Introduction
More than 80% of ovarian cancer cases are discovered
at the advanced stage. When the patients are first diagnosed, they
already have metastasis (1). The
combination of surgery and chemotherapy is common approach to treat
ovarian cancer. Many chemotherapeutic agents including carboplatin,
cisplatin and paclitaxel have been used as a single drug or in
combination. The study conducted by Einzig et al in 1992,
which reported the significant activity of paclitaxel in patients
with advanced ovarian cancer (2),
established the co-administration of paclitaxel and
cisplatin/carboplatin as a standard chemotherapeutic regimen. Since
30% ovarian cancer patients in advanced stages are non-responders
to the first-line chemotherapy and the acquired chemoresistance is
also emerged from the initial responders (3), both intrinsic and acquired
resistances have been the major causes of low survival rates and
poor prognosis. Therefore, it is urgently required to understand
the underlying molecular mechanisms involved in such
chemoresistance, which can introduce novel attempts to overcome
resistance and eventually success to treat ovarian cancer.
In human, there are >90 protein tyrosine kinases
and 58 of them are receptor tyrosine kinases (RTKs) which are, in
turn, divided into 20 families (4). TAM family, a subfamily of RTK, is
comprised of three RTK members, which are Tyro3 (alternatively
called Sky), Axl (alternatively called Ark and Ufo), and Mer
(5). They share common
extracellular structures, two immunoglobin-like domains and two
fibronectin type III repeats, and conserved kinase domain within
cytoplasmic region (6). It has
been reported that the typical ligands to interact with TAM RTKs
are growth arrest-specific 6 (GAS6) and protein S which are vitamin
K-dependent proteins and transduce many intracellular signals
including survival, proliferation, inhibition of apoptosis,
adhesion, morphology, invasion and motility (7–9).
Indeed, overexpression of TAM RTKs as well as GAS6 protein have
been observed in the initiation and progression of many cancers
(10) such as breast cancer
(11), colon cancer (12), gastric cancer (13), leukemias (14), liver cancer (15), melanoma (16,17),
ovarian cancer (18,19) and prostate cancer (20). Therefore, targeting TAM RTKs and
their ligands might be a potent strategy to control these
cancers.
Metformin is the most widely prescribed biguanide to
treat type 2 diabetes (21). In
addition to the primary effect on glucose metabolism, the
antitumorigenic potential of metformin has come to the forefront,
since the accumulating evidence suggests its anticancer effects on
cell proliferation in various cancers and tumor growth in xenograft
model (22–25). The anticancer mechanisms of
metformin have been assessed by its ability to activate
AMP-activated protein kinase (AMPK), resulting in the concomitant
inhibition of mammalian target of rapamycin complex I (mTORC1)
activity which plays an important role in protein synthesis and
cell proliferation (26–29). Metformin has also been reported to
directly inhibit mTORC1 independent of AMPK and to target epidermal
growth factor receptor (EGFR) and Akt/mitogen-activated protein
kinase (MAPK) which mediate pro-survival and anti-apoptotic signals
(30–32).
In this study, we examined the effect of metformin
on the expression of Axl and Tyro3 RTKs in both parental and
chemoresistant ovarian cancer cells, which affects the viability of
these cells. Metformin was found to suppress the expression of both
Axl and Tyro3 RTKs at transcriptional level and deactivate
downstream signaling molecules such as Erk and signal transducers
and activators of transcription 3 (STAT3), which led to significant
decrease of cell proliferation through downregulation of
anti-apoptotic protein such as XIAP.
Materials and methods
Reagents and antibodies
Metformin was obtained from Sigma (St. Louis, MO,
USA). For western blot analysis, specific antibodies against
phospho-Akt, Akt, Axl, phospho-Erk, Erk, phospho-STAT3, STAT3,
Tyro3, XIAP and GAPDH and secondary antibodies were obtained from
Santa Cruz Biotechnology Inc. (Dallas, TX, USA). Primers for PCR to
amplify Axl, Tyro3, and GAPDH were synthesized by Bioneer Inc.
(Daejoun, Korea). TRI reagent was from Solgent (Daejoun, Korea).
AmpliTaq DNA polymerase was obtained from Roche Inc (Indianapolis,
IN, USA).
Cell culture
A2780 and SKOV3 cells were purchased from the
American Type Culture Collection and A2780/Cis cells were from
Sigma. SKOV3/TR cells, a variant of SKOV3 cells with resistance to
taxol, were established as we previously reported. Briefly,
SKOV3/TR cells had acquired the taxol resistance via long-term and
stepwise exposure of the parental cells to the escalating
concentrations of taxol. All of these cells were grown in RPMI-1640
(Gibco BRL, Grand Island, NY, USA) containing 10% FBS, 2 mM
L-glutamine, 10 U/ml penicillin, and 10 g/ml streptomycin at 37°C
in 5% CO2 in a water-saturated atmosphere.
RT-PCR
Cells (3×105) were seeded in 60-mm
culture dishes and grown overnight at 37°C and then treated with
the indicated concentrations of metformin for the 48 h. Total RNA
was extracted using TRI reagent and subjected to cDNA synthesis and
PCR. The specific primers were as follows: Axl, sense
5′-AACCTTCAACTCCTGCCTTCTCG-3′ and antisense
5′-CAGCTTCTCCTTCAGCTCTTCAC-3′; Tyro3, sense
5′-GTGTGTGGCTGACTTCGGAC-3′ and antisense
5′-CACGTCCTCCATACACTCCG-3′; GAPDH, sense 5′-GGAGCCAAAAGGGTCATCAT-3′
and antisense 5′-GTGATGGCATGGACTGTGGT-3′.
Western blot analysis
Cells were treated with the indicated concentrations
of metformin for the 48 h. Whole cell lysates were prepared from
those cells using lysis buffer [1% Triton X-100, 50 mM Tris (pH
8.0), 150 mM NaCl, 1 mM PMSF, 1 mM Na3VO4 and
protease inhibitor cocktail]. Protein concentrations were
determined using Bio-Rad protein assays. Proteins from whole cell
lysates (20–40 μg) were separated on 12% SDS-PAGE, and
electrotransferred to nitrocellulose membranes. Membranes were
blocked for 30 min at room temperature in Tris-buffered
saline-0.05% Tween-20 (TTBS) containing 5% non-fat dry milk, and
then incubated with TTBS containing a primary antibody for 4 h at
room temperature. After 3×10-min washes in TTBS, membranes were
incubated with peroxidase-conjugated secondary antibody for 1 h.
Following 3 additional 10-min washes with TTBS, protein bands of
interest were visualized using an enhanced chemiluminescence
detection system (Amersham™ ECL™ prime Western Blotting detection
reagent; GE Healthcare, Piscataway, NJ, USA).
Colony-forming assay
Cells (1–2×103 cells per dish) were
seeded in 35-mm culture dishes and allowed to grow in the absence
or presence of the metformin for 7–10 days to form colonies.
Colonies of >50 cells were visualized by crystal violet (in 60%
methanol, Junsei Chemical, Japan) staining and images were taken by
RAS 3000 Image Analysis System (Fuji Film, Japan).
Axl promoter activity test
To measure the Axl promoter activity upon metformin
treatment, the plasmid containing Axl promoter-luciferase reporter,
pGL3-Axl, was used. As we previously reported, pGL3-Axl plasmid is
constructed by PCR amplification of Axl promoter region ranging
from −887 to +7 bp of the transcriptional start site and the
following subcloning of the PCR product into the pGL3-basic vector.
The pGL3-Axl plasmid was co-transfected into cells
(3×104 cells in a 60-mm dish) with Renilla
luciferase vectors, pRL-SV40, as an internal control. Luciferase
activity was measured using a Dual-Glo luciferase assay system.
siRNA transfection
RNA interference silencing was performed to suppress
Axl or Tyro3 protein levels. SKOV3 or SKVO3/TR cells
(1×106) were seeded in 100-mm culture dish and grown
overnight and then transfected with 50 nM siRNA against Axl
(sc-36438, Santa Cruz Biotech), Tyro3 (sc-36438, Santa Cruz
Biotech), or control siRNA (sc-37007, Santa Cruz Biotech). For cell
proliferation assay, cells were harvested 24 h after transfection
and re-seeded into 35-mm culture dishes and allowed to grow for 48
h in the absence or presence of the metformin. Cells were then
harvested and the number of viable cells was counted. Whole cell
lysates were prepared and used for western blot analysis to
evaluate Axl and Tyro3 protein levels.
ELISA
The level of IL-6 in culture media was measured
using ELISA kit from R&D Systems according to the
manufacturer’s protocol. Cells were treated with the indicated
concentration of metformin for 48 h. Conditioned media were
harvested and assayed for IL-6. The data are representative of at
least three independent experiments.
Statistical analysis
Data are expressed as the mean ± SD of triplicate
samples or at least three independent experiments. Student’s t-test
was used to determine statistical significance with a threshold
P-value of <0.05.
Results
Metformin inhibits proliferation of both
parental and chemo-resistant ovarian cancer cells
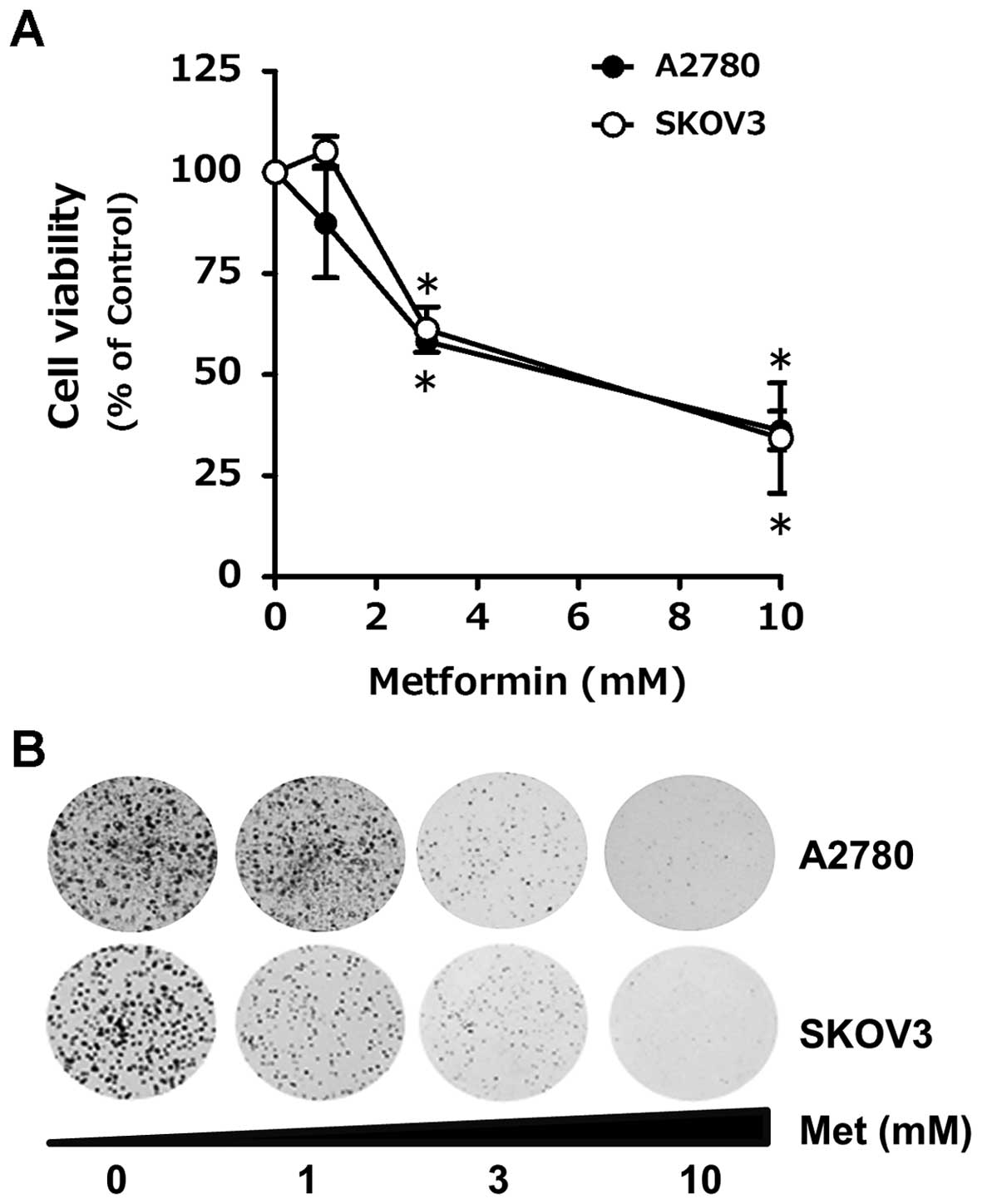
To determine anti-proliferative effect of metformin
on human ovarian cancer cells, A2780 and SKOV3 cells were incubated
with 1, 3 and 10 mM of metformin for 48 h and then cell viability
was measured. As shown in Fig. 1A,
cell counting results showed that metformin treatment caused
dose-dependent reduction of cell proliferation in both cell types.
Of note, when the cells were exposed to 10 mM of metformin for 48
h, the number of viable A2780 and SKOV3 cells were diminished to 36
and 33% of that of untreated cells, respectively. The inhibitory
effect of metformin on cell proliferation was further confirmed by
colony-forming assay. Exposure of A2780 and SKOV3 cells upon 1, 3
and 10 mM metformin for 7 days were found to form less colonies in
a dose-dependent manner (Fig. 1B).
Especially, both cell types failed to grow into colonies in the
presence of 10 mM metformin, indicating cytotoxic effect of
metformin on human ovarian cancer cells.
We also found that metformin decreased proliferation
of chemoresistant ovarian cancer cells. Cisplatin-resistant
A2780/Cis cells and taxol-resistant SKOV3/TR, which are each of
subline derived from the parental A2780 and SKOV3 cells, were
treated with 1, 3, 10 mM of metformin for 48 h for cell counting or
7 days for colony-forming assay. As shown in Fig. 1C, the viability SKOV3/TR cells at
10 mM metformin was 37% of control, while that of A2780/Cis cells
was 55% of control. The result from colony-forming assay also
demonstrated that metformin induced a dose-dependent inhibition of
cell proliferation (Fig. 1D). It
seems to be evident that SKOV3/TR cells were more profoundly
affected by metformin treatment than A2780/Cis cells, since both
the cell viability and the number of colonies of SKOV3/TR cells
incubated with metformin were less than each of those of A2780/Cis
cells exposed to the same concentrations of metformin (Fig. 1C and D). Taken together, these
results indicate that metformin decreases proliferative capacity of
both parental and chemoresistant ovarian cancer cells, SKOV3,
SKOV3/TR, A2780 and A2780/Cis cells.
Metformin suppresses the expression of
Axl and Tyro3 RTKs at transcriptional level
Next, we explored if metformin affects the
expression of receptor tyrosine kinases (RTKs), since TMA RTKs,
Axl, Tyro3, and Mer, have been reported to mediate cell survival
and proliferation signals (33).
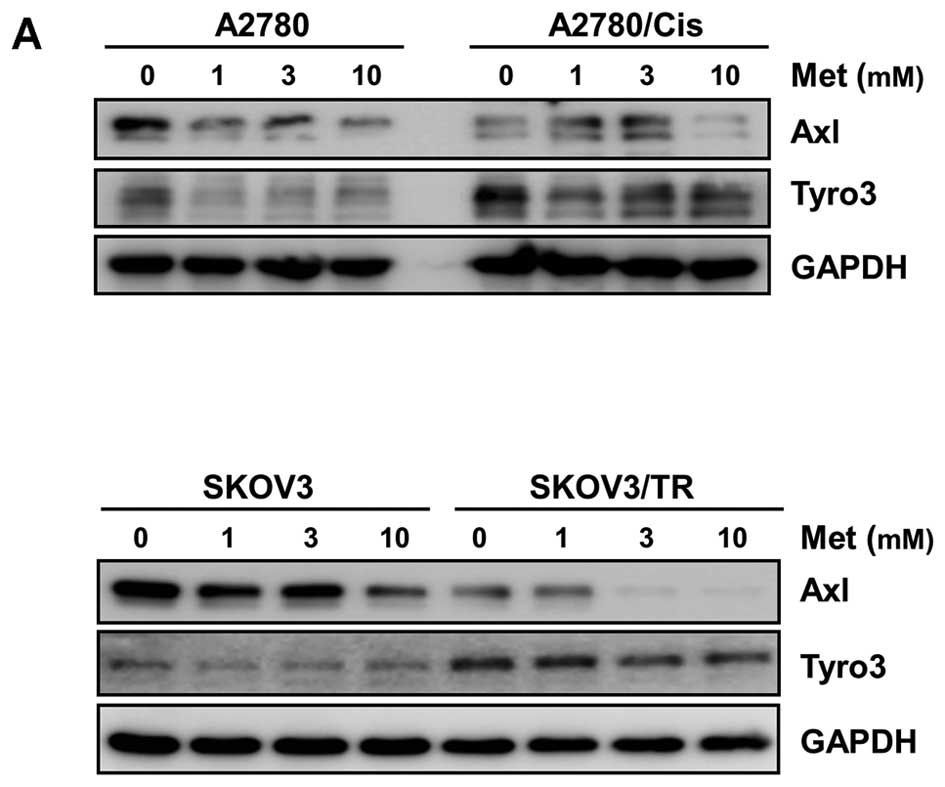
After exposure of cells with the indicated concentrations of
metformin for 48 h, the expression of Axl and Tyro3 was examined at
protein and mRNA levels. As shown in Fig. 2A, metformin treatment was found to
induce dose-dependent downregulation of Axl and Tyro3 expression in
both SKOV3 and taxol-resistant SKOV3/TR cells, while Axl and Tyro3
protein levels in A2780 cells and cisplatin-resistant A2780/Cis
cells were significantly reduced upon 1 or 10 mM metformin
treatment.
The effect of metformin on Axl and Tyro3 expression
was further confirmed by RT-PCR. Consistent with western blot
results, the mRNA levels of Axl and Tyro3 were decreased by
metformin treatment (Fig. 2B). It
is notable that Axl mRNA expression was slightly reduced in
chemoresistant A2780/Cis and SKOV3/TR cells, compared to that in
their parental cells, while Tyro3 mRNA expression was increased in
these two chemoresistant cells, indicating that the acquisition of
cisplatin or taxol resistance resulted in the down/upregulation of
Axl and Tyro3 expression, respectively.
The inhibitory effect of metformin on the
transcription of Axl gene was also demonstrated by promoter
activity test. SKOV3 cells were transfected with pGL3-Axl
construct, a luciferase expressing plasmid under the control of
human Axl promoter and then incubated with 10 mM metformin for 48
h. As illustrated in Fig. 2C,
metformin treatment was found to reduce Axl promoter activity to
46% compared with the untreated cells. The results from RT-PCR and
promoter activity test indicate that metformin suppresses Axl
expression at the transcriptional level.
Anti-proliferative effect of metformin is
mediated by dysregulation of Axl and Tyro3 RTKs and downstream
signaling molecules
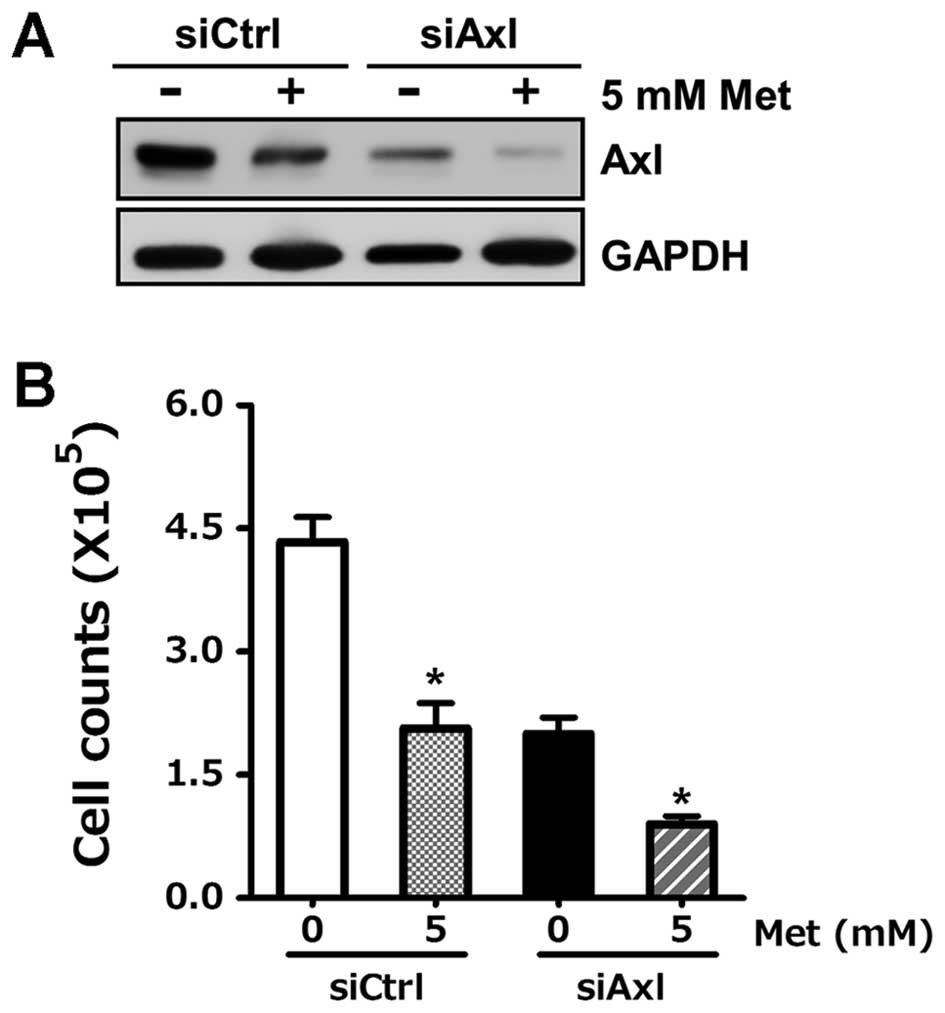
We further examined whether the inhibitory effect of
metformin on Axl and Tyro3 expression is responsible for its
anti-proliferative property. Axl and Tyro3-specific siRNA, siAxl
and siTyro3, was transfected into SKOV3 and SKOV3/TR cells,
respectively, and then cells were treated with metformin to
evaluate the synergistic effect on protein expression as well as
cell viability. Western blot results showed that knock-down of Axl
and Tyro3 protein level by specific siRNA augmented
metformin-induced downregulation of each protein expression
(Fig. 3A and C). The viability of
the cells transfected with siAxl or siTyro3 was found to be 48% in
SKOV3 cells and 45% in SKOV3/TR cells, thus being lower than each
of the cells transfected with control siRNA, siCtrl (Fig. 3B and D). This indicates that
decreased Axl and Tyro3 protein level by siRNA escalated the
inhibitory effect of metformin on cell proliferation. Taken
together, these data demonstrate that Axl and Tyro3 RTKs are novel
targets of metformin for its anti-cancer activity in ovarian cancer
cells.
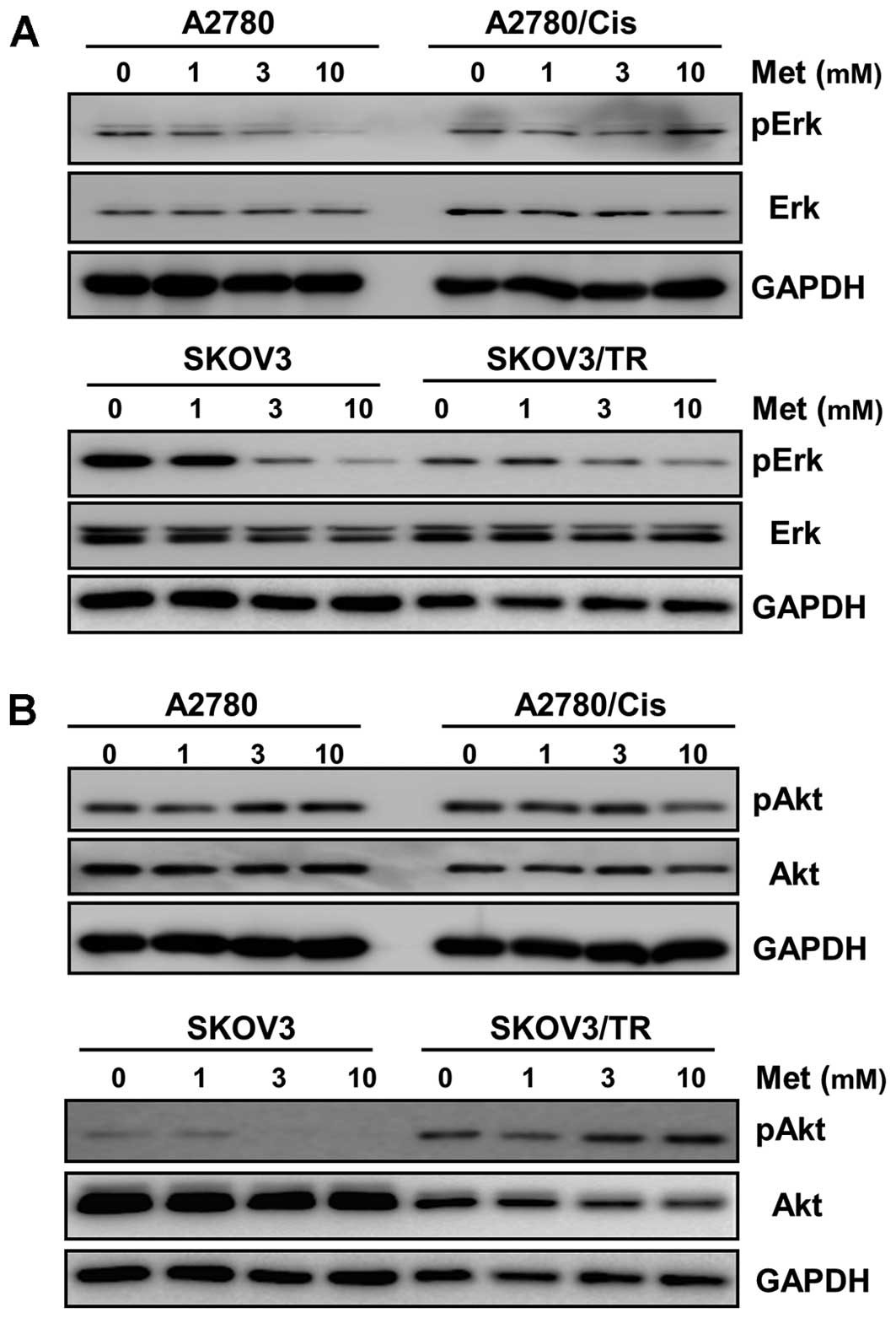
We next examined the effects of metformin on several
downstream effectors known as regulators of cell survival,
proliferation and apoptosis. As shown in Fig. 4A, dose-dependent inhibition of Erk
phosphorylation by metformin was found in A2780, SKOV3 and SKOV3/TR
cells, but not in A2780/Cis cells. On the other hand, metformin was
found to have no effect on phosphorylation of Akt which delivers
proliferative signals and is elevated in SKOV3/TR cells, except in
SKOV3 cells (Fig. 4B).
Western blot analysis further showed that metformin
fairly reduced phosphorylation of signal transducers and activators
of transcription 3 (STAT3), one of the key regulators driving cell
proliferation in both parental and chemoresistant cells (Fig. 4C). Based on metformin-mediated
suppression of STAT3 phosphorylation, we also examined IL-6
production upon metformin treatment, since activation of IL-6/STAT3
axis has been known to play an important role in cell viability
(34,35). ELISA results showed that total IL-6
production was slightly reduced by metformin, whereas IL-6
production per cell was rather increased (Fig. 4D).
In addition, we observed that the metformin reduced
the level of X-linked inhibitor of apoptosis protein (XIAP) which
inhibits apoptosis (Fig. 4E).
Collectively, these data indicate that metformin induces the
downregulation of Axl and Tyro3 expression and the subsequent
decrease of Erk and STAT3 phosphorylation as well as XIAP
expression by which it exerts its anticancer activity in ovarian
cancer cells.
Discussion
Cisplatin has primarily been approved as an
anticancer agent to treat testicular and ovarian cancer in 1978
(36,37), and is the first-line chemotherapy
drug used in many cancers such as bladder, cervical, ovarian
cancers, lymphomas and sarcomas, as a single agent or in
combination with other anticancer agents, including paclitaxel
(38–40). Taxol, a microtubule-stabilizing
drug, which in turn prevents depolymerization of microtubule, has
been frequently used to treat breast, lung, stomach or ovarian
cancers (41). Although the
combination of platinum-based agent and taxol is still standard
chemotherapy regimen in ovarian cancers, most patients eventually
develop tolerance against cisplatin and taxol, leading to a low
5-year survival rate (3,42). However, the molecular mechanisms of
this chemoresistance are not yet fully understood.
We found that metformin suppressed the viability of
ovarian cancer cells (A2780 and SKOV3) and their
cisplatin/taxol-resistant cells (A2780/Cis and SKOV3/TR) (Fig. 1). Consistent with our data, the
anticancer and chemopreventive effects of metformin have been
suggested by many epidemiological and preclinical studies which
demonstrated that metformin treatment is associated with the
reduction of cancer risk and better prognosis in patients with type
II diabetes (43–46). Of note, however, there are also
several studies which failed to demonstrate any beneficial effect
of metformin (47–49).
We then observed that metformin downregulates the
expression of Axl and Tyro3 receptor tyrosine kinases (RTKs) at
transcriptional level (Fig. 2).
TAM family of RTKs, Tyro3, Axl and Mer, have been reported to play
critical roles in cell survival, proliferation, apoptosis, and
adhesion (33). Dysregulation of
these RTKs and their ligands including GAS6 has been shown in many
human cancers. In ovarian adenocarcinoma tissues, the
overexpression and activation of Axl, Mer and ligands have been
observed and found to be correlated with disease stages and
metastasis (19). In acute
lymphoblastic leukemia (ALL) and non-small cell lung cancer (NSCLC)
cell lines, the Mer expression was found to be elevated to promote
cancer cell survival, tumorigenesis and chemoresistance. In
addition, Axl or Mer inhibition using RNA interference, monoclonal
antibodies and shRNA have been demonstrated to abrogate
proliferation, metastasis and tumor progression in xenograft model,
while it enhanced chemosensitivity in ALL and NSCLC (9,50,51).
Consistent with these reports, we also found that the silencing of
Axl and Tyro3 RTKs by specific siRNAs, siAxl and siTyro3, augmented
the anti-proliferative effect of metformin in SKOV3 and SKOV3/TR
cells, respectively (Fig. 3B and
D). Our data suggest that Axl and Tyro3 RTKs might be novel
targets of metformin to inhibit cell proliferation as well as to
overcome chemoresistance.
A recent study by Feng et al demonstrated
that metformin-induced inactivation of STAT3 signaling promotes
autophagy and apoptosis as a mechanism to inhibit cell growth in
esophageal squamous cells (52).
In our setting, metformin was found to decrease STAT3
phosphorylation (Fig. 4C). Since
STAT3 is a transcription factor to induce IL-6 production and IL-6
phosphorylates STAT3, we examined the effect of metformin on IL-6
expression. Interestingly, metformin was found to have almost no
effect of on IL-6 production and IL-6 production per cell was even
shown to be increased upon metformin treatment (Fig. 4D). These results indicate that
metformin directly inhibits STAT3 phosphorylation, while cells must
somehow have a strategy to compensate the IL-6 production which
might provide residual cell viability upon metformin exposure.
In conclusion, our data demonstrate that metformin
has inhibitory effects on Axl and Tyro3 expression and subsequent
activation of Erk and STAT3, resulting in its anti-proliferative
activity in both parental and chemoresistant ovarian cancer cells.
Thus, Axl and Tyro3 and/or their downstream signaling pathway seem
to be novel therapeutic targets of metformin to control
proliferation and chemoresistance of ovarian cancer cells.
Acknowledgements
This study was supported by the by the Basic Science
Research Program through the National Research Foundation of Korea
(NRF) funded by the Ministry of Education, Science and Technology
(grant no. 2006-2005303).
Abbreviations:
|
AMPK
|
AMP-activated protein kinase
|
|
EGFR
|
epidermal growth factor receptor
|
|
ELISA
|
enzyme-linked immunosorbent assay
|
|
GAS6
|
growth arrest-specific 6
|
|
IL-6
|
interleukin-6
|
|
mTORC
|
mammalian target of rapamycin
complex
|
|
RTK
|
receptor tyrosine kinase
|
|
STAT
|
signal transducers and activators of
transcription
|
|
XIAP
|
X-linked inhibitor of apoptosis
protein
|
References
|
1
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Einzig AI, Wiernik PH, Sasloff J, Runowicz
CD and Goldberg GL: Phase II study and long-term follow-up of
patients treated with Taxol for advanced ovarian adenocarcinoma. J
Clin Oncol. 10:1748–1753. 1992.PubMed/NCBI
|
|
3
|
McGuire WP, Hoskins WJ, Brady MF, Kucera
PR, Partridge EE, Look KY, Clarke-Pearson DL and Davidson M:
Cyclophosphamide and cisplatin compared with paclitaxel and
cisplatin in patients with stage III and stage IV ovarian cancer. N
Engl J Med. 334:1–6. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Robinson DR, Wu YM and Lin SF: The protein
tyrosine kinase family of the human genome. Oncogene. 19:5548–5557.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ohashi K, Mizuno K, Kuma K, Miyata T and
Nakamura T: Cloning of the cDNA for a novel receptor tyrosine
kinase, Sky, predominantly expressed in brain. Oncogene. 9:699–705.
1994.PubMed/NCBI
|
|
6
|
Sasaki T, Knyazev PG, Clout NJ, Cheburkin
Y, Göhring W, Ullrich A, Timpl R and Hohenester E: Structural basis
for Gas6-Axl signalling. EMBO J. 25:80–87. 2006. View Article : Google Scholar
|
|
7
|
Stitt TN, Conn G, Gore M, Lai C, Bruno J,
Radziejewski C, Mattsson K, Fisher J, Gies DR, Jones PF, et al: The
anticoagulation factor protein S and its relative, Gas6, are
ligands for the Tyro 3/Axl family of receptor tyrosine kinases.
Cell. 80:661–670. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hafizi S and Dahlbäck B: Gas6 and protein
S: Vitamin K-dependent ligands for the Axl receptor tyrosine kinase
subfamily. FEBS J. 273:5231–5244. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wimmel A, Glitz D, Kraus A, Roeder J and
Schuermann M: Axl receptor tyrosine kinase expression in human lung
cancer cell lines correlates with cellular adhesion. Eur J Cancer.
37:2264–2274. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zwick E, Bange J and Ullrich A: Receptor
tyrosine kinase signalling as a target for cancer intervention
strategies. Endocr Relat Cancer. 8:161–173. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Meric F, Lee WP, Sahin A, Zhang H, Kung HJ
and Hung MC: Expression profile of tyrosine kinases in breast
cancer. Clin Cancer Res. 8:361–367. 2002.PubMed/NCBI
|
|
12
|
Craven RJ, Xu LH, Weiner TM, Fridell YW,
Dent GA, Srivastava S, Varnum B, Liu ET and Cance WG: Receptor
tyrosine kinases expressed in metastatic colon cancer. Int J
Cancer. 60:791–797. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wu CW, Li AF, Chi CW, Lai CH, Huang CL, Lo
SS, Lui WY and Lin WC: Clinical significance of AXL kinase family
in gastric cancer. Anticancer Res. 22B:1071–1078. 2002.
|
|
14
|
Challier C, Uphoff CC, Janssen JW and
Drexler HG: Differential expression of the ufo/axl oncogene in
human leukemia-lymphoma cell lines. Leukemia. 10:781–787.
1996.PubMed/NCBI
|
|
15
|
Tsou AP, Wu KM, Tsen TY, Chi CW, Chiu JH,
Lui WY, Hu CP, Chang C, Chou CK and Tsai SF: Parallel hybridization
analysis of multiple protein kinase genes: Identification of gene
expression patterns characteristic of human hepatocellular
carcinoma. Genomics. 50:331–340. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Györffy B and Lage H: A Web-based data
warehouse on gene expression in human malignant melanoma. J Invest
Dermatol. 127:394–399. 2007. View Article : Google Scholar
|
|
17
|
van Ginkel PR, Gee RL, Shearer RL,
Subramanian L, Walker TM, Albert DM, Meisner LF, Varnum BC and
Polans AS: Expression of the receptor tyrosine kinase Axl promotes
ocular melanoma cell survival. Cancer Res. 64:128–134. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Macleod K, Mullen P, Sewell J, Rabiasz G,
Lawrie S, Miller E, Smyth JF and Langdon SP: Altered ErbB receptor
signaling and gene expression in cisplatin-resistant ovarian
cancer. Cancer Res. 65:6789–6800. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rankin EB, Fuh KC, Taylor TE, Krieg AJ,
Musser M, Yuan J, Wei K, Kuo CJ, Longacre TA and Giaccia AJ: AXL is
an essential factor and therapeutic target for metastatic ovarian
cancer. Cancer Res. 70:7570–7579. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sainaghi PP, Castello L, Bergamasco L,
Galletti M, Bellosta P and Avanzi GC: Gas6 induces proliferation in
prostate carcinoma cell lines expressing the Axl receptor. J Cell
Physiol. 204:36–44. 2005. View Article : Google Scholar
|
|
21
|
Donato NJ: Imatinib is receptive to a
collaboration. Blood. 118:4009–4010. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gotlieb WH, Saumet J, Beauchamp MC, Gu J,
Lau S, Pollak MN and Bruchim I: In vitro metformin anti-neoplastic
activity in epithelial ovarian cancer. Gynecol Oncol. 110:246–250.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang LW, Li ZS, Zou DW, Jin ZD, Gao J and
Xu GM: Metformin induces apoptosis of pancreatic cancer cells.
World J Gastroenterol. 14:7192–7198. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Goodwin PJ, Pritchard KI, Ennis M, Clemons
M, Graham M and Fantus IG: Insulin-lowering effects of metformin in
women with early breast cancer. Clin Breast Cancer. 8:501–505.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ben Sahra I, Laurent K, Loubat A,
Giorgetti-Peraldi S, Colosetti P, Auberger P, Tanti JF, Le
Marchand-Brustel Y and Bost F: The antidiabetic drug metformin
exerts an antitumoral effect in vitro and in vivo through a
decrease of cyclin D1 level. Oncogene. 27:3576–3586. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hadad SM, Appleyard V and Thompson AM:
Therapeutic metformin/AMPK activation promotes the angiogenic
phenotype in the ERalpha negative MDA-MB-435 breast cancer model.
Breast Cancer Res Treat. 114:3912009. View Article : Google Scholar
|
|
27
|
Liu H, Scholz C, Zang C, Schefe JH, Habbel
P, Regierer AC, Schulz CO, Possinger K and Eucker J: Metformin and
the mTOR inhibitor everolimus (RAD001) sensitize breast cancer
cells to the cytotoxic effect of chemotherapeutic drugs in vitro.
Anticancer Res. 32:1627–1637. 2012.PubMed/NCBI
|
|
28
|
Nair V, Sreevalsan S, Basha R, Abdelrahim
M, Abudayyeh A, Rodrigues Hoffman A and Safe S: Mechanism of
metformin-dependent inhibition of mammalian target of rapamycin
(mTOR) and Ras activity in pancreatic cancer: Role of specificity
protein (Sp) transcription factors. J Biol Chem. 289:27692–27701.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Honjo S, Ajani JA, Scott AW, Chen Q,
Skinner HD, Stroehlein J, Johnson RL and Song S: Metformin
sensitizes chemotherapy by targeting cancer stem cells and the mTOR
pathway in esophageal cancer. Int J Oncol. 45:567–574.
2014.PubMed/NCBI
|
|
30
|
Ota S, Horigome K, Ishii T, Nakai M,
Hayashi K, Kawamura T, Kishino A, Taiji M and Kimura T: Metformin
suppresses glucose-6-phosphatase expression by a complex I
inhibition and AMPK activation-independent mechanism. Biochem
Biophys Res Commun. 388:311–316. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ben Sahra I, Regazzetti C, Robert G,
Laurent K, Le Marchand-Brustel Y, Auberger P, Tanti JF,
Giorgetti-Peraldi S and Bost F: Metformin, independent of AMPK,
induces mTOR inhibition and cell-cycle arrest through REDD1. Cancer
Res. 71:4366–4372. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ming M, Sinnett-Smith J, Wang J, Soares
HP, Young SH, Eibl G and Rozengurt E: Dose-dependent AMPK-dependent
and independent mechanisms of berberine and metformin inhibition of
mTORC1, ERK, DNA synthesis and proliferation in pancreatic cancer
cells. PLoS One. 9:e1145732014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Linger RM, Keating AK, Earp HS and Graham
DK: TAM receptor tyrosine kinases: Biologic functions, signaling,
and potential therapeutic targeting in human cancer. Adv Cancer
Res. 100:35–83. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chang KT, Tsai CM, Chiou YC, Chiu CH, Jeng
KS and Huang CY: IL-6 induces neuroendocrine dedifferentiation and
cell proliferation in non-small cell lung cancer cells. Am J
Physiol Lung Cell Mol Physiol. 289:L446–L453. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Block KM, Hanke NT, Maine EA and Baker AF:
IL-6 stimulates STAT3 and Pim-1 kinase in pancreatic cancer cell
lines. Pancreas. 41:773–781. 2012.PubMed/NCBI
|
|
36
|
Rodriguez LH and Johnson DE: Clinical
trial of cisplatinum (NSC 119875) in metastatic renal cell
carcinoma. Urology. 11:344–346. 1978. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Briscoe KE, Pasmantier MW, Ohnuma T and
Kennedy BJ: cis-Dichlorodiammineplatinum(II) and adriamycin
treatment of advanced ovarian cancer. Cancer Treat Rep.
62:2027–2030. 1978.PubMed/NCBI
|
|
38
|
Go RS and Adjei AA: Review of the
comparative pharmacology and clinical activity of cisplatin and
carboplatin. J Clin Oncol. 17:409–422. 1999.PubMed/NCBI
|
|
39
|
Boeckman HJ, Trego KS and Turchi JJ:
Cisplatin sensitizes cancer cells to ionizing radiation via
inhibition of nonhomologous end joining. Mol Cancer Res. 3:277–285.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kelland L: The resurgence of
platinum-based cancer chemotherapy. Nat Rev Cancer. 7:573–584.
2007. View
Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kohler DR and Goldspiel BR: Paclitaxel
(Taxol). Pharmacotherapy. 14:3–34. 1994.PubMed/NCBI
|
|
42
|
Stordal B, Pavlakis N and Davey R: A
systematic review of platinum and taxane resistance from bench to
clinic: An inverse relationship. Cancer Treat Rev. 33:688–703.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Jiralerspong S, Palla SL, Giordano SH,
Meric-Bernstam F, Liedtke C, Barnett CM, Hsu L, Hung MC, Hortobagyi
GN and Gonzalez-Angulo AM: Metformin and pathologic complete
responses to neoadjuvant chemotherapy in diabetic patients with
breast cancer. J Clin Oncol. 27:3297–3302. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lai SW, Chen PC, Liao KF, Muo CH, Lin CC
and Sung FC: Risk of hepatocellular carcinoma in diabetic patients
and risk reduction associated with anti-diabetic therapy: A
population-based cohort study. Am J Gastroenterol. 107:46–52. 2012.
View Article : Google Scholar
|
|
45
|
Garrett CR, Hassabo HM, Bhadkamkar NA, Wen
S, Baladandayuthapani V, Kee BK, Eng C and Hassan MM: Survival
advantage observed with the use of metformin in patients with type
II diabetes and colorectal cancer. Br J Cancer. 106:1374–1378.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Sadeghi N, Abbruzzese JL, Yeung SC, Hassan
M and Li D: Metformin use is associated with better survival of
diabetic patients with pancreatic cancer. Clin Cancer Res.
18:2905–2912. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Bodmer M, Becker C, Meier C, Jick SS and
Meier CR: Use of metformin is not associated with a decreased risk
of colorectal cancer: A case-control analysis. Cancer Epidemiol
Biomarkers Prev. 21:280–286. 2012. View Article : Google Scholar
|
|
48
|
Bodmer M, Becker C, Jick SS and Meier CR:
Metformin does not alter the risk of lung cancer: A case-control
analysis. Lung Cancer. 78:133–137. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Cossor FI, Adams-Campbell LL, Chlebowski
RT, Gunter MJ, Johnson K, Martell RE, McTiernan A, Simon MS, Rohan
T, Wallace RB, et al: Diabetes, metformin use, and colorectal
cancer survival in postmenopausal women. Cancer Epidemiol.
37:742–749. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Linger RM, Keating AK, Earp HS and Graham
DK: Taking aim at Mer and Axl receptor tyrosine kinases as novel
therapeutic targets in solid tumors. Expert Opin Ther Targets.
14:1073–1090. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Lee-Sherick AB, Eisenman KM, Sather S,
McGranahan A, Armistead PM, McGary CS, Hunsucker SA, Schlegel J,
Martinson H, Cannon C, et al: Aberrant Mer receptor tyrosine kinase
expression contributes to leukemogenesis in acute myeloid leukemia.
Oncogene. 32:5359–5368. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Feng Y, Ke C, Tang Q, Dong H, Zheng X, Lin
W, Ke J, Huang J, Yeung SC and Zhang H: Metformin promotes
autophagy and apoptosis in esophageal squamous cell carcinoma by
downregulating Stat3 signaling. Cell Death Dis. 5:e10882014.
View Article : Google Scholar : PubMed/NCBI
|