Introduction
Head and neck squamous cell carcinoma (HNSCC) with
>600,000 newly diagnosed cases per year is the sixth most common
cancer in the world (1). In
developed countries the average 5-year survival rate is ~50%
(2). Improvements during the past
decades in radio- and chemotherapy, and recently introduced
targeted antibody therapy and improved surgical procedures have
resulted in a significantly enhanced quality of life for cancer
patients. However, local and distant recurrence is common in HNSCC
and during the past years rates of survival have only marginally
improved (3).
According to the cancer stem cell (CSC) theory,
tumor formation, relapse after therapy and the development of
metastases is thought to involve CSC in different phenotypic and
functional states. This particular subset of cells was named cancer
stem (like) cells (CSCs), similar to that conceptualised in normal
tissue as stem cells (4). To date,
the existence of CSCs have been identified in several human
hematologic and solid tumors, including leukemia, bladder cancer,
breast cancer, colon carcinoma, HNSCC and pancreatic cancer
(5–9). The CSC theory suggests that only a
subpopulation of cells in a tumor has the ability to self-renew,
differentiate and regenerate similar tumors, as demonstrated in
vitro (10–13) and by xeno-transplantation into
immuno-deficient mice (14). These
cells display also some resistance to conventional therapy
(11,12).
Spheroid culture is an effective and convenient way
to enrich the CSC population present in the cancer cell population
and is thus increasingly being used, sometimes in combination with
further enrichment by specific markers, to evaluate functional stem
cell activity in normal tissue and putative CSC. The ability of CSC
to form spheroids was first observed in cells derived from the
central nervous system in 1992 (15). Todaro et al found that cells
isolated from the striatum of adult mouse brain could be clonally
expanded by culturing spheres, and the cells generated both
astrocytes and neurons (16). The
ability of isolated cells to form anchorage-independent spheres in
culture was demonstrated in breast, colon, HNSCC and melanoma
(6,7,17).
In HNSCC cell lines, CSC cultured with this method showed a high
level of CSC-marker expression such as CD44, CD133 and ALDH
(10,18,19).
Epithelial to mesenchymal transition (EMT) is a key
event in metastasis and increasing evidence suggests there is a
link between EMT and CSCs (20–23)
that has also been demonstrated for HNSCC (10). Mani et al (22) and Hindriksen and Bijlsma (24) reported that induction of an EMT in
human mammary epithelial cells led to an increased expression of
CSC markers and in pancreatic cancer the occurrence of EMT is often
accompanied by the activation of CSC-related pathways. EMT is a
physiological process that plays an important role during embryonic
development and morphogenesis (25). Important characteristics of EMT are
a loss of polarity of the epithelial cells, loss of intercellular
contacts, acquisition of mesenchymal features and phenotype along
with an increased cellular motility (23). Therefore, during EMT the expression
of epithelial markers, such as E-cadherin decreases while the
expression of mesenchymal markers, such as N-cadherin, vimentin and
α-smooth muscle actin increases (10,25,26).
In recent years it has been demonstrated that EMT is involved in
the development of metastasis by increased cellular motility and
invasiveness (23,25,27)
resulting in a more metastatic phenotype (28,29).
During the EMT process, several changes in gene expression, such as
increased Snail1, Snail2 and Twist transcription factors can be
observed (23,30). We and others have demonstrated a
link between EMT and CSCs. For example the expression of the
EMT-marker Snail1/2 or Twist is increased in CSC, that are involved
in the loss of the epithelial phenotype and the acquisition of the
mesenchymal phenotype (10,31–33).
In the past few years, it became evident that the induction of EMT
not only promotes tumor cell metastasis and invasion, but it is
also involved in drug resistance and enrichment of CSCs (21–23,34).
MiRs are a family of 21–25 nucleotide long,
non-coding endogenous RNAs, that act by binding to the target mRNA
3′-untranslated region inducing mRNA degradation or repression of
mRNA translation (35). A single
miR may regulate several mRNAs and one mRNA can be targeted by
dozens of miRs. Although small, miRs play an essential role in
biological processes, such as development, metastasis,
proliferation and apoptosis (35).
Early investigations demonstrated that miRs control embryonic stem
cell (ESC) properties including self-renewal and differentiation.
Later studies have shown that abnormal expression/functions of miRs
are involved in tumorigenesis (36).
MiR34a is a tumor suppressor which directly targets
p53 on a post-transcriptional level. In p53-deficient human
pancreatic cancer cells, overexpression of miR-34a inhibited cell
proliferation, cell cycle progression, self-renewal, EMT and
invasion, indicating that miR-34a may restore p53 function,
potentially directly via the downstream targets Notch and Bcl-2
which are involved in CSC differentiation and self-renewal
(37). In prostate cancer, miR-34a
was reported to directly repress CSC properties and metastasis. In
breast cancer, ectopic miR-34a expression reduced CSC properties
and enhanced sensitivity to chemical treatment. Thus, miR-34a is
considered a tumor suppressor which represses ‘stemness’ features
and function. In HNSCC cell lines, miR34a was found downregulated
in HNSCC and -lines (38).
However, miR-34a expression and function of in HNSCC is not clear
yet.
Materials and methods
Cell lines
Eight HNSCC cell lines were used: UD-SCC1, −2 (gift
of Henning Bier, University of Munich, Germany); UM-SCC9, −11B,
−47, −104 (University of Michigan, Tom Carey, MI, USA) and UT-SCC33
(University of Turku, Reidar A. Grenman, Finland), 93VU147T (VU
Medical Center, John P. de Winter, Amsterdam, The Netherlands). The
HPV types were as follows: HPV+: UD-SCC2, UM-SCC47,
UM-SCC104, 93VU147T; all HPV type 16+. HPV−:
UD-SCC1, UM-SCC9, UM-SCC11B, UT-SCC33. All cell lines were
regularly tested for mycoplasma and found free of any
contamination.
Spheroid culture
Adherent monolayer cells were grown in 75
cm2 cell culture flasks (BD Science, Franklin Lakes, NJ,
USA) in DMEM (Invitrogen, Heidelberg, Germany) supplemented with
10% heat inactivated fetal bovine serum (FBS; Biochrom, Berlin,
Germany) and 1% penicillin/streptomycin (10,000 U/ml and 10,000
μg/ml, respectively; Biochrom) at 37°C in humidified atmosphere
with 5% CO2, until 70–80% confluence. Cells were washed
with PBS twice and detached using trypsin/EDTA solution (T/E;
Biochrom). The reaction was stopped by adding complete culture
medium. After centrifugation at 200 × g for 5 min, cells were
resuspended in serum-free Quantum 263 (PAA), supplemented with 10
ng/ml EGF and 10 ng/ml βFGF (Biochrom). To generate spheroids,
single cells were plated in cell culture dishes (BD Falcon™, 100×20
mm) with a coated surface with 1% agarose at a specific density of
2×104 cells/ml. Cells were maintained at 37°C in
humidified atmosphere with 5% CO2 content.
Every three to four days, half of the medium was
replaced. After 5–7 days, spheroid formation was checked and
representative images were taken. For passaging or following
experiments, culture medium with spheroids was removed from the
dish and filtered through a 40-μm mesh (BD Biosciences, Heidelberg,
Germany) to collect the spheroids. For experiments, typically 2nd
or higher generation were used.
Aldefluor assay and FACS sorting
The ALDH activity of SDC and MDC was determined by
using the Aldefluor assay kit (Stem Cell Technologies, Durham, NC,
USA). For collection of the spheroids a 40 μm mesh was used.
Subsequently cells were dissociated into single cells by
trypsin/EDTA digestion for 10 min at 37°C followed by 30 times up
and down pipetting using a 1,000 μl pipette tip. Then, the
single-cell suspension was washed twice in PBS without
Ca2+/Mg2+ and suspended in 1 ml Aldefluor
assay buffer containing 5 μl ALDH substrate (1 ml/per
1×106 cells) and incubated for 30–40 min at 37°C in the
dark. As a negative control, for each sample, an aliquot was
treated with 5 μl diethylaminobenzaldehyde (DEAB; 50 mmol/l), a
specific ALDH inhibitor. Following incubation, all samples were
centrifuged for 5 min at 250 × g and the supernatant was removed.
After washing twice with buffer, cells were maintained in ALDH
buffer on ice during all subsequent procedures.
For FACS sorting, cells were resuspended in PBS
buffer at 1×107 cells per ml and run on an Aria cell
sorter (BD Biosciences). The sorting gates were established, by
negative control cells which were treated with the ALDH inhibitor
DEAB.
Quantitative real-time PCR
To quantify the expression of mRNA, total RNA was
isolated by TRIzol reagent (Life Technologies, Darmstadt, Germany)
according to the manufacturer's instructions and then cDNA was
prepared using the Omniscript First-Strand synthesis system
(Qiagen, Hilden, Germany) random primers (Qiagen). RT-qPCRs were
carried out using ABI Power SYBR Green mix (ABI, Applied Biosystems
Inc, Foster City, CA, USA) and run on a Bio-Rad Chromo 4 (Bio-Rad,
München, Germany). GAPDH was the reference gene.
For the detection of miR-34a, TRIzol reagent was
used to extract total RNA according to the manufacturer's
instructions. Poly A tailing and cDNA synthesis were performed by
using the Ncode VILO miR cDNA (Invitrogen, Carlsbad, CA, USA)
synthesis kit. The RT-qPCR analysis was performed on a Bio-Rad
Chromo 4 (Bio-Rad) with Express SYBR GreenER™ qPCR SuperMix
Universal (Invitrogen). GAPDH was the reference gene. Reactions
were carried out in triplicate with controls; primer sequences are
listed in Table I. The data were
statistically analyzed by the modified 2−ΔΔCt value
method (39).
 | Table IPrimer sequences used for RT-PCR
(5′-3′). |
Table I
Primer sequences used for RT-PCR
(5′-3′).
| Transcript
name | Forward primer
sequence | Reverse primer
sequence |
|---|
| Nanog | AAT ACC TCA GCC TCC
AGC AGA TG | TGC GTC ACA CCA TTG
CTA TTC TTC |
| Oct3/4 | GAC AGG GGG AGG GGA
GGA GCT AGG | CTT CCC TCC AAC CAG
TTG CCC CAA AC |
| Sox2 | GGG AAA TGG GAG GGG
TGC AAA AGA GG | TTG CGT GAG TGT GGA
TGG GAT TGG TG |
| Snail1 | GGC GCA CCT GCT CGG
GGA GTG | GCC GAT TCG CGC AGC
A |
| Snail2 | GGG GAG AAG CCT TTT
TCT TG | TCC TCA TGT TTG TGC
AGG AG |
| Twist | GGA GTC CGC AGT CTT
ACG AG | TCT GGA GGA CCT GGT
AGA GG |
| GAPDH
(reference) | AGC TCC CAA AAA TAG
ACG CAC | TTC ATA GCA GTA GGC
ACA AAG G |
| Hsa-miR-34a | TGG CAG TGT CTT AGC
TGG TTG T | |
Transfection of miRs
For transfection of miR mimics, spheroids were
harvested using a 40 μm mesh and dispersed into single cells as
described above. The single cell suspension was plated at a density
of 8×104 per well in 6-well ultra-low attachment plates
with complete medium. Then cells were transfected with 50 nmol/l
miR-34a mimic, negative control (NC) or MOCK control using the
Lipofectamine RNAiMAX reagent (Invitrogen) in antibiotic-free
Opti-MEM (Invitrogen) according to the manufacturer's instructions.
After 6 h, the medium was replaced with spheroid culture medium.
Twenty-four hours after transfection, cells were harvested and
processed for further analysis. The transfection efficiency was
determined by cell counting after visualization of transfected
cells with a BLOCK-iT™ Alexa Fluor Red Fluorescent Oligo using
fluorescence microscopy.
The expression of the CSCs marker ALDH was assessed
by FACS as described above at intervals of 24, 48 and 72 h after
transfection. Quantification of EMT- and CSC-related TF expression
by transfected SDC were carried out by RT-qPCR.
Invasion assay
Warm (37°C) culture medium was added to the interior
of the inserts and bottom of BD BioCoat Matrigel Invasion Chambers
(BD) and allowed to rehydrate for 2 h in a humidified tissue
culture incubator, at 37°C in 5% CO2 atmosphere. After
rehydration, the medium was carefully removed without disturbing
the layer of Matrigel™ matrix on the membrane. Next, cell
suspensions (SDC transfected with miR-34a mimics and controls) were
prepared in culture medium containing 5×104 cells/ml for
24-well chambers. DMEM (750 μl) containing 10% FBS serving as
chemoattractant was added to the wells of the plate. Sterile
forceps were used to transfer the chambers to the wells.
Immediately 0.5 ml of cell suspension (2.5×104 cells)
was added to the 24-well chambers that were then incubated for 24 h
in a humidified tissue culture incubator, at 37°C in 5%
CO2 atmosphere.
After incubation, the non-invading cells were
removed from the upper surface of the membrane by gentle scrubbing
and the cells on the lower surface of the membrane were stained
with Giemsa. Cell counting was facilitated by photographing the
membrane through the microscope and 3 fields per membrane of
triplicate membranes were counted under ×200 magnification
(Axiovert, Axiovision, Zeiss, Germany).
Colony formation assay and spheroid
formation assay
A colony formation assay was used to assess the
clonogenicity of miR-34a mimic-transfected UM-SCC9 cells sorted
according to ALDH+ and ALDH−. After FACS
sorting, the ALDH+ and ALDH− cells were
placed in 6-well plates for transfection with miR-34a mimics.
After FACS sorting, 800 ALDH+- and
ALDH− cells, respectively, were inoculated into
Ultra-low attachment 24-well plates (Corning, NY, USA) for
transfection with miR-34a mimics, respectively. Cells were
resuspended in serum-free Quantum 263 medium (Biochrom AG),
supplemented with 10 ng/ml EGF and 10 ng/ml bFGF (Biochrom). Fresh
medium containing growth factors was added each week for 2
weeks.
Two weeks later, colonies were visualized by
staining with Giemsa and viable colonies that contained >50
cells or were >0.1 mm in size were counted with an ocular
micrometer. The clone formation rate was calculated according to
the formula: Clone formation rate = number of formed colony/number
of seeded cells × 100%.
Statistical analysis
For statistical evaluation of flow cytometric
results, SPSS software (version 22; SPSS, Chicago, IL, USA) was
used. Student's t-test was used to analyze statistical significance
of the data.
For all RT-qPCR data, the expression analysis was
performed using the modified ΔΔCt value method. Expression analysis
and statistical evaluations was carried out by using the pair-wise
fixed re-allocation randomization by Qiagen REST 2009 software
(version 2.0.13).
Results
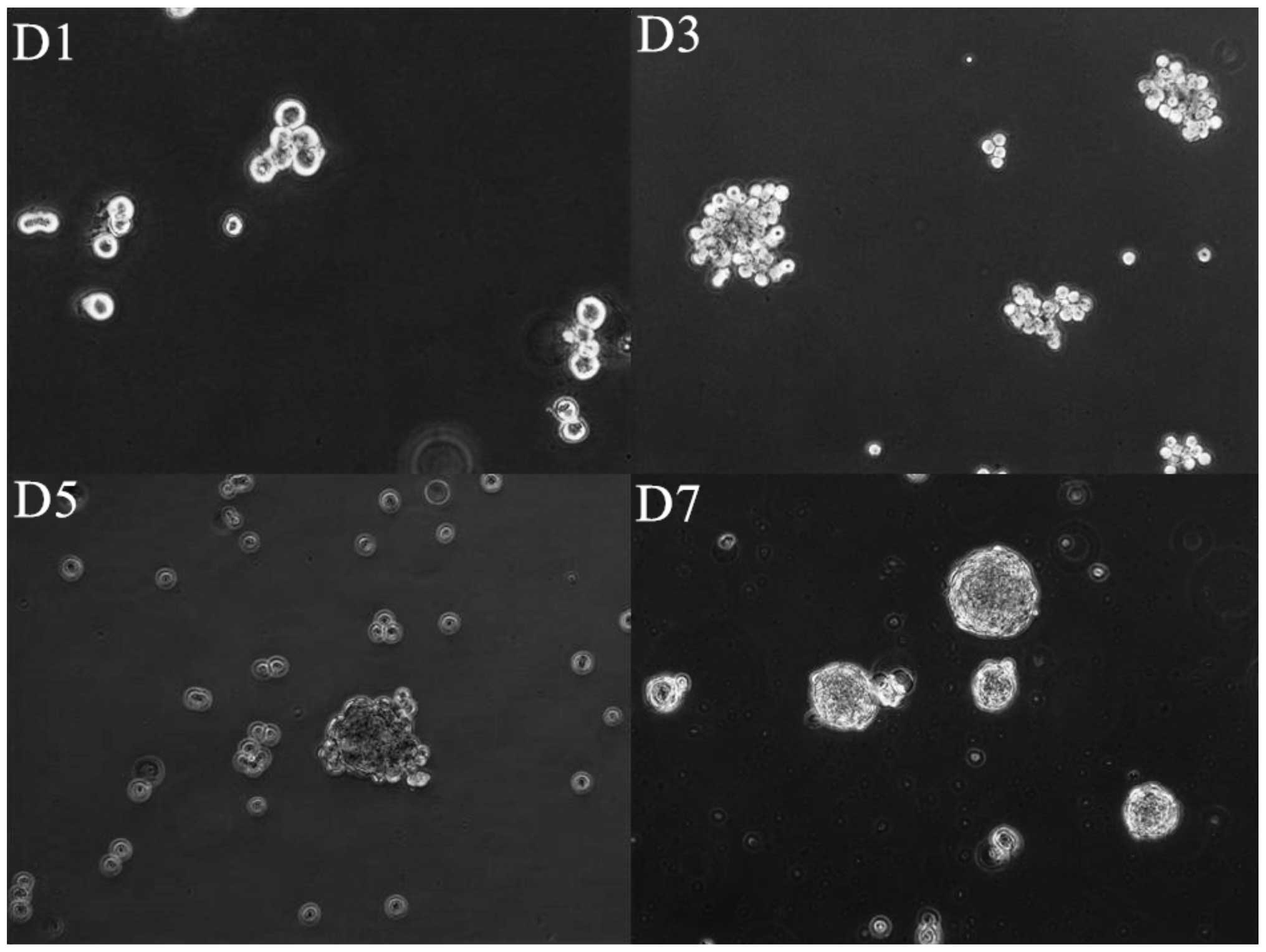
HNSCC cell lines contain cells with
self-renewing capacity and the ability to form spheroids
Cells from 8 HNSCC cell lines were cultured in
suspension for 7–10 days at a specific density of 2×104
cells/ml. The spheroid formation typically started at the first day
after plating suspension cultures and the spheroid size became
progressively larger. After 4–7 days the morphology of the
spheroids did not change in size anymore, but the number of the
spheroids in culture still continued to increase and the cell
clusters became more compact (Fig.
1).
Independent of HPV-association, all HNSCC cell lines
except UD-SCC2, formed highly compact spheroids. Only UD-SCC2
formed loose aggregates of cells easily dispersed by pipetting.
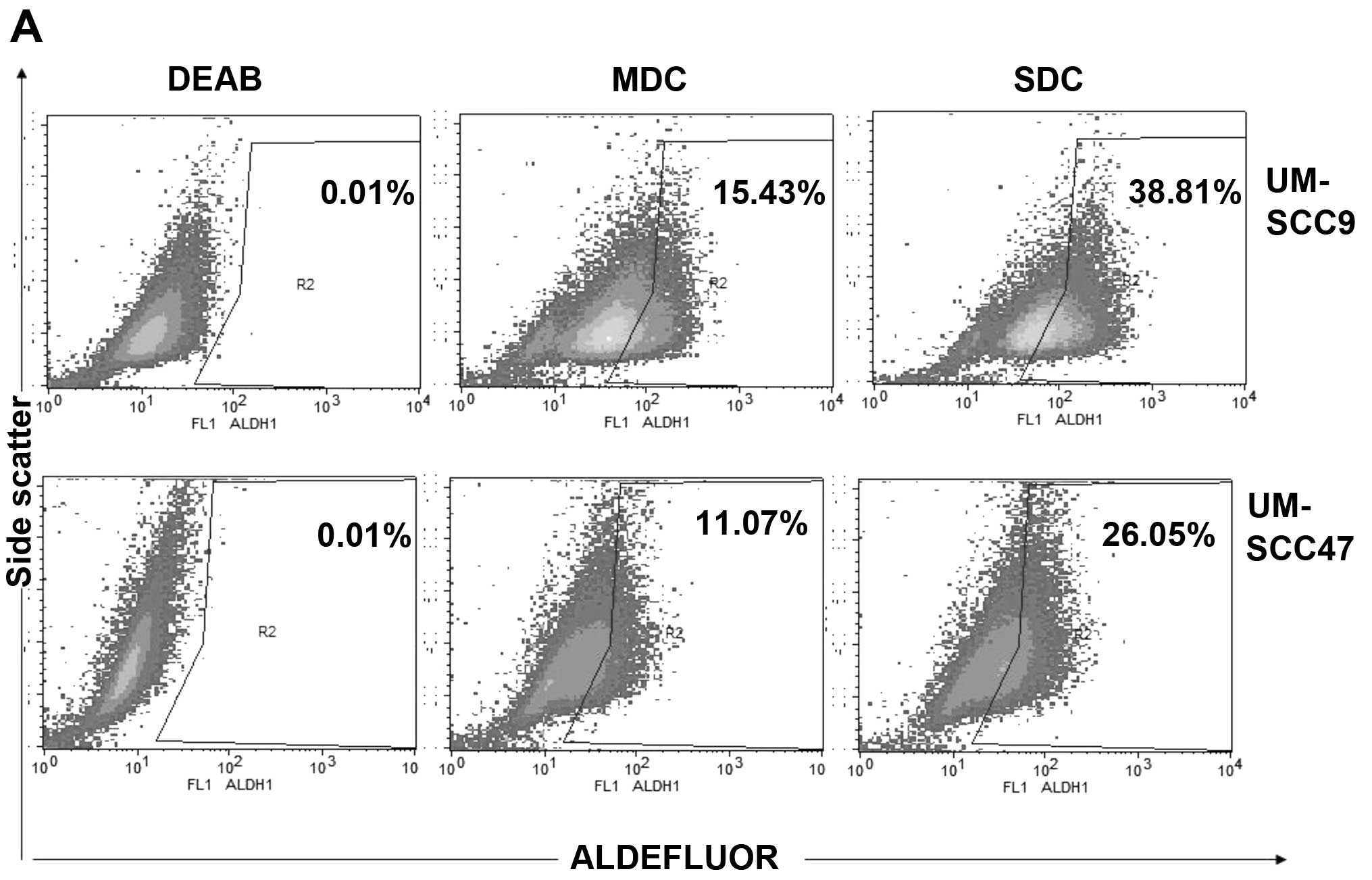
The CSCs marker ALDH shows a higher
expression level in SDC than in MDC
The Aldefluor assay has been successfully applied to
detect ALDH-expression in CSCs from primary tumors or established
cancer cell lines in various solid tumors, including HNSCC
(40). We measured ALDH enzymatic
activity of SDC of the eight HNSCC cell lines and their matched MDC
to identify and quantify the stem cell-like population (Fig. 2A).
All HNSCC-derived SDC showed a significant increase
in the number of ALDH+ cells compared to parental MDC
(P<0.05) (Fig. 2B). In the
HPV+ group, the cell line with the highest ALDH content
was UM-SCC47 SDC (25.62±0.50%) compared to the parental MDC
(11.05±0.16%). In the HPV− group, the highest proportion
of ALDH+ cells was found in SDC derived from UM-SCC11b
(45.05±0.22%) which was 3.3-fold higher than in the corresponding
parental MDC (13.38±0.11%). Interestingly, ALDH expression was
higher in SDC and MDC cultured from HPV− than from
HPV+ cell lines. The average percentage of
ALDH+ in in the MDC cell population derived from the
four HPV− cell lines was 10.64±1.37% while in the four
HPV+ HNSCC cell lines on average 5.74±0.92% of cells
were ALDH+ (P<0.01). In SDC the average percentage of
ALDH+ in the HPV− cell lines was 30.33±4.03
vs. 12.83±2.29% in the HPV+ cell lines (P<0.01).
These data correspond to our findings in HNSCC in vivo
(41) and may also reflect the
poor success rate with HPV+ HNSCC tumor specimen in a
xenograft mouse model due to a relatively low CSC content (42). Despite the fact that UD-SCC2 did
not form compact spheroids, the SDC still showed an increased
expression frequency of ALDH+ as compared to its
MDC.
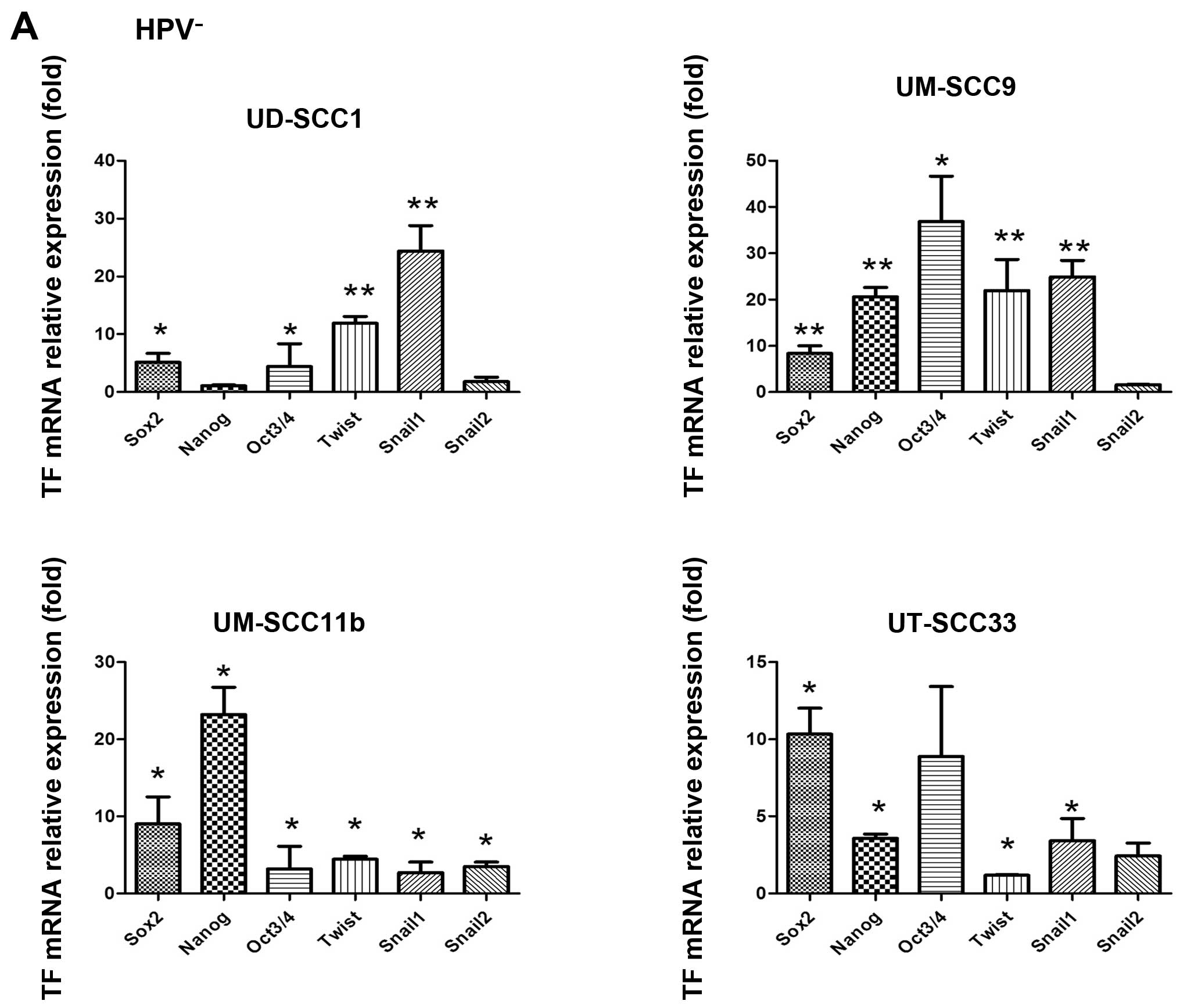
Stemness-related TFs and EMT-related TFs
are overexpressed in SDC
Sox2, Oct3/4 and Nanog were reported as important
TFs in maintaining stemness characteristics such as self-renewal
and pluripotency in human embryonic stem cells and cancer cells
(43). In line with previous
findings (10), we confirmed as
prerequisite for further work, that the mRNA level of Sox2, Oct3/4
and Nanog were significantly increased in SDC of the eight
investigated HNSCC cell lines (Fig.
3). The highest increase was observed in UM-SCC9 SDC, where a
36.81±5.66-fold increase in Oct3/4 expression was found over the
expression present in the parental MDC. The smallest difference was
observed in UD-SCC2, where the Nanog expression showed a
0.28±0.12-fold decrease. With regard to HPV-association, the mean
Sox2 expression ratio in HPV− SDC/MDC was 8.21±0.79- vs.
1.83±0.36-fold in HPV+ cell lines (P<0.01). The mean
Oct3/4 expression ratio in HPV− SDC/MDC was 13.32±4.38-
vs. 4.07±1.34-fold in HPV+ cell lines (P=0.07). Finally,
the mean Nanog expression ratio was in HPV− SDC/MDC
11.93±2.80- vs. 1.80±0.52-fold in HPV+ cell lines
(P<0.01).
Snail1, one of the key TFs involved in EMT, was
significantly increased in all SDC generated from the 8 tested
HNSCC cell lines as compared to MDC independent of their HPV status
(1.04–27.28-fold, P<0.01). Interestingly, we also found that the
mean expression ratio of Snail1 (HPV− SDC/MDC 20.72 vs.
HPV+ 2.90-fold, P<0.05) and of Twist (HPV−
9.87 SDC/MDC vs. HPV+ 2.72-fold, P<0.05) was
significantly higher in HPV− SDC/MDC than
HPV+ cell lines.
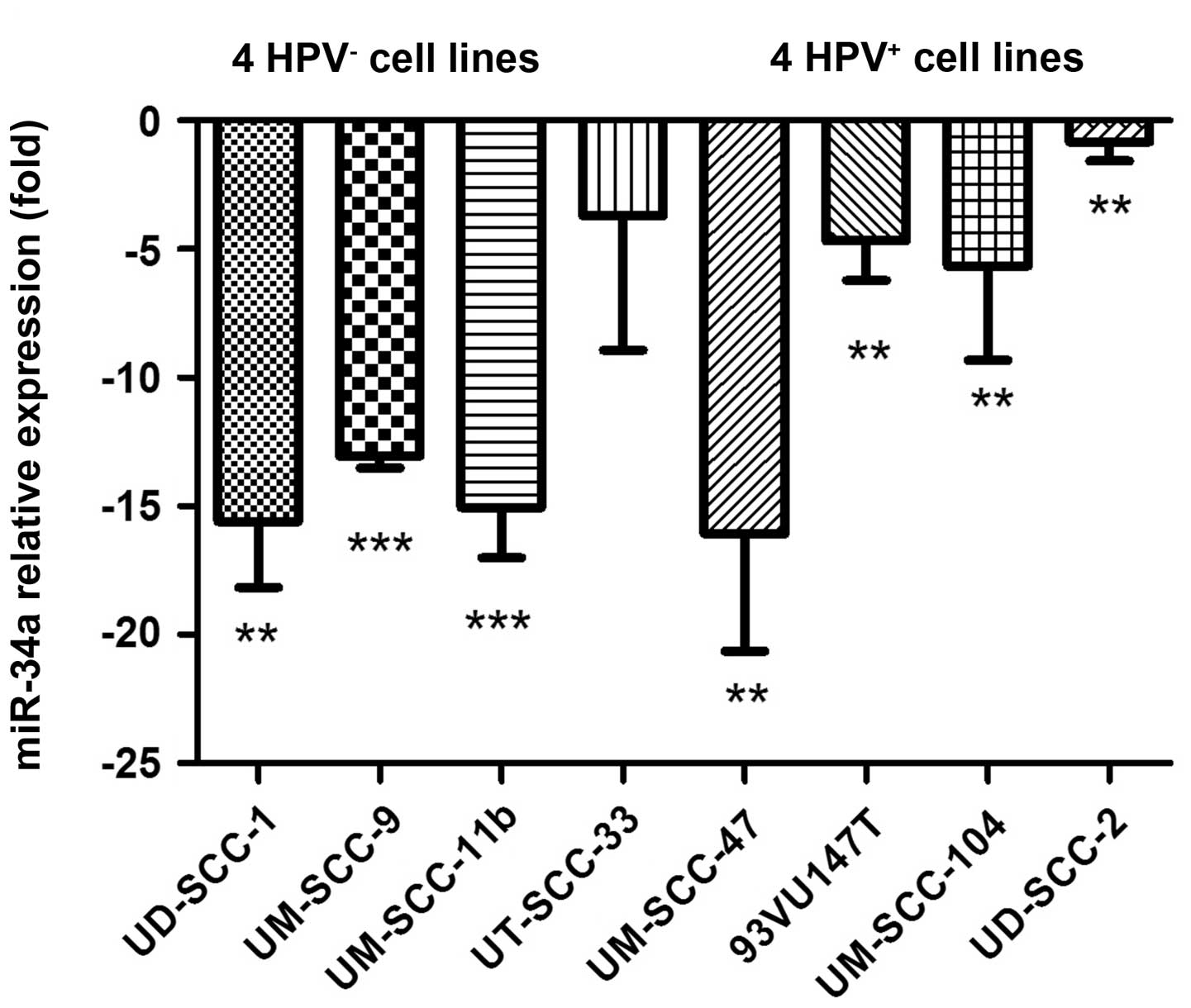
miR-34a is downregulated in HNSCC-derived
SDC
We selected miR-34a due to its role as a tumor
suppressor. We found that the expression of miR-34a was
consistently and significantly downregulated in SDC compared to the
parental MDC in all tested HNSCC cell lines (1.61–16.37-fold,
P<0.05) (Fig. 4).
Overexpression of miR-34a reduces
stemness- and EMT properties in HNSCC-SDC
To investigate the possibility that miR-34a serves
as a link between EMT and CSCs by regulating EMT in HNSCC-SDC the
following experiments were performed. First, in order to test
whether miR-34a expression was sufficient to downregulate EMT and
CSCs marker expression, ectopic miR-34a mimics were transiently
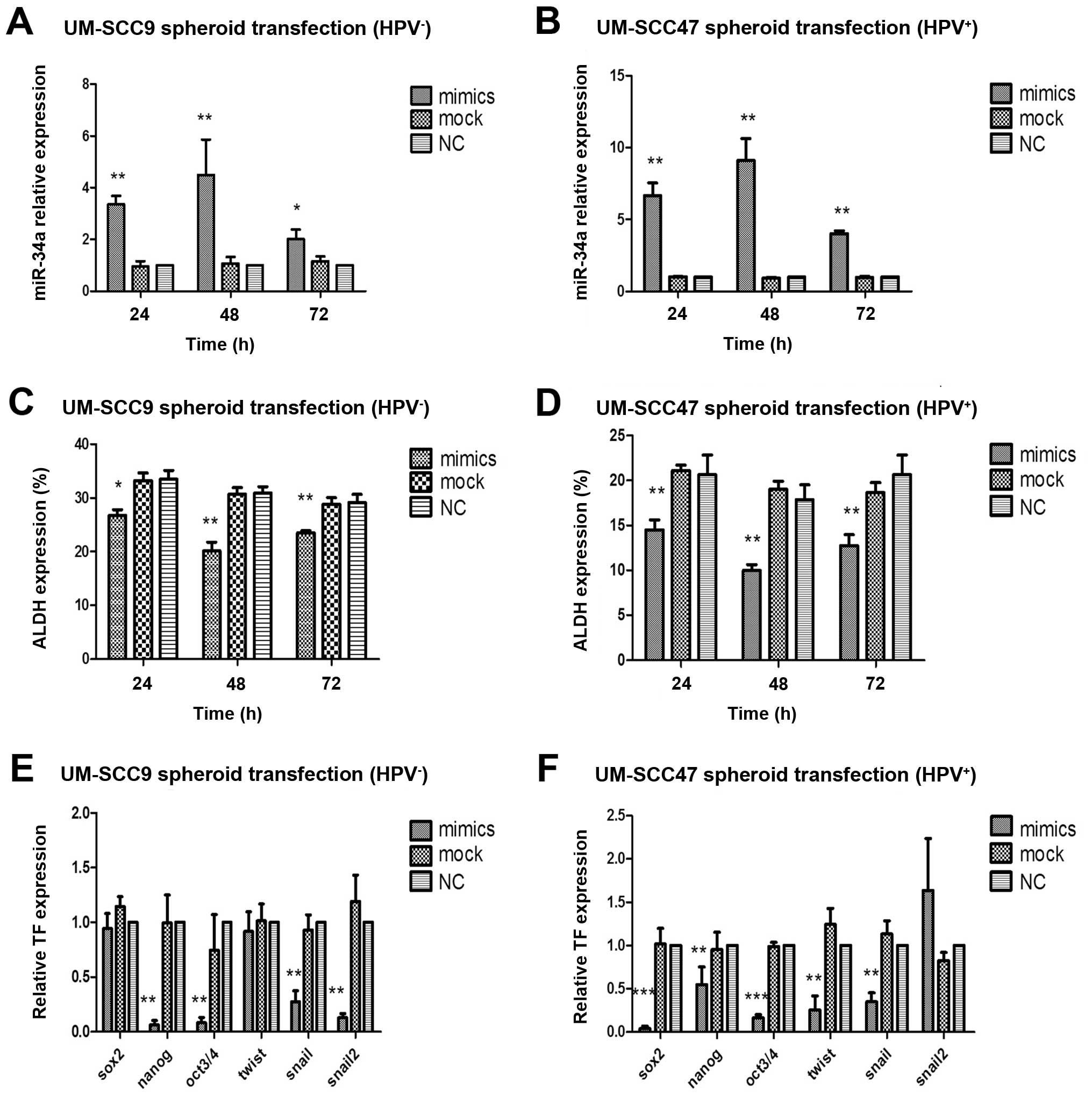
transfected into UM-SCC9 and UM-SCC47 cells.
The transfection efficiency of UM-SCC9 was
94.33±0.57% and of UM-SCC47 93.66±1.52%. We also measured the
transfection efficiency by verifying relative expression level of
miR-34a mRNA after the transfection with miR-34a mimics and found
that miR-34a mRNA level significantly increased after 24, 48 and 72
h, while the mock transfected group showed no differences in
expression levels in comparison to the normal control (NC group)
(Fig. 5A and B). The peak of
miR-34a level was found 48 h post-transfection (UM-SCC9 4.49-fold,
UM-SCC47 9.1-fold, P<0.01).
With respect to the regulation of CSC marker, we
observed downregulation of ALDH expression after miR-34a mimics
transfection after 24, 48 and 72 h (Fig. 5C and D). The strongest change in
ALDH+ cell frequency was found to correlate to the
highest miR-34a levels 48 h post-transfection (UM-SCC9 1.47-fold,
UM-SCC47 1.93-fold, P<0.01). Furthermore, transfection of
miR-34a mimics resulted in a decrease in CSC- and EMT-related TF
mRNA expression which was tested 48 h post-transfection (Fig. 5E and F). Nanog, Oct3/4 and Snail1
were significantly decreased over controls in the two transfected
HNSCC cell lines and Sox2 and Twist showed significant
downregulation in UM-SCC47. While Snail 2 showed decreased
expression in UM-SCC9 and an increase in expression in
UM-SCC47.
Overexpression of miR-34a reduces the
invasive capacity
For evaluation of the invasive capacity of tumor
cells we used the Matrigel invasion assay. After 24 h of culture,
the number of miR-34a mimic-transfected HNSCC-SDC that migrated
through the Matrigel coated chamber was significantly decreased
(P<0.001) compared with NC and mock-transfected cells (Fig. 6). The miR-34a levels after
transfection were significantly increased as described above
(UM-SCC9: 29.75±1.08 vs. 12.87±0.61%, UM-SCC47: 30.25±1.23 vs.
13.12± 0.66%). Transfection of miR-34a significantly decreased the
migration/invasion of UM-SCC9 and UM-SCC47 SDC by ~50%.
Decreased ability of spheroid and colony
formation as well as anchorage-independent growth after miR-34a
transfection
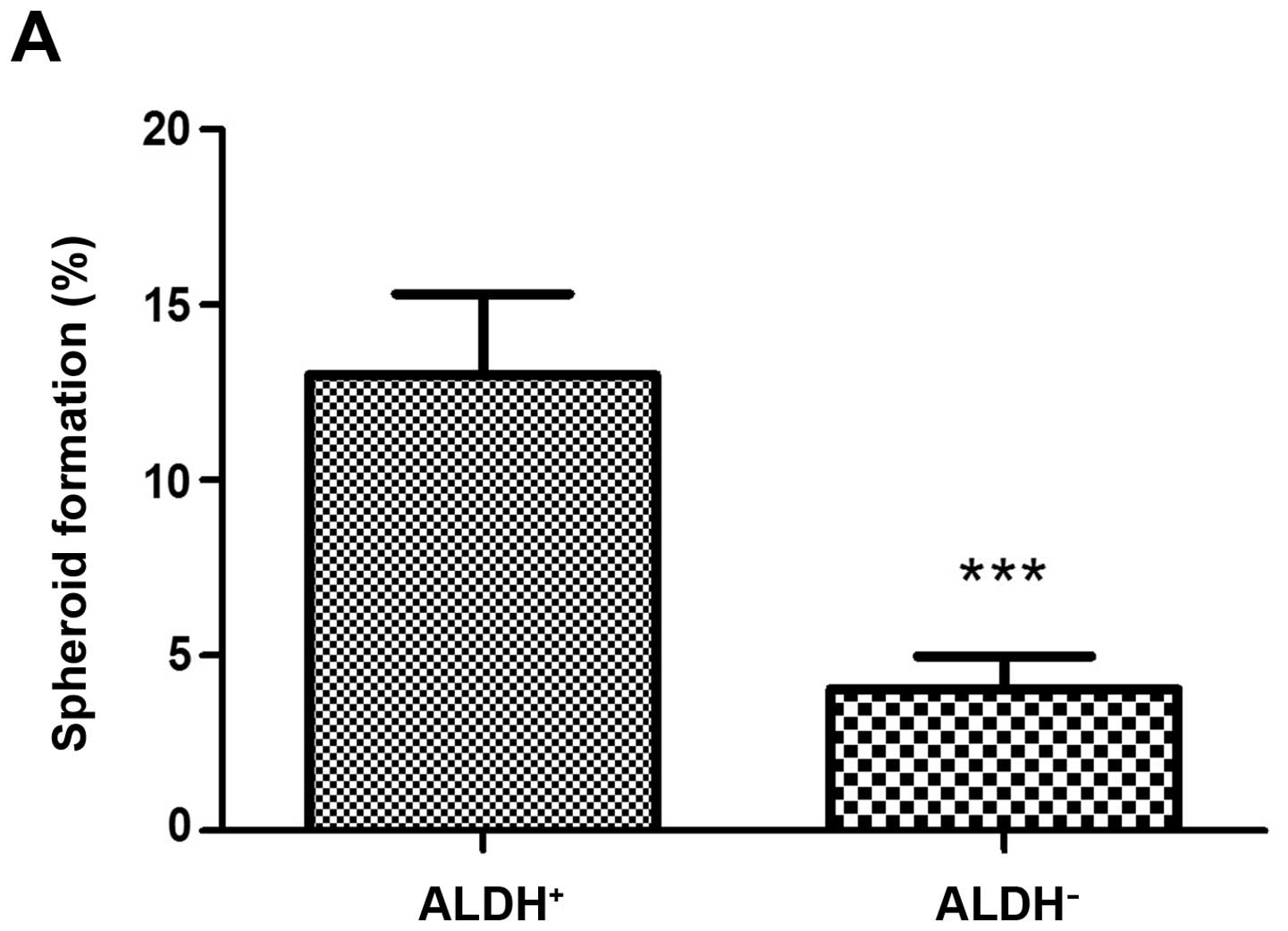
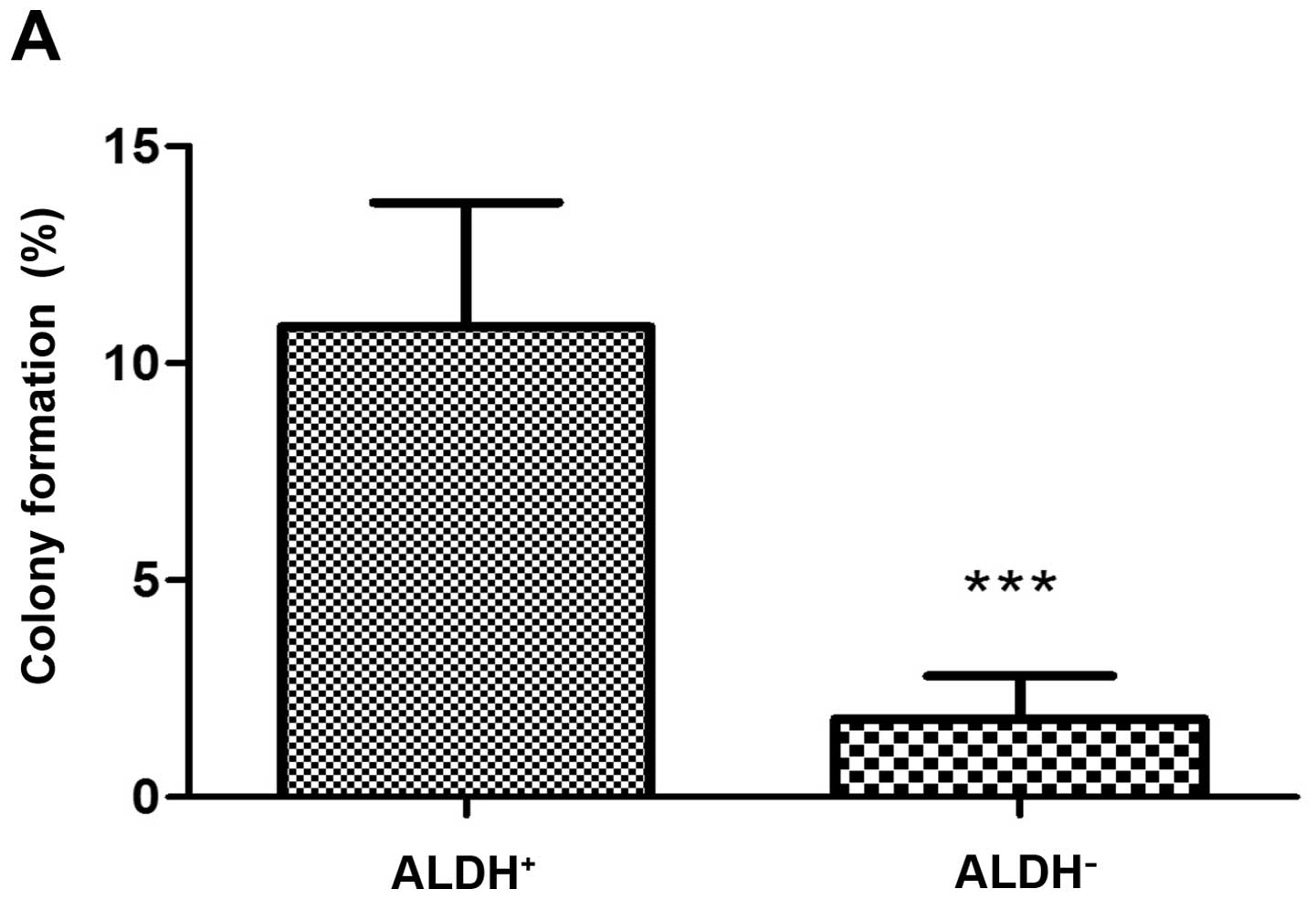
Following flow cytometric sorting of UM-SCC9 for
ALDH-positivity, the role of miR-34a in spheroid and colony
formation and anchorage-independent growth was assessed. Before
transfection of miR-34a, the sorted ALDH+ cells showed a
higher spheroid formation (Fig.
7A) and colony formation ability (Fig. 8A) than ALDH− cells.
Transfection with miR-34a mimics reduced significantly the ability
for anchorage-independent growth of UM-SCC9 cells. Analysis of
spheroid formation showed that cells transfected with miR-34a
mimics displayed much fewer and smaller spheroids than the
control-transfected cells (Fig.
7B–G). The number of spheroids and their size were
significantly decreased in miR-34a mimic-transfected samples as
compared with NC.
Discussion
Spheroid culture and enrichment of
HNSCC-CSCs
Spheroids are spherical three-dimensional tumor cell
clusters that are grown from one or several cell clones. As
compared to cells cultured in monolayer, cell doubling time and the
rate and pattern of spheroid growth in vitro better matches
that observed in tumors in vivo (44). Spheroid cell growth has also been
identified to be a property of normal tissue cells which display
stem cell properties (45).
Spheroid enriched CSCs can be derived from a panel of different
solid malignancies such as HNSCC (10), melanoma (46), breast cancer (47) and gliosarcoma (14). Cells from these entities could be
propagated by anchorage-independent growth and displayed the
phenotype of non-adherent spheroids. The spheroid-forming ability
was found to correspond to the expression of established CSC
markers (48).
The ability of a single cell to regenerate a
malignant tumor consisting of cells with heterogeneous phenotypes
is one characteristic of CSCs that may help to explain some of the
differences which discriminate tumor cells from differentiated
somatic cells such as immortality, quiescence, invasion, metastasis
and relapse after treatment. Initial studies identified
CD44+ HNSCC cells that could generate new tumors in
vivo (<5,000 cells injected into mice) (8) and concluded that CD44 is a CSC
marker. ALDH has been demonstrated to be another useful CSC marker
to identify CSCs in many epithelial cancers including HNSCC
(10,33,41,49–51).
In HNSCC, Chen et al showed in immunocompromised mice that
500 injected ALDH+ HNSCC cells resulted in visible
tumors in all animals after 6 weeks, while 104
ALDH− cells failed to produce tumors (51). According to our own results and in
line with the above mentioned observations, ALDH+ cells
showed increased CSCs properties compared to ALDH− cells
derived from HNSCC cell lines (10). We and others have demonstrated a
varying overlap of ALDH and CD44 populations in HNSCC (10,40).
In our experiments, we cultured spheroids from eight
HNSCC cell lines. The eight cell lines showed varying ability to
form spheroids. Sox2, Nanog and Oct3/4 were chosen as the stemness
related TFs in this project, because they were expressed at high
levels, both in normal stem cells and CSC as predicted by
Chickarmane and Peterson (52) and
demonstrated for HNSCC by us (10). We validated the content of SDC
positive for the HNSCC-CSC marker ALDH by flow cytometry and
CSC-related TFs by RT-qPCR. We found that in SDC compared with
corresponding MDC, the CSC-related transcription factors Sox2,
Oct3/4 and Nanog were significantly upregulated, as was the
proportion of ALDH+ cells.
These findings reveal that the spheroid culture
assay is a useful and efficient method to enrich cells with CSC
characteristics from HNSCC cell lines and that sorting for
ALDH+ cells even allows for further enrichment of cells
with spheroid- and clone-forming abilities. However, there was also
a limitation in the spheroid culture assay to enrich for CSCs. The
most important limitation was that the spheres still represent a
heterogeneous population, with only a part of the cells having the
ability of self-renewal (53).
MiR-34a regulates the stemness and EMT
properties in HNSCC CSCs
MiR-34a has been considered a tumor suppressor which
represses stemness-related features and -functions in prostate
cancer (54), breast cancer
(55) and pancreatic cancer
(37). An important finding of
this study was that the data support the observation that miR-34a
acts as tumor stemness repressor in HNSCC. Recent reports indicate
downregulation of miR-34a in HNSCC cell lines and tumor samples
that may promote tumor growth and tumor angiogenesis (38). Our own data confirm the observation
of dysregulated miR-34a expression in HNSCC cell lines. We
demonstrated that the expression in stem cell-enriched HNSCC-SDC
was significantly lower than in parental HNSCC-MDC. Furthermore,
the frequency of ALDH+ CSC was increased in SDC compared
to MDC. However, transfection of miR-34a mimics into the HNSCC-SDC
replenishing miR-34a levels simultaneously decreased the expression
of ALDH along with the stemness-related TFs Sox2, Nanog and Oct3/4.
After upregulation of miR-34a, we found the colony and sphere
formation ability and invasive capacity decreased in
ALDH+ cells compared to controls.
Although the direct effects of miR-34a have been
studied in a wide range of different cancers, relatively few
studies have investigated other possible cellular functions of
miR-34a. Kim et al reported that the p53/miR-34a axis
regulates Snail1-dependent EMT. Among the proto-type EMT regulators
(ZEB1/2, Snail1, Slug), also miR-34a directly targets Snail1
(56). According to a study by
Siemens et al (57) miR-34a
and Snail forms a double-negative feedback loop to regulate EMT.
Activated p53 downregulates the EMT-inducing transcription factor
Snail1 by upregulation of miR-34a. On the other hand the TF Snail1
binds to the E boxes in miR-34a promoters to repress its
expression. However, indirect downregulation of ZEB1/2 and Slug,
which are miR-200 targets, may occur via de-repression of the
Snail-regulated miR-200 promoters after miR-34a activation
(57).
In this study, we compared EMT-properties of SDC to
MDC after transfection with miR-34a mimics to further characterize
the role of this molecule in EMT. We were able to demonstrate that
the transfection of miR-34a mimics led to downregulation of
EMT-related TFs in SDC. In line with these findings, the invasive
capacity of these cells was reduced, indicating an implication of
miR-34a in the regulation of EMT in HNSCC. This observation may be
of future clinical relevance since, in a prostate cancer mouse
model systemic delivery of miR-34a inhibited formation of
metastasis (54). Therefore, the
mechanisms of miR-34a action and regulation presented here may have
diagnostic and therapeutic relevance in the future.
In our experiments, the EMT-related TF Snail1 and
CSC-related TFs showed decreased expression in HNSCC cell lines
after transfection of miR-34a mimics, indicating that the miR-34a
may play multiple roles in suppressing mesenchymal traits and
inhibiting stemness properties. A similar phenomenon was also
reported in pancreatic cancer, where a restoration of miR-34a
reduced CSC properties and inhibited the EMT (58). In breast cancer cells, it was
demonstrated that re-expression of miR-200 suppressed EMT-related
genes and stemness properties (59). C-MYC and CD44 represent direct
miR-34a targets (54,60–62),
whereas, the effect on the other stemness markers/factors is
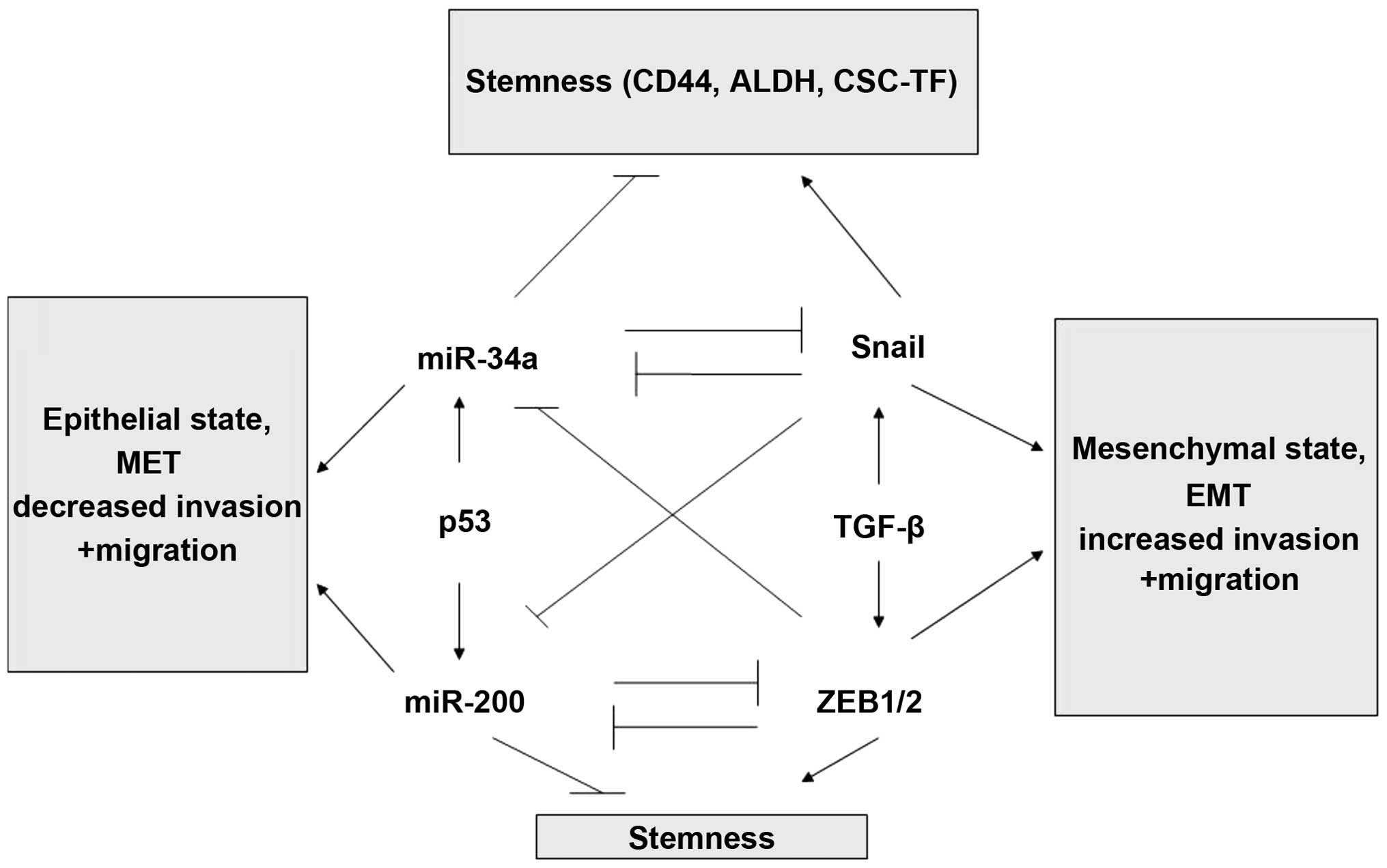
presumably indirect (Fig. 9).
Taken together, we were able to demonstrate that
miR-34a is involved in the regulation of EMT and invasive
properties of CSC by quantifying TFs involved in the regulation of
EMT (Snail and Twist) and by conducting functional assays
displaying colony forming and invasive capacities. Previously, we
demonstrated as a property of HNSCC-CSC an increased invasive
capacity and expression of EMT-markers such as α-smooth muscle
actin and vimentin, while at the same time the expression of the
adhesion molecule E-cadherin was significantly reduced (10). Our experiments demonstrated that
this can be reversed by transfer of miR34a-mimics in HNSCC in
vitro. Evidence for an accumulation of the formulated miR34a
mimics was reported in the spleen, lung and kidney (63), suggesting a potential opportunity
for cancer therapeutic development.
Influence of the HPV on HNSCC-CSC
characteristics
HPV+- and HPV− HNSCC show
distinct characteristics in their biological and clinical behavior.
We found in primary oropharyngeal squamous cell carcinoma, that
HPV-DNA+ tumors had a lower ALDH1A1 expression and the
HPV-DNA− group had higher expression as measured by
immunohistochemistry (41).
ALDH1A1 is not only a CSC marker but it also serves as a prognostic
biomarker in HNSCC (33,64). Moreover, we have assessed the
utility of ALDH1A1 staining-intensity as measure for its expression
as a prognostic biomarker in surgically treated HNSCC patients. We
were able to demonstrate a significant correlation between ALDH1A1
staining intensity and prognosis (33). HNSCC patients with strong ALDH1A1
expression, had a significantly inferior 5-year overall survival
compared with those HNSCC patients who had weak ALDH1A1 expression
(65). On the basis of these
observations, we hypothesized that the improved clinical outcome of
patients with HPV+ HNSCC may be due to the fact that
HPV+ HNSCC has a lower expression level of ALDH than
HPV− HNSCC and a lower CSC content. Our data support
these clinical results, as also in cell culture HPV+
HNSCC cell lines have a lower expression level of ALDH compared to
the HPV− group. However, the number of tested cell lines
was limited and may represent characteristics specific to
HNSCC-lines that have the ability to grow in vitro.
In conclusion, in this study, we found that
increased miR-34a can suppress CSC-like properties of HNSCC and
EMT. This was demonstrated by the downregulation of CSC- and
EMT-related TFs, colony- and spheroid-forming abilities and
decreased invasive capacity. These findings suggest that miR-34a
may play important roles in these processes and may therefore have
a potential in novel therapeutic regimen or in combination with
existing treatments of HNSCC: a) by reducing the frequency of the
CSC-phenotype which is held responsible for therapy-resistance and
b) by reducing the formation of novel metastases by inhibiting EMT.
However, routes to apply this molecule in a therapeutic setting in
humans remain to be explored.
References
|
1
|
Kamangar F, Dores GM and Anderson WF:
Patterns of cancer incidence, mortality, and prevalence across five
continents: Defining priorities to reduce cancer disparities in
different geographic regions of the world. J Clin Oncol.
24:2137–2150. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jemal A, Siegel R, Ward E, Hao Y, Xu J,
Murray T and Thun MJ: Cancer statistics, 2008. CA Cancer J Clin.
58:71–96. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pignon JP, Baujat B and Bourhis J:
Individual patient data meta-analyses in head and neck carcinoma:
what have we learnt? Cancer Radiother. 9:31–36. 2005.(In French).
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Reya T, Morrison SJ, Clarke MF and
Weissman IL: Stem cells, cancer, and cancer stem cells. Nature.
414:105–111. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chan KS, Espinosa I, Chao M, Wong D,
Ailles L, Diehn M, Gill H, Presti J Jr, Chang HY, van de Rijn M, et
al: Identification, molecular characterization, clinical prognosis,
and therapeutic targeting of human bladder tumor-initiating cells.
Proc Natl Acad Sci USA. 106:14016–14021. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Al-Hajj M, Wicha MS, Benito-Hernandez A,
Morrison SJ and Clarke MF: Prospective identification of
tumorigenic breast cancer cells. Proc Natl Acad Sci USA.
100:3983–3988. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dalerba P, Dylla SJ, Park IK, Liu R, Wang
X, Cho RW, Hoey T, Gurney A, Huang EH, Simeone DM, et al:
Phenotypic characterization of human colorectal cancer stem cells.
Proc Natl Acad Sci USA. 104:10158–10163. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Prince ME, Sivanandan R, Kaczorowski A,
Wolf GT, Kaplan MJ, Dalerba P, Weissman IL, Clarke MF and Ailles
LE: Identification of a subpopulation of cells with cancer stem
cell properties in head and neck squamous cell carcinoma. Proc Natl
Acad Sci USA. 104:973–978. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hermann PC, Huber SL, Herrler T, Aicher A,
Ellwart JW, Guba M, Bruns CJ and Heeschen C: Distinct populations
of cancer stem cells determine tumor growth and metastatic activity
in human pancreatic cancer. Cell Stem Cell. 1:313–323. 2007.
View Article : Google Scholar
|
|
10
|
Liu X, Wang C, Chen Z, Jin Y, Wang Y,
Kolokythas A, Dai Y and Zhou X: MicroRNA-138 suppresses
epithelial-mesenchymal transition in squamous cell carcinoma cell
lines. Biochem J. 440:23–31. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nguyen LV, Vanner R, Dirks P and Eaves CJ:
Cancer stem cells: An evolving concept. Nat Rev Cancer. 12:133–143.
2012.PubMed/NCBI
|
|
12
|
Lee HE, Kim JH, Kim YJ, Choi SY, Kim SW,
Kang E, Chung IY, Kim IA, Kim EJ, Choi Y, et al: An increase in
cancer stem cell population after primary systemic therapy is a
poor prognostic factor in breast cancer. Br J Cancer.
104:1730–1738. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jiang W, Peng J, Zhang Y, Cho WC and Jin
K: The implications of cancer stem cells for cancer therapy. Int J
Mol Sci. 13:16636–16657. 2012. View Article : Google Scholar
|
|
14
|
Ghods AJ, Irvin D, Liu G, Yuan X,
Abdulkadir IR, Tunici P, Konda B, Wachsmann-Hogiu S, Black KL and
Yu JS: Spheres isolated from 9L gliosarcoma rat cell line possess
chemoresistant and aggressive cancer stem-like cells. Stem Cells.
25:1645–1653. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Azari H, Rahman M, Sharififar S and
Reynolds BA: Isolation and expansion of the adult mouse neural stem
cells using the neurosphere assay. J Vis Exp.
45:23932010.PubMed/NCBI
|
|
16
|
Todaro M, Alea MP, Di Stefano AB,
Cammareri P, Vermeulen L, Iovino F, Tripodo C, Russo A, Gulotta G,
Medema JP, et al: Colon cancer stem cells dictate tumor growth and
resist cell death by production of interleukin-4. Cell Stem Cell.
1:389–402. 2007. View Article : Google Scholar
|
|
17
|
Schatton T, Murphy GF, Frank NY, Yamaura
K, Waaga-Gasser AM, Gasser M, Zhan Q, Jordan S, Duncan LM,
Weishaupt C, et al: Identification of cells initiating human
melanomas. Nature. 451:345–349. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Okamoto A, Chikamatsu K, Sakakura K,
Hatsushika K, Takahashi G and Masuyama K: Expansion and
characterization of cancer stem-like cells in squamous cell
carcinoma of the head and neck. Oral Oncol. 45:633–639. 2009.
View Article : Google Scholar
|
|
19
|
Chiou SH, Yu CC, Huang CY, Lin SC, Liu CJ,
Tsai TH, Chou SH, Chien CS, Ku HH and Lo JF: Positive correlations
of Oct-4 and Nanog in oral cancer stem-like cells and high-grade
oral squamous cell carcinoma. Clin Cancer Res. 14:4085–4095. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wu KJ and Yang MH: Epithelial-mesenchymal
transition and cancer stemness: The Twist1-Bmi1 connection. Biosci
Rep. 31:449–455. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Santisteban M, Reiman JM, Asiedu MK,
Behrens MD, Nassar A, Kalli KR, Haluska P, Ingle JN, Hartmann LC,
Manjili MH, et al: Immune-induced epithelial to mesenchymal
transition in vivo generates breast cancer stem cells. Cancer Res.
69:2887–2895. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mani SA, Guo W, Liao MJ, Eaton EN, Ayyanan
A, Zhou AY, Brooks M, Reinhard F, Zhang CC, Shipitsin M, et al: The
epithelial-mesenchymal transition generates cells with properties
of stem cells. Cell. 133:704–715. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen C, Zimmermann M, Tinhofer I, Kaufmann
AM and Albers AE: Epithelial-to-mesenchymal transition and cancer
stem(-like) cells in head and neck squamous cell carcinoma. Cancer
Lett. 338:47–56. 2013. View Article : Google Scholar
|
|
24
|
Hindriksen S and Bijlsma MF: Cancer stem
cells, EMT, and developmental pathway activation in pancreatic
tumors. Cancers (Basel). 4:989–1035. 2012. View Article : Google Scholar
|
|
25
|
Micalizzi DS, Farabaugh SM and Ford HL:
Epithelial-mesenchymal transition in cancer: Parallels between
normal development and tumor progression. J Mammary Gland Biol
Neoplasia. 15:117–134. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Reddy BY, Lim PK, Silverio K, Patel SA,
Won BW and Rameshwar P: The microenvironmental effect in the
progression, metastasis, and dormancy of breast cancer: A model
system within bone marrow. Int J Breast Cancer. 2012:7216592012.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Phinney DG: Twist,
epithelial-to-mesenchymal transition, and stem cells. Stem Cells.
29:3–4. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
van der Pluijm G: Epithelial plasticity,
cancer stem cells and bone metastasis formation. Bone. 48:37–43.
2011. View Article : Google Scholar
|
|
29
|
Korpal M, Lee ES, Hu G and Kang Y: The
miR-200 family inhibits epithelial-mesenchymal transition and
cancer cell migration by direct targeting of E-cadherin
transcriptional repressors ZEB1 and ZEB2. J Biol Chem.
283:14910–14914. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Viñas-Castells R, Beltran M, Valls G,
Gómez I, García JM, Montserrat-Sentís B, Baulida J, Bonilla F, de
Herreros AG and Díaz VM: The hypoxia-controlled FBXL14 ubiquitin
ligase targets SNAIL1 for proteasome degradation. J Biol Chem.
285:3794–3805. 2010. View Article : Google Scholar :
|
|
31
|
Martin A and Cano A: Tumorigenesis: Twist1
links EMT to self-renewal. Nat Cell Biol. 12:924–925. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yang MH, Hsu DS, Wang HW, Wang HJ, Lan HY,
Yang WH, Huang CH, Kao SY, Tzeng CH, Tai SK, et al: Bmi1 is
essential in Twist1-induced epithelial-mesenchymal transition. Nat
Cell Biol. 12:982–992. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Qian X, Wagner S, Ma C, Coordes A, Gekeler
J, Klussmann JP, Hummel M, Kaufmann AM and Albers AE: Prognostic
significance of ALDH1A1-positive cancer stem cells in patients with
locally advanced, metastasized head and neck squamous cell
carcinoma. J Cancer Res Clin Oncol. 140:1151–1158. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hugo H, Ackland ML, Blick T, Lawrence MG,
Clements JA, Williams ED and Thompson EW: Epithelial-mesenchymal
and mesenchymal-epithelial transitions in carcinoma progression. J
Cell Physiol. 213:374–383. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ji Q, Hao X, Zhang M, Tang W, Yang M, Li
L, Xiang D, Desano JT, Bommer GT, Fan D, et al: MicroRNA miR-34
inhibits human pancreatic cancer tumor-initiating cells. PLoS One.
4:e68162009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kumar B, Yadav A, Lang J, Teknos TN and
Kumar P: Dysregulation of microRNA-34a expression in head and neck
squamous cell carcinoma promotes tumor growth and tumor
angiogenesis. PLoS One. 7:e376012012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Pfaffl MW: A new mathematical model for
relative quantification in real-time RT-PCR. Nucleic Acids Res.
29:e452001. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Clay MR, Tabor M, Owen JH, Carey TE,
Bradford CR, Wolf GT, Wicha MS and Prince ME: Single-marker
identification of head and neck squamous cell carcinoma cancer stem
cells with aldehyde dehydrogenase. Head Neck. 32:1195–1201. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Qian X, Wagner S, Ma C, Klussmann JP,
Hummel M, Kaufmann AM and Albers AE: ALDH1-positive cancer
stem-like cells are enriched in nodal metastases of oropharyngeal
squamous cell carcinoma independent of HPV status. Oncol Rep.
29:1777–1784. 2013.PubMed/NCBI
|
|
42
|
Klinghammer K, Raguse JD, Plath T, Albers
AE, Joehrens K, Zakarneh A, Brzezicha B, Wulf-Goldenberg A,
Keilholz U, Hoffmann J, et al: A comprehensively characterized
large panel of head and neck cancer patient-derived xenografts
identifies the mTOR inhibitor everolimus as potential new treatment
option. Int J Cancer. 136:2940–2948. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ling GQ, Chen DB, Wang BQ and Zhang LS:
Expression of the pluripotency markers Oct3/4, Nanog and Sox2 in
human breast cancer cell lines. Oncol Lett. 4:1264–1268.
2012.PubMed/NCBI
|
|
44
|
Santini MT, Rainaldi G and Indovina PL:
Multicellular tumour spheroids in radiation biology. Int J Radiat
Biol. 75:787–799. 1999. View Article : Google Scholar
|
|
45
|
Wicha MS, Liu S and Dontu G: Cancer stem
cells: an old idea - a paradigm shift. Cancer Res. 66:1883–1890;
discussion 1895–1886. 2006. View Article : Google Scholar
|
|
46
|
Fang D, Nguyen TK, Leishear K, Finko R,
Kulp AN, Hotz S, Van Belle PA, Xu X, Elder DE and Herlyn M: A
tumorigenic subpopulation with stem cell properties in melanomas.
Cancer Res. 65:9328–9337. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Ponti D, Costa A, Zaffaroni N, Pratesi G,
Petrangolini G, Coradini D, Pilotti S, Pierotti MA and Daidone MG:
Isolation and in vitro propagation of tumorigenic breast cancer
cells with stem/progenitor cell properties. Cancer Res.
65:5506–5511. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Kuch V, Schreiber C, Thiele W, Umansky V
and Sleeman JP: Tumor-initiating properties of breast cancer and
melanoma cells in vivo are not invariably reflected by spheroid
formation in vitro, but can be increased by long-term culturing as
adherent monolayers. Int J Cancer. 132:E94–E105. 2013. View Article : Google Scholar
|
|
49
|
Ginestier C, Hur MH, Charafe-Jauffret E,
Monville F, Dutcher J, Brown M, Jacquemier J, Viens P, Kleer CG,
Liu S, et al: ALDH1 is a marker of normal and malignant human
mammary stem cells and a predictor of poor clinical outcome. Cell
Stem Cell. 1:555–567. 2007. View Article : Google Scholar
|
|
50
|
Ma S, Chan KW, Lee TK, Tang KH, Wo JY,
Zheng BJ and Guan XY: Aldehyde dehydrogenase discriminates the
CD133 liver cancer stem cell populations. Mol Cancer Res.
6:1146–1153. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Chen YC, Chen YW, Hsu HS, Tseng LM, Huang
PI, Lu KH, Chen DT, Tai LK, Yung MC, Chang SC, et al: Aldehyde
dehydrogenase 1 is a putative marker for cancer stem cells in head
and neck squamous cancer. Biochem Biophys Res Commun. 385:307–313.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Chickarmane V and Peterson C: A
computational model for understanding stem cell, trophectoderm and
endoderm lineage determination. PLoS One. 3:e34782008. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Jensen JB and Parmar M: Strengths and
limitations of the neurosphere culture system. Mol Neurobiol.
34:153–161. 2006. View Article : Google Scholar
|
|
54
|
Liu C, Kelnar K, Liu B, Chen X,
Calhoun-Davis T, Li H, Patrawala L, Yan H, Jeter C, Honorio S, et
al: The microRNA miR-34a inhibits prostate cancer stem cells and
metastasis by directly repressing CD44. Nat Med. 17:211–215. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Park EY, Chang E, Lee EJ, Lee HW, Kang HG,
Chun KH, Woo YM, Kong HK, Ko JY, Suzuki H, et al: Targeting of
miR34a-NOTCH1 axis reduced breast cancer stemness and
chemoresistance. Cancer Res. 74:7573–7582. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Kim NH, Kim HS, Li XY, Lee I, Choi HS,
Kang SE, Cha SY, Ryu JK, Yoon D, Fearon ER, et al: A p53/miRNA-34
axis regulates Snail1-dependent cancer cell epithelial-mesenchymal
transition. J Cell Biol. 195:417–433. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Siemens H, Jackstadt R, Hünten S, Kaller
M, Menssen A, Götz U and Hermeking H: miR-34 and SNAIL form a
double-negative feedback loop to regulate epithelial-mesenchymal
transitions. Cell Cycle. 10:4256–4271. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Nalls D, Tang SN, Rodova M, Srivastava RK
and Shankar S: Targeting epigenetic regulation of miR-34a for
treatment of pancreatic cancer by inhibition of pancreatic cancer
stem cells. PLoS One. 6:e240992011. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Chang CJ, Chao CH, Xia W, Yang JY, Xiong
Y, Li CW, Yu WH, Rehman SK, Hsu JL, Lee HH, et al: p53 regulates
epithelial-mesenchymal transition and stem cell properties through
modulating miRNAs. Nat Cell Biol. 13:317–323. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Cannell IG and Bushell M: Regulation of
Myc by miR-34c: A mechanism to prevent genomic instability? Cell
Cycle. 9:2726–2730. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Christoffersen NR, Shalgi R, Frankel LB,
Leucci E, Lees M, Klausen M, Pilpel Y, Nielsen FC, Oren M and Lund
AH: p53-independent upregulation of miR-34a during oncogene-induced
senescence represses MYC. Cell Death Differ. 17:236–245. 2010.
View Article : Google Scholar
|
|
62
|
Kong YW, Cannell IG, de Moor CH, Hill K,
Garside PG, Hamilton TL, Meijer HA, Dobbyn HC, Stoneley M, Spriggs
KA, et al: The mechanism of micro-RNA-mediated translation
repression is determined by the promoter of the target gene. Proc
Natl Acad Sci USA. 105:8866–8871. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Bader AG, Brown D and Winkler M: The
promise of microRNA replacement therapy. Cancer Res. 70:7027–7030.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Koukourakis MI, Giatromanolaki A, Tsakmaki
V, Danielidis V and Sivridis E: Cancer stem cell phenotype relates
to radio-chemotherapy outcome in locally advanced squamous cell
head-neck cancer. Br J Cancer. 106:846–853. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Xu J, Müller S, Nannapaneni S, Pan L, Wang
Y, Peng X, Wang D, Tighiouart M, Chen Z, Saba NF, et al: Comparison
of quantum dot technology with conventional immunohistochemistry in
examining aldehyde dehydrogenase 1A1 as a potential biomarker for
lymph node metastasis of head and neck cancer. Eur J Cancer.
48:1682–1691. 2012. View Article : Google Scholar : PubMed/NCBI
|