Introduction
The incidence of melanoma has increased over the
past decades and this tumor entity is responsible for the majority
of skin cancer-related deaths (1,2).
Melanoma is one of the most challenging types of cancer to be
treated due to its high heterogeneity. Surgery remains the main
treatment option for early-stage melanomas (3). However, the prognosis of patients
diagnosed with metastatic melanoma is still poor, with only a
limited number of agents available for treatment (4,5). One
of the latest successful drugs for the treatment of advanced
melanoma is the BRAF inhibitor vemurafenib (also known as PLX4032)
that showed an increase in overall survival and an extension of
progression-free survival (6,7).
However, incomplete response and acquired drug resistance caused by
multiple abnormally regulated signaling pathways present in
melanoma frequently led to treatment failures and tumor progression
(8–10). Therefore, simultaneously targeting
multiple signaling pathways has great potential for enhancing
therapeutic efficacy and overcoming resistance to vemurafenib
(10,11).
The chondroitin sulfate proteoglycan 4 (CSPG4) is a
cell-surface proteoglycan involved in the activation of several
signaling pathways playing an important role in tumor cell
proliferation, survival, and migration as well as in tumor
progression (12). Since it is
highly expressed on tumor cells and has restricted distribution in
normal tissues, CSPG4 has been used as a target of antibody-based
immunotherapy (13,14). The CSPG4-specific monoclonal
antibody 225.28 was shown to induce regression of tumor metastases,
to inhibit spontaneous metastasis and tumor recurrence in breast
cancer mouse models (15), and to
inhibit the growth and recurrence of melanoma in a human melanoma
xenograft model (16). Recent
findings have shown that the combination of vemurafenib and the
CSPG4-specific mAb 225.28 was capable of blocking the FAK and PKCα
signaling pathways which are important for cell growth, migration,
and survival (17). Antibodies
induced by a mimotope vaccine (anti-225D9+-TT Abs)
against the epitope defined by the mAb 225.28 were capable of
inhibiting melanoma cell growth in vitro and in vivo
(18,19).
As many other cancers, melanomas include regions of
hypoxia caused by an imbalance between oxygen supply and
consumption. The response to treatment is affected by this
microenvironmental factor (20,21).
Tumor hypoxia can negatively influence treatment outcome and
patient survival in various cancer types (22,23).
Melanomas appear to down-regulate signaling pathways associated
with proliferation in order to migrate (24). Hypoxia has been shown to enhance
the cell migratory propensity and invasiveness and to contribute to
cancer metastasis (25) through
the hypoxia inducible factor 1α (HIF1α). HIF1α regulates genes that
are regarded pro-tumorigenic (26,27)
and increases the expression of a number of genes involved in
invasion (28). Carbonic anhydrase
IX (CAIX), a direct transcriptional target of HIF1α, plays an
important role in maintaining pH homeostasis (29). Earlier studies have shown that a
BRAF V600E mutation increased HIF1α expression under hypoxic
conditions (30). Hypoxia also
induced phenotypic plasticity and therapy resistance in melanoma
cells via tyrosine kinase receptors (21).
Herein we report on the response of CSPG4-specific
anti-225D9+-TT Abs to enhance the anti-proliferative
effects of vemurafenib in normoxia. We also describe the role of
hypoxia on the response to vemurafenib and
anti-225D9+-TT Abs and its effect on the
anti-proliferative, migratory and invasive potential of melanoma
cells.
Materials and methods
Cell lines, BRAF inhibitor and polyclonal
antibodies
The human CSPG4 expressing (CSPG4+)
melanoma cell line 518A2 and the human CSPG4 negative
(CSPG4−) melanoma cell line M14, both harboring the
V600E BRAF mutation, were described elsewhere (17,18).
Routine tests to exclude mycoplasma and characterize the origin of
the cells (short tandem repeat analysis) were performed.
Vemurafenib (PLX4032, Selleckchem, Houston, TX, USA) is a potent
selective inhibitor of BRAFV600. Polyclonal
anti-225D9+-TT Abs recognizing CSPG4 were developed and
characterized as previously described (18,19).
Isotype control anti-TT Abs were used as negative control. Exposure
to hypoxia was performed in an anaerobic work station (Ruskin
Technologies, Bridgend, UK) in 2% O2, 5% CO2,
10% H2, and 83% N2 at 37°C.
Cell proliferation assays in normoxic and
hypoxic conditions
The impedance-based x-CELLingence system (ACEA
Bioscience Inc., San Diego, CA, USA) was placed at 37°C in a
humidified 5% CO2 incubator. 518A2 cells
(5×103) were seeded in each well and placed in the
x-CELLingence system and proliferation was measured for 24 h. After
24 h the following compounds were added: i) 5 μM vemurafenib, ii) 1
mM DMOG [dimethyloxalylglycine, N-(methoxyoxoacetyl)-glycine methyl
ester] (Sigma-Aldrich, St. Louis, MO, USA), and iii) 5 μM
vemurafenib plus 1 mM DMOG. The plate was placed back in the
x-CELLingence system and measured for 100 h. DMOG was used to
induce hypoxia in cells when it was technically not possible to use
a hypoxia chamber.
Cell proliferation with
anti-225D9+-TT Abs and vemurafenib
In order to determine the optimal doses of
vemurafenib on 518A2 and M14 melanoma cell lines, dose-titration
experiments were performed. 518A2 (CSPG4+) and M14
(CSPG4−) cells (2×103) per well were seeded
and vemurafenib was added at different concentrations (0.001, 0.01,
0.1, 0.5, 1.0, 10.0 μM). A [3H]-Thymidine incorporation
assay was performed and percentage of inhibition of proliferation
was calculated by comparing the CPM values of treated cells with
those of untreated cells, which were set at 100%.
To test the combinatorial treatment of vemurafenib
and anti-225D9+-TT Abs (2×103) 518A2
(CSPG4+) and M14 (CSPG4−) cells per well were
seeded and incubated with anti-225D9+-TT Abs or isotype
control anti-TT Abs at a concentration of 200 μg/ml. A
[3H]-Thymidine incorporation assay was performed as
previously described (18). To
test the long-term effect of vemurafenib combined with
anti-225D9+-TT Abs, an 8-day proliferation assay was
performed. 518A2 (CSPG4+) and M14 (CSPG4−)
cells (2×103) were seeded. After 24 h, cells were
treated with 1 μM vemurafenib, 1 μM vemurafenib plus 200 μg/ml
anti-225D9+-TT Abs, or 1 μM vemurafenib plus 200 μg/ml
isotype control TT Abs. On day 4, 6 and 8 a
[3H]-Thymidine incorporation assay was performed.
Transwell migration and Transwell
invasion assays
The CIM-Plate 16 (8-μm pore diameter; ACEA
Bioscience Inc.) with or without matrigel [1/10 in medium with 10%
FCS (BD Biosciences, Franklin Lakes, NJ, USA)], was used for
Transwell invasion or migration assays (31). 518A2 cells (4×105) were
seeded in quadruplicates and treated with i) serum-free medium
(SFM), ii) medium supplemented with 10% fetal calf serum (FSC),
iii) vemurafenib (1 μM), and iv) vemurafenib (1 μM) plus
anti-225D9+-TT Abs (200 μg/ml). The plates were placed
in the RTCA DP Analyzer (ACEA Bioscience Inc.) in normoxia or
hypoxia and numbers of migrated or invaded cells measured for 48 h.
Cell index values are directly proportional to the measured
impedances which are automatically and continuously recorded by the
RTCA DP instrument.
Determination of extracellular pH and
oxygen consumption in melanoma cells
Since extracellular pH regulation and oxygen
consumption are affected by hypoxia, which induces a cell-type
specific shift in glycolysis and thereby alters the consumption of
oxygen (32), a SDR optical sensor
system (1450 MicroBeta TriLux; Perkin-Elmer, Waltham, MA, USA)
embedded in the hypoxic box was used for measuring these two
parameters. 518A2 (CSPG4+) and M14 (CSPG4−)
cells (0.2×106) were seeded and incubated with i) 1 μM
vemurafenib, ii) 1 μM vemurafenib plus 200 μg/ml
anti-225D9+-TT Abs, and iii) 200 μg/ml
anti-225D9+-TT Abs. The plates were placed in the
hypoxia chamber with 2% of atmospheric oxygen and the extracellular
pH and oxygen consumption were measured by using the SDR optical
sensor system. The sensor dish reader monitors the pH in real-time
in an OxoDish® for oxygen and in a HydroDish®
for pH using a non-invasive technique that detects the luminescence
lifetime of a sensor spot at the bottom of each well that is
dependent on the pH of the surrounding sample. The extracellular pH
was measured by the SDR every 25 min for 112 h and the oxygen
consumption was measured by the SDR every 5 min for 5 h.
RNA extraction and quantitative reverse
transcription-PCR
Total RNA was extracted with the TRIzol reagent
(Sigma-Aldrich) from cells that had been treated with vemurafenib
(1 μM) and cultured in hypoxic or normoxic conditions for 24 h.
Three micrograms of RNA were reverse-transcribed into cDNA with
High-Capacity cDNA Reverse Transcription kit (Applied Biosystems,
Foster City, CA, USA) using random heptameric primers. Quantitative
RT-PCR analysis of mRNAs of HIF1α, CA9 and β-actin as
internal standard were performed on a StepOne™ Real-time PCR System
(Applied Biosystems) using Power SYBR® Green PCR Master
mix (Applied Biosystems). The following primers were used:
HIF1α sense: 5′-GCTTGGTGCTGATTTGTGAACC-3′, HIF1α
antisense: 5′-GCATCCTGTACTGTCCTGTGGTG-3′; CA9 sense:
5′-CCGAGCGACGCAGCCTTTGA-3′, CA9 antisense:
5′-GGCTCCAGTCTCGGCTACCT-3′; β-actin sense: 5′-TCC
TCCCTGGAGAAGAGCTA-3′, β-actin antisense: 5′-ACATC
TGCTGGAAGGTGGAC-3′. The results were analyzed using the Applied
Biosystems 7500 system v1.4.0 software.
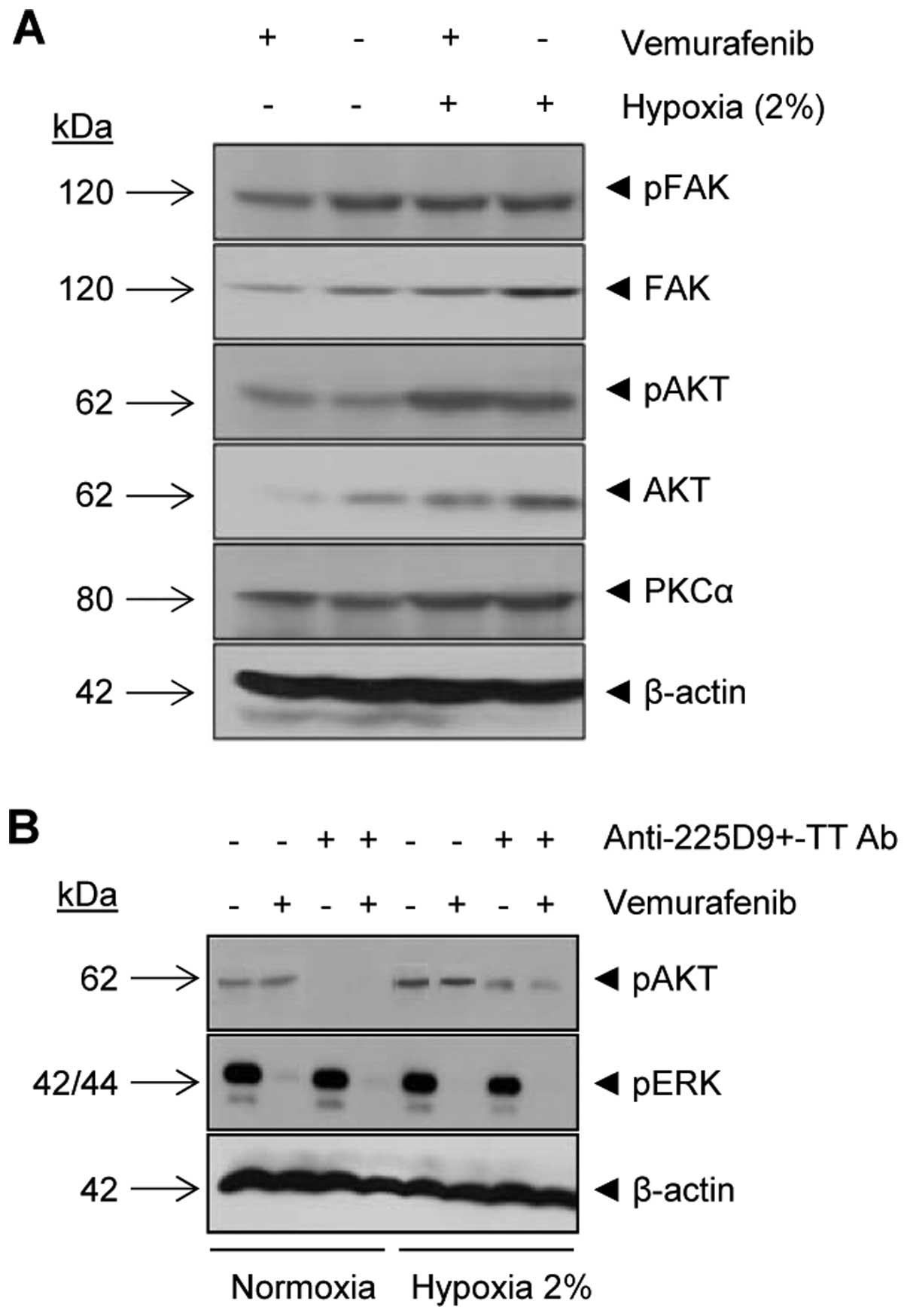
Immunoblot analysis
518A2 melanoma cells were incubated with vemurafenib
(1 μM), anti-225D9+-TT Abs (200 μg/ml) or vemurafenib (1
μM) plus anti-225D9+-TT Abs (200 μg/ml) and placed in
normoxia or hypoxia for 24 h. After treatment cells were lysed and
equal amounts of proteins were separated by SDS-PAGE under reducing
conditions and transferred onto polyvinylidene fluoride (PVDF)
membranes (Immobilon Millipore, Billerica, MA, USA). Cells were
lysed in lysis buffer [1% Triton X-100, 150 mM NaCl, 50 mM Tris, pH
7.5, 0.5% Nonidet P-40, 50 mM NaF, protease inhibitor cocktail
(Roche, Mannheim, Germany) or 10 mM Tris/HCl pH 8.2, 1% NP40, 1 mM
EDTA, 150 mM NaCl, protease inhibitor cocktail (Roche)]. Primary
antibodies used recognized human CAIX [M75, (33)], HIF1α (BD Transduction
Laboratories, San Jose, CA, USA), pFAK(Tyr397), FAK, PKCα (Abcam,
Cambridge, UK) and pAKT (Ser473), AKT, pERK 1/2(Thr202/Tyr204),
ERK1/2 or β-actin mAbs (Cell Signaling Technology, Danvers, MA,
USA). Corresponding peroxidase-conjugated secondary mAbs (Cell
Signaling Technology) were used. Blots were developed using the
LumiGLO chemiluminescencent substrate (Cell Signaling Technology)
and bands visualized using the Filmentwickler CP1000 Processor
(AGFA, Mortsel, Belgium).
Immunohistochemistry in xenografts
The human melanoma C.B.17 SCID/SCID mouse
xenotransplantation model was described elsewhere (16). All experiments were approved by the
Animal Experimentation Committee of the University of Vienna and
the Ministry of Education, Science and Culture, Vienna, Austria.
Untreated 518A2 melanoma tumors were fixed in 4% paraformaldehyde
and embedded in paraffin. For CAIX detection, sections were stained
in the Dako Autostainer using the DakoCytomation
EnVision®+ System-HRP (Dako, Glostrup, Denmark)
according to the manufacturer’s instructions. Antibodies used were
the anti-CAIX antibody M75 and a secondary anti-mouse IgG antibody
(Dako). Staining was visualized with DAB solution (Dako). For HIF1α
immunostaining, antigen retrieval was carried out with citrate
buffer, pH 6.0, for 5 min at 125°C using a Pascal pressure chamber
(Dako). Deparaffinized sections were stained with DakoCytomation
Catalysed Signal Amplification System according to the
manufacturer’s instructions. Sections were incubated with a primary
antibody specific for HIF1α (1:250; BD Transduction Laboratories).
Staining was visualized with DAB solution. The stained sections
were examined with an Olympus BX53 microscope and photographed with
an Olympus DP73 camera (Olympus Electronics, Tokyo, Japan).
Results
Vemurafenib inhibits cell growth of BRAF
V600E mutant 518A2 melanoma cells in normoxic and hypoxic
conditions
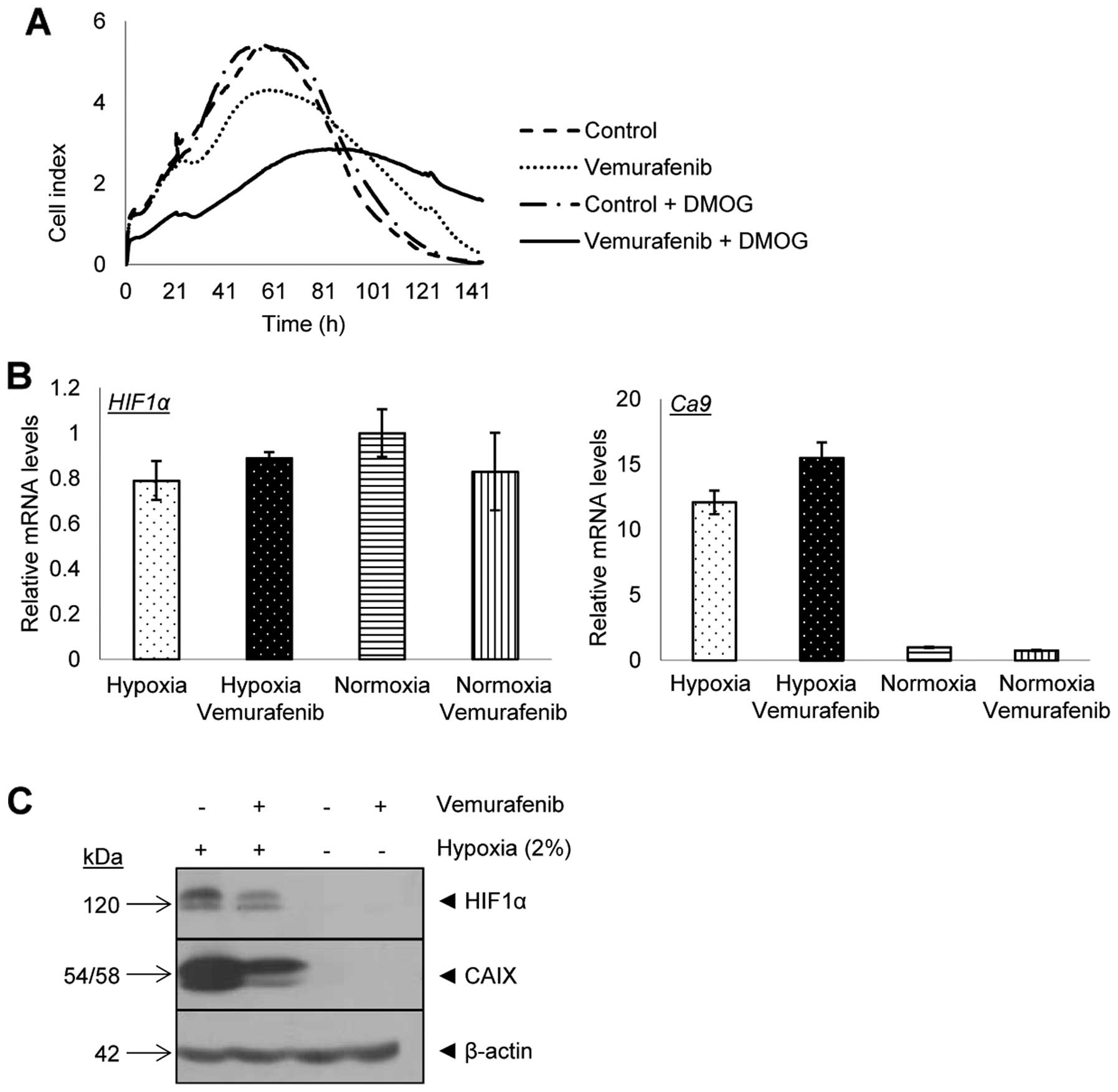
Treatment of melanoma cells with increasing
concentrations of vemurafenib resulted in a dose-dependent
inhibition of proliferation of the BRAF V600E mutant melanoma cell
lines 518A2 and M14. The growth of the BRAF V600E mutant 518A2
melanoma cells and M14 melanoma cells treated with 1 μM vemurafenib
was inhibited by 40±3% and 60±5%, respectively (data not shown). In
order to visualize the time-course of the inhibitory effect of
vemurafenib, a real-time characterization was performed using the
x-CELLingence system. 518A2 cells were treated with 5 μM
vemurafenib and cell proliferation was measured for up to 130 h in
normoxic and hypoxic conditions. 518A2 cells were not susceptible
to hypoxic conditions and the proliferation rate did not change
(maximum cell index of 5.3). After addition of vemurafenib, hypoxic
518A2 cells (maximum cell index of 2.8) reduced cell growth by an
additional 38% compared to normoxic, vemurafenib-treated 518A2
cells (maximum cell index of 4.3) (Fig. 1A). The inhibitory peak after
vemurafenib in normoxic conditions was reached after 60 h, whereas
hypoxia shifted the peak to 90 h (Fig.
1A).
Effect of hypoxia on HIF1α and CAIX
protein expression and mRNA levels in vemurafenib-treated 518A2
melanoma cells
To investigate how hypoxia influences the response
of 518A2 cells to vemurafenib, we focused on important markers of
hypoxia, HIF1α and CAIX. Messenger RNA levels of HIF1α did
not show significant changes. The relative mRNA level of HIF1α in
518A2 cells in hypoxia was 0.8±0.08. Treatment with vemurafenib
increased the mRNA level of HIF1α to 0.9±0.02. In normoxic
conditions the relative mRNA levels of HIF1α were 1.0±0.1
and 0.8±0.17 for untreated and treated 518A2 cells, respectively
(Fig. 1B). The relative CA9
mRNA levels were 1.0±0.5 in normoxic conditions with and without
vemurafenib treatment. In hypoxic conditions CA9 mRNA levels
increased to 12.0±0.9 for untreated cells and to 16.0±1.1 for
vemurafenib-treated cells (Fig.
1B). All mRNA levels have been normalized to the housekeeping
gene coding for β-actin. Both proteins were expressed in
hypoxic 518A2 cells. After treatment with vemurafenib in hypoxic
conditions HIF1α and CAIX protein expression was downregulated,
whereas in normoxic conditions HIF1α and CAIX were not expressed in
either treated or untreated 518A2 cells (Fig. 1C).
Expression of hypoxia markers in melanoma
xenografts
The expression of HIF1α and CAIX in a 518A2 melanoma
xenograft model was evaluated by immunohistochemistry. As shown in
Fig. 2, immunohistochemical
staining of formalin-fixed, paraffin-embedded tumor tissues
demonstrated the presence of HIF1α and CAIX, which showed typical,
hypoxia-related expression patterns. These in vivo results
reflect the expression level of both proteins observed in
vitro.
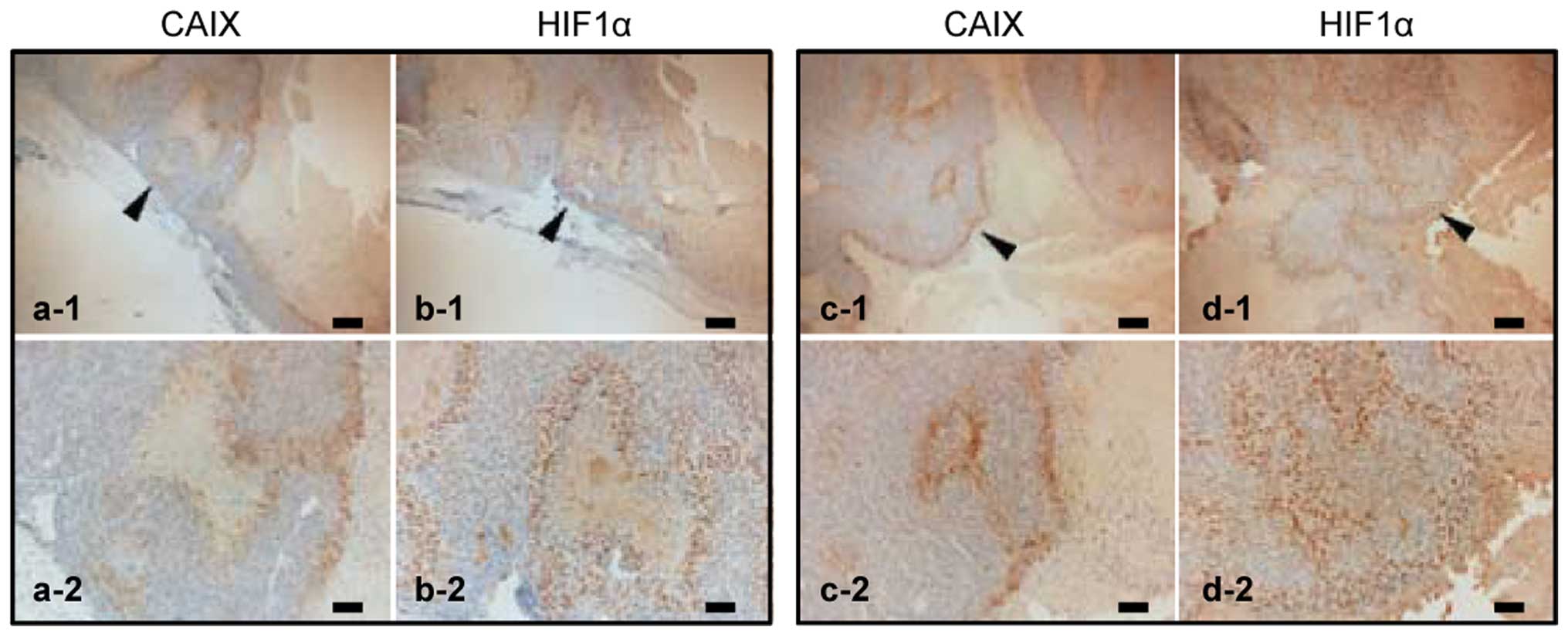 | Figure 2Immunohistochemistry of two
representative 518A2 xenograft tumor samples stained with HIF1α and
CAIX antibodies from corresponding tumor regions. Overview images
of two representative melanoma tumors are shown in the top row
(a-1, b-1 and c-1, d-1); staining for CAIX (a-1, a-2, c-1 and c-2)
and HIF1α (b-1, b-2, d-1 and d-2) shows regions with high
expression. Scale bars, 200 μm (a-1, b-1, c-1 and d-1), and 100 μm
(a-2, b-2, c-2 and d-2). Arrows point to the area of
magnification. |
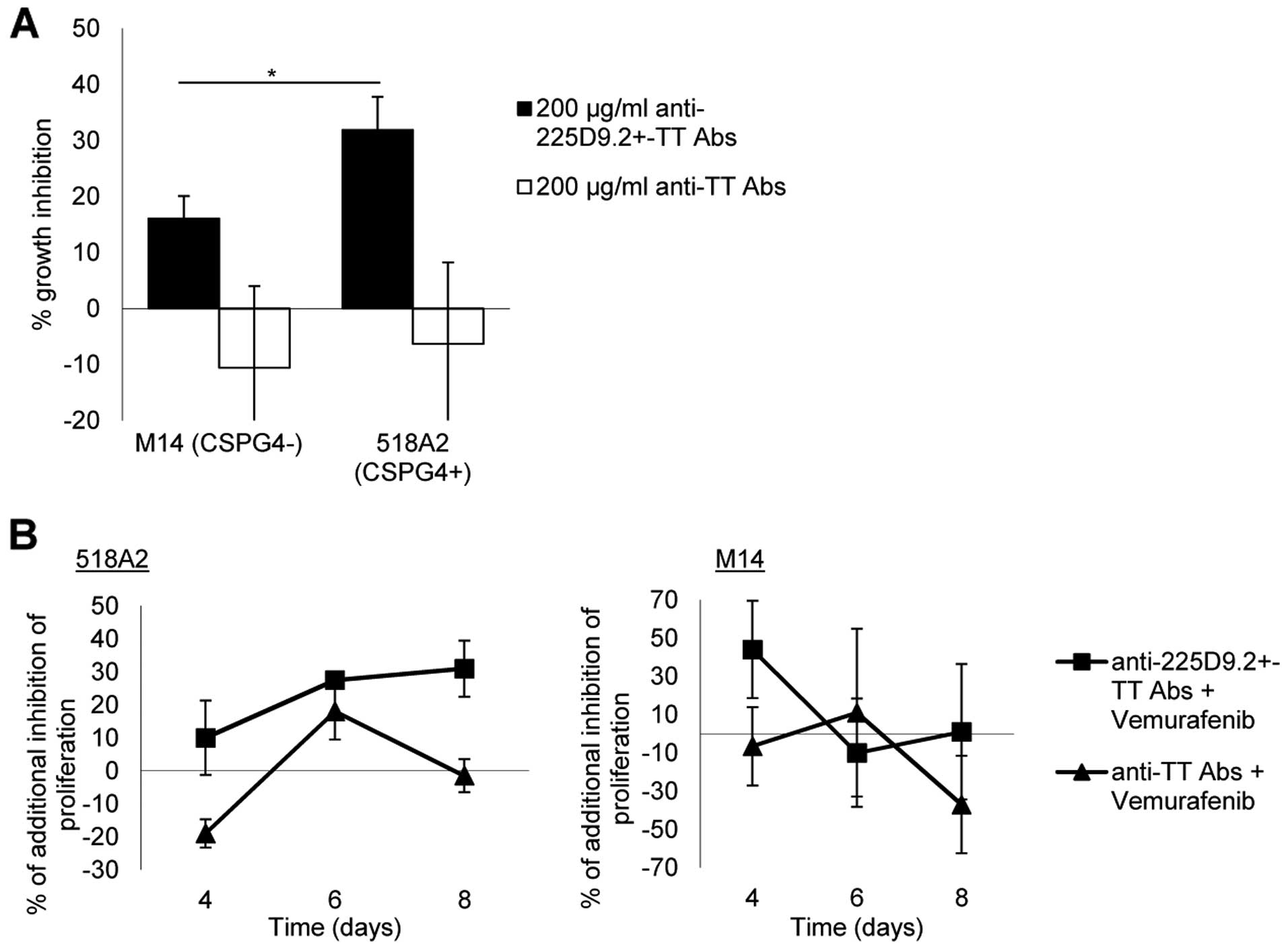
Anti-225D9+-TT Abs enhance the
effects of vemurafenib in CSPG4 expressing 518A2 melanoma
cells
Targeting multiple signaling pathways was more
effective in suppressing the growth of BRAF V600E mutant melanoma
cells (11). Treatment with CSPG4
specific anti-225D9+-TT Abs showed that the growth of
518A2 cells [BRAF(V600E)/CSPG4+] was inhibited by
32±5.9% compared to M14 cells [BRAF(V600E)/ CSPG4−]
(Fig. 3A). When combining
vemurafenib with anti-225D9+-TT Abs, this led to an
additional growth inhibition of the 518A2
[BRAF(V600E)/CSPG4+] by 30±6.0% on day 8 compared to the
CSPG4 negative cell line M14 (Fig.
3B). These results showed that targeting the CSPG4 protein
alone and in combination with vemurafenib influenced the
proliferation rate of 518A2 melanoma cells.
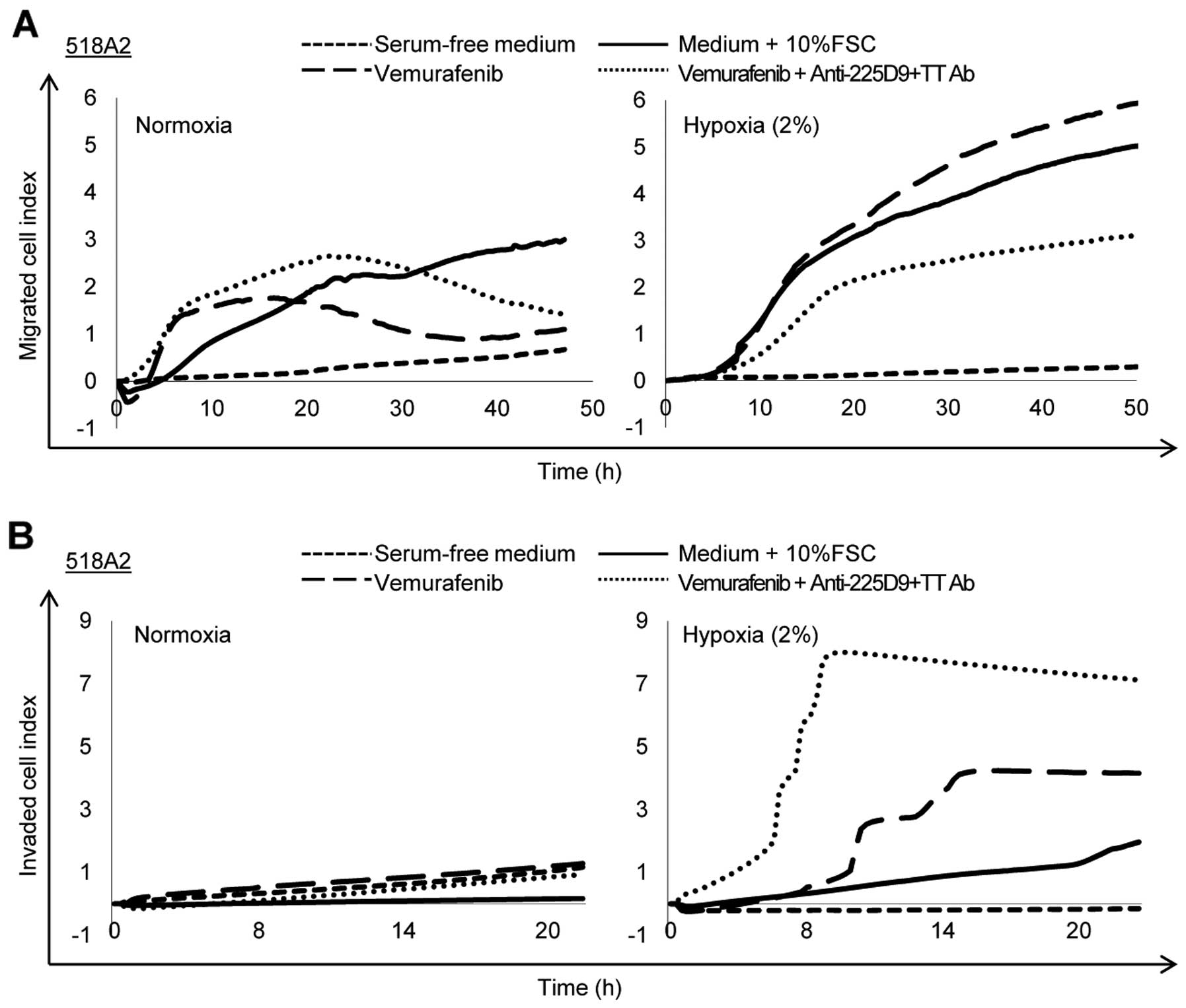
Influence of vemurafenib and
anti-225D9+-TT Abs on the Transwell migration and
Transwell invasion capacity of 518A2 melanoma cells
In order to measure the tumorigenicity of the 518A2
melanoma cells in hypoxic and normoxic conditions, a Transwell
migration assay using the RTCA DP Analyzer was performed (Fig. 4A). In normoxic conditions,
untreated 518A2 cells migrated relatively slowly but continuously
and reached a cell index of 3.3 after 50 h. In the presence of
vemurafenib, the cells initially migrated and reached a cell index
of 1.8 after 15 h, and then their migration index decreased to 0.9
after 38 h. The combination of vemurafenib and
anti-225D9+-TT Abs had a similar effect on the course of
migration, since the cell index initially increased to 2.6 after 22
h and then decreased to 1.6 after 46 h. As expected, in hypoxic
conditions the cell index was markedly increased in untreated 518A2
melanoma cells, where it reached a value of 5.0 after 52 h. The
migration capacity of vemurafenib treated melanoma cells was even
slightly increased and reached a cell index of 6.0 when compared to
untreated cells. In contrast, vemurafenib together with
anti-225D9+-TT Abs decreased the migration capacity of
518A2 cells, reaching a cell index of 3.0 after 50 h.
To test the invasive potential of 518A2 melanoma
cells, a Transwell matrigel invasion assay was performed. In
normoxic conditions, untreated 518A2 cells did not show any
invasion capacity, reaching a cell index of 0.1 after 22 h
(Fig. 4B). Vemurafenib or
vemurafenib together with anti-225D9+-TT Abs increased
the cell indices slightly to 0.85 and 1.4, respectively. However,
in hypoxic conditions the invasion capacity of untreated 518A2
melanoma cells increased and the cell index reached 1.4 after 22 h.
Surprisingly, when the cells were exposed to vemurafenib or
vemurafenib and anti-225D9+-TT Abs, the effect was
intensified with cell indices reaching 4.0 and 7.2 after 22 h,
respectively.
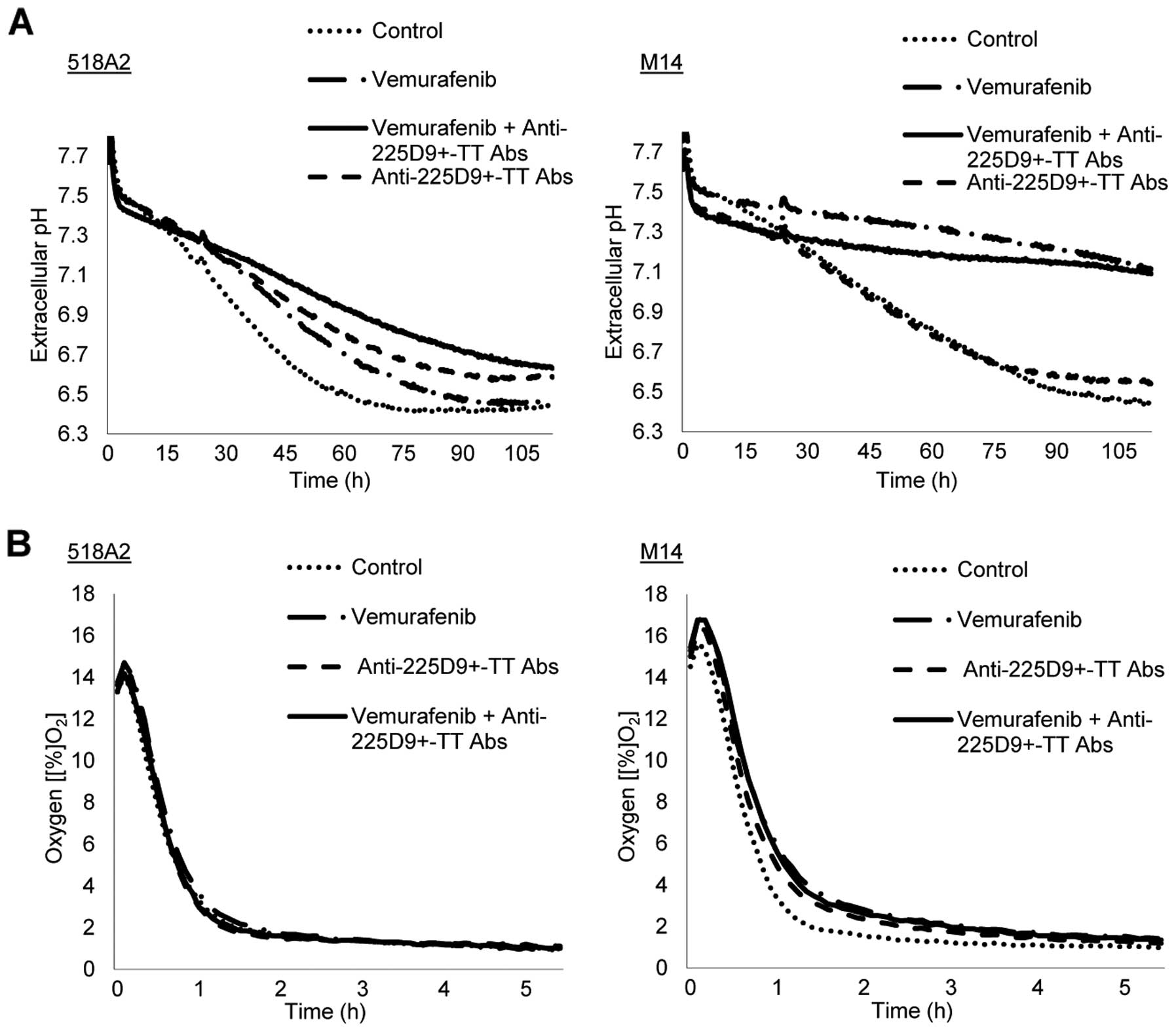
Anti-225D9+-TT Abs influence
the metabolism in hypoxic, vemurafenib-treated melanoma cells
Untreated 518A2 cells showed a decrease in the
extracellular pH. After adding vemurafenib,
anti-225D9+-TT Abs or a combination of both, the
extracellular pH decreased more slowly compared to untreated 518A2
cells (Fig. 5A and Table I). No change was observed in oxygen
consumption between treated and untreated 518A2 cells (Fig. 5B). In contrast, no
anti-225D9+-TT Abs-induced shift in extracellular pH was
observed in the CSPG4 negative M14 melanoma cell line (Fig. 5A and Table I). A minor change for oxygen
consumption from 2 to 4% was observed in this cell line in all
treatment modalities (Fig.
5B).
 | Table IExtracellular pH measured 75 h after
the administration of vemurafenib and anti-225D9+-TT Abs
to hypoxic 518A2 melanoma cells [BRAF(V600E)/CSPG4+] and
hypoxic M14 melanoma cells [BRAF(V600E)/CSPG4−]. |
Table I
Extracellular pH measured 75 h after
the administration of vemurafenib and anti-225D9+-TT Abs
to hypoxic 518A2 melanoma cells [BRAF(V600E)/CSPG4+] and
hypoxic M14 melanoma cells [BRAF(V600E)/CSPG4−].
| Cells | Control | Vemurafenib |
Anti-225D9+-TT Abs | Vemurafenib and
Anti-225D9+-TT Abs |
|---|
| 518A2
[BRAF(V600E)/CSPG4+] | 6.4 | 6.6 | 6.7 | 6.8 |
| M14
[BRAF(V600E)/CSPG4−] | 6.6 | 7.3 | 6.6 | 7.2 |
Vemurafenib and anti-225D9+-TT
Abs differently affect multiple signaling pathways in normoxic and
hypoxic conditions
To understand the molecular heterogeneity in
melanoma we compared the effects on signaling pathways important to
cell growth, migration, invasion and survival in 518A2 melanoma
cells after treatment with vemurafenib in normoxic and hypoxic
conditions. In normoxic conditions, vemurafenib treatment of the
melanoma cell line 518A2 resulted in a decreased protein expression
level of pERK1/2 (Thr202/Tyr204) (Fig.
6B) and decreased expression levels of pFAK (Tyr397), FAK and
AKT compared to untreated 518A2 cells (Fig. 6A). However, in hypoxic conditions
untreated melanoma cells showed an increased protein expression of
FAK and AKT compared to untreated cells in normoxia. The expression
levels of PKCα and pAKT (Ser473) increased in vemurafenib-treated
and untreated hypoxic 518A2 cells compared to normoxic cells
(Fig. 6A).
To understand the molecular mechanisms underlying
the modification of vemurafenib-mediated effects when combined with
anti-225D9+-TT Abs, AKT and ERK signaling pathways were
analyzed. The phosphorylation level of AKT (Ser473) increased in
hypoxic conditions. However, pAKT (Ser473) slightly decreased after
the treatment anti-225D9+-TT Abs and was decreased even
more when vemurafenib was combined with anti-225D9+-TT
Abs. No change was observed in the phosphorylation level of ERK1/2
(Thr202/Tyr204) (Fig. 6B).
Discussion
Melanoma is one of the most aggressive skin cancers.
The poor efficiency of available therapies demands to find new
therapeutic strategies to improve patient survival rates and to
overcome resistance to currently used drugs (8,34).
Oncogenic mutations within the MAPK pathway are frequent in
melanoma and targeting of MAPK signaling has yielded significant
responses in a large number of patients that last for several
months before relapsing (35).
One of the factors which contributes to melanoma
progression is hypoxia through HIF-mediated molecular responses.
HIF1 promotes the upregulation of genes which control a series of
metabolic changes in tumor cells as well as increasing their
invasive properties (36,37). Thereby hypoxia significantly
affects tumor phenotype and in many tumor types is associated with
therapy resistance (38,39). In melanoma, hypoxia is a
microenvironmental stimulus that triggers a switch from a
proliferative to an invasive cell phenotype that is less sensitive
to therapies (21,26). In our studies, hypoxia did not
influence the proliferative capacity of 518A2 melanoma cells,
whereas hypoxia enhanced the antiproliferative capacity of
vemurafenib in a time-dependent manner (Fig. 1A). One of the mediators of hypoxic
responses in many cell types is the HIF1α protein (36,37).
Vemurafenib markedly reduced the protein expression of this hypoxic
marker in 518A2 melanoma cells and this downregulation also
contributed to lower expression levels of CAIX (Fig. 1C). This was not reflected on the
mRNA level of HIF1α, suggesting that the transcription of HIF1α is
not influenced by vemurafenib (Fig.
1B). Widmer et al (26)
showed that knock-down of HIF1α under hypoxic conditions
decreased the invasion capacity of melanoma cells. We demonstrated
that reduced HIF1α expression was responsible for enhanced
Transwell migration and invasion despite unchanged HIF1α
mRNA levels (Figs. 1B, and
4). CAIX is a hypoxia-inducible,
tumor-associated member of the human α-CA family (40). CAIX shows only limited expression
in normal tissues but its expression is highly elevated in various
cancers such as colorectal and lung carcinomas (41,42)
and has not been shown in melanomas (43). To the best of our knowledge, we
demonstrate for the first time CAIX expression and its
downregulation by vemurafenib in 518A2 melanoma cells (Fig. 1C). We also observed CAIX staining
in 518A2 xenografts in many nested tumor regions, preferable near
necrotic areas co-localized with HIF1α (Fig. 2).
Clinical evidence has already shown that melanoma
regression is rarely complete after 6–9 months of therapy with
vemurafenib (7). This is partly
due to acquired resistance that leads to melanoma progression and a
boost in aggressiveness of the disease (34) but also to the tumor
microenvironment and the fact that melanomas are composed of
heterogeneous zones containing proliferative and quiescent cells
(44). A combined approach with
vemurafenib and inhibitors targeting other signaling pathways
determined to be abnormal in melanoma cells would be preferential.
Therefore, we combined vemurafenib with antibodies directed against
CSPG4 and evaluated their therapeutic potential in normoxic and
hypoxic conditions. The CSPG4-specific mAb 225.28S was shown to
inhibit the growth and recurrence of melanoma in mice grafted with
CSPG4-expressing human melanoma cell lines (16) and the antibodies induced by a
mimotope vaccine (anti-225D9+-TT Abs) directed against
CSPG4 were capable of inhibiting melanoma cell growth in a SCID
mouse model (18,19). In line with these data we found
that anti-225D9+-TT Abs were effective in normoxic
conditions against proliferative cells and reduced the growth of
human melanoma 518A2 cells by 30% (Fig. 3A). Here we show that the
combination of vemurafenib and anti-225D9+-TT Abs could
reduce the growth of 518A2 cells by additional 30% (Fig. 3B). Comparable results were achieved
with the CSPG4-specific mAb 225.28 (17).
This finding provided the rationale for the strategy
described in this report that combined vemurafenib with antibodies
directed against CSPG4 as a means of enhancing the magnitude of the
BRAF inhibitor responses that could also influence migration and
invasion. There are varying results regarding the effect of MAPK
pathway inhibition on invasion and migration in the literature
(44,45). We tracked 518A2 cells in the
presence of vemurafenib and anti-225D9+-TT Abs for long
periods of time within a hypoxic microenvironment and obtained a
longitudinal track for invasion and migration. We showed that
Transwell migration of 518A2 cells toward a chemoattractant was
markedly increased in hypoxic conditions and even more after the
treatment with vemurafenib. This effect was partly blocked by
anti-225D9+-TT Abs (Fig.
4A) reflecting the role of CSPG4 in cell migration (12). However, the addition of
anti-225D9+-TT Abs markedly enhanced the invasive
capacity of vemurafenib-treated, hypoxic 518A2 cells (Fig. 4B).
Our data imply that hypoxia is important in
determining the effect of targeting CSPG4 on cell migration and
invasion. Since hypoxia is associated with changes in the
extracellular pH and related to the acidification of the tumor
microenvironment that promotes cancer cell invasion (46), we were able to show that
vemurafenib increased the extracellular pH in 518A2 cells, an
effect that was diminished by the addition of
anti-225D9+-TT Abs (Fig.
5A and Table I). This was not
reflected in the oxygenation levels (Fig. 5B) underlining the role of CSPG4
irrespective of oxygenation in tumor microenvironment and showing
its potential as a therapeutic target. In a next step we focused on
the capacity of vemurafenib and anti-225D9+-TT Ab to
block multiple signaling pathways in hypoxic and normoxic
conditions. Kumar et al (30) showed that the BRAF inhibitor
sorafenib inhibited ERK phosphorylation and suggested that some of
the effect of this compound was mediated through HIF1α inhibition.
Hypoxia per se influenced multiple signaling pathways such
as pAKT (Ser473), AKT and PKCα (Fig.
6A). In this study we found that the expression of pAKT
(Ser473) decreased after treatment with vemurafenib and
anti-225D9+-TT Abs in hypoxic conditions (Fig. 6B) confirming that hypoxia affected
the response to vemurafenib and anti-225D9+-TT Abs in
518A2 melanoma cells.
In this study we showed that targeting CSPG4 in
melanoma cells enhanced antiproliferative effects of vemurafenib in
normoxic conditions and reduced the migratory capacity in hypoxic
conditions. Hypoxia strongly influenced the response to vemurafenib
treatment in melanoma cells which switched to a more invasive and
aggressive phenotype. Therefore, therapeutic efforts will have to
consider that the microenvironment of melanoma cells has an impact
on tumor progression.
Acknowledgements
This project is a part of the EU Marie Curie Initial
Training Network (ITN) Biomedical engineering for cancer and brain
disease diagnosis and therapy development (EngCaBra). Project no.
PITN-GA-2010-264417.
Abbreviations:
|
CSPG4
|
chondroitin sulfate proteoglycan 4
|
|
HIF1α
|
hypoxia-inducible factor 1α
|
|
CAIX
|
carbonic anhydrase IX
|
|
DMOG
|
dimethyloxalylglycine,
N-(methoxyoxoacetyl)-glycine methyl ester
|
|
PVDF
|
polyvinylidene fluoride
|
References
|
1
|
MacKie RM, Hauschild A and Eggermont AMM:
Epidemiology of invasive cutaneous melanoma. Ann Oncol. 20(Suppl
6): vi1–vi7. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Linos E, Swetter SM, Cockburn MG, Colditz
GA and Clarke CA: Increasing burden of melanoma in the United
States. J Invest Dermatol. 129:1666–1674. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Testori A, Rutkowski P, Marsden J,
Bastholt L, Chiarion-Sileni V, Hauschild A and Eggermont AM:
Surgery and radiotherapy in the treatment of cutaneous melanoma.
Ann Oncol. 20(Suppl 6): vi22–vi29. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kee D and McArthur G: Targeted therapies
for cutaneous melanoma. Hematol Oncol Clin North Am. 28:491–505.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gogas H, Polyzos A and Kirkwood J:
Immunotherapy for advanced melanoma: Fulfilling the promise. Cancer
Treat Rev. 39:879–885. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chapman PB, Hauschild A, Robert C, Haanen
JB, Ascierto P, Larkin J, Dummer R, Garbe C, Testori A, Maio M, et
al; BRIM-3 Study Group. Improved survival with vemurafenib in
melanoma with BRAF V600E mutation. N Engl J Med. 364:2507–2516.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Flaherty KT, Puzanov I, Kim KB, Ribas A,
McArthur GA, Sosman JA, O’Dwyer PJ, Lee RJ, Grippo JF, Nolop K, et
al: Inhibition of mutated, activated BRAF in metastatic melanoma. N
Engl J Med. 363:809–819. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fedorenko IV, Paraiso KH and Smalley KS:
Acquired and intrinsic BRAF inhibitor resistance in BRAF V600E
mutant melanoma. Biochem Pharmacol. 82:201–209. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sun C, Wang L, Huang S, Heynen GJ,
Prahallad A, Robert C, Haanen J, Blank C, Wesseling J, Willems SM,
et al: Reversible and adaptive resistance to BRAF(V600E) inhibition
in melanoma. Nature. 508:118–122. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kugel CH III and Aplin AE: Adaptive
resistance to RAF inhibitors in melanoma. Pigment Cell Melanoma
Res. 27:1032–1038. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kudchadkar R, Paraiso KH and Smalley KS:
Targeting mutant BRAF in melanoma: Current status and future
development of combination therapy strategies. Cancer J.
18:124–131. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Price MA, Colvin Wanshura LE, Yang J,
Carlson J, Xiang B, Li G, Ferrone S, Dudek AZ, Turley EA and
McCarthy JB: CSPG4, a potential therapeutic target, facilitates
malignant progression of melanoma. Pigment Cell Melanoma Res.
24:1148–1157. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mittelman A, Chen ZJ, Liu CC, Hirai S and
Ferrone S: Kinetics of the immune response and regression of
metastatic lesions following development of humoral anti-high
molecular weight-melanoma associated antigen immunity in three
patients with advanced malignant melanoma immunized with mouse
anti-idiotypic monoclonal antibody MK2-23. Cancer Res. 54:415–421.
1994.PubMed/NCBI
|
|
14
|
Ferris RL, Jaffee EM and Ferrone S: Tumor
antigen-targeted, monoclonal antibody-based immunotherapy: Clinical
response, cellular immunity, and immunoescape. J Clin Oncol.
28:4390–4399. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang X, Osada T, Wang Y, Yu L, Sakakura K,
Katayama A, McCarthy JB, Brufsky A, Chivukula M, Khoury T, et al:
CSPG4 protein as a new target for the antibody-based immunotherapy
of triple-negative breast cancer. J Natl Cancer Inst.
102:1496–1512. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hafner C, Breiteneder H, Ferrone S,
Thallinger C, Wagner S, Schmidt WM, Jasinska J, Kundi M, Wolff K,
Zielinski CC, et al: Suppression of human melanoma tumor growth in
SCID mice by a human high molecular weight-melanoma associated
antigen (HMW-MAA) specific monoclonal antibody. Int J Cancer.
114:426–432. 2005. View Article : Google Scholar
|
|
17
|
Yu L, Favoino E, Wang Y, Ma Y, Deng X and
Wang X: The CSPG4-specific monoclonal antibody enhances and
prolongs the effects of the BRAF inhibitor in melanoma cells.
Immunol Res. 50:294–302. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wagner S, Hafner C, Allwardt D, Jasinska
J, Ferrone S, Zielinski CC, Scheiner O, Wiedermann U, Pehamberger H
and Breiteneder H: Vaccination with a human high molecular weight
melanoma-associated antigen mimotope induces a humoral response
inhibiting melanoma cell growth in vitro. J Immunol. 174:976–982.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wagner S, Krepler C, Allwardt D, Latzka J,
Strommer S, Scheiner O, Pehamberger H, Wiedermann U, Hafner C and
Breiteneder H: Reduction of human melanoma tumor growth in severe
combined immunodeficient mice by passive transfer of antibodies
induced by a high molecular weight melanoma-associated antigen
mimotope vaccine. Clin Cancer Res. 14:8178–8183. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Walsh JC, Lebedev A, Aten E, Madsen K,
Marciano L and Kolb HC: The clinical importance of assessing tumor
hypoxia: Relationship of tumor hypoxia to prognosis and therapeutic
opportunities. Antioxid Redox Signal. 21:1516–1554. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
O’Connell MP, Marchbank K, Webster MR,
Valiga AA, Kaur A, Vultur A, Li L, Herlyn M, Villanueva J, Liu Q,
et al: Hypoxia induces phenotypic plasticity and therapy resistance
in melanoma via the tyrosine kinase receptors ROR1 and ROR2. Cancer
Discov. 3:1378–1393. 2013. View Article : Google Scholar
|
|
22
|
Vergis R, Corbishley CM, Norman AR,
Bartlett J, Jhavar S, Borre M, Heeboll S, Horwich A, Huddart R,
Khoo V, et al: Intrinsic markers of tumour hypoxia and angiogenesis
in localised prostate cancer and outcome of radical treatment: A
retrospective analysis of two randomised radiotherapy trials and
one surgical cohort study. Lancet Oncol. 9:342–351. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mujcic H, Hill RP, Koritzinsky M and
Wouters BG: Hypoxia signaling and the metastatic phenotype. Curr
Mol Med. 14:565–579. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ghislin S, Deshayes F, Middendorp S,
Boggetto N and Alcaide-Loridan C: PHF19 and Akt control the switch
between proliferative and invasive states in melanoma. Cell Cycle.
11:1634–1645. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Erler JT, Bennewith KL, Nicolau M,
Dornhöfer N, Kong C, Le QT, Chi JT, Jeffrey SS and Giaccia AJ:
Lysyl oxidase is essential for hypoxia-induced metastasis. Nature.
440:1222–1226. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Widmer DS, Hoek KS, Cheng PF, Eichhoff OM,
Biedermann T, Raaijmakers MI, Hemmi S, Dummer R and Levesque MP:
Hypoxia contributes to melanoma heterogeneity by triggering
HIF1α-dependent phenotype switching. J Invest Dermatol.
133:2436–2443. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hanna SC, Krishnan B, Bailey ST, Moschos
SJ, Kuan PF, Shimamura T, Osborne LD, Siegel MB, Duncan LM, O’Brien
ET III, et al: HIF1α and HIF2α independently activate SRC to
promote melanoma metastases. J Clin Invest. 123:2078–2093. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mouriaux F, Sanschagrin F, Diorio C,
Landreville S, Comoz F, Petit E, Bernaudin M, Rousseau AP, Bergeron
D and Morcos M: Increased HIF-1α expression correlates with cell
proliferation and vascular markers CD31 and VEGF-A in uveal
melanoma. Invest Ophthalmol Vis Sci. 55:1277–1283. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ditte P, Dequiedt F, Svastova E, Hulikova
A, Ohradanova-Repic A, Zatovicova M, Csaderova L, Kopacek J,
Supuran CT, Pastorekova S, et al: Phosphorylation of carbonic
anhydrase IX controls its ability to mediate extracellular
acidification in hypoxic tumors. Cancer Res. 71:7558–7567. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kumar SM, Yu H, Edwards R, Chen L,
Kazianis S, Brafford P, Acs G, Herlyn M and Xu X: Mutant V600E BRAF
increases hypoxia inducible factor-1alpha expression in melanoma.
Cancer Res. 67:3177–3184. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Limame R, Wouters A, Pauwels B, Fransen E,
Peeters M, Lardon F, De Wever O and Pauwels P: Comparative analysis
of dynamic cell viability, migration and invasion assessments by
novel real-time technology and classic endpoint assays. PLoS One.
7:e465362012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chiche J, Brahimi-Horn MC and Pouysségur
J: Tumour hypoxia induces a metabolic shift causing acidosis: A
common feature in cancer. J Cell Mol Med. 14:771–794. 2010.
View Article : Google Scholar
|
|
33
|
Pastoreková S, Závadová Z, Kostál M,
Babusíková O and Závada J: A novel quasi-viral agent, MaTu, is a
two-component system. Virology. 187:620–626. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Trunzer K, Pavlick AC, Schuchter L,
Gonzalez R, McArthur GA, Hutson TE, Moschos SJ, Flaherty KT, Kim
KB, Weber JS, et al: Pharmacodynamic effects and mechanisms of
resistance to vemurafenib in patients with metastatic melanoma. J
Clin Oncol. 31:1767–1774. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Cheng Y, Zhang G and Li G: Targeting MAPK
pathway in melanoma therapy. Cancer Metastasis Rev. 32:567–584.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Irwin DC, McCord JM, Nozik-Grayck E,
Beckly G, Foreman B, Sullivan T, White M, T Crossno J Jr, Bailey D,
Flores SC, et al: A potential role for reactive oxygen species and
the HIF-1alpha-VEGF pathway in hypoxia-induced pulmonary vascular
leak. Free Radic Biol Med. 47:55–61. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Dewhirst MW: Relationships between cycling
hypoxia, HIF-1, angiogenesis and oxidative stress. Radiat Res.
172:653–665. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Liao J, Qian F, Tchabo N,
Mhawech-Fauceglia P, Beck A, Qian Z, Wang X, Huss WJ, Lele SB,
Morrison CD, et al: Ovarian cancer spheroid cells with stem
cell-like properties contribute to tumor generation, metastasis and
chemotherapy resistance through hypoxia-resistant metabolism. PLoS
One. 9:e849412014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Azab AK, Hu J, Quang P, Azab F,
Pitsillides C, Awwad R, Thompson B, Maiso P, Sun JD, Hart CP, et
al: Hypoxia promotes dissemination of multiple myeloma through
acquisition of epithelial to mesenchymal transition-like features.
Blood. 119:5782–5794. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
McDonald PC, Winum JY, Supuran CT and
Dedhar S: Recent developments in targeting carbonic anhydrase IX
for cancer therapeutics. Oncotarget. 3:84–97. 2012.PubMed/NCBI
|
|
41
|
Ivanov S, Liao SY, Ivanova A,
Danilkovitch-Miagkova A, Tarasova N, Weirich G, Merrill MJ,
Proescholdt MA, Oldfield EH, Lee J, et al: Expression of
hypoxia-inducible cell-surface transmembrane carbonic anhydrases in
human cancer. Am J Pathol. 158:905–919. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chrastina A, Závada J, Parkkila S, Kaluz
S, Kaluzová M, Rajcáni J, Pastorek J and Pastoreková S:
Biodistribution and pharmacokinetics of 125I-labeled monoclonal
antibody M75 specific for carbonic anhydrase IX, an intrinsic
marker of hypoxia, in nude mice xenografted with human colorectal
carcinoma. Int J Cancer. 105:873–881. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Syrjänen L, Luukkaala T, Leppilampi M,
Kallioinen M, Pastorekova S, Pastorek J, Waheed A, Sly WS, Parkkila
S and Karttunen T: Expression of cancer-related carbonic anhydrases
IX and XII in normal skin and skin neoplasms. APMIS. 122:880–889.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Haass NK, Beaumont KA, Hill DS, Anfosso A,
Mrass P, Munoz MA, Kinjyo I and Weninger W: Real-time cell cycle
imaging during melanoma growth, invasion, and drug response.
Pigment Cell Melanoma Res. 27:764–776. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Vultur A, Villanueva J, Krepler C, Rajan
G, Chen Q, Xiao M, Li L, Gimotty PA, Wilson M, Hayden J, et al: MEK
inhibition affects STAT3 signaling and invasion in human melanoma
cell lines. Oncogene. 33:1850–1861. 2014. View Article : Google Scholar
|
|
46
|
Sedlakova O, Svastova E, Takacova M,
Kopacek J, Pastorek J and Pastorekova S: Carbonic anhydrase IX, a
hypoxia-induced catalytic component of the pH regulating machinery
in tumors. Front Physiol. 4:4002014. View Article : Google Scholar : PubMed/NCBI
|