Introduction
Cancer-associated fibroblasts (CAFs) regulate not
only carcinogenesis, but also the immune evasion of cancer in the
tumor microenvironment (TME), which facilitates cancer cell
proliferation, expansion, and metastasis (1–5). The
TME is comprised of various types of cells, including microvascular
endothelial cells, immune cells, and CAFs, and these cells interact
directly and indirectly with each other maintaining homeostasis of
the TME. Among these cells, CAFs are important regulators of the
immunological microenvironment of the tumor. Sanz-Moreno et
al reported that CAFs regulate immune evasion in the TME by
various mechanisms, including the use of cytokines and cell
attachment (6). They demonstrated
that the ROCK and JAK1 signaling pathway generates a contractile
force in stromal fibroblasts, allowing remodeling of the
extracellular matrix and the creation of tracks for the collective
migration of squamous carcinoma cells. Furthermore, Gaggioli et
al demonstrated that the generation of these tracks by
fibroblasts was sufficient in enabling collective invasion of
squamous cell carcinoma cells (7).
NK cells play an important role in cancer immunity
in the TME. A review by Chan et al identified several
well-known ligands of NK paired or activating receptors that are
expressed on the cell surface of target cells, including malignant
cells (8). NK activating receptors
include NKp30, NKp44, NKp46, NKG2D, DNAX accessory molecule-1
(DNAM-1), and LFA-1 (9). In
addition, indoleamine 2,3-dioxygenase (IDO) is produced by various
malignant cells, inactivates NK cells, and evades the immune system
in the TME (10).
Poliovirus receptor (PVR/CD155) is a ligand of the
paired NK receptors, DNAM-1 (activating) and TIGIT (inhibiting). NK
cells can kill cancer cells expressing PVR via the DNAM-1-mediated
activating signaling (11,12). Several studies have demonstrated
that PVR overexpression in cancer cells significantly affects their
migration, invasion, proliferation, and metastasis (13). Although these previous studies have
investigated the interactions between NK cells and malignant cells,
there are few reports investigating the interaction of CAFs with NK
cells. A previous study reported that CAFs inhibit the IL-2-induced
cell-surface expression of the activating NK receptors, NKp44,
NKp30, and DNAM-1 (9). However,
there have been no studies investigating the association between NK
cell activity and PVR expression in CAFs.
Considering the NK cell-mediated immune evasion
mechanisms in the TME, we hypothesized that in addition to
malignant cells, CAFs may also play a role in the suppression of NK
cell activity in the TME. In this study, we used CAFs and normal
endometrial fibroblasts (NEFs), derived from endometrial cancer and
normal endometrial stroma, respectively. In the uterine
endometrium, endometrial stroma is enriched in fibroblasts and
surrounds the endometrial glandular epithelia, and these NEFs can
be transformed to CAFs in endometrial cancer. Therefore, the use of
endometrial cancer is suitable for comparison between CAFs and
NEFs. In this study, we investigated the inhibitory effect of CAFs
on NK cell-killing activity and the underlying mechanism.
Materials and methods
Patients and establishment of
fibroblasts
Tumor samples were obtained from the patients with
endometrial carcinoma, and normal endometrium were collected from
those without pathology in uterine endometrium, undergoing surgical
resection in our hospital. All women gave written informed consent
and the Research Ethics Committee of the University of Tokyo
approved all aspects of the study.
CAFs were isolated from the cancer tissues of
endometrial cancer while NEFs were from the normal endometrium with
no pathological abnormality. The tissues were minced and digested
in DMEM/F12 medium, (Gibco, Japan), supplemented with 100 IU/ml
penicillin, 100 μg/ml streptomycin, 1 mg/ml collagenase type I
(Wako, Tokyo, Japan), and 25 ng/ml DNase, Roche Diagnostics GmbH at
37°C for 60 min, filtered with 100 and 70 μm cell strainers, BD
Falcon, and centrifuged at 1,500 rpm for 5 min and washed with
D-PBS, Wako. They were resuspended in DMEM/F12 with 10% FBS, 100
IU/ml penicillin, 100 μg/ml streptomycin, and cultured at 37°C in
humidified 5% CO2 environment. Fibroblasts passaged for
2–8 passages were used before the experiments.
Isolation of NK cells and NK
cell/fibroblast co-cultures
NK cells were obtained from healthy volunteers after
Ficoll-Paque gradient and negative magnetic selection, using human
NK cell isolation kit from Miltenyi Biotec. NK cell purity was
>95% as evaluated by flow cytometry. They were cultured in RPMI
with 10% FBS, 100 IU/ml penicillin, 100 μg/ml streptomycin, at 37°C
in humidified 5% CO2 atmosphere, and stimulated 1 ng/ml
IL15 (R&D) for 48 h, before co-culture.
For co-culture experiments, CAFs or NEFs were seeded
at 5×104 in 24-well plates with 500 μl of medium and
cultured for 48 h. NK cells were added at 1×105 well, at
an NK/fibroblasts ratio of 1:1, with or without 1-μm pore of
cell-culture-insert. NK cells alone were also cultured in the
absence of fibroblasts. After 24-h incubation, NK cells were
harvested and analyzed. We confirmed that allogenic NK cells did
not kill these fibroblasts during co-culture (data not shown).
NK cell killing assay
K562 cells (obtained from ATCC; American Type
Culture Collection, VA, USA) were cultured in RPMI-1640
supplemented with 10% FBS containing 100 U/ml penicillin and 100
μg/ml streptomycin. K562 cells were resuspended at
1–2×107 cells/ml in 0.1% FBS/PBS and added CFSE, using
CFSE Cell Division Assay kit from Cayman chemical, to a final
concentration of 2.5 μM CFSE staining solution, incubated cells at
37°C for 30 min. After centrifugation of cells at 300 g for 10 min,
supernatant was discarded and they were resuspended into RPMI
medium and incubated at 37°C for 30 min. The cells were washed with
2% FBS/PBS three times, and 2×104 of stained K562 cells
in 200 μl medium were added to tubes. Harvested NK cells were
diluted at 1×106 cells/ml and prepare dilution series
2×105 cells per 200 μl. After incubation the tubes at
37°C for 4 h, 5 μl of PI (propidium iodide solution, Biolegend, San
Diego, CA, USA) was added for dead cell count. Percentage of
PI-positive dead K562 cells (% K562 PI-positive cell) was evaluated
by flow cytometry. In some experiments, allogenic NK cells were
incubated for 24 h with NEFs, CAFs or no fibroblasts followed by
exposure to target (K562) cells. NK cell killing activities were
indicated by ratio of % K562 PI positive cells under each condition
against that of NK cells only.
Immunocytochemistry
CAFs were grown on coverslips, then serum deprived
and fixed in 4% paraformaldehyde, permeabilized with 0.1% Triton
X-100, washed twice with PBS, and incubated for 60 min at room
temperature with an anti-α-SMA antibody (mouse clone 1A4, ab7817,
Abcam MA, USA), labeled with Alexa Flour 488 (Zenon). After
incubation, the slides were washed with PBS and fixed with 4%
paraformaldehyde. The cells were counterstained with
4′,6-diamidino-2-phenylindole dihydrochloride (DAPI).
Western blotting
CAFs and NEFs grown in chamber slides were fixed and
each of them were collected and total proteins were extracted.
Immunoblotted with anti-beta Actin antibody (rabbit polyclonal
antibody, Abcam), and α-SMA (mouse clone 1A4, ab7817, Abcam).
Flow cytometry
Cultured cells were harvested and incubated with
PerCP Cy5.5-conjugated CD90 (mouse clone Thy1, BioLegend),
FITC-conjugated Vimentin (mouse clone RV202, ab8978, Abcam),
PE-conjugated PVR (mouse clone SKII.4, BioLegend) or appropriate
isotype control at 4°C for 30 min, washed twice, and analyzed using
a BD FACS Calibur cytometer. The results were analyzed using Kaluza
software.
siRNA experiments
NEFs were transfected with 100 pmol of siRNA with
Lipofectamine RNAiMAX. The complexes of RNAi and 1.5 μl
Lipofectamine RNAiMAX in 140 μl Opti-MEM I medium without serum.
Then the complexes were added in the cultured cells at
4×104 in 24-well plates, and 500 μl of complete medium
without antibiotics was gently added in each well, and incubated
for 12 h at 37°C in a CO2 incubator to be ready for the
next assay of gene knockdown. To knock down PVR, siRNA by R&D
was used. The sequences were as follows: human PVR; sense,
5′-rCrArGrCUrAUUrCrGrGrArCUrCrCrArArATT; antisense,
5′-UUArGrGrArCUrCrCrGrArAUrArGrCUrGTT. The negative siRNA controls
were obtained from Life Technologies.
Statistical analysis
Data are presented as means ± SEM. Statistical
analyses were carried out using Student's t-test or Dunnett
analysis using JMP software. A value of p<0.05 was considered
significant. Asterisks indicate those comparisons with statistical
significance (p<0.05).
Results
Isolation of CAFs from endometrial
cancer
To investigate the difference in effect on NK cell
activity between NEFs and CAFs, we isolated fibroblasts derived
from normal endometrium (NEFs) and endometrial cancer tissue
(CAFs). Uterine endometrium is composed of many glands and abundant
stroma. The endometrial stroma is enriched in fibroblasts, and
surrounds the endometrial glandular epithelium. Endometrial cancer
can transform normal fibroblasts in the stroma into CAFs.
Therefore, the comparison between endometrial CAFs and NEFs was
thought to be suitable for the investigation of CAF activity.
Fibroblasts were isolated from cancer tissue or normal endometrium
by standard isolation methods and identified by immunostaining and
western blotting. Fibroblasts isolated from endometrial cancer
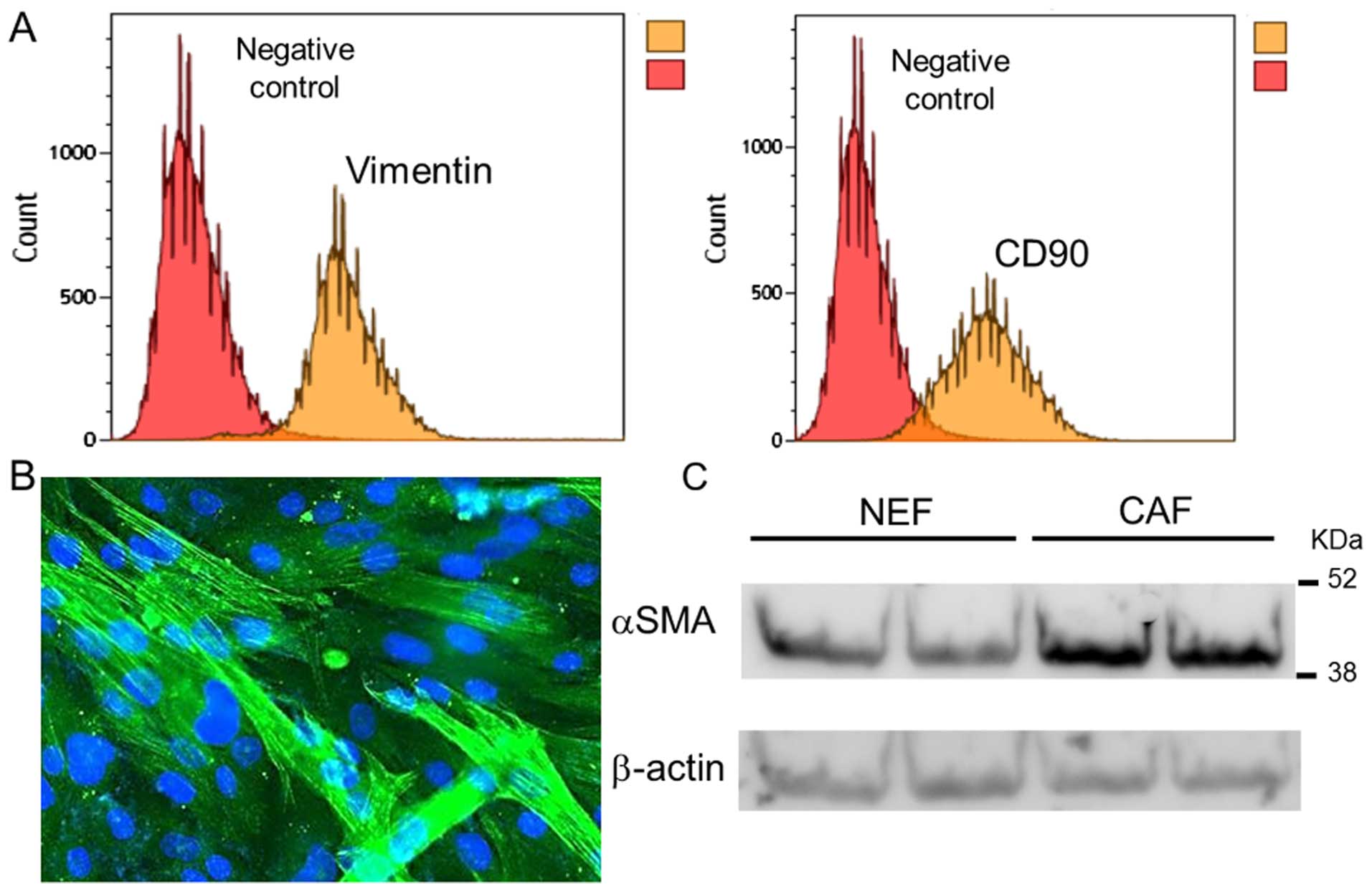
expressed the fibroblastic markers vimentin and CD90 (Fig. 1A). To distinguish the
cancer-derived fibroblasts from the NEFs, presence of α-SMA was
assessed by immunocytochemistry (Fig.
1B), and expression level of α-SMA compared with NEFs was
assessed by western blotting (Fig.
1C). α-SMA localizes to the cytoplasm in cancer-derived
fibroblasts (Fig. 1B). Western
blotting demonstrated increased expression levels of α-SMA in two
cancer-derived fibroblast samples compared with two NEF samples.
Therefore, the cancer-derived fibroblasts isolated in this study
possessed typical CAF characteristics, and were confirmed to be
CAFs.
Suppression of NK cell-killing activity
by CAFs
We assessed the effect of CAFs on the killing
activity of NK cells to investigate the mechanism of CAF-mediated
immune evasion. Assessment of NK cell activity is often performed
by measuring allogenic NK cell killing activity against K562 cells
(14). Using this NK cell-killing
assay, we first examined whether CAFs isolated from endometrial
cancer suppressed the killing activity of allogenic NK cells
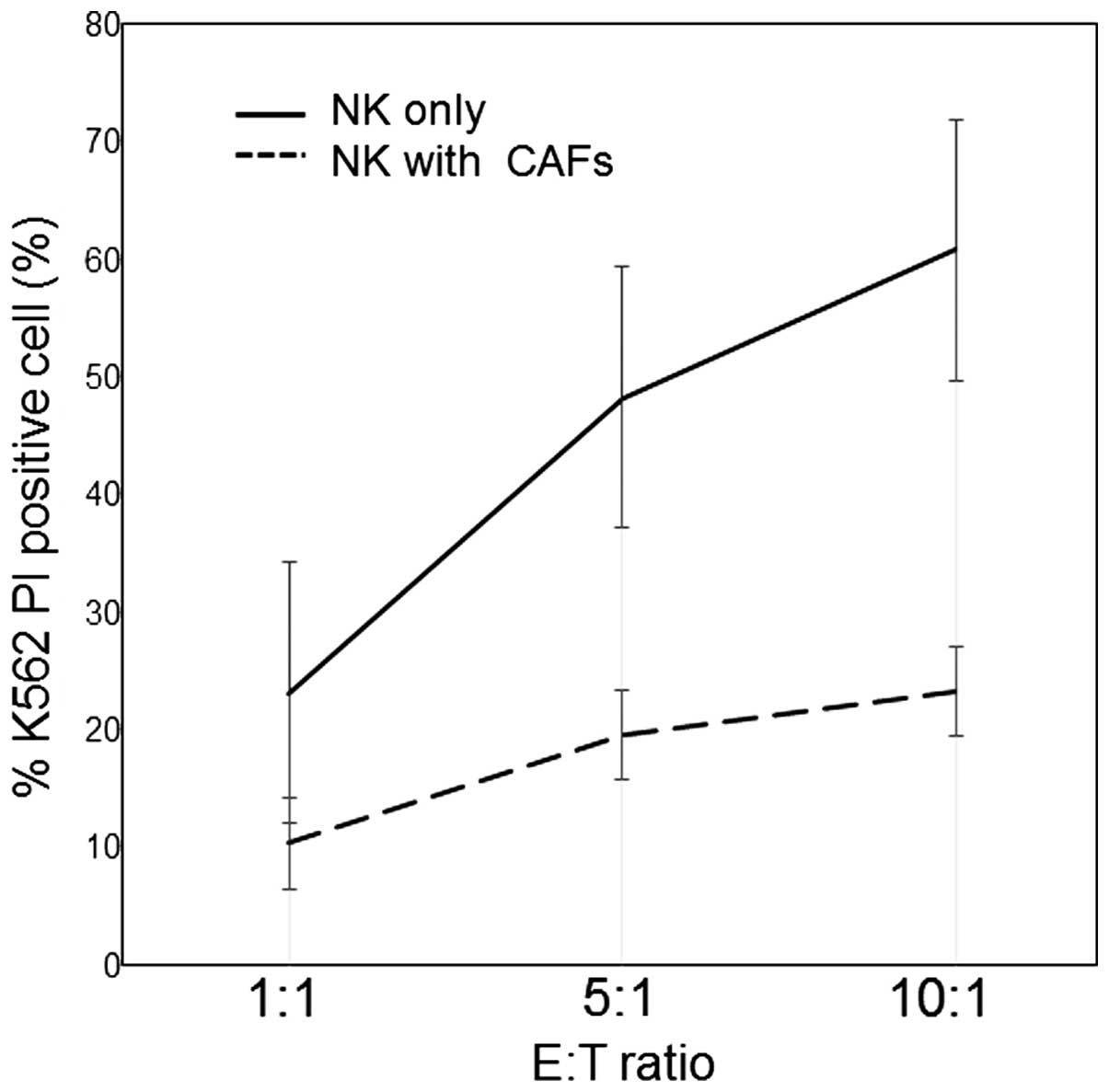
(Fig. 2). NK cells co-cultured
with CAFs were assessed for killing activity by changing the E:T
(NK cell: K562 cell) ratio. The proportion of dead K562 cells
increased in an E:T ratio-dependent manner in the presence of NK
cells alone, or with NK cells co-cultured with CAFs, indicating
that the target (K562) cells were killed by allogenic effector (NK)
cells (Fig. 2). The killing
activity of NK cells co-cultured with CAFs was significantly
reduced, to less than one third the level of NK cells alone
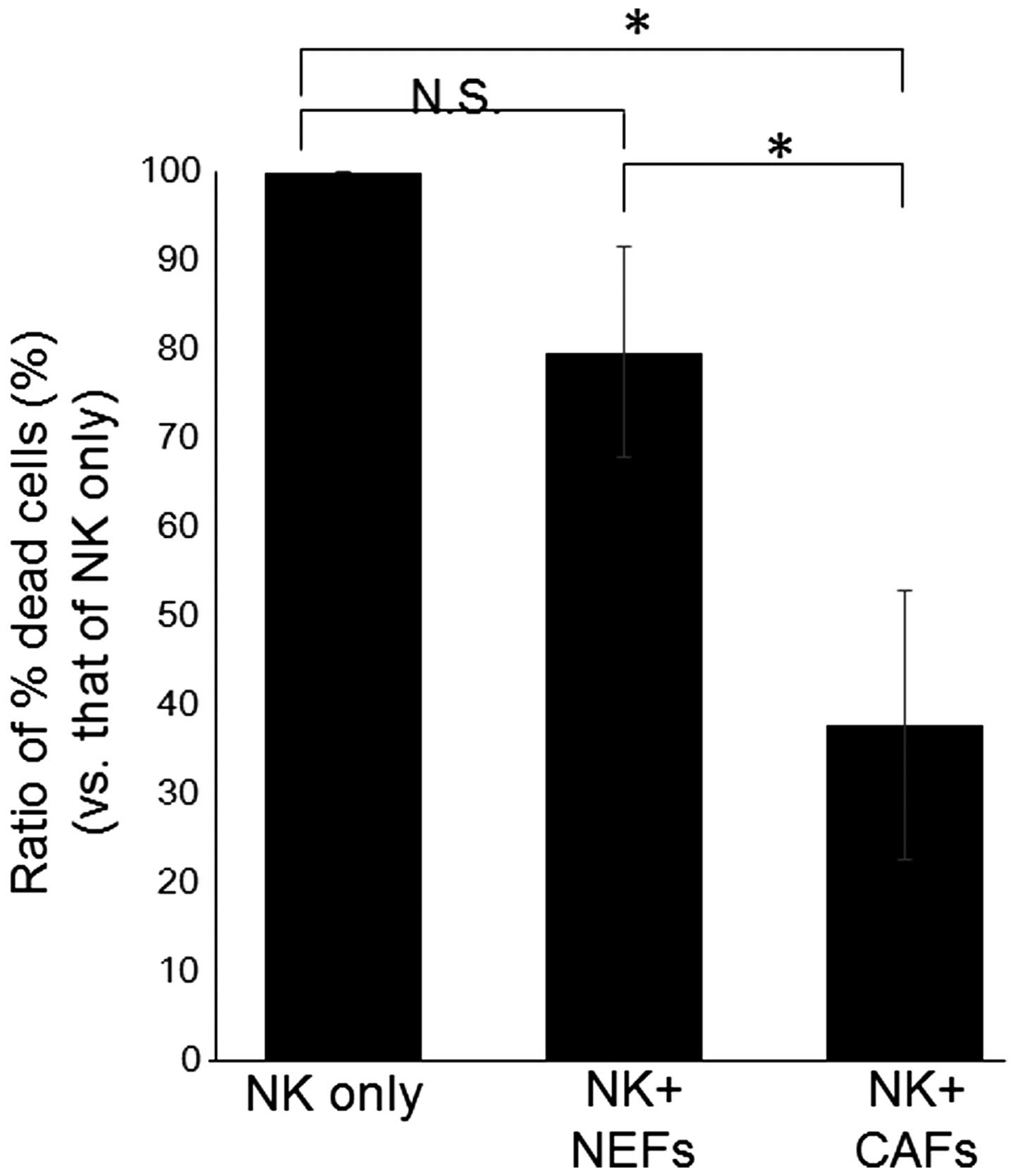
(Fig. 2). Next, NK cell activity
was assessed when co-cultured with either NEFs or CAFs (Fig. 3). Allogenic NK cells were incubated
for 24 hours with NEFs, CAFs, or no fibroblasts, and then exposed
to K562 cells. NK cell-killing activity was determined by
calculating the ratio of PI-positive K562 cells in the presence of
NK cells co-cultured with fibroblasts versus NK cells that had been
cultured alone. There was no significant difference in killing
activity observed between NK cells only and NK cells co-cultured
with NEFs. Again, the killing activity of NK cells co-cultured with
CAFs was significantly decreased, to approximately one-third of the
activity of NK cells cultured with no fibroblasts. However, NK cell
activity was significantly decreased when co-cultured with CAFs,
compared with NEF's (Fig. 3).
The CAF-induced decrease in NK cell
activity was not rescued by indoleamine 2,3-dioxygenase
inhibitor
Some studies have shown that malignant cells
suppress NK cell activity via production of IDO (15,16).
IDO expression was barely detected in non-stimulated CAFs, whereas
it was clearly detected in CAFs co-cultured with NK cells (data not
shown). Previous reports have demonstrated that IDO expression in
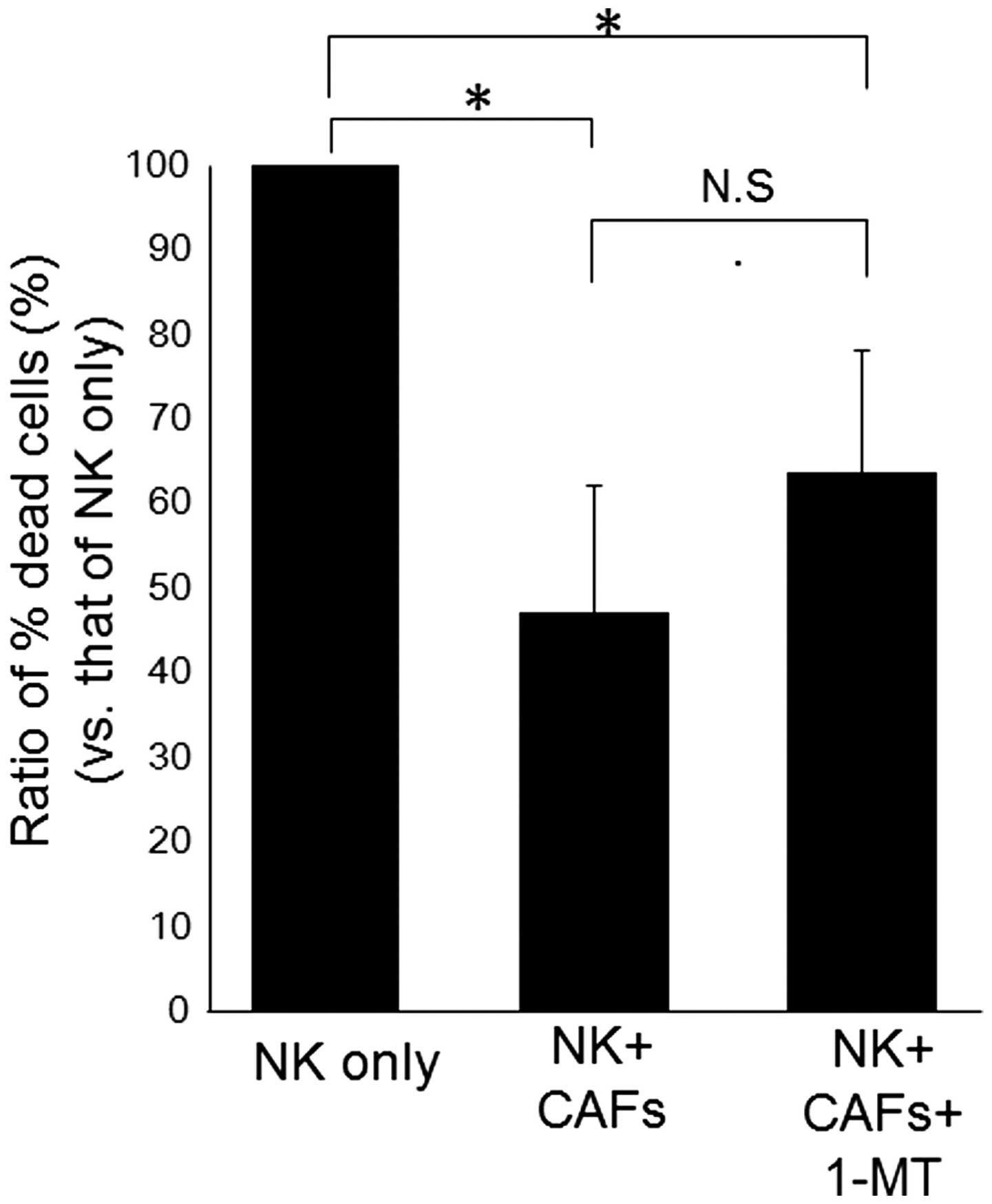
CAFs is likely caused by NK cell-derived IFNγ (17). Therefore, we inhibited IDO activity
using the IDO inhibitor 1-MT, to investigate whether IDO was
involved in the suppression of NK cell activity by CAFs. NK cells
and CAFs were co-cultured in the presence or absence of 1-MT,
followed by assessment of NK cell-killing activities (Fig. 4). While NK cell activity was
decreased by co-culture with CAFs, this suppression was not rescued
by 1-MT treatment.
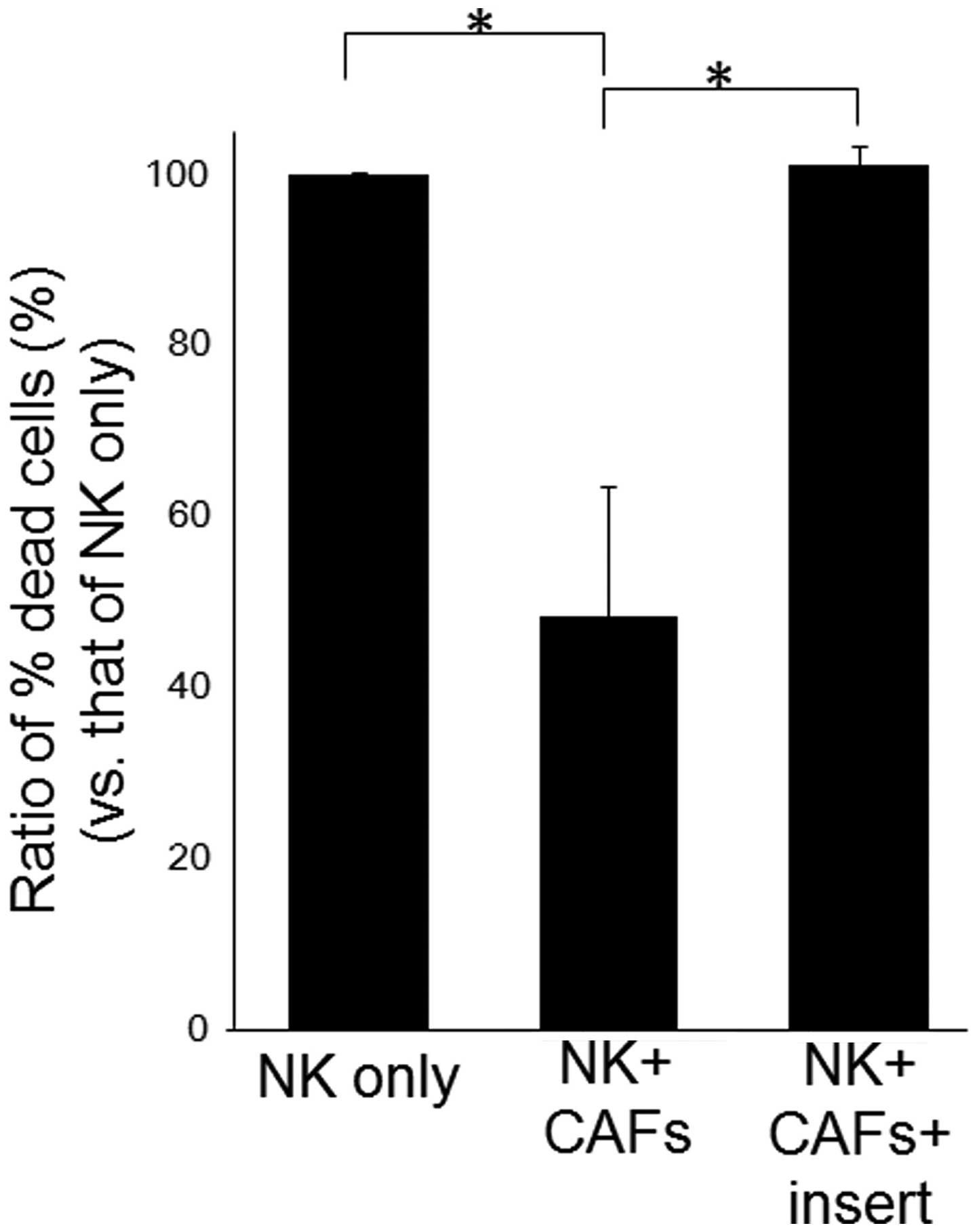
Cell-to-cell interaction was critical for
the CAF-induced decrease in NK cell activity
To examine whether direct cell-to-cell interaction
between NK cells and CAFs was required for the suppression of NK
cell activity, NK cells and CAFs were cultured in a chamber with an
inserted membrane separating these cells (Fig. 5). The suppression of NK cell
activity by CAFs was completely abolished by the physical blockage
of cell-to-cell interaction using the inserted membrane. These data
strongly suggested that CAFs suppressed NK cell activity via
cell-to-cell interaction rather than IDO, or other cytokine,
production.
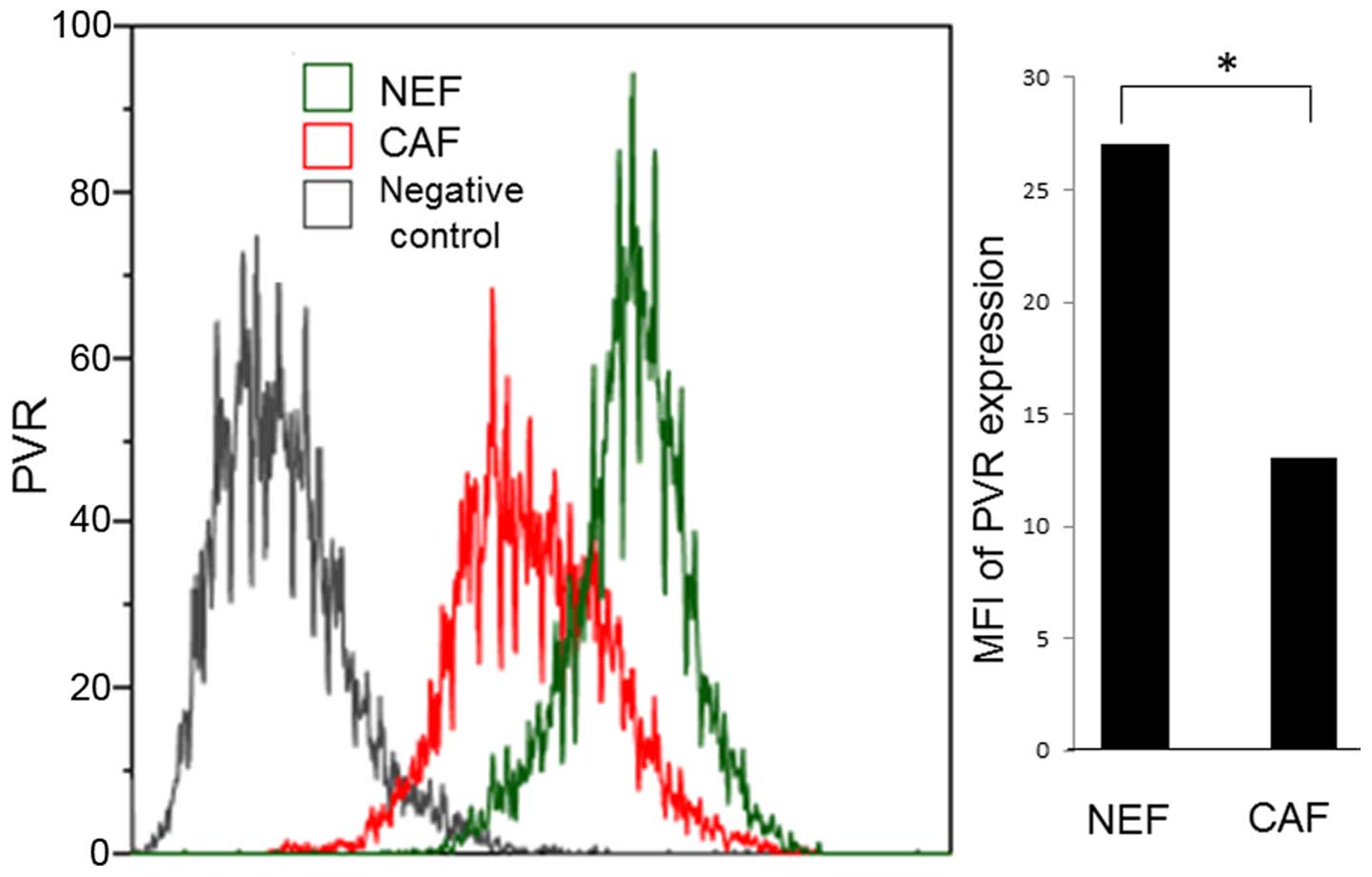
Cell-surface expression of PVR is
decreased in CAFs compared with NEFs
We next focused on the cell-surface ligands
expressed on CAFs that interact with activating NK cell receptors.
Several ligands of paired or activating NK receptors have been
previously demonstrated to be expressed on the cell surface of
target cells, including malignant cells (18). However, few reports have
investigated the cell-surface expression of NK receptor ligands on
CAFs. We investigated the expression of several NK receptor ligands
in CAFs. Due to the difference between CAFs and NEFs in their
ability to induce NK cell activity, we hypothesized that the
expression of NK receptor ligands may differ between CAFs and NEFs.
Expression of poliovirus receptor (PVR/CD155), a ligand of paired
NK receptors DNAM-1 and TIGIT, was observed to differ between CAFs
and NEFs (Fig. 6). Flow cytometry
analysis revealed that the cell-surface expression of PVR was
decreased in CAFs compared with NEFs. This suggested that the
downregulation of PVR in CAFs may mediate the suppression of DNAM-1
signaling.
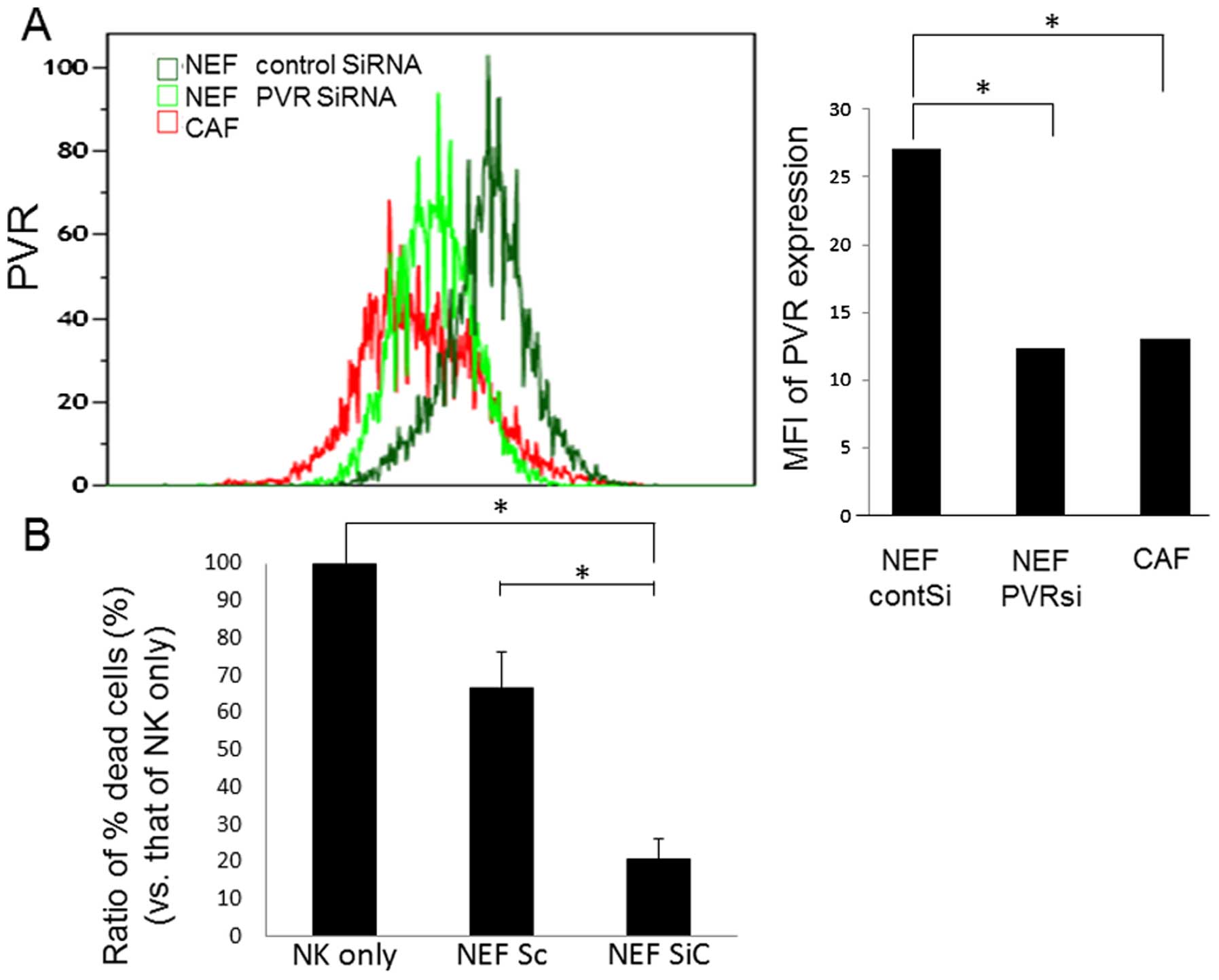
PVR knockdown downregulates NK cell
activity
To confirm whether PVR downregulation results in the
decrease of NK cell-killing activity, PVR expression in NEFs was
knocked down using siRNA against PVR (PVRsi) (Fig. 7A). PVRsi-transfected NEFs showed
decreased cell-surface PVR expression when compared with control
siRNA-transfected NEFs. The median flow cytometry index (MFI) of
PVR expression in PVR knockdown cells was similar to that of CAFs
(Fig. 7A). Interestingly, NK
cell-killing activity of NK cells co-cultured with
PVRsi-transfected NEFs was decreased to approximately one third of
that of control si-transfected NEFs (Fig. 7B). The decreases in both PVR
expression and effect on NK cell activity were approximately
equivalent to those of CAFs. These data suggested that the level of
PVR cell-surface expression in CAFs was critical for the killing
activity of NK cells.
Discussion
We demonstrated that CAFs showed increased
suppression of NK cell-killing activity compared with NEFs, due to
decreased PVR cell surface expression, a ligand of an NK activating
receptor. In this study, human CAFs and NEFs were isolated from the
stroma of endometrial cancer and normal endometrium, respectively,
and their interactions with NK cells were compared. Uterine
endometrium is composed of many glands and abundant stroma. The
endometrial stroma is enriched in fibroblasts, and surrounds the
endometrial glandular epithelium. Endometrial cancer cells can
transform normal fibroblasts in the stroma into CAFs. In the cancer
microenvironment, CAFs acquire cancer-specific characteristics in
addition to their fibroblastic background. Therefore, we believe
that the comparison between endometrial CAFs and normal endometrial
stromal fibroblasts was suitable for investigation of CAF
characteristics. We observed an increased α-SMA expression level in
CAFs compared with NEFs, in confirmation with previous studies
demonstrating the expressing of α-SMA and vimentin in active CAFs
(19). CD90 is a fibroblastic
marker that is also expressed in CAFs of a variety of cancers, such
as lung, prostate, pancreas, and breast cancers (20–22).
CAFs isolated from endometrial cancer in this study were
fibroblasts showing typical CAF characteristics.
Many previous studies have demonstrated the
suppression of NK cell activity by cancer cells by measuring the
killing activity of allogenic NK cell against K562 cells (23). Some studies have shown that
malignant cells suppress NK cell activity via the production of
IDO. In this study, the suppression of NK cell-killing activity by
CAFs was not rescued by exposure to 1-MT (IDO inhibitor),
suggesting suppression of NK cell activity is not due to IDO
production by CAFs. Prevention of cell-to-cell interaction between
NK cells and CAFs completely abolished the suppression of NK cell
activity by CAFs, indicating direct interaction between NK cells
and CAFs was critical for the CAF-induced suppression of NK cell
activity. Several studies have demonstrated that exosomes derived
from CAFs may interact with NK cells to suppress NK activity.
However, exosomes should be able to penetrate the transwell
membrane used in this study, suggesting that the suppression of NK
cell activity was not dependent on CAF exosome exposure.
Our data demonstrated cell-surface expression of PVR
was reduced in CAFs, compared with NEFs. The MFI of PVR in CAFs was
half that in NEFs. NK activating receptors include NKp30, NKp44,
NKp46, NKG2D, DNAM-1, and LFA-1 (8). Among the ligands of these receptors,
PVR, a ligand of DNAM-1, was observed in this study to be
ubiquitously expressed on the cell-surface of CAFs. It is known
that DNAM-1 activates NK cell activity. We observed that co-culture
of NK cells and CAFs did not alter DNAM-1 expression in NK cells
(data not shown). In contrast, PVR interacts with NK paired
receptors, DNAM-1 (activating) and TIGIT (inhibiting).
Interestingly, some studies demonstrated that PVR interacts with
higher affinity to TIGIT than DNAM-1. Our data suggested that the
reduced expression of PVR in CAFs resulted in decreased interaction
with DNAM-1, leading to suppression of NK killing activity.
Furthermore, it is possible that the high affinity of PVR to TIGIT
allowed even low expression of cell-surface PVR in CAFs to bind to
TIGIT, thus inhibiting NK cell activity. We also demonstrated that
the NK killing activity of PVR-knockdown NEFs decreased to levels
comparable with that of CAFs. The data suggested that the reduction
of PVR expression in CAFs is critical for CAF-induced suppression
of NK cell activity.
Many studies have reported PVR involvement in NK
cell-associated immune-evasion by malignant cells, and that low
expression of PVR is associated with poor prognosis (24). However, this study is the first to
demonstrate that the suppression of NK killing activity by CAFs was
mediated by downregulation of cell-surface PVR in CAFs; however,
the mechanism of PVR downregulation remains to be clarified.
Recently, a study by Gong et al reported that ER stress
induces the downregulation of PVR in hepatoma cells via the ATF6
and IRE1a pathways (25). ER
stress, such as that induced by hypoxia in the TME, may result in
downregulation of PVR in CAFs; therefore a low-level of PVR
expression in fibroblasts may be a characteristic of CAFs. Further
study is warranted to bring to light the mechanism by which CAFs
acquire such features in the TME.
CAFs may assist malignant cells in a similar fashion
to maintain the immunosuppressive microenvironment of the tumor.
Additionally, immune evasion may be influenced by both CAFs and
malignant cells with regard to NK cell-mediated killing activity.
Therefore, soluble PVR may be used as a potential agent to activate
NK cell activity in the TME. These data may provide a novel
strategy for inhibiting the immune evasion system in the TME.
In conclusion, this is the first report to
demonstrate that CAF-mediated suppression of NK killing activity is
due to downregulation of PVR cell-surface expression in CAFs. We
discovered that CAFs suppressed NK cells function via a
receptor-ligand interaction, aiding cancer progression. Soluble PVR
may be used as a potential agent to activate NK cell activity in
the TME. These data may provide a novel strategy for inhibiting the
immune evasion system in the TME.
Acknowledgements
The authors would like to thank Dr Terufumi Yokoyama
for expert advice on experimental methodologies. This study was
supported by a grant-in-aid from the Ministry of Health, Labour and
Welfare of Japan (KK) and by a cancer research grant from the
Ministry of Education, Culture, Sports, Science and Technology
(K.K., K.A. and K.T.) of Japan.
Abbreviations:
|
CAFs
|
cancer-associated fibroblasts
|
|
TME
|
tumor microenvironment
|
|
PVR
|
poliovirus receptor
|
|
NEFs
|
normal endometrial fibroblasts
|
|
IDO
|
indoleamine 2,3-dioxygenase
|
References
|
1
|
Karagiannis GS, Poutahidis T, Erdman SE,
Kirsch R, Riddell RH and Diamandis EP: Cancer-associated
fibroblasts drive the progression of metastasis through both
paracrine and mechanical pressure on cancer tissue. Mol Cancer Res.
10:1403–1418. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lorusso G and Rüegg C: The tumor
microenvironment and its contribution to tumor evolution toward
metastasis. Histochem Cell Biol. 130:1091–1103. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Joyce JA and Pollard JW:
Microenvironmental regulation of metastasis. Nat Rev Cancer.
9:239–252. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Franco OE, Shaw AK, Strand DW and Hayward
SW: Cancer associated fibroblasts in cancer pathogenesis. Semin
Cell Dev Biol. 21:33–39. 2010. View Article : Google Scholar :
|
|
5
|
Kalluri R and Zeisberg M: Fibroblasts in
cancer. Nat Rev Cancer. 6:392–401. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sanz-Moreno V, Gaggioli C, Yeo M,
Albrengues J, Wallberg F, Viros A, Hooper S, Mitter R, Féral CC,
Cook M, et al: ROCK and JAK1 signaling cooperate to control
actomyosin contractility in tumor cells and stroma. Cancer Cell.
20:229–245. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gaggioli C, Hooper S, Hidalgo-Carcedo C,
Grosse R, Marshall JF, Harrington K and Sahai E: Fibroblast-led
collective invasion of carcinoma cells with differing roles for
RhoGTPases in leading and following cells. Nat Cell Biol.
9:1392–1400. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chan CJ, Smyth MJ and Martinet L:
Molecular mechanisms of natural killer cell activation in response
to cellular stress. Cell Death Differ. 21:5–14. 2014. View Article : Google Scholar
|
|
9
|
Calon A, Tauriello DV and Batlle E:
TGF-beta in CAF-mediated tumor growth and metastasis. Semin Cancer
Biol. 25:15–22. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Salaroglio IC, Campia I, Kopecka J,
Gazzano E, Orecchia S, Ghigo D and Riganti C: Zoledronic acid
overcomes chemoresistance and immunosuppression of malignant
mesothelioma. Oncotarget. 6:1128–1142. 2015. View Article : Google Scholar :
|
|
11
|
Tahara-Hanaoka S, Shibuya K, Onoda Y,
Zhang H, Yamazaki S, Miyamoto A, Honda S, Lanier LL and Shibuya A:
Functional characterization of DNAM-1 (CD226) interaction with its
ligands PVR (CD155) and nectin-2 (PRR-2/CD112). Int Immunol.
16:533–538. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bottino C, Castriconi R, Pende D, Rivera
P, Nanni M, Carnemolla B, Cantoni C, Grassi J, Marcenaro S, Reymond
N, et al: Identification of PVR (CD155) and Nectin-2 (CD112) as
cell surface ligands for the human DNAM-1 (CD226) activating
molecule. J Exp Med. 198:557–567. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Atsumi S, Matsumine A, Toyoda H, Niimi R,
Iino T and Sudo A: Prognostic significance of CD155 mRNA expression
in soft tissue sarcomas. Oncol Lett. 5:1771–1776. 2013.PubMed/NCBI
|
|
14
|
Tanizaki Y, Kobayashi A, Toujima S, Shiro
M, Mizoguchi M, Mabuchi Y, Yagi S, Minami S, Takikawa O and Ino K:
Indoleamine 2,3-dioxygenase promotes peritoneal metastasis of
ovarian cancer by inducing an immunosuppressive environment. Cancer
Sci. 105:966–973. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kasatori N, Ishikawa F, Ueyama M and
Urayama T: A differential assay of NK-cell-mediated cytotoxicity in
K562 cells revealing three sequential membrane impairment steps
using three-color flow-cytometry. J Immunol Methods. 307:41–53.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yoshida N, Ino K, Ishida Y, Kajiyama H,
Yamamoto E, Shibata K, Terauchi M, Nawa A, Akimoto H, Takikawa O,
et al: Overexpression of indoleamine 2,3-dioxygenase in human
endometrial carcinoma cells induces rapid tumor growth in a mouse
xenograft model. Clin Cancer Res. 14:7251–7259. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Löb S, Königsrainer A, Rammensee HG, Opelz
G and Terness P: Inhibitors of indoleamine-2,3-dioxygenase for
cancer therapy: Can we see the wood for the trees? Nat Rev Cancer.
9:445–452. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pegram HJ, Andrews DM, Smyth MJ, Darcy PK
and Kershaw MH: Activating and inhibitory receptors of natural
killer cells. Immunol Cell Biol. 89:216–224. 2011. View Article : Google Scholar
|
|
19
|
Augsten M: Cancer-associated fibroblasts
as another polarized cell type of the tumor microenvironment. Front
Oncol. 4:622014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yan X, Luo H, Zhou X, Zhu B, Wang Y and
Bian X: Identification of CD90 as a marker for lung cancer stem
cells in A549 and H446 cell lines. Oncol Rep. 30:2733–2740.
2013.PubMed/NCBI
|
|
21
|
True LD, Zhang H, Ye M, Huang CY, Nelson
PS, von Haller PD, Tjoelker LW, Kim JS, Qian WJ, Smith RD, et al:
CD90/THY1 is overexpressed in prostate cancer-associated
fibroblasts and could serve as a cancer biomarker. Mod Pathol.
23:1346–1356. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lu H, Clauser KR, Tam WL, Fröse J, Ye X,
Eaton EN, Reinhardt F, Donnenberg VS, Bhargava R, Carr SA, et al: A
breast cancer stem cell niche supported by juxtacrine signalling
from monocytes and macrophages. Nat Cell Biol. 16:1105–1117. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Alter G, Malenfant JM and Altfeld M:
CD107a as a functional marker for the identification of natural
killer cell activity. J Immunol Methods. 294:15–22. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Qu P, Huang X, Zhou X, Lü Z, Liu F, Shi Z,
Lü L, Wu Y and Chen Y: Loss of CD155 expression predicts poor
prognosis in hepatocellular carcinoma. Histopathology. 66:706–714.
2015. View Article : Google Scholar
|
|
25
|
Gong J, Fang L, Liu R, Wang Y, Xing J,
Chen Y, Zhuang R, Zhang Y, Zhang C, Yang A, et al: UPR decreases
CD226 ligand CD155 expression and sensitivity to NK cell-mediated
cytotoxicity in hepatoma cells. Eur J Immunol. 44:3758–3767. 2014.
View Article : Google Scholar : PubMed/NCBI
|