Introduction
Colorectal carcinoma (CRC) is one of the most common
types of solid malignancies and is a primary cause of
cancer-related mortality worldwide (1). Colorectal carcinogenesis is a complex
multistep process that involves the progressive disruption of
proliferation (2), differentiation
(3), apoptosis (4) and survival mechanisms (5) of intestinal epithelial cells.
Although the survival rates are reasonably good for patients who
are diagnosed with the disease in its very early stages (6), the majority of patients present with
advanced disease. Therefore, chemotherapeutic agents have limited
efficacy in patients who have relapsed and in those who present
with metastatic disease (7).
Therefore, new strategies are needed to improve patient survival
and quality of life in this setting.
With substantial advances in our understanding of
tumour biology, key signalling pathways that are involved in the
mediation of CRC growth and progression have been identified.
Dominant oncogenes and tumour suppressor genes that are involved in
the pathogenesis of CRC have attracted substantial interest
(8), and their central roles and
fundamental contribution to the dysregulation of cancer cells have
been elucidated (9). These genes
offer new targets for biological therapies.
One such target is the v-src avian sarcoma
(Schmidt-Ruppin A-2) viral oncogene homolog (Src) (10), a tyrosine kinase that is frequently
overexpressed in cancer. Expression of this gene leads to the
increased migration and invasiveness of various types of cancer
cells (11,12), including CRC cells (13). Src kinase signalling inhibitor 1
(Srcin1), which is also known as Srcin1 (p140 Cas-associated
protein) (14), contains two
coiled-coil domains, two proline-rich regions and two regions of
highly charged amino acids. According to the characteristic domain
structure, Srcin1 is thought to act as an adaptor protein (15).
Recently, it was determined that Srcin1 negatively
controls tumour cell properties via the inhibition of in
vivo tumour growth and metastasis of human breast cancers
(16,17). This result suggests that Srcin1
functions as a tumour suppressor gene and may slow tumour
progression (18). However, the
manner in which Srcin1 acts at the molecular and cellular levels in
CRC has yet to be elucidated.
Materials and methods
Reagents and antibodies
Sodium butyrate and 5-FU (5-fluorouracil) were
purchased from Sigma-Aldrich (St. Louis, MO, USA). Sodium butyrate
has various effects on cultured mammalian cells including
inhibition of proliferation, induction of differentiation and
induction or repression of gene expression (19). As such, it can be used in lab to
bring about any of these effects. Specifically, butyrate treatment
of cells results in histone hyper acetylation, and butyrate itself
inhibits class I histone deacetylase (HDAC) activity (20), specifically HDAC1, HDAC2, HDAC3 and
HDAC8. Butyrate is an essential vehicle for determining the role of
histone acetylation in chromatin structure and function. Inhibition
of HDAC activity is estimated to affect the expression of only 2%
of mammalian genes (21).
Mouse anti-human Srcin1, cyclin D1, CDK6, cyclin B
and mouse anti-human glyceraldehyde-3-phosphate dehydrogenase
(GAPDH), which were used for western blotting, were purchased from
Santa Cruz Biotechnology (Santa Cruz, CA, USA). Mouse anti-human
Srcin1, which was used for western blotting and/or
immunohistochemistry, was purchased from Novus Biologicals LLC
(Littleton, CO, USA). Goat anti-rabbit immunoglobulins/HRP and
rabbit anti-mouse immunoglobulins/HRP were purchased from Dako
(Carpinteria, CA, USA).
Cell lines, vectors and transfection
Human colorectal carcinoma LS174T, SW620, SW1116,
LoVo, W480, Caco-2, DLD1 and HT29 cell lines were obtained from the
American Type Culture Collection (ATCC; Manassas, VA, USA) and were
cultured in RPMI-1640 medium supplemented with 10% fetal bovine
serum (FBS) and penicillin/streptomycin in a humidified incubator
at 37°C in an atmosphere of 5% CO2. Complementary DNA
(cDNA) that corresponds to full-length Srcin in a pcDNA3.1 plasmid
was obtained by RT-PCR amplification of cDNA from normal human
testis. The clones were digested with NheI and KpnI
(Invitrogen, Carlsbad, CA, USA), and the constructs were
transiently transfected into LoVo cells in the presence of 8
µg/ml polybrene (Sigma-Aldrich) using Lipofectamine 2000
(Invitrogen). Construct expression was then confirmed by western
blotting. For stable selection, 800 µg/ml G418 (Gibco,
Carlsbad, CA, USA) was used.
Tissue microarray (TMA) and
immunohistochemistry (IHC)
A tissue microarray (TMA) containing normal human
tissue samples was purchased from Alenabio, Co., Ltd. (Xian,
China). In accordance with the manufacturer's recommendations, all
tissue samples were fixed in formalin, processed in a tissue
processor (after no more than 24 h of fixation) and finally
embedded in paraffin. The TMA FDA807-1 contains 80 cores (1.5 mm
diameter) of the following 16 normal tissue types (5 cores of each
tissue type): breast, brain, colon, oesophagus, kidney, liver,
lung, ovary, pancreas, prostate, skin, small intestine, stomach,
testis, uterine and rectum. In all, 8 patients with CRC metastatic
lymph nodes and 10 patients with CRC tissues were excised in the
Department of Surgery of Nanfang Hospital, Southern Medical
University. The Ethics Committee of the Southern Medical
University, China approved the experimental protocols.
Histopathological analyses confirmed the malignant and adjacent
normal mucosa tissues.
Western blot analysis
LoVo cell lysates were prepared by homogenization in
RIPA buffer. The protein concentrations were determined with a
bicinchoninic acid protein assay kit (Pierce, Rockford, IL, USA).
The protein lysates were then resolved by SDS-PAGE. The blots were
probed with the appropriate primary antibody followed by a
horseradish peroxidase-conjugated secondary antibody.
Antigen-antibody complexes were visualized by an enhanced
chemiluminescence system (Amersham Biosciences, Ltd., Little
Chalfont, UK).
WST-1 assay
The measurement was based on the ability of viable
cells to cleave the sulfonated tetrazolium salt WST-1
(4-(3-(4-iodophenyl)-2-(4-nitrophenyl)-2H-5-tetrazolio)-1,
3-benzene disulfonate) by mitochondrial dehydrogenases. LoVo cells
(5,000 cells/well) were plated in a 96-well plate in regular growth
medium. After 12, 24 and 48 h 10 µl of WST-1 reagent was
added in each well followed by additional incubation for 1–2 h. The
absorbance at 450 nm was measured using a microplate reader. We
considered OD of vector (or treated) cells at time 12 h as 100%,
and converted OD values by the following calculation: % growth =
[(average OD treated cells/average OD untreated cells) × 100].
Migration and Matrigel invasion
assays
Cell migration was analysed by a conventional
wound-healing assay, as previously described (22). Briefly, Srcin1-siRNA or
scrambled-siRNA (Scr-siRNA) stably infected cells were grown in
6-well plates to confluence. Wounds were generated using the tip of
a sterile micropipette, and the detached cells were removed by
washing with PBS. The medium was then refreshed to remove cell
debris, and the cells were cultured in the presence of 10
µg/ml mitomycin C to inhibit cell proliferation. Then, the
cells were incubated and allowed to migrate for up to 48 h. Images
were captured, and the migration index was calculated as follows:
migration index = [(initial wound width − width of wound at
time-point tested)/initial wound width] × 100%. A Matrigel invasion
assay was performed using Matrigel invasion chambers (BD
Biosciences) according to the manufacturer's instructions. Matrigel
membranes were rehydrated for 2 h in RPMI-1640 medium, and all
inserts were then placed in wells that contained 10% FBS in
RPMI-1640. For each experiment, a total of 5×104 cells
suspended in 0.5 ml of serum-free RPMI-1640 was added to the top of
each of three chambers and incubated at 37°C in 5% CO2.
After 24 h, the chambers were washed with phosphate-buffered saline
(PBS), and the cells were removed from the top of the membranes
with a cotton swab. Invading cells were fixed in 4%
paraformaldehyde for 30 min and stained with 2% crystal violet. In
each chamber, five representative fields were documented by
photo-microscopy and the number of invading cells was counted. The
average number of invading cells in each chamber in the triplicate
samples is presented.
Flow cytometric analysis
To analyse the cell cycle, the cells were collected
and fixed in ice-cold 70% ethanol in PBS and stored at −4°C until
use. After resuspension, the cells were incubated with 100
µl of RNase I (1 g/ml) and 100 µl of propidium iodide
(400 µg/ml) at 37°C and were then analysed by flow cytometry
(FCM; BD Biosciences, San Jose, CA, USA). The cell cycle phase
distribution was calculated from the resultant DNA histogram using
MultiCycle AV software (Phoenix Flow Systems, San Diego, CA, USA).
Apoptosis was detected with an Annexin V-FITC kit according to the
manufacturer's instructions (Trevigen, Inc., Gaithersburg, MD,
USA). Briefly, the cells that received various treatments were
collected and stained with Annexin V-FITC and PI in the dark for 15
min at room temperature. After the addition of binding buffer, the
cells were subjected to flow cytometry and analysed using WinMDI
2.9.
Immunofluorescence and the morphological
detection of apoptosis
For the staining of F-actin, the cells were fixed in
3.7% formaldehyde and incubated with rhodamine-conjugated
phallotoxin (5 U/ml; Molecular Probes, Eugene, OR, USA) in PBS at
room temperature. The coverslips that contained the cells were
washed, mounted and visualized.
A morphological evaluation of apoptotic cell death
was performed as previously described, with some modifications. The
cells were fixed for 5 min in 3% paraformaldehyde in PBS. After the
cells were air-dried, they were stained for 10 min in Hoechst 33258
(10 µg/ml), mounted in 50% glycerol containing 20 mM citric
acid and 50 mM orthophosphate, and stored at -20°C. Prior to
analysis, nuclear morphology was evaluated using a Zeiss IM 35
fluorescence microscope. Illustration of apoptotic cells after
staining with Hoechst 33258. The rate of apoptotic cells was
caculated by the average value of apoptotic cells in 100 cells in
three different fields.
Caspase-3, -8 and -9 activity assay
The activity of caspases-3, -8 and 9 was determined
using the ApoAlert caspase colorimetric assay kit according to the
manufacturer's instructions (One unit is the amount of enzyme that
will cleave 1.0 nmol of the colorimetric substrate Ac-DEVDpNA per
hour at 37°C under saturated substrate concentrations. Both the
concentrations of pNA and the value of OD405 are proportional)
(Clontech Laboratories Inc., Mountain View, CA, USA). Briefly, the
assays were performed in 96-well microtiter plates; 10 µ of
protein from cell lysates of each sample was incubated in 80
µl of the reaction buffer [1% NP-40, 20 mmol/l Tris-HCl (pH
7.5), 137 mmol/l Nad, and 10% glycerol] containing 10 µl of
caspase-3, -8 or -9 substrate (Ac-DEVDpNA; 2 mmol/l). The cell
lysates were incubated at 37°C for 4 h. The samples were then
measured by an enzyme-linked immunosorbent assay (ELISA) reader at
an absorbance of 405 nm. All experiments were performed at least 3
times.
Transient transfection and
Dual-luciferase reporter assay
The reporter constructs pTOPFLASH and pFOPFLASH, in
which the luciferase gene was driven by wild-type or
dominant-negative mutant consensus 3x T-cell factor/lymphoid
enhancer factor binding element (TBE), respectively, were kindly
provided by Professor J. Schneikert of the University
Erlangen-Nürnberg, Germany. Transient transfection was performed
with Lipofectamine 2000 reagent (Invitrogen) according to the
manufacturer's instructions. The firefly and Renilla
luciferase activities were measured using the Dual-luciferase
reporter assay kit (Promega, Madison, WI, USA) with a luminometer
(Lumat LB 9507; Berthold Technologies GmbH, Bad Wildbad,
Germany).
Construction and transfection of
lentiviral vectors with Srcin1 short hairpin RNA
To investigate the effect of small interfering RNA
(siRNA)-induced downregulation of Srcin1 expression on tumour
growth in vivo, a Srcin1 RNAi lentiviral vector
(pGCSIL-Srcin1 shRNA) was constructed (Shanghai GeneChem, Co., Ltd,
Shanghai, China). Double-stranded oligonucleotides encoding human
Srcin1-vshRNA
CCGGAAGCTGTGTCTGTTGAGGCTGTCAAGAGCAGCCTCAACAGACACAGCTTTTTTTG) were
inserted into the short hairpin RNA (shRNA) expression vector
pGCSIL. A scrambled sequence that shared no homology with the
mammalian genome was used as a control. The successful cloning of
these sequences into the pGCSIL-GFP lentivector was confirmed by
Sanger sequencing. The expression of shRNA was driven by a U6
promoter. This vector also encoded green fluorescent protein (GFP).
The recombinant lentiviral vector was produced by co-transfection
of HEK293FT cells with the lentiviral expression vector and the
packing plasmid mix using Lipofectamine 2000 according to the
manufacturer's instructions. LoVo cells were then transduced at a
multiplicity of infection (MOI) of 5.
In vivo tumourigenesis
Female BALB/c nude mice (Laboratory Animal Unit,
Southern Medical University, Guangzhou, China) were housed in a
specific pathogen-free (SPF) environment. In all, 5×106
LoVo cells were subcutaneously injected into twenty 4-week-old male
nude mice (five mice per group). Seven days after subcutaneous
tumour cell injection, mice were anesthetized and
1.5×1011 v.g. of lenti-Src shRNA, lenti-Srcin1 shRNA
lenti-Src shRNA + 5-FU, lenti-Srcin1 shRNA + 5-FU was injected
(23). The size of the tumours was
measured every 7 days using a microcaliper. The volumes of the
tumours were calculated as follows: V = (4/3)
R12R2, where R1 is radius 1, R2 is radius 2,
and R1<R2 (the smallest and the largest tumours were deleted).
The mice were sacrificed by CO2 euthanasia 30 days after
the injection. All tumours were removed and dissociated. The
Committee on the Use of Live Animals in Teaching and Research of
Southern Medical University approved the protocol.
Statistical analyses
The results are representative of at least three
independent experiments that were performed in triplicate and are
expressed as the mean ± SD. A statistical analysis of the data was
performed using Student's t-test. A value of P<0.05 was
considered significant.
Results
Srcin1 expression is significantly
increased in human normal tissues, CRC and metastatic lymph node
tissues
Of the 16 normal tissue types (5 cores of each
tissue type) that were tested for Srcin1 expression in this TMA,
cytoplasmic immunoreactivity was observed in 100% (all 5 cores) of
the breast, cerebrum, liver and skin samples. Strong cytoplasmic
immunoreactivity was observed in 80% of the kidney, testis and
stomach samples; cytoplasmic immunoreactivity was observed in 60%
of the lung and 20% of the pancreas samples (Fig. 1). Cells of the esophagus, ovary,
prostate and uterus were negative. Among them, 80% (4/5) of cores
that represented normal colon and rectal tissues were negative
(Fig. 1). The paraffin tissue
sections of normal colorectal mucosa showed either negative or weak
staining for Srcin1 protein. However, the expression of Srcin1
protein, as exemplified in two patients by IHC, was predominantly
localized within the cytoplasm of colorectal cancer cells (21/25,
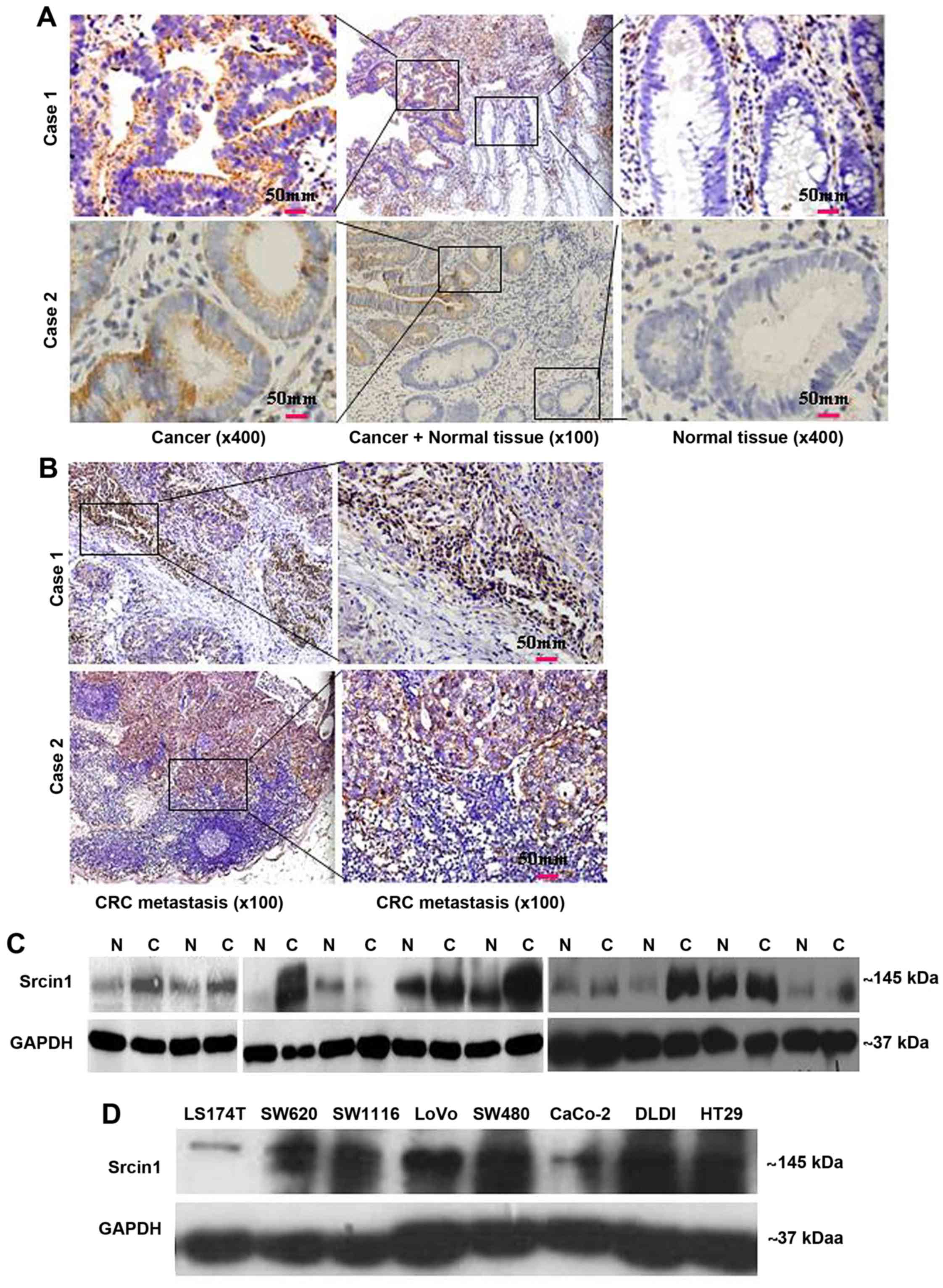
84% of the cases) (Fig. 2A). In
addition, all of CRC metastatic lymph nodes were positive for
Srcin1 protein (8/8, 100% of the cases) (Fig. 2B).
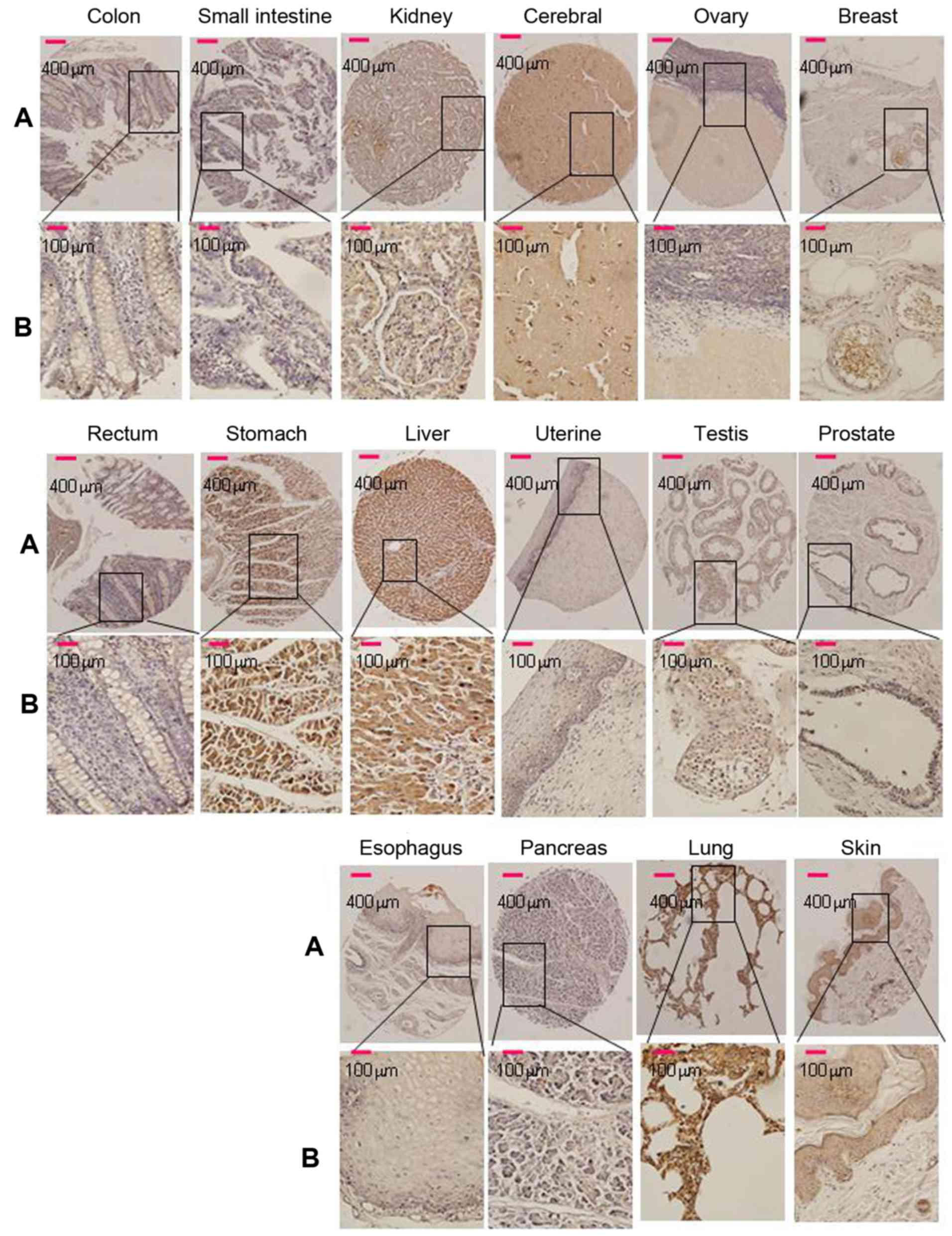 | Figure 1Srcin1 expression in 16 normal human
tissues was detected by immunohistochemistry. Breast, brain, colon,
oesophagus, kidney, liver, lung, ovary, pancreas, prostate, skin,
small intestine, stomach, testis, uterine and rectum, of 16 normal
human tissues stained with anti-Srcin1 antibody by means of TMA.
Original magnification, A, ×100 and B, ×400. |
We then measured Srcin1 expression in 10 pairs of
matched normal colon (N) and cancerous colon (T) tissues by western
blotting. Of the 10 cancerous tissue specimens, 7 expressed higher
levels of Srcin1 than normal tissues (7/10, 70% of the cases)
(Fig. 2C). Next, Srcin1 protein
was observed in the colorectal cells lines LS174T, SW1116, SW620,
LoVo, SW480, CaCo, DLD1 and HT29 by western blotting (Fig. 2D).
Thus, a high level of Srcin1 protein expression
correlates with a transformed phenotype, which indicates that
Srcin1 may play a role in CRC tumourigenesis.
Suppression with siRNA induces cell
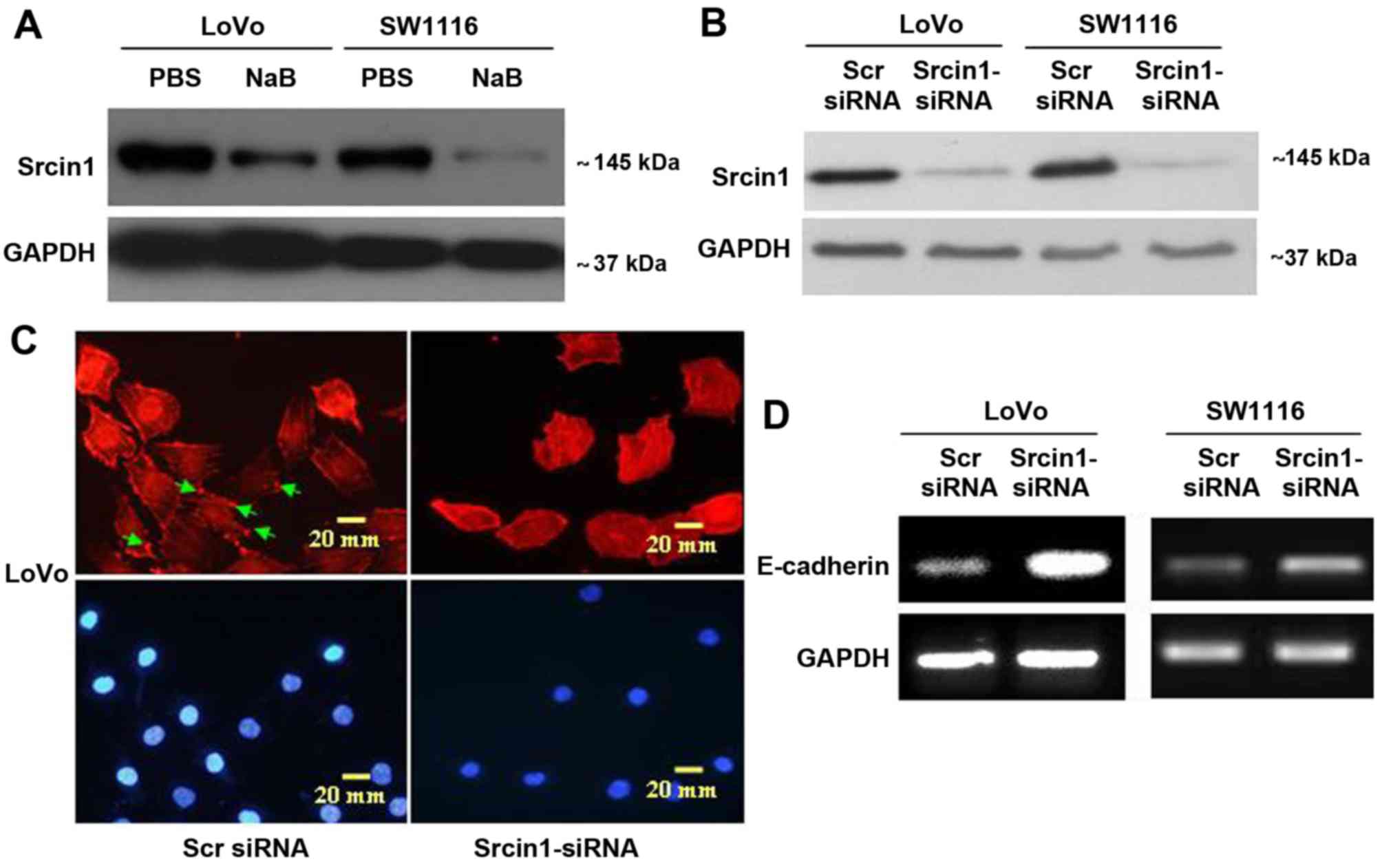
differentiation in CRC cells
To examine the effect of Srcin1 on cell
differentiation (Fig. 3A), we
synthesized Srcin1-siRNA, the expression of which was confirmed by
western blotting (Fig. 3B) in both
CRC cell lines (LoVo and SW1116). We first stained for F-actin in
Srcin-suppressed cells using phalloidin-conjugated rhodamine and
immunofluorescence microscopy. Diffuse and generally uniform
staining was observed throughout the cytoplasm in
Srcin1-siRNA-transfected LoVo and SW1116 cells. However, after Scr
siRNA transfection, F-actin was present throughout the cytoplasm
and at the rim zone of the protrusion (arrow), an area that
features actin polymerization (Fig.
3C).
Furthermore, we showed by reverse-transcription
polymerase chain reaction (RT-PCR) that the knockdown of Srcin1
increased the expression of differentiation marker E-cadherin in
both LoVo and SW1116 cells (Fig.
3D). This result suggests that the knockdown of Srcin1 might
induce the differentiation of CRC cells.
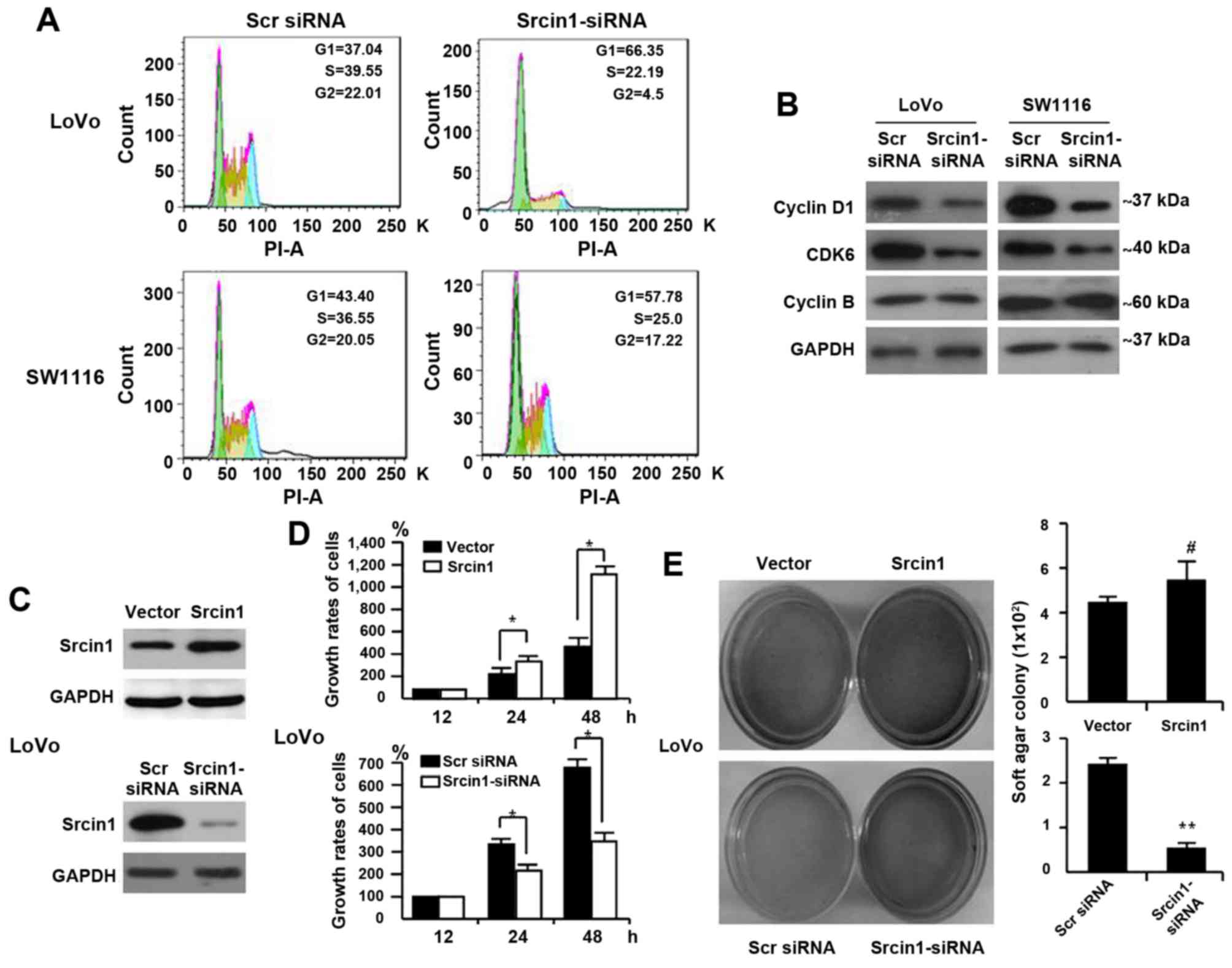
The impact of Srcin1 on cell cycle arrest
in human CRC cells
We performed flow cytometric analyses to detect
whether Srcin1 could change the cell cycle distribution. As shown
in Fig. 4A, Srcin1 knockdown
resulted in an increase in cells in the G0/G1-phase and a
concomitant decrease in cells in the S-phase and G2/M-phase of the
cell cycle.
Furthermore, cell cycle-related protein expression
was assessed by western blotting. Consistent with the accumulation
of cells in G0/G1 phase, the expression of cyclin D1 and CDK6 was
significantly decreased in Srcin1-siRNA-transfected LoVo and SW1116
cells, whereas the expression of cyclin B remained unchanged
compared with scrambled-siRNA-transfected cells (Fig. 4B).
Together, these results indicate that Srcin1
silencing in CRC is associated with a block in cell cycle
progression.
Effect of Srcin1 on the proliferation of
CRC cells
To demonstrate the effect of Srcin1 on cell growth,
we first detected Srcin1 expression in the CRC cell line LoVo using
a WST-1 assay. We then established stable transfectants of LoVo
cells that expressed the vector pcDNA3.1, pcDNA3.1-Srcin1 and Scr
siRNA and Srcin1-siRNA were confirmed by immunoblotting (Fig. 4C). We found that the growth rates
of the vector-transfected cells were 100%, 260.80±9.33 and
477.84±35.09% at 12, 24 and 48 h, respectively, whereas those of
Srcin1-expressing cells were 100%, 371.48±13.87 and
1168.31±176.82%, respectively (Fig.
4D). A significant difference between Srcin1- and the
vector-transfected cells was found at two time-points (24 and 48
h).
Moreover, we assessed the effect of Srcin1
repression on CRC cell proliferation. We showed that the growth
rates of Scr-siRNA-transfected cells were 100%, 357.47±39.76 and
673.97±72.43% at 12, 24 and 48 h, respectively, whereas
Srcin1-siRNA-transfected cells were 100%, 210.98±15.68 and
332.70.97±24.79% at 12, 24 and 48 h, respectively (Fig. 4D). A significant difference between
the Srcin1-siRNA- and Scr-siRNA-transfected cells was found at two
time-points (24 and 48 h). Srcin1 overexpression significantly
inhibited cancer cell growth (P<0.05).
In addition, we examined the anchorage-independent
growth of the stable transfectants (empty vector and Srcin1) in
soft agar after 12 days. As expected, LoVo empty vector cells
formed colonies in soft agar, and the forced expression of Srcin1
in LoVo cells markedly enhanced colony formation in soft agar
(Fig. 4E). In contrast, we tested
the effect of RNAi-mediated repression of Srcin1 in LoVo cells. The
application of Srcin1-siRNA significantly inhibited the ability of
these cells to form colonies (Fig.
4E), which indicates that Srcin1 downregulation suppressed the
proliferation of colon cancer cells.
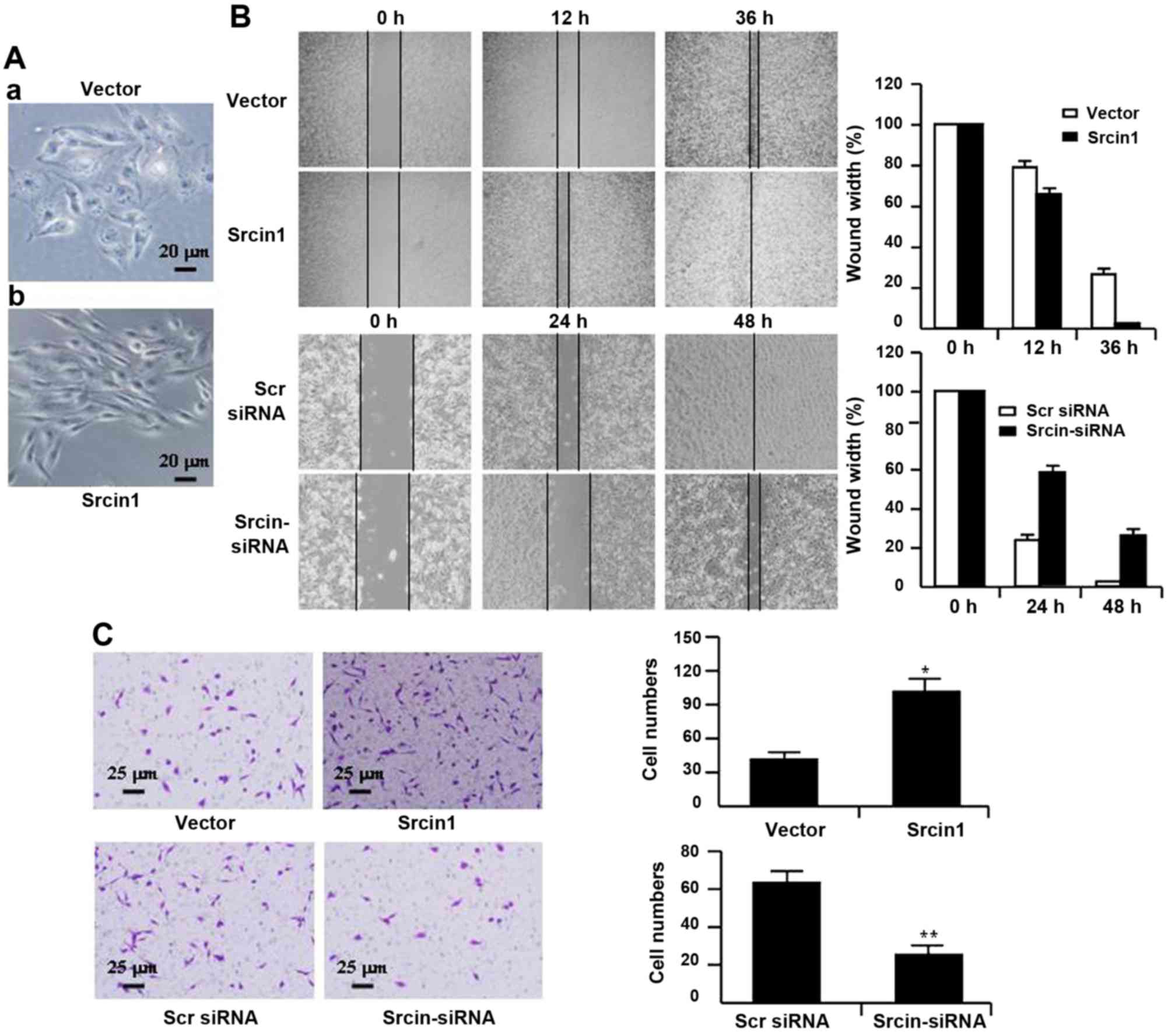
The role of Srcin1 in CRC cell metastasis
and invasion
To characterize Srcin1 transfectants, we first
examined the morphologic features of these cells. The parental
cells (i.e., cells that were stably transfected with vector)
displayed a flat morphology with a short cytoplasmic process
(Fig. 5A–a). However, the cells
that were stably transfected with Srcin1 demonstrated diffuse
Srcin1 expression in the cytoplasm and had an elongated or
shuttle-shape morphology with a loss of cell-cell contacts. In
addition, increased cell scattering was observed according to
phase-contrast microscopy, which may be associated with an EMT-like
conversion (Fig. 5A–b) (24).
Therefore, we studied the effect of Srcin1 on the
metastatic and invasive potential of CRC-derived LoVo cells. Cell
migration was determined using a wound-healing assay. As shown in
Fig. 5B, Srcin1 upregulation
significantly increased LoVo cell migration, whereas Srcin1
downregulation significantly suppressed migration. The migration
index of Srcin1 transfectants was increased by 12 and 24% at 12 and
36 h, respectively. In contrast, the migration index of Srcin1
knockdown cells was decreased by 30 and 28% at 24 and 48 h,
respectively (Fig. 5B).
To examine cell invasion in vitro, we used
Transwell inserts coated with Matrigel matrix. The invasiveness of
LoVo cells that were transfected with Srcin1 increased by 110.6%
compared with the control cells. In contrast, after Srcin1 was
knocked down, the invasiveness of LoVo cells decreased by 269.5%
compared with the Scr siRNA-transfected cells (Fig. 5C).
These data indicate that the role of Srcin1 in
invasion and metastasis are associated with an increase in the
metastatic potential of cancer cells.
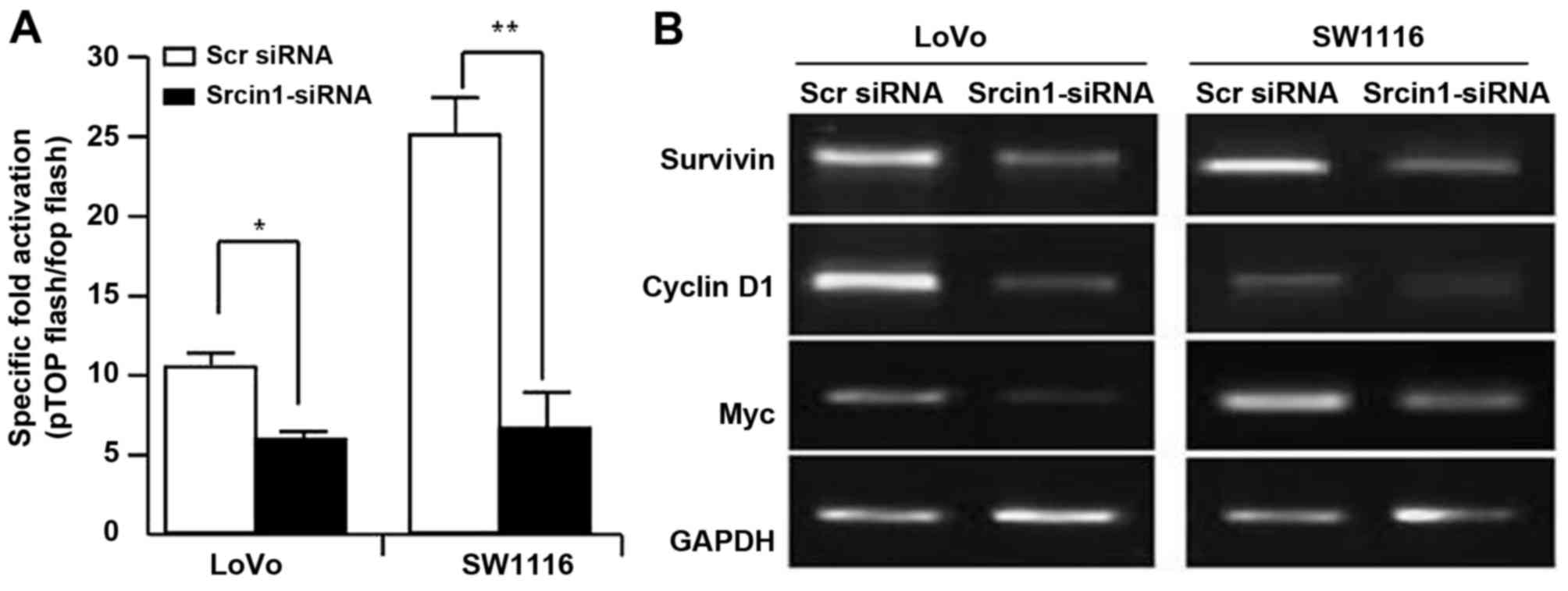
Srcin1 induces the activation of the
Wnt/β-catenin signalling pathway
The Wnt/β-catenin signalling pathway governs cell
proliferation, differentiation, migration and invasion in CRC
(25,26). We assessed the effect of Srcin1 on
the trans-activation of β-catenin. pTOPFLASH and pFOPFLASH,
contained the wild-type and dominant-negative TBE binding element,
respectively. We knocked down the expression of Srcin1 in LoVo and
SW1116 cells using siRNA, and the luciferase reporter assay was
then used. As shown in Fig. 5A,
Srcin1-siRNA decreased the expression of Srcin1 compared with Scr
siRNA, and additionally, the TOP-Flash plasmid, but not the
FOP-Flash plasmid, was downregulated by Srcin1-siRNA, but not by
Scr siRNA (Fig. 6A).
To further investigate whether Srcin1-siRNA
regulated the transcriptional activity of Wnt/β-catenin signalling,
we examined the expression of survivin, c-Myc and cyclin D1.
Indeed, the knockdown of Srcin1 by shRNA also inhibited the
expression of survivin, c-Myc and cyclin D1 (Fig. 6B). These results indicate that
Srcin1 may directly or indirectly regulate Wnt/β-catenin signalling
in CRC.
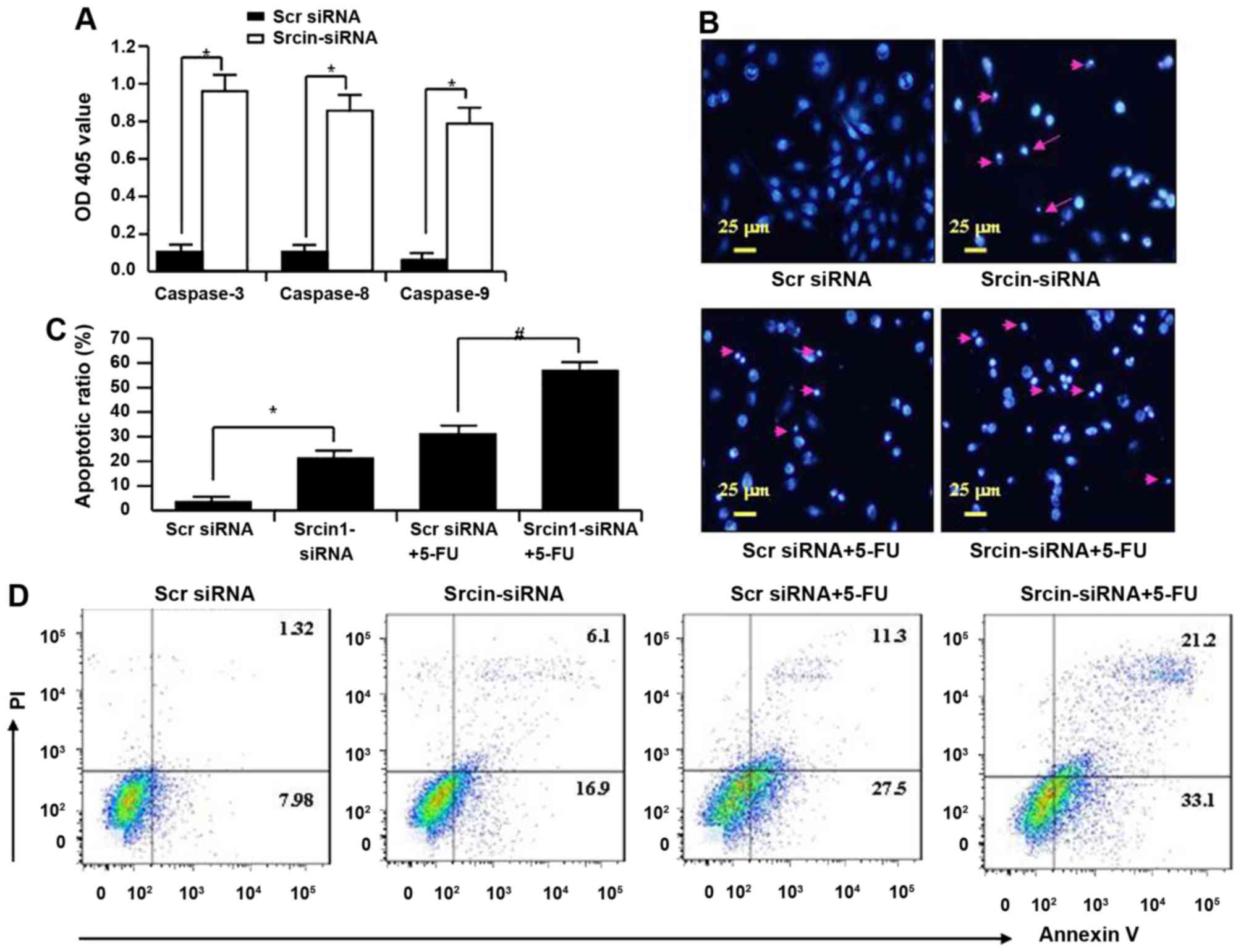
Srcin1-specific silencing induces
apoptosis and sensitizes cancer cells to the chemotherapeutic
5-FU
To investigate the mechanism of knockdown of
Srcin1-induced growth suppression, apoptosis was assayed by ELISA.
The activities of caspase-3, -8 and -9 were also analysed. The
activities of caspase-3, -8 and -9 were significantly increased in
Srcin1-siRNA-transfected cells compared with Src siRNA-transfected
cells (Fig. 7A).
Apoptotic induction was further confirmed by DAPI
staining at the single-cell level. Bright blue-fluorescent
condensed nuclei and chromatin fragmentation were considered
indicative of apoptosis by fluorescence microscopy (Fig. 7B). Similarly, the ratios of cells
with condensed nuclei were higher in Srcin1-siRNA-transfected LoVo
cells compared with the Src siRNA-transfected LoVo cells (P<0.05
compared with Src siRNA; Fig.
7B).
After an evaluation of the role of Srcin1-siRNA in
chemotherapy-induced apoptosis, the ratios of cells with condensed
nuclei were higher in Srcin1 + 5-FU-treated LoVo cells compared
with the Src siRNA + 5-FU-treated controls (P<0.05; Fig. 7C).
To validate the results of DAPI staining, Src
siRNA-transfected cells were treated with 5-FU for 48 h; the cells
were then stained with Annexin V and PI and analysed by flow
cytometry. As shown in Fig. 7D,
the apoptotic index of Srcin1-siRNA + 5-FU-treated cells was
significantly increased relative to Src siRNA-transfected
cells.
These findings suggest that Srcin1-siRNA enhances
the susceptibility of cancer cells to apoptotic triggers induced by
5-FU.
Suppression of Srcin1 sensitizes cancer
cells to 5-FU-induced apoptosis in vivo
Finally, we investigated the potential utility of
lenti-Srcin1 shRNA as a therapy for established tumours.
Lentivirus-transduced LoVo cells were subcutaneously injected into
the right dorsal flank of nude mice, some of which were treated
with 5-FU. When the tumour nodules became visible (~3–5 mm in
diameter), lenti-Src shRNA or lenti-Srcin1 shRNA was injected
directly into the tumour; 5-FU was intraperitoneally injected as a
co-treatment. The tumour sizes were continuously monitored on a
weekly basis. As shown in Fig. 8A and
B, the tumour volumes of the lenti-Srcin1 shRNA-injected mice
were markedly smaller than those of the lenti-Src shRNA-injected
mice. Similar results were obtained in the 5-FU-treated (lenti-Scr
shRNA + 5-FU or lenti-Srcin1 shRNA + 5-FU) mice compared with
untreated (lenti-Scr shRNA or lenti-Srcin1 shRNA) mice (Fig. 8A and B). Importantly, mice that
were injected with both lenti-Src shRNA and 5-FU presented with the
smallest tumour nodules (Fig.
8C).
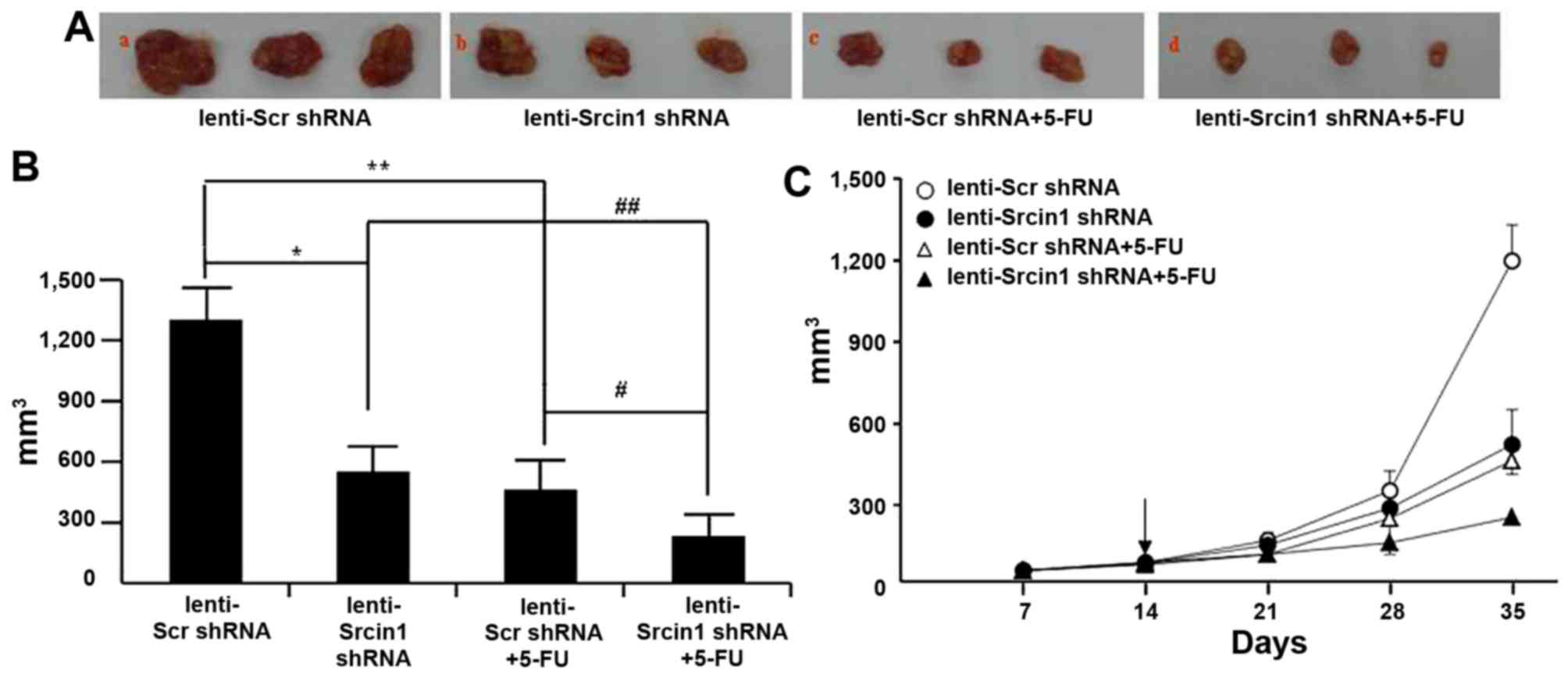 | Figure 8Suppression of Srcin1 with shRNA
sensitized cancer cells to 5-FU-induced apoptosis in vivo.
(A) LoVo cells (5×106) were injected subcutaneously in
the right flanks of the nude mice. When the tumor nodules became
visible, 5-FU (50 µg/kg, once per two days, 4 times) or NS
were administered by intraperitoneal injection. Images shown were
taken on day 35. (B) Tumor growth was monitored in 3 dimensions and
expressed as tumor volume in cubic millimeters. Data are the pooled
average ± standard error of mean of the tumor volumes for each of 3
animals per group. *P<0.05, lenti-Scr shRNA vs.
lenti-Srcin1 shRNA, #P<0.05, lenti-Scr shRNA + 5-FU
vs. lenti-Srcin1 shRNA+ 5-FU, **P<0.05, lenti-Scr
shRNA vs. lenti-Scr shRNA + 5-FU, ##P<0.05,
lenti-Srcin1 shRNA vs. lenti-Srcin1 shRNA+ 5-FU. (C) Tumor size was
measured weekly after tumor cell inoculation in each group. The
arrow indicates the time when intratumoral and intravenous
injections were performed. |
These results demonstrate that targeting with
lenti-Srcin1 shRNA has an inhibitory effect on tumorigenesis in
vivo. Furthermore, inhibition of Srcin1 siRNA has a synergistic
effect with 5-FU treatment in the treatment of colorectal
cancer.
Discussion
In the present study, we describe a novel role of
Srcin1 in the regulation of the pathogenesis and progression of
CRC. The evidence that supports our conclusions is as follows: i)
by western blotting and IHC, CRC cells expressed higher levels of
Srcin1 than normal tissues; CRC metastatic lymph nodes were
positive for Srcin1 protein by IHC; ii) RNAi-mediated silencing of
Srcin1 in LoVo cells suppressed the proliferation, metastasis and
invasiveness of these cells; and iii) lentivirus-Srcin1-shRNA
increased cell susceptibility to apoptotic stimuli by 5-FU in
vitro and in vivo. Together, these findings provide
strong evidence for the oncogenic activity of Srcin1 in CRC.
Despite the high expression of Srcin1 in normal
human breast tissues, as reported in previous studies (15,27),
Srcin1 expression in other tissue types is unknown. This study
showed that Srcin1 is expressed in human somatic tissues according
to IHC of a TMA. The present study revealed the unequivocal
presence of Srcin1 in 7 of 16 tissues examined. In particular, 80%
(4/5) of normal colon and rectal tissues were negative. However,
Scrin1 may be a novel negative regulator of tumour growth because
it strongly impaired breast cancer cell growth (17). Thus, Srcin1 is particularly
intriguing because it can function as either a repressor or an
activator of target proteins in a cell type-dependent fashion.
Further study could be of interest.
It has been reported that Srcin1 is essential for
the regulation of cell proliferation and motility (16,18).
Little is known, however, regarding the role of Srcin1 in CRC.
Studies have reported that Srcin1 expression in normal human breast
tissues inversely correlates with its expression in breast cancer
tissues (18). We showed that
Srcin1 was expressed at higher a level in CRC cells than in cells
from normal tissues. We determined that Srcin1 is a mediator of
NaB-induced pro-differentiation of CRC cells. Our finding that
Srcin1 suppression induced the maturation of F-actin filaments in
cancer cells implicates Srcin1 in the dedifferentiation of cancer
cells. Moreover, the suppression of Srcin1 increased the expression
of a differentiation marker for colorectal epithelial cells
(E-cadherin). Taken together, our data here show that the
suppression of Srcin1 increased differentiation and tumorigenesis
of CRC.
The cell cycle is regulated by a series of
checkpoints that monitor the genomic integrity and ensure that DNA
replication proceeds in a coordinated manner (28). Aberrations in cell cycle
progression occur in the majority of human malignancies (29). Different combinations of cyclin and
CDK subunits operate at checkpoint controls during the cell cycle
to integrate mitogenic and antiproliferative signals (30). Cyclin D1 has a critical role in the
control of the G1/S transition (31). The present study indicates that
downregulation of Srcin1 causes G0/G1 phase cell cycle arrest via a
reduction in cyclin D1 levels, which appears to be the underlying
mechanism in CRC growth inhibition. Therefore, this might also
contribute to G0/G1 phase arrest.
The canonical Wnt/β-catenin signalling pathway plays
an essential role in the regulation of developmental and adult
tissue homeostasis (25,26). This pathway is centred on
β-catenin, which binds to TCF/LEF transcription factors and leads
to the transcription of target genes (32). In this study, we proposed that
Srcin1 could regulate Wnt/β-catenin. The data that were obtained
from both experiments where Srcin1 was overexpressed or knocked
down by siRNA supported our hypothesis. Moreover, Srcin1 expression
affected the location and function of the TCF/β-catenin complex,
which results in the induction of genes that are downstream of
β-catenin. These studies and our results suggest that Srcin1 may
play a critical role in cell to-cell adhesion through its
regulation of the TCF/β-catenin complex. We are currently
evaluating the regulation of Wnt/β-catenin signaling by Srcin1
in vivo.
Recently, the catalytic activity of Src kinases was
found to regulate proliferation, migration and invasiveness of
MDA-MB-231 breast cancer cells, as well as decrease their
susceptibility to the chemotherapy agent oxaliplatin in CRC
(33,34). However, whether Srcin1 contributes
to chemoresistance remains to be elucidated. In the present study,
we demonstrated that Srcin1 knockdown in LoVo cells remarkably
suppressed cell proliferation and enhanced cell apoptosis in
response to 5-FU treatment in vitro. In vivo, Srcin1
knockdown inhibited xenograft tumorigenesis and resulted in the
near eradication of established CRC xenografts when combined with
5-FU. Collectively, these findings imply that Srcin1 inhibition by
RNAi along with 5-FU-based chemotherapy may be exploited as a
potential synergistic therapy for patients with CRC. Srcin1
interacts with Src kinase and Srcin1 and Src may differentially
regulate CRC cell differentiation, proliferation, migration,
invasion, and survival, we are evaluating the effects of Srcin1
overexpression and knockdown on Src activity.
Taken together, the results of the present study
demonstrate that Srcin1 is crucial for cancer growth and
metastasis. Srcin1 knockdown induces differentiation, inhibits cell
growth, causes cell cycle arrest in G0/G1 phase, promotes
spontaneous apoptosis, and enhances the chemosensitivity of CRC
LoVo cells to 5-FU. Our data suggest that Srcin1 may serve as a
promising therapeutic target for CRC.
Abbreviations:
|
ATCC
|
American Type Culture Collection
|
|
qRT-PCR
|
real-time RT-PCR
|
|
CRC
|
colorectal cancer
|
|
IHC
|
immunohistochemistry
|
|
TMA
|
tissue microarray
|
|
AJCC
|
American Joint Committee on Cancer
|
|
siRNA
|
small interfering RNA
|
|
RLU
|
relative luciferase unit
|
|
MOI
|
multiplicity of infection
|
|
Srcin1
|
SRC kinase signalling inhibitor 1
|
Acknowledgments
The present study was supported by grants from the
National Natural Science Funds of China (nos. 81172057, 81272761
and 81470036), the President Foundation of Nanfang Hospital,
Southern Medical University (2012B009 and 2013Z007), the High-Level
Topic-Matching Funds of Nanfang Hospital (201347 and G201227), the
Projects of Science and Technology of Guangdong (2012B050600020),
the Guangdong Provincial Key Laboratory of Gastroenterology,
Department of Gastroenterology, Nanfang Hospital, Southern Medical
University and the Guangzhou Pilot Project of Clinical and
Translational Research Center (early gastrointestinal cancer, no.
7415696196402).
References
|
1
|
Gill S and Sinicrope FA: Colorectal cancer
prevention: Is an ounce of prevention worth a pound of cure? Semin
Oncol. 32:24–34. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chinery R, Beauchamp RD, Shyr Y, Kirkland
SC, Coffey RJ and Morrow JD: Antioxidants reduce cyclooxygenase-2
expression, prostaglandin production, and proliferation in
colorectal cancer cells. Cancer Res. 58:2323–2327. 1998.PubMed/NCBI
|
|
3
|
Wang J, Yang Y, Xia HH, Gu Q, Lin MC,
Jiang B, Peng Y, Li G, An X, Zhang Y, et al: Suppression of FHL2
expression induces cell differentiation and inhibits gastric and
colon carcinogenesis. Gastroenterology. 132:1066–1076. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gu Q, Wang JD, Xia HH, Lin MC, He H, Zou
B, Tu SP, Yang Y, Liu XG, Lam SK, et al: Activation of the
caspase-8/Bid and Bax pathways in aspirin-induced apoptosis in
gastric cancer. Carcinogenesis. 26:541–546. 2005. View Article : Google Scholar
|
|
5
|
He H, Xia HH, Wang JD, Gu Q, Lin MC, Zou
B, Lam SK, Chan AO, Yuen MF, Kung HF, et al: Inhibition of human
telomerase reverse transcriptase by nonsteroidal antiinflammatory
drugs in colon carcinoma. Cancer. 106:1243–1249. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Capozzi E, Della Puppa L, Fornasarig M,
Pedroni M, Boiocchi M and Viel A: Evaluation of the replication
error phenotype in relation to molecular and clinicopathological
features in hereditary and early onset colorectal cancer. Eur J
Cancer. 35:289–295. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sinicrope FA and Sugarman SM: Role of
adjuvant therapy in surgically resected colorectal carcinoma.
Gastroenterology. 109:984–993. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kim IJ, Ku JL, Kang HC, Park JH, Yoon KA,
Shin Y, Park HW, Jang SG, Lim SK, Han SY, et al: Mutational
analysis of OGG1, MYH, MTH1 in FAP, HNPCC and sporadic colorectal
cancer patients: R154H OGG1 polymorphism is associated with
sporadic colorectal cancer patients. Hum Genet. 115:498–503. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li X, Liang L, Huang L, Ma X, Li D and Cai
S: High expression of protein phosphatase 4 is associated with the
aggressive malignant behavior of colorectal carcinoma. Mol Cancer.
14:952015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Maroney AC, Qureshi SA, Foster DA and
Brugge JS: Cloning and characterization of a thermolabile v-src
gene for use in reversible transformation of mammalian cells.
Oncogene. 7:1207–1214. 1992.PubMed/NCBI
|
|
11
|
Liu W, Yue F, Zheng M, Merlot A, Bae DH,
Huang M, Lane D, Jansson P, Lui GY, Richardson V, et al: The
proto-oncogene c-Src and its downstream signaling pathways are
inhibited by the metastasis suppressor, NDRG1. Oncotarget.
6:8851–8874. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Burger KL, Learman BS, Boucherle AK,
Sirintrapun SJ, Isom S, Díaz B, Courtneidge SA and Seals DF:
Src-dependent Tks5 phosphorylation regulates invadopodia-associated
invasion in prostate cancer cells. Prostate. 74:134–148. 2014.
View Article : Google Scholar
|
|
13
|
Baker AM, Cox TR, Bird D, Lang G, Murray
GI, Sun XF, Southall SM, Wilson JR and Erler JT: The role of lysyl
oxidase in SRC-dependent proliferation and metastasis of colorectal
cancer. J Natl Cancer Inst. 103:407–424. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Di Stefano P, Cabodi S, Boeri Erba E,
Margaria V, Bergatto E, Giuffrida MG, Silengo L, Tarone G, Turco E
and Defilippi P: P130Cas-associated protein (p140Cap) as a new
tyrosine-phosphorylated protein involved in cell spreading. Mol
Biol Cell. 15:787–800. 2004. View Article : Google Scholar :
|
|
15
|
Damiano L, Le Dévédec SE, Di Stefano P,
Repetto D, Lalai R, Truong H, Xiong JL, Danen EH, Yan K, Verbeek
FJ, et al: p140Cap suppresses the invasive properties of highly
metastatic MTLn3-EGFR cells via impaired cortactin phosphorylation.
Oncogene. 31:624–633. 2012.
|
|
16
|
Damiano L, Di Stefano P, Camacho Leal MP,
Barba M, Mainiero F, Cabodi S, Tordella L, Sapino A, Castellano I,
Canel M, et al: p140Cap dual regulation of E-cadherin/EGFR
cross-talk and Ras signalling in tumour cell scatter and
proliferation. Oncogene. 29:3677–3690. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sharma N, Repetto D, Aramu S, Grasso S,
Russo I, Fiorentino A, Mello-Grand M, Cabodi S, Singh V, Chiorino
G, et al: Identification of two regions in the p140Cap adaptor
protein that retain the ability to suppress tumor cell properties.
Am J Cancer Res. 3:290–301. 2013.PubMed/NCBI
|
|
18
|
Di Stefano P, Damiano L, Cabodi S, Aramu
S, Tordella L, Praduroux A, Piva R, Cavallo F, Forni G, Silengo L,
et al: p140Cap protein suppresses tumour cell properties,
regulating Csk and Src kinase activity. EMBO J. 26:2843–2855. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kruh J: Effects of sodium butyrate, a new
pharmacological agent, on cells in culture. Mol Cell Biochem.
42:65–82. 1982.PubMed/NCBI
|
|
20
|
Candido EP, Reeves R and Davie JR: Sodium
butyrate inhibits histone deacetylation in cultured cells. Cell.
14:105–113. 1978. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Davie JR: Inhibition of histone
deacetylase activity by butyrate. J Nutr. 133:2485S–2493S.
2003.PubMed/NCBI
|
|
22
|
Kopetz S, Morris VK, Parikh N, Overman MJ,
Jiang ZQ, Maru D, Elvin P and Gallick G: Src activity is modulated
by oxaliplatin and correlates with outcomes after hepatectomy for
metastatic colorectal cancer. BMC Cancer. 14:6602014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wu Y, Guo Z, Zhang D, Zhang W, Yan Q, Shi
X, Zhang M, Zhao Y, Zhang Y, Jiang B, et al: A novel colon cancer
gene therapy using rAAV-mediated expression of human shRNA-FHL2.
Int J Oncol. 43:1618–1626. 2013.PubMed/NCBI
|
|
24
|
Zhang W, Jiang B, Guo Z, Sardet C, Zou B,
Lam CS, Li J, He M, Lan HY, Pang R, et al: Four-and-a-half LIM
protein 2 promotes invasive potential and epithelial-mesenchymal
transition in colon cancer. Carcinogenesis. 31:1220–1229. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yan Q, Zhang W, Wu Y, Wu M, Zhang M, Shi
X, Zhao J, Nan Q, Chen Y, Wang L, et al: KLF8 promotes
tumorigenesis, invasion and metastasis of colorectal cancer cells
by transcriptional activation of FHL2. Oncotarget. 6:25402–25417.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Schneikert J, Grohmann A and Behrens J:
Truncated APC regulates the transcriptional activity of
beta-catenin in a cell cycle dependent manner. Hum Mol Genet.
16:199–209. 2007. View Article : Google Scholar
|
|
27
|
Tetsu O and McCormick F: Beta-catenin
regulates expression of cyclin D1 in colon carcinoma cells. Nature.
398:422–426. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kennedy S, Clynes M, Doolan P, Mehta JP,
Rani S, Crown J and O'Driscoll L: SNIP/p140Cap mRNA expression is
an unfavourable prognostic factor in breast cancer and is not
expressed in normal breast tissue. Br J Cancer. 98:1641–1645. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Eves EM and Rosner MR: MAP kinase
regulation of the mitotic spindle checkpoint. Methods Mol Biol.
661:497–505. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Stewénius Y, Gorunova L, Jonson T, Larsson
N, Höglund M, Mandahl N, Mertens F, Mitelman F and Gisselsson D:
Structural and numerical chromosome changes in colon cancer develop
through telomere-mediated anaphase bridges, not through mitotic
multipolarity. Proc Natl Acad Sci USA. 102:5541–5546. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Van Arsdale T, Boshoff C, Arndt KT and
Abraham RT: Molecular pathways: Targeting the cyclin D-CDK4/6 axis
for cancer treatment. Clin Cancer Res. 21:2905–2910. 2015.
View Article : Google Scholar
|
|
32
|
Kurokawa K, Akaike Y, Masuda K, Kuwano Y,
Nishida K, Yamagishi N, Kajita K, Tanahashi T and Rokutan K:
Downregulation of serine/arginine-rich splicing factor 3 induces G1
cell cycle arrest and apoptosis in colon cancer cells. Oncogene.
33:1407–1417. 2014. View Article : Google Scholar
|
|
33
|
Chan DW, Mak CS, Leung TH, Chan KK and
Ngan HY: Downregulation of Sox7 is associated with aberrant
activation of Wnt/b-catenin signaling in endometrial cancer.
Oncotarget. 3:1546–1556. 2012. View Article : Google Scholar
|
|
34
|
Sánchez-Bailón MP, Calcabrini A,
Gómez-Domínguez D, Morte B, Martín-Forero E, Gómez-López G,
Molinari A, Wagner KU and Martín-Pérez J: Src kinases catalytic
activity regulates proliferation, migration and invasiveness of
MDA-MB-231 breast cancer cells. Cell Signal. 24:1276–1286. 2012.
View Article : Google Scholar : PubMed/NCBI
|