Introduction
Hypopharyngeal squamous cell carcinoma (HSCC), which
accounts for 3–5% of all head and neck cancers, is the most
aggressive head and neck cancer and has very poor prognosis
(1). More than three-quarters of
patients afflicted with this cancer are at an advanced stage at the
time of diagnosis (1). In the
course of the disease, because of the early high rates of cervical
lymph nodes metastasis (2), the
overall survival is relatively poor, with 5-year survival rates
<40%, despite aggressive surgical and adjuvant treatment
(3,4). Smoking cigarettes and drinking
alcohol are the main risk factors for HSCC (5). However, the molecules involved in
tumorigenesis of HSCC are not clearly revealed. Therefore, it is
overwhelmingly urgent to discover new biomarkers to advance
stratification and development of novel therapeutic targets for
HSCC.
The phosphoprotein stathmin1 (STMN1), also known as
oncoprotein 18 (OP18), is a 17-kDa cytoplasmic protein (6). Reports have shown that STMN1 could
regulate microtubule dynamics by participating in microtubule
catastrophe and/or in the sequestering of α/β-tubulin heterodimers,
thereby impacting cell cycle progression, proliferation, motility
and survival (7). Previous studies
have also shown that STMN1 is overexpressed across many different
human cancers, including some hematological malignancy, digestive
system tumors, genitourinary neoplasms, lung cancer, mesothelioma
and some head and neck cancers such as oral squamous-cell
carcinoma, laryngeal squamous-cell carcinoma and primary
nasopharyngeal carcinoma (8).
Furthermore, abundant evidence has shown that STMN1 knockdown could
significantly influence cell cycle progression, apoptosis,
metastasis, and chemoresistance of tumor cells (8). Our previous study confirmed that
STMN1 is significant highly expressed in non-melanoma skin cancers
(NMSCs), and its expression is closely related to the
proliferation, migration, invasion and apoptosis of cSCC cells
(9). However, the effects of STMN1
on human HSCC remain unclear.
In this study, we investigated the expression
pattern of STMN1 in HSCC tissues by immunohistochemistry and its
correlation to clinicopathological characteristics. In addition,
the application of STMN1, a prognostic factor, was assessed.
Furthermore, we examined the biological function of STMN1 in HSCC
FaDu cells. The data revealed that elevated STMN1 could promote
HSCC proliferation and migration and enhance the EMT process, and
it may be used as a prognostic biomarker and therapeutic target of
HSCC.
Materials and methods
Tissue specimens
A total of 51 HSCC biopsy samples with complete
clinical and pathological data were obtained from patients who
underwent operations from 2009 to 2015 in the Affiliated Hospital
of Nantong University. Sixteen non-cancer HSCC tissue samples were
recruited as controls. Prior to surgery, none of the patients
received adjuvant chemotherapy or radiotherapy. The pathological
analysis was carried out according to the TNM system of the
American Joint Committee on Cancer (AJCC 2010). All the detailed
information of these 51 HSCC patients and clinicopathological
parameters in this series are shown in Table I. Furthermore, 7 HSCC and
corresponding 7 para-carcinoma tissues were collected to confirm
the expression of STMN1 protein. This study was approved by the
Ethics Committee of the Affiliated Hospital of Nantong University
and all the participants gave their informed consent.
 | Table IThe clinicopathological
characteristics and IHC staining for STMN1 of the HSCC
patients. |
Table I
The clinicopathological
characteristics and IHC staining for STMN1 of the HSCC
patients.
| N | STMN1 IHC
score | P-value |
|---|
| Age |
| <60 years | 21 | 7.24±3.11 | 0.430 |
| ≥60 years | 30 | 6.46±3.61 | |
| Gender |
| Male | 47 | 6.79±3.36 | 0.824 |
| Female | 4 | 6.33±4.93 | |
| Tobacco
smoking |
| None or
limited | 20 | 6.02±3.46 | 0.214 |
| Excessive | 31 | 7.25±3.35 | |
| Alcohol
consumption |
| None or
limited | 14 | 5.85±3.94 | 0.242 |
| Excessive | 37 | 7.11±3.18 | |
|
Differentiation |
| High
differentiation | 22 | 4.70±2.01 | <0.001** |
| Moderate/poor
differentiation | 29 | 8.33±3.45 | |
| Tumor size
(cm) |
| ≤2 | 24 | 5.22±2.62 | 0.001** |
| >2 | 27 | 8.14±3.48 | |
| Clinical stage |
| I–II | 19 | 4.59±2.23 | <0.001** |
| III–IV | 32 | 8.06±3.35 | |
| Lymph node
metastasis |
| Negative | 25 | 5.50±3.13 | 0.008** |
| Positive | 26 | 7.98±3.27 | |
| Treatment |
| Surgery only | 20 | 4.98±2.89 | 0.010* |
| Surgery and
radiation | 17 | 7.85±2.58 | |
| Surgery,
radiation, chemotherapy | 11 | 7.40±4.16 | |
| Other
treatment | 3 | 10.13±3.23 | |
Immunohistochemical staining
Immunohistochemical staining was performed on
4-µm paraffin tissue sections mounted on slides and dried at
60°C for 8 h. The slides were deparaffinized in xylene and
dehydrated conventionally, then pressure-cooked in sodium citrate
buffer (Sangon Biotech, Shanghai, China) (pH 6.0) to facilitate
antigen retrieval. After natural cooling, endogenous peroxidase was
blocked with 3% hydrogen peroxide (ZSGB-BIO, Beijing, China).
Non-specific bindings were blocked by treating slides with normal
goat serum (ZSGB-BIO) for 40 min, the sections were subsequently
incubated with rabbit anti-STMN1 polyclonal antibody (Abcam,
Cambridge, MA, USA, dilution 1:250) or rabbit anti-Ki-67 polyclonal
antibody (dilution 1:100, sc-15402; Santa Cruz Biotechnology, Inc.
Santa Cruz, CA, USA), overnight at 4°C. After being washed with
PBS, the sections were incubated for 30 min with the 2-step plus
poly-HRP anti-mouse/rabbit IgG detection system (ZSGB-BIO). DAB
detection kit (enhanced polymer) (ZSGB-BIO) was used for 3 min to
show immunolabeling, resulting in a brown precipitate. Following
the above, sections were counterstained with hematoxylin. Positive
and negative controls were performed in parallel.
The immunostaining results were blindly evaluated by
two experienced pathologists without knowledge of the
clinicopathologic outcomes of these patients. Immunostaining for
STMN1 was localized in the cytoplasm and, partly, in the nucleus,
and immunostaining for Ki-67 was limited to the nucleus. The
proportion of epithelial cells positively-stained was divided into
two groups: category A scores from 1 to 4 [(A = 0 (0–4%); 1
(5–25%); 2 (26–50%); 3 (51–75%); 4 (76–100%)]. Intensity of
staining was category B, scores 0 (negative); 1 (weak); 2
(moderate); 3 (strong). For the IHC-score assessment, all fields
were observed at ×400 magnification. A final score was calculated
by multiplying A by B (minimum 0, maximum 12). Moreover, the
average score of each case was obtained. Total scores were devided
into 4 grades: negative, 0–3; weak, 3–6 (contain 3, 6); moderate,
6–9; strong, 9–12 (contain 9).
Cell culture and siRNA transfection
Human HSCC cell line, FaDu cells, was purchased from
ATCC, Shanghai, China. FaDu cells were cultured in Dulbecco's
modified Eagle's medium supplemented with 10% fetal bovine serum at
37°C in a humid atmosphere containing 5% CO2. To silence
the expression of STMN1 (accession no. NM 203401 form NCBI
GenBank), FaDu cells were transfected with four siRNAs which were
designed and obtained from Biomics Biotechnologies Co., Ltd.
(Nantong, China). The sequence alignment (BLAST) was carried out to
ensure no homology with other genes in human. Furthermore, the
negative control siRNA (si-NC) was also designed. The targeted
sequences are shown in Table
II.
| Table IISequences of siRNAs targeting
STMN1. |
Table II
Sequences of siRNAs targeting
STMN1.
| SiRNAs | Sequence
(5′-3′) |
|---|
| STMN1-si1 | Sense:
GCUUCAGAAGGCAAUAGAAdTdT |
| Antisense:
UUCUAUUGCCUUCUGAAGCdTdT |
| STMN1-si2 | Sense:
GGAGGAAAUUCAGAAGAAAdTdT |
| Antisense:
UUUCUUCUGAAUUUCCUCCdTdT |
| STMN1-si3 | Sense:
GCACGAGAAAGAAGUGCUUdTdT |
| Antisense:
AAGCACUUCUUUCUCGUGCdTdT |
| STMN1-si4 | Sense:
GAACAACAACUUCAGUAAAdTdT |
| Antisense:
UUUACUGAAGUUGUUGUUCdTdT |
| Si-NC | Sense:
UUCUCCGAACGUGUCACGUdTdT |
| Antisense:
ACGUGACACGUUCGGAGAAdTdT |
Immunofluorescence
FaDu cells were cultured in 24-well chamber slides,
washed three times with cold 1X PBS, fixed with 4% paraformaldehyde
for 1 h at 4°C, washed three times in PBS, permeabilized with 0.1%
Triton X-100 for 10 min (except for staining the cell
membrane-bound E-cadherin), and blocked with Immunol Staining
Blocking Buffer (Beyotime, China) for 1 h at room temperature.
Cells were then incubated overnight at 4°C with anti-STMN1 (Abcam,
1:100 dilution), anti-E-cadherin (Ruiying Biological, Suzhou,
China, 1:100 dilution) or anti-vimentin (Ruiying Biological, 1:100
dilution) primary antibody and subsequently with Alexa
Fluor-conjugated secondary antibodies (1:1,000; Invitrogen Life
Technologies, Carlsbad, CA, USA) and Hoechst (Sigma-Aldrich Co.,
St. Louis, MO, USA) for 2 h at room temperature. The
immunofluorescence images were acquired with a fluorescence
microscope.
Real-time quantity PCR (RT-qPCR)
The mRNA expression of STMN1, E-cadherin, vimentin
in FaDu cells after STMN1-siRNA transfection were quantified by
real-time RT-PCR. According to the manufacturer's instructions,
total RNA was extracted using a TRIzol® reagent (Vazyme,
USA) and 2 µg RNA was reverse-transcribed into cDNA samples
by using a Transcriptor First Strand cDNA Synthesis kit (Roche,
Germany). Then AceQ® qPCR SYBR Green Master Mix kit
(Vazyme) was used for RT-qPCR. The primers used were obtained from
Biomics Biotechnologies Co. Ltd. and are shown in Table III. GAPDH was used as the
reference gene. The PCR conditions consisted of 10 min at 95°C 1
cycle, 15 sec at 95°C, 30 sec at 60°C and 30 sec at 72°C 40 cycles.
The experiment was performed in triplicate. The 2−ΔΔCT
method was implemented to analyze the results (10).
| Table IIIThe primers. |
Table III
The primers.
| The molecule | Sequence
(5′-3′) |
|---|
| STMN1 | Forward:
TGGAGAAGCGTGCCTCAG |
| Reverse:
TTCATGGGACTTGCGTCTTTC |
| E-cadherin | Forward:
CCAGGAACCTCTGTGATGGA |
| Reverse:
TTTTGTCAGGGAGCTCAGGA |
| Vimentin | Forward:
AAATGGCTCGTCACCTTCGT |
| Reverse:
CAGCTTCCTGTAGGTGGCAA |
| GAPDH | Forward:
GAAGGTGAAGGTCGGAGTC |
| Reverse:
GAAGATGGTGATGGGATTTC |
Protein extraction western blot
analysis
The cells or tissue samples were lysed in lysis
buffer (PMSF:RIPA = 1:100, Beyotime) and then centrifuged at 10,000
rpm for 10 min at 4°C. Protein concentrations were measured with
BCA protein assay kit (Pierce, USA). The supernatant was diluted in
5X SDS-polyacrylamide gel electrophoresis (SDS-PAGE) loading buffer
[50 mM Tris-HCl (pH 6.8), 10% (w/v) SDS, 0.5% (w/v) bromophenol
blue, 50% (w/v) glycerol, 5% (w/v) β-mercaptoethanol] and boiled.
Protein samples (20 µg) were separated by 10% (E-cadherin),
12% (vimentin, β-actin) and 15% (STMN1) polyacrylamide gels and
electrophoresed, then electroblotted onto polyvinylidine difluoride
filter (PVDF) membranes (Millipore, USA), followed by blocking with
5% skim milk in TBST (20 mM Tris, 150 mM NaCl, 0.05% Tween-20, pH
7.5) for 2 h at room temperature. The membranes were incubated with
primary antibody overnight at 4°C (anti-STMN1: 1:5,000, Abcam),
anti-E-cadherin: 1:300, anti-vimentin: 1:300 (Ruiying Biological),
anti-β-actin: 1:2500 (Santa Cruz). After being washed three times
with TBST, horseradish peroxidase (HRP)-conjugated secondary
antibodies (goat anti-rabbit IgG-HRP with 1:1,000 dilution for
STMN1, E-cadherin and vimentin; goat anti-mouse IgG-HRP with
1:2,000 dilution for β-actin) were used for 1.5 h at room
temperature. After three additional washes with TBST for 45 min.
The immunoreactive bands were detected with ECL plus kit (ZSbio,
Beijing, China).
Cell viability studies by CCK-8
assays
FaDu cells (5×103 cells/well) were plated
into a 96-well plate containing complete medium and incubated for
24 h before transfection. After treatment with siRNAs for 0, 12,
24, 48 and 72 h, 10 µl Cell Counting Kit-8 (CCK-8) kit
solution (Beyotime) was added to each well containing original
medium in a final volume of 100 µl and the plate was
incubated for 1.5 h in an incubator at 37°C. Then the optical
density (OD) was measured at 450 nm by using a Fluorescence
Spectrophotometer (Hitachi, Japan). Three wells were used for each
experimental condition and all the experiments were independently
repeated three times.
Flow cytometric cell cycle analysis
For cell cycle analysis, FaDu cells were harvested
at 48 h post-transfection, 6-cm plates. Cells were collected using
cold PBS and fixed with 70% cold ethanol over 24 h in −20°C and
treated with 1 ng/ml RNaseA for 20 min at 4°C. Cellular DNA was
stained with 0.5 mg/ml propidium iodide (PI, 50 mg/ml;
Becton-Dickinson, San Jose, CA, USA) for 20 min at 4°C in the dark.
The cells were then sorted by FACSCalibur Flow Cytometer
(Becton-Dickinson, San Jose, CA, USA) and CellQuest acquisition and
analysis programs (Becton-Dickinson, San Diego, CA, USA). The
experiments were replicated three times.
Transwell assay
A Transwell chamber was used (8 µm, 24-well
format; BD Biosciences) to perform cell migration assays. For the
migration assay, 5×104 cells in 500 µl serum-free
media were added to the upper chamber after transfection for 48 h,
while medium containing 10% FBS was added to the lower chambers to
act as a chemoattractant. The upper surfaces of the chamber were
coated with a growth factor-reduced Matrigel matrix. Cells were
incubated at 37°C with 5% CO2 for 24 h, and the medium
was removed from the upper chamber. The non-invaded cells on the
upper side of the chamber were gently scraped off with a cotton
swab, whereas the cells that had migrated through the membrane were
stained with 4% paraformaldehyde and 0.1% crystal violet, imaged,
and counted using an inverted microscope and analyzed by
visualizing five random fields at a magnification of ×200. Each
experiment was replicated three times.
Observation of morphological changes
Morphological changes of the FaDu cells were
observed using an inverted microscope. The image was taken using a
Leica microscope image system (Leica, Mannheim, Germany) at a
magnification of ×200.
Statistical analysis
Statistical analyses were performed using SPSS
statistical software package, version 22.0. The STMN1 IHC scores of
each subgroup of clinicopathological parameters in Table I are shown as mean ± standard
deviation (SD). Statistical significance was determined using
t-test. Kaplan-Meier and log-rank tests were used for calculating
survival curves. In addition, statistical analysis was performed
using GraphPad Prism 5.0 statistical software, sigmaplot 10.0 and
Adobe Photoshop CC. Statistical significance was assigned for
P<0.05.
Results
Relationship between STMN1 expression and
clinicopathologic parameters in HSCC specimens
We first sought to investigate the role of STMN1 in
HSCC. Sixteen human normal cases and 51 HSCC cases were observed by
us through immunohistochemistry staining. STMN1 protein was
localized in the cytoplasm and, partly, in the nucleus in all tumor
tissues, while negative in all of the normal tissues.
With respect to the staining intensity, normal
tissue (NT), cancer in situ (CIS), and invasive cancer (IC)
displayed negative, moderate and strong immunoreactivity for STMN1
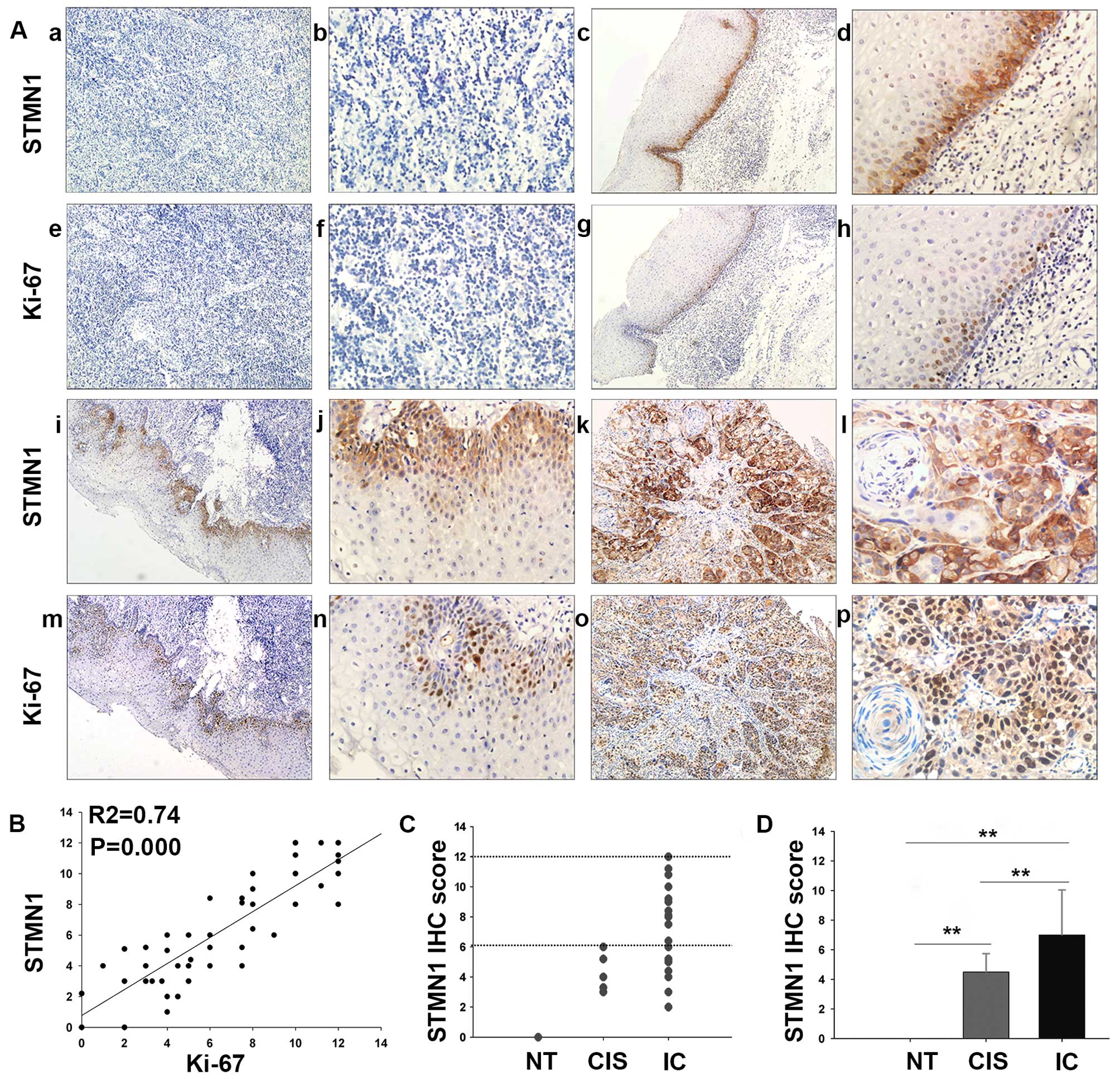
protein, respectively (Fig. 1A).
As showed in Fig. 1C and D, STMN1
IHC score was statistically increased in IC tissue (7.05±3.13)
compared with that in the CIS (4.13±1.38, P=0.009) or normal tissue
(0, P<0.001). Moreover, the correlations between the expression
of STMN1 and clinicopathological characteristics of the patients
with HSCC are summarized in Table
I (Fig. 2). Interestingly, IHC
staining showed that the intensity of STMN1 expression was much
stronger in poorly differentiated than well-differentiated samples
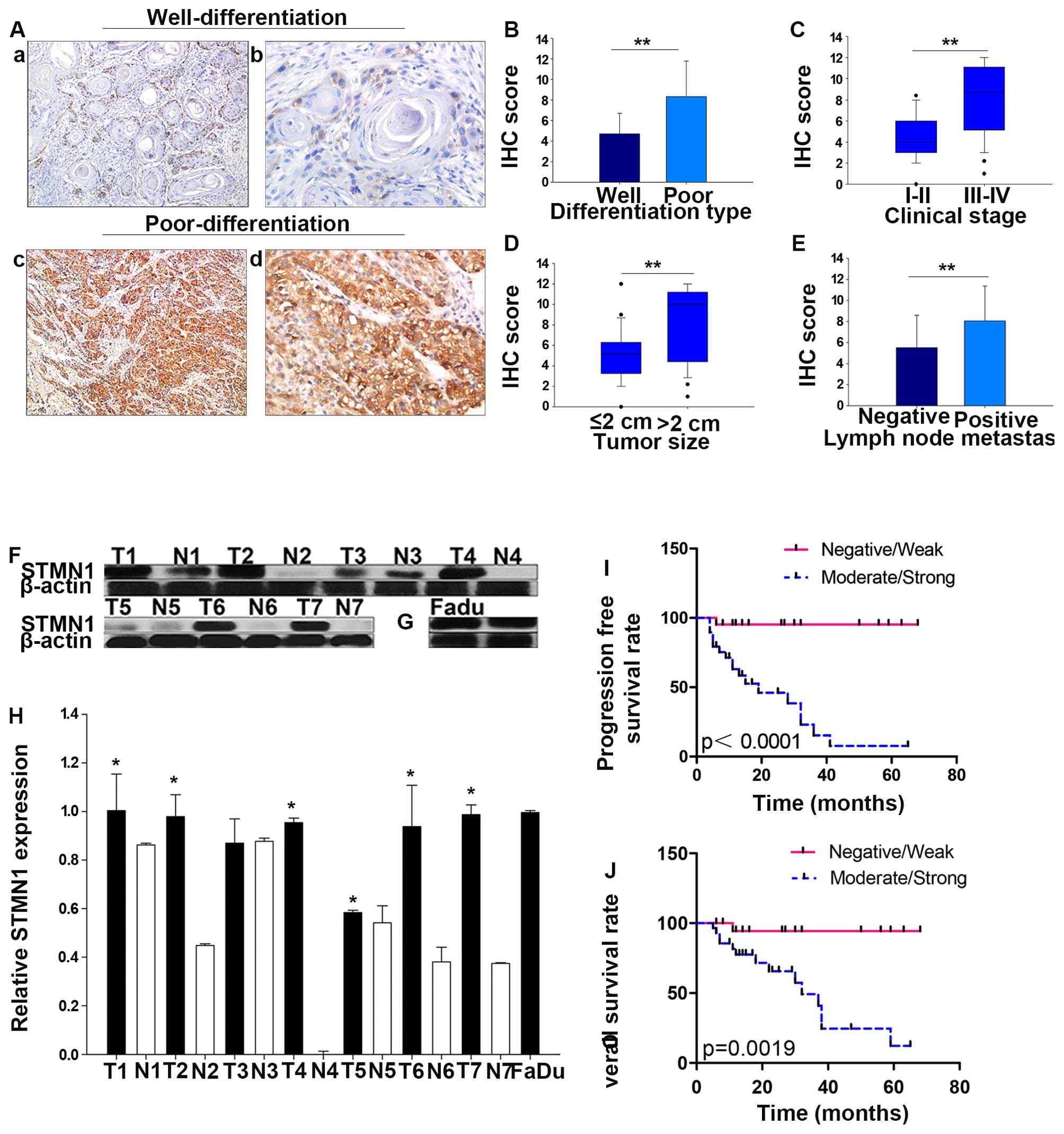
(P<0.001, Fig. 2A and B). In
addition, overexpression of STMN1 was significantly correlated with
advanced clinical stage (stage III and IV compared with stage I and
II, P<0.001, Fig. 2C), large
tumor size (size >2 cm compared with those ≤2 cm, P=0.001,
Fig. 2D), lymph node metastasis
(metastasis to the lymph nodes compared with non-metastasis,
P=0.008, Fig. 2E) and treatment
(P=0.01), but not correlated with age (P=0.430), gender (P=0.824),
tobacco smoking (P=0.242), or alcohol consumption (P=0.242).
STMN1 is overexpressed in human HSCC
tissues and FaDu cells
To further verify the results of
immunohistochemistry staining, STMN1 expression at protein levels
in 7 HSCC tumor tissues (T1–T7) and their adjacent tissues (N1–N7)
were detected using western blot analysis. Results showed that,
compared with their adjacent tissues, the expression of STMN1 in
HSCC tumor was significantly higher (Fig. 2F and H). Besides, the basic
expression and distribution of STMN1 in FaDu cells were examined at
protein levels. As is showed in Fig.
2G, the results of western blot analysis suggested that STMN1
is highly expressed in FaDu cells. Simultaneously, strong
immunoreactivity for STMN1 protein in cytoplasm in FaDu cells was
found by immunocytochemistry (Fig.
3D).
STMN1 expression significantly associates
with tumor proliferation and poor survival of HSCC patients
Furthermore, specimens with positive STMN1 staining
showed significantly higher frequencies of Ki-67 positivity
(Fig. 1A). Spearman correlation
analysis indicates a positive correlation between STMN1 expression
and Ki-67 based on proliferative activity (R2=0.74,
P=0.000; Fig. 1B). Thus, these
findings indicate that overexpression of STMN1 is likely to be
involved in the progression of HSCC.
In addtion, in light of our results that displayed a
multifaceted expression of STMN1 in HSCC patients with different
malignancy grade, we investigated the prognostic significance of
STMN1 in HSCC using Kaplan-Meier analysis. Indeed, increased
expression of STMN1 was significantly associated with worse
prognoses. The choice of treatment is usually associated with
clinical stage and the presence of lymph node metastasis, thus the
expression of STMN1 was different in patients under different
therapy. STMN1 samples highly or moderately stained indicated
shorter overall survival and progression-free survival rate than
those with STMN1 weakly or negatively stained in the 51 HSCC
specimens (P=0.0019, P<0.0001; Fig.
2I and J).
Establishment of siRNAs targeting
STMN1
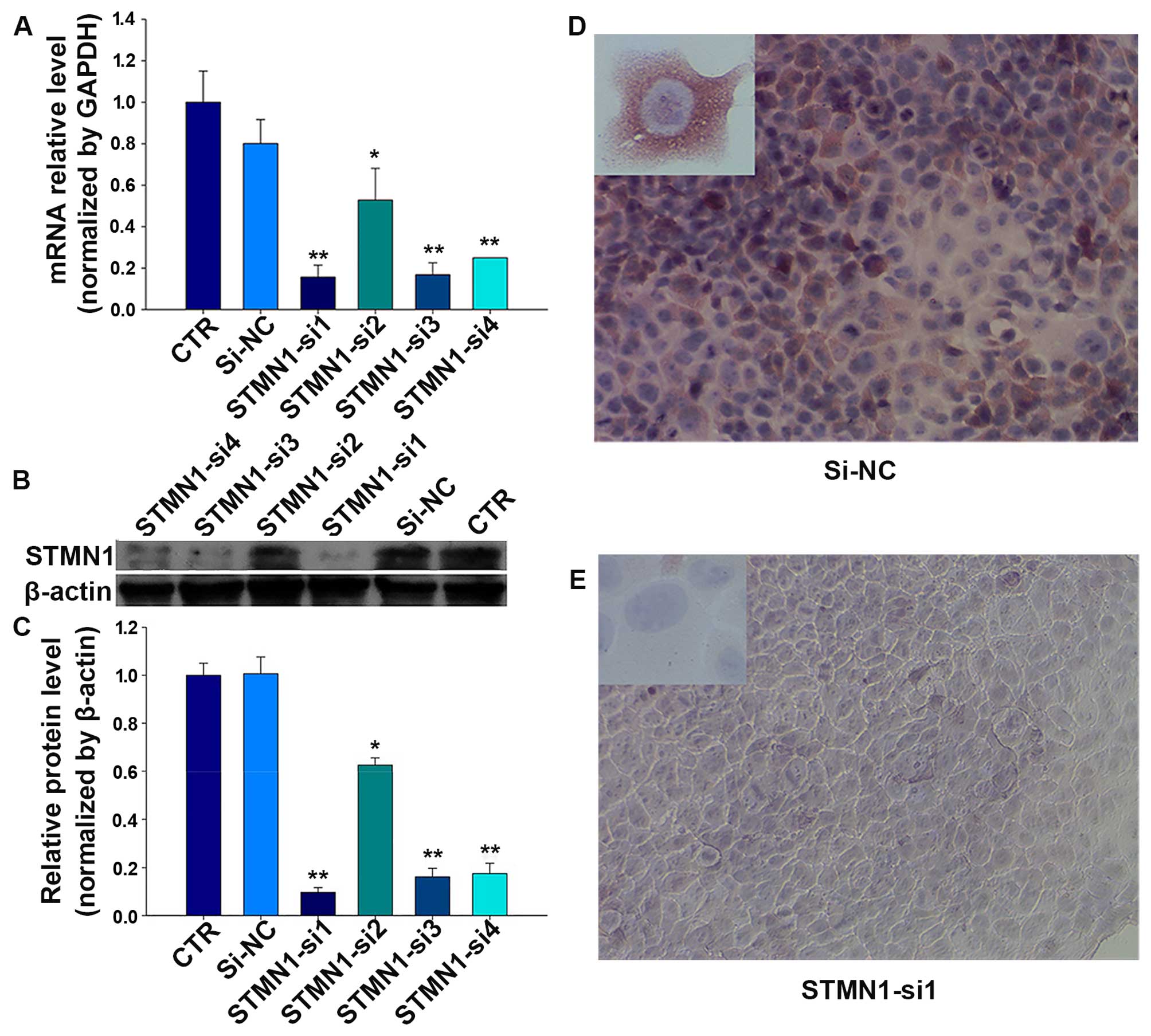
To investigate the effect of STMN1 on HSCC, four
siRNAs were designed to knock down the STMN1 expression in FaDu
cells. The mRNA and protein levels were determined in treated FaDu
cells after 48 h. As shown in Fig.
3A, compared with the control (CTR) and si-NC, after treatments
with FaDu cells for 72 h, STMN1 expression was inhibited by siRNAs
at the mRNA level achieving 87% (STMN1-si1), and the protein level
achieving 90% (STMN1-si1) (Fig. 3B and
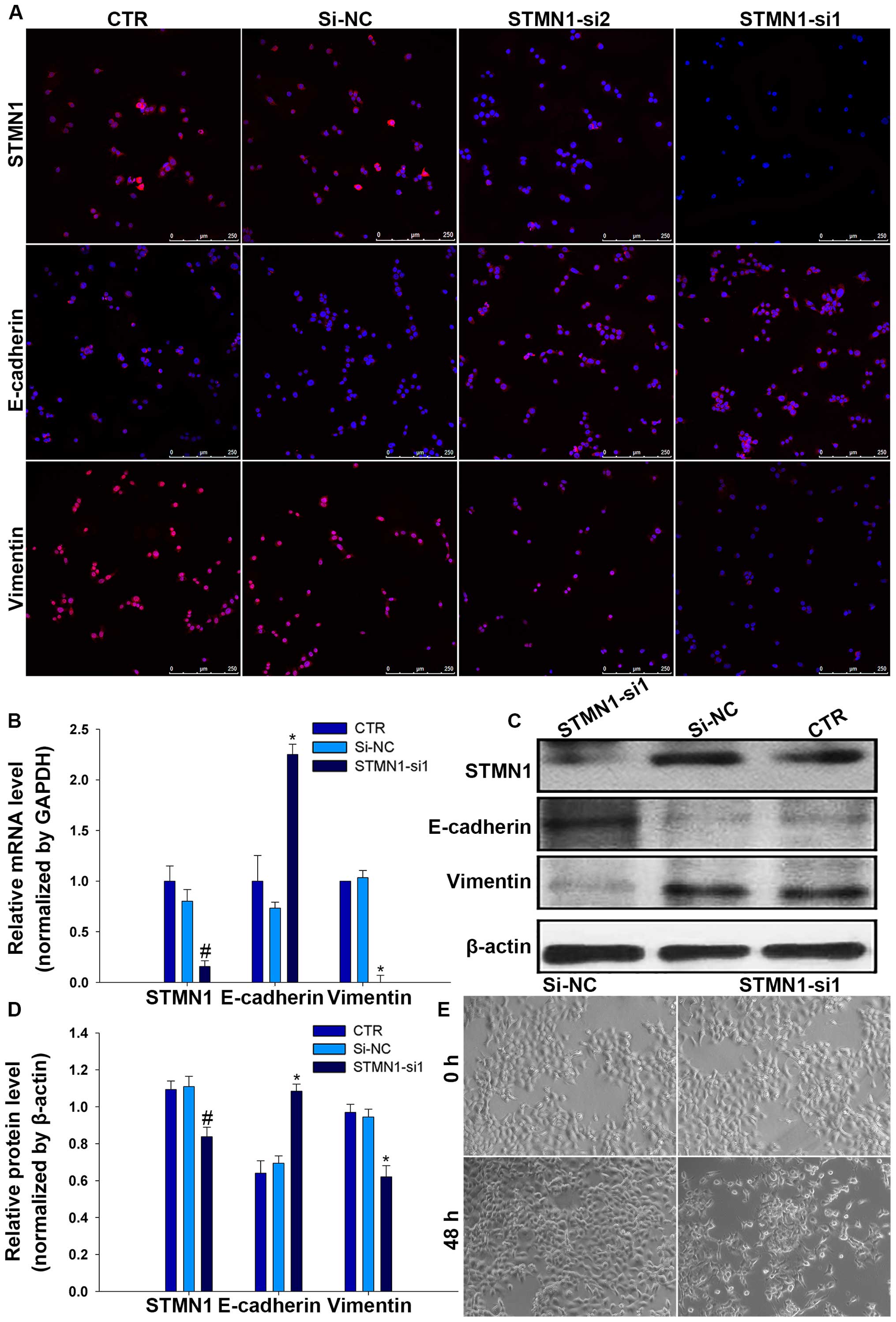
C). Furthermore, the results of immunofluorescence indicated
that the staining intensity of STMN1 in FaDu cells significantly
diminished after treatment with STMN1-si1 (Fig. 3D and E).
STMN1 knockdown inhibits cellular
proliferation and promotes cell cycle arrest
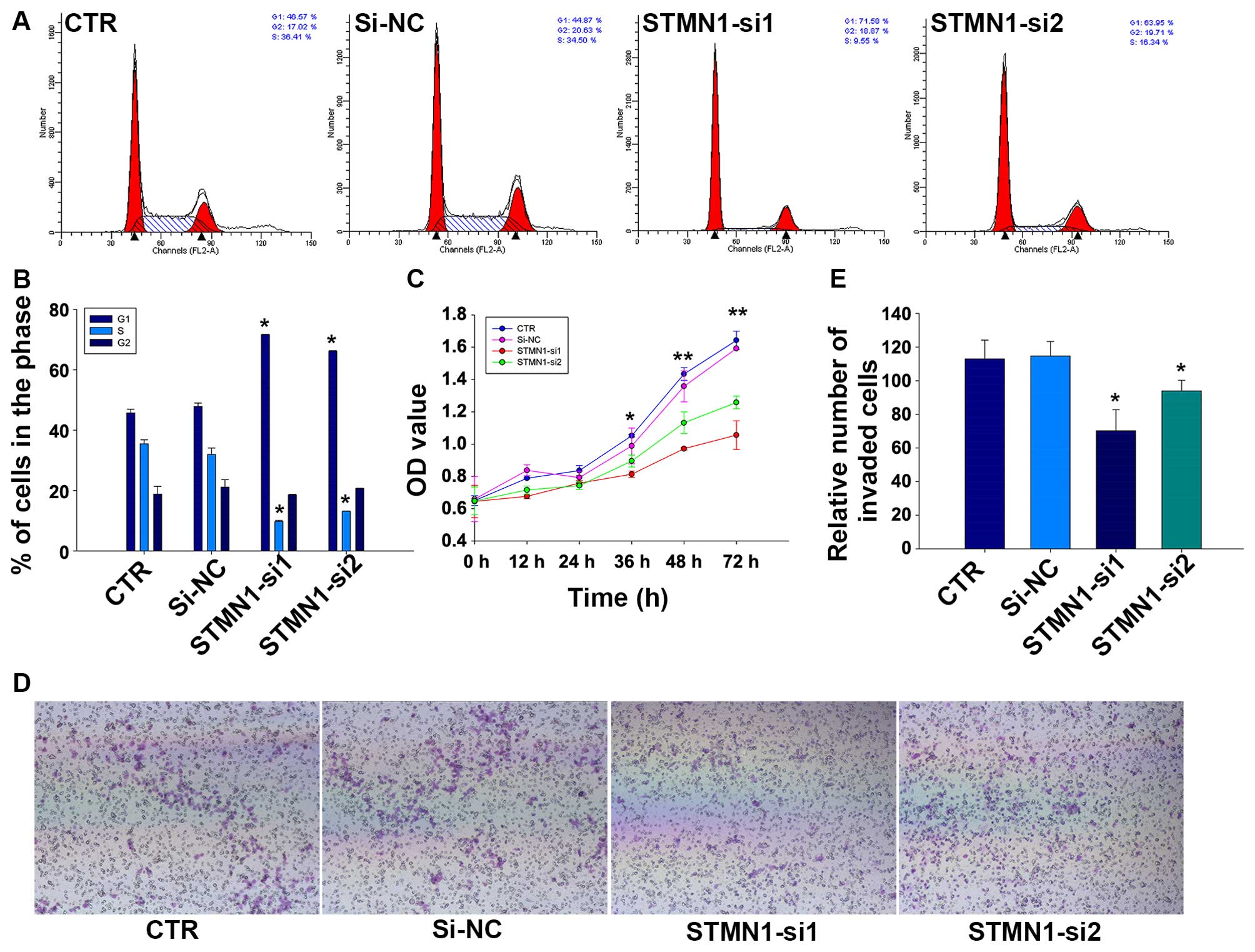
To assess the effect of knocking down STMN1 in
proliferation of FaDu cells, CCK-8 and flow cytometry assays were
implemented. Results of CCK-8 assay showed that the cell
proliferation of FaDu could be greatly reduced by treating with
STMN1 siRNAs in comparison with si-NC or control (CTR) (Fig. 4C). We further examined the effect
of STMN1 siRNAs on cell cycle distribution of FaDu cells. Flow
cytometry analysis revealed that the knockdown of STMN1 expression
in FaDu increased the percentage of cells in G1 phase (45.72±1.2
and 47.88±1.13 vs. 71.67±0.12 and 66.18±0.20%, respectively;
P<0.05) whereas decreased the percentage of cells in S phase
(35.46±1.35 and 31.96±2.14 vs. 9.77±0.31 and 13.15±0.09%,
respectively; P<0.05; Fig. 4A and
B). These results indicated that reduction of STMN1 expression
in FaDu cells could inhibit cellular proliferation and promote cell
cycle arrest.
STMN1 knockdown reduces the migration
activity and induces a change of EMT-related protein expressions in
FaDu cells
The influence of STMN1 knockdown on the migration
activity of FaDu cells was detected by Transwell assays. As is
shown in Fig. 4E, the results
indicated that there were fewer cells migrated to the bottom
chambers in FaDu/STMN1-si1 (70.33±12.50). FaDu/STMN1-si2
(94.00±6.25) compared with FaDu/si-NC (114.67±8.74) and CTR
(113.00±11.14). The representative micrographs are shown in
Fig. 4D.
EMT is a crucial mechanism for migration of multiple
tumor cells, with epithelial cells usually losing the capability of
intercellular adhesion and acquiring a mesenchyme phenotype
(11). To assess whether STMN1
promotes FaDu cell migration via stimulating EMT, EMT-associated
proteins were detected by immunofluorescence in FaDu/STMN1-siRNA
and FaDu/si-NC and CTR. Interestingly, results showed that the
expression level of E-cadherin decreased while the expression level
of vimentin increased by depleting STMN1 levels in FaDu cells
(Fig. 5A), suggestive of a less
epithelial phenotype and a more mesenchyme phenotype. These changes
in mRNA and protein expression were confirmed by RT-qPCR and
western blot analysis (Fig. 5B–D).
During the EMT process, this is usually accompanied by a change in
cell morphology (12). To explore
whether knockdown of STMN1 has an impact on the morphology of FaDu
cells, cell morphological changes were evaluated by an inverted
microscope after treated with STMN1-si1 or si-NC. The results
showed that cells treated with STMN1-si1 were changed from a more
rounded, cell polarity to an elongated, with loss of cell-cell
contacts, but there was no effect on cells treated with si-NC
(Fig. 5E).
Discussion
In this study, we found that STMN1 expression was
linked to tumor grade, differentiation, size, stage and
proliferation (Ki-67 index) of HSCC. Also, higher protein level of
STMN1 was discovered in HSCC fresh tissues compared with non-cancer
tissues. The results of Kaplan-Meier analysis suggested that STMN1
overexpression was closely related to progression-free survival
(PFS) and overall survival (OS) of HSCC patients. Furthermore,
studies in vitro indicated the vital role of STMN1 in FaDu
cell proliferation and migration after STMN1 knockdown.
Previous studies showed that high-expressing and
abnormal activation of STMN1 in multiple human cancers along with
its overexpression usually indicated an unfavorable prognosis in
patients with malignancies (8). It
is suggested that STMN1 overexpression is independently predictive
of DSS (disease-specific survival), DMeFS (distal metastasis-free
survival) and LRFS (local recurrence-free survival) in
nasopharyngeal carcinoma (13).
The study of Trovik et al (14) also showed that STMN1
immunohistochemical staining identified with high grade, lymph node
metastases and poor survival through studies on 1,076 endometrial
cancer patients. In breast cancer, high STMN1 and phospho-STMN1
levels predicted poor survival of patients (15). Similar notions were verified in
non-small cell lung cancer (16),
colorectal cancer (17) and
pheochromocytomas (18). Our data
were in favour of this observation as well. We demonstrated that
STMN1 was differentially expressed in normal, CIS, IC HSCC tissues.
Elevated STMN1 was significantly associated with tumor
differentiation, tumor size, clinical stage, lymph node metastasis
and showed a positive correlation with Ki-67 index (19). Patients with moderate/strong STMN1
staining intensity always underwent poor prognosis. All the above
implied that STMN1 may be used as a potential prognostic marker for
survival in patients with HSCC.
Microtubule is an essential part of the
cytoskeleton, which is required for a wide variety of fundamental
cellular functions, including the maintenance of cell
differentiation, cell morphology, motility, and polarity (20). With proven destabilizing activity
(7), the abnormal activation or
increased expression of STMN1 has enhanced the proliferation, cell
cycle progression, migration and invasion in human cancer cell
lines, including gastric (21),
ovarian (22), and hepatoma cell
lines (23). In hepatoma HCCLM3
cells, studies found that STMN1 siRNA could obviously inhibit cell
proliferation and migration (24).
In human endometrial carcinoma (EC) cell line Ishikawa, STMN1 was
further identified showing a positive effect on cell viability and
migration (25). In agreement with
previous reports, our studies in vitro suggested that
reduced expression of STMN1 in FaDu cells obviously induced a cell
cycle arrest in G1 phase, accompanied by inhibited cell viability.
Our investigations also suggested that overexpressed STMN1 could
accelerate migration in FaDu cells. Interestingly, high level of
STMN1 was noted especially in the tumor fronts and invading tumor
islets of HSCC tissue sections. These results may account for this
higher expression level of STMN1 in large tumor size, lymph node
metastasis and poor prognosis of HSCC patients, simultaneously,
informing us that STMN1 may participate in the progression of
HSCC.
Metastasis of tumor is a multistep and complex
process, yet it remains the most poorly understood component of
cancer pathogenesis (26). A
handful of studies suggest that EMT facilitates cancer epithelial
cells to enter into a mesenchymal-like state, which endows them
with migratory and invasive properties (27). EMT process is accompanied by a loss
of epithelial marker proteins and dissolution of adherent junction
proteins, such as E-cadherin, cytokeratin, γ-catenin and β-catenin
which play a significant role in cell-cell adhesion. Concomitantly,
mesenchymal marker proteins, such as vimentin, N-cadherin,
fibronectin and P-cadherin which contribute to cell migration, are
frequently overexpressed (12,28).
E-cadherin as the best characterized cadherins, in particular, has
a key role in epithelial cell-cell adhesion (29). Decreased E-cadherin expression
usually induces cell migration, morphological changes and cancer
development (30), while
upregulated vimentin are in cancer cells resulting in epithelial
cells to acquire a mesenchymal shape and increased motility
(31). Indeed, EMT has been
reported as a pivotal program in numerous human solid cancers,
including pancreas cancer (32),
prostate cancer (33) and breast
cancer (34), ultimately resulting
in tumor metastasis. Lu et al (35) performed a series of evidence to
elaborate the role of stathmin1 and microtubule dynamics in
promoting EMT. In addition, via the inhibition of stathmin1, Li
et al (36) found Siva1
could enhance the formation of microtubules and impedes focal
adhesion assembly, cell migration, and EMT. Our present findings
are consistent with these previous results. We demonstrated that
reducing the expression of STMN1 would result in activation of
epithelial marker E-cadherin along with blocked mesenchyme markers
vimentin expression in HSCC. These investigations suggested that
overexpressed STMN1 may enhance metastasis and aggressiveness in
HSCC, at least, in part, via facilitating the EMT process.
In conclusion, our results indicate that STMN1
possesses higher degree of malignancy and shorter PFS and OS of
HSCC, suggesting a potential diagnostic biomarker for HSCC.
Furthermore, the results from the present work emphasize the vital
role of STMN1 in promoting proliferation, enhancing migration and
encouraging EMT in FaDu cells. Thus, our data revealed that STMN1
may contribute to the aggressive phenotype of human HSCC, and,
perhaps, act as a promising molecular target for controlling cancer
progression.
References
|
1
|
Hall SF, Groome PA, Irish J and O'Sullivan
B: The natural history of patients with squamous cell carcinoma of
the hypopharynx. Laryngoscope. 118:1362–1371. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chung EJ, Lee SH, Baek SH, Park IS, Cho SJ
and Rho YS: Pattern of cervical lymph node metastasis in medial
wall pyriform sinus carcinoma. Laryngoscope. 124:882–887. 2014.
View Article : Google Scholar
|
|
3
|
Lee MS, Ho HC, Hsiao SH, Hwang JH, Lee CC
and Hung SK: Treatment results and prognostic factors in locally
advanced hypopharyngeal cancer. Acta Otolaryngol. 128:103–109.
2008. View Article : Google Scholar
|
|
4
|
Chan JY and Wei WI: Current management
strategy of hypopharyngeal carcinoma. Auris Nasus Larynx. 40:2–6.
2013. View Article : Google Scholar
|
|
5
|
Zhou L: Standardization of the management
of laryngeal and hypopharyngeal carcinoma. Zhonghua Er Bi Yan Hou
Tou Jing Wai Ke Za Zhi. 44:705–706. 2009.In Chinese.
|
|
6
|
Curmi PA, Gavet O, Charbaut E, Ozon S,
Lachkar-Colmerauer S, Manceau V, Siavoshian S, Maucuer A and Sobel
A: Stathmin and its phosphoprotein family: General properties,
biochemical and functional interaction with tubulin. Cell Struct
Funct. 24:345–357. 1999. View Article : Google Scholar
|
|
7
|
Gupta KK, Li C, Duan A, Alberico EO, Kim
OV, Alber MS and Goodson HV: Mechanism for the
catastrophe-promoting activity of the microtubule destabilizer
Op18/stathmin. Proc Natl Acad Sci USA. 110:20449–20454. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nemunaitis J: Stathmin 1: A protein with
many tasks. New biomarker and potential target in cancer. Expert
Opin Ther Targets. 16:631–634. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li X, Wang L, Li T, You B, Shan Y, Shi S,
Qian L and Cao X: STMN1 overexpression correlates with biological
behavior in human cutaneous squamous cell carcinoma. Pathol Res
Pract. 211:816–823. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
11
|
Acloque H, Adams MS, Fishwick K,
Bronner-Fraser M and Nieto MA: Epithelial-mesenchymal transitions:
The importance of changing cell state in development and disease. J
Clin Invest. 119:1438–1449. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kalluri R: EMT: When epithelial cells
decide to become mesenchymal-like cells. J Clin Invest.
119:1417–1419. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hsu HP, Li CF, Lee SW, Wu WR, Chen TJ,
Chang KY, Liang SS, Tsai CJ and Shiue YL: Overexpression of
stathmin 1 confers an independent prognostic indicator in
nasopharyngeal carcinoma. Tumour Biol. 35:2619–2629. 2014.
View Article : Google Scholar
|
|
14
|
Trovik J, Wik E, Stefansson IM,
Marcickiewicz J, Tingulstad S, Staff AC, Njolstad TS, Vandenput I,
Amant F, Akslen LA, et al: MoMaTec Study Group: Stathmin
overexpression identifies high-risk patients and lymph node
metastasis in endometrial cancer. Clin Cancer Res. 17:3368–3377.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kuang XY, Chen L, Zhang ZJ, Liu YR, Zheng
YZ, Ling H, Qiao F, Li S, Hu X and Shao ZM: Stathmin and
phospho-stathmin protein signature is associated with survival
outcomes of breast cancer patients. Oncotarget. 6:22227–22238.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Nie W, Xu MD, Gan L, Huang H, Xiu Q and Li
B: Overexpression of stathmin 1 is a poor prognostic biomarker in
non-small cell lung cancer. Lab Invest. 95:56–64. 2015. View Article : Google Scholar
|
|
17
|
Tan HT, Wu W, Ng YZ, Zhang X, Yan B, Ong
CW, Tan S, Salto-Tellez M, Hooi SC and Chung MC: Proteomic analysis
of colorectal cancer metastasis: Stathmin-1 revealed as a player in
cancer cell migration and prognostic marker. J Proteome Res.
11:1433–1445. 2012. View Article : Google Scholar
|
|
18
|
Björklund P, Cupisti K, Fryknäs M,
Isaksson A, Willenberg HS, Akerström G, Hellman P and Westin G:
Stathmin as a marker for malignancy in pheochromocytomas. Exp Clin
Endocrinol Diabetes. 118:27–30. 2010. View Article : Google Scholar
|
|
19
|
Ghanim B, Klikovits T, Hoda MA, Lang G,
Szirtes I, Setinek U, Rozsas A, Renyi-Vamos F, Laszlo V, Grusch M,
et al: Ki67 index is an independent prognostic factor in
epithelioid but not in non-epithelioid malignant pleural
mesothelioma: A multicenter study. Br J Cancer. 112:783–792. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Belletti B, Nicoloso MS, Schiappacassi M,
Berton S, Lovat F, Wolf K, Canzonieri V, D'Andrea S, Zucchetto A,
Friedl P, et al: Stathmin activity influences sarcoma cell shape,
motility, and metastatic potential. Mol Biol Cell. 19:2003–2013.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Akhtar J, Wang Z, Zhang ZP and Bi MM:
Lentiviral-mediated RNA interference targeting stathmin1 gene in
human gastric cancer cells inhibits proliferation in vitro and
tumor growth in vivo. J Transl Med. 11:2122013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wei SH, Lin F, Wang X, Gao P and Zhang HZ:
Prognostic significance of stathmin expression in correlation with
metastasis and clinicopathological characteristics in human ovarian
carcinoma. Acta Histochem. 110:59–65. 2008. View Article : Google Scholar
|
|
23
|
Singer S, Ehemann V, Brauckhoff A, Keith
M, Vreden S, Schirmacher P and Breuhahn K: Protumorigenic
overexpression of stathmin/Op18 by gain-of-function mutation in p53
in human hepatocarcinogenesis. Hepatology. 46:759–768. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gan L, Guo K, Li Y, Kang X, Sun L, Shu H
and Liu Y: Up-regulated expression of stathmin may be associated
with hepatocarcinogenesis. Oncol Rep. 23:1037–1043. 2010.PubMed/NCBI
|
|
25
|
He X, Liao Y, Lu W, Xu G, Tong H, Ke J and
Wan X: Elevated STMN1 promotes tumor growth and invasion in
endometrial carcinoma. Tumour Biol. 37:9951–9958. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chaffer CL and Weinberg RA: A perspective
on cancer cell metastasis. Science. 331:1559–15642. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xu Q, Deng F, Qin Y, Zhao Z, Wu Z, Xing Z,
Ji A and Wang QJ: Long non-coding RNA regulation of
epithelial-mesenchymal transition in cancer metastasis. Cell Death
Dis. 7:e22542016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Przybyla L, Muncie JM and Weaver VM:
Mechanical control of epithelial-to-mesenchymal transitions in
development and cancer. Annu Rev Cell Dev Biol. 32:527–554. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
van Roy F and Berx G: The cell-cell
adhesion molecule E-cadherin. Cell Mol Life Sci. 65:3756–3788.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chan SH and Wang LH: Regulation of cancer
metastasis by microRNAs. J Biomed Sci. 22:92015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zeisberg M and Neilson EG: Biomarkers for
epithelial-mesenchymal transitions. J Clin Invest. 119:1429–1437.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Park JS, Lee JH, Lee YS, Kim JK, Dong SM
and Yoon DS: Emerging role of LOXL2 in the promotion of pancreas
cancer metastasis. Oncotarget. Jun 7–2016.Epub ahead of print.
View Article : Google Scholar
|
|
33
|
Bloom JE and McNeel DG: SSX2 regulates
focal adhesion but does not drive the epithelial to mesenchymal
transition in prostate cancer. Oncotarget. Jun 2–2016.Epub ahead of
print. PubMed/NCBI
|
|
34
|
Karaczyn AA, Adams TL, Cheng RY, Matluk NN
and Verdi JM: Human NUMB6 induces epithelial-mesenchymal transition
and enhances breast cancer cells migration and invasion. J Cell
Biochem. Jun 15–2016.Epub ahead of print. PubMed/NCBI
|
|
35
|
Lu Y, Liu C, Xu YF, Cheng H, Shi S, Wu CT
and Yu XJ: Stathmin destabilizing microtubule dynamics promotes
malignant potential in cancer cells by epithelial-mesenchymal
transition. Hepatobiliary Pancreat Dis Int. 13:386–394. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li N, Jiang P, Du W, Wu Z, Li C, Qiao M,
Yang X and Wu M: Siva1 suppresses epithelial-mesenchymal transition
and metastasis of tumor cells by inhibiting stathmin and
stabilizing microtubules. Proc Natl Acad Sci USA. 108:12851–12856.
2011. View Article : Google Scholar : PubMed/NCBI
|