Introduction
Cancer, a serious public health problem worldwide,
is responsible for countless deaths each year and is currently
considered the second leading cause of death on the planet
(1–3). In addition, various malignant tumor
types do not have effective treatment (4–8), due
to the ability of tumor cells to evade death, by presenting changes
in apoptosis pathway protein levels (9–11).
Changes in other cellular pathway proteins such as Pi3k/Akt/mTOR
and MAPK/ERK, which are highly dysregulated in malignant tumors,
also corroborate the ability of malignant cells to evade apoptosis
death, contributing to chemotherapy resistance (12–14).
The expression of multi-drug resistance genes, such as MDR1, has
been implicated as the main cause of chemoresistance (15,16).
Furthermore, conventional chemotherapeutic treatments are known for
their side-effects on non-tumor cells, such as the strong oxidative
stress (17–19).
Moreover, there is a need for alternative treatments
that promote apoptosis in tumor cells and yet do not negatively
affect normal cells. In this regard, GNPs have been highlighted in
the literature as promising agents with large pooling surfaces for
various drugs (20), which
concentrate on tumor tissues (21,22),
are resistant to corrosion and present low toxicity to the
biological system (23–25). In addition, various studies
reported an effective antitumor action with non toxicity to normal
cells (26–29). The antihypertensive carvedilol is
often used as a non-selective inhibitor of adrenergic receptors
(30), is known for its
cardiovascular and antioxidant benefits (31–34)
and has recently presented good antitumor activity, such as growth
inhibition of neuroblastoma cell lines (35) and rat glioma cell line (36), suppressing migration and invasion
of malignant breast cells (37),
preventing carcinogenesis in rat epidermal lineages (38) and promoting apoptosis in tumoral
hepatic and oral cell linages (39,40).
The discovery of new substances and combinations is fundamental to
the process of establishing new treatments against cancer (41–43).
Combining substances has proven to be extremely effective due to
the lower doses used, the decreased adverse effects, and the
possibility of acting on different signaling pathways (44–50).
In addition, the discovery of substances that promote inhibition of
dysregulated pathways such as Pi3k/Akt/mTOR and MAPK/Erk would
positively modulate apoptosis in tumor cells (12,51,52).
In the present study, we investigated the effects of combined
carvedilol and GNPs action on both tumor and non-tumor cells.
Materials and methods
Reagents
The reagents were purchased as indicated: Dulbecco's
modified Eagle's medium (DMEM; Life Technologies, Grand Island, NY,
USA); 10% (v/v) heat-inactivated fetal bovine serum (FBS; Cultilab
Materiais para Cultura de Células Ltda, Campinas, Brazil);
trypsin/EDTA (ethylenediaminetetraacetic acid) (Gibco-BRL, Life
Technologies, Grand Island, NY, USA); cisplatin (citoplax, 50 mg;
Bergamo, Taboão da Serra, Brazil); gold nanoparticles GNPs
(Institute of Chemical, UFRN, Natal, Brazil); carvedilol
(Farmafórmula, Natal, Brazil); gold (III) chloride (30% wt. in
HCl), sodium hydroxide, glycerol, and polyvinylpyrrolidone (PVP,
molecular weight, 10,000 Da) were products from Sigma-Aldrich. The
synthesis and characterization of GNPs was the described by de
Araújo et al (53). GNPs,
carvedilol and cisplatin solutions were filtered using a 0.22-mm
minipore membrane.
Cell culture
The human cell lines hepatocellular carcinoma
(HepG2) and human non-cancerous renal cell line (HEK-293) were
purchased from the Culture Collection of the Federal University of
Rio de Janeiro (RJCB Collection, Rio de Janeiro, Brazil). HepG2 and
Hek-293 cells were maintained in DMEM supplemented with 10% (v/v)
heat-inactivated FBS.
Cell viability
In order to determine GNP and carvedilol doses to
promote and maintain low inhibition in cancer cells, cell viability
was determined by trypan blue exclusion assay. The viability was
determined at 24 and 48 h for HepG2 (1×105 cells) at
different concentrations of GNPs (1–50 µg/ml, aqueous
suspension) and carvedilol [1.5–300 µM, dissolved in
dimethyl sulfoxide (DMSO) 1%]. The cells were placed into 6-well
plates. Briefly, cell aliquots were mixed with the same volume 0.5%
(w/v) trypan blue and incubated at room temperature for 5 min. The
number of viable cells was calculated using a hemocytometer.
Annexin V and propidium iodide
staining
The apoptotic assay was conducted according to
Araújo Jr et al (54).
HepG2 and HEK-293 were plated in 6-well plates (2×105
cells/well) with 2 ml medium/well. After 24 h, concentrations of
GNPs (3 and 6.25 µg/ml), cisplatin (15 µg/ml) and
carvedilol (1.5 and 3 µM) were added (24 and 48 h),
respectively. In parallel, control cells were maintained in culture
medium without GNPs, carvedilol or cisplatin. For observation of
combined action, the cells were treated with GNPs (3 and 6.25
µg/ml) and at 24 h treated with carvedilol (1.5 and 3
µM). After another 24 h, they were analyzed. The cells were
then assayed using the Annexin V-FITC/PI apoptosis detection kit I
(BD Biosciences, San Diego, CA, USA). Annexin V-FITC and propidium
iodide (PI) were added to the cellular suspension according to the
manufacturer's instructions. A total of 1×106 cells from
each sample was then analyzed by FACSCalibur cytometer (BD
Biosciences, Franklin Lakes, NJ, USA), and FlowJo software (BD
Biosciences). Annexin V-FITC-positive/PI-negative cells were
identified as cells in the early stages of apoptosis, while Annexin
V-FITC-positive/PI-positive cells were identified as cells in the
late stages of apoptosis, or as cells undergoing necrosis.
Glutathione (GSH) levels
Antioxidant GSH levels in cell lines were measured
[adapted from Rahman et al (55) and Costa et al (56)]. HepG2 and Hek-293 were plated in
6-well plates (2×105 cells/well) with 2 ml medium/well.
After 24 h, GNPs (6.25 µg/ml) was added and after 24 h, 3
µM carvedilol. A homogenate of cells (100 µl of cell
in 500 µl EDTA 0.02 M) were added to 320 µl of
distilled water and 80 µl of 50% trichloroacetic acid (TCA).
Samples were centrifuged at 3,000 rpm for 15 min at 4°C. The
supernatant (100 µl) was added to 200 µl of 0.4 M
Tris buffer at pH 8.9 and 20 µl of 0.01 M 5,5′-dithiobis
(2-nitrobenzoic acid) (DTNB). The absorbance of each sample was
measured at 420 nm, in a spectrophotometric/microplate reader
Polaris and the results were reported as units of GSH per
milligram.
Malondialdehyde levels
Malondialdehyde (MDA) is an end product of lipid
peroxidation. To quantify the increase in free radicals in
non-cancer (HEK-293) and cancer (HepG2) cells, MDA content was
measured via the assay described by Esterbauer and Cheeseman
(57). Cell samples were suspended
in buffer; Tris HCl 1:5 (w/v), and minced with scissors for 15 sec
on an ice-cold plate. The resulting suspension was homogenized for
2 min with an automatic Potter homogenizer and centrifuged at
11,000 rpm at 4°C for 10 min. The supernatants were assayed in
order to determine the MDA content. The absorbance of each sample
was measured at 586 nm. The results are expressed as nanomoles of
MDA per cell.
Immunofluorescence, Bcl-2, MAPK/ERK,
caspase-3 and caspase-8 activity
HepG2 cells were plated on glass coverslips in
24-well plates (5×104 cells/well). After 24 h, they were
treated with the GNPs (6.25 µg/ml), cisplatin (15
µg/ml) and carvedilol (3 µM) for 48 h. For combined
action, we used GNPs (6.25 µg/ml) + carvedilol (3
µM). The cells were then washed, and fixed with
paraformaldehyde, permeabilized by Triton-X, and incubated with
anti-Bcl-2 mouse polyclonal antibody, rabbit polyclonal
anti-caspase-3 antibody (Abcam, San Francisco, CA, USA), rabbit
anti-caspase-8 monoclonal antibody (Santa Cruz Biotechnology, Santa
Cruz, CA, USA), and monoclonal mouse antibody anti-MAPK/ERK
(Invitrogen, Carlsbad, CA, USA) diluted 1:500 in phosphate-buffered
saline (PBS) containing bovine serum albumin (BSA; 5%; Life
Technologies do Brasil Ltda, São Paulo, Brazil) for 1 h at RT in a
humid atmosphere. The primary antibody was detected with Alexa
Fluor 488 goat anti-rabbit or anti-mouse secondary antibody
(Abcam), and 4′,6-diamidino-2-phenylindole (Life Technologies do
Brasil Ltda) was used for nuclear staining. The immunostained
coverslips were examined under Axio Observer z.1, inverted
fluorescence and brightfield.
Fluorescent images were obtained on a Carl Zeiss
Laser Scanning Microscope (LSM 710, 20X objectives; Carl Zeiss,
Oberkochen, Germany). Negative controls and treated groups were
included in each batch of samples. Cell reactivity in all groups
(negative, GNPs, carvedilol, GNPs + carvedilol and cisplatin) was
assessed by computerized densitometric analysis of the captured
digital images with the aforementioned immunofluorescence
microscope. Average densitometric values were calculated in ImageJ
software (http://rsb.info.nih.gov/ij/).
Contrast index measurements were obtained from the formula
[(selected area × 100)/total area] after removal of background in
regions of interest (three samples per group).
Real-time PCR
HepG2 cells were plated in 6-well plates
(2×105 cells/well) with 2 ml medium/well. After 24 h,
concentrations of gold nanoparticles (6.25 µg/ml), cisplatin
(15 µg/ml) and carvedilol (3 µM) were added for 48 h.
For combined action, GNPs (6.25 µg/ml) + (3 µM)
carvedilol was used. The cells were collected with cell scrapers
and total RNA was isolated from cells using TRIzol reagent. The
total RNA was extracted from cell samples using RNeasy Mini kit
(Qiagen, Tokyo, Japan) from QIAcube following the manufacturer's
guidelines. The total RNA extracted underwent reverse transcriptase
activity using the High capacity RNA-to-cDNA kit (Applied
Biosystems, Ltd., Tokyo, Japan). Real-time quantitative PCR
analyses of EGFR, Akt, mTOR, survivin, MDR-1, FADD, Apaf-1 and
GAPDH mRNAs were performed with SYBR-Green Mix in the Applied
Biosystems® 7500 FAST system (Applied Biosystems, Foster
City, CA, USA), according to a standard protocol with the following
primers: GAPDH (forward, 5′-AAC TTT GGC ATC GTG GAA GG-3′
and reverse, 5′-GTG GAT GCA GGG ATG ATG TTC-3′, annealing primer
temperature, 60°C); EGFR (forward, 5′-TGA TAG ACG CAG ATA GTC
GCC-3′ and reverse, 5′-TCA GGG CAC GGT AGA AGT TG-3′, annealing
primer temperature, 56.6°C); Akt (forward, 5′-ACG GCA TGG ACT TTA
CCA AG-3′ and reverse, 5′-GCG GGT GAA AGA CAG GAA TA-3′, annealing
primer temperature, 55°C); mTOR (forward, 5′-TTG AGG TTG CTA TGA
CCA GAG AGA A-3′ and reverse, 5′-TTA CCA GAA AGG ACA CCA GCC AAT
G-3′, annealing primer temperature, 58.3°C); survivin (forward,
5′-TAC AGC TTC GCT GGA AAC CT-3′ and reverse, 5′-AGC CCG GAT GAT
ACA AAC AG-3′, annealing primer temperature, 55.6°C); MDR1
(forward, 5′-GTG TGG TGA GTC AGG AAC CTG TAT-3′ and reverse, 5′-TCT
CAA TCT CAT CCA TGG TGA CA-3′, annealing primer temperature, 57°C);
FADD (forward, 5′-TCT CCA ATC TTT CCC CAC AT-3′ and reverse, 5′-GAG
CTG CTC GCC TCC CT-3′, annealing primer temperature, 58.7°C); and
Apaf-1 (forward, 5′-CCT CTC ATT TGC TGA TGT CG-3′ and reverse,
5′-TCA CTG CAG ATT TTC ACC AGA-3′, annealing primer temperature,
56.9°C). The experiments were performed in triplicate. The standard
PCR conditions were as follow: 50°C for 2 min and 95°C for 10 min,
followed by 40 of 30-sec cycles at 94°C, a variable annealing
primer temperature for 30 sec and at 72°C for 1 min. Mean Ct values
were used to calculate the relative expression levels of the target
genes for the experimental groups as relative to those in the
negative control group; expression data were normalized relative to
the housekeeping gene GAPDH using the 2−ΔΔCt
formula.
Western blot analysis
Cells (HepG2) were lysed in buffer [Tris-HCl 50 mM,
NaCl 150 mM, Triton X-100 1%, EDTA 1 mM, sodium pyrophosphate 20
mM, pH 7.4 containing a protease inhibitors cocktail (Roche), NaF
(10 mM), DTT (1 mM), PMSF (0.1 mM) and sodium vanadate (1 mM)] on
ice. To confirm equal loadings, total protein concentration was
determined using the Bradford method (Bio-Rad Laboratories,
Hercules, CA, USA). Proteins were resolved using SDS-PAGE and then
transferred to a polyvinylidene diflouride (PVDF) membrane.
Non-specific binding sites on the membrane were blocked using 5%
non-fat skimmed milk and incubated with the primary antibody
anti-Akt (1:500; Abcam), anti-mTOR (1:500; Abcam), and MAPK/ERK
(1:200; Abcam), overnight at 4°C, followed by incubation with the
appropriate secondary antibodies: Akt α-rat peroxidase 1:1,000;
mTOR-rabbit peroxidase 1:2,000 and MAPK/ERK α-rat peroxidase
1:1,000. Proteins were detected using the ECL Plus kit
(Perkin-Elmer, San Jose, CA, USA).
Transmission electronic microscopy
(TEM)
Cells (HepG2) at a density of 3×105 were
plated into (GNPs sensitized and treated) 6-well plates, and after
48 h were collected with trypsin, centrifuged at 1,500 rpm for 5
min, and washed with PBS. The cell pellet was fixed with 2.5%
glutaraldehyde + paraformaldehyde 4% + sodium cacodylate buffer
(0.1 M) for 4 h at 4–8°C. Afterwards, the samples were washed in
0.05 M sodium cacodylate (3×30 min), post-fixed in 1%
OsO4 + 1% potassium ferrocyanide (2 h), washed again
(3×) with 0.05 M sodium cacodylate (3×30 min), and then serially
dehydrated in ethanol 50, 70, 90 and 100, for 30 min each.
Polymerization of the resin was performed at 60°C for 48 h.
Finally, ultramicrotomy was performed followed by staining (uranyl
acetate 1% + 1% lead citrate 1 h), and electron microscope
visualization (Tescan transmission, Vega 3 model).
Statistical analysis
All experiments were performed in triplicate, and
the significant differences between the groups were calculated
using the analysis of variance and the Bonferroni's test, as
indicated. A P<0.05 was considered statistically
significant.
Results
Cell viability
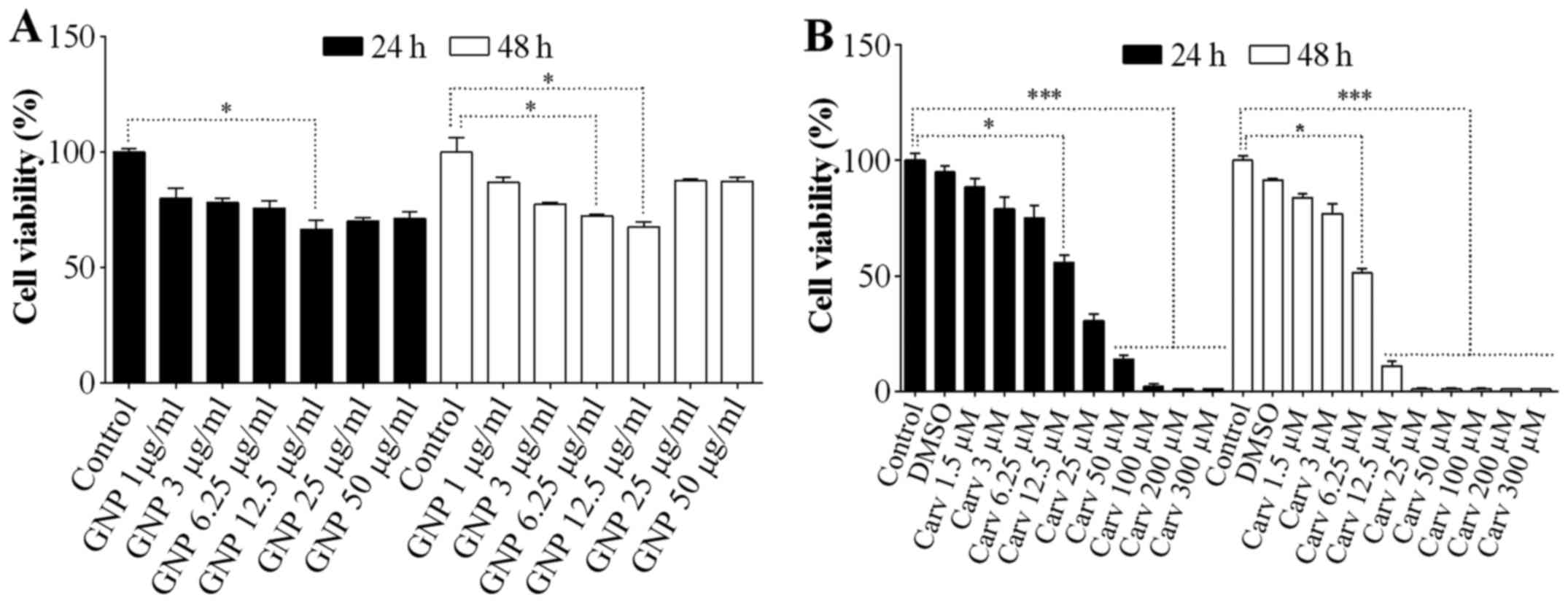
The GNPs doses 1, 3 and 6.25 µg/ml, at 24 h
(Fig. 1A) promoted low inhibition
of cellular viability. The other GNPs doses promoted greater
cellular growth inhibition, when compared to lower doses. The most
significant was 12.5 µg/ml (P<0.05). At 48 h, GNPs doses
of 3 and 6.25 µg/ml maintained low inhibition similar to
that at 24 h (Fig. 1A), and for
GNPs doses of 1, 25 and 50 µg/ml, there was cellular growth
when compared to the same doses at 24 h. The carvedilol doses that
promoted low inhibition of cellular viability were 1.5 and 3
µM at 24 h (Fig. 1B). The
other carvedilol doses promoted greater cellular growth inhibition
(P<0.001). At 48 h, the 1.5 and 3 µM carvedilol doses
maintained low inhibition (Fig.
1B). The other doses of carvedilol promoted greater cellular
growth inhibition (P<0.001). DMSO at 1% was used as the
carvedilol vehicle.
Detection of apoptosis, by flow
cytometer
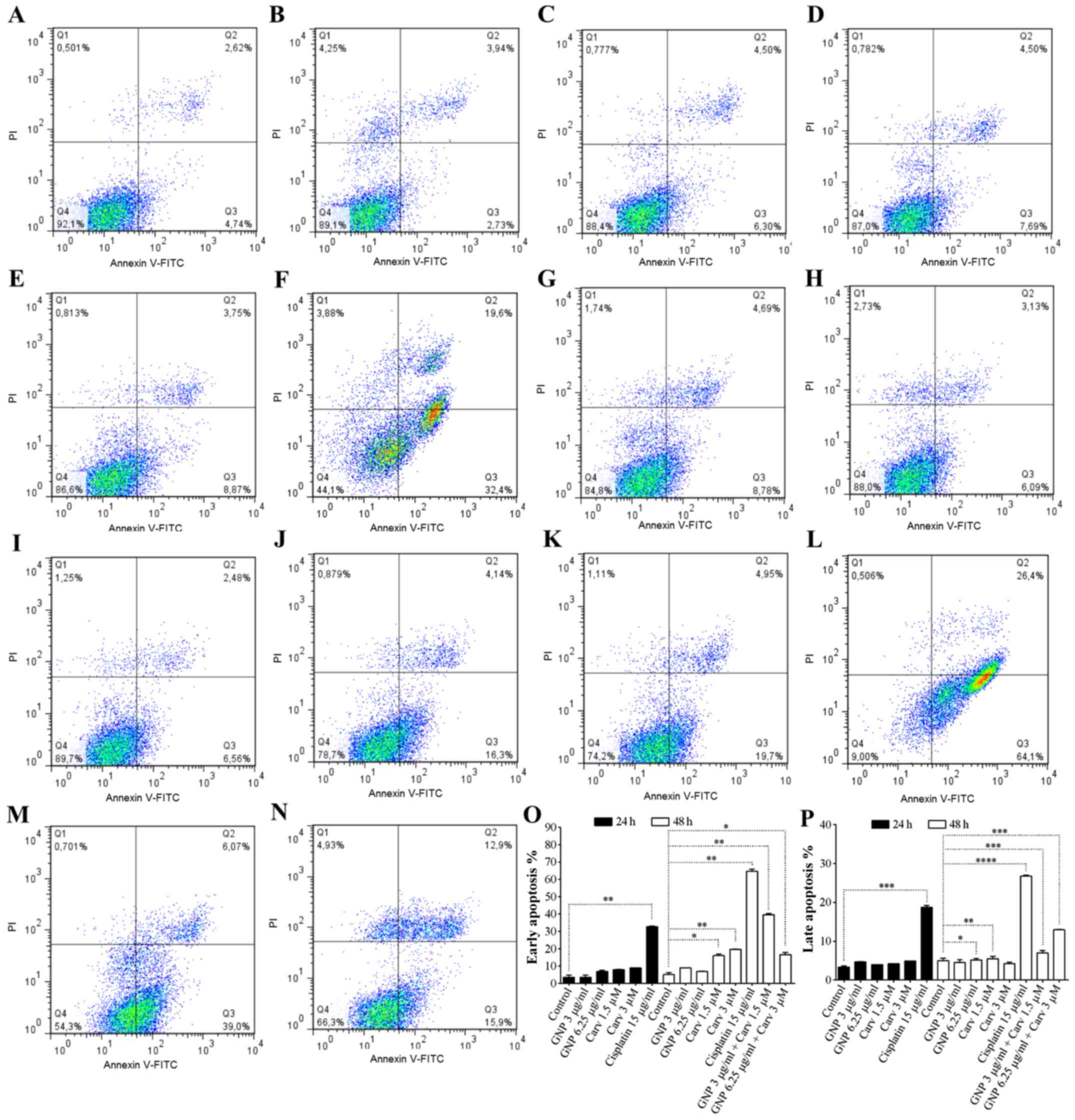
For HepG2 cells, combined treatment (GNP 6.25
µg/ml + carvedilol 3 µM) induced early (P<0.05)
and late (P<0.001) apoptosis at 48 h (Fig. 2O–P). The isolated dose of
carvedilol (3 µM) induced early apoptosis at 48 h (Fig. 2O; P<0.01). GNPs (6.25
µg/ml) induced late apoptosis at 48 h, P<0.05. Doses of
GNPs and carvedilol did not induce statistically significant
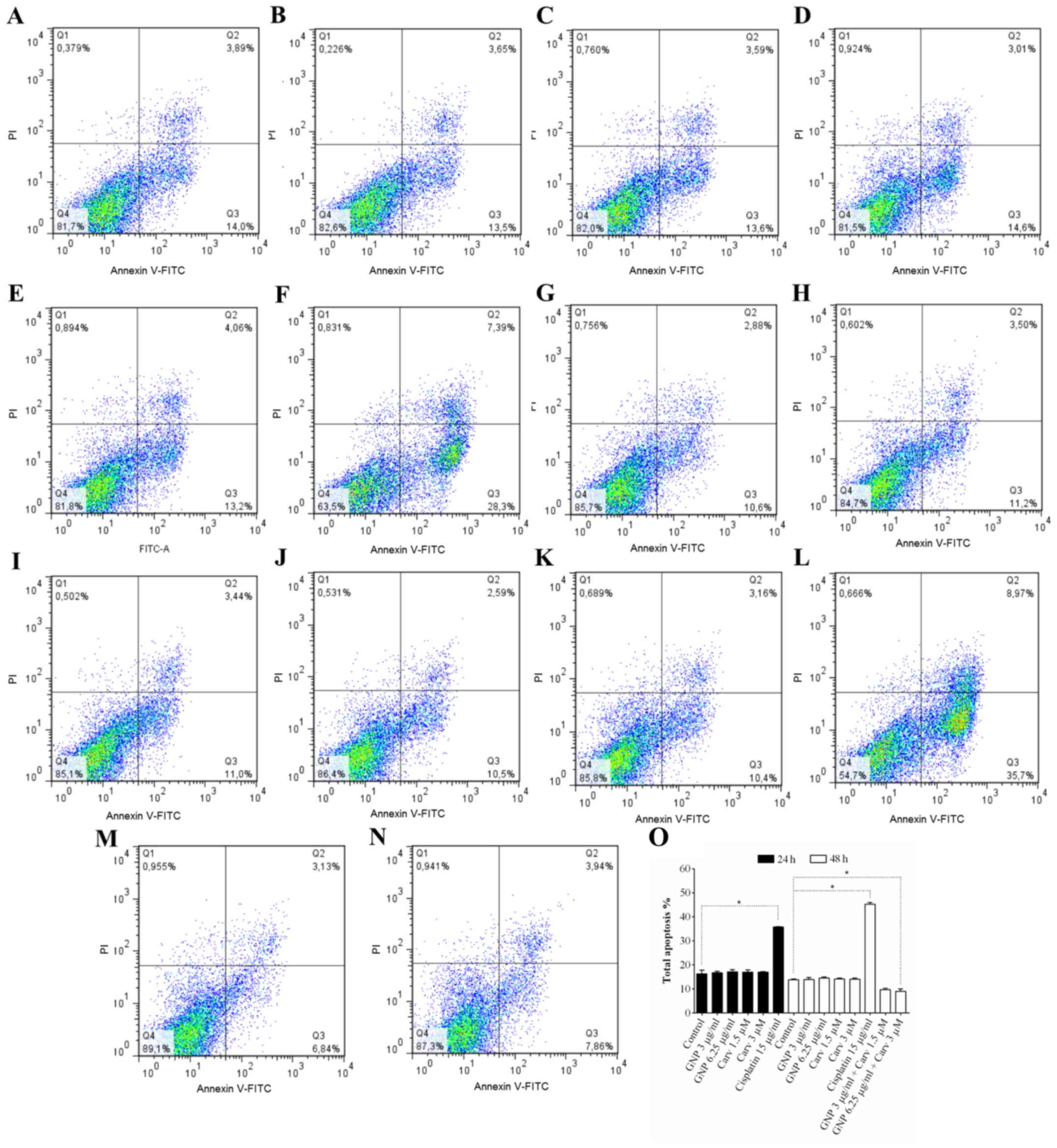
apoptosis in non-tumor cells (Fig.
3). Combined treatment (GNP 6.25 µg/ml + carvedilol 3
µM) promoted cytoprotection (P<0.05; Fig. 3O). When HepG2 and HEK-293 cells
were treated with 15 µg/ml cisplatin, apoptosis was detected
at 24 and 48 h after treatment (Figs.
2F and L and 3F and L,
respectively). The goal of using a non-tumor cell is to have as a
control a non-tumoral lineage to analyze its behavior against the
treatment. HEK-293 is used to study cytotoxicity because of its
reliable growth despite its karyotype complex with alterations.
This makes toxicity studies more consistent (54,58–62).
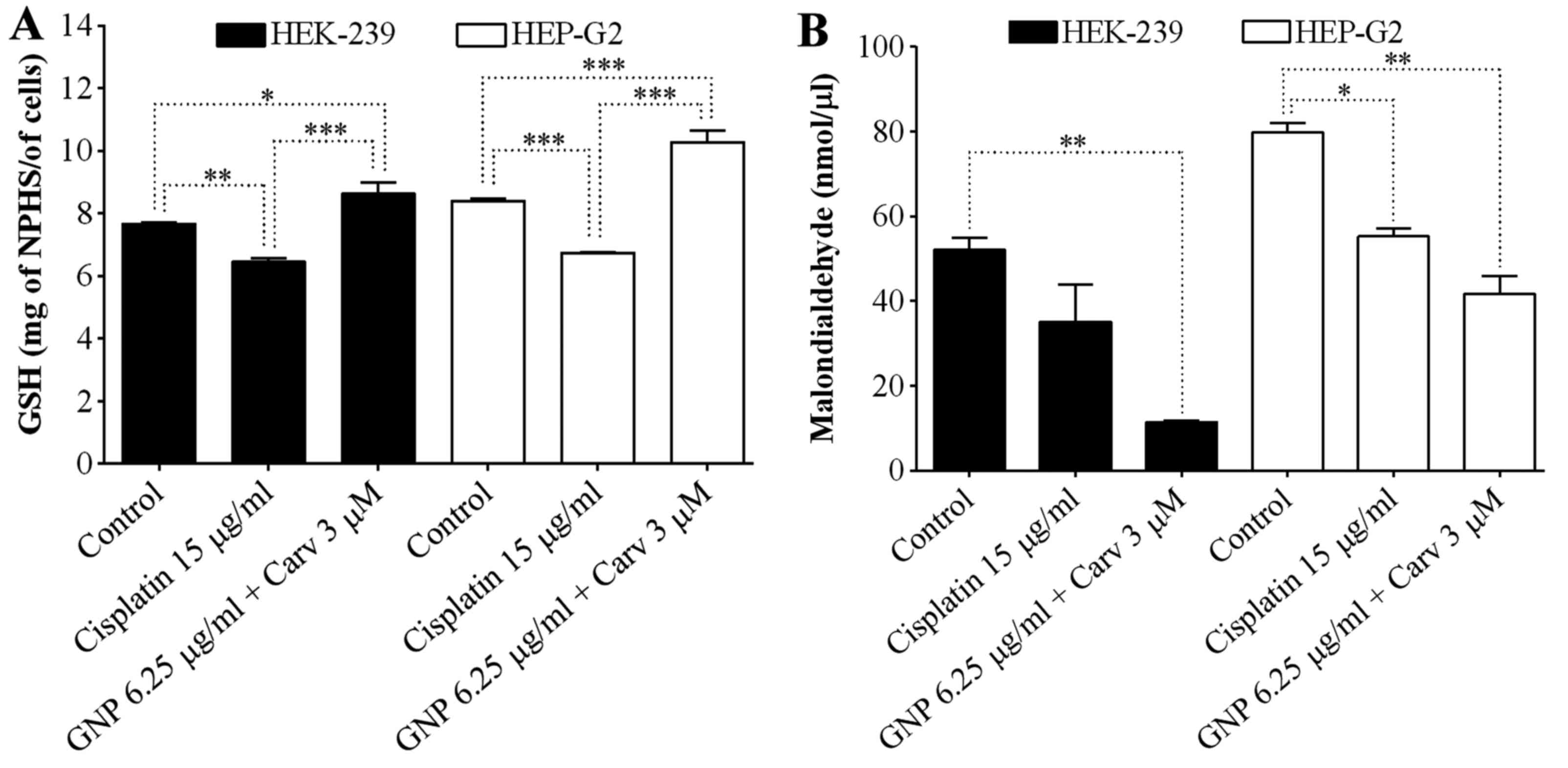
Oxidative stress
The reduced glutathione and lipid peroxidation
levels were observed from GSH and MDA assays, respectively. Due to
the fact that the combined treatment with GNPs (6.25 µg/ml)
+ carvedilol (3 µM), at 48 h increased the protection for
HEK-293, it was observed that the levels of GSH increased
statistically significantly for both HEK-293 (P<0.05) and HepG2
(P<0.001), as shown in Fig. 4A.
Cisplatin promoted reduction of GSH levels in both cell lines.
Regarding lipid peroxidation for the non-tumoral lineage, there was
a great reduction in malondialdehyde levels using the combined
treatment with GNPs (6.25 µg/ml) + carvedilol (3 µM)
(P<0.01), overcoming the control and cisplatin treated groups.
This same result was observed for the HepG2 tumor cells (Fig. 4B).
Immunofluorescence of caspase-3,
caspase-8, Bcl-2 and MAPK/ERK
After combined treatment with GNPs (6.25
µg/ml) + carvedilol (3 µM), caspase-3 and caspase-8
marking was noted in all treated groups (Fig. 5A). Densitometric analysis confirmed
significant increases in caspase-3 (P<0.01; Fig. 5B), and caspase-8 (P<0.001;
Fig. 5C). In addition, there was
no statistically significant change in Bcl-2 expression for the
group treated with GNPs and carvedilol (Fig. 5D; P>0.05). There was a decrease
in MAPK/ERK immunoreactivity in the HepG2 cells treated with GNPs
and carvedilol and GNP (6.25 µg/ml) + carvedilol (3
µM) for 48 h (P<0.0001; Fig.
5E).
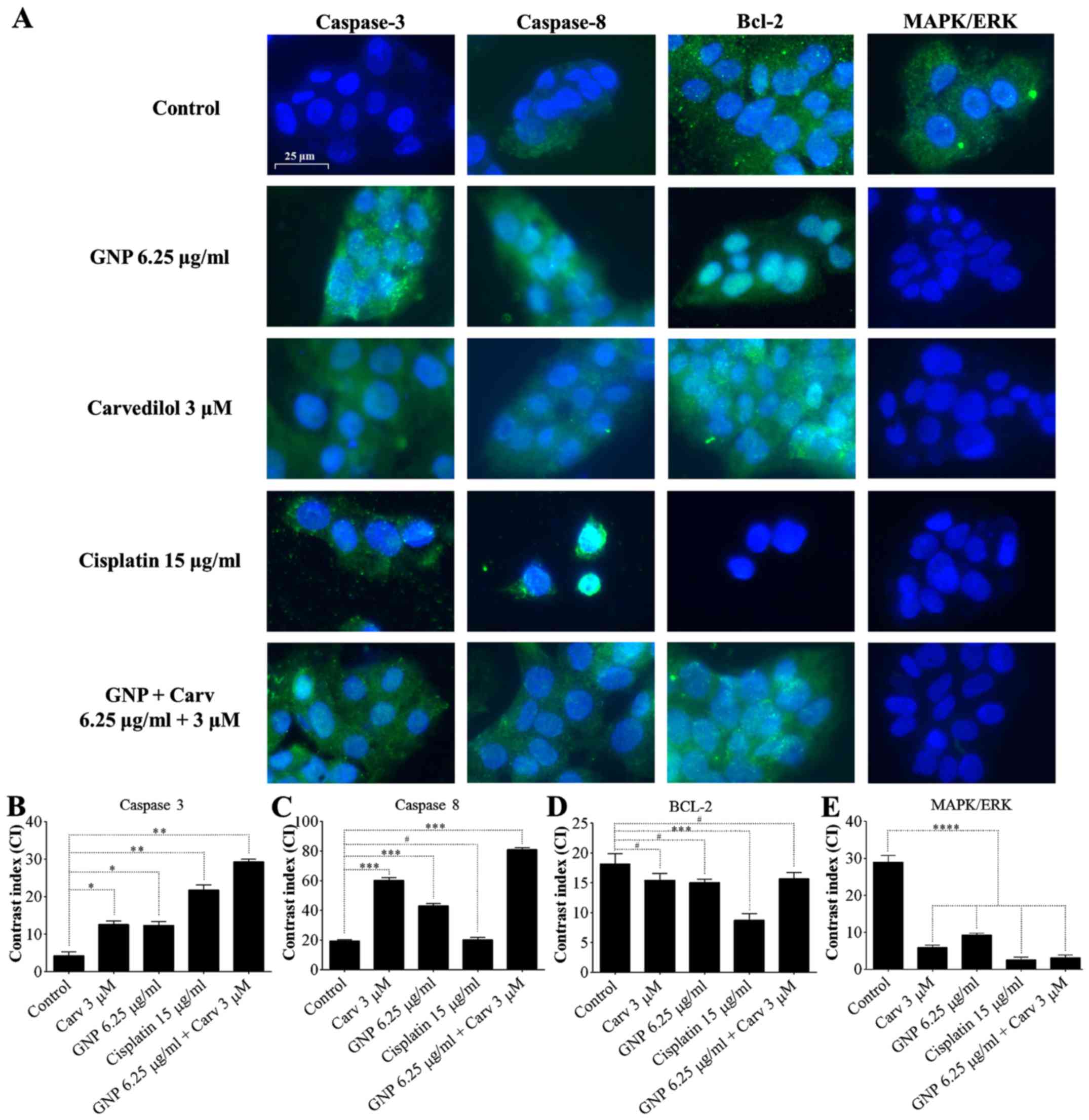 | Figure 5Detection of caspase-3, caspase-8,
Bcl-2 and MAPK/ERK. HepG2 cells stained with DAPI (blue),
anti-caspase-3, anti-caspase-8, anti-Bcl-2, and anti-MAPK/ERK
antibodies (green) (A). Caspase-3 and caspase-8 were detected in
all treated groups, yet MAPK/ERK was not. Bcl-2 did not alter
expression. Contrast index for caspase-3, *P<0.05 and
**P<0.01 (B); caspase-8, ***P<0.001 and
#P>0.05 (C); Bcl-2, #P>0.05 and
***P<0.001 (D) and MAPK/ERK, ****P<0.0001 (E). All
groups treated with GNPs + Carv showed high immunoreactivity for
caspase-3, caspase-8 and low immunoreactivity for MAPK/ERK. |
Gene and protein expression by RT-PCR and
western blot analysis
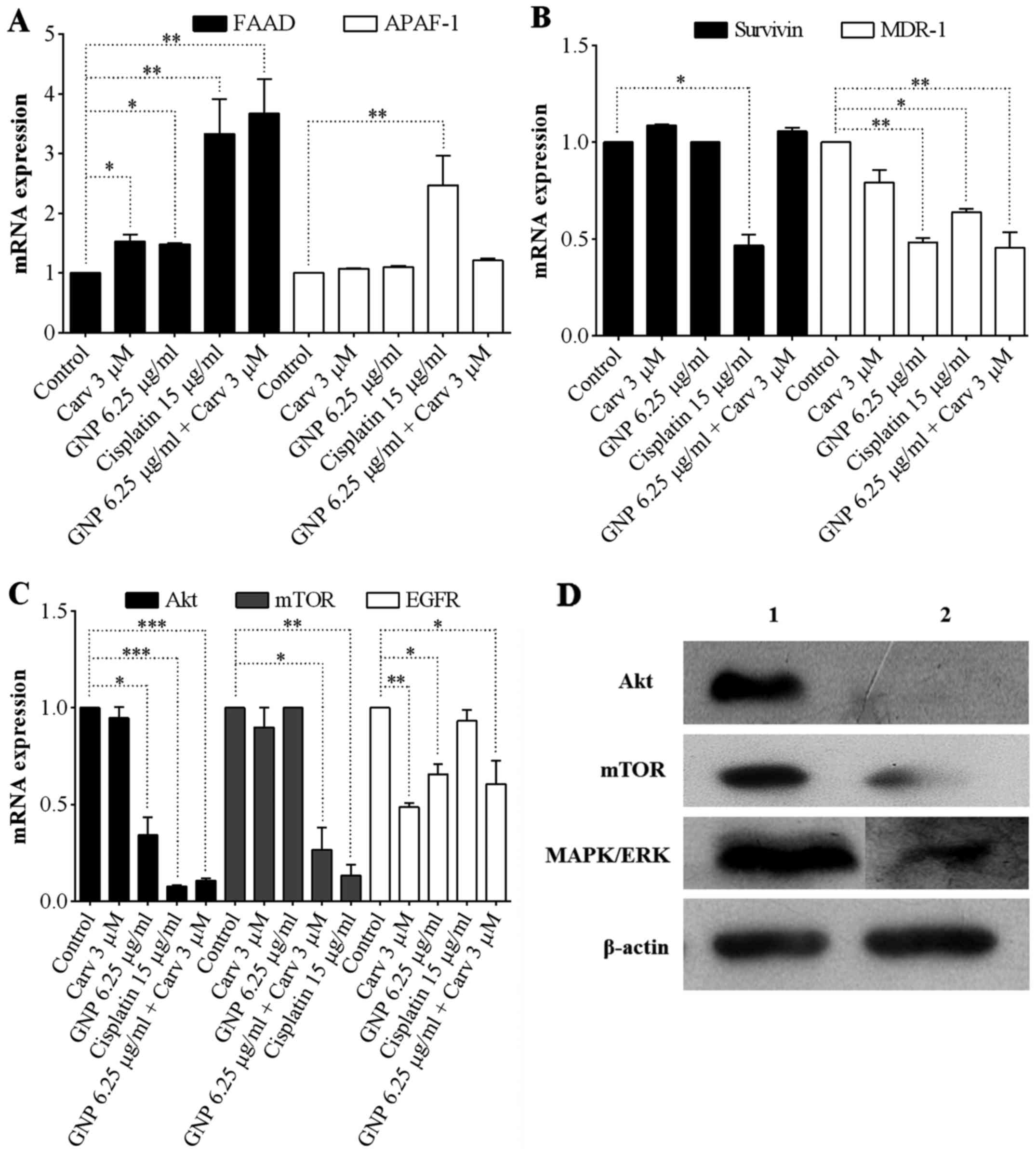
From the gene expression analysis, it was observed
that FADD elevation was statistically significant for all groups,
those treated separately with GNPs (6.25 µg/ml) and
carvedilol (3 µM) (P<0.05), and in combined treatment
GNPs (6.25 µg/ml) + carvedilol (3 µM) (P<0.01),
yet not observed for APAF-1 levels (Fig. 6A). In relation to survivin levels,
no group treated with GNPs and carvedilol demonstrated a reduction,
meanwhile MDR-1 levels showed statistically significant reductions
for those treated with GNPs (6.25 µg/ml), and for combined
treatment GNPs (6.25 µg/ml) + carvedilol (3 µM)
(P<0.01; Fig. 6B). Regarding
the Akt, mTOR and EGFR levels, the groups treated with combined
treatment GNPs (6.25 µg/ml) + carvedilol (3 µM)
showed statistically significant expression declines P<0.001,
P<0.05 and P<0.05, respectively (Fig. 6C). The groups treated with
carvedilol (3 µM) showed statistically significant
reductions only for EGFR (P<0.01); and those treated with GNP
showed statistically significant reductions only for Akt
(P<0.05) and EGFR (P<0.05).
The anti-apoptotic protein levels after combined
treatment of GNPs (6.25 µg/ml) + carvedilol (3 µM)
are highlighted in Fig. 6D. The
results showed a large reduction of Akt and mTOR. MAPK/ERK has a
good reduced protein expression when compared to the control group.
β-actin was used as internal control.
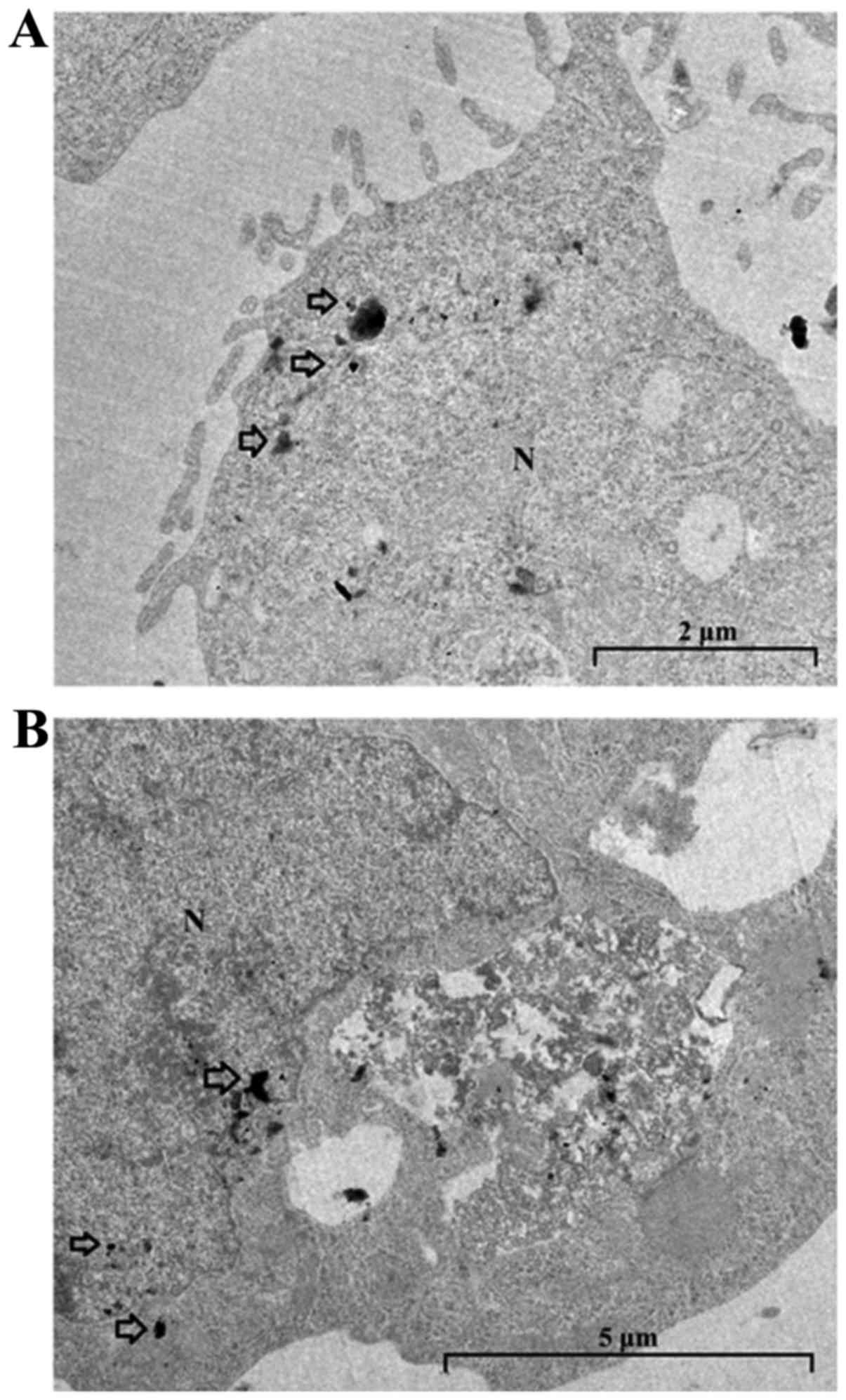
Transmission electron microscopy
TEM was performed in order to ascertain locations
where gold nanoparticles would concentrate, applied without
carvedilol (Fig. 7A) and in
combination with carvedilol (Fig.
7B). Applied alone, GNPs (6.25 µg/ml) was concentrated
in the vicinity of the plasma membrane. In the combined treatment
with GNPs (6.25 µg/ml) + carvedilol (3 µM), GNPs
displayed intra-nuclear and perinuclear concentration.
Discussion
Data from the literature demonstrate the importance
of using low doses for drug combination studies that aim to reduce
side-effects and increase the field of action in different
signaling pathways (63–65). In the present study, low doses of
GNPs (3 and 6.25 µg/ml) and carvedilol (1.5 and 3 µM)
were selected using the cell viability test, in order to use them
in combination. Studies in the literature demonstrate that low
doses of GNPs are more effective in inhibiting cell proliferation
than larger doses (53,66). In relation to carvedilol, very high
doses cause high inhibition of cell growth (67); a risk for a combined treatment. The
pro-apoptotic activities of GNPs, carvedilol, and their combined
use were analyzed by flow cytometry, both for tumor and non-tumoral
lines. The combination of GNPs (6.25 µg/ml) + carvedilol (3
µM) promoted significant initial and late apoptosis in
hepatic tumor cells, while promoting reduction of total apoptosis
in the non-tumoral lineage. Studies in the literature report the
ability of GNPs to induce apoptosis in several tumor cell lines
without interfering with non-tumoral lineages (27,29,68,69).
Carvedilol has itself also been shown to be an inducer of apoptosis
in tumor cells and to protect normal cells (36,70,71).
The cytoprotective effect of the combined treatment
for non-cancerous cells was evidenced through GSH dosage; an
important balancing antioxidant system peptide (72,73).
After treatment, GSH levels were elevated, which is important for
reducing oxidative stress, which is related to apoptosis induction
in several tissues (74–77). In addition, GNPs and carvedilol
have the ability to promote elevation of GSH levels (78–81).
Furthermore, a decrease in MDA levels were evidenced, an indirect
biomarker for oxidative stress, as related to plasma membrane
damage (82–84). This is a significant result, since
one of the main side-effects of chemotherapy is lipid peroxidation
(85,86).
In the tumoral lineage, we observed GSH level
elevation and decreased MDA levels, demonstrating the antioxidant
action of combined treatment when compared to cisplatin, which is
known for its strong oxidative stress promotion (18,19).
This result was highly positive, since tumor cells have high rates
of reactive oxygen species, which trigger the activation of
proteins such as Pi3K, Akt and MAPK/ERK; responsible for both cell
proliferation and survival (87,88).
The pro-apoptotic activity of the treatment was
confirmed by the positive immunoreactivity of caspase-3 and
caspase-8, proteins involved in the extrinsic apoptosis pathway
(89), these were the most
significant in the group treated with the combination. In addition,
after treatments with GNPs (6.25 µg/ml) + carvedilol (3
µM), there was a decrease in immunoreactivity for the
MAPK/ERK, anti-apoptotic protein. A similar result is observed in
western blot analysis. There were no significant changes for Bcl-2.
These results indicate a prominent participation of the extrinsic
apoptosis pathway, since Bcl-2 is an anti-apoptotic protein related
to the intrinsic apoptosis pathway (mitochondrial) (54,90).
GSH elevation and MDA decreases, promoted by the combined treatment
with GNPs (6.25 µg/ml) + carvedilol (3 µM), may have
corroborated with mitochondria protection, justifying
non-activation of the intrinsic apoptosis pathway (91–96).
The statistically significant elevation of FADD, mainly by combined
treatment with GNPs (6.25 µg/ml) + carvedilol (3 µM),
confirms activation of the extrinsic apoptosis pathway, since it
participates therein (97–99). Levels of APAF-1, a protein involved
in the intrinsic pathway (100),
did not present a significant differences.
The combined treatment with GNPs (6.25 µg/ml)
+ carvedilol (3 µM) was extremely effective on gene
expression of anti-apoptotic proteins such as Akt and mTOR,
matching or overcoming the action of cisplatin. Similar effect was
observed in western blot analysis. Inhibition of these proteins, as
well as MAPK/ERK, is critical for induction of apoptosis (12,51,52,101). Furthermore, other studies
indicate that treatments with targeting of the Pi3K/Akt/mTOR
pathway and the MAPK/ERK protein induce early and late apoptosis
(102–104), as also obtained in the present
study.
The decrease in EGFR levels is of great relevance
since it is related to activation of Akt/mTOR and MAPK/ERK
(105–108). However, it is possible that the
combined treatment may be acting on EGFR-independent pathways that
activate Akt and mTOR; and causing depletion (14,109–115). Levels of survivin were unchanged
for treatment. However, in the same figure, there was a
statistically significant reduction in MDR1 levels for both the
combined treatment and the gold nanoparticles alone. Studies have
shown that the use of nanoscale systems has excellent effects (this
includes inorganic nanoparticles, such as gold nanoparticles), on
the expression of genes related to multi-drug resistance, such as
MDR1 (116,117). The decrease in the expression of
this protein presents potential as a new strategy to combat one of
the main problems of cancer treatment, resistance to treatment
(118). Carvedilol, although
showing no effect on MDR1 expression, has been reported in the
literature as a potential MDR1 inhibitor (119,120). Studies in the literature have
demonstrated that treatments acting on drug resistance genes
promote both early and late apoptosis (121).
TEM did reveal GNPs internalization. However, we
noted that when given alone, GNPs accumulated near the plasmatic
membrane. Previous studies have reported that endosomal/lysosomal
vesicles can imprison GNPs preventing complete action in the
cellular interior, and are the greatest barrier that GNPs need to
overcome to reach the cellular nucleus (main target) (122–125). However, after administration of
carvedilol, concentration of GNPs was observed in the vicinity of
the cell nucleus, both intra- and peripherally. The data possibly
explain the fact that the results for GNPs administered alone show
smaller indices as compared to the combined treatment, which was
more effective. But even the isolated action of the gold
nanoparticle can reduce, although less than combined treatment, the
levels of survival and proliferation proteins such as Akt, EGFR,
MDR-1 and MAPK/ERK, demonstrated in the present study, and suggest
the way in which the GNPs inhibit proliferation. Concerning the
combined treatment, carvedilol may be acting through other
signaling pathways, or may be facilitating gold nanoparticle escape
from endosomal/lysosomal vesicles. Han et al (126) reported that carvedilol has a role
during receptor recycling in endosomal/lysosomal transiting of
vascular smooth muscle cell beta-adrenergic receptors. However,
there remains the need for further chemical interaction and
intracellular unfolding studies.
The present study demonstrated the ability of the
combined GNPs and carvedilol treatment to induce apoptosis in the
tumor cells by exclusively activating the extrinsic pathway.
Activation of this pathway is advantageous compared to the
intrinsic pathway, since it can be triggered independently of the
p53 gene, which in many tumors is inactive or absent (127). In addition, studies have
demonstrated that treatments with targeting intrinsic pathway
induction may also promote positive selection of tumor cells, while
evading the mitochondrial pathway (128). In addition to these results, the
combined treatment protected non-tumor cells, reducing oxidative
stress and consequently, apoptosis. The novel findings in these
studies highlight a promising alternative for future cancer
treatments.
Acknowledgments
The authors are grateful to the Brain Institute
(Federal University of Rio Grande do Norte), the BIOPOL (Department
of Biochemistry, Federal University of Rio Grande do Norte), the
Federal University of Ceará and the Leiden University Medical
Center for their contributions to the present study. Universal
476996/2013-9 CNPq 2013. CAPES 88881.119850/201601. This study was
also supported by the European Commission where R.F. De Araújo,
A.B. Chan and L.J. Cruz have received funding from a
MSCA-ITN-2015-ETN action grant (proposal no. 675743; project:
ISPIC).
Abbreviations:
|
GNPs
|
gold nanoparticles
|
|
Carv
|
carvedilol
|
|
EGFR
|
epidermal growth factor receptor
|
|
Erk
|
extracellular signal-regulated
kinases
|
|
FADD
|
fas-associated protein with death
domain
|
|
GAPDH
|
glyceraldehyde 3-phosphate
dehydrogenase
|
|
MDA
|
malondialdehyde
|
|
MAPK
|
mitogen activated protein kinases
|
|
MDR1
|
multidrug resistance genes-1
|
|
mTOR
|
mechanistic target of rapamycin
|
|
Pi3K
|
phosphatidylinositide 3-kinases
|
References
|
1
|
Hales S, Chiu A, Husain A, Braun M, Rydall
A, Gagliese L, Zimmermann C and Rodin G: The quality of dying and
death in cancer and its relationship to palliative care and place
of death. J Pain Symptom Manage. 48:839–851. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar
|
|
4
|
Özdemir F, Akalın G, Şen M, Önder NI,
Işcan A, Kutlu HM and Incesu Z: Towards novel anti-tumor strategies
for hepatic cancer: ε-viniferin in combination with vincristine
displays pharmacodynamic synergy at lower doses in HepG2 cells.
OMICS. 18:324–334. 2014. View Article : Google Scholar
|
|
5
|
Ling CQ: Problems in cancer treatment and
major research of integrative medicine. Zhong Xi Yi Jie He Xue Bao.
1:168–170. 2003.In Chinese. View Article : Google Scholar
|
|
6
|
Agrawal S: Late effects of cancer
treatment in breast cancer survivors. South Asian J Cancer.
3:112–115. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Oberstein PE and Olive KP: Pancreatic
cancer: Why is it so hard to treat? Therap Adv Gastroenterol.
6:321–337. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Michaelson MD, Cotter SE, Gargollo PC,
Zietman AL, Dahl DM and Smith MR: Management of complications of
prostate cancer treatment. CA Cancer J Clin. 58:196–213. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fernald K and Kurokawa M: Evading
apoptosis in cancer. Trends Cell Biol. 23:620–633. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Labi V and Erlacher M: How cell death
shapes cancer. Cell Death Dis. 6:e16752015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ran LK, Chen Y, Zhang ZZ, Tao NN, Ren JH,
Zhou L, Tang H, Chen X, Chen K, Li WY, et al: SIRT6 verexpression
Potentiates apoptosis evasion in hepatocellular carcinoma via
BCL2-associated X protein-dependent apoptotic pathway. Clin Cancer
Res. 22:3372–3382. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cai Y, Tan X, Liu J, Shen Y, Wu D, Ren M,
Huang P and Yu D: Inhibition of PI3K/Akt/mTOR signaling pathway
enhances the sensitivity of the SKOV3/DDP ovarian cancer cell line
to cisplatin in vitro. Chin J Cancer Res. 26:564–572.
2014.PubMed/NCBI
|
|
13
|
Lu Z and Xu S: ERK1/2 MAP kinases in cell
survival and apoptosis. IUBMB Life. 58:621–631. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang C, Cigliano A, Delogu S, Armbruster
J, Dombrowski F, Evert M, Chen X and Calvisi DF: Functional
crosstalk between AKT/mTOR and Ras/MAPK pathways in
hepatocarcinogenesis: Implications for the treatment of human liver
cancer. Cell Cycle. 12:1999–2010. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Katayama K, Noguchi K and Sugimoto Y:
Regulations of P-glycoprotein/ABCB1/MDR1 in human cancer cells. New
J Sci. 2014:e4769742014. View Article : Google Scholar
|
|
16
|
Yang X, Uziely B, Groshen S, Lukas J,
Israel V, Russell C, Dunnington G, Formenti S, Muggia F and Press
MF: MDR1 gene expression in primary and advanced breast cancer. Lab
Invest. 79:271–280. 1999.PubMed/NCBI
|
|
17
|
Chiara F, Gambalunga A, Sciacovelli M,
Nicolli A, Ronconi L, Fregona D, Bernardi P, Rasola A and Trevisan
A: Chemotherapeutic induction of mitochondrial oxidative stress
activates GSK-3α/β and Bax, leading to permeability transition pore
opening and tumor cell death. Cell Death Dis. 3:e4442012.
View Article : Google Scholar
|
|
18
|
Dasari S and Tchounwou PB: Cisplatin in
cancer therapy: Molecular mechanisms of action. Eur J Pharmacol.
740:364–378. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Marullo R, Werner E, Degtyareva N, Moore
B, Altavilla G, Ramalingam SS and Doetsch PW: Cisplatin induces a
mitochondrial-ROS response that contributes to cytotoxicity
depending on mitochondrial redox status and bioenergetic functions.
PLoS One. 8:e811622013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dreaden EC, Austin LA, Mackey MA and
El-Sayed MA: Size matters: Gold nanoparticles in targeted cancer
drug delivery. Ther Deliv. 3:457–478. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jain S, Hirst DG and O'Sullivan JM: Gold
nanoparticles as novel agents for cancer therapy. Br J Radiol.
85:101–113. 2012. View Article : Google Scholar :
|
|
22
|
Lee J, Chatterjee DK, Lee MH and Krishnan
S: Gold nanoparticles in breast cancer treatment: Promise and
potential pitfalls. Cancer Lett. 347:46–53. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Alkilany AM and Murphy CJ: Toxicity and
cellular uptake of gold nanoparticles: What we have learned so far?
J Nanopart Res. 12:2313–2333. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Alvarenga ÉC, Caires A, Ladeira LO, Gamero
EJP, Andrade LM and Paz MTL: Potenciais alvos terapêuticos contra o
câncer. Cienc Cult. 66:43–48. 2014. View Article : Google Scholar
|
|
25
|
Naha PC, Chhour P and Cormode DP:
Systematic in vitro toxicological screening of gold nanoparticles
designed for nanomedicine applications. Toxicol In Vitro.
29:1445–1453. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Butterworth KT, Coulter JA, Jain S, Forker
J, McMahon SJ, Schettino G, Prise KM, Currell FJ and Hirst DG:
Evaluation of cytotoxicity and radiation enhancement using 1.9 nm
gold particles: Potential application for cancer therapy.
Nanotechnology. 21:2951012010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Coulter JA, Jain S, Butterworth KT,
Taggart LE, Dickson GR, McMahon SJ, Hyland WB, Muir MF, Trainor C,
Hounsell AR, et al: Cell type-dependent uptake, localization, and
cytotoxicity of 1.9 nm gold nanoparticles. Int J Nanomedicine.
7:2673–2685. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Murawala P, Tirmale A, Shiras A and Prasad
BLV: In situ synthesized BSA capped gold nanoparticles: Effective
carrier of anticancer drug methotrexate to MCF-7 breast cancer
cells. Mater Sci Eng C. 34:158–167. 2014. View Article : Google Scholar
|
|
29
|
Patra HK, Banerjee S, Chaudhuri U, Lahiri
P and Dasgupta AK: Cell selective response to gold nanoparticles.
Nanomedicine (Lond). 3:111–119. 2007. View Article : Google Scholar
|
|
30
|
Budni P, Pedrosa RC, Dalmarco EM, Dalmarco
JB, Frode TS and Wilhelm Filho D: Carvedilol enhances the
antioxidant effect of vitamins E and C in chronic Chagas heart
disease. Arq Bras Cardiol. 101:304–310. 2013.PubMed/NCBI
|
|
31
|
Arozal W, Watanabe K, Veeraveedu PT, Ma M,
Thandavarayan RA, Sukumaran V, Suzuki K, Kodama M and Aizawa Y:
Protective effect of carvedilol on daunorubicin-induced
cardiotoxicity and nephrotoxicity in rats. Toxicology. 274:18–26.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Arumanayagam M, Chan S, Tong S and
Sanderson JE: Antioxidant properties of carvedilol and metoprolol
in heart failure: A double-blind randomized controlled trial. J
Cardiovasc Pharmacol. 37:48–54. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Dandona P, Ghanim H and Brooks DP:
Antioxidant activity of carvedilol in cardiovascular disease. J
Hypertens. 25:731–741. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li YC, Ge LS, Yang PL, Tang JF, Lin JF,
Chen P and Guan XQ: Carvedilol treatment ameliorates acute
coxsackievirus B3-induced myocarditis associated with oxidative
stress reduction. Eur J Pharmacol. 640:112–116. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Pasquier E, Street J, Pouchy C, Carre M,
Gifford AJ, Murray J, Norris MD, Trahair T, Andre N and Kavallaris
M: β-blockers increase response to chemotherapy via direct
antitumour and anti-angiogenic mechanisms in neuroblastoma. Br J
Cancer. 108:2485–2494. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Erguven M, Yazihan N, Aktas E, Sabanci A,
Li CJ, Oktem G and Bilir A: Carvedilol in glioma treatment alone
and with imatinib in vitro. Int J Oncol. 36:857–866. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Dezong G, Zhongbing M, Qinye F and Zhigang
Y: Carvedilol suppresses migration and invasion of malignant breast
cells by inactivating Src involving cAMP/PKA and PKCδ signaling
pathway. J Cancer Res Ther. 10:998–1003. 2014. View Article : Google Scholar
|
|
38
|
Chang A, Yeung S, Thakkar A, Huang KM, Liu
MM, Kanassatega RS, Parsa C, Orlando R, Jackson EK, Andresen BT, et
al: Prevention of skin carcinogenesis by the β-blocker carvedilol.
Cancer Prev Res (Phila). 8:27–36. 2015. View Article : Google Scholar
|
|
39
|
Hsieh YD, Chi CC, Chou CT, Cheng JS, Kuo
CC, Liang WZ, Lin KL, Tseng LL and Jan CR: Investigation of
carvedilolevoked Ca2+ movement and death in human oral
cancer cells. J Recept Signal Transduct Res. 31:220–228. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Cheng JS, Huang CC, Chou CT and Jan CR:
Mechanisms of carvedilol-induced [Ca2+]i
rises and death in human hepatoma cells. Naunyn Schmiedebergs Arch
Pharmacol. 376:185–194. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Cohen DJ and Hochster HS: Rationale for
combining biotherapy in the treatment of advanced colon cancer.
Gastrointest Cancer Res. 2:145–151. 2008.
|
|
42
|
Patutina OA, Mironova NL, Vlassov VV and
Zenkova MA: New approaches for cancer treatment: Antitumor drugs
based on gene-targeted nucleic acids. Acta Naturae. 1:44–60.
2009.PubMed/NCBI
|
|
43
|
Siddiqui M and Rajkumar SV: The high cost
of cancer drugs and what we can do about it. Mayo Clin Proc.
87:935–943. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Tannock IF: Combined modality treatment
with radiotherapy and chemotherapy. Radiother Oncol. 16:83–101.
1989. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Brito AF, Ribeiro M, Abrantes AM, Pires
AS, Teixo RJ, Tralhão JG and Botelho MF: Quercetin in cancer
treatment, alone or in combination with conventional therapeutics?
Curr Med Chem. 22:3025–3039. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Mierzwa ML, Nyati MK, Morgan MA and
Lawrence TS: Recent advances in combined modality therapy.
Oncologist. 15:372–381. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Li J, Wang Y, Zhu Y and Oupický D: Recent
advances in delivery of drug-nucleic acid combinations for cancer
treatment. J Control Release. 172:589–600. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Collery P, Mohsen A, Kermagoret A,
D'Angelo J, Morgant G, Desmaele D, Tomas A, Collery T, Wei M and
Badawi A: Combination of three metals for the treatment of cancer:
Gallium, rhenium and platinum. 1. Determination of the optimal
schedule of treatment. Anticancer Res. 32:2769–2781.
2012.PubMed/NCBI
|
|
49
|
Law MR, Wald NJ, Morris JK and Jordan RE:
Value of low dose combination treatment with blood pressure
lowering drugs: Analysis of 354 randomised trials. BMJ.
326:14272003. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Morton CO, Chau M and Stack C: In vitro
combination therapy using low dose clotrimazole and photodynamic
therapy leads to enhanced killing of the dermatophyte Trichophyton
rubrum. BMC Microbiol. 14:2612014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Chang L, Graham PH, Ni J, Hao J, Bucci J,
Cozzi PJ and Li Y: Targeting PI3K/Akt/mTOR signaling pathway in the
treatment of prostate cancer radioresistance. Crit Rev Oncol
Hematol. 96:507–517. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Chappell WH, Steelman LS, Long JM, Kempf
RC, Abrams SL, Franklin RA, Bäsecke J, Stivala F, Donia M, Fagone
P, et al: Ras/Raf/MEK/ERK and PI3K/PTEN/Akt/mTOR inhibitors:
Rationale and importance to inhibiting these pathways in human
health. Oncotarget. 2:135–164. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
de Araújo RF, de Araújo AA, Pessoa JB,
Freire Neto FP, da Silva GR, Leitão Oliveira AL, de Carvalho TG,
Silva HF, Eugênio M, Sant'Anna C, et al: Anti-inflammatory,
analgesic and anti-tumor properties of gold nanoparticles.
Pharmacol Rep. 69:119–129. 2017. View Article : Google Scholar
|
|
54
|
de Araújo Júnior RF, Leitão Oliveira ALC,
de Melo Silveira RF, de Oliveira Rocha HA, de França Cavalcanti P
and de Araújo AA: Telmisartan induces apoptosis and regulates Bcl-2
in human renal cancer cells. Exp Biol Med (Maywood). 240:34–44.
2015. View Article : Google Scholar
|
|
55
|
Rahman I, Kode A and Biswas SK: Assay for
quantitative determination of glutathione and glutathione disulfide
levels using enzymatic recycling method. Nat Protoc. 1:3159–3165.
2006. View Article : Google Scholar
|
|
56
|
da Costa CM, dos Santos RC and Lima ES: A
simple automated procedure for thiol measurement in human serum
samples. J Bras Patol Med Lab. 42:345–350. 2006. View Article : Google Scholar
|
|
57
|
Esterbauer H and Cheeseman KH:
Determination of aldehydic lipid peroxidation products:
Malonaldehyde and 4-hydroxynonenal. Methods Enzymol. 186:407–421.
1990. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Al-Sheddi ES, Al-Oqail MM, Saquib Q,
Siddiqui MA, Musarrat J, Al-Khedhairy AA and Farshori NN: Novel all
trans-retinoic Acid derivatives: Cytotoxicity, inhibition of cell
cycle progression and induction of apoptosis in human cancer cell
lines. Molecules. 20:8181–8197. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Kimura H, Sakai K, Arao T, Shimoyama T,
Tamura T and Nishio K: Antibody-dependent cellular cytotoxicity of
cetuximab against tumor cells with wild-type or mutant epidermal
growth factor receptor. Cancer Sci. 98:1275–1280. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Świątek Ł, Rajtar B, Pawlak K, Ludwiczuk
A, Głowniak K and Polz-Dacewicz M: In vitro evaluation of
cytotoxicity of n-hexane extract from Alnus sieboldiana male
flowers on VERO and HEK293 cell lines. JPCCR. 7:110–107. 2014.
|
|
61
|
Lapique N and Benenson Y: Digital
switching in a biosensor circuit via programmable timing of gene
availability. Nat Chem Biol. 10:1020–1027. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Selvaraj V, Bodapati S, Murray E, Rice KM,
Winston N, Shokuhfar T, Zhao Y and Blough E: Cytotoxicity and
genotoxicity caused by yttrium oxide nanoparticles in HEK293 cells.
Int J Nanomed. 9:1379–1391. 2014. View Article : Google Scholar
|
|
63
|
Jia J, Zhu F, Ma X, Cao Z, Cao ZW, Li Y,
Li YX and Chen YZ: Mechanisms of drug combinations: Interaction and
network perspectives. Nat Rev Drug Discov. 8:111–128. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Richardson PG, Siegel DS, Vij R,
Hofmeister CC, Baz R, Jagannath S, Chen C, Lonial S, Jakubowiak A,
Bahlis N, et al: Pomalidomide alone or in combination with low-dose
dexamethasone in relapsed and refractory multiple myeloma: A
randomized phase 2 study. Blood. 123:1826–1832. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Nijhof IS, Franssen LE, Levin M-D, Bos
GMJ, Broijl A, Klein SK, Koene HR, Bloem AC, Beeker A, Faber LM, et
al: Phase 1/2 study of lenalidomide combined with low-dose
cyclophosphamide and prednisone in lenalidomide-refractory multiple
myeloma. Blood. 128:2297–2306. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Gatoo MA, Naseem S, Arfat MY, Dar AM,
Qasim K and Zubair S: Physicochemical properties of nanomaterials:
Implication in associated toxic manifestations. BioMed Res Int.
2014:4984202014. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Coelho M, Moz M, Correia G, Teixeira A,
Medeiros R and Ribeiro L: Antiproliferative effects of β-blockers
on human colorectal cancer cells. Oncol Rep. 33:2513–2520. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Baharara J, Ramezani T, Divsalar A,
Mousavi M and Seyedarabi A: Induction of apoptosis by green
synthesized gold nanoparticles Through activation of caspase-3 and
9 in human cervical cancer cells. Avicenna J Med Biotechnol.
8:75–83. 2016.PubMed/NCBI
|
|
69
|
Connor EE, Mwamuka J, Gole A, Murphy CJ
and Wyatt MD: Gold nanoparticles are taken up by human cells but do
not cause acute cytotoxicity. Small. 1:325–327. 2005. View Article : Google Scholar
|
|
70
|
Zhao Y, Xu Y, Zhang J and Ji T:
Cardioprotective effect of carvedilol: Inhibition of apoptosis in
H9c2 cardiomyocytes via the TLR4/NF-κB pathway following
ischemia/reperfusion injury. Exp Ther Med. 8:1092–1096. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Carvalho Rodrigues MA, Gobe G, Santos NA
and Santos AC: Carvedilol protects against apoptotic cell death
induced by cisplatin in renal tubular epithelial cells. J Toxicol
Environ Health A. 75:981–990. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Lu SC: Glutathione synthesis. Biochim
Biophys Acta. 1830:3143–3153. 2013. View Article : Google Scholar :
|
|
73
|
Townsend DM, Tew KD and Tapiero H: The
importance of glutathione in human disease. Biomed Pharmacother.
57:145–155. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Kannan K and Jain SK: Oxidative stress and
apoptosis. Pathophysiology. 7:153–163. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Takahashi A, Masuda A, Sun M, Centonze VE
and Herman B: Oxidative stress-induced apoptosis is associated with
alterations in mitochondrial caspase activity and Bcl-2-dependent
alterations in mitochondrial pH (pHm). Brain Res Bull. 62:497–504.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Chang WK, Yang KD, Chuang H, Jan JT and
Shaio MF: Glutamine protects activated human T cells from apoptosis
by up-regulating glutathione and Bcl-2 levels. Clin Immunol.
104:151–160. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Estrela JM, Ortega A and Obrador E:
Glutathione in cancer biology and therapy. Crit Rev Clin Lab Sci.
43:143–181. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Zubairi MB, Ahmed JH and Al-Haroon SS:
Effect of adrenergic blockers, carvedilol, prazosin, metoprolol and
combination of prazosin and metoprolol on paracetamol-induced
hepatotoxicity in rabbits. Indian J Pharmacol. 46:644–648. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Sgobbo P, Pacelli C, Grattagliano I,
Villani G and Cocco T: Carvedilol inhibits mitochondrial complex I
and induces resistance to H2O2-mediated
oxidative insult in H9C2 myocardial cells. Biochim Biophys Acta.
1767:222–232. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Barathmanikanth S, Kalishwaralal K, Sriram
M, Pandian SR, Youn HS, Eom S and Gurunathan S: Anti-oxidant effect
of gold nanoparticles restrains hyperglycemic conditions in
diabetic mice. J Nanobiotech. 8:162010. View Article : Google Scholar
|
|
81
|
Yakimovich NO, Ezhevskii AA, Guseinov DV,
Smirnova LA, Gracheva TA and Klychkov KS: Antioxidant properties of
gold nanoparticles studied by ESR spectroscopy. Russ Chem Bull.
57:520–523. 2008. View Article : Google Scholar
|
|
82
|
Nielsen F, Mikkelsen BB, Nielsen JB,
Andersen HR and Grandjean P: Plasma malondialdehyde as biomarker
for oxidative stress: Reference interval and effects of life-style
factors. Clin Chem. 43:1209–1214. 1997.PubMed/NCBI
|
|
83
|
Gaweł S, Wardas M, Niedworok E and Wardas
P: Malondialdehyde (MDA) as a lipid peroxidation marker. Wiad Lek.
57:453–455. 2004.In Polish.
|
|
84
|
Ho E, Karimi Galougahi K, Liu CC, Bhindi R
and Figtree GA: Biological markers of oxidative stress:
Applications to cardiovascular research and practice. Redox Biol.
1:483–491. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Sangeetha P, Das UN, Koratkar R and
Suryaprabha P: Increase in free radical generation and lipid
peroxidation following chemotherapy in patients with cancer. Free
Radic Biol Med. 8:15–19. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Esfahani A, Ghoreishi Z, Nikanfar A,
Sanaat Z and Ghorbanihaghjo A: Influence of chemotherapy on the
lipid peroxidation and antioxidant status in patients with acute
myeloid leukemia. Acta Med Iran. 50:454–458. 2012.PubMed/NCBI
|
|
87
|
Cabello CM, Bair WB III and Wondrak GT:
Experimental therapeutics: Targeting the redox Achilles heel of
cancer. Curr Opin Investig Drugs. 8:1022–1037. 2007.PubMed/NCBI
|
|
88
|
Liou GY and Storz P: Reactive oxygen
species in cancer. Free Radic Res. 44:479–496. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
García M and Vecino E: Vías de
señalización intracelular que conducen a la apoptosis de las
células de la retina. Arch Soc Esp Oftalmol. 78:351–364. 2003.
View Article : Google Scholar
|
|
90
|
Kang MH and Reynolds CP: Bcl-2 inhibitors:
Targeting mitochondrial apoptotic pathways in cancer therapy. Clin
Cancer Res. 15:1126–1132. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Ahmad S, White CW, Chang LY, Schneider BK
and Allen CB: Glutamine protects mitochondrial structure and
function in oxygen toxicity. Am J Physiol Lung Cell Mol Physiol.
280:L779–L791. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Drake J, Sultana R, Aksenova M, Calabrese
V and Butterfield DA: Elevation of mitochondrial glutathione by
gamma-glutamylcysteine ethyl ester protects mitochondria against
peroxynitrite-induced oxidative stress. J Neurosci Res. 74:917–927.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Marí M, Morales A, Colell A, García-Ruiz C
and Fernández-Checa JC: Mitochondrial glutathione, a key survival
antioxidant. Antioxid Redox Signal. 11:2685–2700. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Cheng J, Wang F, Yu DF, Wu PF and Chen JG:
The cytotoxic mechanism of malondialdehyde and protective effect of
carnosine via protein cross-linking/mitochondrial
dysfunction/reactive oxygen species/MAPK pathway in neurons. Eur J
Pharmacol. 650:184–194. 2011. View Article : Google Scholar
|
|
95
|
Ayala A, Muñoz MF and Argüelles S: Lipid
peroxidation: Production, metabolism, and signaling mechanisms of
malondialdehyde and 4-hydroxy-2-nonenal. Oxid Med Cell Longev.
2014:3604382014. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Qin J, Kang Y, Xu Z, Zang C, Fang B and
Liu X: Dioscin prevents the mitochondrial apoptosis and attenuates
oxidative stress in cardiac H9c2 cells. Drug Res (Stuttg).
64:47–52. 2014.
|
|
97
|
Bang S, Jeong EJ, Kim IK, Jung YK and Kim
KS: Fas- and tumor necrosis factor-mediated apoptosis uses the same
binding surface of FADD to trigger signal transduction. A typical
model for convergent signal transduction. J Biol Chem.
275:36217–36222. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Osborn SL, Sohn SJ and Winoto A:
Constitutive phosphorylation mutation in Fas-associated death
domain (FADD) results in early cell cycle defects. J Biol Chem.
282:22786–22792. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Xerri L, Devilard E, Bouabdallah R, Stoppa
AM, Hassoun J and Birg F: FADD expression and caspase activation in
B-cell lymphomas resistant to Fas-mediated apoptosis. Br J
Haematol. 106:652–661. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Campioni M, Santini D, Tonini G, Murace R,
Dragonetti E, Spugnini EP and Baldi A: Role of Apaf-1, a key
regulator of apoptosis, in melanoma progression and
chemoresistance. Exp Dermatol. 14:811–818. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Brazil DP, Yang ZZ and Hemmings BA:
Advances in protein kinase B signalling: AKTion on multiple fronts.
Trends Biochem Sci. 29:233–242. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Daniele S, Costa B, Zappelli E, Da Pozzo
E, Sestito S, Nesi G, Campiglia P, Marinelli L, Novellino E,
Rapposelli S, et al: Combined inhibition of AKT/mTOR and MDM2
enhances glioblastoma multiforme cell apoptosis and differentiation
of cancer stem cells. Sci Rep. 5:99562015. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Li C, Xin P, Xiao H, Zheng Y, Huang Y and
Zhu X: The dual PI3K/mTOR inhibitor NVP-BEZ235 inhibits
proliferation and induces apoptosis of burkitt lymphoma cells.
Cancer Cell Int. 15:652015. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Liu Z, Ruan HJ, Gu PQ, Ding WY, Luo XH,
Huang R, Zhao W and Gao LJ: The roles of p38 MAPK and ERK1/2 in
coplanar polychlorinated biphenyls-induced apoptosis of human
extravillous cytotrophoblast-derived transformed cells. Cell
Physiol Biochem. 36:2418–2432. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Freudlsperger C, Burnett JR, Friedman JA,
Kannabiran VR, Chen Z and Van Waes C: EGFR-PI3K-AKT-mTOR signaling
in head and neck squamous cell carcinomas: Attractive targets for
molecular-oriented therapy. Expert Opin Ther Targets. 15:63–74.
2011. View Article : Google Scholar
|
|
106
|
Gan Y, Shi C, Inge L, Hibner M, Balducci J
and Huang Y: Differential roles of ERK and Akt pathways in
regulation of EGFR-mediated signaling and motility in prostate
cancer cells. Oncogene. 29:4947–4958. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Seshacharyulu P, Ponnusamy MP, Haridas D,
Jain M, Ganti AK and Batra SK: Targeting the EGFR signaling pathway
in cancer therapy. Expert Opin Ther Targets. 16:15–31. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Kidger AM and Keyse SM: The regulation of
oncogenic Ras/ERK signalling by dual-specificity mitogen activated
protein kinase phosphatases (MKPs). Semin Cell Dev Biol.
50:125–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Gao Y, Moten A and Lin HK: Akt: A new
activation mechanism. Cell Res. 24:785–786. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Liu P, Begley M, Michowski W, Inuzuka H,
Ginzberg M, Gao D, Tsou P, Gan W, Papa A, Kim BM, et al:
Cell-cycle-regulated activation of Akt kinase by phosphorylation at
its carboxyl terminus. Nature. 508:541–545. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Aeder SE, Martin PM, Soh JW and Hussaini
IM: PKC-eta mediates glioblastoma cell proliferation through the
Akt and mTOR signaling pathways. Oncogene. 23:9062–9069. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Fan QW, Cheng C, Knight ZA, Haas-Kogan D,
Stokoe D, James CD, McCormick F, Shokat KM and Weiss WA: EGFR
signals to mTOR through PKC and independently of Akt in glioma. Sci
Signal. 2:ra42009. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Mendoza MC, Er EE and Blenis J: The
Ras-ERK and PI3K-mTOR pathways: Cross-talk and compensation. Trends
Biochem Sci. 36:320–328. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Denduluri SK, Idowu O, Wang Z, Liao Z, Yan
Z, Mohammed MK, Ye J, Wei Q, Wang J, Zhao L, et al: Insulin-like
growth factor (IGF) signaling in tumorigenesis and the development
of cancer drug resistance. Genes Dis. 2:13–25. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Farabaugh SM, Boone DN and Lee AV: Role of
IGF1R in breast cancer subtypes, stemness and lineage
differentiation. Front Endocrinol (Lausanne). 6:592015.
|
|
116
|
Kapse-Mistry S, Govender T, Srivastava R
and Yergeri M: Nanodrug delivery in reversing multidrug resistance
in cancer cells. Front Pharmacol. 5:1592014.PubMed/NCBI
|
|
117
|
Salomon JJ and Ehrhardt C: Nanoparticles
attenuate P-glycoprotein/MDR1 function in A549 human alveolar
epithelial cells. Eur J Pharm Biopharm. 77:392–397. 2011.
View Article : Google Scholar
|
|
118
|
Callaghan R, Luk F and Bebawy M:
Inhibition of the multidrug resistance P-glycoprotein: Time for a
change of strategy? Drug Metab Dispos. 42:623–631. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Kakumoto M, Sakaeda T, Takara K, Nakamura
T, Kita T, Yagami T, Kobayashi H, Okamura N and Okumura K: Effects
of carvedilol on MDR1-mediated multidrug resistance: Comparison
with verapamil. Cancer Sci. 94:81–86. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Wessler JD, Grip LT, Mendell J and
Giugliano RP: The P-glycoprotein transport system and
cardiovascular drugs. J Am Coll Cardiol. 61:2495–2502. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Mitsiades CS, Treon SP, Mitsiades N, Shima
Y, Richardson P, Schlossman R, Hideshima T and Anderson KC:
TRAIL/Apo2L ligand selectively induces apoptosis and overcomes drug
resis-tance in multiple myeloma: Therapeutic applications. Blood.
98:795–804. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Huang X, Kang B, Qian W, Mackey MA, Chen
PC, Oyelere AK, El-Sayed IH and El-Sayed MA: Comparative study of
photothermolysis of cancer cells with nuclear-targeted or
cytoplasm-targeted gold nanospheres: Continuous wave or pulsed
lasers. J Biomed Opt. 15:0580022010. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Yang CJ and Chithrani DB: Nuclear
targeting of gold nanoparticles for improved therapeutics. Curr Top
Med Chem. 16:271–280. 2016. View Article : Google Scholar
|
|
124
|
Kodiha M, Wang YM, Hutter E, Maysinger D
and Stochaj U: Off to the organelles - killing cancer cells with
targeted gold nanoparticles. Theranostics. 5:357–370. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Yanes RE, Tarn D, Hwang AA, Ferris DP,
Sherman SP, Thomas CR, Lu J, Pyle AD, Zink JI and Tamanoi F:
Involvement of lysosomal exocytosis in the excretion of mesoporous
silica nanoparticles and enhancement of the drug delivery effect by
exocytosis inhibition. Small. 9:697–704. 2013. View Article : Google Scholar
|
|
126
|
Han SO, Xiao K, Kim J, Wu JH, Wisler JW,
Nakamura N, Freedman NJ and Shenoy SK: MARCH2 promotes endocytosis
and lysosomal sorting of carvedilol-bound β2-adrenergic
receptors. J Cell Biol. 199:817–830. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
El-Deiry WS: Insights into cancer
therapeutic design based on p53 and TRAIL receptor signaling. Cell
Death Differ. 8:1066–1075. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Sayers TJ: Targeting the extrinsic
apoptosis signaling pathway for cancer therapy. Cancer Immunol
Immunother. 60:1173–1180. 2011. View Article : Google Scholar : PubMed/NCBI
|