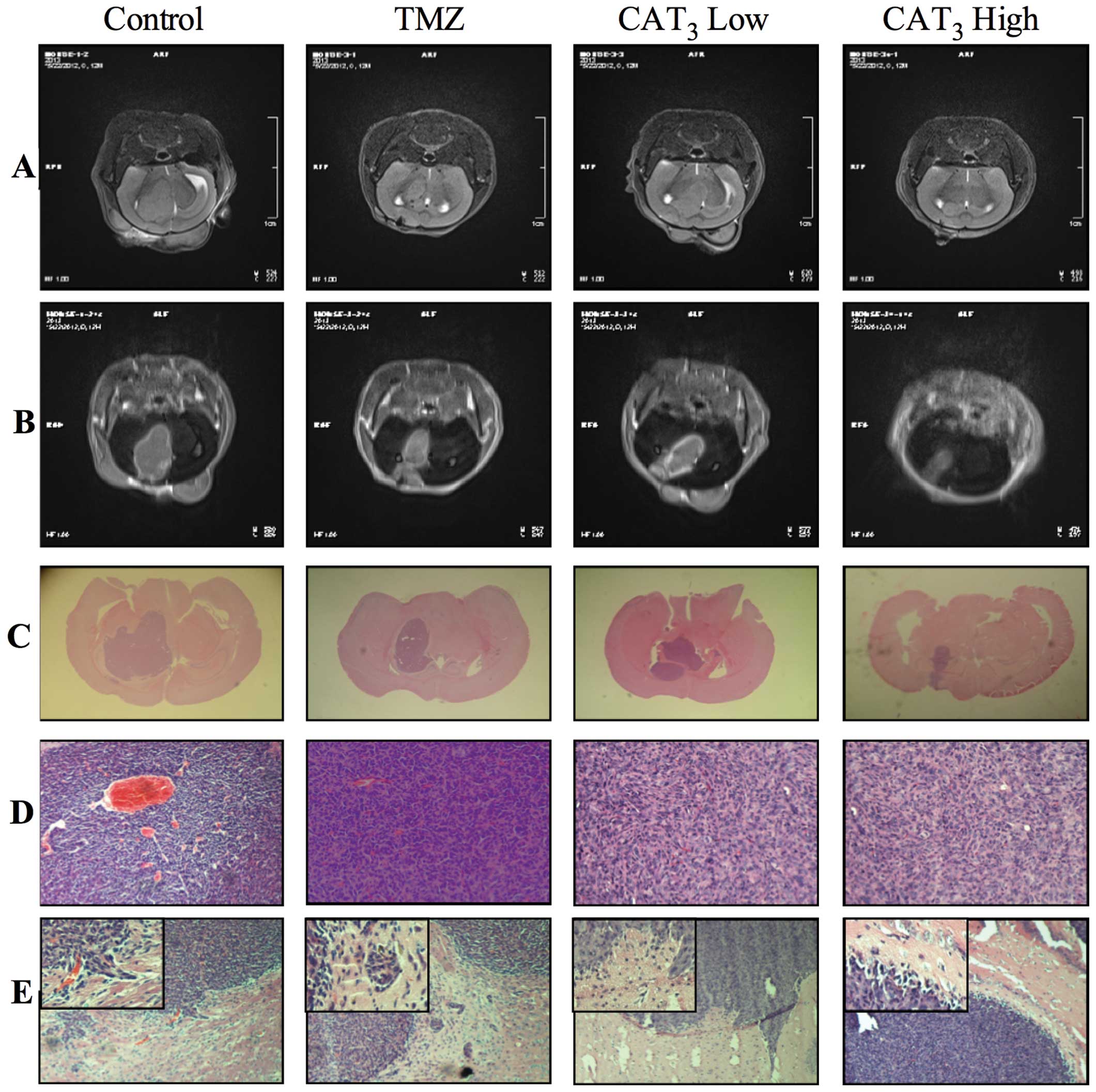
In order to observe the anti-neuroblastoma activity
of PF403 in vivo, an in situ model was enrolled. The
pro-drug of PF403, CAT3, was utilized to improve the
bioavailability. MRI was chosen to observe the development of tumor
in brain, and H&E staining in diagnosis and in analysis of the
tumor. Coronal plane T2 weighted images demonstrated mildly
hyper-intense intracranial tumor and hard to observe invasion of
the surrounding brain tissue of mice in the experimental group.
Control group T2 weighted images showed larger area of
hyper-intense intracranial tumor than the experimental groups, and
clear invasion of the surrounding brain tissue (Fig. 5A). Coronal plane enhanced T1
weighted images demonstrated markedly small intracranial tumors of
mice in the experimental group, whereas tumors occupied almost the
whole brain of mice in the control group (Fig. 5B). Tumor was observed in 9
successive slices for control group, and 8 slices for TMZ (120
mg/kg) group, but only 6 and 4 slices for CAT3 low group
(6 mg/kg) and CAT3 high group (12 mg/kg).
Histopathological examination of control mice showed that
experimental groups developed smaller tumors in the brain (Fig. 5C). H&E staining proved the
tumor as neuroblastoma, tumor cells packed closely, cell morphology
was basically the same, mostly the small round cells were
distributed like islands. Shape and boundary of the tumors were
consistent with MRI results. The tumor cells had less cytoplasm and
their nucleus was clear. Capillaries surrounding tumor cells were
arranged in a rosette structure (Fig.
5D). Tumor cells invaded into the surrounding normal tissue,
but in CAT3 treated groups this phenomenon was not
observed (Fig 5E). The volume of
tumor through MRI and H&E stain sections were calculated, and
showed consistency of the two methods (Table III). These results proved that
the pro-drug of PF403 had potent anti-neuroblastoma activity in
situ.
| Table IIIAntitumor activity of CAT3
on neuroblastoma in situ. |
Table III
Antitumor activity of CAT3
on neuroblastoma in situ.
| Body weight | H&E
staining | MRI |
|---|
|
|
|
|
|---|
| Groups | Beginning | End | Lmax (mm) | W (mm) | VH
(mm) | Inhibition rate
(%) | Lmax (mm) | W (mm) | T (mm) | VM
(mm) | Inhibition rate
(%) |
|---|
| Control | 16.2±0.9 | 17.3±2.9 | 5.4±1.3 | 4.4±1.6 | 104.5 | - | 5.2±2.1 | 3.7±1.1 | 4.5±0.5 | 86.6 | - |
| TMZ | 16.7±1.1 | 17.5±2.1 | 4.3±1.2 | 2.4±1.0 | 24.8a | 76.3 | 3.9±1.3 | 2.4±0.8 | 4.0±0.5 | 37.4a | 56.8 |
| CAT3
low | 16.5±1.1 | 17.7±2.3 | 4.0±1.5 | 3.1±1.1 | 38.4a | 63.3 | 3.9±1.1 | 2.9±0.9 | 3.0±0.5 | 33.9a | 60.9 |
| CAT3
high | 16.3±1.2 | 17.9±1.9 | 2.8±1.0 | 1.7±0.6 | 8.1a | 92.2 | 2.7±1.1 | 2.0±0.5 | 2.0±0.5 | 10.8a | 87.5 |
PF403 induces the change of multiple cell
signaling pathways involved in tumor cell invasion and cell
apoptosis
We explored which factors were involved in the
events where PF403 inhibited invasion and induced apoptosis of
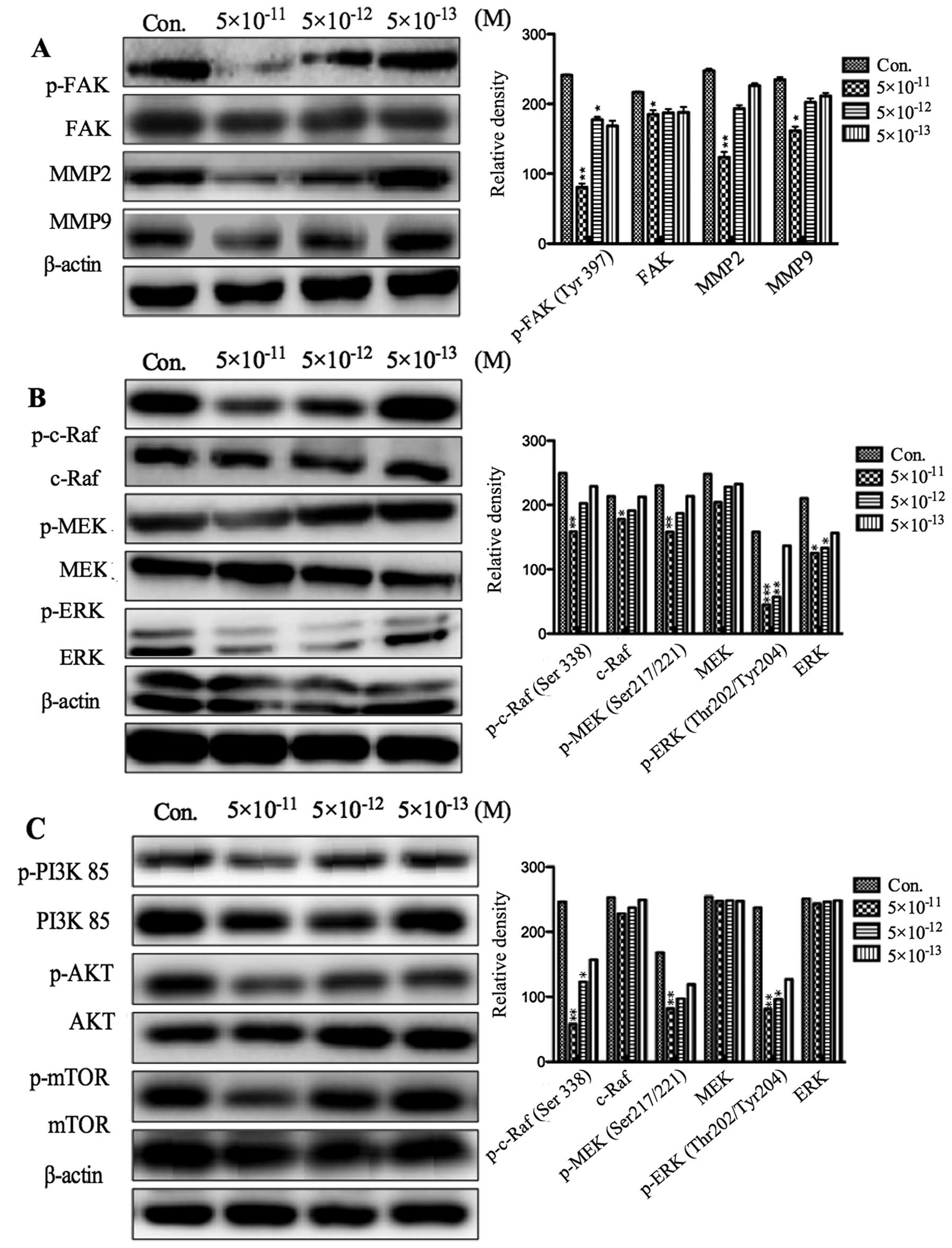
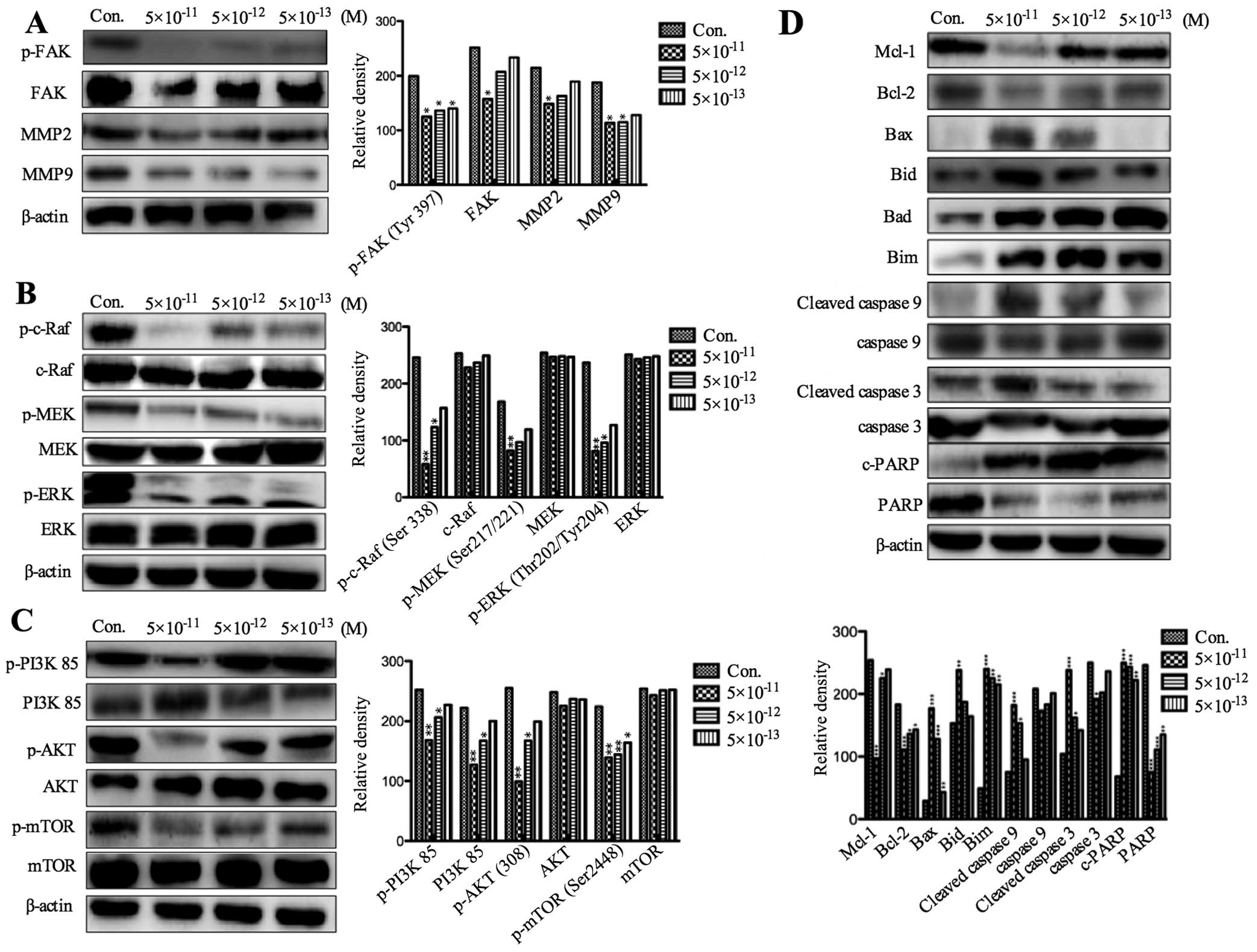
neuroblastoma cells. In the process of tumor cell invasion, focal
adhesion kinase (FAK) and matrix metalloproteinases (MMPs) play an
important role (10). The levels
of MMP-2, MMP-9, phosphorylated and total FAK in PF403
(5×10−11 M, 5×10−12 M and 5×10−13
M) treated SH-SY5Y cells were detected by western blot analysis at
different times. The results showed that the expression of p-FAK,
MMP-2 and MMP-9 was, respectively, significantly decreased after
treatment with PF403 for 4 and 72 h (Figs. 6A and 7A). However, the total of FAK did not
alter (Figs. 6A and 7A). The expression of FAK is related to
the Ras-mitogen-activated protein kinases (MAPKs) and PI3K/AKT cell
signaling pathway (11,12). To assess whether the PF403
inhibited phosphorylation of FAK was associated with these cell
signaling pathways, SH-SY5Y cells were treated with various
concentrations of PF403 (5×10−11 M, 5×10−12 M
and 5×10−13 M) for 4 and 72 h. The results of protein
expression of Raf/MEK/ERK and PI3K/AKT/mTOR pathway demonstrated
that PF403 downregulated the proteins expression of Raf/MEK/ERK and
PI3K/AKT/mTOR pathways at the concentration of 5×10−11 M
in 4 h (Fig. 6B and C), and PF403
downregulated proteins expression of Raf/MEK/ERK and PI3K/AKT/mTOR
pathway at 5×10−13 M for 72 h in a concentration
dependent manner (Fig. 7B and C).
PF403 accelerated apoptosis by upregulated expression of
pro-apoptotic proteins (Bad, Bid, Bim and Bax), downregulated
anti-apoptosis proteins (Mcl-1 and Bcl-2), and activated
caspase-dependent mitochondrial apoptosis pathways
(cleave-caspase-9, cleave-caspase-3 and c-PARP) in cells
administrated with PF403 for 72 h (Fig. 7D).
Discussion
The clinical characteristics of NB are
heterogeneity, metastasis and high malignancy, resulting in shorter
survival in patients (2,13). Few chemotherapeutic or molecularly
targeted antineoplastic agents have shown efficacy in brain tumors,
therefore, development of novel chemotherapy agents are urgent. Our
earlier study had found that PF403 exerted potent antitumor
activity in various tumors in vitro, such as, melanoma,
hepatoma, ovarian cancer, glioma and neuroblastoma (9). As this compound could penetrate the
blood-brain barrier (8), we
supposed that it could suppress growth of brain tumors, and
neuroblastoma was chosen in this study. The results of MTT, colony
formation and flow cytometry illustrated that PF403-induced SH-SY5Y
cell death was dose- and time-dependent. Significant increase in
cell deaths was observed at the lowest concentration of PF403
treated groups than controls. PF403 induced SH-SY5Y cell apoptosis
with EC50 value of 10−11–10−12 M
at 72 h by MTT assay. This is much lower than its precursor
compound CAT (at 72 h IC50 value was
10−7–10−8 M) (7), and the traditional cytotoxic drug
taxol (at 72 h IC50 value at
10−8–10−9 M). Colony forming assay showed
that PF403 inhibited SH-SY5Y NB cell proliferation in a long-term,
and it had a good dose-effect relationship. Flow cytometry assay
showed that SH-SY5Y cells cultured with PF403 for 72 h induced
intense apoptosis.
Invasion of growth is the most characteristic
biological phenotype of NB, which is also the major reason causing
poor prognosis. The present study found that PF403 could inhibit
the invasion ability of NB cells in a dose-dependent manner, which
is essential for NB treatment.
As PF403 displayed a strong anti-neuroblastoma and
anti-invasion activity in vitro, subsequently its antitumor
activity in vivo was studied. The barrier function of the
BBB is critical for regulating transport to the brain, but also
represents a significant roadblock in developing therapies for CNS
diseases including brain cancer, and most convention of
chemotherapeutic agents cannot penetrate BBB (14–16).
Besides, the microenvironment in brain is different from other
tissues in human body. Thus, an in situ NB model was used to
evaluate the proliferation inhibitory activity of PF403. Our
results exhibited that, 14 days after SH-SY5Y cell implantation,
the tumor could be located by MRI in mouse brain, and
CAT3 significantly inhibited NB cell proliferation.
H&E staining confirmed its potent anti-proliferation and
anti-invasion ability in situ.
FAK is a mediator of cell-extracellular matrix
signaling events and its expression plays an important role in the
process of NB invasion (17).
Compiling evidence implies that FAK regulates focal adhesion
signaling by phosphorylating various substrates, as well as the
MMPs-mediated matrix degradation, which consecutively regulates
downstream signaling cascades (18). FAK signaling played a critical role
in the production of MMPs such as MMP-2 and MMP-9, and subsequently
activated tumor invasion (10).
Sun et al (19)
demonstrated that inhibition of the expression and phosphorylation
of FAK, activities of MMP-2 and MMP-9 could be downregulated. FAK
is also essential for the Ras/ERK signaling pathway (20). The residues surrounding
Tyr397 (FAK) can also constitute a sequence that binds
to the Ras/ERK signaling pathway (21). FAK and MAPK signaling involved in
MMP-2 secretion has been shown in QG90 lung cancer cell (22). Moreover, it was found that the
MMP-9 gene promoter is partially regulated through activation of
the Ras/ERK pathway (23). Also,
PI3K is one of the critical downstream signal molecules of the FAK
pathway (24). Activated FAK binds
the SH2 of PI3K, thereby transporting the catalytic subunit of PI3K
to the membrane, where it catalyzes the phosphorylation of inositol
lipids in lung cancer cell migration (25). Several studies have indicated that
FAK/PI3K/Akt is involved in the regulation of MMP-2 and MMP-9
activities on different cell types (12,26).
In the present study, PF403 inhibited the activation of FAK,
reduced phosphorylation of FAK, and regulated the expression of
MMP-2 and MMP-9. In addition, it showed that PF403 inhibited
PI3K/AKT/mTOR and Raf/MEK/ERK pathways in SH-SY5Y cells. Thus, it
seems that PF403 inhibited SH-SY5Y NB cell migration in concert
with downregulated protein expression of PI3K/AKT and Raf/ERK
signaling pathways, and associated with p-FAK, MMP-2 and MMP-9.
PI3K/AKT/mTOR and Raf/MEK/ERK pathways influence
protein expression of the Bcl-2 family, they play a central role in
apoptosis by regulating mitochondrial outer membrane
permeabilisation (MOMP) and releasing apoptosis-inducing proteins
such as cytochrome c, and SMAC sequestered within the
mitochondria (27). The Bcl-2
family is divided into three groups based on functionality and
presence of conserved Bcl-2 homology (BH1-4) domains: multi-domain
anti-apoptotic proteins (Bcl-2 and Mcl-1), multi-domain
pro-apoptotic proteins (BAX) and BH3-only proteins (Bad, Bid and
Bim). Interactions between these groups of proteins dictate whether
the cells die or not. Multi-domain anti-apoptotic proteins prevent
MOMP by interacting with and sequestering the multi-domain
pro-apoptotic proteins (28,29).
BAX allows for oligomerization at the mitochondrial outer membrane
and subsequent MOMP through pore formation (30). The BH3-only proteins, competitively
bind and neutralize anti-apoptotic proteins, allowing BAX
oligomerization and promoting cell death, whereas Bid and Bim can
also interact with and activate BAX, facilitating membrane
insertion and MOMP (27,31). In this study, upregulation of BAX
and downregulation of Bcl-2 was observed in a PF403
concentration-dependent manner. PF403 could upregulate expression
of Bim, Bid and Bad, and downregulate expression of Mcl1. The
regulation of expression of the Bcl-2 family proteins was a
response to the accelerated SH-SY5Y cell death.
Apoptotic pathways in PF403-mediated cell death in
SH-SY5Y neuroblastoma cells were examined. The activity of PI3K/AKT
and Ras/ERK pathways was downregulated in PF403 treated cells,
which were important to proliferation and apoptosis of NB. Our
results also strongly demonstrated caspase-9, downstream of Bcl-2
family, was activated in PF403 treated SH-SY5Y cells, suggesting
activation of the intrinsic apoptotic pathway. Downstream target of
caspase-9, caspase-3, was also activated in PF403 treated SH-SY5Y
cells. Activation of caspase-3 has been reported to have targets in
nuclear compartment for the initiation of DNA damage suggesting its
nuclear localization (32). PARP
is a nuclear target of cleaved caspase-3 and proteolysis cleavage
of PARP is considered to be a hallmark feature of apoptosis. Our
results demonstrated the presence of only full-length PARP protein
in untreated cells but a strong PARP fragment was produced in all
PF403 treated cells, strongly suggesting involvement of apoptosis.
Taken together, our results clearly suggest that the intrinsic
apoptotic pathway was contributed to PF403-mediated SH-SY5Y cell
death.
In conclusion, the results presented in the present
study demonstrated the effect and novel mechanisms associated with
cytotoxic property of PF403 on NB cells. Our studies show that
PF403, a novel antitumor agent, could induce apoptosis and inhibit
invasion and metastasis of SH-SY5Y NB cells in vitro;
CAT3, the pro-drug of PF403, had anti-neuroblastoma
activity in situ; PF403 inhibited invasion of NB cells
through downregulating protein expression of p-FAK, following
MMP-2, MMP-9 and Raf/ERK and PI3K/AKT pathways; PF403-mediated
neuroblastoma cell death partly through downregulating Raf/MEK/ERK
and PI3K/AKT/mTOR pathways, and then activating apoptotic pathway
through regulation of proteins expression of Bcl-2 family
activating the intrinsic apoptotic pathway. Thus, the primary
biochemical mechanism of action of PF403 was as speculated. In
summary, our results strongly suggested that PF403 has potential as
a new anticancer drug for the treatment of neuroblastoma.
References
|
1
|
Brodeur GM: Neuroblastoma: biological
insights into a clinical enigma. Nat Rev Cancer. 3:203–216. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Maris JM, Hogarty MD, Bagatell R and Cohn
SL: Neuroblastoma. Lancet. 369:2106–2120. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chemler SR: Phenanthroindolizidines and
phenanthroquinolizidines: promising alkaloids for anti-cancer
therapy. Curr Bioact Compd. 5:2–19. 2007. View Article : Google Scholar
|
|
4
|
Fu Y, Lee SK, Min HY, Lee T, Lee J, Cheng
M and Kim S: Synthesis and structure-activity studies of antofine
analogues as potential anticancer agents. Bioorg Med Chem Lett.
17:97–100. 2007. View Article : Google Scholar
|
|
5
|
Gao W, Bussom S, Grill SP, Gullen EA, Hu
YC, Huang X, Zhong S, Kaczmarek C, Gutierrez J, Francis S, et al:
Structure-activity studies of phenanthroindolizidine alkaloids as
potential antitumor agents. Bioorg Med Chem Lett. 17:4338–4342.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xi Z, Zhang R, Yu Z and Ouyang D: The
interaction between tylophorine B and TMV RNA. Bioorg Med Chem
Lett. 16:4300–4304. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liu ZJ, Lv HN, Li HY, Zhang Y, Zhang HJ,
Su FQ, Si YK, Yu SS and Chen XG: Anticancer effect and
neurotoxicity of S-(+)-deoxytylophorinidine, a new
phenanthroindolizidine alkaloid that interacts with nucleic acids.
J Asian Nat Prod Res. 13:400–408. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lv H, Ren J, Ma S, Xu S, Qu J, Liu Z, Zhou
Q, Chen X and Yu S: Synthesis, biological evaluation and mechanism
studies of deoxytylophorinine and its derivatives as potential
anticancer agents. PLoS One. 7:e303422012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yu P, Li C, Ren J, Ma S, Song X, Chen X
and Yu S: Stereospecific synthesis and biological evaluation of
monodesmethyl metabolites of (+)-13a-(S)-deoxytylophorinine as
potential antitumor agents. Synthesis. 44:3757–3764. 2012.
View Article : Google Scholar
|
|
10
|
Mon NN, Ito S, Senga T and Hamaguchi M:
FAK signaling in neoplastic disorders: a linkage between
inflammation and cancer. Ann NY Acad Sci. 1086:199–212. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shibata K, Kikkawa F, Nawa A, Thant AA,
Naruse K, Mizutani S and Hamaguchi M: Both focal adhesion kinase
and c-Ras are required for the enhanced matrix metalloproteinase 9
secretion by fibronectin in ovarian cancer cells. Cancer Res.
58:900–903. 1998.PubMed/NCBI
|
|
12
|
Zeng ZZ, Jia Y, Hahn NJ, Markwart SM,
Rockwood KF and Livant DL: Role of focal adhesion kinase and
phosphatidylinositol 3′-kinase in integrin fibronectin receptor-
mediated, matrix metalloproteinase-1-dependent invasion by
metastatic prostate cancer cells. Cancer Res. 66:8091–8099. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
De Bernardi B, Nicolas B, Boni L, Indolfi
P, Carli M, Cordero Di Montezemolo L, Donfrancesco A, Pession A,
Provenzi M, Di Cataldo A, et al: Disseminated neuroblastoma in
children older than one year at diagnosis: comparable results with
three consecutive high-dose protocols adopted by the Italian
Co-Operative Group for Neuroblastoma. J Clin Oncol. 21:1592–1601.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pardridge WM: The blood-brain barrier:
bottleneck in brain drug development. NeuroRx. 2:3–14. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Pardridge WM: Molecular Trojan horses for
blood-brain barrier drug delivery. Curr Opin Pharmacol. 6:494–500.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Pardridge WM: Biopharmaceutical drug
targeting to the brain. J Drug Target. 18:157–167. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yount G, Taft RJ, Luu T, Rachlin K, Moore
D and Zhang W: Independent motile microplast formation correlates
with glioma cell invasiveness. J Neurooncol. 81:113–121. 2007.
View Article : Google Scholar
|
|
18
|
Canel M, Secades P, Garzón-Arango M,
Allonca E, Suarez C, Serrels A, Frame M, Brunton V and Chiara MD:
Involvement of focal adhesion kinase in cellular invasion of head
and neck squamous cell carcinomas via regulation of MMP-2
expression. Br J Cancer. 98:1274–1284. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sun T, Zhao N, Ni CS, Zhao XL, Zhang WZ,
Su X, Zhang DF, Gu Q and Sun BC: Doxycycline inhibits the adhesion
and migration of melanoma cells by inhibiting the expression and
phosphorylation of focal adhesion kinase (FAK). Cancer Lett.
285:141–150. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mon NN, Hasegawa H, Thant AA, Huang P,
Tanimura Y, Senga T and Hamaguchi M: A role for focal adhesion
kinase signaling in tumor necrosis factor-alpha-dependent matrix
metalloproteinase-9 production in a cholangiocarcinoma cell line,
CCKS1. Cancer Res. 66:6778–6784. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lai KC, Huang AC, Hsu SC, Kuo CL, Yang JS,
Wu SH and Chung JG: Benzyl isothiocyanate (BITC) inhibits migration
and invasion of human colon cancer HT29 cells by inhibiting matrix
metalloproteinase-2/-9 and urokinase plasminogen (uPA) through PKC
and MAPK signaling pathway. J Agric Food Chem. 58:2935–2942. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang Y, Thant AA, Hiraiwa Y, Naito Y,
Sein TT, Sohara Y, Matsuda S and Hamaguchi M: A role for focal
adhesion kinase in hyluronan-dependent MMP-2 secretion in a human
small-cell lung carcinoma cell line, QG90. Biochem Biophys Res
Commun. 290:1123–1127. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chang YM, Shih YT, Chen YS, Liu CL, Fang
WK, Tsai CH, Tsai FJ, Kuo WW, Lai TY and Huang CY: Schwann cell
migration induced by earthworm extract via activation of PAs and
MMP-2/9 mediated through ERK1/2 and p38. Evid Based Complement
Alternat Med. 2011:3954582011. View Article : Google Scholar
|
|
24
|
Choi YA, Lim HK, Kim JR, Lee CH, Kim YJ,
Kang SS and Baek SH: Group IB secretory phospholipase A2 promotes
matrix metalloproteinase-2-mediated cell migration via the
phosphatidylinositol 3-kinase and Akt pathway. J Biol Chem.
279:36579–36585. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Meng XN, Jin Y, Yu Y, Bai J, Liu GY, Zhu
J, Zhao YZ, Wang Z, Chen F, Lee KY, et al: Characterisation of
fibronectin-mediated FAK signalling pathways in lung cancer cell
migration and invasion. Br J Cancer. 101:327–334. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chan KC, Ho HH, Huang CN, Lin MC, Chen HM
and Wang CJ: Mulberry leaf extract inhibits vascular smooth muscle
cell migration involving a block of small GTPase and Akt/NF-kappaB
signals. J Agric Food Chem. 57:9147–9153. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Green DR and Kroemer G: The
Pathophysiology of mitochondrial cell death. Science. 305:626–629.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kuwana T and Newmeyer DD: Bcl-2-family
proteins and the role of mitochondria in apoptosis. Curr Opin Cell
Biol. 15:691–699. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ren D, Tu HC, Kim H, Wang GX, Bean GR,
Takeuchi O, Jeffers JR, Zambetti GP, Hsieh JJ and Cheng EH: BID,
BIM, and PUMA are essential for activation of the BAX- and
BAK-dependent cell death program. Science. 330:1390–1393. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wei MC, Zong WX, Cheng EH, Lindsten T,
Panoutsakopoulou V, Ross AJ, Roth KA, MacGregor GR, Thompson CB and
Korsmeyer SJ: Proapoptotic BAX and BAK: a requisite gateway to
mitochondrial dysfunction and death. Science. 292:727–730. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yang E, Zha J, Jockel J, Boise LH,
Thompson CB and Korsmeyer SJ: Bad, a heterodimeric partner for
Bcl-XL and Bcl-2, displaces Bax and promotes cell death. Cell.
80:285–291. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhivotovsky B, Samali A, Gahm A and
Orrenius S: Caspases their intracellular localization and
translocation during apoptosis. Cell Death Differ. 6:644–651. 1999.
View Article : Google Scholar : PubMed/NCBI
|