Introduction
Cholangiocarcinoma (CCA) is a bile-duct tumor that
is rare in the majority of countries (1,2), but
which has a far greater incidence in the Greater Mekong sub-region
of southeast Asia, particularly in northeastern Thailand, where the
prevalence of the small human liver fluke Opisthorchis
viverrini is high (3). O.
viverrini is classified as a group I carcinogen by the
International Agency for Research on Cancer (4). Infection with this fluke causes
chronic inflammation, leading to periductal fibrosis and ultimately
contributing to the development of CCA (5,6). In
northeastern Thailand, CCA is the major primary liver cancer, with
its management costing US$120 million annually (5,7).
Clinical presentation is typically observed at a late stage and
therefore the majority of patients with CCA cannot be cured by
surgical resection (8,9). Furthermore, the disease is able to
develop resistance to standard chemotherapeutic drugs over time
(10). Therefore, using
phytochemicals with anti-inflammatory and anticancer activities to
treat cancer or to enhance the efficacy of other chemotherapeutic
drugs, may be an alternative approach for the management of CCA and
help to avoid drug-resistance (11–13).
Anthocyanins are the water-soluble flavonoids
responsible for the blue, purple and red colors in a number of
fruits, flowers and leaves (14).
Anthocyanins exhibit anti-inflammatory, anti-angiogenesis,
antioxidant and anti-proliferative effects, and thus have a number
of medical applications (15–18).
Among these are prevention and treatment of cancer (19,20).
Previous studies have demonstrated that the consumption of
anthocyanin-rich foods is associated with a decreased risk of
chronic diseases, including cardiovascular disease, arthritis and
diabetes mellitus, and development of esophageal, colon, lung and
skin cancers (17,21).
Cyanidin and delphinidin glycosides are the most
abundant and well-studied anthocyanins with potential anticancer
activity (22–24). However, a major obstacle in the use
of anthocyanins is that they are unstable and prone to degradation
(25). Previously, cyanidin and
delphinidin were isolated from cobs of purple waxy corn (Zea
mays L. var. ceritina Kulesh) and petals of blue
butterfly pea (Clitoria ternatea L.) and were manipulated to
form a complex with turmeric extract and other trace elements
(26) to increase their stability
and activity. This novel anthocyanin complex (AC) nanoparticle
exhibited anti-inflammatory and anti-fibrotic effects in an O.
viverrini-infected hamster model, establishing its potential
for chemoprevention of CCA (26).
However, the utility for CCA treatment has not yet been
investigated.
The aim of the present study was to demonstrate the
application of AC for CCA treatment. Anticancer activities of AC
and the potential underlying molecular mechanisms against CCA were
investigated in vitro using a CCA cell line. The effect of
combined treatment of AC and gemcitabine against the
gemcitabine-resistant CCA cell line (KKU214GemR) was
also investigated. The results of the present study provide an
insight into the promising use of AC phytochemical products for the
treatment of CCA.
Materials and methods
Chemicals and reagents
AC was prepared as described previously (26). In brief, aqueous extracts of purple
waxy corn cobs and blue butterfly pea petals were mixed with
turmeric (Curcuma longa) extract (7:2:1) in the presence of
100 mM caffeic acid and piperine (Sigma Aldrich; Merck KGaA,
Darmstadt, Germany) and 2 mM zinc sulfate (Ajax Finechem; Thermo
Fisher Scientific, Inc., Waltham, MA, USA). Thereafter, the mixture
was cooled and dried to yield of AC. Only one batch of AC was used
throughout the present study to avoid batch-to-batch variation.
Dulbecco's modified Eagle's medium (DMEM), penicillin/streptomycin,
trypsin-EDTA and fetal bovine serum (FBS) were purchased from
Gibco; Thermo Fisher Scientific, Inc. 4-(2-Aminoethyl)
benzenesulfonyl fluoride hydrochloride (AEBSF), dimethyl sulfoxide
(DMSO), sulforhodamine B (SRB) and the broad-spectrum caspase
inhibitor quinolone-Val-Asp-difluorophenoxymethyl ketone (Q-VD-OPh)
were purchased from Sigma-Aldrich; Merck KGaA. Rabbit anti-protein
kinase RNA-like endoplasmic reticulum (ER) kinase (PERK; cat. no.
5683), anti-p65 (cat. no. 8242), anti-activating transcription
factor 4 (ATF4; cat. no. 11815), anti-phosphorylated eukaryotic
initiation factor 2α (p-eIF2α; cat. no. 3398) (Ser51),
anti-eIF2α (cat. no. 9722), anti-poly(ADP-ribose) polymerase (PARP;
cat. no. 9542), anti-B-cell lymphoma-2 (Bcl-2; cat. no. 2872),
anti-caspase-3 (cat. no. 9662) and anti-β-tubulin (cat. no. 2128)
and radioimmunoprecipitation assay (RIPA) buffer were purchased
from Cell Signaling Technology, Inc. (Danvers, MA, USA). Rabbit
anti-forkhead box M1 (FOXM1; cat. no. sc-502) (C-20) was obtained
from Santa Cruz Biotechnology, Inc. (Dallas, TX, USA). Horseradish
peroxidase (HRP)-conjugated goat anti-rabbit IgG (cat. no.
111-035-003) secondary antibody was purchased from Jackson
ImmunoResearch, Inc. (West Grove, PA, USA). A dead-cell apoptosis
kit [Annexin V/propidium iodide (PI)], Pierce™ bicinchoninic acid
(BCA) protein assay kit, Hank's balanced salt solution (HBSS) and
MitoSOX™ Red mitochondrial superoxide indicator were obtained from
Thermo Fisher Scientific, Inc. ECL™ Prime western blotting
detection reagent and polyvinylidene difluoride (PVDF) membrane
were obtained from GE Healthcare (Chicago, IL, USA).
Human CCA cell lines
The KKU213 CCA cell line was established from Thai
CCA patients as described previously (27). The gemcitabine-resistant CCA cell
line KKU214GemR was established previously by exposure
to stepwise increases in the concentration of gemcitabine as
described previously (28). It
should be noted that the parental line of KKU214GemR
cells appears to be a KKU213 cell derivative (web.expasy.org/cellosaurus/CVCL_M264).
However, due to the fact that KKU214GemR cells were
induced to be a gemcitabine-resistant CCA cell line, it would not
have any bearing on the results of the present study. The cell
lines were maintained in DMEM supplemented with 10% FBS, 100 U/ml
penicillin and 100 µg/ml streptomycin at 37°C in a
humidified incubator containing 10% CO2.
KKU214GemR cells were cultured in the presence of
gemcitabine to maintain its resistant phenotype, but were cultured
in a drug-free medium for one passage prior to performing
experiments.
Assessment of half-maximal inhibitory
concentration (IC50) of AC
The IC50 was assessed using the SRB
assay. For instance, KKU213 cells were seeded at 2,000 cells/well
in flat-bottomed 96-well plates (Corning Inc., Corning, NY, USA).
The following day, the cells were incubated with either DMSO
(diluent control) or various concentrations of AC (100–800
µg/ml) dissolved in DMSO for 12, 24, 36 and 48 h at 37°C in
a humidified incubator containing 10% CO2. Cells were
fixed with ice-cold 40% trichloroacetic acid at 4°C for 1 h.
Following washing three times with running tap water, 0.4% (w/v)
SRB solution in 1% acetic acid was added and incubated further for
1 h at room temperature. Excess SRB solution was removed by washing
with 1% acetic acid and SRB was dissolved by adding 10 mM Tris
buffer. Absorbance at 492 nm was determined using an ELISA plate
reader (Tecan Group Ltd., Männedorf, Switzerland). The absorbance
at 492 nm of DMSO-treated cells was used as control. For
KKU214GemR cells, the same procedure was performed, but
the cells were treated with either single agent (300 µg/ml
AC; 20 or 40 µM gemcitabine) or a combination of AC (300
µg/ml) and gemcitabine (20 or 40 µM) for 24, 48 and
72 h.
Assessment of cellular apoptosis
Cellular apoptosis was determined by Annexin V/PI
staining according to the manufacturer's protocol. In brief,
following trypsinization and washing with sterile PBS,
~106 KKU213 cells were resuspended in 1X binding buffer.
Annexin V/PI solution was added prior to incubation at room
temperature in the dark for 15 min. Stained cells were detected
using a BD FACSCanto II flow cytometer and analyzed using BD
FACSDiva software (version 6.1.3) (both from BD Biosciences, San
Jose, CA, USA).
Plasmid transfection
The pCMV4-p65 plasmid was a gift from Professor
Warner Greene, Gladstone Institute of Virology and Immunology, San
Francisco, CA, USA (Addgene plasmid #21966). In total
~2×106 KKU213 cells were seeded into 10-cm cell culture
dishes for 24 h, prior to transfection with pCMV4-p65 plasmid using
X-tremeGENE HP (Roche Diagnostics, Basel, Switzerland), according
to the manufacturer's protocol. Cells were then harvested at 24 h
post-transfection for further experiments.
Clonogenic assay
CCA cells were seeded into 6-well plates at a
density of 1,000 cells/well overnight prior to drug treatment.
Cells were treated with different concentrations of AC (100–800
µg/ml) and incubated for 48 h at 37°C in a humidified
incubator containing 10% CO2. In the case of
KKU214GemR cells, cells were treated with either a
single agent (300 µg/ml AC; 20 or 40 µM gemcitabine)
or a combination of AC (300 µg/ml) and gemcitabine (20 or 40
µM). The culture medium was changed every 2 days and cells
were cultured for a further 14 days. Finally, cells were fixed with
4% paraformaldehyde and stained with 0.5% crystal violet. Stained
cells were dissolved with 33% acetic acid and absorbance at 620 nm
was measured using an ELISA reader (Tecan Group Ltd.).
Measurement of mitochondrial superoxide
production
Mitochondrial superoxide production was determined
using MitoSOX Red mitochondrial superoxide indicator, according to
the manufacturer's protocol. In brief, CCA cells were trypsinized,
washed once and resuspended in HBSS with
Ca2+/Mg2+. Subsequently, 4 µM MitoSOX
Red solution was added and the mixture was incubated at 37°C for 30
min in the dark. CCA cells were centrifuged at 600 × g for 5 min at
room temperature and washed with 1 ml
HBSS/Ca2+/Mg2+. Finally, CCA cells were
resuspended in HBSS/Ca2+/Mg2+ and the
fluorescence intensity at 575 nm was measured using a flow
cytometer (BD Biosciences).
Western blot analysis
Protein was extracted from CCA cells using RIPA
buffer (50 mM Tris/HCl, 150 mM NaCl, 1% Nonidet P-40, 0.5% sodium
deoxycholate, 0.1% SDS and 1 mM AEBSF) and the protein
concentration was determined using the BCA assay. Subsequently, 20
µg protein was separated by SDS-PAGE (7 or 12% gels) and
then transferred onto a PVDF membrane. Following blocking with 5%
bovine serum albumin in Tris-buffered saline containing 0.05%
Tween-20, membranes were incubated with primary antibodies against
PERK, p65, ATF4, p-eIF2α (Ser51), eIF2α, PARP,
caspase-3, Bcl-2 and β-tubulin (all 1:1,000) overnight at 4°C.
Following washing, membranes were incubated with secondary antibody
(1:3,000) and the chemiluminescent reaction was developed using
ECL™ Prime blotting detection reagent. Immunoreactivity bands were
captured using the ImageQuant™ LAS4000 mini imager (GE
Healthcare).
Statistical analysis
Data are expressed as the mean ± standard deviation.
Student's t-test and analysis of variance and Tukey's test were
performed to determine differences among experimental groups using
SPSS (version 13.0; SPSS, Inc., Chicago, IL, USA). P<0.05 was
considered to indicate a statistically significant difference.
Non-linear regression analysis was performed to determine the
IC50 values using GraphPad Prism (version 6; GraphPad
Software, Inc., La Jolla, CA, USA).
Results
AC treatment inhibits proliferation and
induces caspaseindependent apoptosis of the CCA cell line
Inhibition of tumor growth and induction of cellular
apoptosis are important modes of action of the majority of
phytochemical agents. To determine whether AC exerts
proliferation-inhibitory effects on CCA cells, and to determine
relevant IC50 values, KKU213 cells were treated with
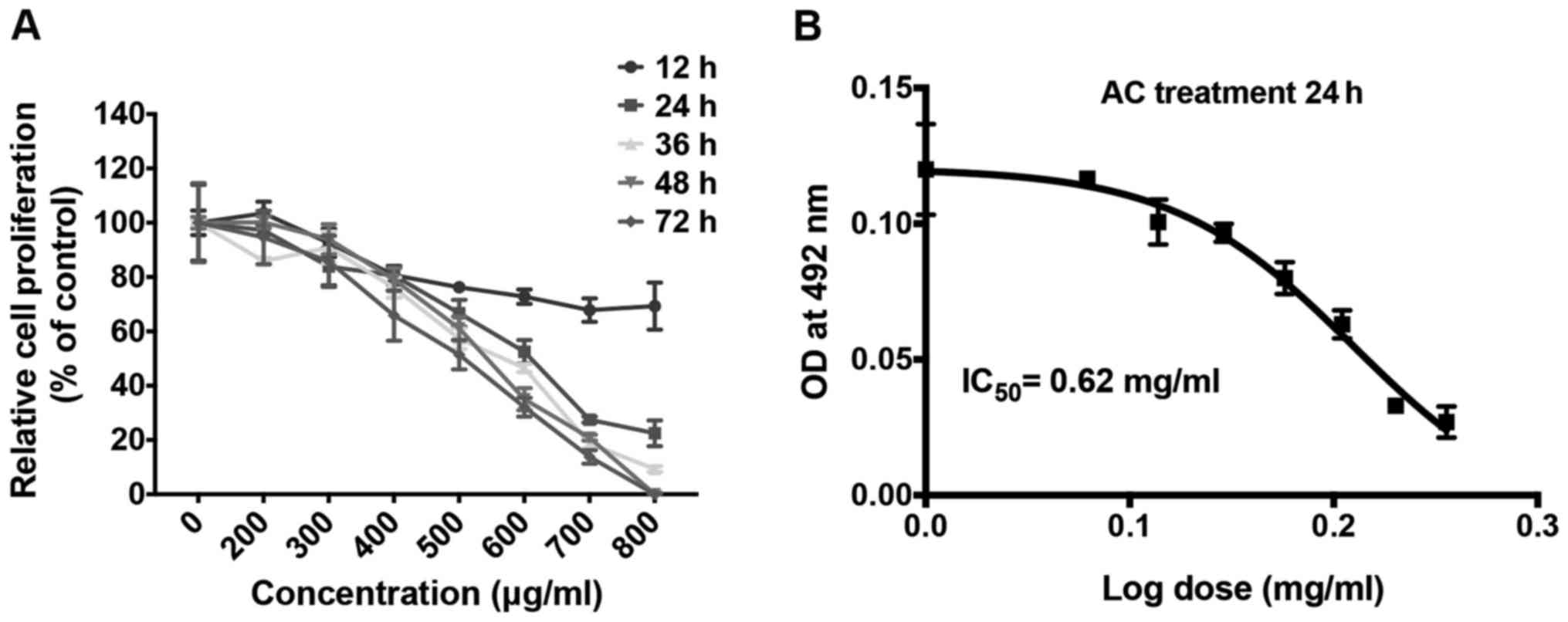
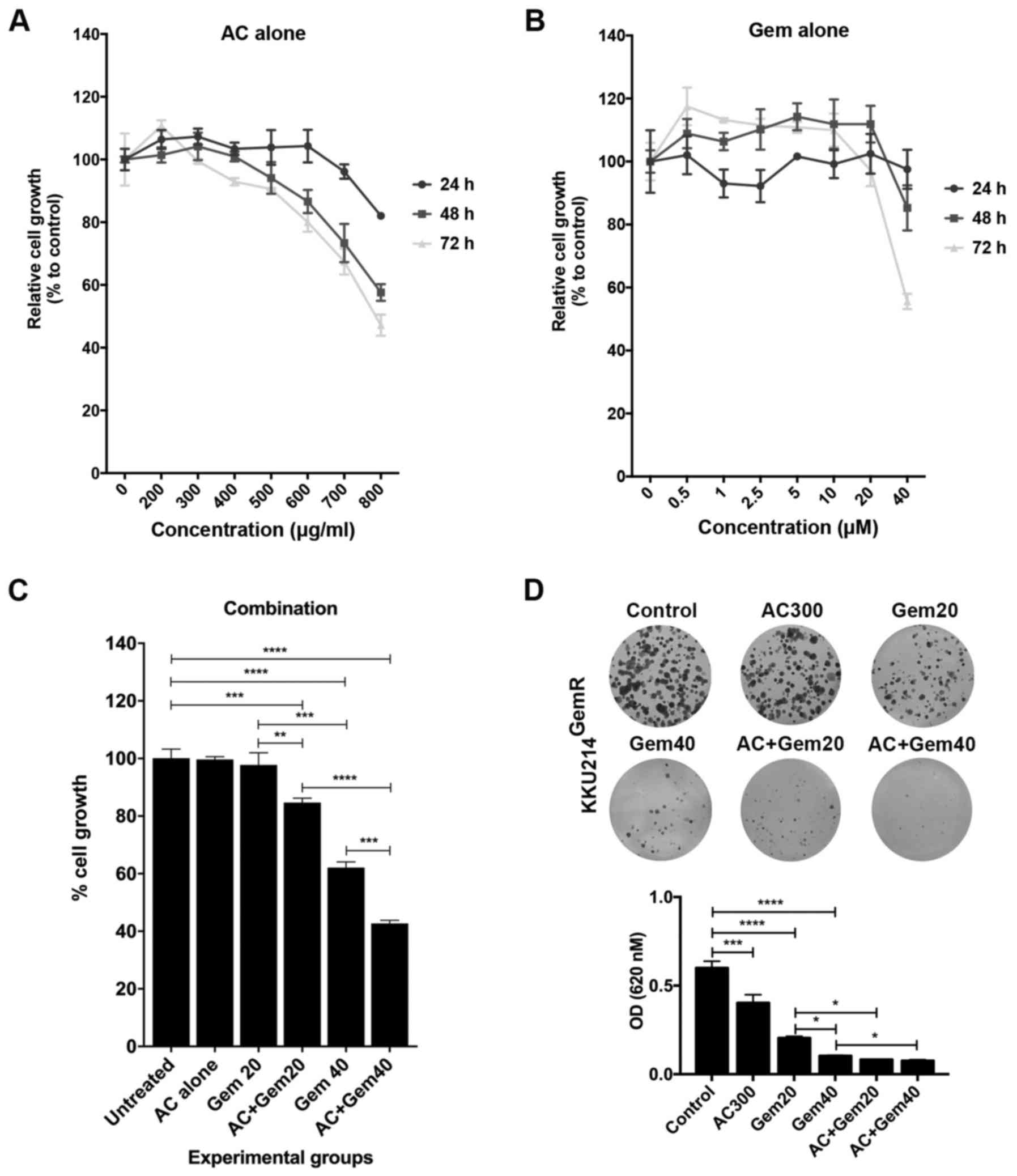
various concentrations of AC. The SRB assay revealed that AC
inhibited cell proliferation in this cell line in a dose- and
time-dependent manner (Fig. 1A).
The proliferation of KKU213 cells was completely inhibited by
treatment with 800 µg/ml AC after 48 h. Non-linear
regression analysis identified that the IC50 value of AC
for KKU213 was 620 µg/ml (Fig.
1B).
It was subsequently investigated whether suppression
of CCA cell proliferation by AC treatment is associated with
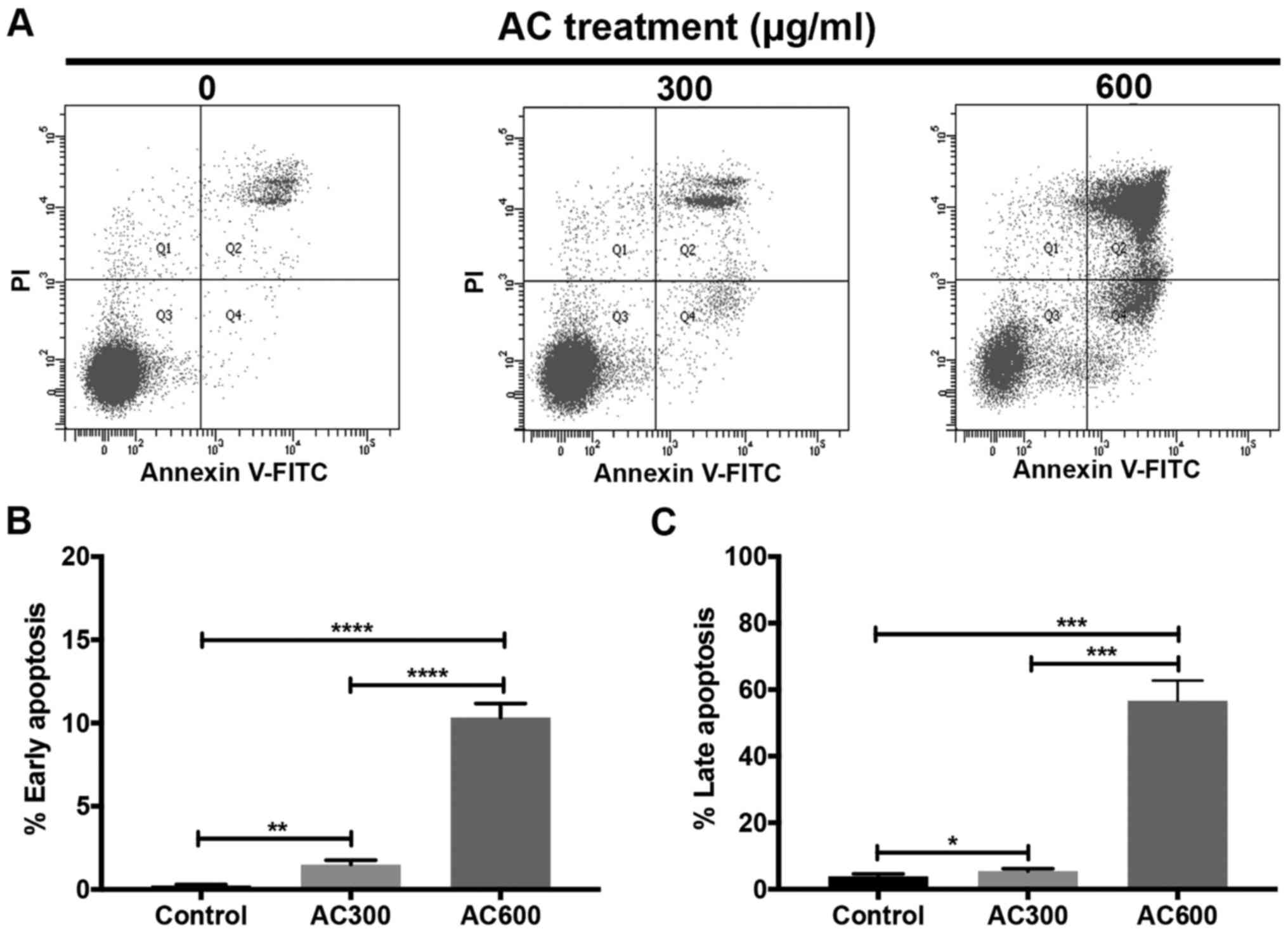
induction of apoptosis. KKU213 cells were treated with 300 or 600
µg/ml AC for 24 h, and cellular apoptosis was investigated
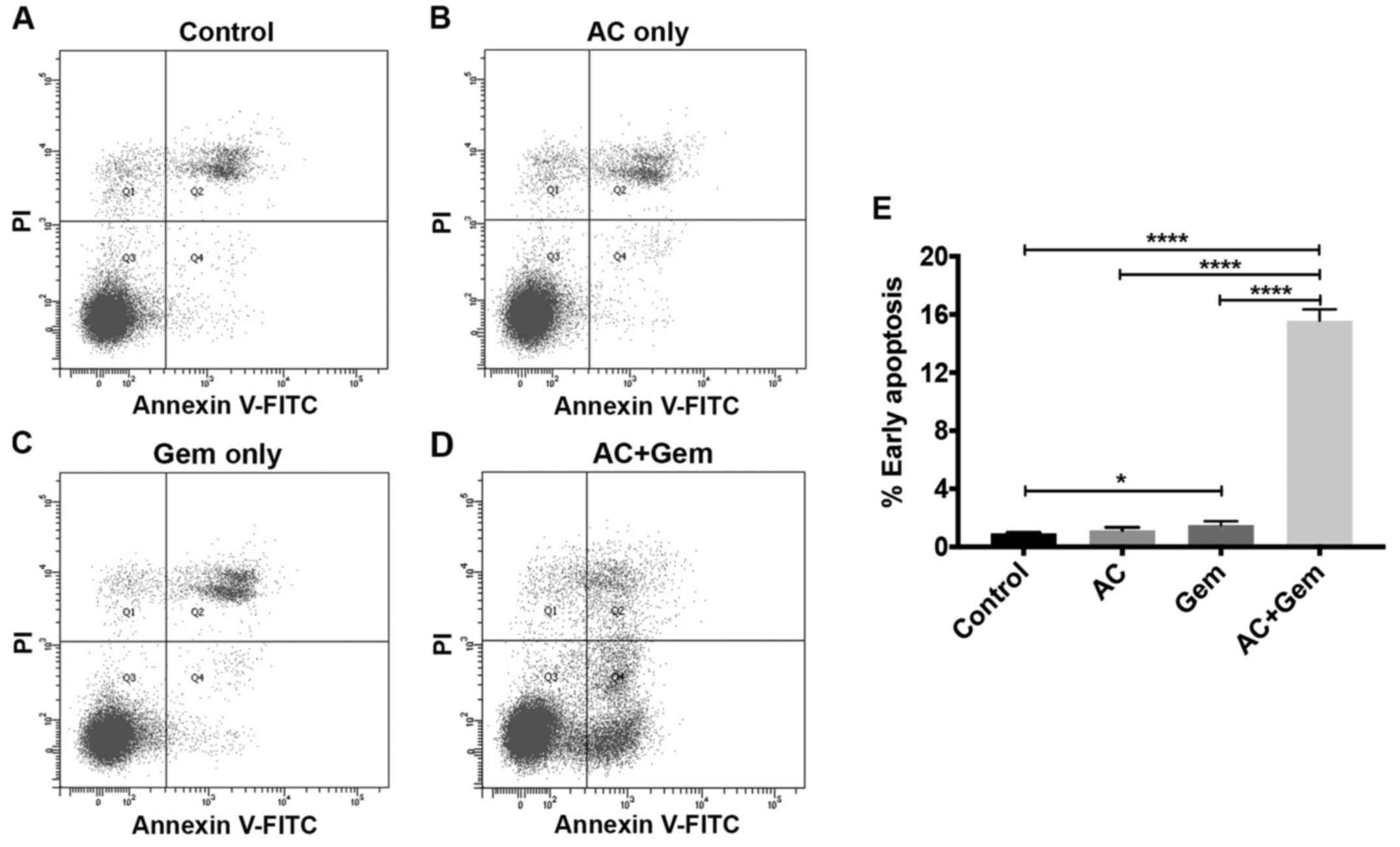
using flow cytometry (Fig. 2A).
The results revealed that early (1.50±0.27% for AC300 and
10.33±0.83% for AC600) (Fig. 2B)
and late (5.50±0.66% for AC300 and 56.70±6.05% for AC600) (Fig. 2C) apoptosis was significantly
induced dose-dependently in AC-treated groups compared with the
control group. These results indicated that AC exhibited
cytotoxicity against CCA cells through the induction of cellular
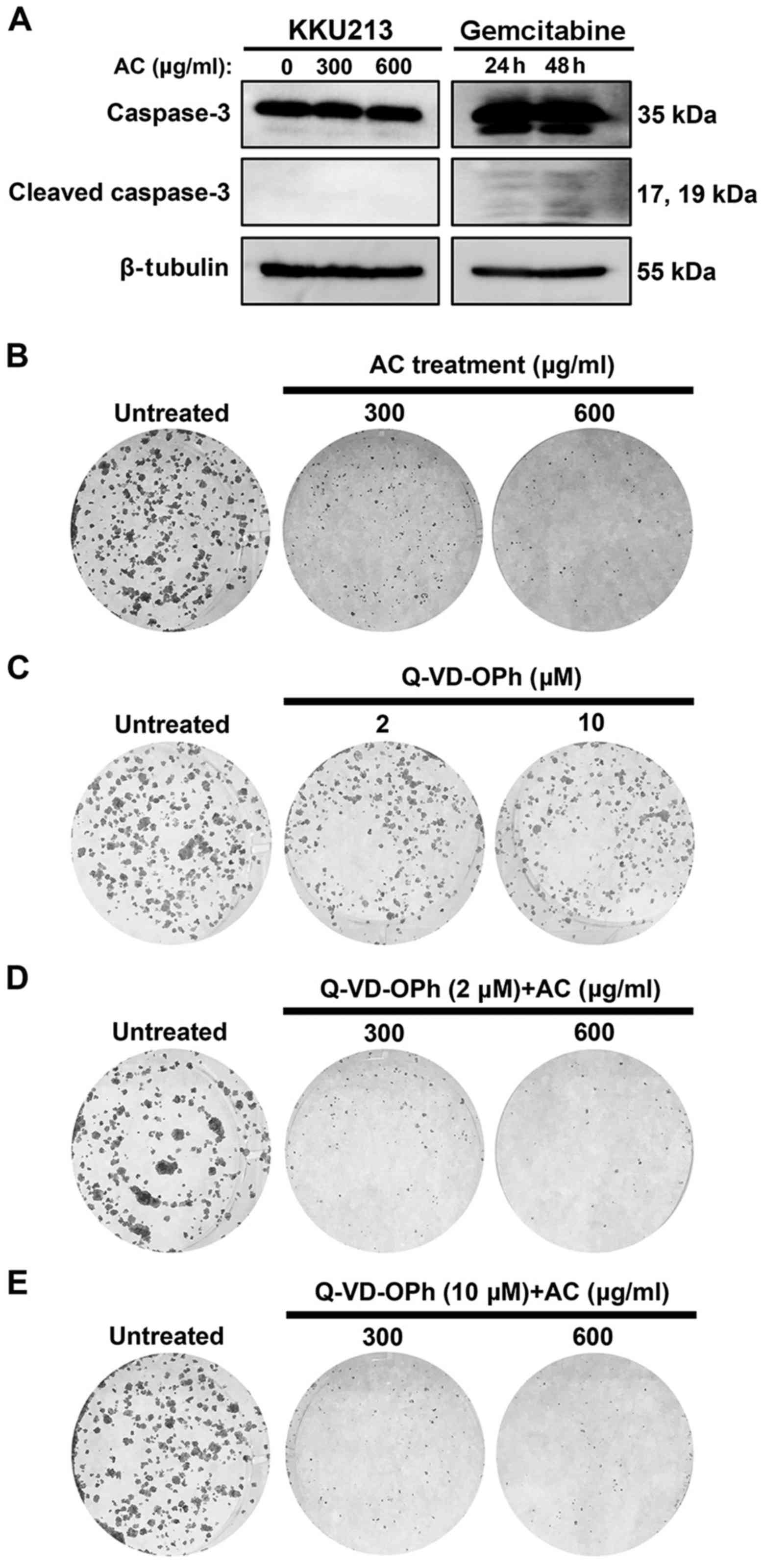
apoptosis. Levels of cleaved caspase-3, a protein marker for
cellular apoptosis, was also assayed using western blotting.
Cleaved caspase-3 was not observed in AC-treated KKU213 cells, but
was detected in a gemcitabine-treated group (Fig. 3A). In addition, a pan-caspase
inhibitor (Q-VD-OPh) was combined with AC treatment to test whether
cell death was due to a caspase-independent pathway. A clonogenic
assay demonstrated that treatment with 2 and 10 µM Q-VD-OPh,
identified to prevent caspase activation (29), did not affect cell viability of the
KKU213 cells (Fig. 3C). Notably,
Q-VD-OPh treatment was not able to prevent cell death when combined
with AC treatment (Fig. 3D and E),
indicating that caspase-independent cell death was occurring in
AC-treated CCA cells.
AC treatment inhibits colony formation of
the CCA cell line
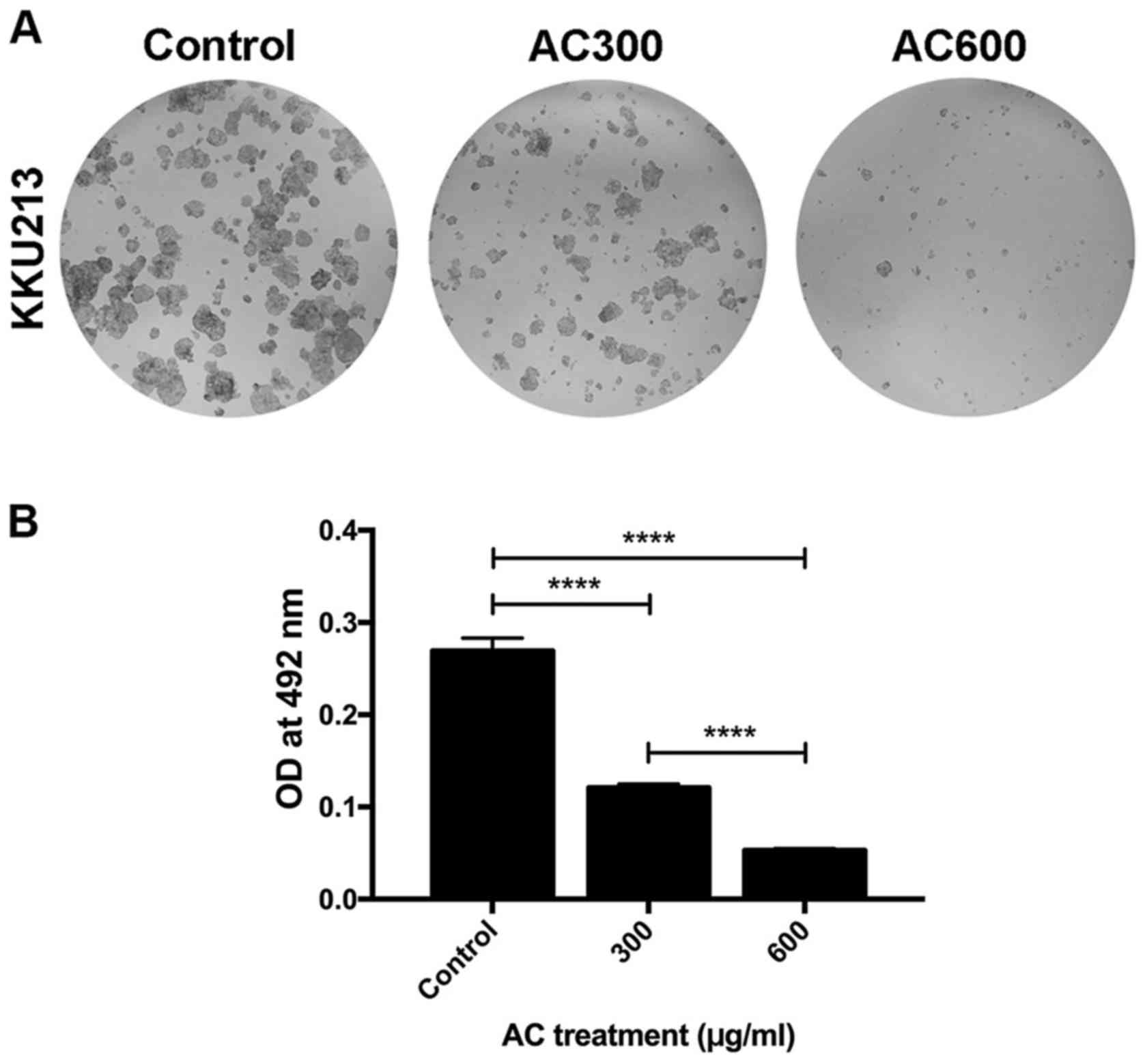
The clonogenic assay is the method of choice to
determine cell reproductive viability (ability of cells to produce
progeny or ability of a single cell to form a colony) following
treatment with radiation or a cytotoxic agent (30,31).
This approach was employed to determine the cytotoxic effects of AC
treatment on a CCA cell line (Fig.
4A). Consistent with the results of the SRB assay, the presence
of AC significantly inhibited KKU213 colony formation in a
dose-dependent manner compared with the control group (P<0.0001)
(Fig. 4B). This result indicated
that the AC-mediated decrease in reproductive viability is one of
the underlying molecular mechanisms of AC against CCA.
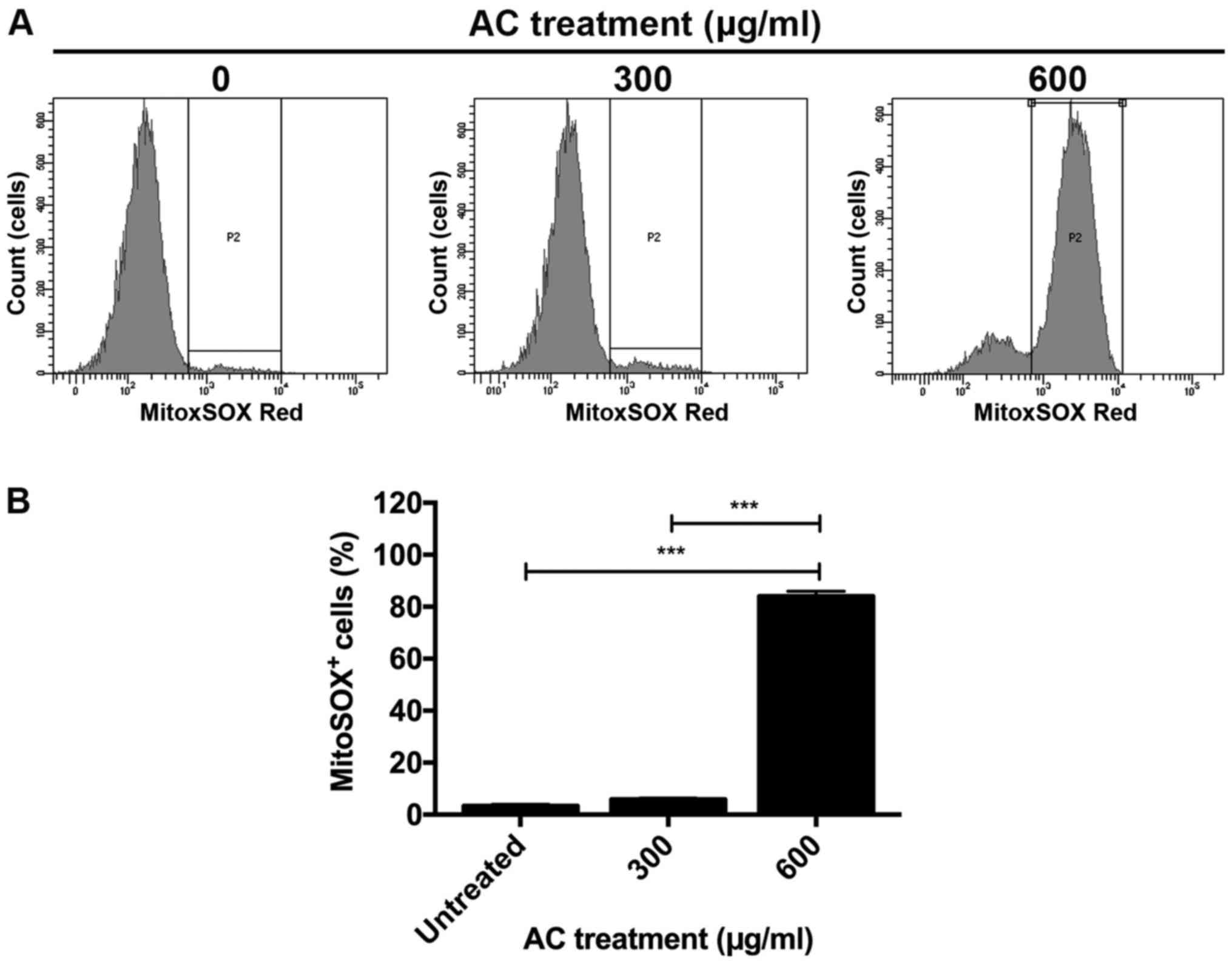
AC treatment induces mitochondrial
superoxide production partly via suppression of Bcl-2
expression
Induction of superoxide production in mitochondria
is an important event in the induction of apoptosis (32). To investigate whether induction of
mitochondrial superoxide production is involved in AC-induced
apoptosis of CCA cells, KKU213 cells were treated with AC, and
levels of mitochondria-specific superoxide were determined using
flow cytometry. Minimal superoxide production was detected in the
control group; however, production was slightly increased in KKU213
cells treated with 300 µg/ml AC (Fig. 5). Treatment with 600 µg/ml
AC significantly induced superoxide production in the mitochondria
of KKU213 cells relative to the control group (P<0.001)
(Fig. 5).
Since superoxide production may be inhibited by the
anti-apoptotic Bcl-2 protein (32), the expression of Bcl-2 protein was
investigated further in KKU213 CCA cells. Western blot analysis
revealed that the expression of Bcl-2 protein mirrored superoxide
production. Expression of Bcl-2 protein decreased in the AC-treated
group, particularly when 600 µg/ml AC was used, relative to
the DMSO-treated control (Fig. 6).
These results indicated that AC induced superoxide
production-mediated apoptosis partly via the inhibition of Bcl-2
protein expression.
AC treatment targets pro-survival and
endoplasmic reticulum stress (ER stress) response of the CCA cell
line
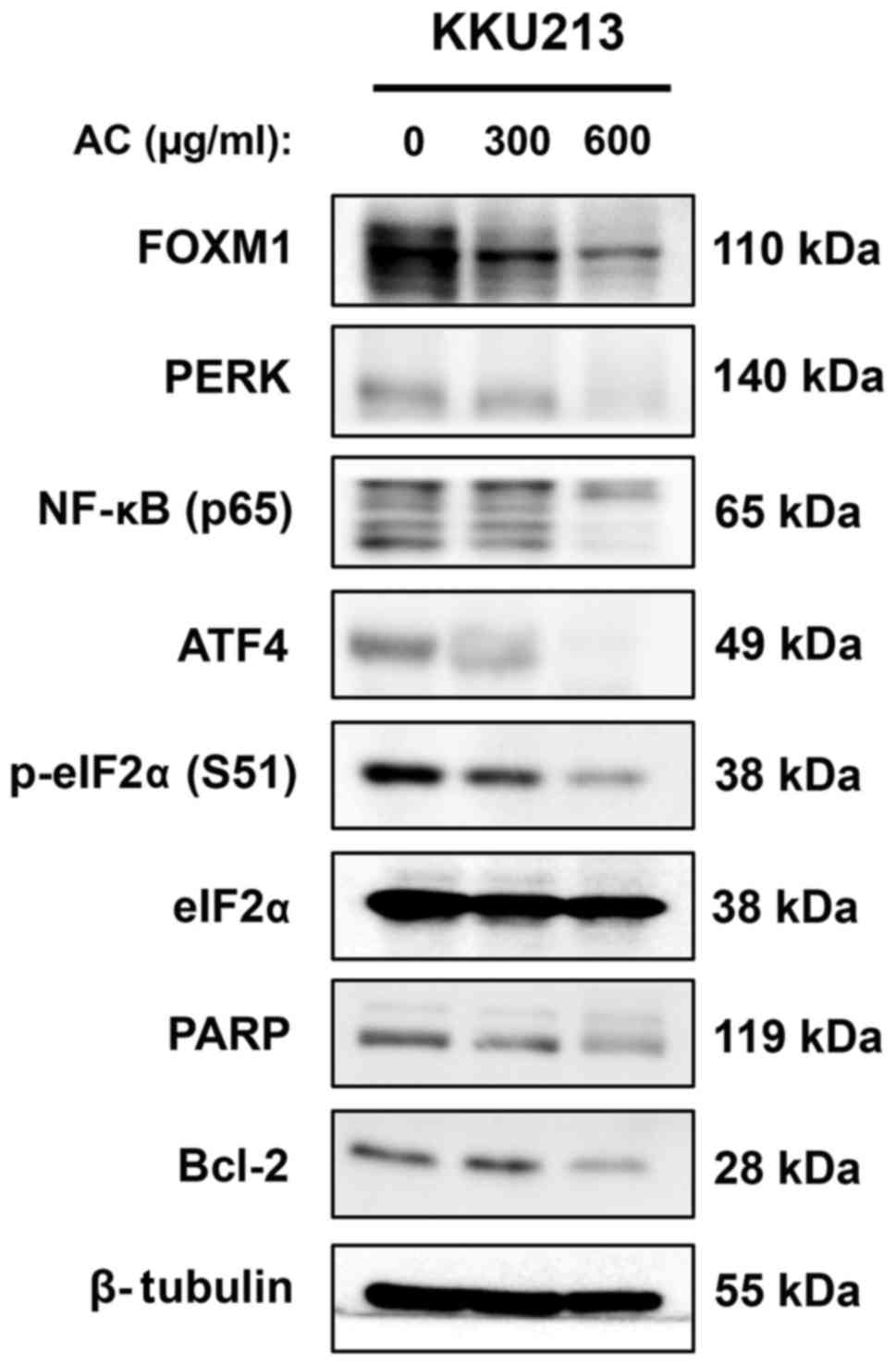
FOXM1 and NF-κB are well-known oncogenic proteins
involved in the survival of cancer cells (33,34).
The expression of these proteins was determined in the CCA cell
line following treatment with AC for 24 h. Western blot analysis
revealed that expression of FOXM1 and the p65 subunit of NF-κB
decreased in a dose-dependent manner in KKU213 cells treated with
AC (Fig. 6). Notably, expression
of these proteins was almost completely inhibited in KKU213 cells
treated with 600 µg/ml AC compared with the DMSO-treated
control (Fig. 6). Additionally,
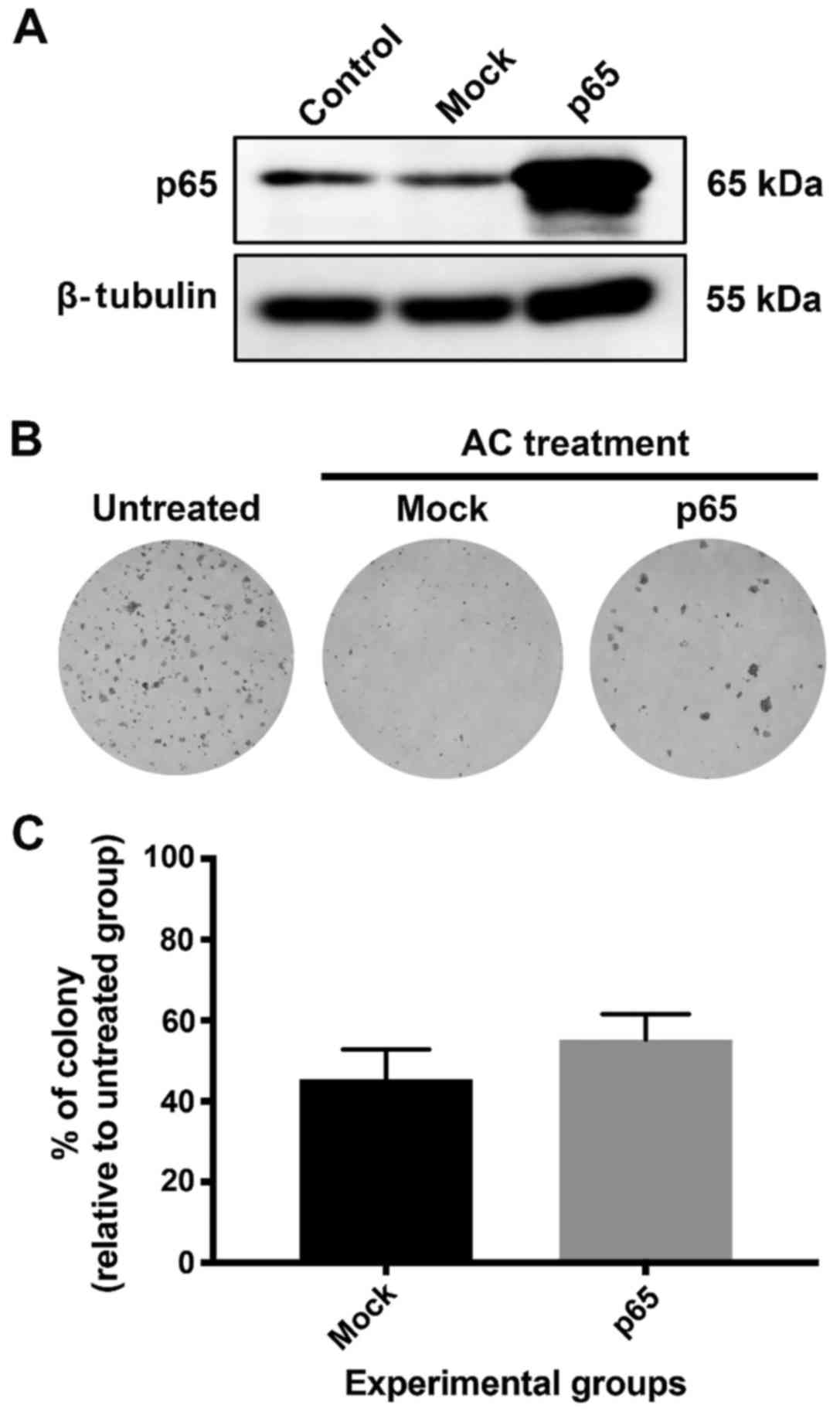
KKU213 cells were transfected with p65 plasmid DNA and the
influence of p65 induction on AC treatment of KKU213 cells was
determined using a clonogenic assay. The p65 subunit of NF-κB was
successfully induced in KKU213 as identified using western blot
analysis (Fig. 7A). However, the
clonogenic assay revealed that p65 overexpression did not affect
the viability of KKU213 cells compared with non-transfected and
mock-transfected controls (Fig. 7B and
C). This implies that AC is a potent inducer of apoptosis
against CCA cells, and that induction of a pro-survival
transcription factor was not able to protect CCA cells from cell
death following AC treatment. A previous study has identified that
AC treatment dramatically induced mitochondrial superoxide
production; excessive superoxide production may cause protein
misfolding and ultimately induce ER stress (35). Therefore, the expression of
proteins in the PERK/eIF2α/ATF4 axis, which is an important ER
stress-response pathway, was investigated. Western blot analysis
revealed that expression of PERK, p-eIF2α (Ser51) and
ATF4 in KKU213 cells decreased following AC treatment (Fig. 6). Notably, total eIF2α expression
was not affected by AC treatment, suggesting that AC suppresses
phosphorylation of the eIF2α protein. In addition to FOXM1, p65 and
the PERK/eIF2α/ATF4 axis, it was identified that PARP expression
was also suppressed by AC treatment, particularly at the highest
dose used (Fig. 6).
AC treatment increases gemcitabine
sensitivity of the gemcitabine-resistant KKU214GemR CCA
cell line
Resistance to chemotherapeutic treatment is an
important obstacle for the treatment of various types of cancer,
including CCA (36). Therefore,
whether co-treatment with AC was able to enhance the effect of
gemcitabine was investigated. The SRB assay revealed that the
IC50 of gemcitabine against the KKU214GemR
CCA cell line was 32.11 µM at 72 h, whereas the
IC50 of gemcitabine against the parental KKU214 cell
line was 0.40 µM at 72 h (data not shown), agreeing with a
previous study (28). Treatment of
KKU214GemR cells with AC inhibited cell proliferation in
a dose- and time-dependent manner (Fig. 8A). For gemcitabine treatment alone,
KKU214GemR cell proliferation was markedly inhibited by
treatment with 40 µM for 72 h (Fig. 8B). AC at a dose of 300
µg/ml, which did not exert a proliferation-inhibitory effect
on KKU214GemR cells, was selected for co-treatment with
20 or 40 µM gemcitabine. As expected, the two combinations
significantly enhanced gemcitabine-mediated proliferation
inhibition compared with treatments with single agents (P<0.01
for 20 µM gemcitabine vs. 300 µg/ml AC+20 µM
gemcitabine; P<0.001 for 40 µM gemcitabine vs. 300
µg/ml AC+40 µM gemcitabine) (Fig. 8C). Furthermore, the clonogenic
assay also identified a significant enhancement of gemcitabine
treatment when co-treated with AC (P<0.05) (Fig. 8D). Annexin V/PI staining coupled
with flow cytometry was used to detect apoptotic cells following
combination treatment of KKU214GemR cells.
Administration of AC (300 µg/ml) and gemcitabine (40
µM) significantly induced early apoptosis of these cells
compared with single treatments and the control group (P<0.0001)
(Fig. 9). These results indicated
that AC treatment significantly enhanced the efficacy of
gemcitabine against the gemcitabine-resistant KKU214GemR
cell line.
Discussion
Several previous studies have demonstrated that
anthocyanins exert anticancer activity against various types of
cancer, including HCC, melanoma, colon cancer, lung cancer and
breast cancer (37–39). In our previous study, we developed
a novel AC, which primarily consisted of extracts of purple corn
cobs, blue butterfly peas and turmeric (26). The combination of these exhibited
increased antioxidant activity and increased thermal stability
compared with that of individual extracts (26). In the present study, to the best of
our knowledge, we have identified, for the first time, anti-CCA
activity of AC, which exhibited cytotoxicity against a CCA cell
line through the suppression of cell proliferation and induction of
caspase-independent apoptosis probably via increased mitochondrial
superoxide production. Furthermore, AC also suppressed the
expression of a number of oncogenic proteins that have previously
been reported to be upregulated in CCA, including FOXM1, NF-κB and
ER stress-response proteins. The potential underlying molecular
mechanisms of AC treatment on CCA cells are summarized in Fig. 10. AC was able to potentiate
gemcitabine treatment against the gemcitabine-resistant
KKU214GemR cell line. We therefore hypothesize that AC
has a potential function as an alternative or supplementary
treatment for CCA.
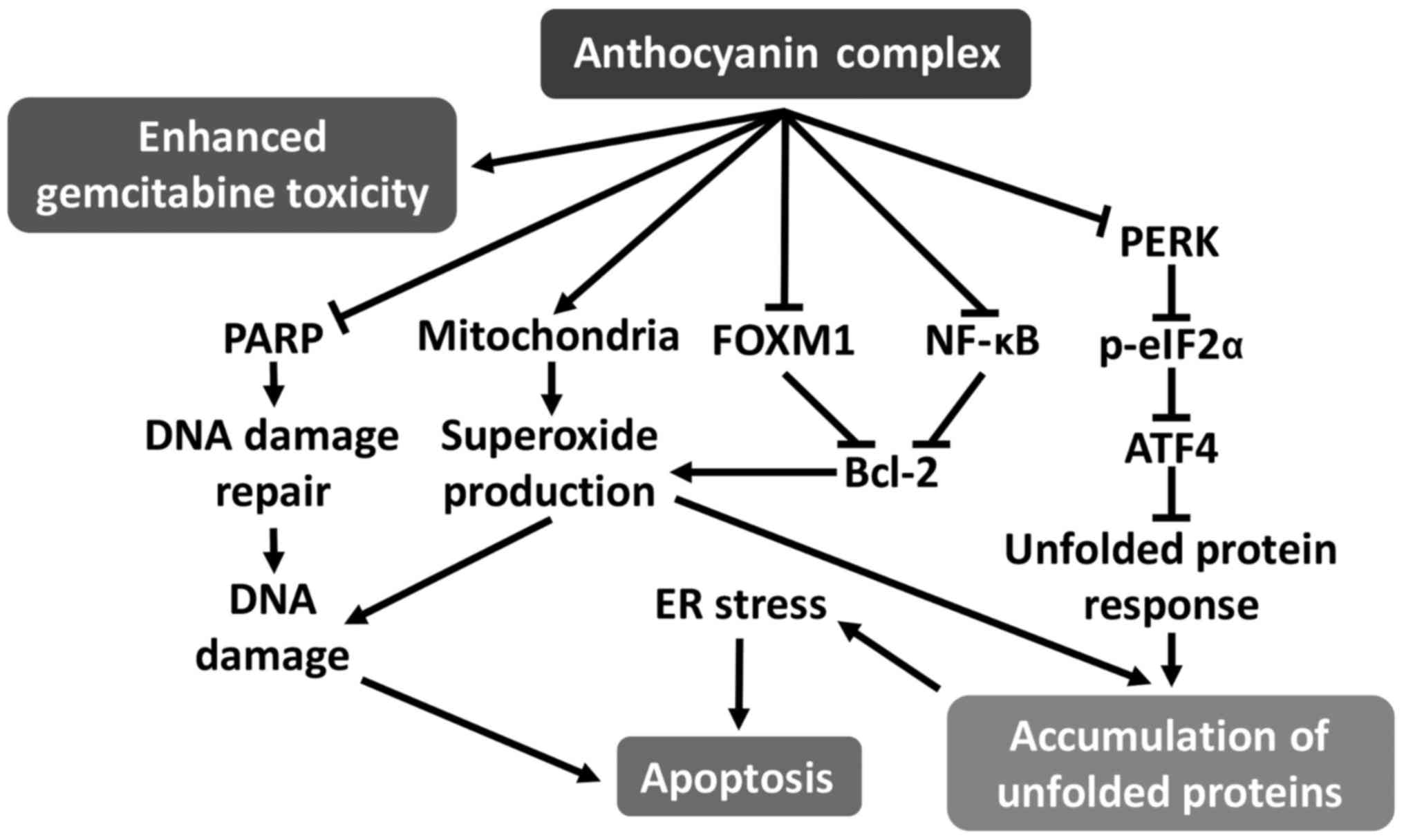 | Figure 10Schematic diagram of the postulated
mechanism of AC against CCA cell lines. AC treatment downregulates
FOXM1 and NF-κB expression, leading to suppression of Bcl-2
expression. AC treatment also stimulates superoxide production from
mitochondria, in part due to Bcl-2 suppression, since this protein
serves a function in inhibiting superoxide production from
mitochondria. Massive superoxide production leads to DNA damage.
Inhibition of the DNA damage-repair protein PARP by AC treatment
may induce DNA damage-mediated apoptosis. In addition to DNA
damage, superoxide production may result in the induction of
protein misfolding or of unfolded proteins, accumulation of which
stimulates the UPR in the ER. AC treatment inhibits the primary UPR
pathway (PERK/eIF2α/ATF4), leading to ER stress and induction of
apoptosis. AC, anthocyanin complex; CCA, cholangiocarcinoma; FOXM1,
forkhead box M1; NF-κB, nuclear factor-κB; Bcl-2, B-cell
lymphoma-2; PARP, poly(ADP-ribose) polymerase; UPR, unfolded
protein response; ER, endoplasmic reticulum; PERK, protein kinase
RNA-like ER kinase; eIF2α, eukaryotic initiation factor 2α; ATF4,
activating transcription factor 4. |
cDNA analysis of O. viverrini-associated CCA
clinical samples revealed that FOXM1 was highly expressed, being
the second most abundant gene in these samples (40). FOXM1 has been implicated primarily
in cell proliferation and survival, as well as in cancer
progression (34). In the present
study, suppression of cell proliferation and induction of cellular
apoptosis following AC treatment were observed alongside
downregulation of FOXM1 expression. These results suggest that
suppression of FOXM1 is, in part, the mode of action of AC against
CCA. To the best of our knowledge, the present study is the first
to identify the effects of cyanidin and delphinidin on FOXM1
expression. However, a previous study demonstrated that curcumin,
an important bioactive compound in turmeric extract, was able to
suppress FOXM1 (41). Therefore,
suppression of FOXM1 using AC treatment is potentially mediated by
either anthocyanin(s) (cyanidin and/or delphinidin) or turmeric
extract or both. In addition to FOXM1, western blot analysis also
demonstrated the downregulation of NF-κB in AC-treated CCA cells.
NF-κB is a well-known transcription factor associated with
inflammation and cancer (42–44).
A previous study has identified that NF-κB, particularly its p65
subunit, was overexpressed in O. viverrini-associated CCA
tumor tissues. Furthermore, treatment with
dehydroxymethylepoxyquin-omicin inhibited CCA cell proliferation
and induced apoptosis in KKU213 cells via the suppression of NF-κB
and Bcl-2 proteins (45). The
results of the present study are in general agreement with this
previous study, demonstrating that the p65 subunit of NF-κB and
Bcl-2 were also suppressed concurrently with induction of apoptosis
in the AC-treated KKU213 CCA cell line. Cyanidin and delphinidin
are known to suppress the expression of NF-κB and Bcl-2 (46,47).
Furthermore, the turmeric extract in AC was also identified to
suppress NF-κB and Bcl-2 (48,49).
Therefore, we hypothesized that cyanidin, delphinidin and turmeric
extract in AC may synergistically inhibit NF-κB and Bcl-2.
Curcumin, an important active ingredient in turmeric extract, is
able to inhibit NF-κB expression and induce cancer cell apoptosis.
AC, composed of purple corn cob, blue butterfly pea and turmeric
extracts at a ratio of 7:2:1, contains ~60 µg turmeric
extract per 600 µg/ml AC. In its standard form, turmeric
contains 5% curcumin, implying that there is ~3 µg or 8
µM curcumin in 60 µg turmeric extract. In our
previous study, we identified that significant suppression of
proliferation, induction of apoptosis (4% of the cell population
compared with the untreated control) and suppression of
pro-survival proteins in the KKU213 cell line were only observed
when treated with 50 µM curcumin (49). The synergistic effect of combining
different anthocyanins was also demonstrated previously (50,51).
These results support our hypothesis that the cytotoxicity of AC
against CCA cells is mediated by synergism of cyanidin, delphinidin
and turmeric extract. Notably, in the present study, the function
of p65 on AC treatment of KKU213 cells was also determined using
p65 plasmid DNA transfection coupled with a clonogenic assay.
Although p65 was successfully induced in KKU213 cells, the
clonogenic assay revealed that induction of pro-survival
transcription factor expression was not able to protect KKU213
cells from apoptosis following AC treatment. Failure of NF-κB to
prevent cell death has also been demonstrated in tumor necrosis
factor α-induced apoptosis in the HeLa cell line (52). Furthermore, failure to escape
apoptosis following p65 induction may be because AC treatment
induces CCA cell apoptosis through multiple pathways. Therefore, AC
is a potent apoptosis inducer against CCA cells.
AC treatment also markedly induced superoxide
production from mitochondria in KKU213 cells. Mitochondrial
super-oxide has been demonstrated as an inducer of apoptosis
(32). Apoptosis induced in this
manner is consistent with a previous study demonstrating that
diphenyleneiodonium induces mitochondrial superoxide-mediated
apoptosis (53). Notably, in the
present study, cleaved PARP (data not shown) and cleaved caspase-3
were not observed after 24 h of AC treatment in CCA cell lines,
suggesting that caspase-independent cell death occurred without
activating Bcl-2-associated X protein, cytochrome c and
caspase-3 (54).
Caspase-independent cell death was also indicated by treatment with
a combination of AC and pan-caspase inhibitor (Q-VD-OPh): addition
of Q-VD-OPh was not able to protect CCA cells from AC-mediated cell
death. The defects in the caspase activation pathway are common in
cancer, leading to resistance to certain pro-apoptotic stimuli
(55). Therefore, triggering
caspase-independent cell death is one of the novel approaches to
treat cancer (56). Therefore, in
addition to CCA, the AC may be useful for the treatment of cancer,
particularly that with the ability to evade caspase-dependent cell
death. Decreasing PARP expression in CCA cell lines was observed
following AC treatment. This result may be explained by the fact
that this protein is involved in DNA damage repair (57). Induction of mitochondrial
super-oxide production was able to induce DNA damage (58,59),
suggesting that decreasing of PARP expression after AC treatment
could lead to a decrease in DNA damage repair by PARP. Therefore,
we hypothesize that AC-treated cells underwent apoptosis via DNA
damage-induced caspase-independent cell death (60).
Superoxide in mitochondria may be converted into
H2O2 and diffuse into the cytoplasm.
H2O2 may be catalyzed further to form other
highly reactive oxygen species (ROS). Under basal physiological
conditions, ROS accumulation may be prevented by cellular
antioxidant defense mechanisms (61). However, excessive ROS production
caused by excessive mitochondrial superoxide production triggers
disturbance of ER redox homeostasis, thus aggravating the
accumulation of misfolded or unfolded proteins in the ER or ER
stress (62). The process known as
the unfolded protein response (UPR) is thus activated to restore ER
homeostasis (63). However, if ER
stress is severe or prolonged, it may induce cell death (64). The PERK/eIF2α/ATF4 axis is
important in UPR signaling during ER stress (65). PERK activates the phosphorylation
of eIF2α at Ser51, resulting in global translation
inhibition (66), but induces the
expression of ATF4 (67) to
overcome ER stress. The results of the present study identified
downregulation of PERK/eIF2α/ATF4 in an AC-treated CCA cell line.
Therefore, we hypothesize that AC treatment induces ER stress and
eventual cell death via the stimulation of mitochondrial superoxide
production and suppression of PERK/eIF2α/ATF4 axis-mediated UPR.
This hypothesis is supported by previous studies demonstrating that
inhibition of PERK or eIF2α rendered cells susceptible to ER
stress-mediated cell death (68–70).
Furthermore, a recent study has demonstrated that activation of
PERK and eIF2α was inhibited in cyanidin-treated cells (71).
Drug resistance is a major barrier for chemotherapy
in a number of types of cancer, including CCA. Enhancement of
chemotherapeutic drug treatment and chemosensitization of cancer
cells by plant polyphenols have been the focus of much work and
discussion (72). In the present
study, the effect of co-treatment of AC and gemcitabine against a
gemcitabine-resistant KKU214GemR cell line was
investigated. It was identified that KKU214GemR cells
were ~80-fold more resistant to gemcitabine compared with its
parental cell line. However, the combination of AC with gemcitabine
significantly enhanced the efficacy of gemcitabine against
KKU214GemR cells compared with single agent treatment.
Although the potential of AC against a CCA cell line as well as its
potential as a chemo-sensitizer in a gemcitabine-resistant CCA cell
line have been demonstrated in the present study, these results
have not been verified in an animal model of CCA. Therefore, the
anticancer activity of AC should be investigated further in
xenograft mouse or liver fluke-induced hamster CCA models.
Furthermore, the anticancer potential of AC requires testing in
diverse cancer cell types to support its potential use as an
alternative or supplementary treatment for cancer, particularly
CCA. Additionally, high-throughput approaches, including RNA
sequencing, are required to explore the precise mechanisms
underlying anticancer and chemosensitization activities of AC.
In conclusion, the results of the present study
demonstrated that AC possesses cytotoxicity against CCA cell lines
by suppression of cell proliferation and induction of
caspase-independent apoptosis possibly via downregulation of FOXM1,
NF-κB and the ER stress response, and by induction of mitochondrial
superoxide production. AC also sensitizes KKU214GemR
cells to gemcitabine treatment. Therefore, AC has potential as an
alternative treatment agent and may assist in overcoming drug
resistance of CCA when co-administered with other chemotherapeutic
agents.
Acknowledgments
The authors thank Miss Sucharat Limsitthichaikoon
for assisting with AC preparation and the Research Instrument Unit,
Faculty of Medicine, Khon Kaen University, Khon Kaen, Thailand, for
technical and facility support.
Abbreviations:
|
AC
|
anthocyanin complex
|
|
AEBSF
|
4-(2-aminoethyl) benzenesulfonyl
fluoride hydrochloride
|
|
ATF4
|
activating transcription factor 4
|
|
BCA
|
bicinchoninic acid
|
|
CCA
|
cholangiocarcinoma
|
|
DMEM
|
Dulbecco's modified Eagle's medium
|
|
DMSO
|
dimethyl sulfoxide
|
|
ER
|
endoplasmic reticulum
|
|
FBS
|
fetal bovine serum
|
|
FOXM1
|
forkhead box protein M1
|
|
HBSS
|
Hank's balanced salt solution
|
|
NF-κB
|
nuclear factor-κB
|
|
PERK
|
protein kinase RNA-like endoplasmic
reticulum kinase
|
|
eIF2α
|
eukaryotic initiation factor 2α
|
|
PVDF
|
polyvinylidene difluoride
|
|
RIPA
|
radioimmunoprecipitation assay
|
|
SRB
|
sulforhodamine B.
|
Notes
[1]
Funding
This study was supported in part by the Thailand
Research Fund (TRF) and Khon Kaen University through Royal Golden
Jubilee PhD joint funding program (grant no. PHD/0166/2553) and
also by the TRF and Medical Research Council (MRC) UK via TRF-MRC
Joint Health Research (grant no. DBG5980004) and Khon Kaen
University Research Fund (grant no. KKU600701).
[2] Availability
of data and materials
The data sets generated during the study are
available from the corresponding author on reasonable request.
[3] Authors'
contributions
SP and KI mainly designed, performed the research
and wrote the manuscript. KI analyzed the data, and AP, CP, CH, KV
and PP helped to prepare and provided materials, and analyzed the
data. All authors have read and approved the final version of the
manuscript.
[4] Ethics
approval and consent to participate
Not applicable.
[5] Consent for
publication
Not applicable.
[6] Competing
interests
The authors declare that they have no competing
interests.
References
|
1
|
Banales JM, Cardinale V, Carpino G,
Marzioni M, Andersen JB, Invernizzi P, Lind GE, Folseraas T, Forbes
SJ, Fouassier L, et al: Expert consensus document:
Cholangiocarcinoma: Current knowledge and future perspectives
consensus statement from the European Network for the Study of
Cholangiocarcinoma (ENS-CCA). Nat Rev Gastroenterol Hepatol.
13:261–280. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kirstein MM and Vogel A: Epidemiology and
risk factors of cholangiocarcinoma. Visc Med. 32:395–400. 2016.
View Article : Google Scholar
|
|
3
|
Sripa B and Pairojkul C:
Cholangiocarcinoma: Lessons from Thailand. Curr Opin Gastroenterol.
24:349–356. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
IARCNA: A Review of Human Carcinogens:
Opisthorchis viverrini and Clonorchis sinensis. IARC Monogr Eval
Carcinog Risks Hum. 100B:341–370. 2012.
|
|
5
|
Sripa B, Brindley PJ, Mulvenna J, Laha T,
Smout MJ, Mairiang E, Bethony JM and Loukas A: The tumorigenic
liver fluke Opisthorchis viverrini - multiple pathways to cancer.
Trends Parasitol. 28:395–407. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Prakobwong S, Pinlaor S, Yongvanit P,
Sithithaworn P, Pairojkul C and Hiraku Y: Time profiles of the
expression of metalloproteinases, tissue inhibitors of
metalloproteases, cytokines and collagens in hamsters infected with
Opisthorchis viverrini with special reference to peribiliary
fibrosis and liver injury. Int J Parasitol. 39:825–835. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Loaharanu P and Sornmani S: Preliminary
estimates of economic impact of liver fluke infection in Thailand
and the feasibility of irradiation as a control measure. Southeast
Asian J Trop Med Public Health. 22(Suppl 22): 384–390.
1991.PubMed/NCBI
|
|
8
|
Khan SA, Davidson BR, Goldin R, Pereira
SP, Rosenberg WM, Taylor-Robinson SD, Thillainayagam AV, Thomas HC,
Thursz MR and Wasan H; British Society of Gastroenterology:
Guidelines for the diagnosis and treatment of cholangiocarcinoma:
Consensus document. Gut. 51(Suppl 6): VI1–VI9. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Anderson CD, Pinson CW, Berlin J and Chari
RS: Diagnosis and treatment of cholangiocarcinoma. Oncologist.
9:43–57. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Valle J, Wasan H, Palmer DH, Cunningham D,
Anthoney A, Maraveyas A, Madhusudan S, Iveson T, Hughes S, Pereira
SP, et al ABC-02 Trial Investigators: Cisplatin plus gemcitabine
versus gemcitabine for biliary tract cancer. N Engl J Med.
362:1273–1281. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Surh YJ: Cancer chemoprevention with
dietary phytochemicals. Nat Rev Cancer. 3:768–780. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang H, Khor TO, Shu L, Su ZY, Fuentes F,
Lee JH and Kong AN: Plants vs. cancer: A review on natural
phytochemicals in preventing and treating cancers and their
druggability. Anticancer Agents Med Chem. 12:1281–1305. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Singh CK, Siddiqui IA, El-Abd S, Mukhtar H
and Ahmad N: Combination chemoprevention with grape antioxidants.
Mol Nutr Food Res. 60:1406–1415. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Castañeda-Ovando A, Pacheco-Hernández ML,
Páez-Hernández ME, Rodríguez JA and Galán-Vidal CA,
Pacheco-Hernández MdL, Páez-Hernández ME, Rodríguez JA and
Galán-Vidal CA: Chemical studies of anthocyanins: A review. Food
Chem. 113:859–871. 2009. View Article : Google Scholar
|
|
15
|
Cerletti C, De Curtis A, Bracone F, Digesù
C, Morganti AG, Iacoviello L, de Gaetano G and Donati MB: Dietary
anthocyanins and health: Data from FLORA and ATHENA EU projects. Br
J Clin Pharmacol. 83:103–106. 2017. View Article : Google Scholar :
|
|
16
|
Lin BW, Gong CC, Song HF and Cui YY:
Effects of anthocyanins on the prevention and treatment of cancer.
Br J Pharmacol. 174:1226–1243. 2017. View Article : Google Scholar
|
|
17
|
Li D, Wang P, Luo Y, Zhao M and Chen F:
Health benefits of anthocyanins and molecular mechanisms: Update
from recent decade. Crit Rev Food Sci Nutr. 57:1729–1741. 2017.
View Article : Google Scholar
|
|
18
|
Yousuf B, Gul K, Wani AA and Singh P:
Health benefits of anthocyanins and their encapsulation for
potential use in food systems: A Review. Crit Rev Food Sci Nutr.
56:2223–2230. 2016. View Article : Google Scholar
|
|
19
|
Nichenametla SN, Taruscio TG, Barney DL
and Exon JH: A review of the effects and mechanisms of
polyphenolics in cancer. Crit Rev Food Sci Nutr. 46:161–183. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Umar Lule S and Xia W: Food phenolics,
pros and cons: A Review. Food Rev Int. 21:367–388. 2005. View Article : Google Scholar
|
|
21
|
Wang LS and Stoner GD: Anthocyanins and
their role in cancer prevention. Cancer Lett. 269:281–290. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang X, Luo E, Liu X, Han B, Yu X and Peng
X: Delphinidin-3-glucoside suppresses breast carcinogenesis by
inactivating the Akt/HOTAIR signaling pathway. BMC Cancer.
16:4232016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen PN, Chu SC, Chiou HL, Kuo WH, Chiang
CL and Hsieh YS: Mulberry anthocyanins, cyanidin 3-rutinoside and
cyanidin 3-glucoside, exhibited an inhibitory effect on the
migration and invasion of a human lung cancer cell line. Cancer
Lett. 235:248–259. 2006. View Article : Google Scholar
|
|
24
|
Kamei H, Kojima T, Hasegawa M, Koide T,
Umeda T, Yukawa T and Terabe K: Suppression of tumor cell growth by
anthocyanins in vitro. Cancer Invest. 13:590–594. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Eiro MJ and Heinonen M: Anthocyanin color
behavior and stability during storage: Effect of intermolecular
copigmentation. J Agric Food Chem. 50:7461–7466. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Intuyod K, Priprem A, Limphirat W,
Charoensuk L, Pinlaor P, Pairojkul C, Lertrat K and Pinlaor S:
Anti-inflammatory and anti-periductal fibrosis effects of an
anthocyanin complex in Opisthorchis viverrini-infected hamsters.
Food Chem Toxicol. 74:206–215. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tepsiri N, Chaturat L, Sripa B, Namwat W,
Wongkham S, Bhudhisawasdi V and Tassaneeyakul W: Drug sensitivity
and drug resistance profiles of human intrahepatic
cholangiocarcinoma cell lines. World J Gastroenterol. 11:2748–2753.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wattanawongdon W, Hahnvajanawong C, Namwat
N, Kanchanawat S, Boonmars T, Jearanaikoon P, Leelayuwat C,
Techasen A and Seubwai W: Establishment and characterization of
gemcitabine-resistant human cholangiocarcinoma cell lines with
multidrug resistance and enhanced invasiveness. Int J Oncol.
47:398–410. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kuželová K, Grebeňová D and Brodská B:
Dose-dependent effects of the caspase inhibitor Q-VD-OPh on
different apoptosis-related processes. J Cell Biochem.
112:3334–3342. 2011. View Article : Google Scholar
|
|
30
|
Franken NA, Rodermond HM, Stap J, Haveman
J and van Bree C: Clonogenic assay of cells in vitro. Nat Protoc.
1:2315–2319. 2006. View Article : Google Scholar
|
|
31
|
Puck TT and Marcus PI: Action of x-rays on
mammalian cells. J Exp Med. 103:653–666. 1956. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cai J and Jones DP: Superoxide in
apoptosis. Mitochondrial generation triggered by cytochrome c loss.
J Biol Chem. 273:11401–11404. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Piva R, Belardo G and Santoro MG:
NF-kappaB: A stress-regulated switch for cell survival. Antioxid
Redox Signal. 8:478–486. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Myatt SS and Lam EW: The emerging roles of
forkhead box (Fox) proteins in cancer. Nat Rev Cancer. 7:847–859.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Cao SS and Kaufman RJ: Endoplasmic
reticulum stress and oxidative stress in cell fate decision and
human disease. Antioxid Redox Signal. 21:396–413. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Holohan C, Van Schaeybroeck S, Longley DB
and Johnston PG: Cancer drug resistance: An evolving paradigm. Nat
Rev Cancer. 13:714–726. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Longo L, Platini F, Scardino A, Alabiso O,
Vasapollo G and Tessitore L: Autophagy inhibition enhances
anthocyanin-induced apoptosis in hepatocellular carcinoma. Mol
Cancer Ther. 7:2476–2485. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Neto CC, Amoroso JW and Liberty AM:
Anticancer activities of cranberry phytochemicals: An update. Mol
Nutr Food Res. 52(Suppl 1): S18–S27. 2008.PubMed/NCBI
|
|
39
|
Rugină D, Hanganu D, Diaconeasa Z, Tăbăran
F, Coman C, Leopold L, Bunea A and Pintea A: Antiproliferative and
apoptotic potential of cyanidin-based anthocyanins on melanoma
cells. Int J Mol Sci. 18:949–959. 2017. View Article : Google Scholar
|
|
40
|
Jinawath N, Chamgramol Y, Furukawa Y,
Obama K, Tsunoda T, Sripa B, Pairojkul C and Nakamura Y: Comparison
of gene expression profiles between Opisthorchis viverrini and
non-Opisthorchis viverrini associated human intrahepatic
cholangiocarcinoma. Hepatology. 44:1025–1038. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhang JR, Lu F, Lu T, Dong WH, Li P, Liu
N, Ma DX and Ji CY: Inactivation of FoxM1 transcription factor
contributes to curcumin-induced inhibition of survival,
angiogenesis, and chemosensitivity in acute myeloid leukemia cells.
J Mol Med (Berl). 92:1319–1330. 2014. View Article : Google Scholar
|
|
42
|
Karin M: Nuclear factor-kappaB in cancer
development and progression. Nature. 441:431–436. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Hoesel B and Schmid JA: The complexity of
NF-κB signaling in inflammation and cancer. Mol Cancer. 12:862013.
View Article : Google Scholar
|
|
44
|
Baldwin AS Jr: The NF-kappa B and I kappa
B proteins: New discoveries and insights. Annu Rev Immunol.
14:649–683. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Seubwai W, Wongkham C, Puapairoj A,
Khuntikeo N, Pugkhem A, Hahnvajanawong C, Chaiyagool J, Umezawa K,
Okada S and Wongkham S: Aberrant expression of NF-κB in liver fluke
associated cholangiocarcinoma: Implications for targeted therapy.
PLoS One. 9:e1060562014. View Article : Google Scholar
|
|
46
|
Hafeez BB, Siddiqui IA, Asim M, Malik A,
Afaq F, Adhami VM, Saleem M, Din M and Mukhtar H: A dietary
anthocyanidin delphinidin induces apoptosis of human prostate
cancer PC3 cells in vitro and in vivo: Involvement of nuclear
factor-kappaB signaling. Cancer Res. 68:8564–8572. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Hecht SS, Huang C, Stoner GD, Li J, Kenney
PM, Sturla SJ and Carmella SG: Identification of cyanidin
glycosides as constituents of freeze-dried black raspberries which
inhibit anti-benzo[a]pyrene-7,8-diol-9,10-epoxide induced NFkappaB
and AP-1 activity. Carcinogenesis. 27:1617–1626. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Kim JH, Gupta SC, Park B, Yadav VR and
Aggarwal BB: Turmeric (Curcuma longa) inhibits inflammatory nuclear
factor (NF)-κB and NF-κB-regulated gene products and induces death
receptors leading to suppressed proliferation, induced
chemo-sensitization, and suppressed osteoclastogenesis. Mol Nutr
Food Res. 56:454–465. 2012. View Article : Google Scholar
|
|
49
|
Prakobwong S, Gupta SC, Kim JH, Sung B,
Pinlaor P, Hiraku Y, Wongkham S, Sripa B, Pinlaor S and Aggarwal
BB: Curcumin suppresses proliferation and induces apoptosis in
human biliary cancer cells through modulation of multiple cell
signaling pathways. Carcinogenesis. 32:1372–1380. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Rahman MM, Ichiyanagi T, Komiyama T,
Hatano Y and Konishi T: Superoxide radical- and
peroxynitrite-scavenging activity of anthocyanins;
structure-activity relationship and their synergism. Free Radic
Res. 40:993–1002. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Kausar H, Jeyabalan J, Aqil F, Chabba D,
Sidana J, Singh IP and Gupta RC: Berry anthocyanidins
synergistically suppress growth and invasive potential of human
non-small-cell lung cancer cells. Cancer Lett. 325:54–62. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Casanelles E, Gozzelino R,
Marqués-Fernández F, Iglesias-Guimarais V, Garcia-Belinchón M,
Sánchez-Osuna M, Solé C, Moubarak RS, Comella JX and Yuste VJ:
NF-κB activation fails to protect cells to TNFα-induced apoptosis
in the absence of Bcl-xL, but not Mcl-1, Bcl-2 or Bcl-w. Biochim
Biophys Acta. 1833:1085–1095. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Li N, Ragheb K, Lawler G, Sturgis J, Rajwa
B, Melendez JA and Robinson JP: DPI induces mitochondrial
superoxide-mediated apoptosis. Free Radic Biol Med. 34:465–477.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Kondo K, Obitsu S, Ohta S, Matsunami K,
Otsuka H and Teshima R: Poly(ADP-ribose) polymerase
(PARP)-1-independent apoptosis-inducing factor (AIF) release and
cell death are induced by eleostearic acid and blocked by
alpha-tocopherol and MEK inhibition. J Biol Chem. 285:13079–13091.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Fernald K and Kurokawa M: Evading
apoptosis in cancer. Trends Cell Biol. 23:620–633. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Mathiasen IS and Jäättelä M: Triggering
caspase-independent cell death to combat cancer. Trends Mol Med.
8:212–220. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Herceg Z and Wang ZQ: Functions of
poly(ADP-ribose) polymerase (PARP) in DNA repair, genomic integrity
and cell death. Mutat Res. 477:97–110. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Schieber M and Chandel NS: ROS function in
redox signaling and oxidative stress. Curr Biol. 24:R453–R462.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Dizdaroglu M, Jaruga P, Birincioglu M and
Rodriguez H: Free radical-induced damage to DNA: Mechanisms and
measurement. Free Radic Biol Med. 32:1102–1115. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Borges HL, Linden R and Wang JY: DNA
damage-induced cell death: Lessons from the central nervous system.
Cell Res. 18:17–26. 2008. View Article : Google Scholar
|
|
61
|
Murphy MP: How mitochondria produce
reactive oxygen species. Biochem J. 417:1–13. 2009. View Article : Google Scholar
|
|
62
|
Higa A and Chevet E: Redox signaling loops
in the unfolded protein response. Cell Signal. 24:1548–1555. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Schröder M and Kaufman RJ: ER stress and
the unfolded protein response. Mutat Res. 569:29–63. 2005.
View Article : Google Scholar
|
|
64
|
Xu C, Bailly-Maitre B and Reed JC:
Endoplasmic reticulum stress: Cell life and death decisions. J Clin
Invest. 115:2656–2664. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Ron D: Translational control in the
endoplasmic reticulum stress response. J Clin Invest.
110:1383–1388. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Shi Y, Vattem KM, Sood R, An J, Liang J,
Stramm L and Wek RC: Identification and characterization of
pancreatic eukaryotic initiation factor 2 alpha-subunit kinase,
PEK, involved in translational control. Mol Cell Biol.
18:7499–7509. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Harding HP, Novoa I, Zhang Y, Zeng H, Wek
R, Schapira M and Ron D: Regulated translation initiation controls
stress-induced gene expression in mammalian cells. Mol Cell.
6:1099–1108. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Scheuner D, Song B, McEwen E, Liu C,
Laybutt R, Gillespie P, Saunders T, Bonner-Weir S and Kaufman RJ:
Translational control is required for the unfolded protein response
and in vivo glucose homeostasis. Mol Cell. 7:1165–1176. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Harding HP, Zhang Y, Bertolotti A, Zeng H
and Ron D: Perk is essential for translational regulation and cell
survival during the unfolded protein response. Mol Cell. 5:897–904.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Rajesh K, Krishnamoorthy J, Kazimierczak
U, Tenkerian C, Papadakis AI, Wang S, Huang S and Koromilas AE:
Phosphorylation of the translation initiation factor eIF2α at
serine 51 determines the cell fate decisions of Akt in response to
oxidative stress. Cell Death Dis. 6:e15912015. View Article : Google Scholar
|
|
71
|
Thummayot S, Tocharus C, Suksamrarn A and
Tocharus J: Neuroprotective effects of cyanidin against Aβ-induced
oxidative and ER stress in SK-N-SH cells. Neurochem Int. 101:15–21.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Garg AK, Buchholz TA and Aggarwal BB:
Chemosensitization and radiosensitization of tumors by plant
polyphenols. Antioxid Redox Signal. 7:1630–1647. 2005. View Article : Google Scholar : PubMed/NCBI
|