Introduction
Pancreatic cancer is an aggressive and lethal
malignancy with an overall 5-year survival rate <8%, and its
death rate continues to increase by 0.3% per annum (1). Although surgery remains a potentially
curative treatment, chemotherapy is an important and indispensable
therapy in maximizing the life span for patients with both
resectable and unresectable tumours. Currently, gemcitabine-based
therapies, either alone or in combination with agents as such
nab-paclitaxel (2), are favored
approaches for treatment of patients with locally advanced or
metastatic pancreatic cancer. Although the regimen, folic acid,
5-fluorouracil, irinotecan, oxalaplatin (FOLFIRINOX), has increased
survival compared to gemcitabine, it often cannot be tolerated due
to its high toxicity and the poor performance status of patients
(3). This necessitates a continued
search for more effective and less toxic combination therapies for
the treatment of pancreatic cancer.
Immunotherapy to activate antitumour immunity has
delivered promising results in various tumours (4). However, immunotherapy has little
effect in pancreatic ductal adenocarcinoma (PDA) because of the
potently immunosuppressive microenvironment in which immune cells,
PDA cells and stroma interact to facilitate cancer progression
(5). A combination approach
involving chemotherapy, immunotherapy, targeted therapy against
stromal elements, and other modalities, may be required in order to
stimulate antitumour immunity, increase treatment efficacy and
improve survival.
The p21 activated kinase (PAK) family of
serine/threonine kinases, the downstream effector proteins of Rho
GTPases, are categorized into two groups. Both group I (PAK1-3) and
group II (PAK4-6) PAKs are involved in multiple cellular signaling
pathways that regulate cell survival, proliferation, and migration
(6). Overexpression and
hyper-activation of PAKs contribute significantly to the initiation
and progression of human cancers, and PAK1 and PAK4 are the
best-studied members in this area. PAK1 expression is upregulated
in both human pancreatic cancer cells and tissues (7). Inhibition of PAK1 in pancreatic
cancer cell lines by shRNA knock-down or a PAK1 specific inhibitor
(Frax-597) decreased cancer cell growth, synergistically with
gemcitabine (7). PAK4 upregulation
enhanced pancreatic cancer cell proliferation and survival through
AKT- and ERK-dependent activation of the NF-κB pathway (8), and stimulated migration and
invasiveness. In addition, PAK4 has been linked to the maintenance
of a stem cell-like phenotype (9)
and is a predictive marker of gemcitabine sensitivity in pancreatic
cancer cells (10).
Kras mutations have been observed in >95% of PDA
(11). The failure to identify a
drug-binding domain in the Kras protein has made it difficult to be
targeted therapeutically. Therefore, the ability to target other
downstream molecules in Kras signaling pathways has become
increasingly important. PAK1 can be activated via a Ras-dependent
pathway and at least in part, mediates the effect of activated Kras
in pancreatic cancer (6). In this
study, a PAK inhibitor [PF-3758309, IC50, 13.7 nM for
PAK1 and 1.3 nM for PAK4 (12)]
was used to evaluate the combined effect of PAK inhibition with
gemcitabine on pancreatic cancer growth and on the tumour immune
response.
Materials and methods
Cells and reagents
Human PANC-1 and MiaPaCa-2 pancreatic cancer cells
(purchased from Sigma-Aldrich, Melbourne, Australia) and a murine
PAN02 pancreatic cancer cell [obtained from National Cancer
Institute (Frederick, MD, USA)] were cultured in Dulbecco's
modified Eagle's medium (DMEM) supplemented with 5% FBS (fetal
bovine serum; Hyclone Laboratories Inc., Melbourne, Australia) in a
37°C incubator with a humidified atmosphere of 5% CO2.
PF-3758309 was purchased from Active Biochemical Co. Maplewood, NJ,
USA and gemcitabine from Sigma-Aldrich. PAK1 knock-down (KD) PANC-1
and MiaPaCa-2 cells were generated by transfection with
SureSilencing shRNA plasmids for human PAK1 (SABioscience,
Doncaster, Australia), or with a scrambled sequence as a negative
control, using Lipofectamine 2000 (Invitrogen, Mulgrave,
Australia), according to the manufacturer's instructions. Stable
clones were selected with geneticin (G418; 1 mg/ml). PAK protein
expression was detected by western blotting.
Cell proliferation
Cell proliferation was measured using a
3H-thymidine incorporation assay. Cells
(5×103 cells/well) were seeded in a 96-well plate and
incubated in DMEM containing 5% FBS, with 1 μCi/well
[methyl-3H]-thymidine (Perkin-Elmer, Boston, MA, USA) in
the presence of PF-3758309 at the concentrations indicated in the
Results section. After 24 h cells were harvested using a NUNC cell
harvester (Nunc, Roskilde, Denmark). The amount of
3H-thymidine incorporated through DNA synthesis was
detected with a β-counter (Packard, Meriden, CT, USA). For
combinational treatment, cells were pre-treated with PF-3758309 for
24 h, and then PF-3758309 was removed. The cells were incubated
with increasing concentrations of gemcitabine and 1 μCi/well
[methyl-3H]-thymidine for a further 24 h and harvested
and detected as above.
Migration/invasion assay
Cell migration/invasion was measured using a
Transwell Boyden Chamber assay as described previously (13). Membranes (8-μm pore size;
Becton-Dickinson, NJ, USA) were coated with 3 μg of human
fibronectin on the lower surfaces and placed into a 24-well plate
containing 600 μl/well of serum-free DMEM with 0.1% bovine
serum albumin (BSA). Cells were added to the upper chambers at
5×104/100 μl with or without PF-3758309 at the
concentrations indicated. Cells were then incubated for 24 h. Cells
that had not penetrated were removed from the upper surface by
wiping with a cotton swab. The membranes were then fixed and
stained with Quick-Dip (Fronine, Sydney, Australia). The cells that
had migrated to the lower surface of the membranes were counted in
24–48 fields, depending on the cell line, at ×40 magnification
using a Nikon Coolscope (Coherent Scientific, Adelaide, Australia).
The combined effects of gemcitabine and PF-3758309 were evaluated
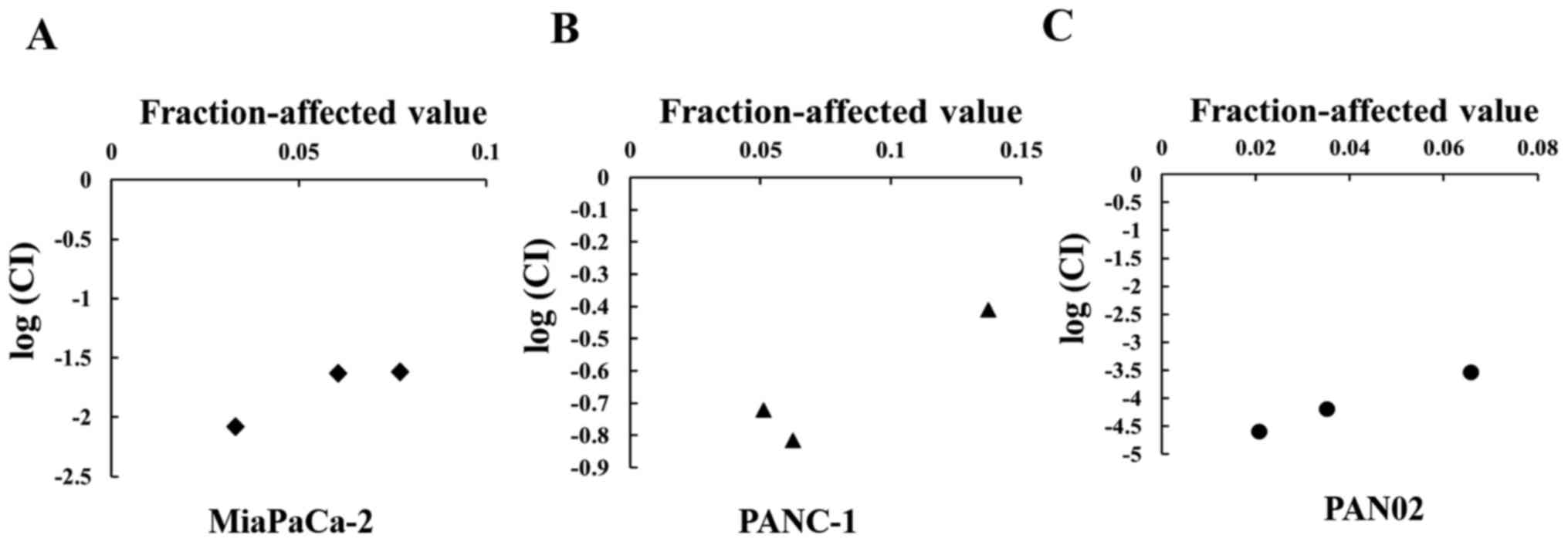
using the Chou-Talalay method as previously described (14). The combination index (CI) was
calculated with the CalcuSyn program (T.C. Chou and M.P Hayball;
Biosoft, Cambridge, UK) using the mutually non-exclusive (α=1)
isobologram equation. The CI value is interpreted as follows:
<1.0, synergistic; 1.0, additive and >1.0, antagonistic.
Anchorage-independent assay
Anchorage-independent growth of pancreatic cancer
cell lines was determined by soft agar assay as described
previously (15). Briefly, cells
(2,000/well) were seeded in top agar layer containing 0.4%
bacto-agar/DMEM (0.5 ml) over a bottom agar layer of 0.6%
bacto-agar/DMEM (1 ml) in 12-well plates. Both layers of soft agar
were allowed to set at room temperature for 20–30 min before 0.5 ml
of DMEM containing treatment reagents was added to the wells. For
wild-type MiaPaCa-2 and PANC-1 cells were divided into four groups:
control, PF-3758309, gemcitabine, and combination of PF-3758309
with gemcitabine. For PAK1 knock-down MiaPaCa-2 and PANC-1, each
cell line was divided into two groups: control and gemcitabine. The
concentrations of PF3758309 and gemcitabine were chosen close to
the IC50 values from the proliferation assay. Cell
colonies were grown in a humidified 5% CO2 incubator at
37°C for 4 weeks. Culture medium containing treatment reagents was
changed twice weekly. At the endpoint, colonies were fixed and
stained with 4% formalin and 0.005% crystal violet in
phosphate-buffered saline (PBS). Colonies were counted manually and
images were captured using a Leica microscope at ×4
magnification.
Animal experiments
All mouse experiments were approved by the Austin
Health Research Ethics Committee (A2013/04898). An orthotopic
pancreatic cancer tail model was used to assess tumour growth in
the pancreatic tails of C57BL/6 mice as previously described
(16). PAN02 (a murine pancreatic
cancer cell line) cells were injected into the pancreatic tails of
24 PAK1 wild-type (WT) mice, which were allocated randomly into
four treatment groups (6 mice/group): control, intraperitoneal
(i.p.) injection of saline every other day; gemcitabine (40 mg/kg),
1.p. twice weekly; PF-3758309 (25 mg/kg), i.p. every other day;
combination of gemcitabine with PF-3758309, following the
individual treatment as described above. For assessment of tumour
growth in PAK1 knockout (KO) C57BL/6 mice, 9 mice were injected
with PAN02 cells in the pancreatic tail and were allocated randomly
into two groups: control (n=4) and gemcitabine (n=5), and treated
as described above. All treatments commenced one week after
surgical induction and continued for 4 weeks. All mice were
monitored based on strict health monitoring criteria and euthanized
when a poor health score was reached. At the endpoint, tumour
dimensions were measured using micro-calipers. The tumour volume
was calculated by using the formula: V = L × W × H × (π/6), where L
was the length; W the width; and H the height.
Flow cytometric analysis
Spleens were harvested from tumour-bearing mice as
described above, and homogenised in 0.5% BSA in PBS. The splenic
cells were collected by centrifugation after red cells had been
removed using red cell lysis buffer (150 mM NH4C1, 10 mM
NaHCO3, 0.1 mM EDTA). After further washing in 0.5% BSA
in PBS, the remaining cells were incubated for 1 h on ice with
FITC- or APC-labelled antibodies against B220, CD3, CD8 (1:100) (BD
Biosciences, North Ryde, Australia) or CD4 (1:10) (Miltenyi Biotec,
Macquarie Park, Australia), or with respective isotype controls.
Cells were washed 3 times in 0.5% BSA in PBS before analysis by
FACSCanto II (BD Biosciences). Data were analysed using Weasel
software (Cytometry Laboratory, Walter and Eliza Hall Institute,
Parkville, Australia).
Immunohistochemistry stain
Tumour tissues, collected from the orthotopic tail
model as described above, were embedded in paraffin and stained
with Ki67 antibody (Thermo Fisher Scientific, Melbourne, Australia)
for proliferation, active caspase 3 antibody (Cell Signaling
Technology, Arundel, Australia) for apoptosis, or CD3 antibody
(Abcam, Melbourne, Australia) for total T cell surface marker.
Samples were imaged using a Leica microscope for at least 4 fields
at ×40 magnification. For the CD3+ stain, cell numbers
were counted per ×40 field. For the Ki67 and caspase 3 stains, the
ratio of positive cells to total number of cells in each field was
calculated.
Western blotting
Cells were treated with PF-3758309 at the
IC50 concentration for 24 h, lysed in SDS sample buffer,
and denatured by heating at 95°C for 5 min. Proteins in the cell
lysates were detected with antibodies against phospho-PAK1, PAK1,
phospho-PAK4, PAK4 and GAPDH. All primary antibodies were purchased
from Cell Signaling Technology. The bound antibodies were
visualized using ECL reagents (GE Healthcare, Amersham, UK) and the
density of each band was analyzed using Multigauge computer
software (Berthold, Bundoora, Australia).
Statistical analysis
All values were expressed as mean ± standard error.
Results were analyzed by Student's t-test or one-way ANOVA (SPSS,
IBM, New York, NY, USA). Differences between two means with
p<0.05 were considered statistically significant.
Results
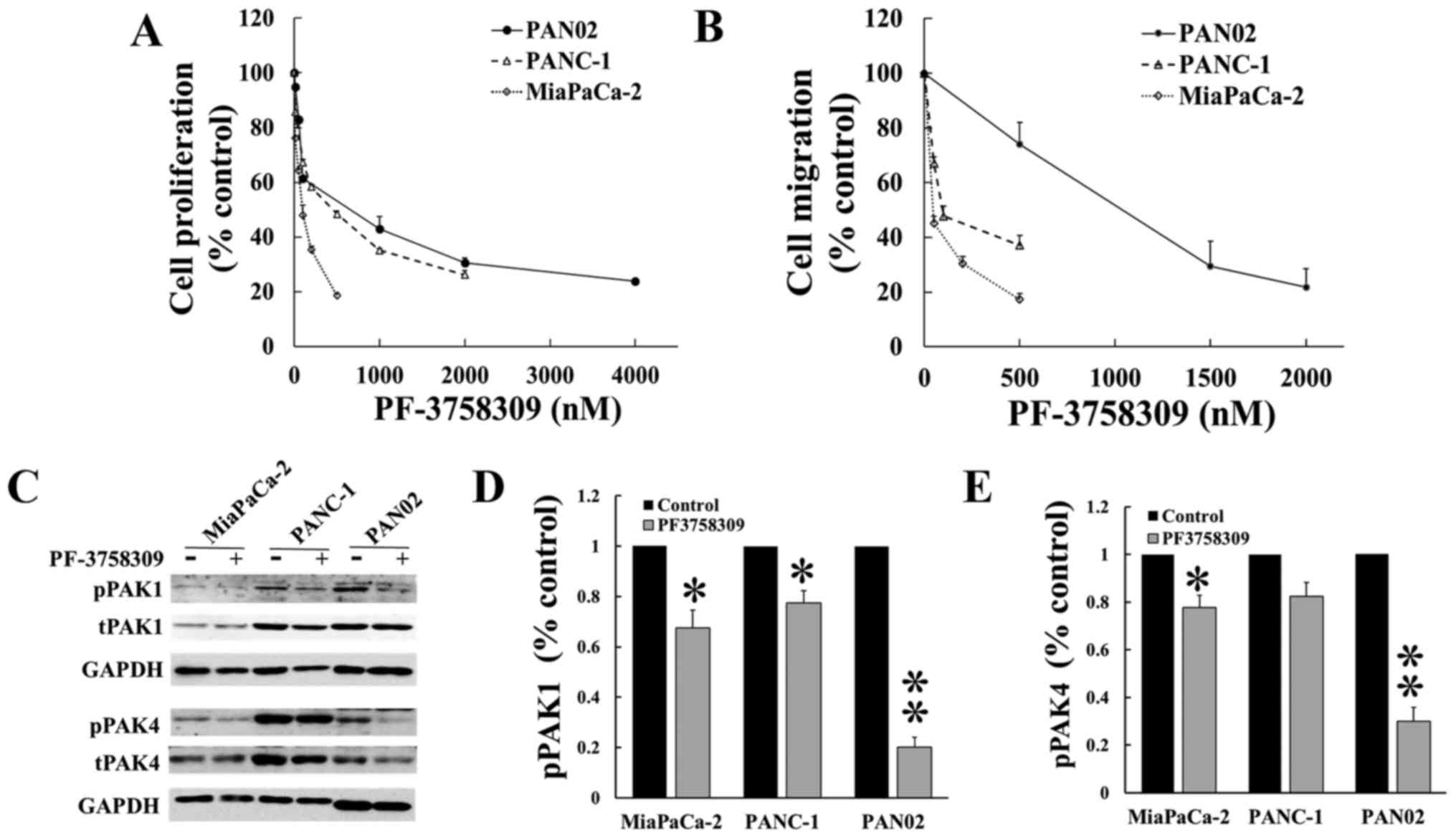
PF-3758309 inhibits pancreatic cancer
cell proliferation and migration
Human (MiaPaCa-2 and PANC-1) and murine (PAN02)
pancreatic cancer cells were cultured with increasing doses of
PF-3758309 (Fig. 1A and B) for 24
h. Cell proliferation and migration/invasion were measured by
3H-thymidine incorporation and Boyden chamber assays,
respectively. PF-3758309 decreased the proliferation and
migration/invasion of both human and murine pancreatic cancer cells
(Fig. 1A and B) in a
dose-dependent manner, the IC50 values are given in
Table I. The three pancreatic
cancer cell lines were then incubated for 24 h with PF-3758309 at
the IC50 value calculated from the proliferation assays
(Fig. 1A), and the expression of
phosphorylated and active PAK1 and PAK4, as well as total PAK1 and
PAK4, was measured by western blotting. PF-3758309 inhibited the
activity of PAK1 in the three cell lines tested (Fig. 1C), and the activity of PAK4 in
MiaPaCa-2 and PAN02. These data indicated that PF-3758309 supressed
pancreatic cancer cell proliferation and migration probably via
inhibition of the activities of both PAK1 and PAK4.
 | Table IThe IC50 for inhibition of
proliferation and migration/invasion of pancreatic cancer cell
lines by PF-3758309. |
Table I
The IC50 for inhibition of
proliferation and migration/invasion of pancreatic cancer cell
lines by PF-3758309.
| MiaPaCa-2 | PANC-1 | PAN02 |
|---|
| Proliferation
(nM) | 87+15 nM | 500+70 nM | 805+155 nM |
| Migration/invasion
(nM) | 60+10 nM | 141+35 nM | 1230+220 nM |
PF-3758309 inhibits pancreatic cancer
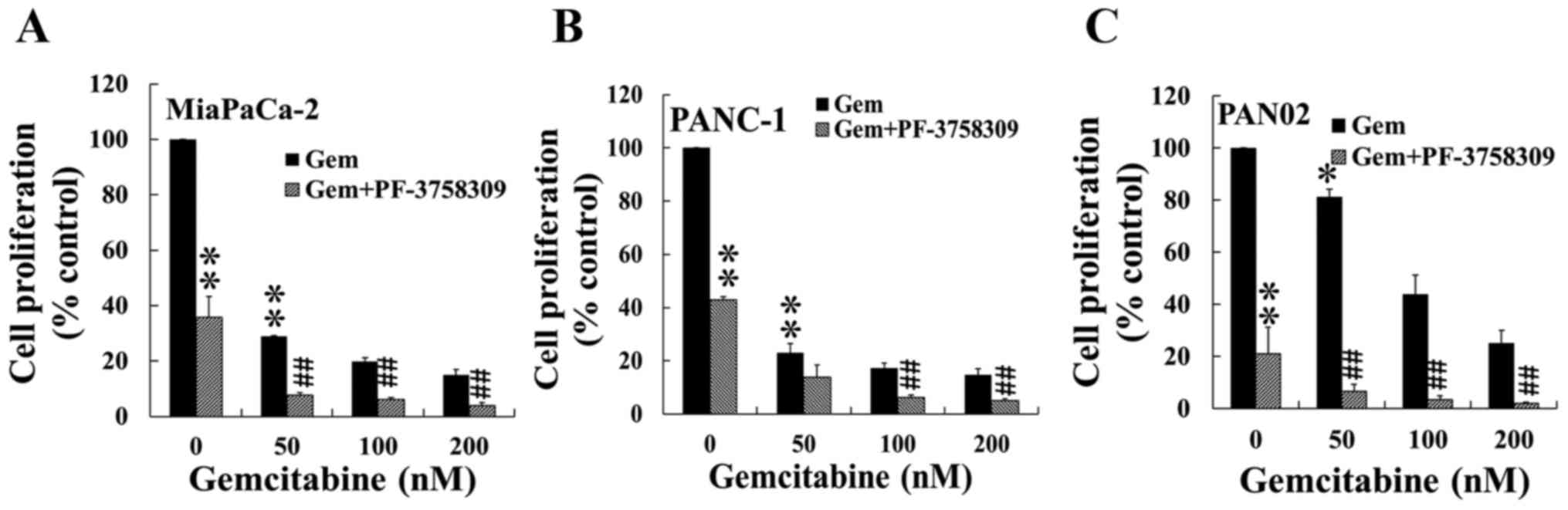
cell proliferation synergistically with gemcitabine
The combined effects of PF-3758309 and gemcitabine
on proliferation were measured by incubation of the pancreatic
cancer cell lines with different concentrations of gemcitabine,
alone or in combination with pre-treatment of PF-3758309, at
concentrations around the IC50 values given in Table I). Gemcitabine on its own inhibited
proliferation in the three pancreatic cancer cell lines in a
dose-dependent manner (Fig. 2). A
further reduction of proliferation was observed in all three cell
lines when the cells were treated with PF-3758309 and gemcitabine
in combination, compared to treatment with gemcitabine alone
(Fig. 2). The combined effect of
PF-3758309 and gemcitabine was shown to be synergistic rather than
additive when analysed by the Chou-Talalay method (17) (Fig.
3). These results demonstrated that PF-3758309 and gemcitabine
synergistically inhibited pancreatic cancer cell growth.
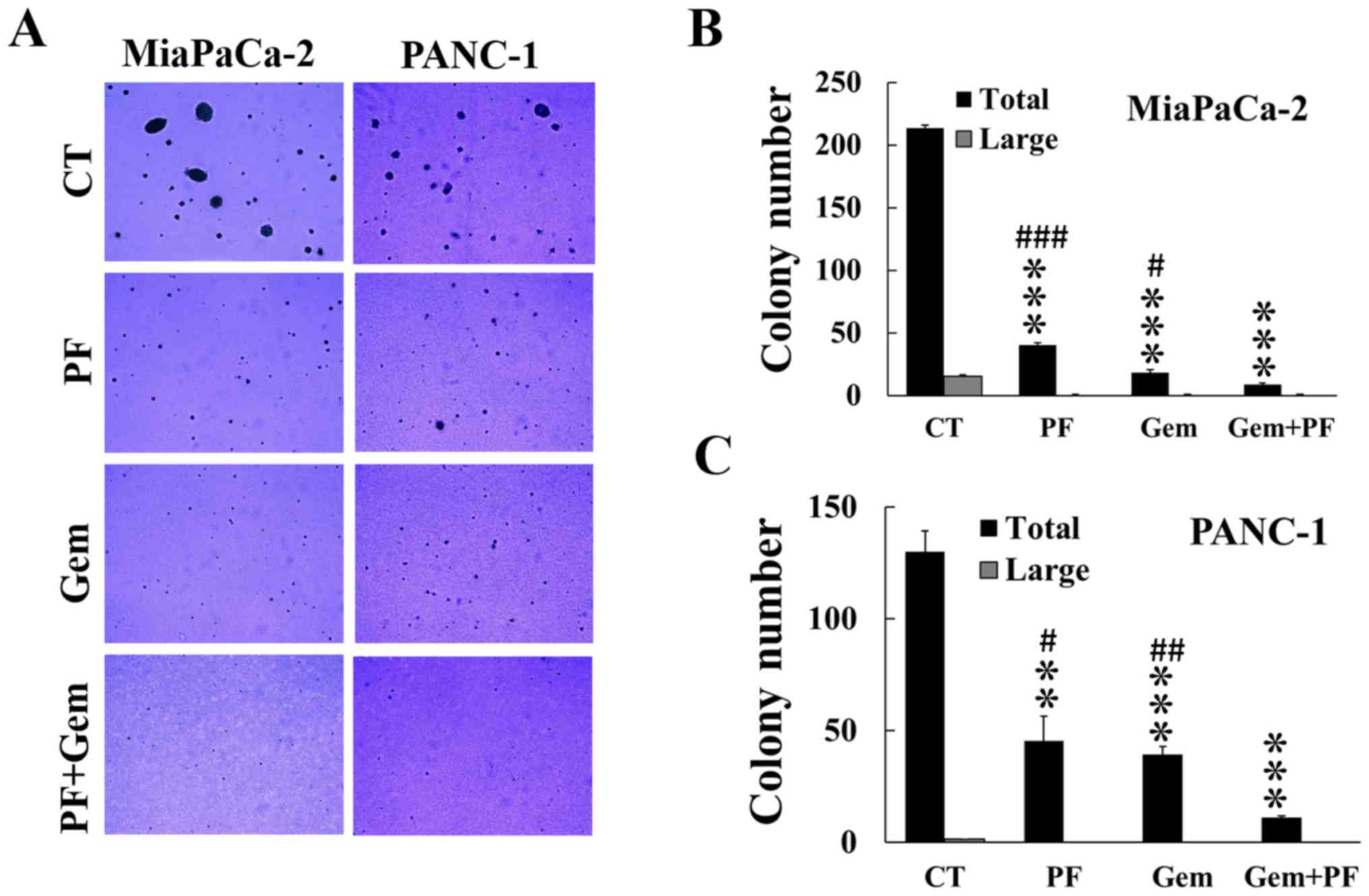
Inhibition of PAKs promotes the
suppression by gemcitabine of anchorage-independent growth of
pancreatic cancer cells
The combined effect of inhibition of PAK by
PF-3758309 with gemcitabine on anchorage-independent growth of
pancreatic cancer cells was measured by growing cells in soft agar,
followed by treatment with gemcitabine alone or in combination with
PF-3758309 at concentrations around the IC50 values
calculated from the proliferation assay. Either PF-3758309 or
gemcitabine alone decreased the numbers of colonies formed in the
agar in both MiaPaCa-2 and PANC-1 cells (Fig. 4). Combined treatment with
PF-3758309 and gemcitabine resulted in a significant further
decrease in the numbers of colonies formed compared with either
PF-3758309 or gemcitabine alone (Fig.
4). PAN02 did not grow in the soft agar under current
condition.
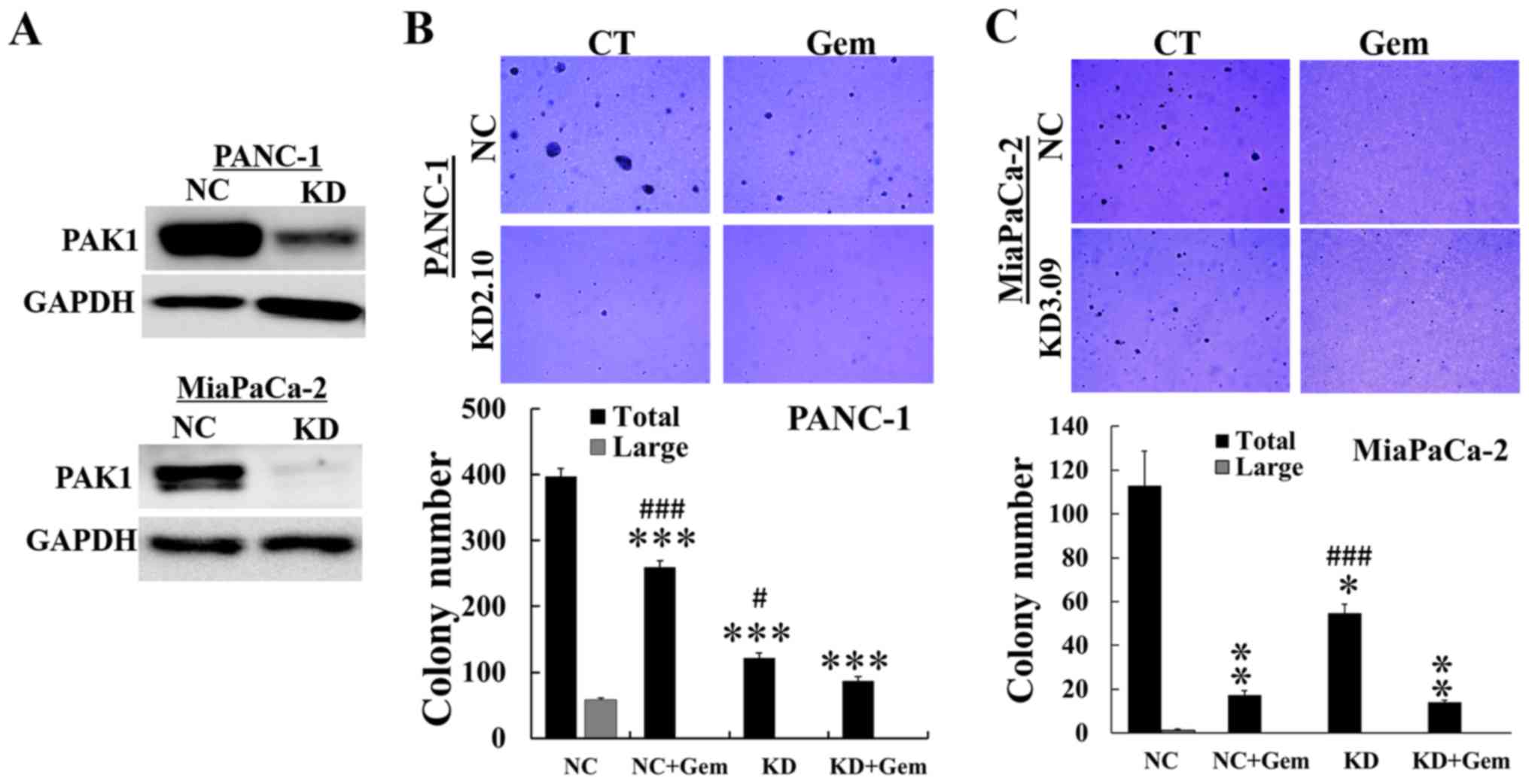
In order to confirm this result, PAK1 expression was
reduced by knockdown (KD) with shRNA (Fig. 5A), and the numbers of colonies
formed in PAK1 KD with or without gemcitabine were analysed. Cells
transfected with a scrambled shRNA sequence served as a negative
control (NC). The number of colonies formed in PAK1 KD cells was
significantly reduced compared to NC cells. Gemcitabine treatment
further decreased the colony numbers in PANC-1 PAK1 KD cells
(Fig. 5B), but not in MiaPaCa-2
PAK1 KD cells (Fig. 5C).
Furthermore, large colonies containing >100 cells only formed in
untreated NC cells. These data demonstrated that inhibition of PAKs
by PF-3758309 or by shRNA knockdown enhanced the suppressive effect
of gemcitabine on anchorage-independent growth of pancreatic cancer
cells.
Inhibition of PAKs sensitizes the
response of pancreatic cancers to gemcitabine in vivo
A mouse orthotopic pancreatic cancer model was used
to determine the effect of inhibition of PAKs on the response of
pancreatic cancers to gemcitabine treatment in vivo. Tumours
were induced by injection of pancreatic cancer cells into the tail
of the pancreas as previously described (16). The tumour take rate was 100% in
both PAK1 wild-type (WT) and PAK1 knockout (KO) C57BL/6 mice. None
of the mice died from surgery. In PAK1 WT mice, either PF-3758309
or gemcitabine alone significantly reduced tumour growth by
decreasing the tumour volume (Fig. 6A
and C) and tumour weight (Fig.
6B), compared to untreated controls. A further reduction in
both tumour volume and weight was observed in mice treated with the
combination of PF-3758309 and gemcitabine. The tumour volume, but
not the tumour weight, was significantly decreased in PAK1 KO mice
when compared to PAK1 WT mice. Gemcitabine treatment of PAK1 KO
mice caused a further reduction in both tumour volume and tumour
weight when compared to the tumours in untreated PAK1 KO mice.
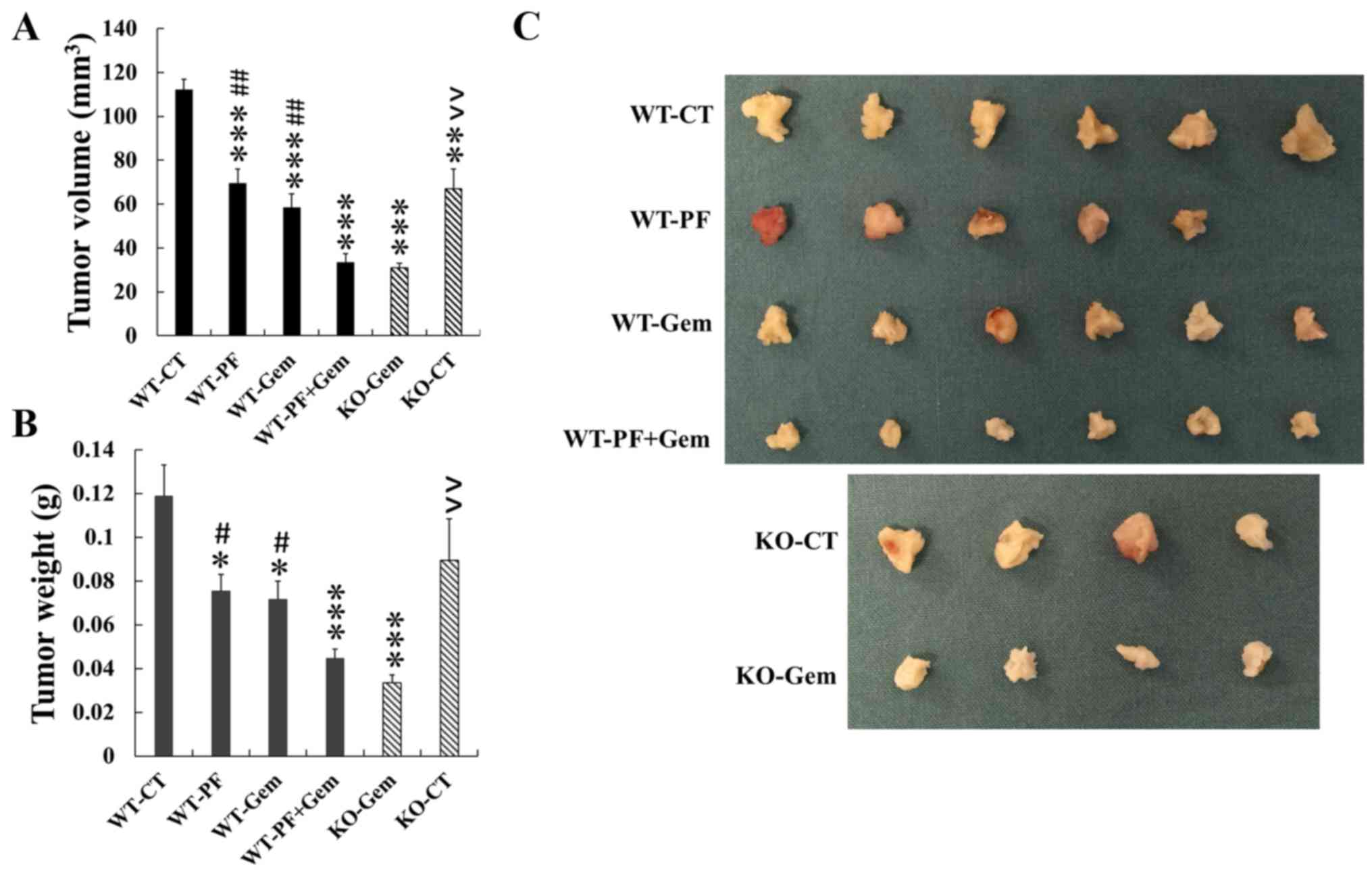 | Figure 6Inhibition of PAK enhances the
antitumour effect of gemcitabine on pancreatic cancer growth in an
orthotopic cancer model. Twenty-four wild-type (WT) male C57BL/6
mice were injected PAN02 cells (5×105 cells/50
μl/mouse) into the tail of pancreas. One week later the mice
were divided into 4 groups, 6 per group, and were treated by
intra-peritoneal injection with 5% DMSO saline (CT, control),
PF-3758309 (PF, 25 mg/kg), gemcitabine (Gem, 40 mg/kg), or PF plus
Gem for 4 weeks. Nine PAK1 knockout (KO) mice were subjected to
similar procedure. The PAK1 KO mice were divided into control (CT,
n=4) and gemcitabine-treated (Gem, n=5) groups. The mice were
sacrificed at the endpoint, their pancreata isolated, and tumour
volume (A) and weight (B) were measured. Tumour images were taken
(C). *p<0.05, **p<0.01,
***p<0.001, compared to the values from WT control
mice. #p<0.05, ##p<0.01, compared to
the values from WT mice treated with PF plus Gem,
^^p<0.01, ^^^p<0.001, compared to the
values from PAK1 KO mice treated with Gem. |
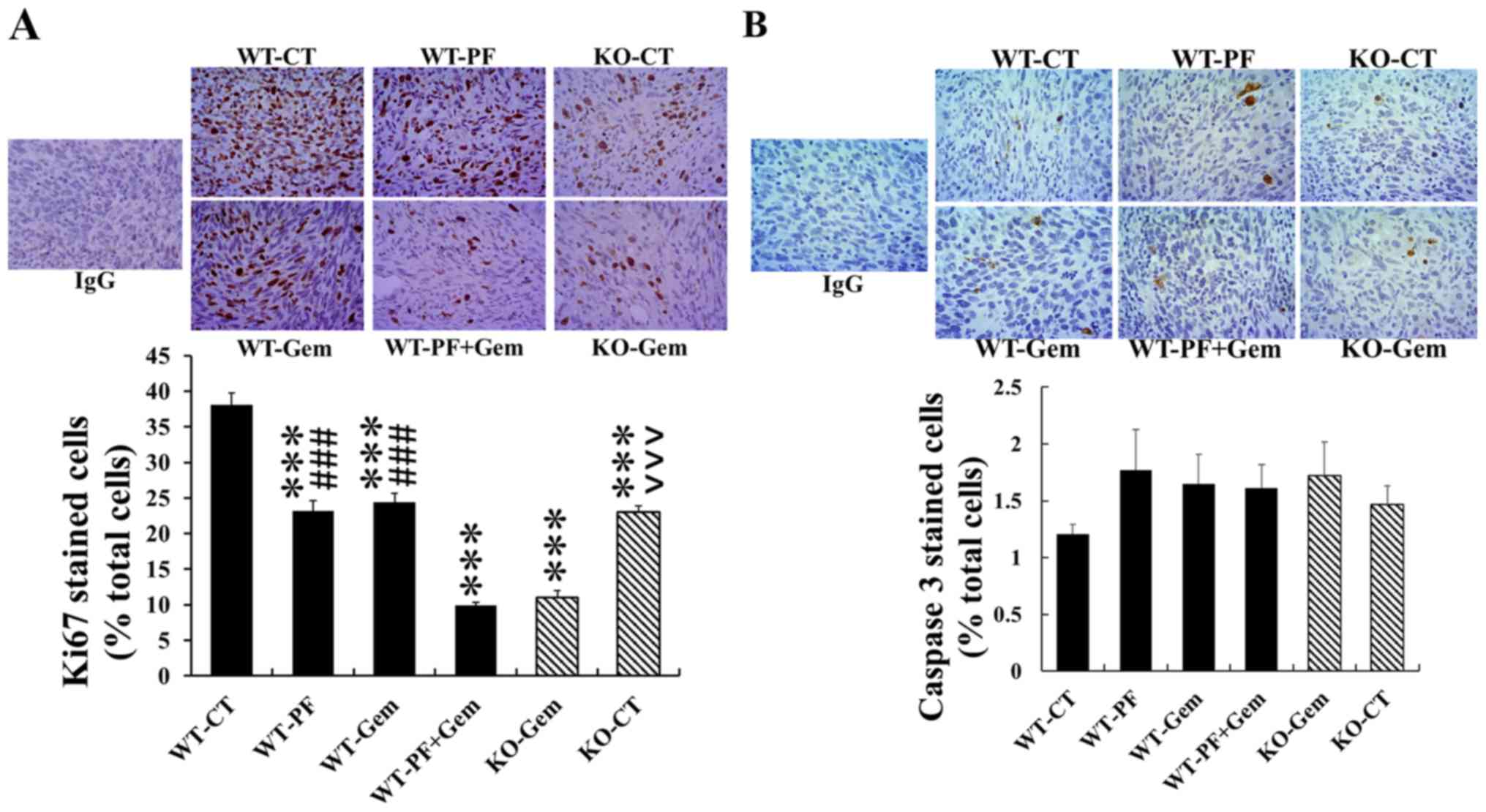
Either PF-3758309 or gemcitabine alone inhibited
tumour growth by decreasing cell proliferation as measured by Ki67
immunohistochemistry (Fig. 7A).
The proliferation in tumours from PAK1 WT mice treated with either
PF-3758309 or gemcitabine was reduced to 60.6% and 60.1% of that in
tumours from untreated mice. The combination of PF-3758309 and
gemcitabine further reduced the proliferation to 25.9% of the
untreated PAK1 WT mice. Importantly, the proliferation in tumours
from PAK1 KO mice was also significantly reduced compared to
tumours from untreated PAK1 WT mice. Gemcitabine treatment of PAK1
KO mice further decreased cell proliferation in tumours when
compared to the corresponding untreated PAK1 KO mice (Fig. 7A). No significant difference was
detected in apoptosis in tumours from all six groups although
compared to control of PAK1 WT mice, there were increased trend in
all other groups of treatment mice (Fig. 7B). These results indicate that
inhibition of PAKs by PF-3758309 or by PAK1 knockout sensitized
pancreatic cancers to gemcitabine in vivo at least by
decreasing cell proliferation.
Inhibition of PAKs enhances tumour immune
response in vivo
To determine the effect of PAK inhibition on tumour
immune response, spleens were dissected from the tumour-bearing
mice treated as shown in Fig. 6.
The red blood cells were removed, and the splenic cells were
extracted and subjected to FACS analysis as described in the
Materials and methods. In the PAK1 WT mice, both CD3+
and CD8+ T lymphocytes were increased by treatment with
PF-3758309 alone or in combination with gemcitabine (Fig. 8A). Both CD3+ and
CD8+ T lymphocytes were increased in untreated PAK1 KO
mice compared to untreated PAK1 WT mice. Gemcitabine treatment did
not cause any significant changes in CD3+ or
CD8+ T lymphocytes of either PAK1 WT or PAK1 KO mice
(Fig. 8A).
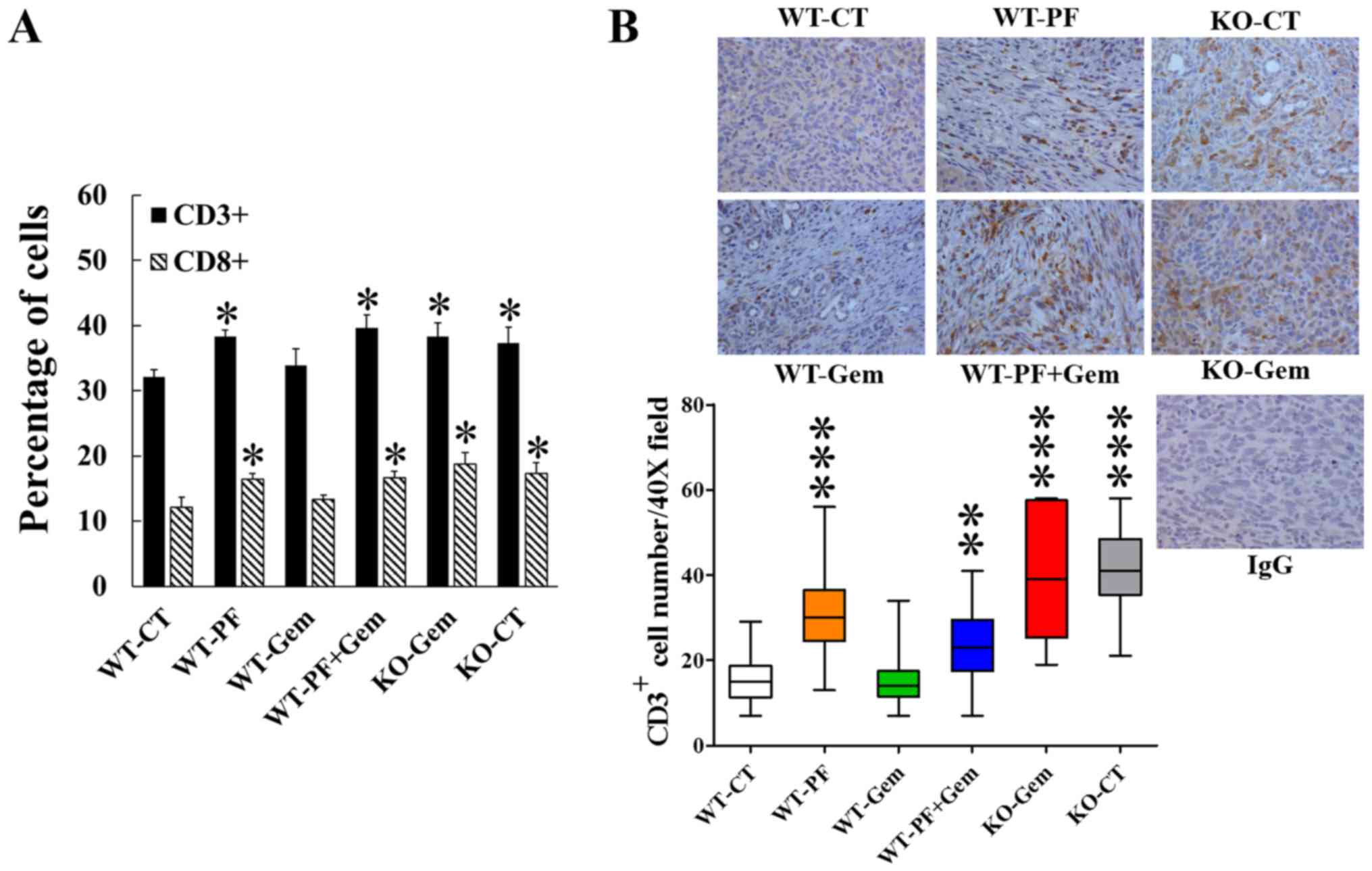 | Figure 8Inhibition of PAK upregulates the
immune system of mice bearing pancreatic tumours and stimulates
tumour immune infiltration. Fresh spleens were dissected out from
the mice described in Fig. 5. The
splenic lymphocytes were analysed by flow cytometry (A). The
percentage of CD3+ or CD8+ cells was
calculated using the Weasel computer program (Cytometry Laboratory,
Walter and Eliza Hall Institute, Parkville, Australia). The
pancreatic tumour tissues were taken from all 6 groups of mice and
stained for CD3+ cells, which were then counted under
the microscope at ×40 magnification (B). WT, wild-type PAK1; KO,
knockout PAK1; CT, control; Gem, gemcitabine; PF, PF-3758309.
*p<0.05, **p<0.01,
***p<0.001, compared to the values from PAK1 WT
control mice. |
CD3+ T lymphocytes were stained in
pancreatic tumour tissues from all 6 groups of mice to determine
the tumour-infiltrating immune cells. In the tumours in PAK1 WT
mice, CD3+ T lymphocytes were increased by treatment
with PF-3758309 alone or in combination with gemcitabine (Fig. 8B). CD3+ T lymphocytes
were also increased in tumours in untreated PAK1 KO mice compared
to tumours in untreated PAK1 WT mice. Gemcitabine treatment alone
did not cause any significant changes in CD3+ T
lymphocytes in tumours from either PAK1 WT or PAK1 KO mice
(Fig. 8B). These results indicate
that inhibition of PAKs by PF-3758309 or by PAK1 KO upregulated the
immune response of tumour-bearing mice by stimulation of both
splenic CD3+ and CD8+ T lymphocytes and
tumour-infiltrating CD3+ T lymphocytes.
Discussion
Our most important findings in this study are that,
in an orthotopic pancreatic cancer mouse model, inhibition of PAK1
and PAK4 by PF-3758309 or depletion of PAK1 not only suppressed the
growth of pancreatic cancer in vivo, but also stimulated the
immune response by increasing the numbers of splenic
CD3+ and CD8+ T lymphocytes as well as by
enhancing the tumour-infiltrating CD3+ T lymphocytes.
Tumour-infiltrating leukocytes (TILs) are critical determinants of
clinical outcome of cancer patients, and play key roles in cancer
progression and in therapeutic responses (18–20).
Among TILs, the activation and location of T lymphocytes are
particularly important. The location and density of intratumoural
CD8+ T lymphocytes alters their prognostic significance
(21). Infiltration of
CD3+ T lymphocytes is associated with a significantly
higher rate of progression-free survival of gastrointestinal cancer
patients (22). In PDA patients,
the presence of both CD4+ and CD8+ T
lymphocytes in tumour tissues correlated with an improved prognosis
and increased 5-year survival rate (23–25).
These pieces of evidence stimulated the initiation and development
of immune-therapeutic approaches to enhance the antitumour immune
response through regulation of TILs. Our finding that inhibition of
PAK1 and PAK4 by PF-3758309 or depletion of PAK1 increased the
number of the tumour infiltrating CD3+ T lymphocytes in
an orthotopic pancreatic cancer mouse model indicates the potential
role of PAK inhibitors in modulation of TILs in order to improve
antitumour immunity. Our finding also prompted us to further
investigate the role of PAKs in tumour immune response and tumour
microenvironment using KPC mouse (KrasG12D/+;
Trp53R172H/+; Pdx-1-Cre) which incorporates expression
of KrasG12D/+ and Trp53R172H/+ alleles
targeted to the mouse pancreas by Cre recombinase under the control
of the pancreas-specific Pdx-1 promoter and reproduces key features
of human PDA (26).
Gemcitabine, as the base-line therapeutic agent for
pancreatic cancer, has been reported to reduce the number of
immunosuppressive T regulatory cells (Treg) and myeloid-derived
suppressor cells (MDSC), while restoring the ratio of T effector
cells to Treg in the blood of pancreatic cancer patients (27). When comparing the number of
intratumoural immune cells from patients undertaking gemcitabine
treatment to the non-treated controls, gemcitabine did not change
the number of intratumoural CD3+ T lymphocytes, but
increased the ratios of CD3+/Treg and
CD4+/Treg, possibly by reducing the number of
FOXP3+ cells (28). In
agreement with these clinical observations in patients, in our
orthotopic pancreatic cancer model in mice, gemcitabine on its own
did not change the number of either splenic CD3+ or
CD8+ T lymphocytes or of intratumoural CD3+ T
lymphocytes (Fig. 8). The effect
of gemcitabine on Treg and MDSC in our model remains to be
determined.
Although targeted therapies and immunotherapy have
significantly improved the outcome of many solid malignancies such
as melanoma and lung cancer, no such effective therapy has been
developed for PDA, despite the knowledge of key mutations and an
increasing understanding of the tumour microenvironment. Current
treatment options for PDA mainly involve combination cytotoxic
chemotherapies, which provide a marginal survival benefit at the
cost of significant toxicity. Gemcitabine-based treatment is still
most commonly applied to pancreatic cancer patients because of its
general good patient tolerability and lack of better alternative
less toxic combinations of chemotherapies (29). Indeed the combination of targeted
therapies of gemcitabine, combined with agents, such as cetuximab
or bevacizumab, has not added any additional benefit (30,31).
Consistent with previous reports, we have shown that
inhibition of both PAK1 and PAK4 by PF-3758309 suppressed the
growth and migration/invasion of pancreatic cancer cells. Although
the Boyden chamber assay is recognised as a standard method to
measure cell migration/invasion (in the case of cancer cells). The
measurement of MMP-2 and MMP-9 will certainly strengthen our data
here, which will be added in our future study. Further we have
demonstrated that PF-3758309, either on its own or synergistically
with gemcitabine, reduced the growth of pancreatic cancer both
in vitro and in vivo. The inhibitory effect of
PF-3758309 on three cell lines tested seems to be correlated well
with the activity of PAK1 as MiaPaCa-2 with lowest level of active
phosphorylated PAK1 (pPAK1) responded to PF-378309 most sensitively
while PAN02 with highest level of pPAK1 responded to PF-3758309
least sensitively (Fig. 1 and
Table I). That the inhibitory
effect of PF-3758309 is mainly mediated by a PAK1-dependent pathway
is clearly demonstrated by our in vivo observation in the
orthotopic pancreatic cancer model that tumour growth was inhibited
by PF-3758309 in PAK1 wild-type mice to a similar degree to the
tumour growth inhibition by PAK1 depletion in PAK1 knockout mice
(Fig. 6). When determining the
hematologic toxicities by full blood count, PF-3758309 and
gemcitabine alone or in combination reduced the haemoglobin by 10%
while PF-3758309 increased the platelet and neutrophils by 50% and
110%, respectively (data not shown). PF-3758309 and/or gemcitabine
did not affect any other hematologic index tested. Our findings
that inhibition of PAKs not only suppressed pancreatic cancer
growth (both alone and synergistically with gemcitabine), but also
upregulated the tumour immune response by increasing the splenic
and intratumoural lymphocytes, make PAKs in general, and PAK1 in
particular, an attractive target(s) in developing effective
therapeutic regimen.
In conclusion, our results that inhibition of PAK1
and PAK4 suppressed pancreatic cancer growth/invasion, stimulated
the tumour immune response, and promoted the inhibitory effect of
gemcitabine, indicate the potential role of PAKs in the immune
response to pancreatic tumours, and provide a solid base for the
development of a combination treatment of PAK targeted therapy with
gemcitabine to improve the outcome for PDA patients.
Acknowledgments
The authors would like to thank Professor Mauro S.
Sandrin for help in the flow cytometry analysis. This study was
supported by grants from the National Health and Medical Research
Council of Australia (1041831, GSB/AS), and the Pancare Foundation
(GSB, MN). H.H. was supported by Baldwin trust fund. K.W. wass
supported by Melbourne International Fee Remission Scholarship
(MIFRS), Melbourne International Research Scholarship (MIRS) and
Pancare Foundation (Moshe Sambor Scholarship).
Glossary
Abbreviations
Abbreviations:
|
PAK
|
p21 activated kinase
|
|
PDA
|
pancreatic ductal adenocarcinoma
|
|
Treg
|
T regulatory cells
|
|
MDSC
|
myeloid-derived suppressor cells
|
|
TILs
|
tumour-infiltrating leukocytes
|
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
Statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Von Hoff DD, Ervin T, Arena FP, Chiorean
EG, Infante J, Moore M, Seay T, Tjulandin SA, Ma WW, Saleh MN, et
al: Increased survival in pancreatic cancer with nab-paclitaxel
plus gemcitabine. N Engl J Med. 369:1691–1703. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang Y, Hochster H, Stein S and Lacy J:
Gemcitabine plus nab-paclitaxel for advanced pancreatic cancer
after first-line FOLFIRINOX: Single institution retrospective
review of efficacy and toxicity. Exp Hematol Oncol. 4:292015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sharma P and Allison JP: The future of
immune checkpoint therapy. Science. 348:56–61. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zheng L, Xue J, Jaffee EM and Habtezion A:
Role of immune cells and immune-based therapies in pancreatitis and
pancreatic ductal adenocarcinoma. Gastroenterology. 144:1230–1240.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
He H and Baldwin GS: p21-activated kinases
and gastrointestinal cancer. Biochim Biophys Acta. 1833:33–39.
2013. View Article : Google Scholar
|
|
7
|
Yeo D, He H, Patel O, Lowy AM, Baldwin GS
and Nikfarjam M: FRAX597, a PAK1 inhibitor, synergistically reduces
pancreatic cancer growth when combined with gemcitabine. BMC
Cancer. 16:242016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tyagi N, Bhardwaj A, Singh AP, McClellan
S, Carter JE and Singh S: p-21 activated kinase 4 promotes
proliferation and survival of pancreatic cancer cells through AKT-
and ERK-dependent activation of NF-κB pathway. Oncotarget.
5:8778–8789. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tyagi N, Marimuthu S, Bhardwaj A, Deshmukh
SK, Srivastava SK, Singh AP, McClellan S, Carter JE and Singh S:
p-21 activated kinase 4 (PAK4) maintains stem cell-like phenotypes
in pancreatic cancer cells through activation of STAT3 signaling.
Cancer Lett. 370:260–267. 2016. View Article : Google Scholar
|
|
10
|
Moon SU, Kim JW, Sung JH, Kang MH, Kim SH,
Chang H, Lee JO, Kim YJ, Lee KW, Kim JH, et al: p21-activated
kinase 4 (PAK4) as a predictive marker of gemcitabine sensitivity
in pancreatic cancer cell lines. Cancer Res Treat. 47:501–508.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Biankin AV, Waddell N, Kassahn KS, Gingras
MC, Muthuswamy LB, Johns AL, Miller DK, Wilson PJ, Patch AM, Wu J,
et al Australian Pancreatic Cancer Genome Initiative: Pancreatic
cancer genomes reveal aberrations in axon guidance pathway genes.
Nature. 491:399–405. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Murray BW, Guo C, Piraino J, Westwick JK,
Zhang C, Lamerdin J, Dagostino E, Knighton D, Loi CM, Zager M, et
al: Small-molecule p21-activated kinase inhibitor PF-3758309 is a
potent inhibitor of oncogenic signaling and tumor growth. Proc Natl
Acad Sci USA. 107:9446–9451. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Huynh N, Liu KH, Baldwin GS and He H:
p21-activated kinase 1 stimulates colon cancer cell growth and
migration/invasion via ERK- and AKT-dependent pathways. Biochim
Biophys Acta. 1803:1106–1113. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yeo D, Huynh N, Beutler JA, Christophi C,
Shulkes A, Baldwin GS, Nikfarjam M and He H: Glaucarubinone and
gemcitabine synergistically reduce pancreatic cancer growth via
down-regulation of P21-activated kinases. Cancer Lett. 346:264–272.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bang D, Wilson W, Ryan M, Yeh JJ and
Baldwin AS: GSK-3α promotes oncogenic KRAS function in pancreatic
cancer via TAK1-TAB stabilization and regulation of noncanonical
NF-κB. Cancer Discov. 3:690–703. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Nikfarjam M, Yeo D, He H, Baldwin G, Fifis
T, Costa P, Tan B, Yang E, Wen Sw and Christophi C: Comparison of
two syngeneic orthotopic murine models of pancreatic
adenocarcinoma. J Invest Surg. 26:352–359. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chou TC and Talalay P: Quantitative
analysis of dose-effect relationships: The combined effects of
multiple drugs or enzyme inhibitors. Adv Enzyme Regul. 22:27–55.
1984. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Newman AM and Alizadeh AA: High-throughput
genomic profiling of tumor-infiltrating leukocytes. Curr Opin
Immunol. 41:77–84. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Junttila MR and de Sauvage FJ: Influence
of tumour microenvironment heterogeneity on therapeutic response.
Nature. 501:346–354. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Rooney MS, Shukla SA, Wu CJ, Getz G and
Hacohen N: Molecular and genetic properties of tumors associated
with local immune cytolytic activity. Cell. 160:48–61. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Peske JD, Woods AB and Engelhard VH:
Control of CD8 T-cell infiltration into tumors by vasculature and
microenvironment. Adv Cancer Res. 128:263–307. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rusakiewicz S, Semeraro M, Sarabi M,
Desbois M, Locher C, Mendez R, Vimond N, Concha A, Garrido F,
Isambert N, et al: Immune infiltrates are prognostic factors in
localized gastrointestinal stromal tumors. Cancer Res.
73:3499–3510. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sideras K, Braat H, Kwekkeboom J, van
Eijck CH, Peppelenbosch MP, Sleijfer S and Bruno M: Role of the
immune system in pancreatic cancer progression and immune
modulating treatment strategies. Cancer Treat Rev. 40:513–522.
2014. View Article : Google Scholar
|
|
24
|
Ino Y, Yamazaki-Itoh R, Shimada K, Iwasaki
M, Kosuge T, Kanai Y and Hiraoka N: Immune cell infiltration as an
indicator of the immune microenvironment of pancreatic cancer. Br J
Cancer. 108:914–923. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fukunaga A, Miyamoto M, Cho Y, Murakami S,
Kawaradaz Y, Oshikiri T, Kato K, Kurokawa T, Suzuoki M, Nakakubo Y,
et al: CD8+ tumor-infiltrating lymphocytes together with
CD4+ tumor-infiltrating lymphocytes and dendritic cells
improve the prognosis of patients with pancreatic adenocarcinoma.
Pancreas. 28:e26–e31. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hingorani SR, Wang L, Multani AS, Combs C,
Deramaudt TB, Hruban RH, Rustgi AK, Chang S and Tuveson DA:
Trp53R172H and KrasG12D cooperate to promote chromosomal
instability and widely metastatic pancreatic ductal adenocarcinoma
in mice. Cancer Cell. 7:469–483. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Eriksson E, Wenthe J, Irenaeus S, Loskog A
and Ullenhag G: Gemcitabine reduces MDSCs, tregs and TGFβ-1 while
restoring the teff/treg ratio in patients with pancreatic cancer. J
Transl Med. 14:2822016. View Article : Google Scholar
|
|
28
|
Shibuya KC, Goel VK, Xiong W, Sham JG,
Pollack SM, Leahy AM, Whiting SH, Yeh MM, Yee C, Riddell SR, et al:
Pancreatic ductal adenocarcinoma contains an effector and
regulatory immune cell infiltrate that is altered by multimodal
neoadjuvant treatment. PLoS One. 9:e965652014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Manji GA, Olive KP, Saenger YM and
Oberstein P: Current and emerging therapies in metastatic
pancreatic cancer. Clin Cancer Res. 23:1670–1678. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Philip PA, Benedetti J, Corless CL, Wong
R, O'Reilly EM, Flynn PJ, Rowland KM, Atkins JN, Mirtsching BC,
Rivkin SE, et al: Phase III study comparing gemcitabine plus
cetuximab versus gemcitabine in patients with advanced pancreatic
adenocarcinoma: Southwest Oncology Group-directed intergroup trial
S0205. J Clin Oncol. 28:3605–3610. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kindler HL, Niedzwiecki D, Hollis D,
Sutherland S, Schrag D, Hurwitz H, Innocenti F, Mulcahy MF,
O'Reilly E, Wozniak TF, et al: Gemcitabine plus bevacizumab
compared with gemcitabine plus placebo in patients with advanced
pancreatic cancer: Phase III trial of the Cancer and Leukemia Group
B (CALGB 80303). J Clin Oncol. 28:3617–3622. 2010. View Article : Google Scholar : PubMed/NCBI
|