Introduction
Osteosarcomas (OSs) are highly aggressive primary
bone tumors of osteoblastic origin that predominantly affect
children and young adults (1).
Early treatment paradigms in the 1960s entailed aggressive local
control with amputation. More recently, a multidisciplinary
treatment approach has enhanced both the overall survival and
preservation of the organ/limb function (2). Ionizing radiation (IR) therapy is a
facet of the modern approach to treat OS (3–6);
however, recent reports have emphasized the limited therapeutic
efficacy of local low-LET (gamma-radiation) radiotherapy for the
treatment of OS, as well as an increased risk of pulmonary
metastasis (5–8). Consequently, conventional therapies
are unable to prevent metastatic progression in 30–40% of the
patients with OS (9). To generate
a more effective and comprehensive treatment methodology,
utilization of high-LET radiation might be necessary to treat OS
that has become resistant to low-LET radiation. High-LET radiation
has several advantages for treating radio-resistant human cancers
because of its higher relative biological effectiveness (RBE),
lower oxygen enhancement ratio (OER), and decreased cell
cycle-dependent radiosensitivity. In addition, because cells are
less likely to repair radiation-induced damage, charged particle
radiation may exert highly lethal effects on radioresistant tumors
as compared to conventional low-LET X-ray or gamma-ray irradiations
(10–13).
As compared to conventional approaches, carbon ion
high-LET radiotherapy is highly efficacious in treating deep-seated
malignant OSs (14,15). In addition, boron neutron capture
therapy (BNCT) has been used to successfully treat a patient with
recurrent radiation-induced OS (16). However, the molecular mechanisms
underlying carbon ion- or neutron-induced OS cytotoxicity are
largely unknown. Therefore, this study investigated the therapeutic
effects of high-LET neutron radiation on OS in vitro and
in vivo.
Materials and methods
Antibodies and chemicals
Anti-p53 and β-actin were purchased from Santa Cruz
Biotechnology (Santa Cruz, CA, USA). Anti-cleaved PARP, caspase-3,
caspase-9, cleaved caspase-3, cleaved caspase-9, anti-LC3,
anti-ATM, anti-p95/NBS1, anti-Ku70, anti-Ku80, anti-DNA-PKcs, and
anti-ERCC1 were purchased from Cell Signaling Technology (Danvers,
MA, USA), and anti-γ-H2AX was obtained from Millipore (Billerica,
MA, USA).
Cell culture
The OS cell lines, U2O2 and KHOS/NP [American Type
Culture Collection (ATCC), Rockville, MD, USA], were maintained in
α minimum essential medium (α-MEM; Gibco Life Technologies,
Carlsbad, CA, USA) supplemented with 10% (v/v) fetal bovine serum
(FBS; Gibco Life Technologies) and 1% (v/v) penicillin-streptomycin
solution (Gibco Life Technologies).
Irradiation
Cells were cultured in 60- or 100-mm dishes until
70–80% confluence at 37°C in a humidified atmosphere of 5%
CO2. Irradiations were performed using a
137Cs gamma-ray source (Atomic Energy of Canada, Ltd.,
Ontario, Canada) at a dose rate of 3.81 Gy/min. Fast neutrons (9.8
MeV, 30–40 keV/μm) were produced by the bombardment of
proton on beryllium 9Be(p,n)10B as a nuclear
reaction in the cyclotron (MC-50; Scanditronix, Uppsala, Sweden).
Paired ionization chambers were used to measure the absorbed dose
and dose distribution of fast neutron beams or gamma-rays (17). Dosimetric measurements were done
before in vitro study to calculate neutron dose using RBE,
2.2, which has been used for neutron therapy in our institute and
it showed cell killing efficacy equivalent to that of gamma-ray as
determined by clonogenic assay (18).
Colony-forming assay
Cells (500–1000) were seeded into 60-mm dishes in
triplicate and stained with 0.4% crystal violet (Sigma, St. Louis,
MO, USA) after 14–20 days to determine plating efficiency (PE),
defined as the percentage of seeded cells that formed colonies
under the specific culture conditions. The surviving fraction was
expressed as a function of irradiation as follows: survival
fraction = colonies counted / (cells seeded × PE/100). The plating
efficiencies of U2O2 and KHOS/NP cells were 0.48±0.18 and
0.34±0.02, respectively. RBE is defined as the ratio of the doses
of the two radiations required to cause the effect to the same
degree. To evaluate the RBE, the ratio of the doses of the two
types of radiations required for similar effect at a survival
fraction of 50% was determined. RBE was evaluated and calculated as
the dose (Gy) for gamma-ray radiation divided by the dose for
neutron radiation that yielded a surviving fraction of 50%
(D50).
Water-soluble tetrazolium (WST-1)
assay
For the cytotoxicity assay, cells were seeded in
96-well culture plastic plates at a density of 1×103
cells per well. Each well was exposed to radiation at varying doses
(0–5 Gy) and the cells were incubated for 72 h, followed by
application of the water-soluble tetrazolium (WST)-1 cytotoxicity
assay reagent (Roche Diagnostics, Laval, Quebec, Canada) according
to the manufacturer's recommendations. Cell viability was assessed
by determining the A450 nm of the cell culture media after addition
of WST-1 for 2 h. The results are reported as a percentage of the
optical density of the untreated control cells, which was
designated as 100% cell viability. Percentage of cytotoxicity was
calculated as (1-Aexp/Acon) ×100, where Aexp and Acontrol are the
absorbance values of the experimental IR-treated and control
untreated cells, respectively.
Analysis of cell cycle progression
Cells were seeded in 60-mm dishes at 60% confluency.
After 24 h, cells were trypsinized, harvested, and fixed in 1 ml
70% cold ethanol in test tubes and then incubated at 4°C overnight.
The fixed cells were centrifuged at 2,000 rpm for 3 min, and the
pellets were resuspended in 500 μl propidium iodine (10
μg/ml) containing 300 μg/ml RNase (Sigma). Cell cycle
distribution was determined using 10,000 cells with CellQuest™
software using a FACSCaliber flow cytometer (Becton Dickinson, San
Jose, CA, USA).
Detection of apoptotic cells by Annexin V
staining
Cells were irradiated and subsequently incubated for
48 h. Cells (1×106 cells/ml) were then washed with
ice-cold PBS, trypsinized, and resuspended in 1X binding buffer [10
mM HEPES/NaOH (pH 7.4), 140 mM NaCl, and 2.5 mM CaCl2].
Aliquots (100 μl) of the cell solution were mixed with 5
μl Annexin V/fluorescein isothiocyanate (FITC) (BD
Biosciences, Franklin Lakes, NJ, USA) and 10 μl propidium
iodide (PI) stock solution (50 μg/ml in PBS) by gentle
vortexing, followed by 15-min incubation at room temperature under
dark conditions. Next, 400 μl 1X binding buffer was added to
each sample, and the samples were analyzed on a FACScan™ flow
cytometer (BD Biosciences). A minimum of 10,000 cells were counted
for each sample, and data were analyzed using CellQuest™ software
(BD Biosciences).
Western blotting
Irradiated OS cells were cultured and then lysed
with RIPA buffer. Proteins were separated by SDS-PAGE and
transferred onto nitrocellulose membranes. The membranes were
blocked with 1% (v/v) nonfat dry milk in Tris-buffered saline with
0.05% Tween-20 and incubated with the indicated primary and
secondary antibodies (1:1,000 and 1:5,000 dilution, respectively).
Immunoreactive protein bands were visualized by enhanced
chemiluminescence (Amersham Biosciences, Little Chalfont, UK).
Caspase activity
Caspase-3 and -9 activities were determined by using
detection kits (R&D Systems, Minneapolis, MN, USA). The assay
is based on spectrophotometric detection of the chromophore
p-nitroanilide (pNA) after cleavage from the labeled
substrates of DEVD-pNA (for caspase-3) and LEHD-pNA
(for caspase-9). The pNA light emission can be quantified
using a spectrophotometer or microtiter plate reader at 405 nm.
Comparison of the pNA absorbance of apoptotic and control
samples allows determination of the fold increase in caspase
activity.
Intracellular reactive oxygen species
detection
Reactive oxygen species (ROS) were monitored using
the fluorescent ROS indicator, C2′,7′-dichlorodihydrofluorescein
diacetate (H2DCFDA; 5 μM; Molecular Probes,
Eugene, OR, USA). Cell-associated fluorescence was detected by FACS
using a FACSort™ flow cytometer with CellQuest software (BD
Biosciences).
Quantitative detection of acid vesicular
organelles by acridine orange staining
Autophagy is the process of sequestrating
cytoplasmic proteins and organelles into the lysosomal component
and is characterized by development of acid vesicular organelles
(AVOs). To detect and quantify AVOs in IR-treated cells, we
performed vital staining with acridine orange (Polysciences,
Warrington, PA, USA) as previously described. U2OS and KHOS/NP
cells were stained with 1.0 μg/ml acridine orange for 15 min
at room temperature and processed for flow cytometry using the
FACScan cytometer and analyzed with CellQuest software (Becton
Dickinson).
Immunocytochemistry
Immunocytochemistry was performed to determine the
nuclear distribution of γ-H2AX in individual cells. Cells were
grown on chambered slides for 1 day prior to IR. After exposure,
cells were irradiated and incubated for 1 or 24 h. All treatments
were performed while cells remained attached to the slides,
followed by fixation with 4% (w/v) paraformaldehyde and
permeabilization with 0.5% (v/v) Triton™ X-100 in PBS. Detection
was performed after the slides were blocked in 10% (v/v) FBS/1%
(v/v) bovine serum albumin for 1 h, followed by incubation with a
1:1,000 dilution of FITC-labeled mouse monoclonal antibody against
γ-H2AX (Millipore).
Wound healing (scratch) assay
Human OS cells were seeded onto 6-well plates
(Corning) at a density of 2.5×104 cells/well, 3 ml
medium was added, and the cells were irradiated. On day 2, the
monolayers were mechanically disrupted with a sterile 200-μl
pipette tip. The assay was performed in duplicates, and wells were
photographed every 48 h prior to staining with 0.2% (w/v) crystal
violet. Cell migration was monitored using an Eclipse Ti microscope
with a DS-Fi1 camera (Nikon, Tokyo, Japan). The cells were counted
using ImageJ (US National Institutes of Health, Bethesda, MD,
USA).
Transwell chamber assay
The in vitro invasive ability of OS cells was
measured using transwell chambers according to the manufacturer's
protocol. Briefly, cells were seeded onto the membrane of the upper
chamber of the transwell at a density of 4×105/ml in 150
μl medium and were left untreated or irradiated with IR for
24 h. The medium in the upper chamber was serum-free, whereas the
medium in the lower chamber contained 10% (v/v) FBS as a source of
chemoattractants. Cells that passed through the
Matrigel®-coated membrane were stained with cell stain
solution containing crystal violet supplied in the Transwell
chamber assay (Chemicon; Millipore) and photographed after 24 h of
incubation.
Orthotopic model and histological
analysis
Twelve 4-week-old female BALB/c nude mice (average
body weight, 12.1 g; range 11.3–13.1 g) were obtained from Orient
Bio Inc. (Seoul, Korea) and quarantined for 1 week prior to
experimentation. KHOS/NP orthotopic tumors were established as
previously described (19).
Cells were implanted into the tibias of mice
anesthetized by intraperitoneal injection of Zoletil (Virbac,
Carros, France) and Roumpun (Bayer Korea, Seoul, Korea). Briefly,
the left tibia was wiped with 70% (v/v) ethanol and an 18-gauge
needle was inserted through the tibial plateau with the knee
flexed. Then, 1×105 KHOS/NP cells in 10 μl PBS
were injected into the marrow space of the proximal tibia with a
26-gauge needle coupled with a Hamilton syringe. Two weeks after OS
cell inoculation, mice were randomly assigned into IR or control
groups (n=4/group). IR was given as a single dose of 8 Gy of
low-LET gamma-ray or high-LET neutron radiotherapy. Animals were
sacrificed 6 weeks after inoculation by CO2
asphyxiation. Tumor length (L) and width (W) were measured with
calipers after their sacrifice and used to calculate tumor volume
as follows: volume = (L × W2)/2. All experimental
protocols were approved by the Institutional Animal Care and Use
Committee of the Korea Institute of Radiological and Medical
Sciences. Histological analysis was performed using hematoxylin and
eosin (H&E)-stained paraffin sections (20,21).
Statistical analysis
All analyses were performed by Student's t-test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
U2OS and KHOS/NP clonogenicity following
gamma-ray or neutron irradiation
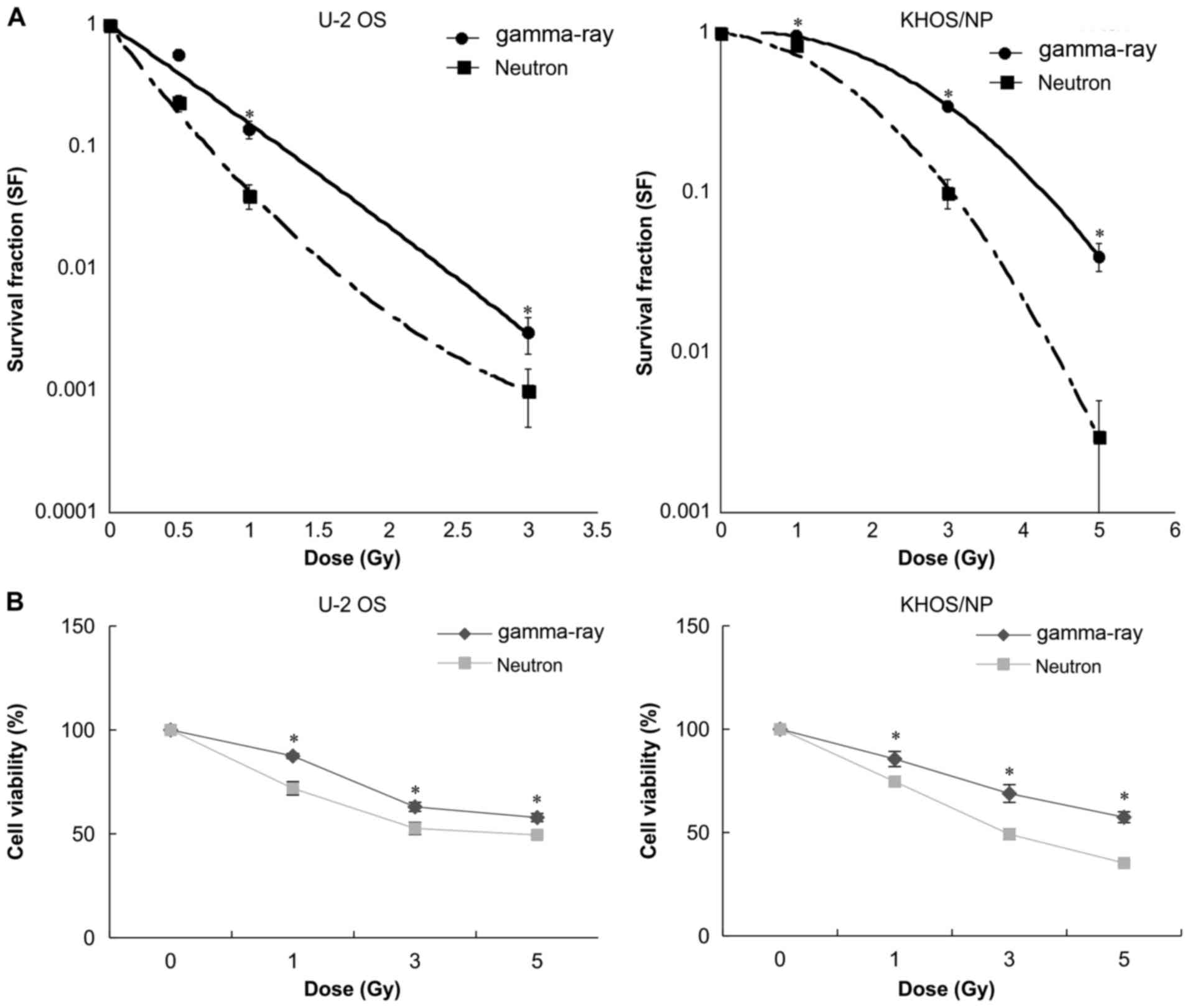
U2O2 and KHOS/NP OS clonogenic survival decreased as
a function of exposure to gamma-ray or neutron irradiation dose
(Fig. 1). The results of the
colony-forming assay of irradiated KHOS/NP cells are shown in
Fig. 1A in the form of survival
curves. The surviving fraction (S/S0) data for irradiated OS cells
were fitted with the linear quadratic dose (D)-dependent relation
given by S/S0 = exp − (αD + βD2), where α and β are
constants. The fitted values of α and β for irradiated KHOS/NP
cells are provided in Table I. RBE
depends on the dose and the biological endpoints. The average
endpoint D50 for U2O2 and KHOS/NP cells was 0.81 and 3.11 Gy for
gamma-irradiation, and 0.45 and 2.01 Gy for neutron irradiation,
respectively (Table II).
Similarly, the dose ratio required for attaining D50 with gamma-ray
or neutron irradiation is extracted from Fig. 1A and summarized in Table II. The values from the fitted
curves suggest that as compared to that of gamma-ray irradiation,
neutron irradiation was far more effective in reducing the survival
fraction. Additionally, U2OS cells showed more radiosensitive than
KHOS/NP cells. Therefore, the RBE values of neutron irradiation
relative to gamma-ray irradiation were 1.80 and 1.55, respectively,
indicating that the neutron beam exerts more effective cell killing
effects compared to the gamma-ray irradiation. The two OS cell
lines were also subjected to cell viability analysis by WST-1 assay
after either form of irradiation (Fig.
1B), which verified that neutron irradiation had a more potent
radiosensitizing effect.
 | Table IFitting parameters α and β for
survival assay data. |
Table I
Fitting parameters α and β for
survival assay data.
| Cell type | Radiation type | α
(Gy−1) | β
(Gy−1) |
|---|
| U2OS | gamma-ray | 0.182±1.260 | 0.042±0.447 |
| Neutron | 3.552±1.258 | 0.416±0.445 |
| KHOS/NP | gamma-ray | 0.097±0.728 | 0.148±0.162 |
| Neutron | 0.107±0.729 | 0.212±0.164 |
| Table IIRadiation dose required for 50%
cytotoxicity and relative biological effectiveness (RBE) values
(data extracted from Fig. 1). |
Table II
Radiation dose required for 50%
cytotoxicity and relative biological effectiveness (RBE) values
(data extracted from Fig. 1).
| Cell type | Radiation type | D50 | RBE |
|---|
| U2OS | gamma-ray | 0.81 | 1.80 |
| Neutron | 0.45 | |
| KHOS/NP | gamma-ray | 3.11 | 1.55 |
| Neutron | 2.01 | |
Effect of low- or high-LET IR-induced
apoptosis and cell cycle
We next analyzed the effects of gamma-ray and
neutron irradiation on cell cycle progression using flow cytometry
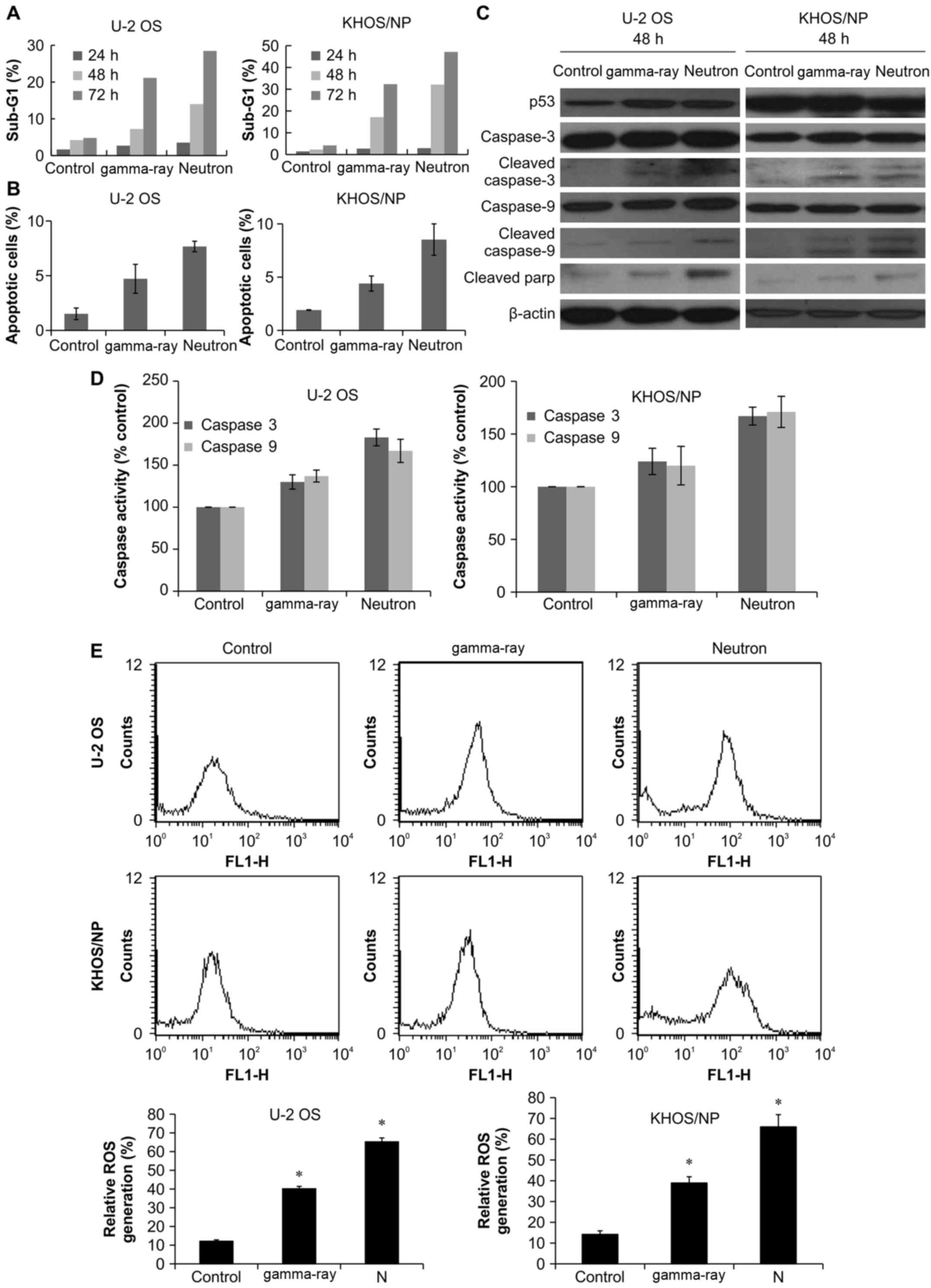
to assess the relative changes in apoptotic sub-G1 cells. Notably,
neutron irradiation resulted in a higher overall increase in sub-G1
cells than that in gamma-ray-treated counterparts in a
time-dependent manner (Fig. 2A).
To determine the fraction of OS cells induced to undergo apoptosis
by the radiation treatment, we detected apoptosis by Annexin V and
PI staining. Notably, 48 h treatment with neutron increased the
percentage of early apoptotic cells as compared to that after
treatment with gamma-ray in the two OS cell lines (Fig. 2B). Cleavage-induced caspase
activation is a defining characteristic of apoptotic cell death
(22). Therefore, we examined
caspase-3 and caspase-9 expression and cleavage by western
blotting. As shown in Fig. 2C,
cell irradiation resulted in increased caspase-3 and -9 expression
in OS cells, with a more potent effect observed with neutron
radiation. Caspase-3 activation leads to PARP-1 cleavage (23). As expected, western blot analysis
showed a marked increase in cleaved PARP-1 level up to 48 h after
exposure, and a more substantial effect was observed with neutron
radiation (Fig. 2C). We measured
radiation-induced activation of caspase-3 and -9 protease
activities. As shown in Fig. 2D,
neutron radiation significantly enhanced caspase-3 and -9 as
compared to that by gamma-ray radiation. Annexin V staining and
caspase activation indicated that neutron irradiation-induced cell
death is dependent on apoptosis induction. To investigate the
relationship between ROS production and radiation-induced
apoptosis, the effects of ROS production on OS cells were examined.
These results revealed elevated ROS production in cells treated
with high-LET radiation as compared to that in cells treated with
low-LET radiation (Fig. 2E).
Effect of radiation-induced autophagy on
OS cells
Autophagy was determined after irradiation by
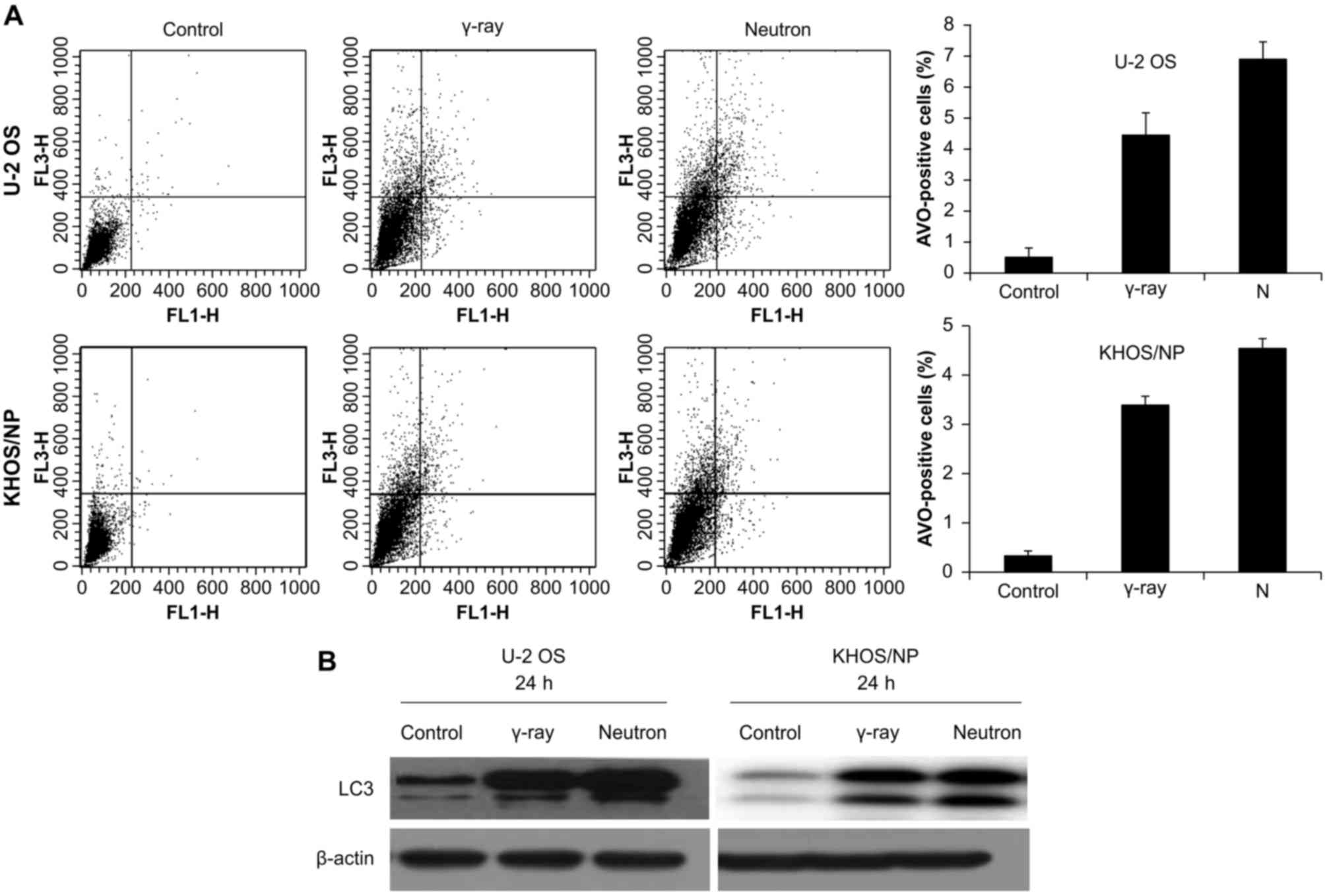
counting AVO-containing cells at given time-points (Fig. 3A). Higher increase in autophagy
after neutron irradiation was demonstrated in the two cell lines.
The expression of LC3 protein, which is known to be a standard
indicator of autophagy, was assessed. Among two separate bands
observed on western blot (an upper one of 18 kDa for LC3-bI and a
lower one of 16 kDa for LC3-bII), the lower band representing
fusion of the autophagosome to lysosome increased with increasing
time after neutron irradiation (Fig.
3B).
Effect of low- or high-LET IR on DNA
damage
To explore the involvement of cellular DNA repair in
high-LET radiation-induced OS cytotoxicity, we analyzed the
abundance of the damage-response protein, H2AX, by
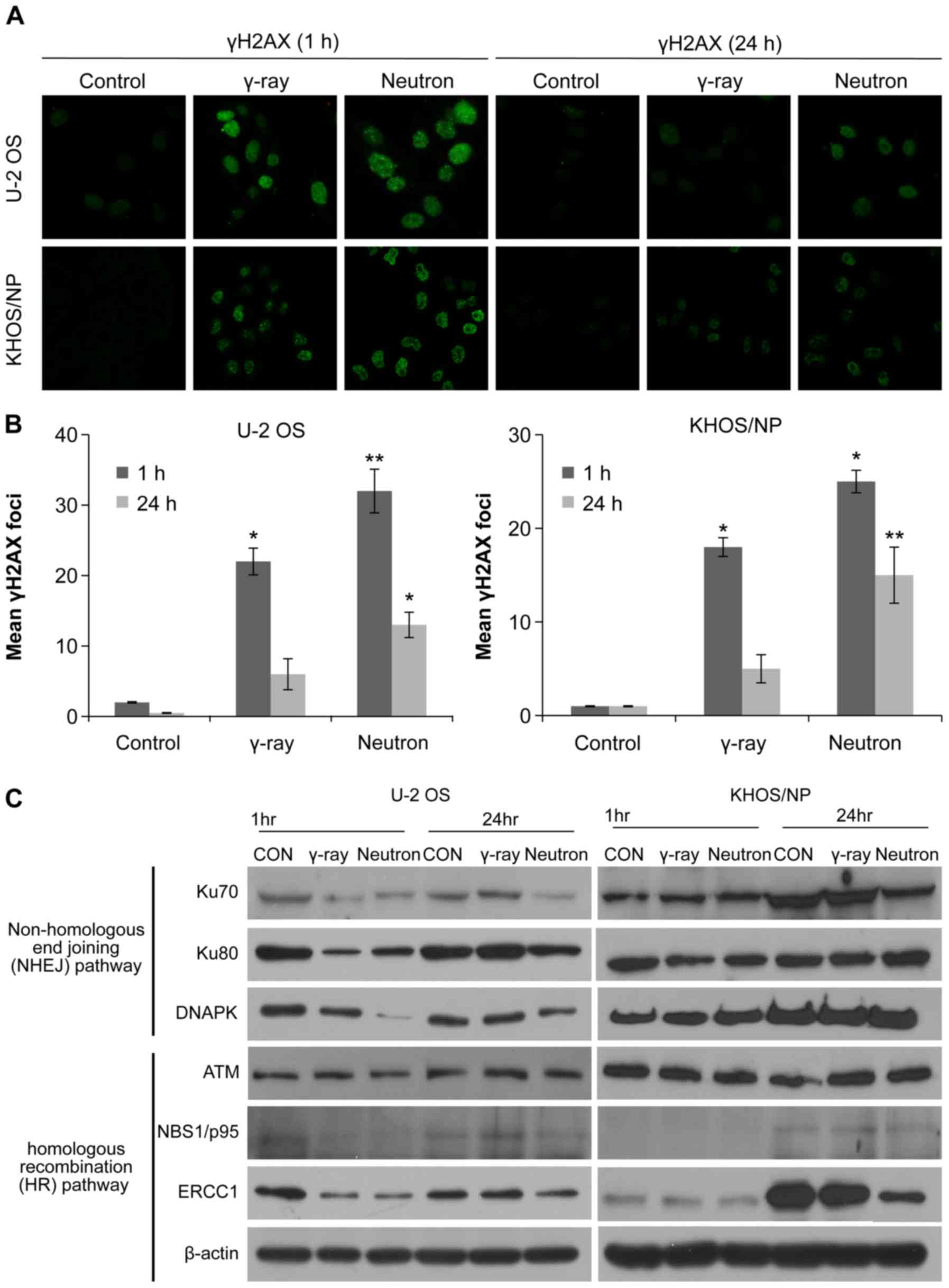
immunocytochemistry and western blotting. The number of γH2AX foci
was initially increased 1 h after irradiation with either gamma-ray
or neutron beams, but then substantially declined after 24 h. The
two OS cell lines treated with neutron radiation exhibited damaged
DNA foci, which appeared 1 h after treatment exposure and were
retained even 24 h after exposure; therefore, the effect was
greater with neutron radiation than with gamma-ray irradiation
(Fig. 4A and B). Moreover,
neutron-irradiated cells exhibited higher degree of IR-induced
γ-H2AX staining, indicative of increased DNA damage (Fig. 4A and B). To investigate whether
reduced double-stranded break (DSB) repair activity detected in
IR-treated cells was due to the suppression of DSB repair-related
proteins, the effect of IR treatment on the expression of key
proteins in the homologous recombination (HR) and non-homologous
end-joining (NHEJ) pathways was examined. The relative abundance of
DNA damage repair-associated proteins in OS cells under various
treatment conditions is shown in Fig.
4C. The proteins Ku70, Ku80, DNA-PKcs, NBS1, and ERCC1 were
downregulated 24 h after neutron irradiation in U2OS cells.
However, Ku70, NBS1, and ERCC1 were downregulated 24 h after
neutron irradiation in KHOS/NP cells. These results suggest that
neutron irradiation inhibits the expression of both HR- and
NHEJ-related proteins, inhibiting the repair of IR-induced DSBs,
depending upon the cell lines.
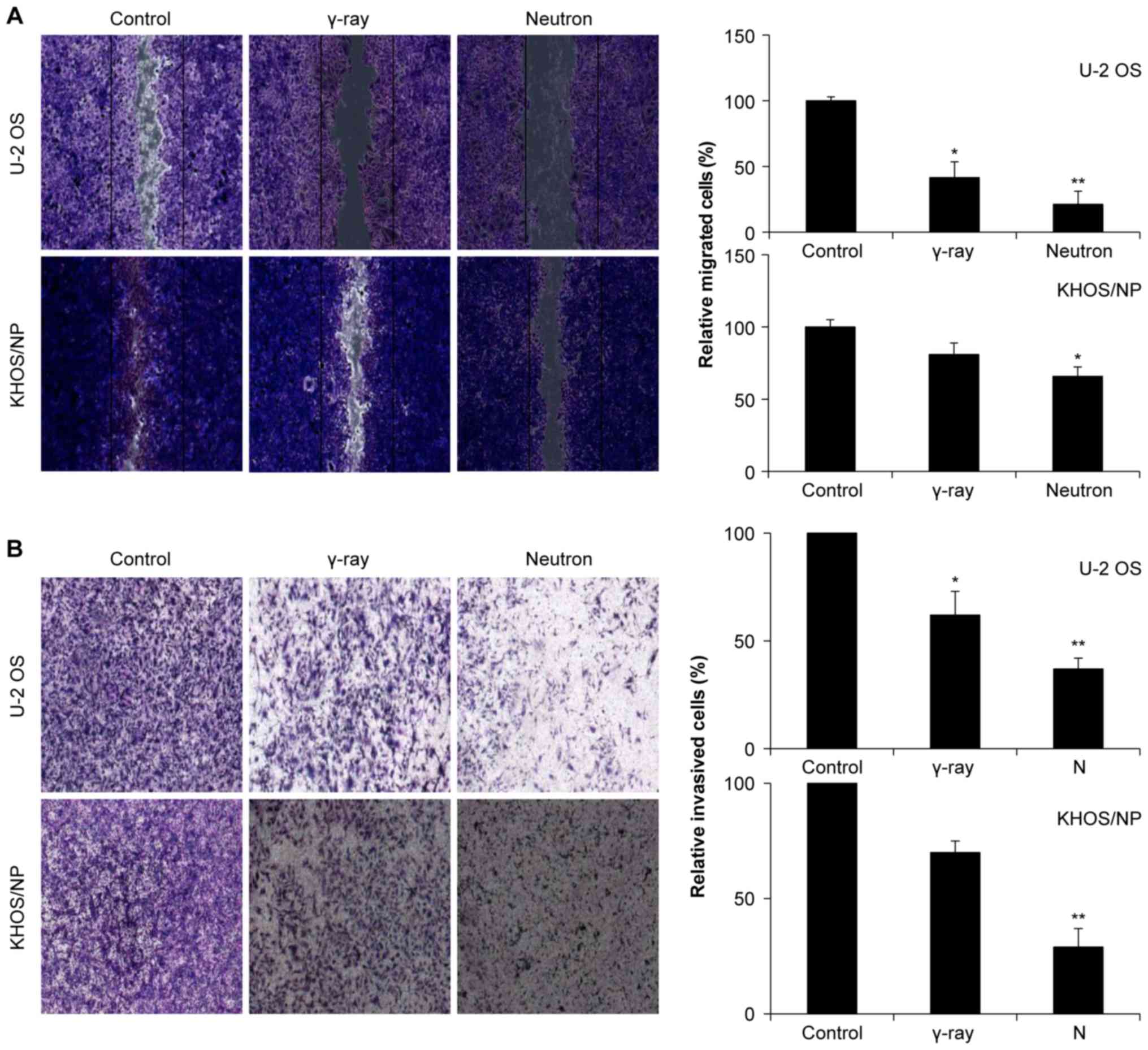
Effect of IR on cell motility and cell
invasion
Next, the effects of gamma-ray and neutron
irradiation on the migratory abilities of OS cells were estimated
using a scratch assay. Remarkably, compared to gamma-ray radiation,
neutron radiation significantly inhibited cell migration (Fig. 5A). Matrigel invasion assay was also
performed to examine the effect of IR on tumor cell invasiveness in
OS cell lines, and it was found that neutron irradiation
effectively inhibited tumor cell invasion (Fig. 5B).
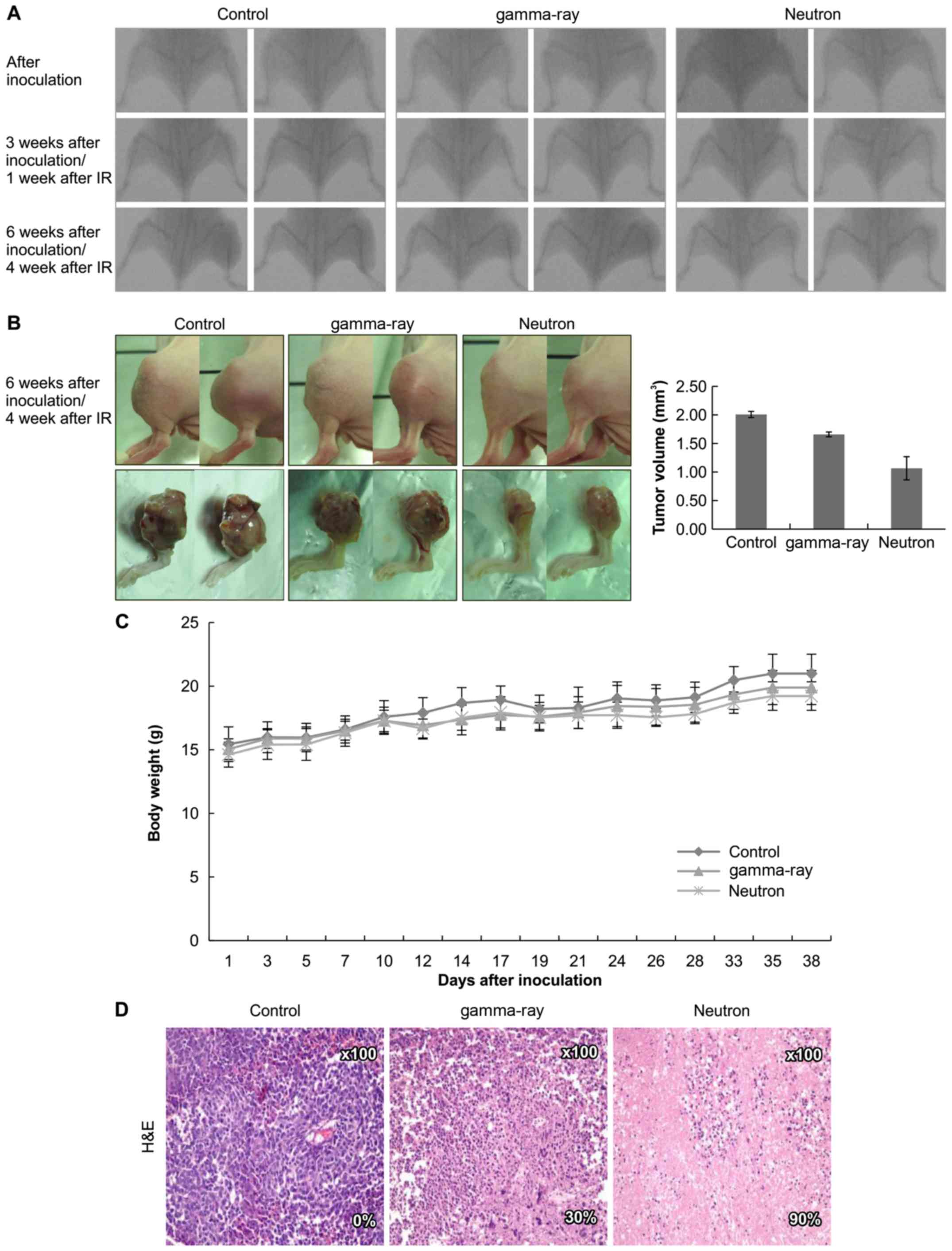
Effects of low- or high-LET IR on
orthotopic tumors in vivo
Based on these data, the therapeutic efficacy of
high-LET radiation was assessed in vivo using an orthotopic
mouse model (Fig. 6A).
Significantly, neutron irradiation decreased tumor growth in mice
as compared to that in gamma-ray-treated mice (Fig. 6B) with no visible signs of toxicity
as evidenced by the lack of a difference in body weight (Fig. 6C). Additionally, H&E staining
revealed that tumor from high-LET radiation-treated mice showed
higher apoptosis rate (Fig.
6D).
Discussion
Osteosarcomas (OSs) are primary malignant bone
tumors identified by the production of osteoid or immature bone
(1). These tumors generally
exhibit a high potential for pulmonary metastasis and significant
radioresistance (5–8). Based on this evidence, high-LET
neutron and carbon ion beam radiations have been the preferred
treatment modality for these patients. Preclinical evidence shows
that the biological effects of IR depend on the quality-type and
dose of radiation, as well as the cell or tumor type (24). High-LET radiation induces dense
ionization along the particle track sufficient for highly localized
DNA damage (25). Conversely,
low-LET radiation beams are sparse, resulting in more diffuse,
homogeneous dosing (25).
Therefore, the biological effect of high-LET radiation is generally
much higher than that of low-LET radiation at the same dose
(26), generally, because high-LET
radiation deposits most of its energy within one cell, resulting in
more extensive DNA damage (27,28).
Thus, high-LET radiation generates more complicated and varied
cellular effects; however, the molecular mechanisms underlying
high-LET radiation-induced cytotoxicity remain a topic of
investigation.
Herein, we investigated the effects of high- and
low-LET irradiation on the two OS cell lines. Notably, high-LET
radiation was significantly more effective at inhibiting OS cell
proliferation and inducing caspase-3/9 activation (Figs. 1 and 2). Additionally, an increase in AVOs was
detected by acridine orange staining of lysosomes as well as
conversion of LC3B-I into LC3B-II by immunoblotting in neutron
treated cells. These results supported the induction of enhanced
autophagy in response to neutron radiation in OS cells compared to
that with gamma-ray radiation (Fig.
3). Our results also showed that high-LET radiation results in
stable free radicals and more DSBs than that observed with
gamma-ray radiation. Neutron irradiation also sensitizes OS cells
by interfering with the NHEJ and HR pathways, thus limiting DSB
repair in OS cells (Fig. 4).
Finally, we studied the metastatic effect of IR treatment and found
that neutron radiation effectively inhibited invasion and migration
of OS cells (Fig. 5). These unique
findings improve our understanding about the indirect effects of
high-LET radiation and its clinical application to patients with
OS.
Orthotopic models are essential for the preclinical
evaluation of therapeutic agents and they are used for studying the
pathobiology of tumor progression and metastasis. Accordingly, we
examined the effects of each type of irradiation on an orthotopic
mouse model of OS. Tumor measurements using caliper and H&E
staining showed significantly smaller tumors with increased
apoptosis in high-LET radiation-treated mice, consistent with our
in vivo data (Fig. 6).
However, only three medical centers worldwide use high-LET neuron
radiotherapy to treat cancer, perhaps because of the difficulties
in obtaining funding and regulatory approval. A potentially more
viable alternative approach is to use other forms of high-LET
radiation, such as carbon ion beams, which are widely used in both
Japan and Europe. A previous study has shown similar RBE between
neutron and carbon ion beams, but there are differences in the
physical dose distribution and damage inflicted on normal tissues,
which we intend to study further (10). Moreover, we believe that carbon ion
beam radiotherapy results in a higher RBE than that with neutron
beam causing less damage to normal tissues. Hence, carbon ion beam
has proven to be a safe and effective modality to manage
unresectable OS of the trunk, where it provides good local control
and long-term functional results without ensuing morbidity
(15,16).
Carbon ion high-LET radiation therapy has provided
dramatic local control in Japanese hospitals and has become more
popular in radiation treatment. Substantial research has shown that
molecular mechanisms differ between high LET and low-LET radiation.
These differences should be further investigated to delineate any
potential rationale or clinical utility for low- versus high-LET
radiation therapy, such as neutron or carbon beam treatment, in
patients with OS.
In conclusion, our study revealed that as compared
to low-LET gamma-ray radiotherapy, high-LET neutron radiotherapy
provided a stronger therapeutic benefit by increasing OS cell
apoptosis and DNA damage in vitro and in orthotopic mouse
model in vivo, and thus, it provides further preclinical
rationale for high-LET radiotherapy.
Acknowledgments
This study was supported by a National Research
Foundation (NRF) of Korea grant (nos. NRF-2017R1D1A1B03028923)
funded by the Korean Government (MSIP) and by a grant of the Korea
Institute of Radiological and Medical Sciences (KIRAMS), funded by
Ministry of Science, ICT, and Future Planning, Republic of Korea
(1711045557;1711045538; 1711045554/50531-2017,50473-2017).
References
|
1
|
Unni KK and Dahlin DC: Osteosarcoma:
Pathology and classification. Semin Roentgenol. 24:143–152. 1989.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ciernik IF, Niemierko A, Harmon DC,
Kobayashi W, Chen YL, Yock TI, Ebb DH, Choy E, Raskin KA, Liebsch
N, et al: Proton-based radiotherapy for unresectable or
incompletely resected osteosarcoma. Cancer. 117:4522–4530. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ta HT, Dass CR, Choong PF and Dunstan DE:
Osteosarcoma treatment: State of the art. Cancer Metastasis Rev.
28:247–263. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Schwarz R, Bruland O, Cassoni A, Schomberg
P and Bielack S: The role of radiotherapy in oseosarcoma. Cancer
Treat Res. 152:147–164. 2009. View Article : Google Scholar
|
|
5
|
Beck JC, Wara WM, Bovill EG Jr and
Phillips TL: The role of radiation therapy in the treatment of
osteosarcoma. Radiology. 120:163–165. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
DeLaney TF, Park L, Goldberg SI, Hug EB,
Liebsch NJ, Munzenrider JE and Suit HD: Radiotherapy for local
control of osteosarcoma. Int J Radiat Oncol Biol Phys. 61:492–498.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Imai R, Kamada T, Tsuji H, Tsujii H,
Tsuburai Y and Tatezaki S; Working Group for Bone and Soft Tissue
Sarcomas: Cervical spine osteosarcoma treated with carbon-ion
radiotherapy. Lancet Oncol. 7:1034–1035. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mankin HJ, Hornicek FJ, Rosenberg AE,
Harmon DC and Gebhardt MC: Survival data for 648 patients with
osteosarcoma treated at one institution. Clin Orthop Relat Res.
429:286–291. 2004. View Article : Google Scholar
|
|
9
|
Lisle JW, Choi JY, Horton JA, Allen MJ and
Damron TA: Metastatic osteosarcoma gene expression differs in vitro
and in vivo. Clin Orthop Relat Res. 466:2071–2080. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Skarsgard LD: Radiobiology with heavy
charged particles: A historical review. Phys Med. 14(Suppl 1):
1–19. 1998.
|
|
11
|
Okayasu R: Repair of DNA damage induced by
accelerated heavy ions - a mini review. Int J Cancer. 130:991–1000.
2012. View Article : Google Scholar
|
|
12
|
Kamada T, Tsujii H, Blakely EA, Debus J,
De Neve W, Durante M, Jäkel O, Mayer R, Orecchia R, Pötter R, et
al: Carbon ion radiotherapy in Japan: An assessment of 20 years of
clinical experience. Lancet Oncol. 16:e93–e100. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kanai T, Endo M, Minohara S, Miyahara N,
Koyama-ito H, Tomura H, Matsufuji N, Futami Y, Fukumura A, Hiraoka
T, et al: Biophysical characteristics of HIMAC clinical irradiation
system for heavy-ion radiation therapy. Int J Radiat Oncol Biol
Phys. 44:201–210. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang W, Tanaka M, Sugimoto Y, Takigawa T
and Ozaki T: Carbon-ion radiotherapy of spinal osteosarcoma with
long-term follow. Eur Spine J. 25(Suppl 1): 113–117. 2016.
View Article : Google Scholar
|
|
15
|
Matsunobu A, Imai R, Kamada T, Imaizumi T,
Tsuji H, Tsujii H, Shioyama Y, Honda H and Tatezaki S; Working
Group for Bone and Soft Tissue Sarcomas: Impact of carbon ion
radiotherapy for unresectable osteosarcoma of the trunk. Cancer.
118:4555–4563. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Futamura G, Kawabata S, Siba H, Kuroiwa T,
Suzuki M, Kondo N, Ono K, Sakurai Y, Tanaka M, Todo T, et al: A
case of radiation-induced osteosarcoma treated effectively by boron
neutron capture therapy. Radiat Oncol. 9:2372014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lee DH, Seo SH and Ji YH: Characteristics
of radiation generated by BNCT irradiator of Hanaro nucleasr
reactor. J Korean Soc Ther Radiol Oncol. 24:s1582006.
|
|
18
|
Eom KY, Wu HG, Park HJ, Huh SN, Ye SJ, Lee
DH and Park SW: Evaluation of biological characteristics of neutron
beam generated from MC50 cyclotron. J Korean Soc Ther Radiol Oncol.
24:280–284. 2006.
|
|
19
|
Luu HH, Kang Q, Park JK, Si W, Luo Q,
Jiang W, Yin H, Montag AG, Simon MA, Peabody TD, et al: An
orthotopic model of human osteosarcoma growth and spontaneous
pulmonary metastasis. Clin Exp Metastasis. 22:319–329. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Song WS, Jeon DG, Cho WH, Kong CB, Cho SH
and Lee SY and Lee SY: Spontaneous necrosis and additional tumor
necrosis induced by preoperative chemotherapy for osteosarcoma: A
case-control study. J Orthop Sci. 20:174–179. 2015. View Article : Google Scholar
|
|
21
|
Kong CB, Byun BH, Lim I, Choi CW, Lim SM,
Song WS, Cho WH, Jeon DG, Koh JS, Yoo JY, et al: 18F-FDG
PET SUVmax as an indicator of histopathologic response after
neoadjuvant chemotherapy in extremity osteosarcoma. Eur J Nucl Med
Mol Imaging. 40:728–736. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
McIlwain DR, Berger T and Mak TW: Caspase
functions in cell death and disease. Cold Spring Harb Perspect
Biol. 5:a0086562013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Los M, Mozoluk M, Ferrari D, Stepczynska
A, Stroh C, Renz A, Herceg Z, Wang ZQ and Schulze-Osthoff K:
Activation and caspase-mediated inhibition of PARP: A molecular
switch between fibroblast necrosis and apoptosis in death receptor
signaling. Mol Biol Cell. 13:978–988. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Goodhead DT, Thacker J and Cox R: Weiss
Lecture. Effects of radiations of different qualities on cells:
Molecular mechanisms of damage and repair. Int J Radiat Biol.
63:543–556. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Goodhead DT: Energy deposition stochastics
and track structure: What about the target? Radiat Prot Dosimetry.
122:3–15. 2006. View Article : Google Scholar
|
|
26
|
Anderson RM, Marsden SJ, Wright EG, Kadhim
MA, Goodhead DT and Griffin CS: Complex chromosome aberrations in
peripheral blood lymphocytes as a potential biomarker of exposure
to high-LET alpha-particles. Int J Radiat Biol. 76:31–42. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Brenner DJ and Ward JF: Constraints on
energy deposition and target size of multiply damaged sites
associated with DNA double-strand breaks. Int J Radiat Biol.
61:737–748. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tanaka K, Gajendiran N, Endo S, Komatsu K,
Hoshi M and Kamada N: Neutron energy-dependent initial DNA damage
and chromosomal exchange. J Radiat Res (Tokyo). 40(Suppl): S36–S44.
1999. View Article : Google Scholar
|