Introduction
Cholangiocarcinoma is a common malignancy of the
biliary tract. It has a high mortality rate, and is difficult to
diagnose (1). The morbidity and
mortality of intrahepatic cholangiccarcinoma in the world are on
the rise (2). The cause for this
increase could be related to an interplay between genetic factors
and environmental triggers (3).
Chemotherapy can lead to drug resistance and thus worse prognosis
(4). Surgical resection is the
only chance for cure. The 5-year survival rate is <5% making it
one of the leading causes of death (5). The conventional chemotherapy drugs
used are Oxaliplatin and Mitomycin, which are prone to resistance
by cancer cells thus compromising the efficacy of the drugs
(6,7). Therefore, it is necessary to search
for new drugs that are more effective and with less side
effects.
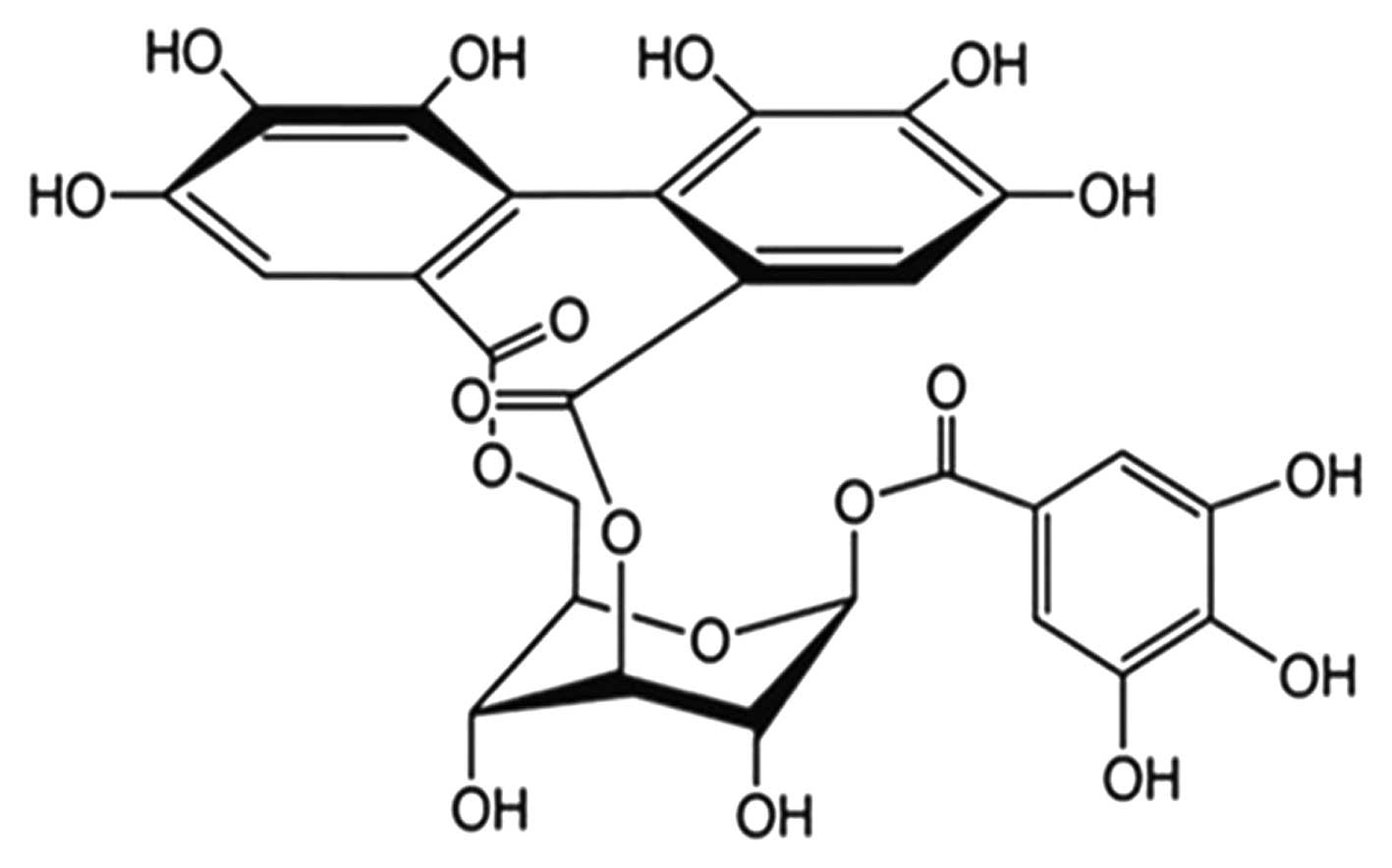
Corilagin
(β-D-glucopyranose,cyclic3,6-[(1R)-4,4′,5,5′,
6,6′-hexahydroxy(1,1′-biphenyl)-2,2′-dicarboxylate]1-(3,4,5-trihydroxybenzoate)
is a polyphenol tannic acid from plant extracts. It has both
anti-inflammatory and anti-oxidant properties, and is commonly used
for treating bacterial and viral infections, and diabetes (8,9). The
molecular formula of corilagin is
C27H22O18, and its molecular
weight is 634.45 (structure is shown in Fig. 1). Corilagin inhibited the growth of
E. coli and C. albicans by disrupting their membrane
permeability but not acting on membrane integrity (10). Corilagin could attenuate
TBHP-induced oxidative stress in microcells, and its protective
effect may be due to its antioxidant and anti-apoptotic properties,
which made corilagin a potential candidate for the treatment of
oxidative stress-induced neurodegenerative diseases (11). Corilagin significantly reduced the
production of pro-inflammatory cytokines and mediators such as
TNF-α, IL-1β, IL-6 and COX-2 (12).
In the past few years, research on corilagin was
focused on its use as an antiviral, hypolipemic, hypotensive and
anticoagulation agent (13).
However, there is scarce data on the effect of corilagin on cancer.
A study shows that corilagin arrests SMMC7721 cells at the G2/M
phase by downregulating p-Akt and cyclin B1/Cdc2 and upregulating
p-p53 and p21Cip1 (14). Another
study (14) of corilagin in human gastric cancer
demonstrated that corilagin nanoparticle induced cell apoptosis via
the mitochondrial pathway. These observations suggest that
corilagin possesses a potential anticancer effect. However, the
role of corilagin in cholangiccarcinoma remains largely unknown. In
our preliminary studies, we demonstrated that CCA cell
proliferation, migration, invasion and apoptosis were affected by
corilagin treatment.
The Notch signaling pathway is an evolutionarily
conserved pathway which plays an important role in cell fate
determination, proliferation, differentiation an survival (16,17).
The Notch system in vertebrates comprises four receptors: Notch1,
Notch2, Notch3 and Notch4 and at least five ligands from the
families Delta and JAG/Serrate (DSL): JAG1, JAG2, Delta-like
(Dll)-1, Dll-3 and Dll-4 (18,19).
Membrane localization of Notch requires S1 cleavage of precursor of
the Notch receptor. This event occurs in the Golgi network by the
action of a furin-like convertase.
Activation of Notch consists of two consecutive
cleavages of the transmembrane receptor upon the binding of a Notch
ligand, which triggers S2 cleavage. S2 cleavage releases the Notch
extracellular domain (NECD) from the heterodimer and creates a
membrane tethered Notch extracellular truncation (NEXT), which
becomes a substrate for γ-secretase. S3 is cleaved by γ-secretase
at sites 3 and 4. This last cleavage occurs on the plasma membrane
and/or in endosome. The new mobile cytoplasmic subunit [Notch
intracellular domain (NICD)] is translocated to the nucleus, where
it interacts with members of the DNA-binding protein, recombination
signal binding protein for immunoglobulin kappa J (RBP-Jκ) or
CBF1/Su(H)/Lag1 (CSL) family of transcription factors (20,21).
Now we known that there are several mechanisms of corilagin in a
tumor, such as MAPK, NF-κB and TGF-β1 signaling (22). However the involvement of corilagin
in the regulation of Notch pathway remains unknown. In this study,
we investigated the effect of corilagin regulated tumor development
and occurrence through Notch signaling pathway in vitro and
in vivo.
Materials and methods
Cancer cell culture
The human cancer cells QBC9939, MZ-Cha-1 were
obtained from the American Type of Culture Collection (ATCC,
Rockville, MD, USA). They were separately cultured in 100-mm
culture dish (Corning Glass Works, Corning, NY, USA) using
RPMI-1640 and DMEM medium (Gibco, Grand Island, NY, USA) with 5%
fetal bovine serum (Gibco), 100 μg/ml streptomycin and 100 U/ml
penicillin at 37°C in 5% CO2 humidified incubator.
Cell proliferation assay
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide
(MTT, Amresco, Solon, OH, USA) was used to detect the effect of
corilagin on the proliferation of CCA cell lines. CCA cells
(1.5–2.0×103 cells per well in 100 μl medium) were
seeded in 96-well plates and incubated with corilagin (0, 10, 20,
30, 40, 50, 60, 70, 80, 90 μmol/l) starting the following day and
continuing for 48 h. Then a solution of 5 mg/ml MTT was dissolved
in PBS and 20 μl of MTT solution was added from step one to each
well containing cells. The plate was incubated at 37°C for 4 h.
Media was remove and 150 μl DMSO (Sigma-Aldrich, St. Louis, MO,
USA) was added to wells with vibration to dissolve crystals, and
the plate was incubated for 5 min. The absorbance at wavelength 570
nm was measured using a plate reader.
Wound-healing assay
CCA cells (2×105 cells/well) were plated
in a 6-well plate, and a scratch created using a sterile 10 μl
pipette tip when they were confluent, then washed twice to remove
non-adherent cells. Cells were cultured in normal medium and
treated with corilagin at 40 μM for 48 h. Photomicrographs were
captured at 48 h after wounding using a microscope.
Invasion assay
The invasion assay was performed in 24-well
transwell plates (Millipore, Billerica, MA, USA) with uncoated,
8-mm pore, polycarbonate membranes. In brief, serum-starved cells
were harvested and re-suspended at a concentration of
2×105 cells/ml in serum-free medium, and 200 μl was
added to the upper chamber. After treatment with corilagin for 48
h, CCA cells that had not invaded were discarded using a cotton
swab. Cells that had invaded the lower compartment were stained
with 0.1% crystal violet. The number of cells on the lower side of
the membrane was quantified by average cell counts from six random
fields in each well, under a microscope at ×200 magnification, and
images were captured.
Cell cycle analysis
CCA cells were seeded in 60-mm plates
(1–2×105 cells/plate) and incubated with corilagin
(20–40 μmol/l) or DMSO as a control the next day. Cells were
harvested after 24 h of treatment, washed with PBS, fixed with
ice-cold 70% ethanol, and incubated at 4°C overnight. The cells
were then washed with PBS and treated with 10 μg/ml RNase (Amresco)
for 30 min at room temperature. Cell nuclei were stained with 50
μg/ml propidium iodide (PI, Sigma) for 15 min at room temperature
in the dark and detected by flow cytometry (BD Biosciences,
Franklin Lakes, NJ, USA). The cell cycle distribution was analyzed
using ModFit3.0 software.
Apoptosis analysis
CCA cells were seeded in a 60-mm dish
(1–2×105 cells/dish) and incubated with corilagin (20–40
μmol/l) or DMSO as a control. The cells were harvested after 24 h
of incubation, washed with PBS, and stained with Annexin
V-fluorescein and PI according to the manufacturer's instructions
for the Annexin V-FLUOS Staining kit (Roche, Mannheim, Germany).
The stained cells were analyzed by flow cytometry.
Real-time PCR analysis
Total RNA was extracted by the guanidinium
thiocyanate-phenol-chloroform method, purified, and subjected to
reverse transcription with random hexanucleotide primers (Takara,
Foster City, CA, USA). The resulting cDNA was then subjected to
real-time PCR analysis with SYBR Green PCR master mix (Applied
Biosystems, Foster City, CA, USA) and 200 nM gene-specific primers.
Assays were performed in triplicate with a StepOnePlus real-time
PCR system (Applied Biosystems). The amplification protocol
comprised initial incubation at 60°C for 30 sec and at 95°C for 3
sec, followed by 40 cycles. The sequences of the various primers
(sense and antisense, respectively) are listed below:
5′-AGCTGCCTTTCATTTAGCACTCTAC-3′ and
5′-TTAAGACTTTCCAGGGTATATCCAGTC-3′ for Cyclin A,
5′-TACCTATGCTGGTGCCAGTG-3′ and 5′-CACATCCAGATGTTTCCATTG-3′ for
Cyclin B, 5′-AGAGGCGGAGGAGAACAAAC-3′ and 5′-TGAGGCGGTAGTAGGACAGG-3′
for Cyclin D, 5′-GGATTGTGGCCTTCTTTGAGT-3′ and
5′-TCAAACAGAGGTCGCATGCT-3′ for Bcl-2, 5′-GCCGAGGCTTGAGGTATATT-3′
and 5′-TCC TTCTTCAGAGGCAGCAT-3′ for Caspase 3,
5′-TATCATGGAGAAGAGGCGAAGG-3′ and 5′-TTCTCTAGCTTGGAATGCCGG-3′ for
Hes1, 5′-CTACCCAAAAGTAATCATCTTAAGTG-3′ and
5′-CCCAACCATGACAAGATTTTCC-3′ for Fbw7 exon8,
5′-TTTCTGTTTCTCCCTCTG-3′ and 5′-GAGCATTTAAGGGAGAGATAAGAG-3′ for
Fbw7 exon9, 5′-TCTTCCAGCCTTCCTTCCT-3′ and
5′-AGCACTGTGTTGGCGTACAG-3′ for β-actin.
Western blot analysis
The CCA cells were harvested and lysed in RIPA
buffer (1% Triton, 0.1% SDS, 0.5% deoxycholate, 1 mM EDTA, 20 mM
Tris (pH 7.4), 150 mM NaCl, 10 mM NaF, 1 mM
Na3VO4, 0.1 mM phenylmethylsulfonyl
fluoride). Then, cell lysates were cleared by centrifugation and a
BCA assay (Pierce, Rockford, IL, USA) was used to detect the
concentration of protein in the supernatants using a Varioskan
multimode microplate spectrophotometer (Thermo Fisher Scientific,
Waltham, MA, USA). Protein was prepared for the same concentration.
The prepared protein was separated by SDS-PAGE and transferred onto
nitrocellulose (NC) membranes. The nitrocellulose membranes were
blocked with 10% non-fat milk in PBS at 37°C for 1 h and incubated
with primary antibodies (Cell Signaling Technology, Danvers, MA,
USA) overnight at 4°C. After washing three times for 10 min each in
PBST, membranes were incubated with HRP-conjugated secondary
antibody for 1 h at 37°C. Chemiluminescence detection was performed
using SuperSignal WestDura Extended Duration Substrate (Pierce),
and bands were visualized using the Automatic digital Imaging
Analysis System (Tanoo-5500, Shanghai, China).
Animals and treatment
Eight weeks old athymic nude mice, weighing
approximately 15–20 g, were purchased from the animal unit of
Xiamen University and maintained in a sterile facility, in
accordance with the institutional guidelines on animal care, with
the required consistent temperature and relative humidity. All the
procedures were approved by the Animal Research Ethics Committee.
Sixteen athymic nude mice were injected subcutaneously with the
human CCA cells. They were housed in sterile conditions. Tumor size
was measured by an electronic calliper daily. When tumor size
reached a mean volume of ~200 mm3, where tumor volume
was calculated by the formula (length × width × width)/2, they were
randomly divided into four groups. Corilagin at a concentration of
20 mg/kg were administrated intraperitoneally for a continuous
period of 3 days starting from day 1 to day 30. Control group
received equal volume of vehicle intraperitoneally. Each group
consisted of 4 mice. On day 30, all the mice were sacrificed and
tumors were collected for further investigation.
Statistical analysis
The data was analyzed from at least three
independent experiments. The statistical significance of
differences between two groups was evaluated using Student's
t-test. SPSS 13.0 software (SPSS Inc., Chicago, IL, USA) was used
to carry out all the statistical analyses. P-values were two-sided,
and v P-value <0.05 was considered to indicate significance. The
data are presented as the mean ± SD.
Results
Corilagin effectively suppresses CCA cell
proliferation
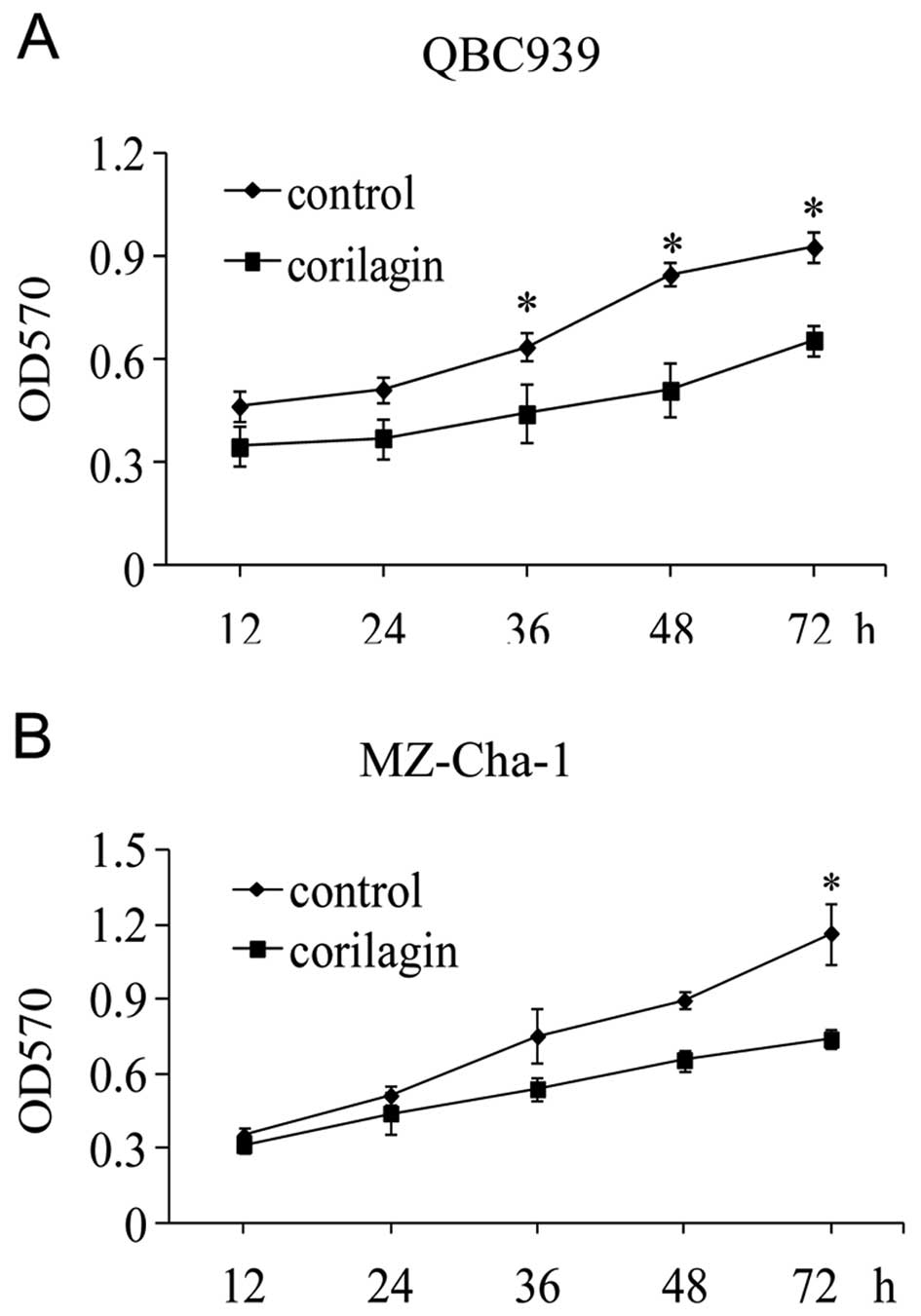
To investigate the effect of corilagin on CCA cell
proliferation, we first determined the 50% cell inhibitory
concentration (IC50) of corilagin. QBC939 and MZ-Cha-1
cells were treated in vitro with increasing concentrations
of corilagin for 48 h and then analyzed by the MTT assay. The means
± standard errors of IC50 were 39.73±1.28 μmol/l for
QBC939 cells and 36.88±1.32 μmol/l for MZ-Cha-1 cells,
respectively. Then we treated the cell lines with 30 μmol/l of
corilagin for the indicated times (Fig. 2A and B). The result showed that
corilagin could efficiently suppress CCA cell proliferation at 24
and 72 h.
Corilagin decreases CCA cell cycle
progression
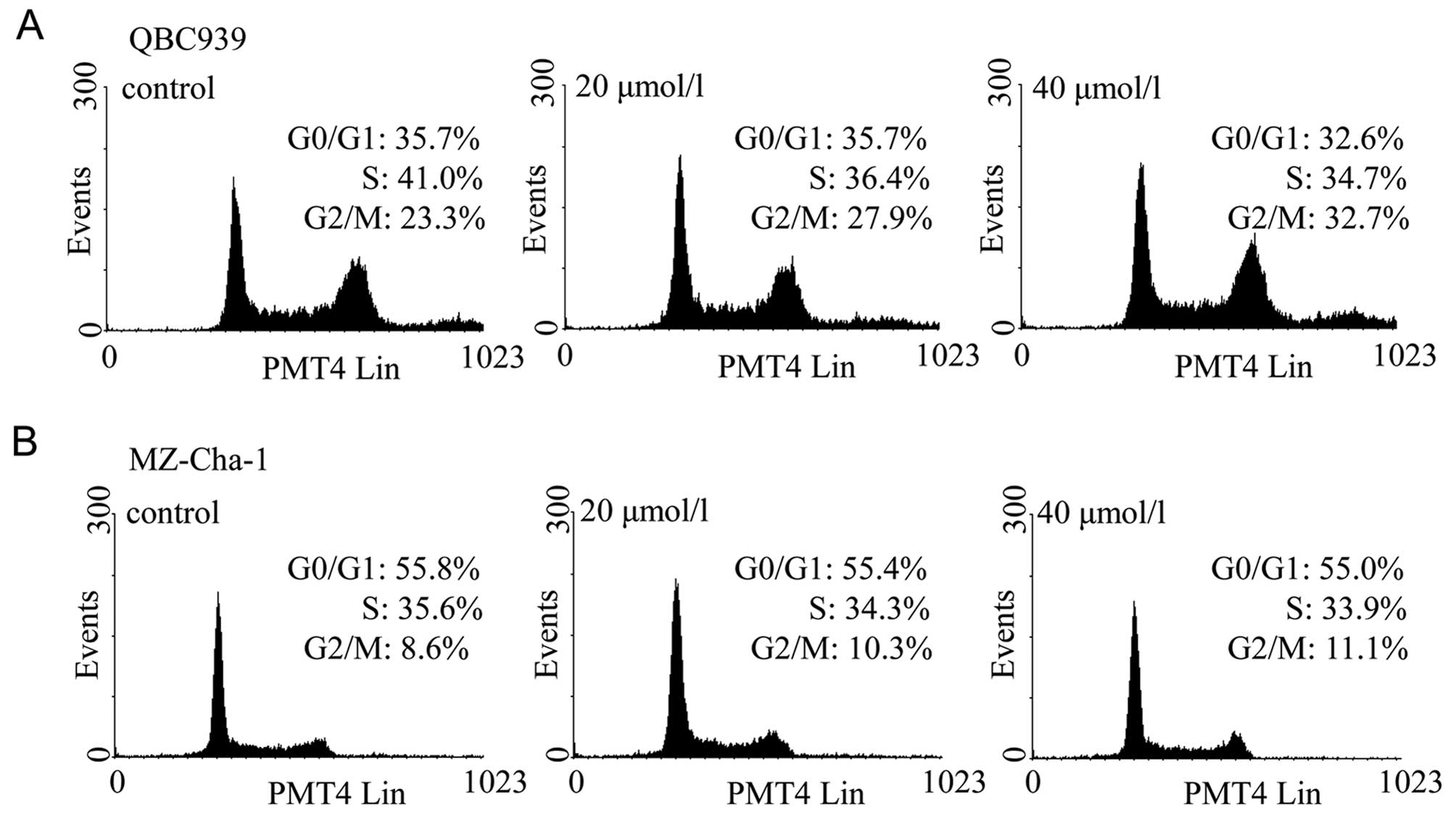
As corilagin inhibited cell proliferation, cell
cycle analysis was performed to examine whether corilagin could
induce cell cycle arrest. Two CCA cell lines were treated with
corilagin at increasing concentrations for 24 h and then stained
with PI. Flow cytometric analysis showed that the proportions of
CCA cells treated with corilagin at G2/M phase was increased
compared with control cells, and this was associated with a
decrease in proportion of cells at S phase (Fig. 3). In QBC939 cells, the proportion
of cells in the G2/M phase increased from 23.3% to 27.9% and 32.7%
with 20 and 40 μM corilagin, respectively (Fig. 3A). Subsequently, inhibition of the
G1 phase and S phase was observed. Corilagin has the same effect in
MZ-Cha-1 cells, but with less potency (Fig. 3B). These results suggest that
corilagin regulates cell cycle by arresting CCA cells in the G2/M
phase, and inhibiting cell growth.
To explore the mechanism underlying the changes in
cell cycle progression, several cell cycle-related genes were
examined by real-time PCR. The mRNA levels were compared between
control cells and corilagin treated cells. The result showed that
there was no significant changes in the expression of cyclin A,
cyclin B and cyclin D. The exact mechanism of corilagin regulation
of cell cycle progression needs to be explored in future
studies.
Corilagin inhibits migration and invasion
of CCA cells
The processes of migration and invasion are integral
to embryonic development and the functioning of adult organisms.
Deregulation of these processes contributes to the development of
tumor and ultimately metastasis (23,24).
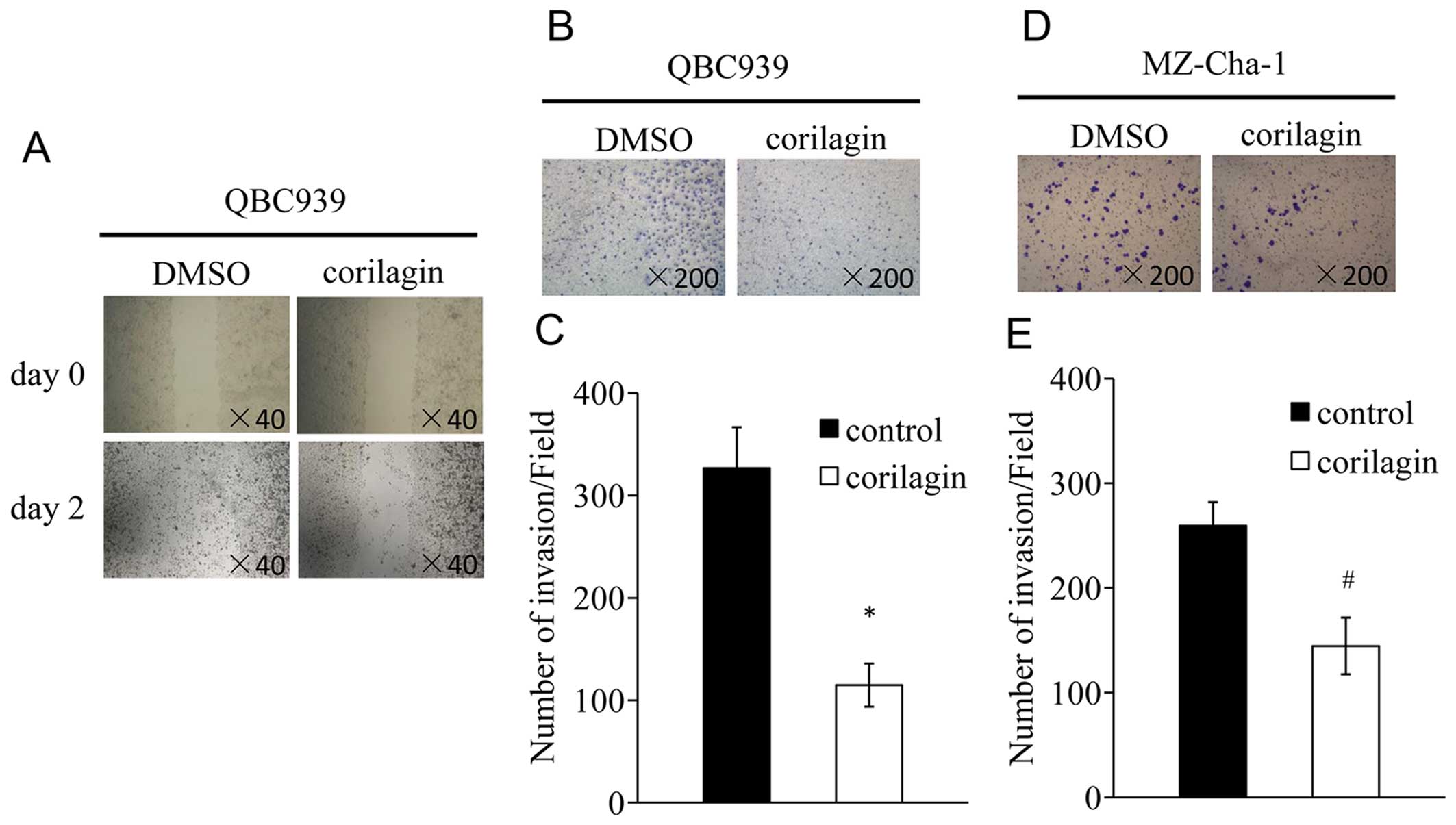
We tested whether corilagin could inhibit the migration and
invasion of CCA cells. We performed monolayer wound healing assay
to evaluate the effect of corilagin on migration of QBC939 cells. A
scratch was created with a pipette tip when the cells were
confluent, and treated with corilagin at 40 μM for 48 h. The result
indicated that corilagin significantly decreased cell migration
(Fig. 4A). For transwell invasion
assay, QBC939 and MZ-Cha-1 cells were treated with corilagin at
indicated concentrations for 12 h. The number of cells that had
invaded was less after corilagin treatment compared with control in
QBC939 cells (Fig. 4B). Similar
result was observed in MZ-Cha-1 cells (Fig. 4D). Together, these results suggest
that corilagin could significantly decrease the migration and
invasion of CCA cells.
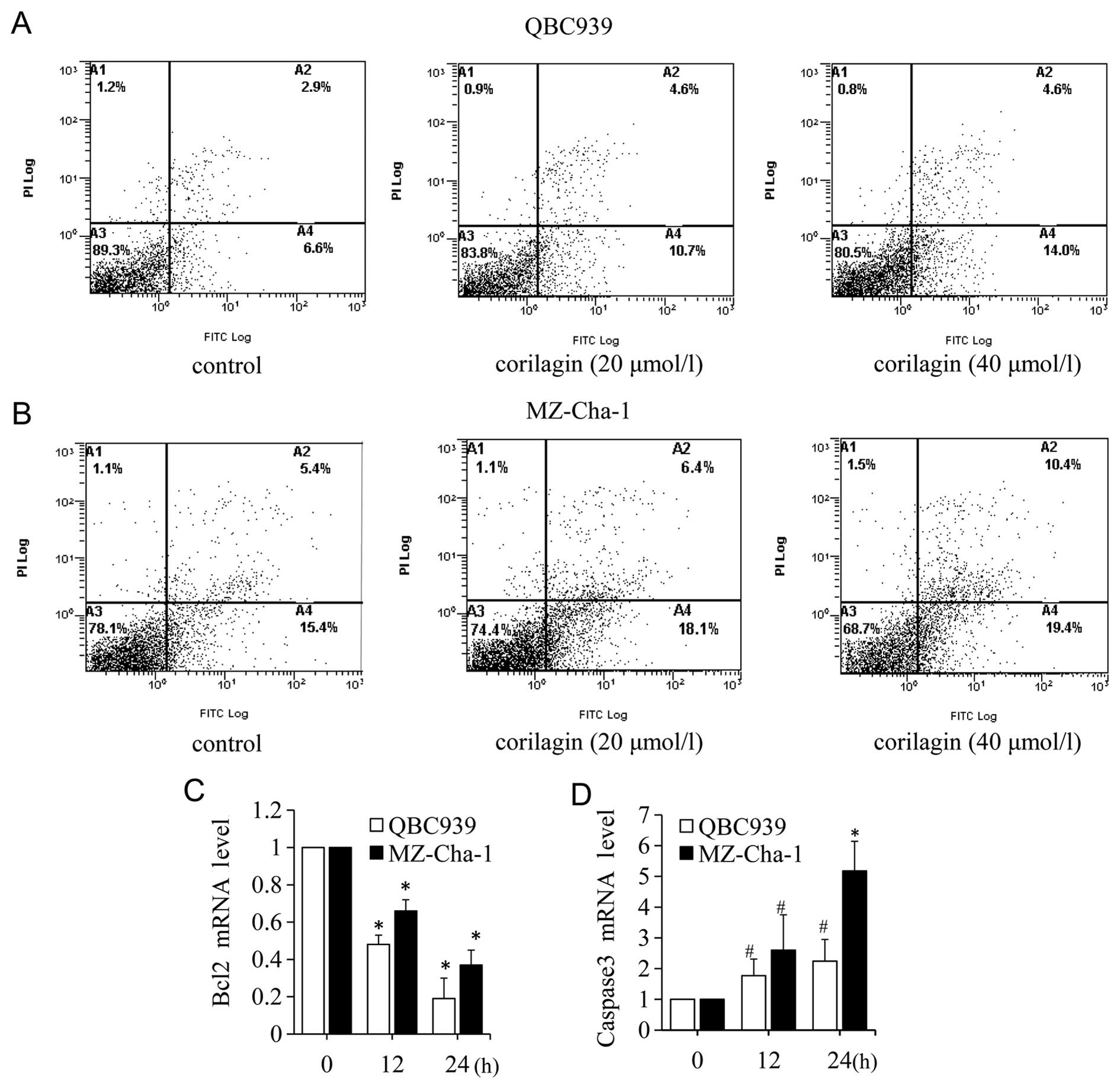
Corilagin induces CCA cell apoptosis
The flow cytometry analysis of Annexin V-Fluos and
PI staining revealed the quantity of viable (Annexin
V−/PI−), early apoptotic (Annexin
V+/PI−), late apoptotic (Annexin
V+/PI+) and necrotic (Annexin
V−/PI+) cells. To assess the antitumor
effects of corilagin on CAA cells, we investigated the impact of
corilagin on apoptosis. QBC939 and MZ-Cha-1 cells were incubated
with increasing concentrations of corilagin. As shown in Fig. 5A and B, for both QBC939 and
MZ-Cha-1 cells, the percentages of early apoptotic cells increased
after treatment with corilagin. After 40 μM corilagin treatment in
QBC939 cells, the early apoptotic population was elevated from 6.9
to approximately 14.0%, and the late apoptotic population was
elevated from 2.9% to approximately 4.6%, whereas, in MZ-Cha-1
cells the early apoptotic population was 19.4% and the late
apoptotic was 10.4%, and compared with control 15.4 and 5.4%,
respectively. To further investigate the apoptotic molecular
mechanism, we detected the apoptosis-related gene bcl-2 and caspase
3 using real-time PCR in CCA cells. We examined the genes at early
(12 h) and late (24 h) stages of corilagin treatment. As shown in
Fig. 5C, the expression of bcl-2
mRNA was significantly decreased at 12, 24 h after corilagin
treatment. While caspase 3 mRNA was strongly increased at 12, 24 h
(Fig. 5D). These results indicated
that bcl-2 and caspase 3 were involved in the corilagin-induced CCA
cell apoptotic process.
Corilagin effectively suppresses
Notch-mTOR pathway
Notch-mTOR pathway plays an important role in the
development and cell fate determination (25). To determine whether corilagin was
involved in Notch-mTOR pathway, we firstly identified the protein
expression patterns of Notch-mTOR signaling in CCA cells. Western
blot analyses showed that corilagin had significant effects on
Notch1 at 12, 24, 48 h after corilagin treatment; moreover,
corilagin strongly inhibited the expression of mTOR at 48 h
(Fig. 6A). In addition, we
investigated the expression of p-Akt and p-Erk1/2, the direct
target gene of Notch that function as an important downstream
effector in Notch-mTOR pathway. As shown in Fig. 6A, the expression of p-Akt and
p-Erk1/2 was significantly reduced at 48 h after corilagin
treatment.
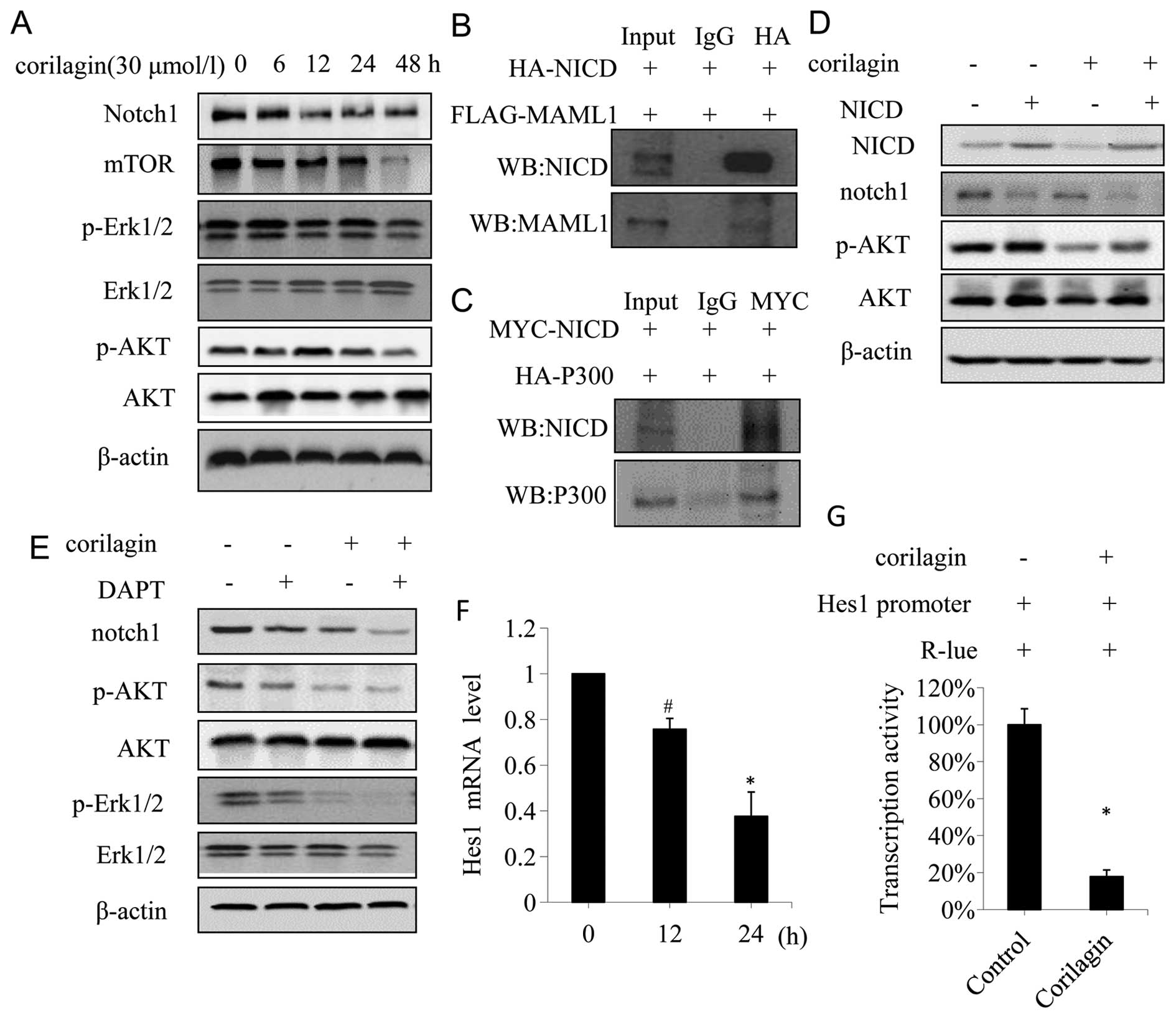 | Figure 6Corilagin suppress Notch-mTOR pathway
and inhibits Notch1 transcriptional activity in CCA cells. (A)
QBC939 cells were treated with 30 μmol/l corilagin for different
times, the expressions of notch1, mTOR, pERK1/2 and pAKT were
detected by western blot analyses. (B) QBC939 cells were
co-transfected with HA-NICD and FLAG-MAML1, and cell lysates were
IP for HA, the immunoprecipitates were anlyzed by western blot for
NICD and MAML1. (C) QBC939 cells were co-transfected with MYC-NICD
and HA-P300, and cell lysates were IP for MYC, the
immunoprecipitates were analyzed by western blotting for NICD and
P300. (D) QBC939 cells transfected with either control vector or
NICD-expressing construct were treated with or without corilagin,
and the expression of NICD, notch1, AKT and pAKT were detected by
western blotting. (E) QBC939 cells were treated with corilagin (30
μmol/l) alone or in combination with DAPT (1 nmol/l) for 24 h, cell
lysates were subjected to western blot analysis for the expression
of notch1, pERK1/2 and pAKT. (F) QBC939 cells were treated with 30
μmol/l corilagin for indicated time and the mRNA level was detected
by real-time PCR. (G) Corilagin increased the activities of
HES1-promoter in QBC939 cells, reporter activities were measured
and normalized. *P<0.01, #P<0.05. |
Notch receptors are synthesized in the endoplasmic
reticulum as an inactive single peptide precursor, which is
proteolytically cleaved by a furin-like convertase in the
trans-Golgi network before it reaches the plasma membrane (26). The third cleavage production, Notch
intracellular domain (NICD), binding to ubiquitous transcription
factor CSL, mastermind-like protein (MAML1) and P300 has been
reported (27). In order to
determine the interactions between NICD, MAML1 and P300, we
performed a co-immunoprecripitation assay in CCA cells
co-transfected with HA-NICD, Flag-MAML1, and MYC-NICD, HA-P300,
respectively. The result showed that Flag-MAML1 was
co-immunoprecipitated with HA-NICD (Fig. 6B), and HA-P300 was
co-immunoprecipitated with MYC-NICD (Fig. 6C), suggesting that NICD interacted
with MAML1 and P300 in CCA cells. To further confirm the effect of
corilagin on Notch signaling, 293T cells transfected with NICD were
treated with or without corilagin. As shown in Fig. 6D, NICD enhanced the phosphorylation
of AKT. In contrast to NICD, corilagin decreased the
phosphorylation of AKT, and counteracted the phosphorylation of AKT
induced by NICD (Fig. 6D, lane 4).
It is noteworthy that NICD inhibited the expression of Notch1
(Fig. 6D, lane 2), and this might
be due to the negative feedback regulation (28). Secretase inhibitor DAPT, which
affects Notch signal pathway, is used to treat Notch-mediated
cancer such an breast cancer (29).
So far there is scarce data reported on DAPT used as
a therapy in CCA. In order to confirm the role of corilagin in
Notch active CCA cells, we assessed the effect of DAPT in CCA cells
treated with corilagin and DAPT. As shown in Fig. 6E, DAPT inhibited the expression of
Notch1, p-AKT and p-Erk1/2, while corilagin reduced expression of
these proteins, which was consistent with previous studies.
Corilagin and DAPT co-procession had a more significant inhibitory
effect on Notch1, p-AKT, and p-Erk1/2. To determine whether
corilagin was involved in the regulation of the transcriptional
activity of Notch1 target genes, a reporter assay was conducted in
QBC939 cells using the luciferase reporter gene. In mammals, the
best-characterized Notch target gene belongs to the Hes family
(30).
To confirm that corilagin affected the CCA cells
through the Notch-mTOR pathway, we investigated the mRNA level of
Hes1, which is one of hairy/enhancer of split (HES) family
proteins. We found that the mRNA level of Hes1 was significantly
decreased after corilagin treatment compared with the control group
(Fig. 6F). Furthermore, we
examined the transcriptional activity of Hes1 promoter after
corilagin treatment. As shown in Fig.
6G, corilagin reduced the transcriptional activity of Hes1
promoter. These results suggested that corilagin could reduce the
Hes1 mRNA level through inhibiting its promoter activity. All the
results above suggested that corilagin might affect the notch
signal pathway by restraining the active cleavage NICD, thereby
affecting the transcriptional activity upon binding of NICD and
ubiquitous transcription factor CSL which regulated the expression
of target gene Hes1, and eventually altered the expression of
downstream proteins such as p-AKT, p-Erk1/2. Thus, corilagin
regulated Notch signal pathway in CCA cells.
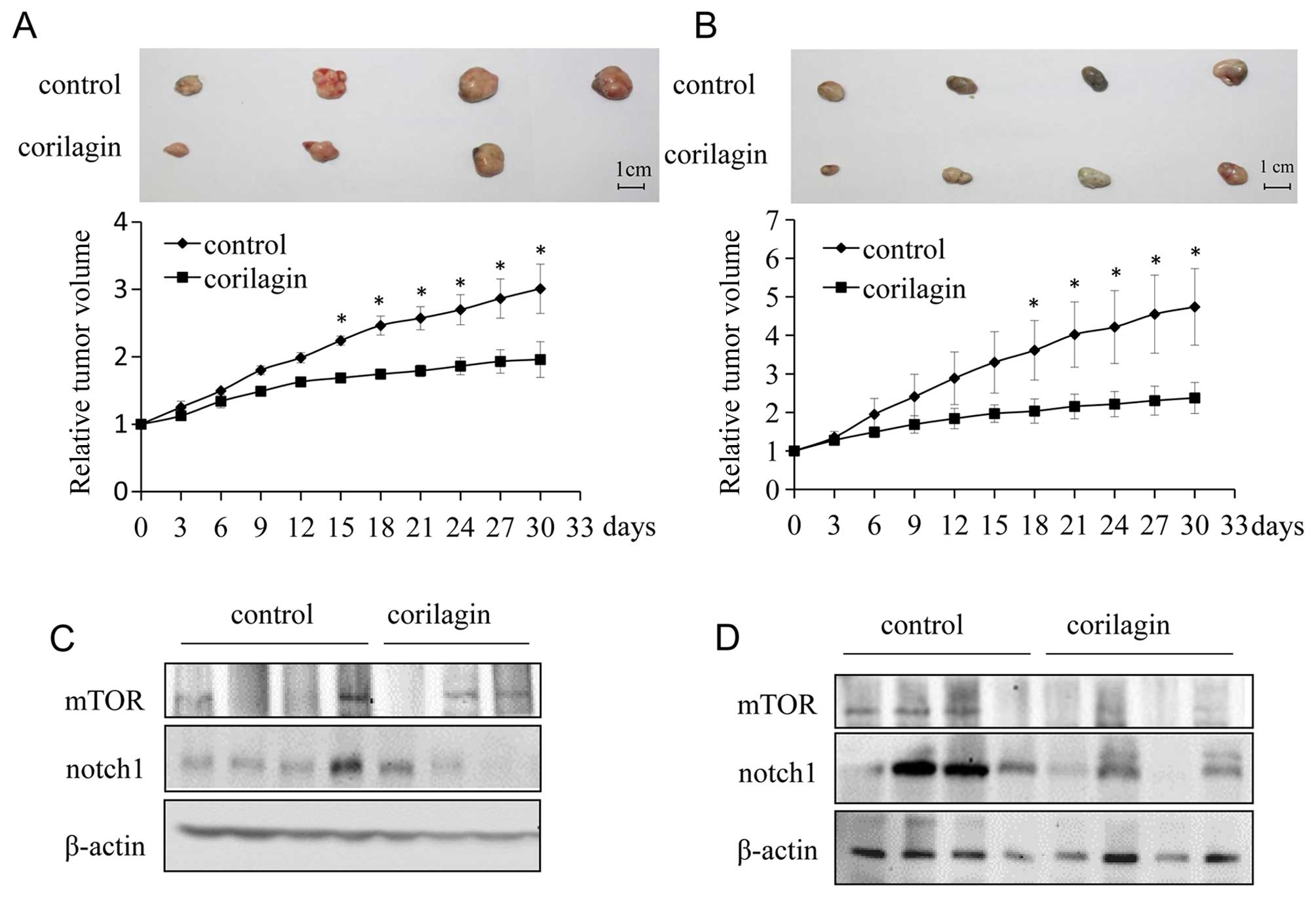
Corilagin inhibits tumor growth
progression in vivo
To investigate the effect of corilagin on the growth
of CCA xenograft tumors in nude mice, we injected either vehicle or
corilagin intraperitoneally. Tumor volume was quantified using
electronic calliper. Corilagin treated CCA tumors grew much slower
than control tumors. At the end of study (30 days),
corilagin-treated mice exhibited markedly reduced tumors as
compared with control mice (Fig. 7A
and B). To further study the role of corilagin in CCA, we
investigated the expression of Notch1 and mTOR in vivo.
Proteins were extracted from xenograft tumors in a tumor-bearing
nude mouse model. As shown in Fig. 7C
and D, the expression of Notch1 and mTOR were significantly
reduced in tumors from corilagin treated group compared with the
control group, which was consistent with the in vitro
experiments described above. Therefore, corilagin regulated the
Notch-mTOR pathway in vivo as well as in vitro.
Discussion
Corilagin is a gallotannin identified in several
plants and is the major active component of Phyllanthus
niruri L. Phyllanthus niruri L. is a well-known
medicinal plant that has shown hepatoprotective, antiviral,
antibacterial, analgesic, anti-spasmodic and anti-diabetic effects;
however, there are few studies describing its antitumor activity.
Herein we investigated the effect of corilagin on CCA and found its
distinct antitumor activity. First we examined the effect of
corilagin on phenotype of CCA cell lines and found that corilagin
was able to inhibit the proliferation and cell cycle, reduce
migration and invasion, and promote apoptosis of CCA cells, which
suggested that corilagin play an important role in the development
of CCA. Corilagin inhibited CCA cell proliferation by inducing G2/M
phase arrest, but had no significant changes in G2/M phase in
MZ-Cha-1 cells after corilagin treatment. These results indicated
that cell growth inhibition might be mainly through some other
ways, such as promoting MZ-Cha-1 cells apoptosis and necrosis.
According to the following results we confirmed that corilagin
simultaneously promoted early and late apoptosis in CCA cells.
Corilagin inhibited the mRNA level of bcl-2 and promoted caspase 3
associated with apoptotic gene detection.
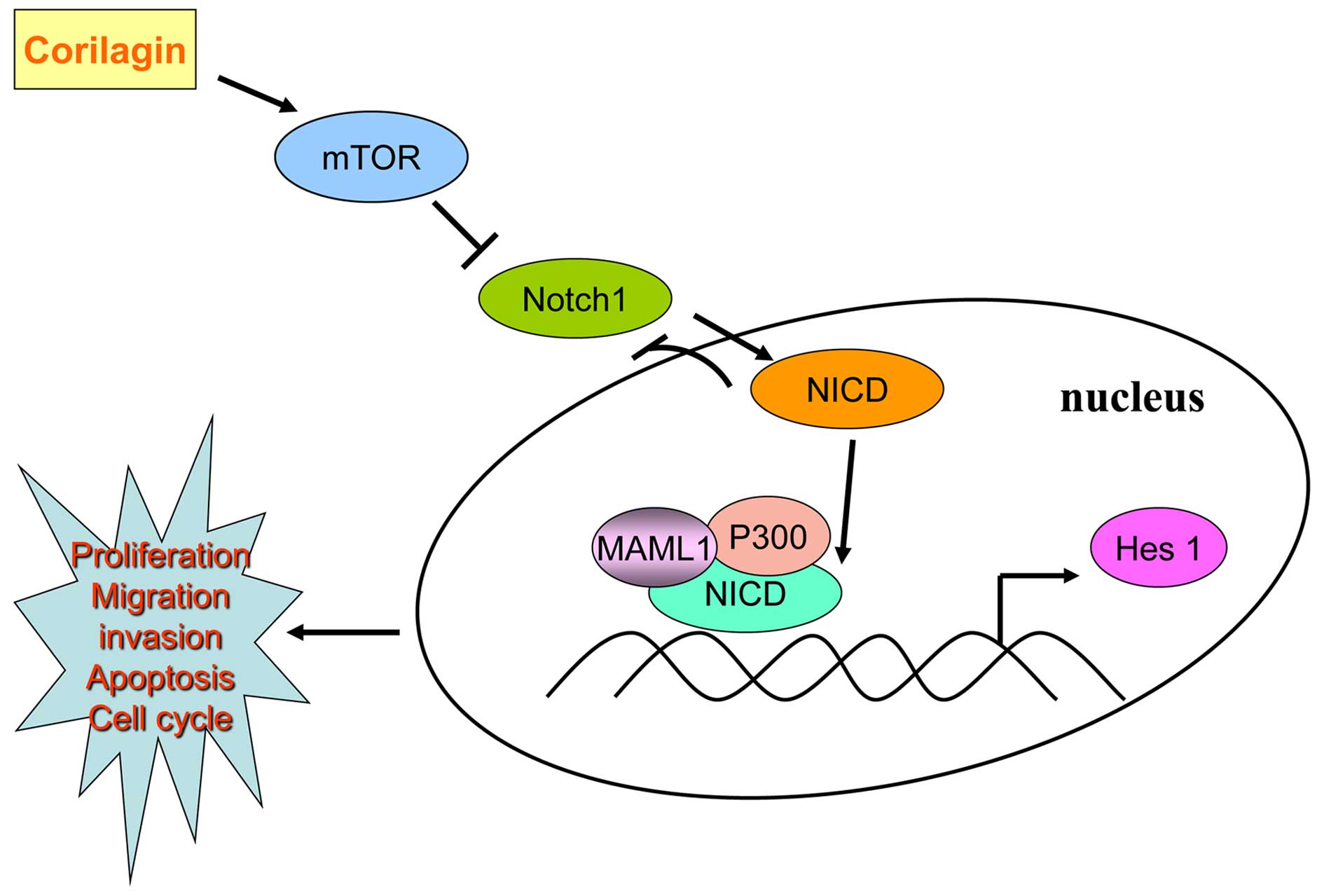
According to our results, corilagin could restrain
the expression of Notch1, which showed a time gradient decreases.
We found corilagin not only reduced Hes1 promoter activity, but
also inhibited the Hes1 mRNA level. Using co-immunoprecipitation
method we found that the active form of Notch intracellular
hydrolysis of NICD was able to form a complex with P300 and MAML1
which could combine with CSL to initiate transcription. Corilagin
regulating Notch signal pathway possibly inhibited the expression
of Notch1 thus reducing the NICD form. In the absence of NICD, CSL
performed transcriptional suppression state which restrained the
activity of PI3K/AKT expression. After addition of the Notch signal
pathway inhibitor DAPT, we found that DAPT treatment alone could
inhibit the protein expression of Notch1, p-AKT and p-Erk1/2. When
corilagin was combined with DAPT, the inhibition became more
enhanced. Furthermore, NICD plasmid was transfected in CCA cells,
and p-AKT and p-Erk1/2 expression increased as compared to control
group, which was a direct effect of NICD promotion on the
expression of downstream target genes; whereas, exogenous NICD
caused decreased expression of Notch1, which was a negative
feedback inhibition mechanism that has been reported (31). These results suggested that
corilagin effect the expression of downstream target protein
Notch1, mTOR, p-AKT and p-Erk1/2 to regulate the occurrence and
development of CCA through Notch signal pathway, as shown in
Fig. 8.
In consideration of the effect of corilagin on CCA
cells in vitro, we further demonstrated that corilagin
profoundly suppressed CCA growth and significantly inhibited Notch1
and mTOR protein expression in vivo. It is well established
that, angiogenesis is an underlying promoter of tumor growth,
invasion, and metastases (32). We
transfected 293T cells with NICD plasmid and found that NICD did
not effectively restore corilagin reduced p-AKT protein level. This
showed, corilagin could regulate other pathways besides NICD such
as NF-κb (12). However which
other pathways are involved in CCA treatment with corilagin still
needs further exploration.
Current antitumor drugs have poor selectivity and
high toxicity. Research on the action of natural product drugs
promise to reveal the intricate interplay of biology, that will
provide new opportunities for drug discovery. In our research,
corilagin significantly inhibited CCA cell growth with lower
cytotoxic effects on normal cells. To further investigate corilagin
role in CCA, we confirmed Notch signal pathway involved in CCA
occurrence and development treated with corilagin. corilagin
enhanced the cytotoxic effect of CCA cells combined with common
chemotherapeutic drugs. Corilagin, a monomer component of
traditional Chinese medicine, is now pushed to the front, and could
became a major herbal antitumor drug based on its safety and
effective features. With the recent technological advances, a new
golden age of nature products drug discovery is dawning (33). We consider that corilagin is a new
promising candidate, as a potential antitumor drug for treating
cancer.
Acknowledgements
This work was supported by funds from the National
Natural Science Foundation of China (grant number: 81272246 to
W.L.), the Xiamen Municipal Science and Technology Innovation Fund
Project (grant number: 3502Z20124049 to W.L.). This work was also
supported by grants from the Natural Science Foundation of China
(grant number: 81274149 to Y.M.). We thank Professor C.D. Yu (Key
Laboratory of Xiamen University) for providing a number of regents.
We also thank Mr. D.Q. Zeng (School of Pharmacy, Xiamen University)
for technical support in analysis.
Abbreviations:
|
CCA
|
cholangiocarcinoma
|
|
NICD
|
Notch intracellular domain
|
|
CSL
|
CBF1/Su(H)/Lag-1
|
References
|
1
|
Singal AK, Vauthey JN, Grady JJ and
Stroehlein JR: Intrahepatic cholangiocarcinoma - frequency and
demographic patterns: Thirty-year data from the M.D. Anderson
Cancer Center. J Cancer Res Clin Oncol. 137:1071–1078. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
McGlynn KA, Tarone RE and El-Serag HB: A
comparison of trends in the incidence of hepatocellular carcinoma
and intrahepatic cholangiocarcinoma in the United States. Cancer
Epidemiol Biomarkers Prev. 15:1198–1203. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Al-Bahrani R, Abuetabh Y, Zeitouni N and
Sergi C: Cholangiocarcinoma: Risk factors, environmental influences
and oncogenesis. Ann Clin Lab Sci. 43:195–210. 2013.PubMed/NCBI
|
|
4
|
Tepsiri N, Chaturat L, Sripa B, Namwat W,
Wongkham S, Bhudhisawasdi V and Tassaneeyakul W: Drug sensitivity
and drug resistance profiles of human intrahepatic
cholangiocarcinoma cell lines. World J Gastroenterol. 11:2748–2753.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shaib Y and El-Serag HB: The epidemiology
of cholangiocarcinoma. Semin Liver Dis. 24:115–125. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Poggi G, Quaretti P, Minoia C, Palumbo I,
Villani L, Amatu A, Teragni C, Scelsi M, Zappoli F and Bernardo G:
Oxaliplatin-eluting microspheres for the treatment of intrahepatic
cholangiocarcinoma: A case report. Anticancer Res. 28:2987–2990.
2008.PubMed/NCBI
|
|
7
|
Shitara K, Ikami I, Munakata M, Muto O and
Sakata Y: Hepatic arterial infusion of mitomycin C with degradable
starch microspheres for unresectable intrahepatic
cholangiocarcinoma. Clin Oncol (R Coll Radiol). 20:241–246. 2008.
View Article : Google Scholar
|
|
8
|
Yeo SG, Song JH, Hong EH, Lee BR, Kwon YS,
Chang SY, Kim SH, Lee SW, Park JH and Ko HJ: Antiviral effects of
Phyllanthus urinaria containing corilagin against human enterovirus
71 and Coxsackievirus A16 in vitro. Arch Pharm Res. 38:193–202.
2015. View Article : Google Scholar
|
|
9
|
Rangkadilok N, Sitthimonchai S,
Worasuttayangkurn L, Mahidol C, Ruchirawat M and Satayavivad J:
Evaluation of free radical scavenging and antityrosinase activities
of standardized longan fruit extract. Food Chem Toxicol.
45:328–336. 2007. View Article : Google Scholar
|
|
10
|
Li N, Luo M, Fu YJ, Zu YG, Wang W, Zhang
L, Yao LP, Zhao CJ and Sun Y: Effect of corilagin on membrane
permeability of Escherichia coli, Staphylococcus aureus and Candida
albicans. Phytother Res. 27:1517–1523. 2013.
|
|
11
|
Chen Y and Chen C: Corilagin prevents
tert-butyl hydroperoxide-induced oxidative stress injury in
cultured N9 murine microglia cells. Neurochem Int. 59:290–296.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gambari R, Borgatti M, Lampronti I, Fabbri
E, Brognara E, Bianchi N, Piccagli L, Yuen MC, Kan CW, Hau DK, et
al: Corilagin is a potent inhibitor of NF-kappaB activity and
downregulates TNF-alpha induced expression of IL-8 gene in cystic
fibrosis IB3-1 cells. Int Immunopharmacol. 13:308–315. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Prasad KN, Yang B, Shi J, Yu C, Zhao M,
Xue S and Jiang Y: Enhanced antioxidant and antityrosinase
activities of longan fruit pericarp by ultra-high-pressure-assisted
extraction. J Pharm Biomed Anal. 51:471–477. 2010. View Article : Google Scholar
|
|
14
|
Wang B: Corilagin nanoparticle-induced
apoptosis in human gastric cancer SGC-7901 cells via the
mitochondrial pathway. Acta Pharmacol Sin. 34:14. 2013.
|
|
15
|
Ming Y, Zheng Z, Chen L, Zheng G, Liu S,
Yu Y and Tong Q: Corilagin inhibits hepatocellular carcinoma cell
proliferation by inducing G2/M phase arrest. Cell Biol Int.
37:1046–1054. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ranganathan P, Weaver KL and Capobianco
AJ: Notch signalling in solid tumours: A little bit of everything
but not all the time. Nat Rev Cancer. 11:338–351. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Takebe N, Harris PJ, Warren RQ and Ivy SP:
Targeting cancer stem cells by inhibiting Wnt, Notch, and Hedgehog
pathways. Nat Rev Clin Oncol. 8:97–106. 2011. View Article : Google Scholar
|
|
18
|
Miele L, Golde T and Osborne B: Notch
signaling in cancer. Curr Mol Med. 6:905–918. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Miele L, Miao H and Nickoloff BJ: NOTCH
signaling as a novel cancer therapeutic target. Curr Cancer Drug
Targets. 6:313–323. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhou W, Wang G and Guo S: Regulation of
angiogenesis via Notch signaling in breast cancer and cancer stem
cells. Biochim Biophys Acta. 1836.304–320. 2013.
|
|
21
|
Borggrefe T and Oswald F: The Notch
signaling pathway: Transcriptional regulation at Notch target
genes. Cell Mol Life Sci. 66:1631–1646. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang Z, Guo QY, Zhang XJ, Li X, Li WT, Ma
XT and Ma LJ: Corilagin attenuates aerosol bleomycin-induced
experimental lung injury. Int J Mol Sci. 15:9762–9779. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kimpton WG, McKenzie GA, Muller HK, Ruby
JC and Poskitt DC: Lymphocyte migration during the development of
regional lymph node anergy in experimental tumor growth. Cell
Immunol. 75:13–21. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Takahashi S, Hasebe T, Oda T, Sasaki S,
Kinoshita T, Konishi M, Ueda T, Ochiai T and Ochiai A: Extra-tumor
perineural invasion predicts postoperative development of
peritoneal dissemination in pancreatic ductal adenocarcinoma.
Anticancer Res. 21:1407–1412. 2001.PubMed/NCBI
|
|
25
|
El-Habr EA, Levidou G, Trigka EA,
Sakalidou J, Piperi C, Chatziandreou I, Spyropoulou A, Soldatos R,
Tomara G, Petraki K, et al: Complex interactions between the
components of the PI3K/AKT/mTOR pathway, and with components of
MAPK, JAK/STAT and Notch-1 pathways, indicate their involvement in
meningioma development. Virchows Arch. 465:473–485. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Takebe N, Nguyen D and Yang SX: Targeting
notch signaling pathway in cancer: Clinical development advances
and challenges. Pharmacol Ther. 141:140–149. 2014. View Article : Google Scholar :
|
|
27
|
Oswald F, Täuber B, Dobner T, Bourteele S,
Kostezka U, Adler G, Liptay S and Schmid RM: p300 acts as a
transcriptional coactivator for mammalian Notch-1. Mol Cell Biol.
21:7761–7774. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Stella MC, Trusolino L, Pennacchietti S
and Comoglio PM: Negative feedback regulation of Met-dependent
invasive growth by Notch. Mol Cell Biol. 25:3982–3996. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhao L, Ma Y, Gu F and Fu L: Inhibition of
Notch1 increases paclitaxel sensitivity to human breast cancer.
Chin Med J (Engl). 127:442–447. 2014.
|
|
30
|
Kuang SQ, Fang Z, Zweidler-McKay PA, Yang
H, Wei Y, Gonzalez-Cervantes EA, Boumber Y and Garcia-Manero G:
Epigenetic inactivation of Notch-Hes pathway in human B-cell acute
lymphoblastic leukemia. PLoS One. 8:e618072013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Huppert SS, Jacobsen TL and Muskavitch MA:
Feedback regulation is central to Delta-Notch signalling required
for Drosophila wing vein morphogenesis. Development. 124:3283–3291.
1997.PubMed/NCBI
|
|
32
|
Trédan O, Lacroix-Triki M, Guiu S,
Mouret-Reynier MA, Barrière J, Bidard FC, Braccini AL, Mir O,
Villanueva C and Barthélémy P: Angiogenesis and tumor
microenvironment: Bevacizumab in the breast cancer model. Target
Oncol. 10:189–198. 2015. View Article : Google Scholar
|
|
33
|
Shen B: A new gloden age of natural
products drug discovery. Cell. 163:1297–1300. 2015. View Article : Google Scholar : PubMed/NCBI
|