Introduction
Glioblastomas have an age adjusted incidence of 3
per 100,000 persons per year in the United States and comprise
approximately 45% of all gliomas with a relative survival rate of
5%, 5-years post-diagnosis (1),
which can be attributed in part to high rates of tumor recurrence.
A major contributor to the low survival rates of patients with
these cancers is the incredibly complex glioblastoma genome, which
has a vast number of uncharacterized expressed genetic
abnormalities that contribute to intra- and intertumor variation
that negatively impact therapeutic response and patient outcomes
(2). The poor prognosis of brain
tumor patients, particularly those with glioblastoma, can be
attributed in part to the limited number of aberrantly expressed
factors used clinically as diagnostic and prognostic indicators.
These have included genetic mutations, deletions, amplifications,
and epigenetic changes in isocitrate dehydrogenase (IDH), EGFR and
O6-methylguanine DNA methyltransferase (MGMT) (3–8). In
the present study, we investigated microtubule actin cross-linking
factor 1 (MACF1), which functions to crosslink microtubules and
actin filaments, for its role in tumor cell biology and as a novel
target in glioblastoma. Although to date MACF1 has received little
consideration in tumor cell biology and as a target in human
cancers, its function as a cytoskeleton integrator has been shown
to be involved in organization of the cytoskeleton (9), tissue repair (10) and vertebrate development (11). These studies provided correlative
evidence that MACF1 is involved in processes such as metastatic
invasion in which cytoskeleton organization is a key element that
contributes to tumor progression in various human cancers, as well
as cellular and physiological processes such as tissue repair when
dysregulated in normal tissue can lead to tumorigenesis.
Additionally, studies by Sjöblom et al (12) identified MACF1 as a candidate
breast cancer gene in a mutational analysis study, while
Misquitta-Ali et al (13)
detected alternatively spliced exons in MACF1 transcripts from lung
cancer patients providing further evidence, albeit circumstantial,
that MACF1 plays a role in human cancer. In the present study, we
demonstrated that genetic inhibition of MACF1, which we found to be
expressed at high levels in glioblastoma relative to normal brain
tissue, had anti-tumorigenic effects on glioblastoma cell behavior
and a synergistic effect with temozolomide, an alkylating agent
used clinically to treat this disease. To the best of our
knowledge, this is the first investigation to evaluate the
functional role of MACF1 in human cancers and as a target that
enhances the efficacy of therapeutic treatment modalities.
Materials and methods
Cells culture conditions and
reagents
U251 human glioblastoma cells were purchased from
Sigma-Aldrich (St. Louis, MO, USA) and the U87 and A172 human
glioblastoma cells were from the American Type Culture Collection
(ATCC; Manassas, VA, USA). Normal human astrocytes were obtained
from Sciencell Research Laboratories (Carlsbad, CA, USA). All cell
lines were maintained in Dulbecco's modified Eagle's medium-DMEM
(Invitrogen, Carlsbad, CA, USA) containing 10% fetal bovine serum
(FBS; Invitrogen), 2 mM L-glutamine (Invitrogen), 100 nM MEM
non-essential amino acids (Invitrogen) and penicillin-streptomycin
(Invitrogen) at 37°C and 5% CO2. Accell SMARTpool siRNAs
targeting MACF1 were purchased from GE Dharmacon (Chicago, IL,
USA). Temozolomide was purchased from Sigma-Aldrich. GBM76 patient
derived xenograft cells were established at the Mayo Clinic
(Rochester, MN, USA) (14), while
primary xenograft lines T4105 and T4302 were provided by Jeremy
Rich from the Cleveland Clinic (Cleveland, OH, USA). Patient
derived xenograft cell lines (GBM76, T4105 and T4302) were serially
passaged as subcutaneous tumors in immunocompromised mice and
maintained as ex vivo cultures in neurobasal medium
supplemented with B27 serum substitute, 20 ng/ml EGF and 20 ng/ml
bFGF (Life Technologies, Carlsbad, CA, USA).
Immunohistochemistry
Brain tumor tissue arrays were purchased from US
Biomax, Inc., (Rockville, MD, USA) to evaluate MACF1 expression.
Each slide contained the following number and array of brain tumor
tissue sections provided by the supplier: 10 grade (I–II)
astrocytomas, 10 grade (III–IV) astrocytomas, 33 grade (IV)
glioblastomas, 3 oligoastrocytomas, 6 grade (I–II)
oligodendrogliomas, 3 medulloblastomas, 3 anaplastic
oligodendrogliomas, 4 ependymomas and cancer adjacent normal tissue
from 5 brains. Paraffin-embedded tissue sections were dewaxed in
xylene for 5 min, rehydrated in 100% ethanol, 95% ethanol and
water. Next tissue was fixed with 100% methanol at −20°C for 15 min
and washed three times with phosphate-buffered saline (PBS).
Detection of MACF1 was conducted using the ImmunoCruz rabbit ABC
Staining system (Santa Cruz Biotechnology, Dallas, TX, USA). Tissue
sections were blocked for 1 h in 1.5% normal horse serum in PBS at
room temperature, permeabilized with 0.05% Triton-X in PBS for 10
min, incubated overnight at 4°C with a polyclonal MACF1 antibody
(Santa Cruz Biotechnology) and rinsed three times with PBS.
Subsequently, sections were incubated with a biotinylated secondary
antibody for 3 h at room temperature and washed in PBS three times
5 min each. Sections were then incubated with an
avidin-biotinylated HRP complex for 30 min, rinsed again three
times with PBS and MACF1 protein visualized as diaminobenzidine
(DAB) positive cells in tissue sections with a peroxidase substrate
solution containing substrate buffer, DAB and peroxidase substrate
according to the manufacturer's instructions. Images were then
taken with a Leica DM4000 microscope and QImaging system (Leica
Microsystems, Inc., Buffalo Grove, IL, USA).
MACF1 silencing
For silencing experiments 3.5×104 U251,
A172, or patient derived xenograft cells were plated in 6-well
plates, grown for two days to 70% confluency, and treated with 1
µM non-targeting control siRNAs, 1 µM siRNAs
targeting MACF1 (GE Dharmacon), or shRNAs targeting MACF1 from the
MISSION shRNA Library (TRCN0000294117 and TRCN0000294123;
Sigma-Aldrich) and allowed to incubate for 6 days in Accell media
for siRNAs or neurobasal medium for shRNAs. MACF1 expression was
subsequently examined in control and MACF1 silenced cells using
immunoblotting procedures.
Cell proliferation assay
To assess the effects of silencing MACF1 on cell
proliferation and viability, cells were treated with 1 µM of
non-targeting control siRNAs or siRNAs targeting MACF1 (GE
Dharmacon) and allowed to incubate for 6 days in 6-well plates as
described above. At the end of the 6-day incubation period
1.5×104 cells treated with control or siRNAs targeting
MACF1 were replated in 12-well plates and allowed to incubate in
the absence of siRNAs for 2, 4, and 6 days in DMEM media
(Invitrogen) containing 10% FBS (Invitrogen). At the end of each
time-point cells were evaluated using the crystal violet cell
proliferation assay, one of three standard assays that can be used
to evaluate cytotoxicity and/or cell viability that provide a
measure of cell proliferation (15). Tissue culture medium was removed
and the cell monolayer was fixed with 100% methanol for 5 min and
stained with 0.5% crystal violet in 25% methanol for 10 min. Cells
were then washed three times 5 min each with distilled water to
remove excess dye and allowed to dry overnight at room temperature.
The incorporated dye was then solubilized in 0.1 M sodium citrate
(Sigma-Aldrich) in 50% ethanol. Next, 100 µl of experimental
samples were transferred to 96-well plates and optical densities
read at 540 nm using a xMark microplate absorbance
spectrophotometer (Bio-Rad Laboratories, Hercules, CA, USA). For
combinatorial experiments glioblastoma cells were also first
treated for 6 days with control non-targeting siRNAs or siRNAs
targeting MACF1, as previously described. Next 2.5×104
siRNA treated cells were replated in 12-well plates in the absence
of siRNAs. Controls and conditions for combinatorial experiments
included: replated siRNA treated cells grown for 72 h, replated
siRNA treated cells exposed to TMZ or DMSO 6 h after replating and
allowed to incubate in TMZ or DMSO for 72 h, and non-treated siRNA
cells exposed to TMZ or DMSO 6 h after plating and grown for 72 h
in the presence of TMZ or DMSO. DMSO is a solvent used to dissolve
drugs and was used as a vehicle control to discern the effects of
TMZ from the drug solvent. To evaluate the combinatorial effects of
silencing MACF1 and TMZ treatment on cell proliferation and
viability the cell proliferation assay, as described above, was
also used at the end of the 72-h incubation time-point.
Cell motility
Motility assays were conducted according to the
manufacturer's instructions (Cell Biolabs, Inc., San Diego, CA,
USA). MACF1 silencing was performed as described above. Cell
suspensions containing 0.5–1.0×106 cells/ml of siRNA
control or siRNA targeted MACF1 treated cells were prepared in
serum-free media, while 500 µl of media containing 10% FBS
was added to the lower chamber of the migration plate. A total of
300 µl of siRNA treated cell suspensions were then added to
the inside of each insert and were allowed to incubate for 24 h at
37°C and 5% CO2. Subsequently, non-migratory cells were
removed from plate inserts (per manufacturer's instructions),
migratory cells were stained with crystal violet and the dye eluted
as described above. Optical densities were read at 595 nm using a
xMark microplate absorbance spectrophotometer (Bio-Rad
Laboratories) and statistical evaluation of the data was performed
as described below.
Western blotting
Cells treated for 6 days with control siRNAs or
siRNAs targeting MACF1 were rinsed with PBS and lysed with CelLytic
M Cell lysis reagent (Sigma-Aldrich). Protein concentrations were
subsequently determined using the Bradford method. Proteins were
separated by SDS-PAGE in 4-15% polyacrylamide gels (Bio-Rad
Laboratories), then transferred to nitrocellulose membranes. For
immunoblotting, nitrocellulose membranes were incubated with MACF1
(Santa Cruz Biotechnology), Axin1 (Cell Signaling Technology,
Danvers, MA, USA), phospho β-catenin (Cell Signaling Technology)
and GAPDH (Cell Signaling Technology) antibodies recognizing target
proteins overnight at 4°C. The membranes were then incubated with
an HRP-conjugated secondary antibody for 1 h at room temperature,
analyzed by enhanced chemiluminescence (ECL) detection system
(Thermo Fisher Scientific, Nashville, TN, USA) and visualized using
a UVP BioSpectrum imaging system (UVP, LLC, Upland, CA, USA).
Densitometry analysis was performed with the Image Studio Lite
software (LI-COR Biosciences, Lincoln, NE, USA).
Statistical analysis
Cell proliferation, migration, time-course and
combinatorial experiments were each performed at least three times
with duplicate or triplicate samples. Means were determined by
averaging duplicate or triplicate samples within each independent
experiment. Students t-tests and ANOVA analysis were used to
evaluate the significance between the experimental conditions with
a P-value of 0.05 and 0.01, respectively.
Results
Differential expression of MACF1 in brain
tumors
Identification and expression profiling of tumor
specific biomarkers are essential for clinical diagnostic and
prognostic purposes. Using immunohistochemistry procedures we
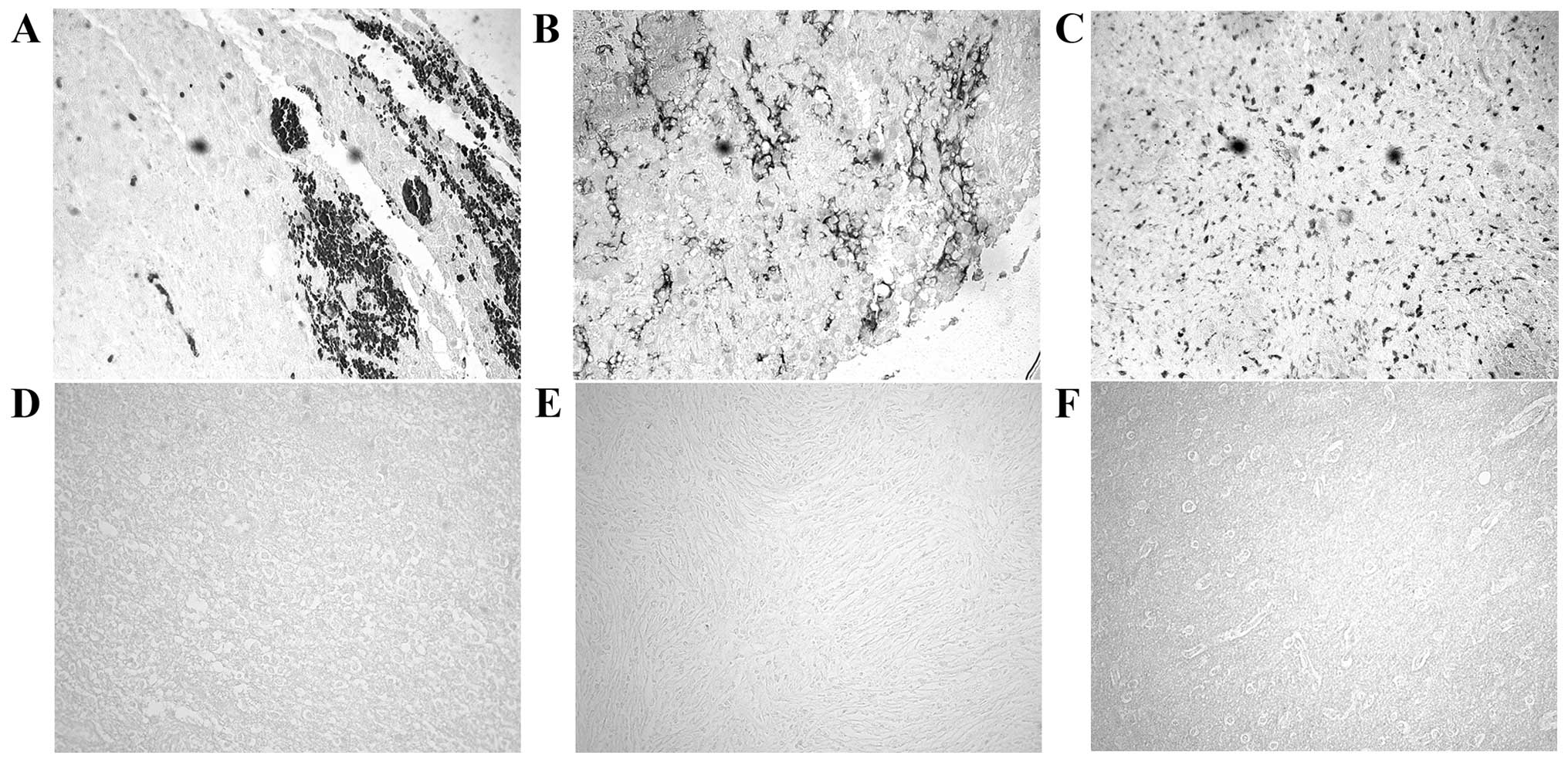
examined MACF1 expression in low and high grade brain tumors. Our
data showed that MACF1 was expressed diffusely and at the periphery
in 40% of grade IV glioblastoma tissue sections as compared to its
absence in normal brain tissue (Fig.
1A–C). In contrast to its expression in glioblastoma, MACF1 was
absent in lower grade brain tumors (oligodendroglioma and
medulloblastoma) and normal brain tissue (Fig. 1D–F), but present in grade II–IV
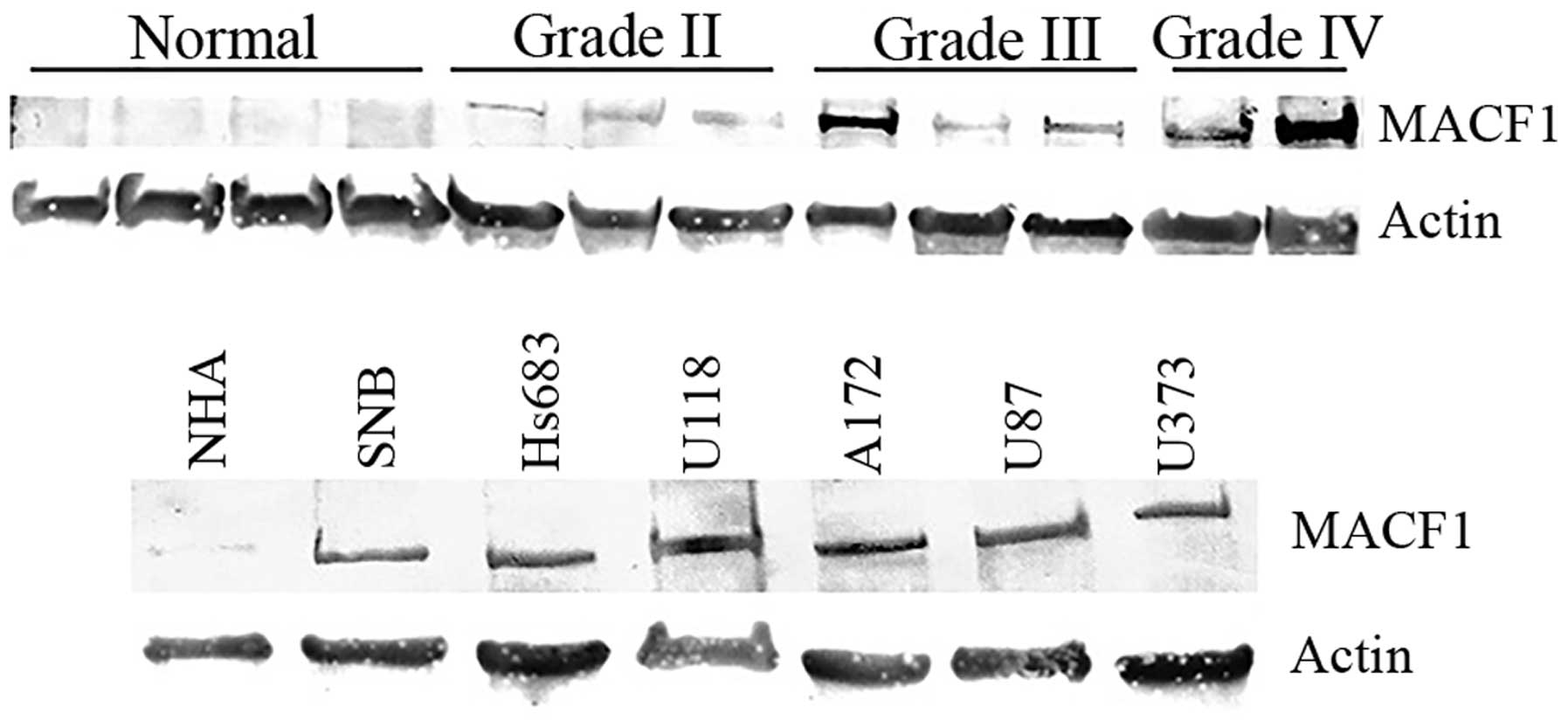
astrocytomas as compared to normal brain tissue (Fig. 2). Complementing data observed in
human tissue, immunoblotting procedures of MACF1 in glioblastoma
cell lines also displayed significantly higher levels of MACF1 as
compared to its expression in normal human astrocyte cells
(Fig. 2). Collectively, these data
provide evidence that MACF1 is predominately expressed in high
grade malignant brain tumors and suggest that it is a potential
biomarker and target in these cancers.
Anti-tumorigenic effects of suppressing
MACF1 in glioblastoma
MACF1 to date has received no consideration as a
target in cancer and associated cell behavior. Using siRNAs to
inhibit MACF1 function we assessed the effects of downregulating
MACF1 expression on glioblastoma cell proliferation and migration.
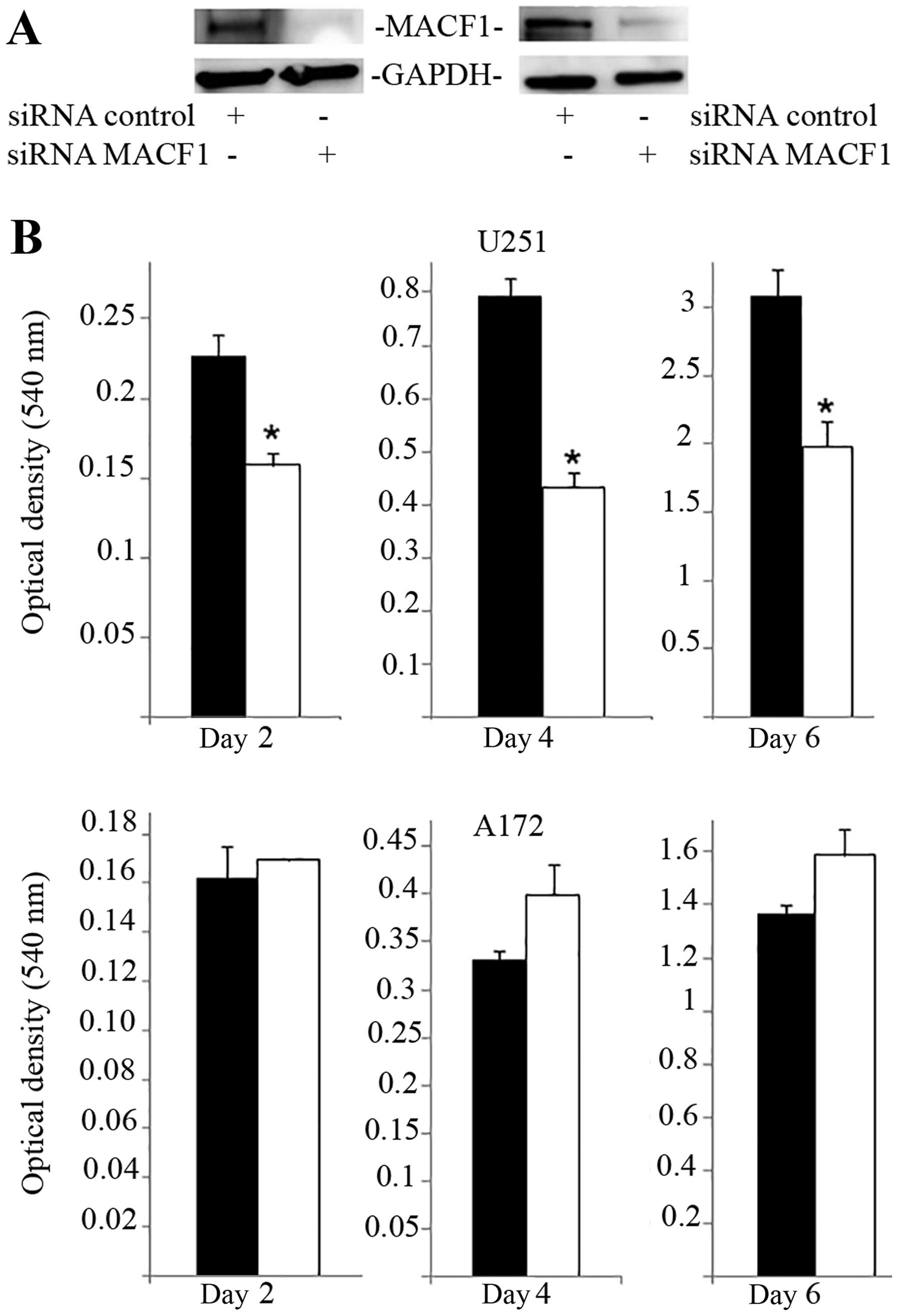
Western blot experiments demonstrated the efficacy of the siRNAs in
silencing MACF1 in U251 and A172 glioblastoma cells. Our data also
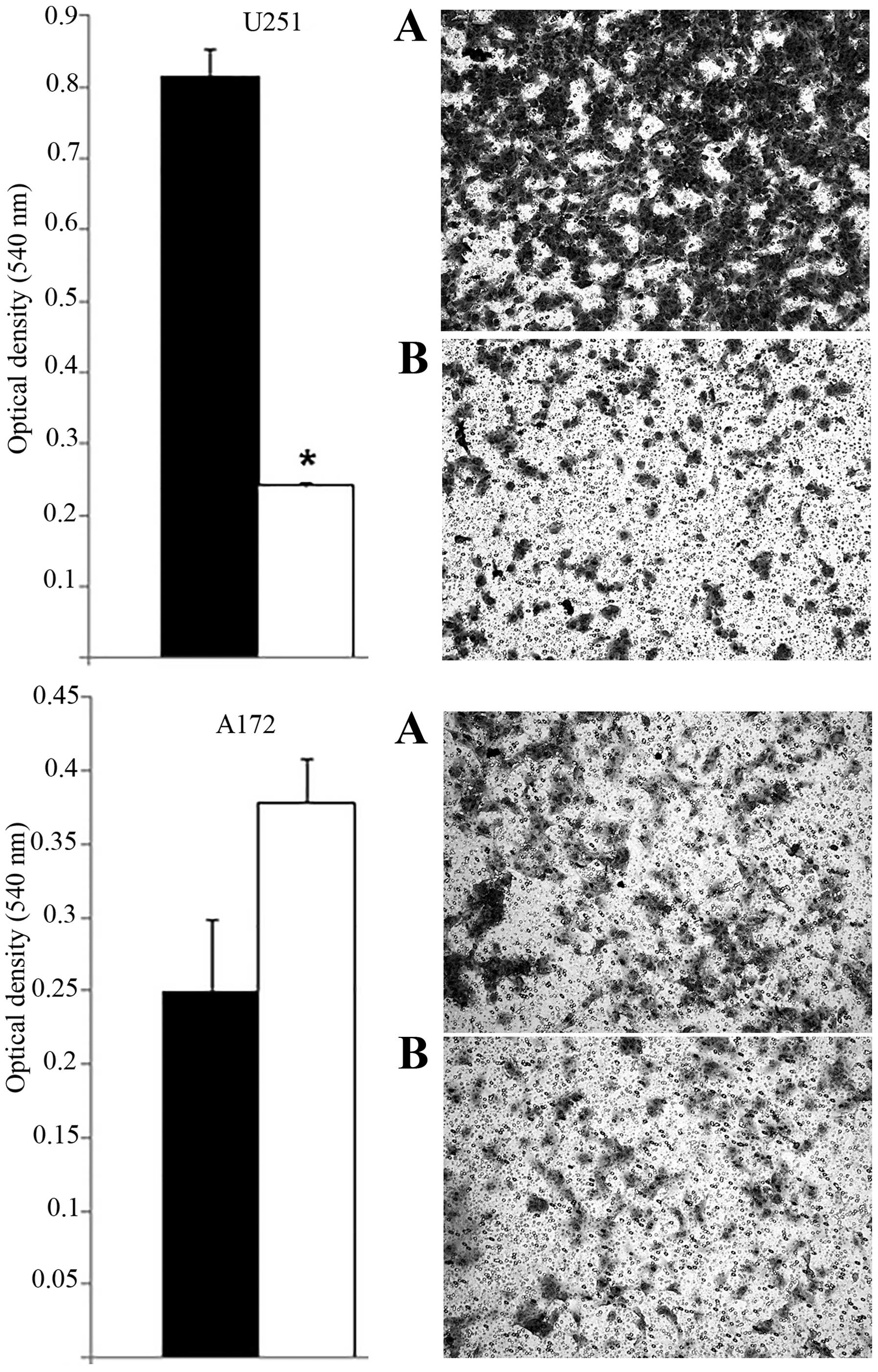
revealed that silencing MACF1 in U251 glioblastoma cells caused a
45 and 37% decrease in cell viability 4 and 6 days post-treatment,
respectively (Fig. 3). However,
suppressing MACF1 expression did not reduce A172 glioblastoma cell
viability (Fig. 3). We further
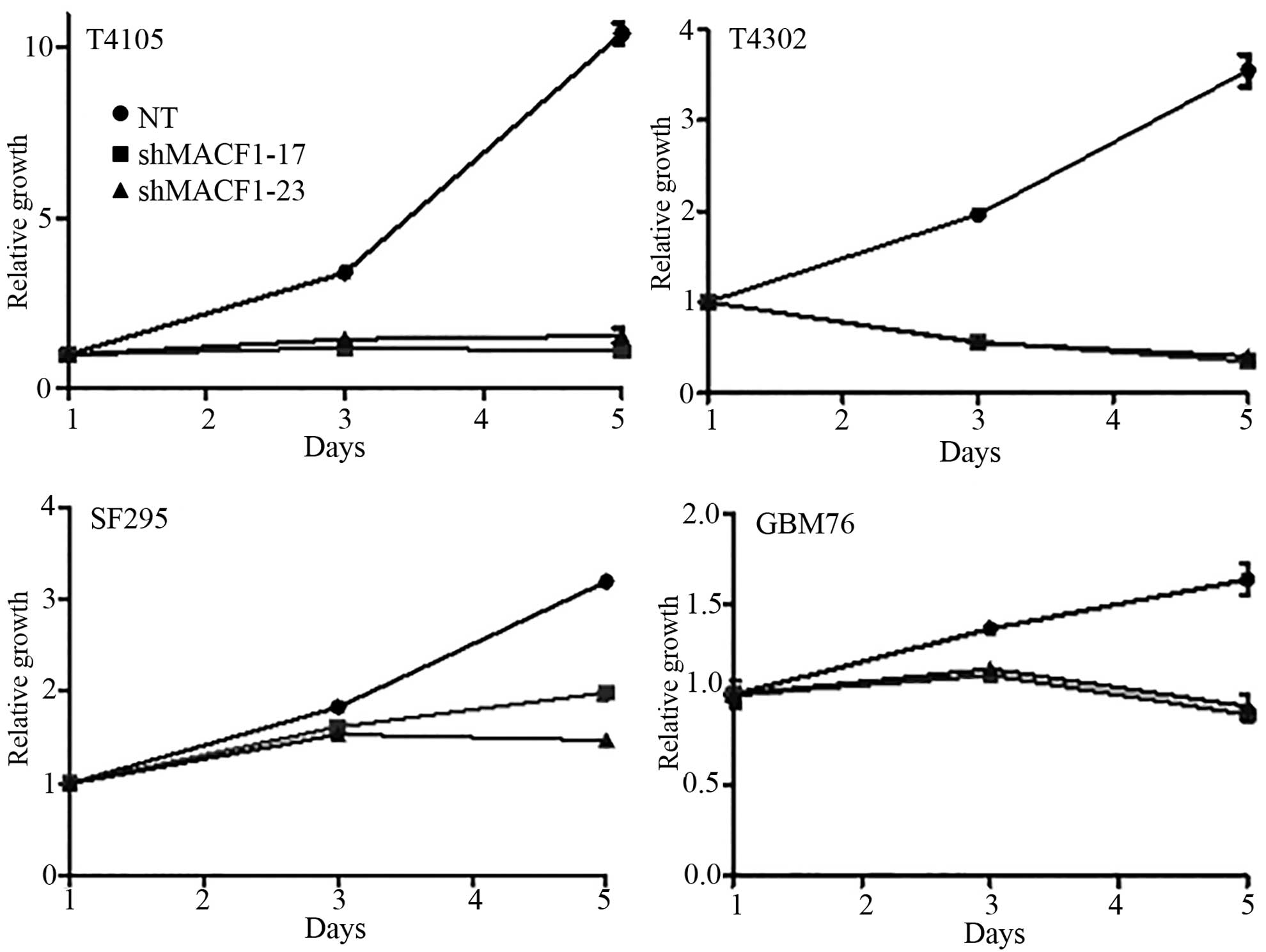
evaluated the effects of inhibiting MACF1 in patient derived
xenograft cell lines (Fig. 4).
Time course analysis displayed that shRNAs targeting MACF1
significantly inhibited the growth of T4105 and T4302 xenograft
cell lines, while SF295 and GBM76 cells were decreased to a much
lesser extent (Fig. 4). Consistent
with the anti-proliferative effects of downregulating MACF1 protein
levels in U251 cells, repression of MACF1 also significantly
reduced U251 cell migration but not A172 cells (Fig. 5). Furthermore, because MACF1 has
been implicated as a modulatory signaling contributor of the Wnt
signaling pathway (16,17), whose dysregulation is well known to
be involved in tumor proliferation and migratory invasion in a
number of human cancers (18–20)
including malignant brain tumors (21–24)
we evaluated the impact of silencing MACF1 on Wnt-signaling
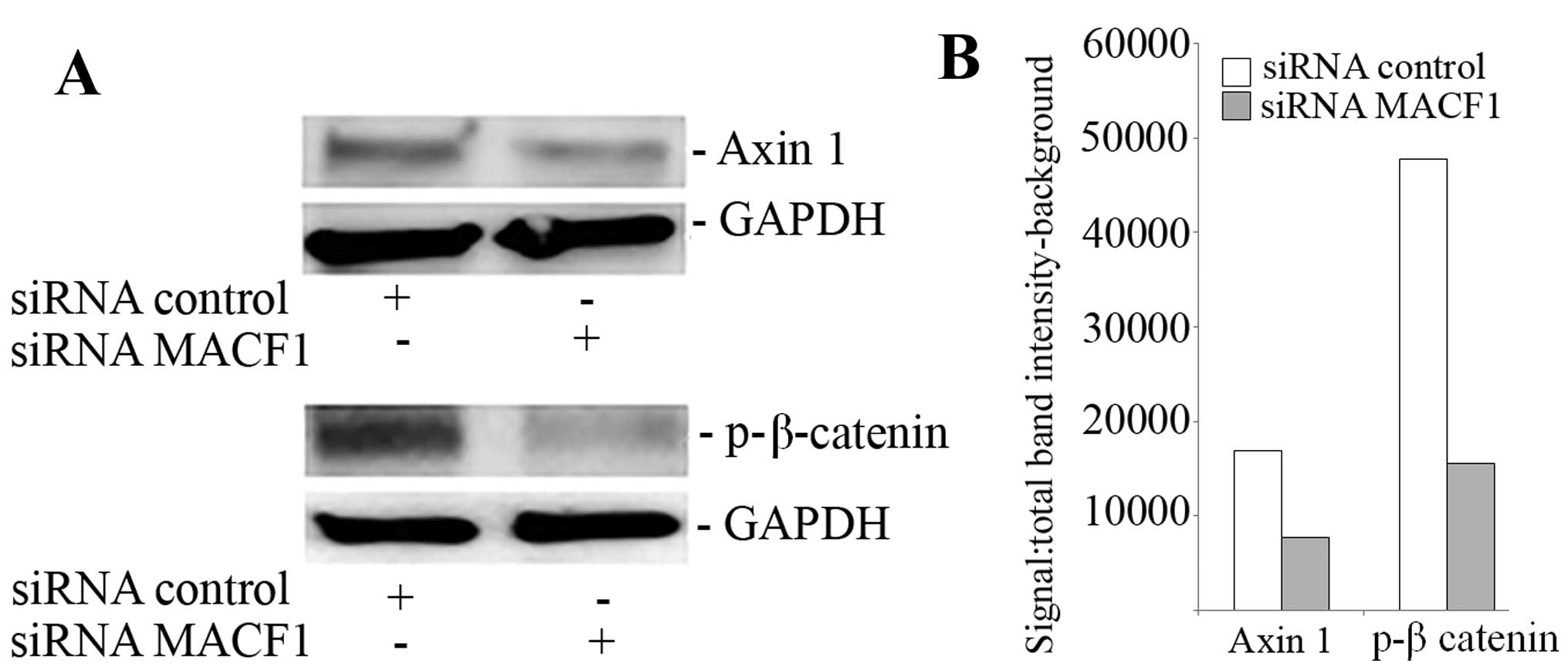
proteins. Our data showed that downregulation of MACF1 protein
levels was accompanied by decreased Axin1 and activated form of
β-catenin protein levels (Fig. 6),
suggesting that reduction of these Wnt-signaling mediators
contribute to the anti-tumorigenic response of impairing MACF1 in
glioblastoma.
Synergistic effects of MACF1 inhibition
and TMZ
Chemotherapeutic agents used to treat glioblastoma
have been minimally effective in part as a consequence of expressed
genetic aberrations that contribute to intrinsic and acquired tumor
resistance (25). We subsequently
performed combinatorial studies to test the effectiveness of
downregulating MACF1 along with the effects of the alkylating
agent, temozolomide (TMZ) in glioblastoma cells. For combinatorial
experiments, glioblastoma cells were treated with siRNAs targeting
MACF1 and 100 µM TMZ and compared to cells treated with TMZ
alone or cells treated with siRNAs targeting MACF1 alone. ANOVA
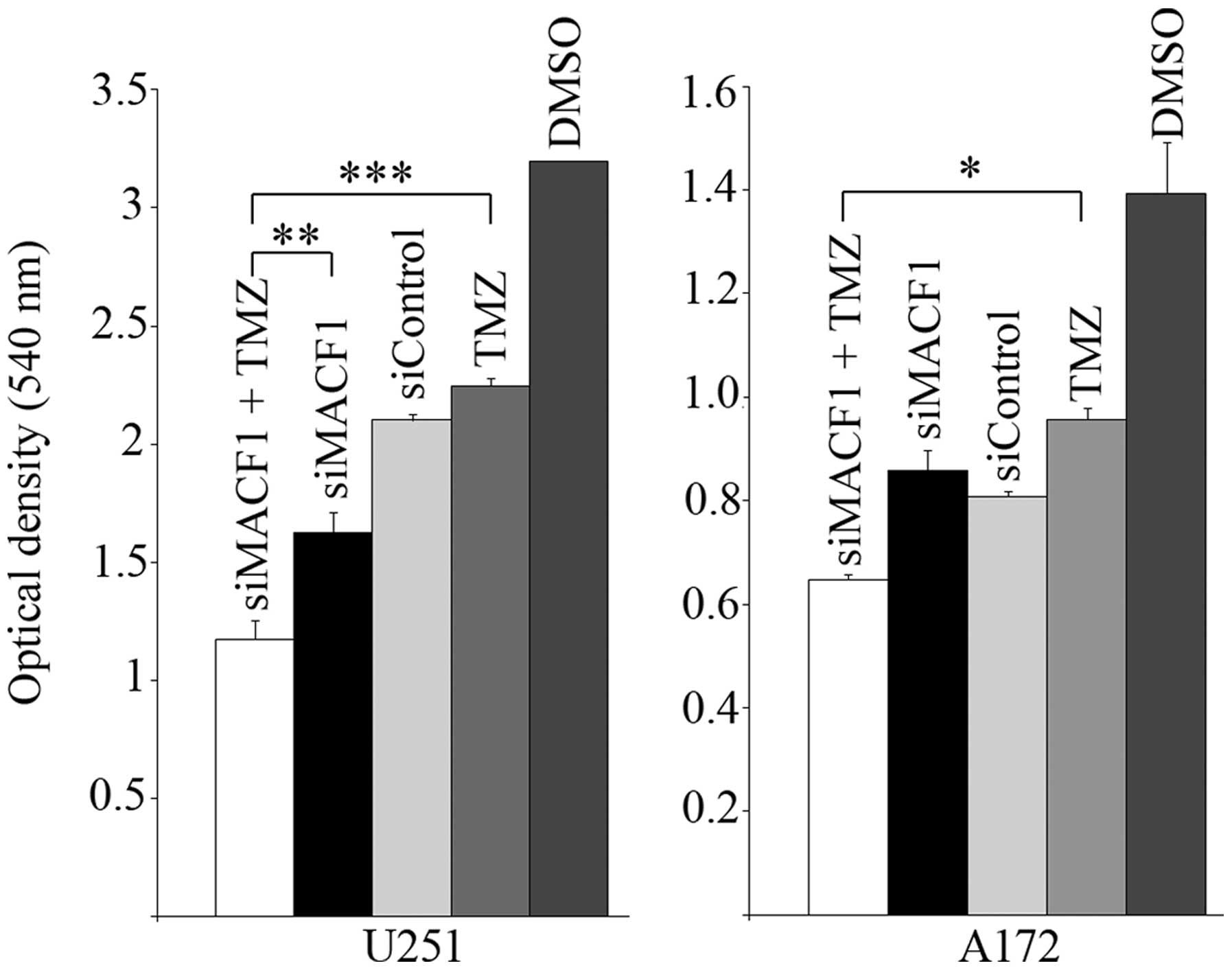
analysis (P<0.01) of data from cell proliferation assays
revealed that the combination of MACF1 and TMZ treatment had the
strongest effect on decreasing glioblastoma cell viability when
compared to other treatment conditions and controls. Additionally,
concurrent treatment of U251 glioblastoma cells with MACF1
targeting siRNAs and TMZ exhibited a 1.5- and 2-fold cell viability
decrease as compared to cells treated with either MACF1 siRNAs or
TMZ alone (Fig. 7). A similar
response was observed in A172 cells, which displayed a 1.32- and
1.48-fold decrease in cell viability when treated concurrently with
TMZ and siRNAs targeting MACF1 as compared to treatment with either
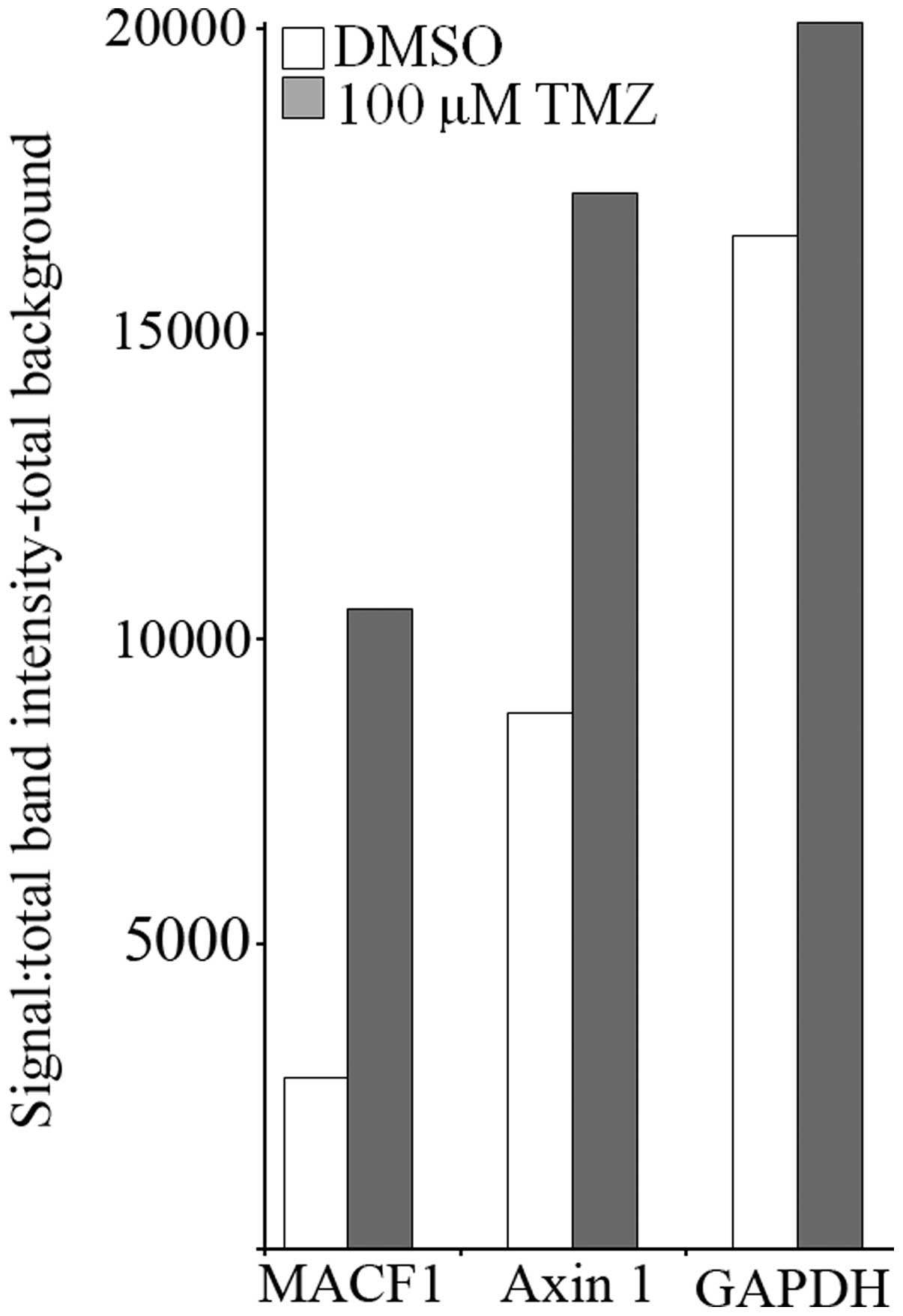
of these conditions alone. We further examined by western blotting
the effects of TMZ on the expression levels of MACF1 and Axin1 in
U251 cells treated for 3 h with 100 µM TMZ. These
experiments showed that TMZ increased the protein levels of MACF1
and the Wnt-cytoplasmic complex protein, Axin1 in U251 cells
(Fig. 8), further supporting the
combinatorial approach and need to inhibit MACF1 in glioblastoma
cells treated with TMZ to achieve a synergistic effect.
Discussion
The scope of genetic abnormalities in the
glioblastoma genome was recently supported in a broad genome
sequence study that revealed 512 mutated genes in U87 glioblastoma
cells, the most commonly studied glioblastoma in vitro cell
line model (26). This study
provided perspective on the instability of the glioblastoma genome
and on the scope of functional studies that must be undertaken to
decipher the roles of uncharacterized genetic aberrations and their
protein products in glioblastoma.
Cytoskeletal proteins play a key role in motility,
invasion and proliferation (27).
In this study we investigated the cytoskeletal spectraplakin
protein, MACF1, as a potential diagnostic and prognostic marker and
have evaluated its potential to be a target for glioblastoma
therapy. Our findings that MACF1 was predominantly expressed in
glioblastoma but absent in normal brain tissue and lower grade
brain tumors suggest that MACF1 is a potential tumor-specific
marker of this grade IV brain tumor. Taken together, our finding
that MACF1 downregulation with RNA interference reduces the
proliferation and migration of glioblastoma cells, indicates that
MACF1 expression positively influences the proliferation and
migration of glioblastoma cells. A similar conclusion was reached
in studies by Hu et al (28) and Wu et al (17) which demonstrated that genetic
inhibition and absence of MACF1 causes a cell cycle arrest
associated decrease in osteoblastic cell proliferation and impaired
keratinocyte migration, respectively. It should also be mentioned
that the role of cell death was not examined as an underlying cause
of the decrease in glioblastoma cell proliferation and viability in
response to negatively regulating MACF1 function in this study,
which is complicated by the established evasiveness of
glioblastomas from apoptotic cell death in response to various
treatment modalities. In this regard forthcoming studies will
examine whether inhibition of MACF1 promotes apoptotic cell death
or autophagy, the preferred mode of cell death in
glioblastomas.
Along with its influence on cell behavior, an early
investigation by Chen et al (16) also revealed that MACF1 was involved
in the Wnt-signaling axis and showed that downregulation of MACF1
decreased nuclear β-catenin and led to an inhibition of Wnt induced
β-catenin-dependent transcriptional activation in P19 and Rat-1
cells. Paralleling these observations, our results demonstrate that
the scaffolding protein Axin1 and the transcriptional activator
β-catenin of the Wnt signaling pathway are decreased in cells with
suppressed MACF1. These data indicate that MACF1 contributes to the
regulation of glioblastoma cell proliferation and migration by
intervening in the Wnt-signaling pathway.
Our results also show that although MACF1 is
expressed at similar levels in U251 and A172 glioblastoma cells,
its silencing impedes motility and proliferation in U251, but not
A172 cells. This could be due to heterogeneous genetic alterations
between these two cell lines, such as p53 which is mutated in U251
and wild-type in A172 or differential expression of MACF1 isoforms.
In support of this hypothesis, glioblastoma cell lines have been
shown to be disparate in their genetic alterations (29). In addition, several MACF1 isoforms
with different 5′UTRs and variations in the N-terminal
actin-binding domain region have been described (30), but their functions have yet to be
deciphered. Furthermore, it has been demonstrated that isoform
specific silencing of the actin-binding protein α-actinin in
glioblastoma cells has been shown to lead to different cellular
outcomes (31), raising the
possibility that differential expression of MACF1 isoforms may
affect glioblastoma cell behavior. To this end, future studies will
investigate the inhibitory effects of silencing different MACF1
isoforms that could impair glioblastoma cell proliferation and
migration, particularly in A172 cells that were impervious to MACF1
inhibition in this study.
Clinically, TMZ is among the lead chemotherapeutic
agents used for the treatment of nervous system cancers,
particularly malignant brain tumors, because of its bioavailability
and limited toxicity (32).
However, a caveat of its efficacy has been intrinsic and acquired
resistance as a result in part of activating pro-survival signaling
mediators (33–35). From a therapeutic precision
medicine standpoint, our data show that in glioblastoma cells in
which MACF1 downregulation decreases proliferation and motility,
reduced MACF1 levels also enhances the anti-proliferative effect of
TMZ. Interestingly, our results also show that TMZ induces the
expression of MACF1 and Axin1, indicating that MACF1-Wnt signaling,
may contribute to TMZ resistance and perpetuate glioblastoma cell
survival. These data are in line with findings that combinatorial
targeted therapy approaches with TMZ is efficient at enhancing the
progression-free survival of patients with genetically diverse
glioblastomas (36–38). Taken together, our results identify
MACF1 as a unique target with the potential to broaden
combinatorial treatment paradigms in glioblastomas with diverse
genetic profiles.
Acknowledgments
The present study was supported by the Tennessee
State University Department of Biology and Division of Research and
Sponsored Programs.
Abbreviations:
|
MACF1
|
microtubule actin cross-linking factor
1
|
|
TMZ
|
temozolomide
|
References
|
1
|
Ostrom QT, Bauchet L, Davis FG, Deltour I,
Fisher JL, Langer CE, Pekmezci M, Schwartzbaum JA, Turner MC, Walsh
KM, et al: The epidemiology of glioma in adults: A 'state of the
science' review. Neuro Oncol. 16:896–913. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Reardon DA and Wen PY: Glioma in 2014:
Unravelling tumour heterogeneity - implications for therapy. Nat
Rev Clin Oncol. 12:69–70. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Parsons DW, Jones S, Zhang X, Lin JC,
Leary RJ, Angenendt P, Mankoo P, Carter H, Siu IM, Gallia GL, et
al: An integrated genomic analysis of human glioblastoma
multiforme. Science. 321:1807–1812. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Verhaak RG, Hoadley KA, Purdom E, Wang V,
Qi Y, Wilkerson MD, Miller CR, Ding L, Golub T, Mesirov JP, et al
Cancer Genome Atlas Research Network: Integrated genomic analysis
identifies clinically relevant subtypes of glioblastoma
characterized by abnormalities in PDGFRA, IDH1, EGFR, and NF1.
Cancer Cell. 17:98–110. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Heimberger AB, Hlatky R, Suki D, Yang D,
Weinberg J, Gilbert M, Sawaya R and Aldape K: Prognostic effect of
epidermal growth factor receptor and EGFRvIII in glioblastoma
multiforme patients. Clin Cancer Res. 11:1462–1466. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pelloski CE, Ballman KV, Furth AF, Zhang
L, Lin E, Sulman EP, Bhat K, McDonald JM, Yung WK, Colman H, et al:
Epidermal growth factor receptor variant III status defines
clinically distinct subtypes of glioblastoma. J Clin Oncol.
25:2288–2294. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hegi ME, Diserens AC, Gorlia T, Hamou MF,
de Tribolet N, Weller M, Kros JM, Hainfellner JA, Mason W, Mariani
L, et al: MGMT gene silencing and benefit from temozolomide in
glioblastoma. N Engl J Med. 352:997–1003. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rivera AL, Pelloski CE, Gilbert MR, Colman
H, De La Cruz C, Sulman EP, Bekele BN and Aldape KD: MGMT promoter
methylation is predictive of response to radiotherapy and
prognostic in the absence of adjuvant alkylating chemotherapy for
glioblastoma. Neuro Oncol. 12:116–121. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sanchez-Soriano N, Travis M,
Dajas-Bailador F, Gonçalves-Pimentel C, Whitmarsh AJ and Prokop A:
Mouse ACF7 and drosophila short stop modulate filopodia formation
and microtubule organisation during neuronal growth. J Cell Sci.
122:2534–2542. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Munemasa Y, Chang CS, Kwong JM, Kyung H,
Kitaoka Y, Caprioli J and Piri N: The neuronal EGF-related gene
Nell2 interacts with Macf1 and supports survival of retinal
ganglion cells after optic nerve injury. PLoS One. 7:e348102012.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bernier G, Pool M, Kilcup M, Alfoldi J, De
Repentigny Y and Kothary R: Acf7 (MACF) is an actin and microtubule
linker protein whose expression predominates in neural, muscle, and
lung development. Dev Dyn. 219:216–225. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sjöblom T, Jones S, Wood LD, Parsons DW,
Lin J, Barber TD, Mandelker D, Leary RJ, Ptak J, Silliman N, et al:
The consensus coding sequences of human breast and colorectal
cancers. Science. 314:268–274. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Misquitta-Ali CM, Cheng E, O'Hanlon D, Liu
N, McGlade CJ, Tsao MS and Blencowe BJ: Global profiling and
molecular characterization of alternative splicing events
misregulated in lung cancer. Mol Cell Biol. 31:138–150. 2011.
View Article : Google Scholar :
|
|
14
|
Carlson BL, Pokorny JL, Schroeder MA and
Sarkaria JN: Establishment, maintenance and in vitro and in vivo
applications of primary human glioblastoma multiforme (GBM)
xenograft models for translational biology studies and drug
discovery. Curr Protoc Pharmacol Chapter. 14:162011.
|
|
15
|
Chiba K, Kawakami K and Tohyama K:
Simultaneous evaluation of cell viability by neutral red, MTT and
crystal violet staining assays of the same cells. Toxicol In Vitro.
12:251–258. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen HJ, Lin CM, Lin CS, Perez-Olle R,
Leung CL and Liem RK: The role of microtubule actin cross-linking
factor 1 (MACF1) in the Wnt signaling pathway. Genes Dev.
20:1933–1945. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wu X, Shen QT, Oristian DS, Lu CP, Zheng
Q, Wang HW and Fuchs E: Skin stem cells orchestrate directional
migration by regulating microtubule-ACF7 connections through GSK3β.
Cell. 144:341–352. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yin X, Xiang T, Li L, Su X, Shu X, Luo X,
Huang J, Yuan Y, Peng W, Oberst M, et al: DACT1, an antagonist to
Wnt/β-catenin signaling, suppresses tumor cell growth and is
frequently silenced in breast cancer. Breast Cancer Res.
15:R232013. View
Article : Google Scholar
|
|
19
|
Narayanan BA, Doudican NA, Park J, Xu D,
Narayanan NK, Dasgupta R and Mazumder A: Antagonistic effect of
small-molecule inhibitors of Wnt/β-catenin in multiple myeloma.
Anticancer Res. 32:4697–4707. 2012.PubMed/NCBI
|
|
20
|
Bakker ER, Das AM, Helvensteijn W, Franken
PF, Swagemakers S, van der Valk MA, ten Hagen TL, Kuipers EJ, van
Veelen W and Smits R: Wnt5a promotes human colon cancer cell
migration and invasion but does not augment intestinal
tumorigenesis in Apc1638N mice. Carcinogenesis. 34:2629–2638. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yu JM, Jun ES and Jung JS, Suh SY, Han JY,
Kim JY, Kim KW and Jung JS: Role of Wnt5a in the proliferation of
human glioblastoma cells. Cancer Lett. 257:172–181. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pu P, Zhang Z, Kang C, Jiang R, Jia Z,
Wang G and Jiang H: Downregulation of Wnt2 and beta-catenin by
siRNA suppresses malignant glioma cell growth. Cancer Gene Ther.
16:351–361. 2009. View Article : Google Scholar
|
|
23
|
De Robertis A, Valensin S, Rossi M, Tunici
P, Verani M, De Rosa A, Giordano C, Varrone M, Nencini A, Pratelli
C, et al: Identification and characterization of a small-molecule
inhibitor of Wnt signaling in glioblastoma cells. Mol Cancer Ther.
12:1180–1189. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kaur N, Chettiar S, Rathod S, Rath P,
Muzumdar D, Shaikh ML and Shiras A: Wnt3a mediated activation of
Wnt/β-catenin signaling promotes tumor progression in glioblastoma.
Mol Cell Neurosci. 54:44–57. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ramirez YP, Weatherbee JL, Wheelhouse RT
and Ross AH: Glioblastoma multiforme therapy and mechanisms of
resistance. Pharmaceuticals (Basel). 6:1475–1506. 2013. View Article : Google Scholar
|
|
26
|
Clark MJ, Homer N, O'Connor BD, Chen Z,
Eskin A, Lee H, Merriman B and Nelson SF: U87MG decoded: The
genomic sequence of a cytogenetically aberrant human cancer cell
line. PLoS Genet. 6:e10008322010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Quick Q, Paul M and Skalli O: Roles and
potential clinical applications of intermediate filament proteins
in brain tumors. Semin Pediatr Neurol. 22:40–48. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hu L, Su P, Li R, Yan K, Chen Z, Shang P
and Qian A: Knockdown of microtubule actin crosslinking factor 1
inhibits cell proliferation in MC3T3-E1 osteoblastic cells. BMB
Rep. 48:583–588. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ishii N, Maier D, Merlo A, Tada M,
Sawamura Y, Diserens AC and Van Meir EG: Frequent co-alterations of
TP53, p16/CDKN2A, p14ARF, PTEN tumor suppressor genes in
human glioma cell lines. Brain Pathol. 9:469–479. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hu L, Su P, Li R, Yin C, Zhang Y, Shang P,
Yang T and Qian A: MACF1, a versatile spectraplakin: Isoforms,
unique structures, and functions. BMB Rep. 49:37–44. 2016.
View Article : Google Scholar :
|
|
31
|
Quick Q and Skalli O: Alpha-actinin 1 and
alpha-actinin 4: Contrasting roles in the survival, motility, and
RhoA signaling of astrocytoma cells. Exp Cell Res. 316:1137–1147.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
von Neubeck C, Seidlitz A, Kitzler HH,
Beuthien-Baumann B and Krause M: Glioblastoma multiforme: Emerging
treatments and stratification markers beyond new drugs. Br J
Radiol. 88:201503542015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Loftus JC, Dhruv H, Tuncali S, Kloss J,
Yang Z, Schumacher CA, Cao B, Williams BO, Eschbacher JM, Ross JT,
et al: TROY (TNFRSF19) promotes glioblastoma survival signaling and
therapeutic resistance. Mol Cancer Res. 11:865–874. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Choi EJ, Cho BJ, Lee DJ, Hwang YH, Chun
SH, Kim HH and Kim IA: Enhanced cytotoxic effect of radiation and
temozolomide in malignant glioma cells: Targeting PI3K-AKT-mTOR
signaling, HSP90 and histone deacetylases. BMC Cancer. 14:172014.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Vo VA, Lee JW, Lee HJ, Chun W, Lim SY and
Kim SS: Inhibition of JNK potentiates temozolomide-induced
cytotoxicity in U87MG glioblastoma cells via suppression of Akt
phosphorylation. Anticancer Res. 34:5509–5515. 2014.PubMed/NCBI
|
|
36
|
Hainsworth JD, Shih KC, Shepard GC,
Tillinghast GW, Brinker BT and Spigel DR: Phase II study of
concurrent radiation therapy, temozolomide, and bevacizumab
followed by bevacizumab/everolimus as first-line treatment for
patients with glioblastoma. Clin Adv Hematol Oncol. 10:240–246.
2012.PubMed/NCBI
|
|
37
|
Chinot OL, Wick W, Mason W, Henriksson R,
Saran F, Nishikawa R, Carpentier AF, Hoang-Xuan K, Kavan P, Cernea
D, et al: Bevacizumab plus radiotherapy-temozolomide for newly
diagnosed glioblastoma. N Engl J Med. 370:709–722. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Clarke JL, Molinaro AM, Phillips JJ,
Butowski NA, Chang SM, Perry A, Costello JF, DeSilva AA, Rabbitt JE
and Prados MD: A single-institution phase II trial of radiation,
temozolomide, erlotinib, and bevacizumab for initial treatment of
glioblastoma. Neuro Oncol. 16:984–990. 2014. View Article : Google Scholar : PubMed/NCBI
|