Introduction
Recurrent and/or metastatic head and neck squamous
cell carcinoma (R/M HNSCC) is a devastating malignancy with a poor
prognosis. Treatment is limited to chemotherapeutic approaches. In
recent studies, the programmed-death receptor-1 (PD-1 and
CD279)/programmed-death ligand 1 (PD-L1, B7-H1 and CD274) pathway
has been indicated to be critical for regulating T-cell responses
and maintaining immune suppression (1–4).
HNSCC is a malignant tumour found most often in the head and neck
region involving the mandibular gingiva and oral floor (5,6); it
is generally highly immunosuppressive, for which PD-L1 expression
has been proposed as a potential causative mechanism (7–9). In
many tumours, expression of PD-L1 is constitutive and is induced by
interferon-γ (IFN-γ). PD-L1 binds to an inhibitory receptor (PD-1),
which is a member of the B7 family of receptors (4), and to the co-stimulatory molecule,
CD80 (B7-1) (10). PD-1 is
expressed on activated T cells; when PD-1 ligates with
tumour-associated PD-L1, the apoptosis or downregulation of
effector cytotoxic T lymphocytes is induced, thereby resulting in
an escape from T-cell-mediated immune surveillance (11). Tumour growth is reduced and
survival is improved when the PD-L1/PD-1 pathway is blocked
(7,12–15).
Moreover, clinical studies have indicated that tumour regression by
the blockade of the PD-1/PD-L1 checkpoint is durable (1,2),
leading to the recent registration of an anti-PD-1 antibody for
head and neck cancer. In treated tumours, the response to the PD-1
blockade has been commonly been associated with PD-L1 expression
(1).
However, anti-PD-1 treatment appears to be
beneficial only in certain patients. Therefore, the mechanisms
underlying the unknown regulation of PD-L1 expression in the HNSCC
microenvironment remain to be elucidated. PD-L1 expression has been
shown to be lower in R/M HNSCC cell lines than in a low-grade
invasive and less metastatic HNSCC cell lines, and PD-L1 expression
and epithelial-mesenchymal transition (EMT) have been found to be
closely related (16). PD-L1
expression has been shown to be upregulated in macrophages and
dendritic cells (DCs) in human R/M HNSCC tissues or co-cultured
with R/M HNSCC cells in vitro (16). However, the mechanisms responsible
for the fact that PD-L1-expressing HNSCC cells exhibit low
invasiveness and are less metastatic remain to be determined.
The immunosuppressive capacity of PD-1 ligands on
fibroblasts may be limited by their matrix metalloproteinase
(MMP)-dependent cleavage, thereby contributing to the aggravation
of inflammation in tissues (17).
Conversely, MMP activity seems to deplete PD-1 ligands in
carcinoma-associated fibroblasts, which may impair the physical
deletion of exhausted defective memory T cells through apoptosis
and may facilitate their regulatory functions (17). As MMPs are a group of proteolytic
enzymes that can degrade principal components of the extracellular
matrix, they are widely believed to play an important role in
tissue degradation. Several sets of experimental and clinical data
concerning MMPs in the contexts of cancer have been reported
(18,19). Numerous MMP inhibitors have
exhibited efficacy in animal models of disease and have been used
in clinical trials in the treatment of cancer, with some studies
focusing on rheumatoid arthritis and osteoarthritis. However, MMP
inhibitors have not exhibited significant therapeutic effects in
any of these human clinical trials (20). The use of these inhibitors also
results in adverse effects, including musculoskeletal pain,
tendonitis and mild anaemia with elevated liver enzyme levels
(20). Therefore, the function of
MMP needs to be redefined. MMPs influence basic processes, such as
cell proliferation, differentiation, angiogenesis and apoptosis
(18). Notably, the MMP family of
proteins exert dual roles in the pathogenesis of inflammation:
Stimulating protective innate and/or adaptive immune functions, as
well as tissue destruction (21).
To predict the efficacy of and optimise anti-PD-1
therapy, alone or in combination with other treatment options, it
is important to elucidate the mechanisms controlling PD-L1
expression. In this study, we thus focused on the regulation of
PD-L1 expression in HNSCC, and discussed the mechanism of this
regulation of PD-L1 expression in the tumour micro-environment.
Materials and methods
Cell culture
Three HNSCC cell lines originally established from
tumour biopsies with different grades of invasive or metastatic
abilities were used, including OSC-20 cells (with low
invasiveness), OSC-19 cells (intermediate invasiveness) and HOC313
cells (recurrent high-grade invasiveness and metastasis). The
OSC-20 cell line was originally derived from a 58-year-old female
with tongue cancer (22). OSC-19
was derived from a 61-year-old male with tongue cancer metastatic
to the cervical lymph nodes (23).
HOC313 was derived from a 51-year-old female with HNSCC (involving
the mandibular gingiva and oral floor) that metastasised to the
cervical lymph nodes and recurred (24). The HOC313 cells were a kind gift
from Dr M. Nagayama (Tokushima University, Tokushima, Japan). The
OSC-20 (JCRB #0197) and OSC-19 (JCRB #0198) cells, and normal human
oral fibroblasts of the lip mucosa (KD; JCRB #9103) were obtained
from the JCRB Cell Bank (Osaka, Japan). DCs were generated from
human peripheral blood mononuclear cells (PBMCs), as previously
described (25,26). Experiments using human samples were
approved by the Ethics Committee of the Kanazawa University
Graduate School of Medical Science (IRB no. 352-2), and written
informed consent was obtained from persons providing human samples.
Peripheral blood was voluntarily donated by 3 healthy individuals.
PBMCs were obtained by venepuncture into an 8-ml Vacutainer CPT
Cell-Preparation Tube (BD Vacutainer Systems, Franklin Lakes, NJ,
USA). Monocyte-derived DCs were generated by incubating monocytes
at 1×106 cells/ml in G4 medium (G4 Dendritic Cell
Generation kit; HumanZyme, Chicago, IL, USA) at 37°C in a
CO2 (5%) incubator for 7 days. The induced DCs were
examined using an anti-DC antibody (CD83; Abcam, Tokyo, Japan).
Eribulin (also known as Halaven; HAL) was purchased from Eisai Co.,
Ltd. (Tsukuba, Japan). Vinblastine (VBL) and paclitaxel (PTX;
Taxol) were purchased from Nihon Kayaku (Tokyo, Japan).
RNA extraction, cDNA synthesis and
quantitative (real-time) PCR (qPCR)
The mRNA expression levels of PD-L1, and MMP-1, -2,
-3, -7, -8, -9, -10, -11, -12, -13 and -14 were analysed using a
Rotor-Gene Q 2plex System (Qiagen, Hilden, Germany) with
FAM/ZEN/IBFQ probes (Integrated DNA Technologies, Inc., Coralville,
IA, USA; DNA sequences not available). Total RNA was extracted
using the PureLink RNA mini kit (Thermo Fisher Scientific, Waltham,
MA, USA), and cDNA was obtained using the PrimeScript First-Strand
cDNA synthesis kit (Takara, Tokyo, Japan). All reactions were
performed in accordance with the manufacturer's instructions. We
amplified 18S rRNA as an internal standard using HEX/ZEN/IBFQ
probes (Integrated DNA Technologies, Inc.; DNA sequences not
available). Relative expression levels were calculated using the
ΔΔCt method for qPCR (27), which
presents the data as fold differences in expression level relative
to a calibrator sample, in this case represented by the mean
expression of 3 experimental measurements of 18S rRNA in the
control cells or vehicle (buffer only control)-treated cells.
Western blot analysis
The concentration of total extracted protein was
measured after 100-fold dH2O dilution by the Qubit
Protein assay, in accordance with the manufacturer's instructions
(Thermo Fisher Scientific). An equal amount (30 μg of
protein) of lysate or culture medium was mixed with loading buffer,
with this mixture then being electrophoretically separated and
transferred onto membranes. The membranes were blocked with
Blocking One (Nacalai Tesque, Kyoto, Japan), followed by incubation
with anti-PD-L1 or MMP-7, -12, -13 antibodies (PD-L1, cat. no.
210931; MMP-7, cat. no. 205525; MMP-12, cat. no. 52897; MMP-13,
cat. no. 51072; Abcam) and an anti-human β-actin antibody (cat. no.
4970; Cell Signaling Technology, Tokyo, Japan). After washing with
Tris-buffered saline (TBS) with 0.05% Tween, the membranes were
incubated with horseradish peroxidase-conjugated anti-mouse IgG.
After washing with TBS-0.05% Tween-20, membranes were incubated
with the ECL Prime Western Blotting Detection Reagent (GE
Healthcare, Little Chalfont, UK). Signals were detected and
analysed using C-DiGit (M&S TechnoSystems, Tokyo, Japan).
Enzyme-linked immunosorbent assay
(ELISA)
The membrane-associated proteins were extracted
separately from the cytosolic proteins using the Mem-PER Plus
Membrane Protein Extraction kit (Thermo Fisher Scientific). The
extracted membrane-associated proteins were analysed for the
presence of PD-L1 using the ELISA kit (cat. no. DY156; R&D
Systems Europe Ltd., Abingdon, UK), in accordance with the
manufacturer's instructions. The total membrane-associated protein
concentration of the sample was measured by the Qubit Protein assay
in accordance with the manufacturer's instructions (Thermo Fisher
Scientific). Values were calculated as pg/mg total protein or ng/mg
total protein. Data are presented as the means ± standard error of
the mean (SEM).
Expression and purification of
recombinant human PD-L1
Human PD-L1 cDNA (cat. no. RDC1087) was purchased
from R&D Systems Europe Ltd. The human PD-L1 coding sequence
and 6×His-Tag were fused using PCR and the InFusion HD kit (Takara
Bio) combined with pSG5 (Agilent Technologies Japan, Tokyo, Japan)
to construct the expression plasmid pSG-hPDL1-His. The sequence was
confirmed by sequencing.
The Expi293 cells were cultured in Expi293
Expression Medium (both from Thermo Fisher Scientific) and
transfected with the pSG-hPDL1-His using FuGENE 6 transfection
reagent (Promega, Tokyo, Japan). The culture media were harvested
at 6 days after transfection and used for purification. They were
concentrated using an Amicon Diaflo apparatus fitted with a YM-10
membrane (both from Merck Japan, Tokyo, Japan) and applied to
complete His-Tag Purification Resin (Roche Diagnostics, Basel,
Switzerland) equilibrated with 50 mM Tris-HCl (pH 7.5), 0.15 M NaCl
and 0.05% Brij 35. Recombinant human PD-L1 was eluted with 500 mM
imid-azole in 50 mM Tris-HCl (pH 7.5) buffer after washing the
column with 50 mM Tris-HCl (pH 7.5) buffer containing 5 mM
imidazole and 0.3 M NaCl. The combined fractions of human PD-L1
were dialysed against 50 mM Tris-HCl (pH 7.5), 0.15 M NaCl and
0.05% Brij 35 buffer to remove imidazole. The protein concentration
was determined using the Qubit Protein assay, in accordance with
the manufacturer's instructions (Thermo Fisher Scientific). Eluted
fractions containing a major species of Mr 56,000 and minor protein
bands of Mr 46,000 were used as the source of PD-L1. Both minor
species were recognised by an anti-PD-L1 antibody (PD-L1, cat. no.
210931; Abcam), while only major species were recognised by an
anti-His antibody (cat. no. D291-3; MBL, Tokyo, Japan).
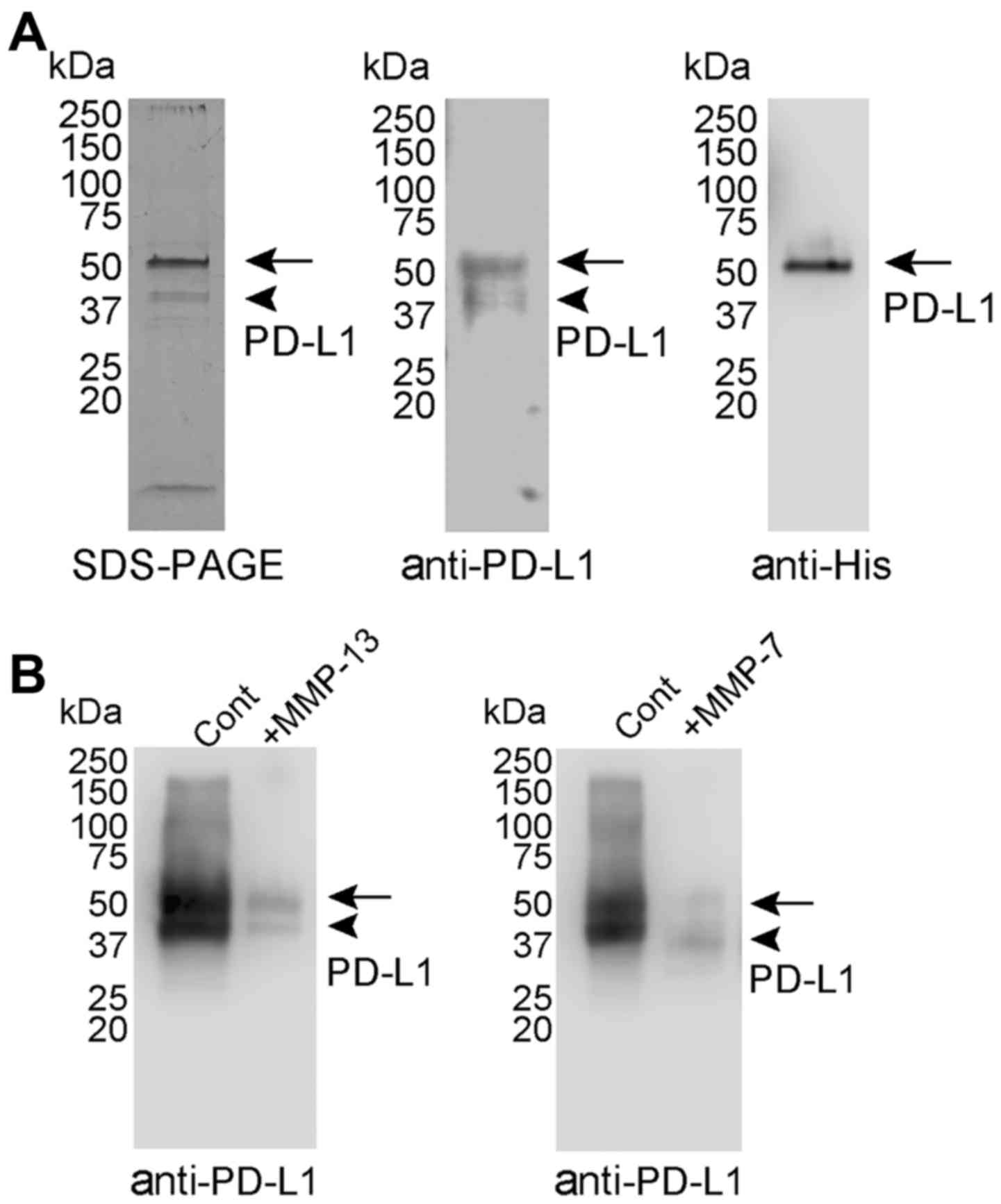
Degradation of PD-L1 by MMP-13 and
-7
The digestion of PD-L1 was examined by incubation of
the purified PD-L1 at 37°C for 24 h with MMP-13 or -7 at an
enzyme/substrate molar ratio of 1:30 in 50 mM Tris-HCl (pH 7.5),
0.15 M NaCl, 10 mM CaCl2 and 0.05% Brij 35 (TNCB
buffer). Human recombinant MMP-13 (cat. no. 444287) and MMP-7 (cat.
no. CC1057) were purchased from Merck Japan. MMPs were activated by
incubation with p-aminophenylmercuric acetate (Merck Japan), in
accordance with previously reported methods (28,29).
These reactions were terminated with 20 mM
ethylenediaminetetraacetic acid (EDTA) and the degradation products
were examined by western blot analysis using anti-PD-L1 antibody
(PD-L1, cat. no. 210931; Abcam).
Inhibition of PD-L1 cell surface
degradation in HNSCC cells
For the incubation of HNSCC cells with MMP
inhibitors, the HNSCC cells were seeded at a concentration of
5×104 cells/cm2 in P6 plates in DMEM medium
(Sigma-Aldrich Japan, Tokyo, Japan) supplemented with 5% FCS and
were then incubated on day 1 in the presence or absence of 2 MMP
inhibitors (10 μg/ml CL82198, cat. no. 141576; and 10
μM BB94, cat. no. 142087; Abcam) and maintained in culture
for 4 days at 37°C without a medium change. Subsequently,
membrane-associated proteins were extracted separately using the
Mem-PER Plus Membrane Protein Extraction kit (Thermo Fisher
Scientific). The extracted membrane-associated proteins were
analysed for the presence of PD-L1 using the ELISA kit (cat. no.
DY156; R&D Systems Europe Ltd.), in accordance with the
manufacturer's instructions. The total protein concentration of the
sample was measured after 100-fold dH2O dilution using
the Qubit Protein assay, in accordance with the manufacturer's
instructions (Thermo Fisher Scientific). Values were calculated as
pg/mg total protein or ng/mg total protein. The PD-L1 expression
recovery (%) was expressed as follows: Ratio (%) = difference
between the amount of PD-L1 in HNSCC cells incubated in the
presence of MMP inhibitors on day 4 and the amount of PD-L1 in
HNSCC cells in the absence of MMP inhibitors on day 4/the
difference between the amount of PD-L1 in HNSCC cells in the
absence of MMP inhibitors on day 1 and the amount of PD-L1 in HNSCC
cells in the absence of MMP inhibitors on day 4 ×100.
Statistical analysis
For comparisons between samples, data were analysed
by ANOVA and Tukey's multiple comparison tests using SPSS, version
23 software (IBM SPSS Statistics, IBM Corp., NY, USA). A P-value
≤0.05 or less was accepted as statistically significant.
Results
PD-L1 expression in HNSCC cell lines and
cell surface PD-L1 degradation in OSC-20 cell lines
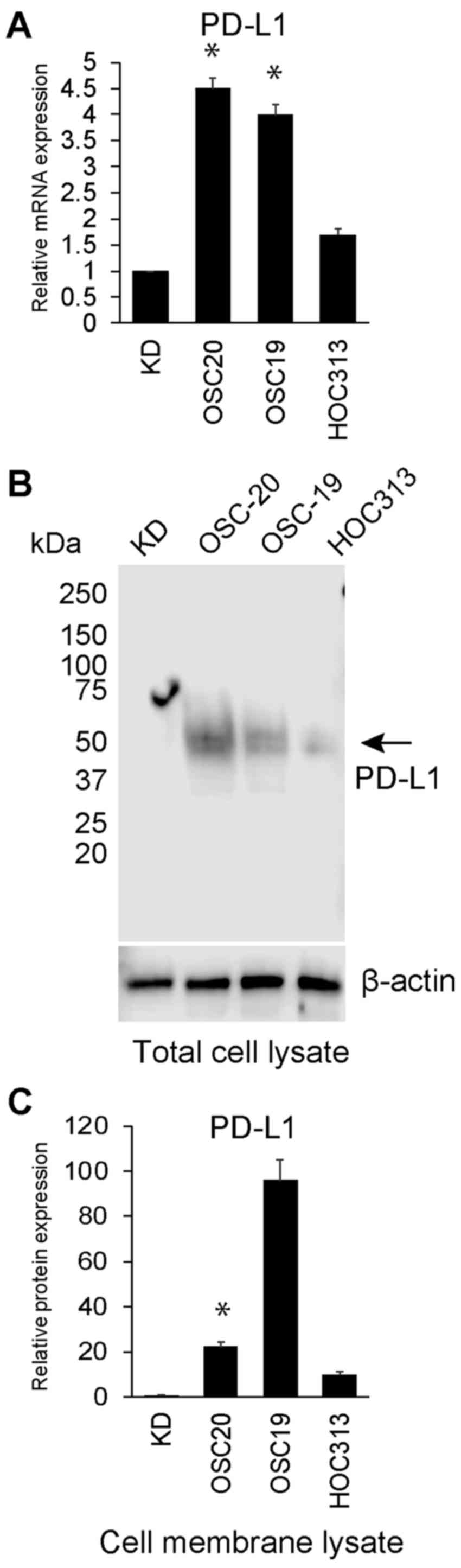
As shown in Fig.
1A, qPCR revealed that PD-L1 mRNA expression was upregulated in
the OSC-20 and OSC-19 cells compared with that in the KD cells.
Among these 3 HNSCC cell lines, the PD-L1 mRNA expression level
ranged from 1.7- to 4.5-fold higher than that in KD cells (Fig. 1A). PD-L1 protein extracted from the
total cell lysate was also upregulated in the OSC-20 and OSC-19
cells compared with the level in the HOC313 and KD cells, as
determined by western blot anaysis (Fig. 1B). However, the amount of PD-L1
protein extracted from the OSC-20 cell membrane (22.3-fold higher
than that in the KD cells) was less than that of PD-L1 protein from
the OSC-19 cell membrane (96.08-fold higher than that in KD cells)
(Fig. 1C). PD-L1 protein in the
culture supernatant from the OSC-20, OSC-19, HOC313 and KD cells
was not detected by ELISA (data not shown).
MMP expression in HNSCC cell lines
(OSC-20, OSC-19 and HOC313)
As shown in Fig.
2A, qPCR was used to determine MMP-1, -2, -3, -7, -8, -9, -10,
-11, -12, -13 and -14 mRNA expression in the OSC-20, OSC-19, HOC313
and KD cells. The expression levels of MMP-7, -10, -12 and -13 in
the OSC-20 cells were 2801.5-, 41.57-, 513.7- and 2522.2-fold
higher than those in the KD cells, respectively. Moreover, the
expression levels of MMP-9, -10 and -13 in the OSC-19 cells were
317.6-, 38.6- and 1380.7-fold higher than those in the KD cells,
respectively. MMP-3 expression in the HOC313 cells was also
7.82-fold higher than that in KD cells. Overall, MMP expression in
the HOC313 cells was not markedly high compared with the levels in
the OSC-20 and OSC-19 cells. Among these MMPs that were upregulated
in these HNSCC cells, the expression of MMP-7, -12 and -13 in
particular was markedly upregulated in the OSC-20 cells compared
with that in the OSC-19 cells. As shown in Fig. 2B, western blot analysis revealed
that MMP-7 and -13 protein expression levels in the culture
supernatant were upregulated in the OSC-20 cells compared with
those in the OSC-19 cells. MMP-12 protein expression was negligible
(Fig. 2B).
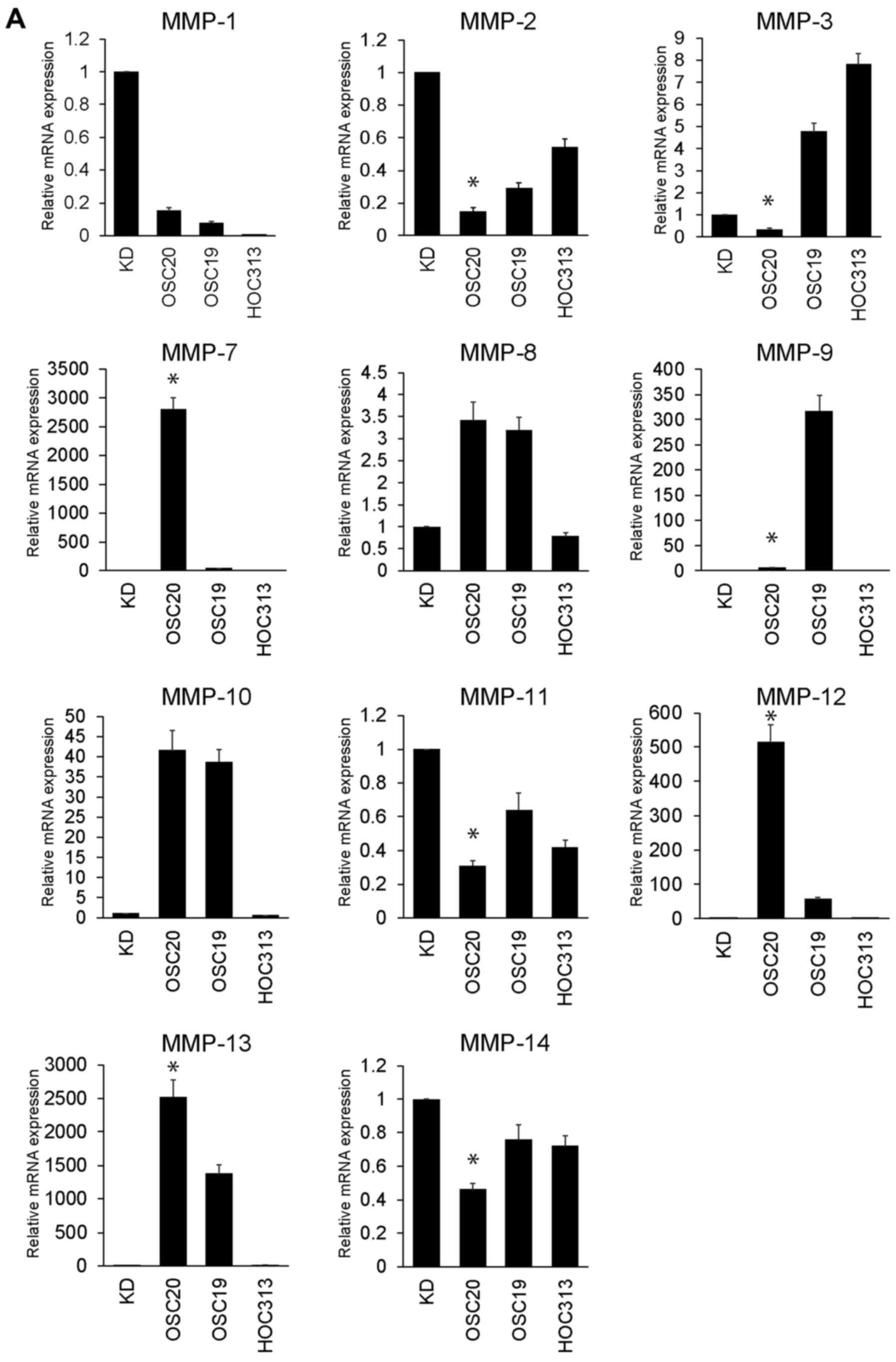 | Figure 2Matrix metalloproteinase (MMP)
expression in head and neck squamous cell carcinoma (HNSCC) cell
lines (OSC-20, OSC-19 and HOC313). (A) Relative mRNA expression
levels of MMP-1, -2, -3, -7, -8, -9, -10, -11, -12, -13 and -14 in
the OSC-20, OSC-19, HOC313 and KD cell lines. Expression levels are
displayed as fold differences relative to control cells (KD cells).
Each data point represents the mean of 3 independent experiments.
The vertical bars show standard deviations. Error bars correspond
to SEM. *P<0.05 vs. OSC-19 cells. (B) MMP-7, -12 and
-13 proteins in the culture supernatant of OSC-20, OSC-19, HOC313
and KD cells were assessed by western blot analysis. The arrows
indicate MMP-7, -12 and -13. β-actin was detected as a loading
control. |
Purification and digestion of PD-L1 by
recombinant MMP-7 and MMP-13
PD-L1 purified from the culture medium of Expi293
cells transfected with the human PD-L1 expression vector contained
a major protein band of Mr 56,000 and a minor band of Mr 46,000
(Fig. 3A), both of which were
recognised by anti-PD-L1 antibody. However, only the major species
of Mr 56,000 was recognised by anti-His antibody. These findings
suggested that the species of Mr 46,000 was generated by C-terminal
truncation of the species of Mr 56,000.
As shown in Fig. 2,
MMP-7 and MMP-13 expression was upregulated in the OSC-20 cells, in
which PD-L1 cell surface degradation was enhanced, and MMP-12
protein expression was negligible (Fig. 2B). Although MMP-12 mRNA was
upregulated in the OSC-20 cells, the absolute amount of mRNA may be
low. These results suggested that MMP-7 and MMP-13 were the
candidates of proteinases involved in PD-L1 degradation.
Recombinant MMP-7 and MMP-13 were activated by incubation with
p-aminophenylmercuric acetate, in accordance with previously
described methods (28,29). PD-L1 was digested for 24 h with
MMP-7 or -13, and the digestion fragments were examined by western
blot analysis. MMP-7 and -13 degraded PD-L1 successfully as the
amounts of PD-L1 species of Mr 56,000 and 46,000 were reduced
(Fig. 3B). The digested fragments
of PD-L1 were not recognised by the anti-PD-L1 antibody.
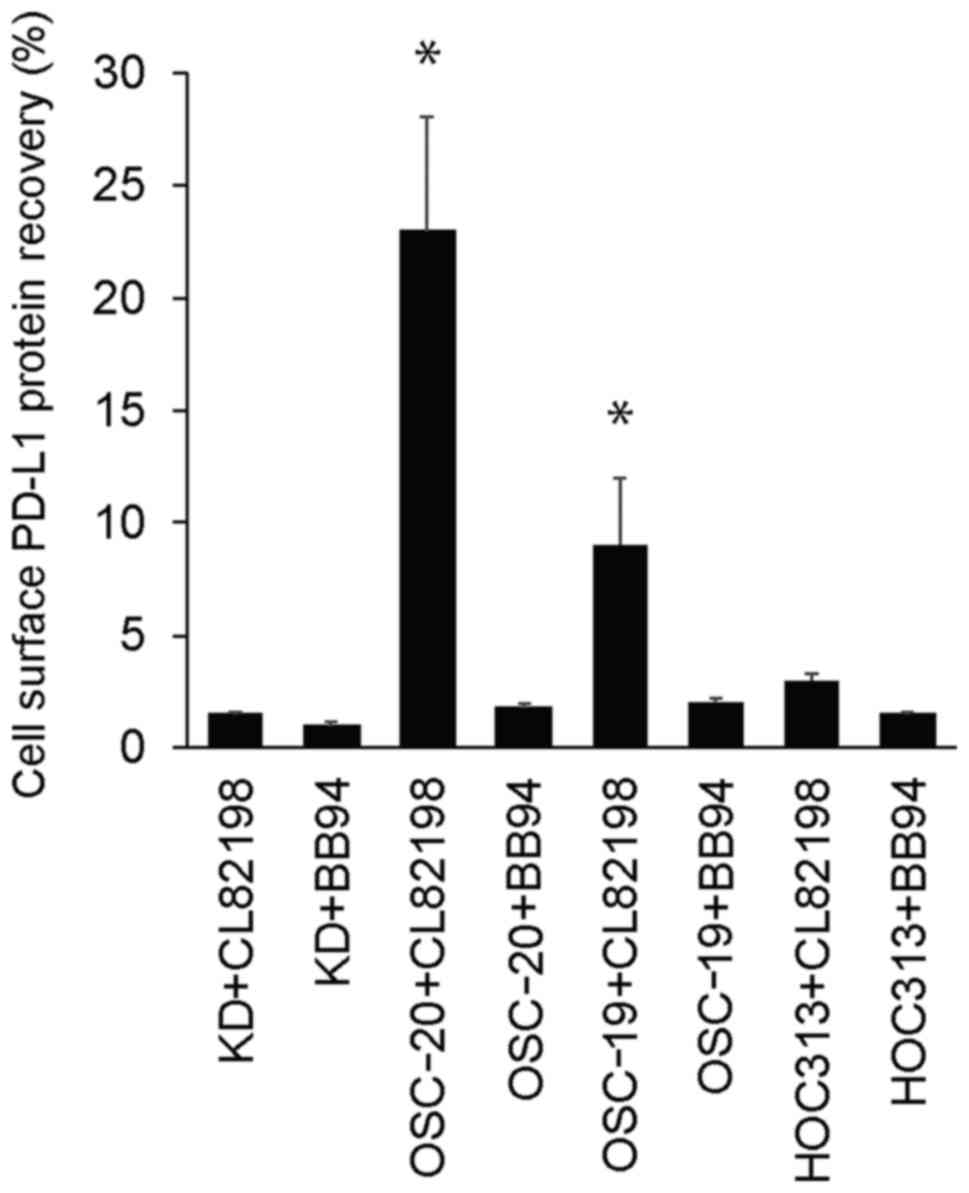
Inhibition of PD-L1 cell surface
degradation in HNSCC cells
To determine which MMP contributes to PD-L1
degradation in HNSCC cells, we used MMP inhibitors with different
specificities. As shown in Fig. 4,
only the MMP-13-specific inhibitor (CL82198) significantly
(P<0.05) restored PD-L1 expression in the OSC-20 cells
(23.2±5.1% recovery from day 1 to 4 in the presence of 10 mg/ml
CL82198) when compared with the BB94 inhibitor. PD-L1 expression
was also significantly (P<0.05) restored by CL82198 in the
OSC-19 cells (9.1±3.2% recovery from day 1 to 4 in the presence of
10 mg/ml CL82198). BB94 did not restore PD-L1 expression in the
HNSCC cells.
Tubulin inhibitor increased MMP-13
expression in R/M HNSCC cells (HOC313 cells) co-cultured with
DCs
The results shown in Fig. 4 suggest that MMP-13 plays a key
role in the shedding/cleavage of PD-L1. We wished to deterimine
which anti-head and neck cancer drug would successfully upregulated
MMP-13 expression. For this purpose, we used tubulin inhibitors
(HAL, PTX and VBL) that are generally used in the treatment of head
and neck cancer. As shown in Fig.
5A–C, following treatment of the OSC-20, OSC-19 and HOC313
cells with tubulin inhibitors, the PD-L1 and MMP13 expression
levels were analysed. MMP-13 expression was significantly
downregulated in the HAL-treated OSC-20 and OSC-19 cells. Moreover,
the MMP-13 and PD-L1 expression levels were significantly
downregulated in the VBL-treated OSC-19 cells. No tubulin inhibitor
used in the OSC-20 and OSC-19 cells increased MMP-13 expression
(Fig. 5A and B). Only treatment
with PTX upregulated MMP-13 expression in the HOC313 cells when
compared with the other tubulin inhibitors used (Fig. 5C). No tubulin inhibitor used to
treat the HOC313 cells altered the expression level of PD-L1. As
shown in Fig. 5D, we examined the
effects of tubulin inhibitor on DCs co-cultured with HOC313 cells.
Under co-culture conditions using DCs and HOC313 cells, treatment
with PTX also upregulated MMP-13 expression when compared with the
other tubulin inhibitors used. Treatment with HAL significantly
upregulated PD-L1 expression in the HOC313 cells co-cultured with
DCs.
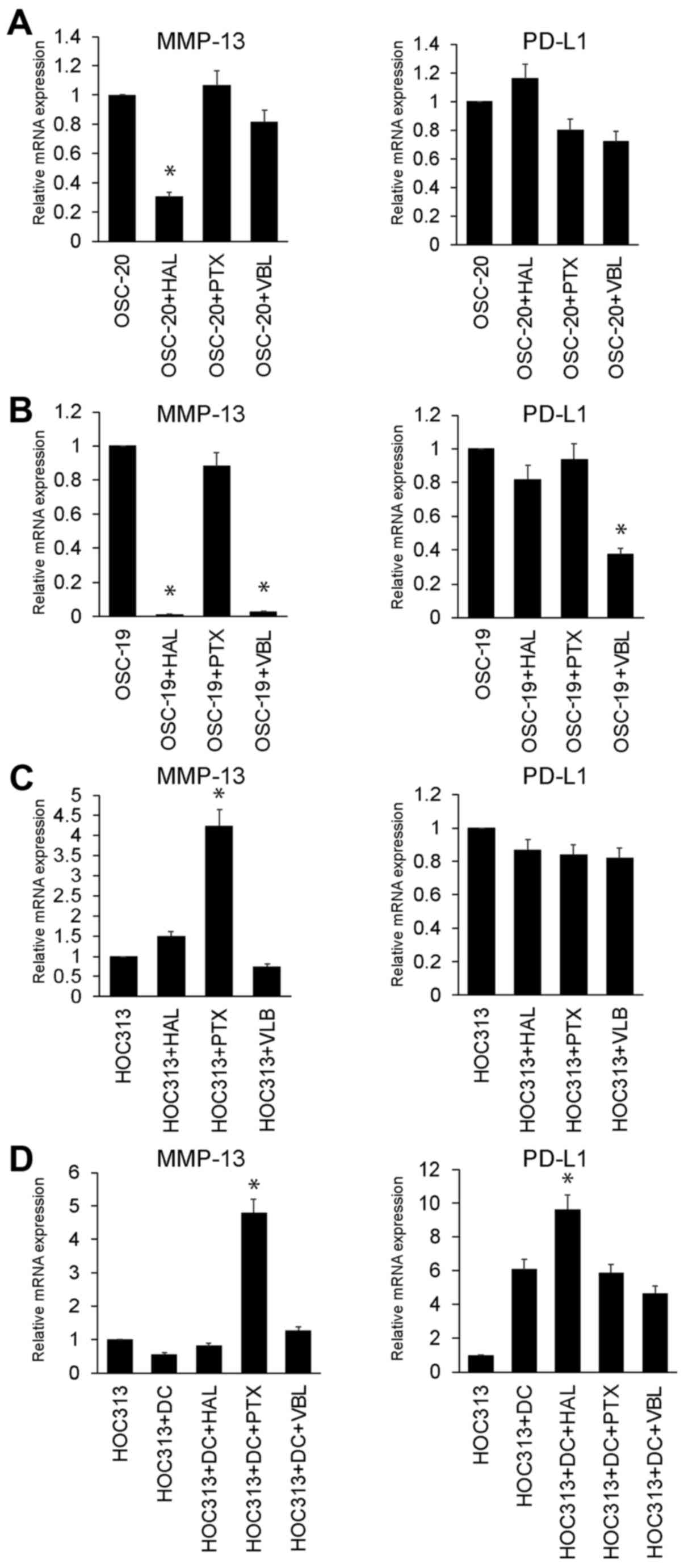 | Figure 5Tubulin inhibitor increases matrix
metalloproteinase-13 (MMP-13) expression on R/M head and neck
squamous cell carcinoma (HNSCC) cells (HOC313 cells) co-cultured
without/with dendritic cells (DCs). Relative mRNA expression levels
of MMP-13 and programmed death-ligand 1 (PD-L1) in the (A) OSC-20,
(B) OSC-19 and (C) HOC313 cell lines treated with tubulin inhibitor
(HAL, eribulin; PTX, paclitaxel; and VBL, vinblastine). Expression
levels are displayed as fold differences relative to the control
cells [vehicle (control buffer only)-treated OSC-20, OSC-19 and
HOC313 cells, respectively]. Each data point represents the mean of
3 independent experiments. The vertical bars show standard
deviations. Error bars correspond to SEM. *P<0.05 vs.
control vehicle-treated OSC-20, OSC-19 and HOC313 cells,
respectively. (D) Relative mRNA expression levels of MMP-13 and
PD-L1 in the DCs co-cultured with HOC313 cell lines treated with
tubulin inhibitor (HAL, eribulin; PTX, paclitaxel; and VBL,
vinblastine). Expression levels are displayed as fold differences
relative to the control cells (control vehicle-treated HOC313
cells). Each data point represents the mean of 3 independent
experiments. The vertical bars show standard deviations. Error bars
correspond to SEM. *P<0.05 vs. control
vehicle-treated DCs co-cultured with HOC313 cells. |
Discussion
PD-L1 is a ligand of the inhibitory receptor, PD-1,
and the PD-1/PD-L1 pathway induces and maintains peripheral
immu-notolerance in cancer tissues (11,30).
PD-L1 expression is constitutive in various types of human cancer,
including squamous cell carcinomas of the lung, oesophagus, and
head and neck, as well as other types of carcinomas of the colon,
ovaries, bladder and breast, and melanoma and glioma (7,12,13,30-38).
Our previous study demonstrated that PD-L1 expression in HNSCC cell
lines with low invasiveness (OSC-20 and OSC-19 cells) was higher
than that in the R/M HNSCC line (TSU cells) (16). These findings contradicted previous
results for other tumour types (7,12,13,30–38).
Although the molecular mechanisms underlying this difference remain
unclear, we hypothesise that they are confined to cancer subtypes
with a unique molecular profile. In this study, we demonstrated
that PD-L1 mRNA and total protein expression was downregulated in
R/M HNSCC (HOC313 cells) and upregulated in cells with low
invasiveness (OSC-20 and OSC-19). These findings are consistent
with our previous results (16).
Of note, in this study, PD-L1 protein extracted from the cell
membrane was downregulated in OSC-20 cells compared with OSC-19
cells, despites higher PD-L1 expression in the total cell lysate of
OSC20 cells compared with that in OSC-19 cells. These findings
suggested that PD-L1 could be degraded on the OSC-20 cell surface
by some proteinases.
A recent study from another group demonstrated that
PD-L1 was cleaved by recombinant MMP-13 and MMP-9, which may limit
the immunosuppressive capacity and may thus contribute to the
exacerbation of inflammation in tissues (17). MMP family proteins are the most
plausible PD-L1 degradation proteinases as they play dual roles in
the pathogenesis of inflammation: Stimulating protective innate
and/or adaptive immune functions, as well as tissue destruction
(21). The results shown in
Fig. 1 suggest that proteinases
that were upregulated in OSC-20 may play a key role in the cell
surface degradation of PD-L1. The results shown in Fig. 2 suggest that several MMPs were
upregulated on HNSCC, and in particular, MMP-7 and -13 were
upregulated in OSC-20 cells rather than in OSC-19 cells. On the
other hand, the OSC-19 cells exhibited a significant upregulation
in MMP-9. Indeed, MMP-9 may play important roles in the invasion
and metastasis of cancer cells (39–41).
However, MMP-9 inhibitors did not exert a significant therapeutic
effect in any of these human clinical trials (20). Recent studies have shown a
protective role of MMP-9 in carcinogenesis, tumour growth or
metastasis, in experiments using MMP-9 KO mice (42,43);
however, the detailed mechanisms behind this protective effect
remain unclear. The results from the present study demonstrated
that PD-L1 shedding in OSC-19 cells was less than that in the
OSC-20 cells. We hypotheised that the shedding/cleavage of PD-L1 by
MMP-9 was not the main mechanism behind the protective effects on
invasion and metastasis. The other mechanisms behind the protective
effects of MMP-9 warrant further investigation. All of these
results suggest that MMP-7 and -13 are strong candidate proteinases
for involvement in the cell surface degradation of PD-L1. As shown
in Fig. 3, the amount of intact
purified PD-L1 was reduced by recombinant MMP-13 and -7 following
incubation for 24 h at 37°C. There were no digested
low-molecular-weight fragments. These results suggest that MMP-13
and MMP-7 have cleavage activity against PD-L1 and PD-L1 digested
at multiple regions may be degraded at the PD-1 binding domain.
However, the issue of which MMP, MMP-13 or MMP-7, is critical for
the cell surface digestion of PD-L1 is remains unclear.
To define which MMP contributes to PD-L1 degradation
in HNSCC cells, we used MMP inhibitors with different
specificities. As shown in Fig. 4,
as the expression of PD-L1 was significantly restored by a specific
inhibitor of MMP-13 (CL82198), we hypothesised that MMP-13 was
involved in the shedding/cleavage of PD-L1 in the OSC-20 cells. The
role of MMP-7 in the shedding/cleavage of PD-L1 may be minor in
vitro as the MMP-7 inhibitor, BB94, could not restore cell
surface PD-L1 expression. MMP-13 may play a protective role in
melanoma cell lung metastasis as the lung metastases of B16BL6
melanoma cells were found to be increased in MMP-13 KO mice after
intravenous injection (44).
MMP-2, MMP-7, MMP-9 and MMP-14 (MT1-MMP) play important roles in
the invasion and metastasis of cancer cells (39–41).
Therefore, the targeted deletion of these MMPs may inhibit cancer
cell proliferation, invasion and metastasis. Indeed, tumourigenesis
and metastasis are decreased in MMP-2 KO (45), MMP-7 KO (46) or MMP-9 KO mice (47,48).
However, contradictory results have also been observed; that is, a
protective role of host MMPs in carcinogenesis, tumour growth or
metastasis, in experiments using MMP-3 KO (49), MMP-7 KO (48), MMP-9 KO (42,43)
and MMP-12 KO mice (50). In
addition, a completely protective role in cancer has been reported
in MMP-8 KO mice, which exhibited enhanced carcinogenesis induced
by chemical carcinogens (51).
Therefore, the data from previous studies suggest that MMP-13 and
some MMPs play dual roles in invasion and metastasis, that is, a
promoting effect and a protective effect. However, in these
previous studies, the detailed mechanisms behind the protective
effects on invasion and metastasis were not fully elucidated. The
results of this study suggest that the shedding/cleavage of PD-L1
by MMP-13 may be one of the mechanisms behind the protective
effects on invasion and metastasis. Further in vivo studies
are required to test and confirm this hypothesis.
As shown in Fig. 5,
only paclitaxel treatment upregulated MMP-13 expression in HOC313
cells compared with the other tubulin inhibitors tested. No tubulin
inhibitor used to treat the OSC-20 and OSC-19 cells increased
MMP-13 expression. In a previous study, we determined the in
vitro anti-proliferative activities in eribulin-treated OSC-20,
OSC-19 and HOC313 cells (52). Of
note, the HOC313 cells were highly sensitive to eribulin compared
with other cell lines. Previously, another group of researchers
demonstrated that the reduced expression levels of 4 tubulins
(TUBA1C, TUBA4A, TUBB3 and TUBB6) was significantly associated with
eribulin sensitivity and that the expression of one tubulin
(TUBB2A) was significantly associated with paclitaxel sensitivity
(53). The tubulin inhibitors are
broadly classified into microtubule-stabilizing (such as
paclitaxel) and microtubule-destabilizing (such as vinblastine and
eribulin) drugs (54). SMAD
proteins, which are essential mediators of the TGF-β signalling
pathway, normally bind microtubules (55). SMAD signalling plays an important
role in promoting the invasive phenotype of human HNSCC cells by
upregulating their MMP-13 expression (56). These data suggest that the
expression profiles of tubulins may differ among the OSC-20, OSC-19
and HOC313 cells, and that tubulin inhibitors may stabilise or
destabilise the association between SMAD proteins and microtubules
in OSC-20, OSC-19 and HOC313 cells, consequently resulting in the
differential induction of MMP13 expression. However, we cannot
exclude the possibility that paclitaxel may target other molecules
to upregulate MMP-13 expression.
Although anti-PD-1 treatment can produce durable
responses, it appears to benefit only a subset of patients. If
anti-PD-1 therapy alone were not so effective, it would be
appropriate to combine this approach with another therapeutic
option' to aid readability. However, at present, no candidate drugs
for such a combination therapy are known. In this study, we
demonstrated that MMP-13 may play a critical role in the
shedding/cleavage of PD-L1. In addition, among the anticancer drugs
generally used in the treatment of head and neck cancer patients,
paclitaxel increased MMP-13 expression in R/M HNSCC cells (HOC313
cells) co-cultured without/with DCs. These results suggested that
paclitaxel is a strong candidate for use in combination with
anti-PD-1 therapy. Clearly, further in vivo studies are
required to test this hypothesis.
Acknowledgments
We are grateful to the members of the Department of
Oral and Maxillofacial Surgery of Kanazawa University for their
helpful suggestions and assistance. We would also like to thank
Enago (www.enago.jp) for assisting with
language editing. This study was supported by Grants-in-Aid for
Scientific Research from the Ministry of Education, Science, Sports
and Culture, Japan (#17K11870 to H.K.; #17K11869 to H.N.; #15H05042
to S.K.). The funders had no role in the study design, data
collection and analysis, decision to publish, or preparation of the
manuscript.
Abbreviations:
|
DCs
|
dendritic cells
|
|
HAL
|
eribulin mesylate (also known as
Halaven)
|
|
HNSCC
|
head and neck squamous cell
carcinoma
|
|
PTX
|
paclitaxel
|
|
MMP
|
matrix metalloproteinase
|
|
PD-1
|
programmed death receptor-1
|
|
PD-L1
|
programmed death-ligand 1
|
|
PBMCs
|
peripheral blood mononuclear cells
|
|
qPCR
|
quantitative (real-time) polymerase
chain reaction
|
|
R/M HNSCC
|
recurrent and/or metastatic head and
neck squamous cell carcinoma
|
|
VBL
|
vinblastine
|
References
|
1
|
Topalian SL, Hodi FS, Brahmer JR,
Gettinger SN, Smith DC, McDermott DF, Powderly JD, Carvajal RD,
Sosman JA, Atkins MB, et al: Safety, activity, and immune
correlates of anti-PD-1 antibody in cancer. N Engl J Med.
366:2443–2454. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Brahmer JR, Tykodi SS, Chow LQ, Hwu WJ,
Topalian SL, Hwu P, Drake CG, Camacho LH, Kauh J, Odunsi K, et al:
Safety and activity of anti-PD-L1 antibody in patients with
advanced cancer. N Engl J Med. 366:2455–2465. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Topalian SL, Drake CG and Pardoll DM:
Targeting the PD-1/B7-H1(PD-L1) pathway to activate anti-tumor
immunity. Curr Opin Immunol. 24:207–212. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Freeman GJ, Long AJ, Iwai Y, Bourque K,
Chernova T, Nishimura H, Fitz LJ, Malenkovich N, Okazaki T, Byrne
MC, et al: Engagement of the PD-1 immunoinhibitory receptor by a
novel B7 family member leads to negative regulation of lymphocyte
activation. J Exp Med. 192:1027–1034. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Oliveira-Neto HH, Gleber-Netto FO, de
Sousa SF, França CM, Aguiar MC, Silva TA and Batista AC: A
comparative study of microvessel density in squamous cell carcinoma
of the oral cavity and lip. Oral Surg Oral Med Oral Pathol Oral
Radiol. 113:391–398. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liang X, Zhou H, Liu X, He Y, Tang Y, Zhu
G, Zheng M and Yang J: Effect of local hyperthermia on
lymphangiogenic factors VEGF-C and -D in a nude mouse xenograft
model of tongue squamous cell carcinoma. Oral Oncol. 46:111–115.
2010. View Article : Google Scholar
|
|
7
|
Strome SE, Dong H, Tamura H, Voss SG,
Flies DB, Tamada K, Salomao D, Cheville J, Hirano F, Lin W, et al:
B7-H1 blockade augments adoptive T-cell immunotherapy for squamous
cell carcinoma. Cancer Res. 63:6501–6505. 2003.PubMed/NCBI
|
|
8
|
Lyford-Pike S, Peng S, Young GD, Taube JM,
Westra WH, Akpeng B, Bruno TC, Richmon JD, Wang H, Bishop JA, et
al: Evidence for a role of the PD-1:PD-L1 pathway in immune
resistance of HPV-associated head and neck squamous cell carcinoma.
Cancer Res. 73:1733–1741. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zandberg DP and Strome SE: The role of the
PD-L1:PD-1 pathway in squamous cell carcinoma of the head and neck.
Oral Oncol. 50:627–632. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Butte MJ, Keir ME, Phamduy TB, Sharpe AH
and Freeman GJ: Programmed death-1 ligand 1 interacts specifically
with the B7-1 costimulatory molecule to inhibit T cell responses.
Immunity. 27:111–122. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Keir ME, Butte MJ, Freeman GJ and Sharpe
AH: PD-1 and its ligands in tolerance and immunity. Annu Rev
Immunol. 26:677–704. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dong H, Strome SE, Salomao DR, Tamura H,
Hirano F, Flies DB, Roche PC, Lu J, Zhu G, Tamada K, et al:
Tumor-associated B7-H1 promotes T-cell apoptosis: A potential
mechanism of immune evasion. Nat Med. 8:793–800. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tsushima F, Tanaka K, Otsuki N, Youngnak
P, Iwai H, Omura K and Azuma M: Predominant expression of B7-H1 and
its immunoregulatory roles in oral squamous cell carcinoma. Oral
Oncol. 42:268–274. 2006. View Article : Google Scholar
|
|
14
|
Iwai Y, Terawaki S and Honjo T: PD-1
blockade inhibits hema-togenous spread of poorly immunogenic tumor
cells by enhanced recruitment of effector T cells. Int Immunol.
17:133–144. 2005. View Article : Google Scholar
|
|
15
|
Hirano F, Kaneko K, Tamura H, Dong H, Wang
S, Ichikawa M, Rietz C, Flies DB, Lau JS, Zhu G, et al: Blockade of
B7-H1 and PD-1 by monoclonal antibodies potentiates cancer
therapeutic immunity. Cancer Res. 65:1089–1096. 2005.PubMed/NCBI
|
|
16
|
Hirai M, Kitahara H, Kobayashi Y, Kato K,
Bou-Gharios G, Nakamura H and Kawashiri S: Regulation of PD-L1
expression in a high-grade invasive human oral squamous cell
carcinoma microenvironment. Int J Oncol. 50:41–48. 2017. View Article : Google Scholar
|
|
17
|
Dezutter-Dambuyant C, Durand I, Alberti L,
Bendriss-Vermare N, Valladeau-Guilemond J, Duc A, Magron A, Morel
AP, Sisirak V, Rodriguez C, et al: A novel regulation of PD-1
ligands on mesenchymal stromal cells through MMP-mediated
proteolytic cleavage. Oncoimmunology. 5:e10911462015. View Article : Google Scholar
|
|
18
|
Egeblad M and Werb Z: New functions for
the matrix metal-loproteinases in cancer progression. Nat Rev
Cancer. 2:161–174. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Overall CM and López-Otín C: Strategies
for MMP inhibition in cancer: Innovations for the post-trial era.
Nat Rev Cancer. 2:657–672. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Milner JM and Cawston TE: Matrix
metalloproteinase knockout studies and the potential use of matrix
metalloproteinase inhibitors in the rheumatic diseases. Curr Drug
Targets Inflamm Allergy. 4:363–375. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Le NT, Xue M, Castelnoble LA and Jackson
CJ: The dual personalities of matrix metalloproteinases in
inflammation. Front Biosci. 12:1475–1487. 2007. View Article : Google Scholar
|
|
22
|
Yokoi T, Hirata S, Nishimura F, Miyakawa
A, Odajima T and Kohama G: Some properties of a newly established
human cell line derived from an oral squamous carcinoma. Tumor Res.
25:93-91–93. 1990.
|
|
23
|
Yokoi T, Homma H and Odajima T:
Establishment and characterization of OSC-19 cell line in serum and
protein free culture. Tumor Res. 24:1–17. 1988.
|
|
24
|
Ishisaki A, Oida S, Momose F, Amagasa T,
Rikimaru K, Ichijo H and Sasaki S: Identification and
characterization of autocrine-motility-factor-like activity in oral
squamous-cell-carcinoma cells. Int J Cancer. 59:783–788. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Arrighi JF, Hauser C, Chapuis B, Zubler RH
and Kindler V: Long-term culture of human CD34(+) progenitors with
FLT3-ligand, thrombopoietin, and stem cell factor induces extensive
amplification of a CD34(−)CD14(−) and a CD34(−) CD14(+) dendritic
cell precursor. Blood. 93:2244–2252. 1999.PubMed/NCBI
|
|
26
|
Yang D, Chen Q, Le Y, Wang JM and
Oppenheim JJ: Differential regulation of formyl peptide
receptor-like 1 expression during the differentiation of monocytes
to dendritic cells and macrophages. J Immunol. 166:4092–4098. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
28
|
Nakamura H, Fujii Y, Ohuchi E, Yamamoto E
and Okada Y: Activation of the precursor of human stromelysin 2 and
its interactions with other matrix metalloproteinases. Eur J
Biochem. 253:67–75. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nakamura H, Fujii Y, Inoki I, Sugimoto K,
Tanzawa K, Matsuki H, Miura R, Yamaguchi Y and Okada Y: Brevican is
degraded by matrix metalloproteinases and aggrecanase-1 (ADAMTS4)
at different sites. J Biol Chem. 275:38885–38890. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zou W and Chen L: Inhibitory B7-family
molecules in the tumour microenvironment. Nat Rev Immunol.
8:467–477. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wintterle S, Schreiner B, Mitsdoerffer M,
Schneider D, Chen L, Meyermann R, Weller M and Wiendl H: Expression
of the B7-related molecule B7-H1 by glioma cells: A potential
mechanism of immune paralysis. Cancer Res. 63:7462–7467.
2003.PubMed/NCBI
|
|
32
|
Konishi J, Yamazaki K, Azuma M, Kinoshita
I, Dosaka-Akita H and Nishimura M: B7-H1 expression on non-small
cell lung cancer cells and its relationship with tumor-infiltrating
lymphocytes and their PD-1 expression. Clin Cancer Res.
10:5094–5100. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ohigashi Y, Sho M, Yamada Y, Tsurui Y,
Hamada K, Ikeda N, Mizuno T, Yoriki R, Kashizuka H, Yane K, et al:
Clinical significance of programmed death-1 ligand-1 and programmed
death-1 ligand-2 expression in human esophageal cancer. Clin Cancer
Res. 11:2947–2953. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Thompson RH and Kwon ED: Significance of
B7-H1 overexpression in kidney cancer. Clin Genitourin Cancer.
5:206–211. 2006. View Article : Google Scholar
|
|
35
|
Hamanishi J, Mandai M, Iwasaki M, Okazaki
T, Tanaka Y, Yamaguchi K, Higuchi T, Yagi H, Takakura K, Minato N,
et al: Programmed cell death 1 ligand 1 and tumor-infiltrating
CD8+ T lymphocytes are prognostic factors of human
ovarian cancer. Proc Natl Acad Sci USA. 104:3360–3365. 2007.
View Article : Google Scholar
|
|
36
|
Nomi T, Sho M, Akahori T, Hamada K, Kubo
A, Kanehiro H, Nakamura S, Enomoto K, Yagita H, Azuma M, et al:
Clinical significance and therapeutic potential of the programmed
death-1 ligand/programmed death-1 pathway in human pancreatic
cancer. Clin Cancer Res. 13:2151–2157. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ghebeh H, Tulbah A, Mohammed S, Elkum N,
Bin Amer SM, Al-Tweigeri T and Dermime S: Expression of B7-H1 in
breast cancer patients is strongly associated with high
proliferative Ki-67-expressing tumor cells. Int J Cancer.
121:751–758. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yao Y, Tao R, Wang X, Wang Y, Mao Y and
Zhou LF: B7-H1 is correlated with malignancy-grade gliomas but is
not expressed exclusively on tumor stem-like cells. Neuro Oncol.
11:757–766. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Liotta LA and Kohn EC: The
microenvironment of the tumour-host interface. Nature. 411:375–379.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Shiomi T and Okada Y: MT1-MMP and MMP-7 in
invasion and metastasis of human cancers. Cancer Metastasis Rev.
22:145–152. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Deryugina EI and Quigley JP: Matrix
metalloproteinases and tumor metastasis. Cancer Metastasis Rev.
25:9–34. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Cornelius LA, Nehring LC, Harding E,
Bolanowski M, Welgus HG, Kobayashi DK, Pierce RA and Shapiro SD:
Matrix metalloproteinases generate angiostatin: Effects on
neovascularization. J Immunol. 161:6845–6852. 1998.PubMed/NCBI
|
|
43
|
Hamano Y, Zeisberg M, Sugimoto H, Lively
JC, Maeshima Y, Yang C, Hynes RO, Werb Z, Sudhakar A and Kalluri R:
Physiological levels of tumstatin, a fragment of collagen IV alpha3
chain, are generated by MMP-9 proteolysis and suppress angiogenesis
via alphaV beta3 integrin. Cancer Cell. 3:589–601. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Fukuda H, Mochizuki S, Abe H, Okano HJ,
Hara-Miyauchi C, Okano H, Yamaguchi N, Nakayama M, D'Armiento J and
Okada Y: Host-derived MMP-13 exhibits a protective role in lung
metastasis of melanoma cells by local endostatin production. Br J
Cancer. 105:1615–1624. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Itoh T, Tanioka M, Yoshida H, Yoshioka T,
Nishimoto H and Itohara S: Reduced angiogenesis and tumor
progression in gela-tinase A-deficient mice. Cancer Res.
58:1048–1051. 1998.PubMed/NCBI
|
|
46
|
Wilson CL, Heppner KJ, Labosky PA, Hogan
BL and Matrisian LM: Intestinal tumorigenesis is suppressed in mice
lacking the metalloproteinase matrilysin. Proc Natl Acad Sci USA.
94:1402–1407. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Itoh T, Tanioka M, Matsuda H, Nishimoto H,
Yoshioka T, Suzuki R and Uehira M: Experimental metastasis is
suppressed in MMP-9-deficient mice. Clin Exp Metastasis.
17:177–181. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Acuff HB, Carter KJ, Fingleton B, Gorden
DL and Matrisian LM: Matrix metalloproteinase-9 from bone
marrow-derived cells contributes to survival but not growth of
tumor cells in the lung microenvironment. Cancer Res. 66:259–266.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
McCawley LJ, Crawford HC, King LE Jr,
Mudgett J and Matrisian LM: A protective role for matrix
metalloproteinase-3 in squamous cell carcinoma. Cancer Res.
64:6965–6972. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Acuff HB, Sinnamon M, Fingleton B, Boone
B, Levy SE, Chen X, Pozzi A, Carbone DP, Schwartz DR, Moin K, et
al: Analysis of host- and tumor-derived proteinases using a custom
dual species microarray reveals a protective role for stromal
matrix metal-loproteinase-12 in non-small cell lung cancer. Cancer
Res. 66:7968–7975. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Balbín M, Fueyo A, Tester AM, Pendás AM,
Pitiot AS, Astudillo A, Overall CM, Shapiro SD and López-Otín C:
Loss of collagenase-2 confers increased skin tumor susceptibility
to male mice. Nat Genet. 35:252–257. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Kitahara H, Hirai M, Kato K, Bou-Gharios
G, Nakamura H and Kawashiri S: Eribulin sensitizes oral squamous
cell carcinoma cells to cetuximab via induction of
mesenchymal-to-epithelial transition. Oncol Rep. 36:3139–3144.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Dezső Z, Oestreicher J, Weaver A, Santiago
S, Agoulnik S, Chow J, Oda Y and Funahashi Y: Gene expression
profiling reveals epithelial mesenchymal transition (EMT) genes can
selectively differentiate eribulin sensitive breast cancer cells.
PLoS One. 9:e1061312014. View Article : Google Scholar
|
|
54
|
Kavallaris M: Microtubules and resistance
to tubulin-binding agents. Nat Rev Cancer. 10:194–204. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Dong C, Li Z, Alvarez R Jr, Feng XH and
Goldschmidt-Clermont PJ: Microtubule binding to Smads may regulate
TGF-beta activity. Mol Cell. 5:27–34. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Leivonen SK, Ala-Aho R, Koli K, Grénman R,
Peltonen J and Kähäri VM: Activation of Smad signaling enhances
collagenase-3 (MMP-13) expression and invasion of head and neck
squamous carcinoma cells. Oncogene. 25:2588–2600. 2006. View Article : Google Scholar : PubMed/NCBI
|