Introduction
Chronic myeloid leukemia (CML), a clonal disease
affecting hematopoietic stem cells, is driven by the BCR/ABL
oncopro-tein, a constitutively active tyrosine kinase. Patients
with CML in the chronic phase are treated with imatinib (IM) or
other tyrosine kinase inhibitors (TKIs), which are highly effective
at inducing remission and prolonging survival. However, TKIs do not
completely eliminate leukemia stem cells (LSCs), even in patients
who achieve deep molecular responses (1–3).
Thus, the identification of drugs that can target these LSCs is of
primary importance in order to acheive the eradication of CML.
Transcriptomic and proteomic analyses have revealed that p53 and
c-Myc play defining roles in CML-LSC survival (4). These results suggest that the dual
targeting of p53 and c-Myc may selectively eliminate LSCs in
patients with CML.
Human and murine myeloid leukemia cells can be
induced to differentiate into mature granulocytes and macrophages
by various differentiation-inducing agents (5,6).
There are a number of methods with which to induce granulocytic or
monocytic differentiation in leukemia cells, since
differentiation-inducing agents act by different mechanisms to
induce the production of more or less identical end-stage cells
(7–9). However, the downregulation of c-Myc
and the upregulation of p21 (a target gene of p53) are commonly
observed during the differentiation of myeloid leukemia cells
induced by various inducers of differentiation (10–12).
These results suggest that differentiation-inducing agents may be
useful regulators of c-Myc and p53 expression in leukemia
cells.
In this study, we examined the effects of agents
that induce the differentiation of acute myeloid leukemia (AML)
cells on the proliferation of CML cells in the presence of TKIs.
Among the former, cotylenin A (CN-A) was found to be the most
effective at inhibiting the clonogenic potential and proliferation
of CML cells in long-term culture in the presence of TKIs. CN-A,
which is a novel fusicoccane-diterpene glycoside with a complex
sugar moiety, has been shown to affect the differentiation of
leukemia cells that have been freshly isolated from patients with
AML in primary culture (6,13). The administration of CN-A has been
shown to significantly prolonged the survival of mice inoculated
with retinoid-resistant human promyelocytic leukemia NB4 cells, and
no appreciable adverse effects were observed (14). These findings thus suggest that
CN-A may be useful in CML therapy when combined with TKIs.
Materials and methods
Materials
RPMI-1640 medium, all-trans retinoic acid
(ATRA), doxorubicin, rapamycin and cytosine arabinoside (AraC) were
purchased from Sigma-Aldrich Japan (Tokyo, Japan). Dimethyl
sulfoxide (DMSO), 1α,25-dihydroxy vitamin D3 (VD3) and
sodium butyrate were purchased from Wako Chemicals (Osaka, Japan).
CN-A was a gift from Professor Takeshi Sassa, phycoerythrin
(PE)-labeled anti-CD38 antibody (cat. no. 555460) was obtained from
BD Immunocytometry Systems (San Jose, CA, USA). IM, dasatinib (DAS)
and nilotinib (NIL) were obtained from Selleck Chemicals (Houston,
TX, USA).
Patients with leukemia
Leukemic bone marrow specimens were collected at
diagnosis, after the patients provided written informed consent for
sample collection in accordance with institutional policy. The
Shimane University Institutional Committee on Ethics (Shimane,
Japan) approved the present study. The samples were obtained from
the following patients: Case 1 was a 71-year-old Japanese male who
was admitted to the Department of Oncology/Hematology, Shimane
University Hospital (Shimane, Japan), presenting with
hyperleukocytosis. Laboratory data upon admission were white blood
cell count (WBC) 169,040/μl; myelocytes, 13.5%;
metamyelocytes, 15.5%; and lactate dehydrogenase (LDH), 861 U/l.
Samples from case 1 were obtained in November, 2017. Case 2 was a
64-year-old Japanese female who was admitted to the Department of
Oncology/Hematology, Shimane University Hospital, presenting with
hyperleukocytosis. Laboratory data upon admission were WBC,
60,420/μl; myelocytes, 12.7%; metamyelocytes, 4.2%; and LDH,
674 U/l. Samples from case 2 were collected in December, 2017. In
both cases, CML was diagnosed on the basis of bone marrow
morphology and the standard cytogenetic translocation, t(9;22), in
98% of the cells by fluorescence in situ hybridization
(FISH) analysis.
Cells and cell culture
K562 (15) and
KU812 (16) cells were obtained
from the respective founders and were maintained in RPMI-1640
medium supplemented with 10% fetal bovine serum (Biowest, Nauille,
France) and 80 μg/ml gentamicin (MSD Co., Ltd., Tokyo,
Japan) at 37°C in a humidified atmosphere of 5% CO2 in
air.
Assay of cell growth and properties of
differentiated cells
Suspensions of cells (2×104 cells/ml) in
1 ml of culture medium were incubated with or without the test
compounds in multidishes. Cell numbers were counted using a Model
Z1 Coulter Counter (Beckman Coulter, Tokyo, Japan). The growth
inhibitory effects of the drugs were examined by determining the
concentrations of drugs that were required to reduce the cell
number to one-half of that in untreated cells (IC50).
Morphological changes were examined in cell smears stained with
May-Grünwald-Giemsa solution (Merck Japan, Tokyo, Japan). The
surface expression of CD38 was determined by monoclonal antibody
labeling and flow cytometry using a FACScan flow cytometer (BD
Biosciences, Franklin Lakes, NJ, USA), as previously described
(13).
Assay of cumulative cell number
The cells (2×104/ml) were cultured in
medium with the test compounds. Thereafter, the cell density of the
treated cells was kept at 1–8×105/ml to maintain the
growth phase. The medium of treated cultures was replaced with
fresh medium with or without the test compounds at least every 7
days. The cumulative cell number was calculated from the cell
counts and the dilution used when feeding the culture. Cell numbers
were counted using a Beckman Coulter Z1 Particle Counter.
Colony-forming assay
The K562 or KU812 cells (3×103 per dish)
were plated into 2 ml of a semi-solid medium with 0.8%
methylcellulose and 20% fetal bovine serum in triplicate multi-well
plates (12 wells, 3.5 cm2 growth area/well) for 7–14
days. A solution of 0.1 ml of PBS containing various concentrations
of the drugs was added to the semi-solid medium. To determine the
colony-forming ability of the leukemia cells from patients with
CML, heparinized bone marrow aspirations were diluted with
RPMI-1640 medium supplemented with 10% fetal bovine serum, overlaid
on 15 ml of Ficoll-Paque Plus (GE Healthcare Biosciences, Uppsala,
Sweden) and centrifuged at 500 × g for 30 min. The mononuclear
cells were washed twice and suspended in RPMI-1640 medium
supplemented with 10% fetal bovine serum, plated in semi-solid
culture medium with 20% serum at 105 cells/dish for
colony formation, and incubated at 37°C in a humidified atmosphere
of 5% CO2 in air. Colonies were photographed under an
inverted microscope (Model CKX41; Olympus, Tokyo, Japan). In serial
colony formation assays, the cells were serially replated after 7
days of culture.
Western blot analysis
The cells were packed after being washed with cold
PBS, and then lysed at 1.5×107 cells/ml in sample buffer
[63 mM Tris-HCl (pH 6.8), 15% glycerol, 2% sodium dodecyl sulfate
(SDS), 5% 2-mercaptoethanol and 0.005% bromophenol blue]. The
resultant lysates were resolved on 10% SDS-polyacrylamide gels.
Protein concentration was quantified using the Protein
Quantification kit-Rapid (Wako Pure Chemical Industries, Ltd.).
Equal amounts of protein (10 μg) were separated by SDS/PAGE
(10% gels) prior to transfer to polyvinylidene fluoride membranes
(Bio-Rad Laboratories, Hercules, CA, USA), and then blocked with
Block Ace (DS Pharma Biomedical Co., Ltd., Osaka, Japan) for 60 min
at room temperature. The membranes were then immunoblotted with
anti-p21 (#2947), anti-cMyc (#5606) and anti-β-actin (#4970)
antibodies (1:500 dilution) antibodies which were purchased from
Cell Signaling Technology Japan (Tokyo, Japan). Horseradish
peroxidase (HRP)-conjugated antibody (#7074S and 7076S; Cell
Signaling Technology, Danvers, MA, USA) was used as a secondary
antibody (1:2,000 dilution). The bands were developed by treatment
with the Immun-Star HRP Chemiluminescent kit (Bio-Rad Laboratories)
for 5 min at room temperature, and detected using a Fuji Lumino
Image Analyzer LAS-4000 system (Fuji Film, Tokyo, Japan).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from the cells using TRIzol
reagent (Sigma-Aldrich). Total RNA was converted to first-strand
cDNA primed with random hexamer in a reaction volume of 20
μl using an RNA PCR kit (qPCR RT Master Mix; Toyobo Co.,
Ltd., Osaka, Japan), and 2 μl of this reaction was used as a
template in real-time PCR. The primers were used as previously
described (17). The quantitative
PCR reaction was performed using a Takara TP860 Real-Time PCR
system (Takara Bio, Tokyo, Japan) according to the manufacturer’s
instructions (40 cycles of duration at 95°C for 15 sec and
annealing and extension at 60°C for 60 sec). The threshold cycle
values were normalized to the threshold value of
glyceraldehyde-3-phosphate dehydrogenase. Data analysis was
performed using the 2−ΔΔCq method (18).
Statistical analysis
The results are expressed as the means ± standard
deviation (SD). Statistical analysis was conducted with SPSS 19.0
software (IBM Japan, Tokyo, Japan). Statistical significance
between multiple groups was determined by ANOVA with a Bonferroni
post-hoc test, and between 2 groups using a Student’s t-test.
Significant differences were considered to exist for probabilities
below 5% (P<0.05).
Results
Combined effects of IM and
differentiation-inducing agents on the proliferation of CML
cells
IM inhibited the growth of the K562 and KU812 cells
in a concentration dependent manner; the IC50 values at
7 days were 25 and 18 nM, respectively (data not shown). The KU812
cells were treated with 30, 60 and 100 nM IM for 14 days, washed
with PBS, resuspended in drug-free culture medium, and cultured for
an additional 7 days. The cells were first treated with 100 nM IM
for 14 days. After washing, the cells were further treated without
or with IM. While continuous IM treatment effectively suppressed
proliferation, the depletion of IM from the medium allowed the
cells to re-grow, indicating that IM did not completely block the
ability of the cells to repopulate (Fig. 1A). Similar results were obtained
when the K562 cells were treated with 100 nM IM (data not shown).
We then examined the effects of several agents that are known to
induce the differentiation of AML cells on the proliferation of CML
cells (Table I). KU812 cells were
treated with various concentrations of the inducers in the presence
or absence of 30 nM IM. Most of the compounds we used effectively
synergized with IM to inhibit the growth of KU812 cells. Low
concentrations of ATRA and IM synergistically inhibited the
proliferation of KU812 cells, but ATRA did not affect the
proliferation of K562 cells even in the presence of IM. Combination
of butyrate and IM was not evident, suggesting that butyrate is not
a good partner of IM. On the other hand, CN-A alone greatly
inhibited the long-term proliferation of KU812 cells. In this
condition, therefore, the combined effects with IM were modest.
These results suggest that CN-A at concentrations less than
IC50 are enough to inhibit the long-term proliferation
of CML cells in the presence of IM. The results of treatment with
IC50 concentrations for 4 days are shown in Table I. CN-A was less cytotoxic than the
other compounds tested, but was the most potent at inhibiting cell
proliferation for 14 days (Table
I), suggesting that CN-A effectively suppressed the
self-renewal ability of these cells. CN-A and IM cooperatively
inhibited cell proliferation (Fig.
1B). These compounds also exerted combined effects on the K562
cells. While the K562 cells that were treated with 2 μg/ml
of CN-A or 30 nM IM for 14 days still grew in the drug-free
culture, those treated with CN-A plus IM lost the ability to
proliferate (Fig. 1C). Although
CN-A alone hardly affected the apoptosis of the CML cells, it
markedly enhanced the IM-induced apoptosis of the K562 cells (data
not shown). These compounds also exerted combined effects on the
clonogenic activity of the cells (Fig.
1D). The colony-forming ability of the K562 cells was markedly
inhibited by treatment with CN-A alone at a concentration of >4
μg/ml, and IM cooperatively inhibited the colony-forming
ability of the K562 cells treated with CN-A. Treatment with CN-A at
1 μg/ml completely suppressed the colony-forming ability of
the KU812 cells and IM did not alter this effect of CN-A on colony
formation (data not shown).
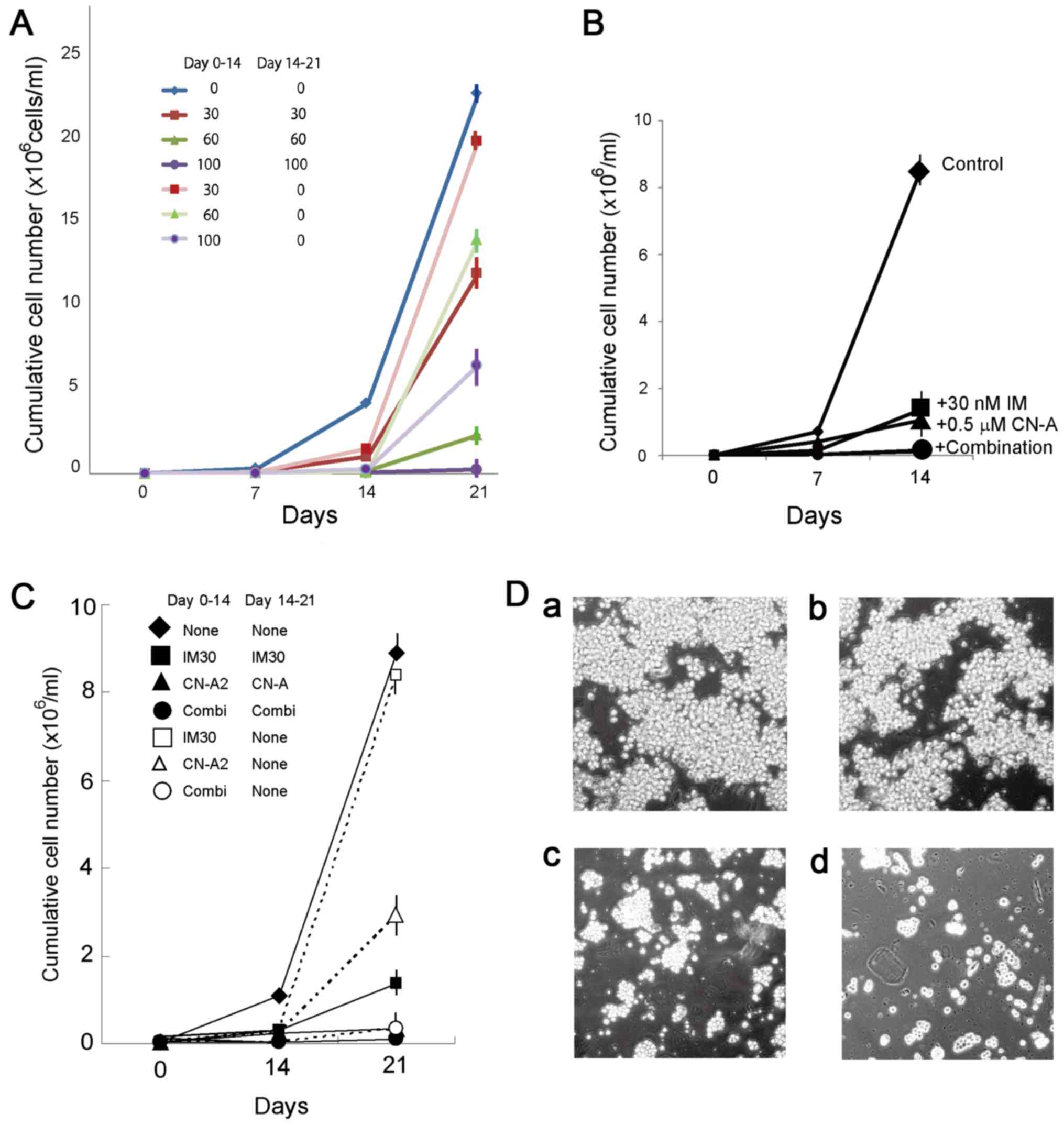 | Figure 1(A) Proliferation of KU812 cells in
long-term culture with IM. The KU812 cells were first treated with
0, 30, 60 or 100 nM IM (indicated in the color key) for 14 days.
After washing, the cells were treated with or without IM. The cell
density of the IM-treated cells was kept at 1–8×105
cells/ml. Data are representative of the means ± SD of 3
determinations. (B) Combined effects of CN-A and IM on the growth
of KU812 cells. Cells were treated with 30 nM IM (■), 0.5
μg/ml CN-A (▲), or 30 nM IM plus 0.5 μg/ml CN-A (●).
◆, Untreated cells. Data are representative of the means ± SD of 3
determinations. (C) Irreversible suppression of proliferation of
K562 cells by combined treatment with CN-A and IM. Cells were
treated with 30 nM IM (■), 2 μg/ml CN-A (▲), or 30 nM IM
plus 2 μg/ml CN-A (●) continuously. Cells were treated with
30 nM IM (■), 2 μg/ml CN-A (△), or 30 nM IM plus 2
μg/ml CN-A (○) for 14 days, and then washed and cultured
without drugs. ◆, Untreated cells. Data are representative of the
means ± SD of 3 determinations. (D) Colony formation of K562 cells
with IM and/or CN-A. Cells were cultured (a) without or (b) with 30
IM, (c) 6 μg/ml CN-A, (d) or 30 IM plus 6 μg/ml CN-A
for 7 days. IM, imatinib; CN-A, cotylenin A. |
 | Table IEffects of differentiation inducers
on the growth of KU812 cells in the presence or absence of IM. |
Table I
Effects of differentiation inducers
on the growth of KU812 cells in the presence or absence of IM.
| Compound |
Concentrationa | Cumulative cell
number (/ml)b
|
|---|
| None | +30 nM IM |
|---|
| None | |
3,778,000±265,000 | 920,000±73,000 |
| ATRA | 3 nM | 412,000±26,000 | 84,000±7,000 |
| CN-A | 1.2 μg/ml
(1.92 μM) | 54,000±4,200 | 42,000±3,400 |
| VD3 | 300 nM | 590,000±62,000 | 186,400±12,000 |
| Butyrate | 730 μM | 238,000±19,200 | 178,000±12,600 |
| DMSO | 105 mM | 265,000±22,000 | 138,600±11,600 |
| Rapamycin | 0.13 nM |
1,269,000±104,000 | 138,800±14,200 |
| Doxorubicin | 12.7 nM | 233,600±20,200 | 167,200±13,600 |
| AraC | 1.7 nM | 216,400±18,200 | 76,000±6,200 |
The effectiveness of combined treatment with CN-A
and IM on primary CML cells was examined and compared with that of
treatment with either IM or CN-A alone. CN-A significantly
inhibited colony formation by leukemia cells from a patient with
CML (case 1) (Fig. 2A). Although
IM alone did not significantly suppress colony formation, it
significantly enhanced the inhibitory effects of CN-A on colony
formation (Fig. 2A and B). The
clonogenic activity of the leukemia cells from case 2 was also
inhibited by treatment with IM plus CN-A, although the combined
effects on the cells from case 2 were less prominent than those on
the cells from case 1. The inhibitory effects of CN-A on the stem
cell potential of primary cells from case 2 were confirmed using
serial colony formation assays (Fig.
2C). CN-A, alone or in combination with IM, completely reduced
the replating efficiency of the cells from case 2. By contrast,
treatment with 30 nM IM did not reduce colony formation in
secondary cultures from case 2.
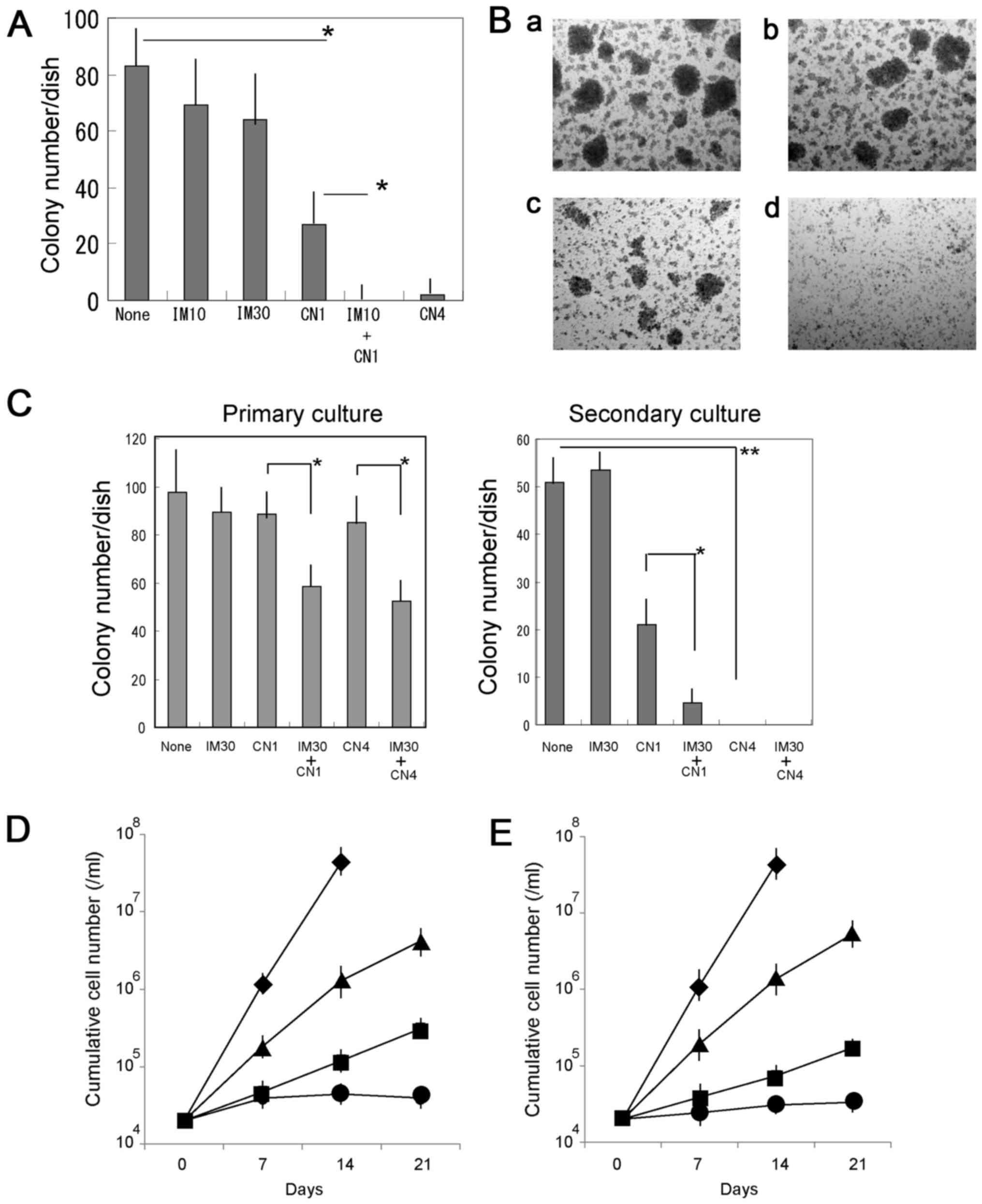 | Figure 2Combined effects of CN-A and IM on
colony formation of primary CML cells. (A) Bone marrow mononuclear
cells from a patient with CML (case 1) were plated in
methylcellulose-containing medium and treated with 10 nM IM (IM10),
30 nM IM (IM30), 1 μg/ml CN-A (CN1) or 4 μg/ml CN-A
(CN4) for 7 days. The results are presented as the means ± SD of 3
separate experiments. *P<0.05. (B) Colony formation
by primary CML (case 1) cells with IM and/or CN-A. Cells were
cultured (a) without or with (b) 10 nM IM, (c) 1 μg/ml CN-A,
or (d) 10 nM IM plus 1 μg/ml CN-A for 7 days. (C) Combined
treatment with CN-A and IM impaired serial colony formation by
primary CML (case 2) cells. Cells were cultured without or with
CN-A and/or IM for 7 days. The colony number was scored and cells
were replated in secondary cultures with or without drugs for an
additional 7 days. Results are presented as the means ± SD of 3
separate experiments. *P<0.05,
**P<0.01. (D and E) Combined effects of CN-A and
second-generation TKIs on the growth of KU812 cells in long-term
culture. (D) Cells were cultured without (◆) or with 0.2 nM DAS
(■), 1 μg/ml CN-A (▲) or 0.2 nM DAS plus 1 μg/ml CN-A
(●). (E) Cells were cultured without (◆) or with 0.3 nM NIL (■), 1
μg/ml CN-A (▲) or 0.3 nM NIL plus 1 μg/ml CN-A (●).
Results are presented as the means ± SD of 3 separate experiments.
IM, imatinib; CN-A, cotylenin A; CML, chronic myeloid leukemia. |
Two second-generation TKIs have been developed and
represent viable alternatives to IM. Thus, we examined the combined
effects of CN-A and these TKIs. Similar results were observed when
the cells were treated with CN-A plus other TKIs, such as DAS and
NIL (Fig. 2D and E).
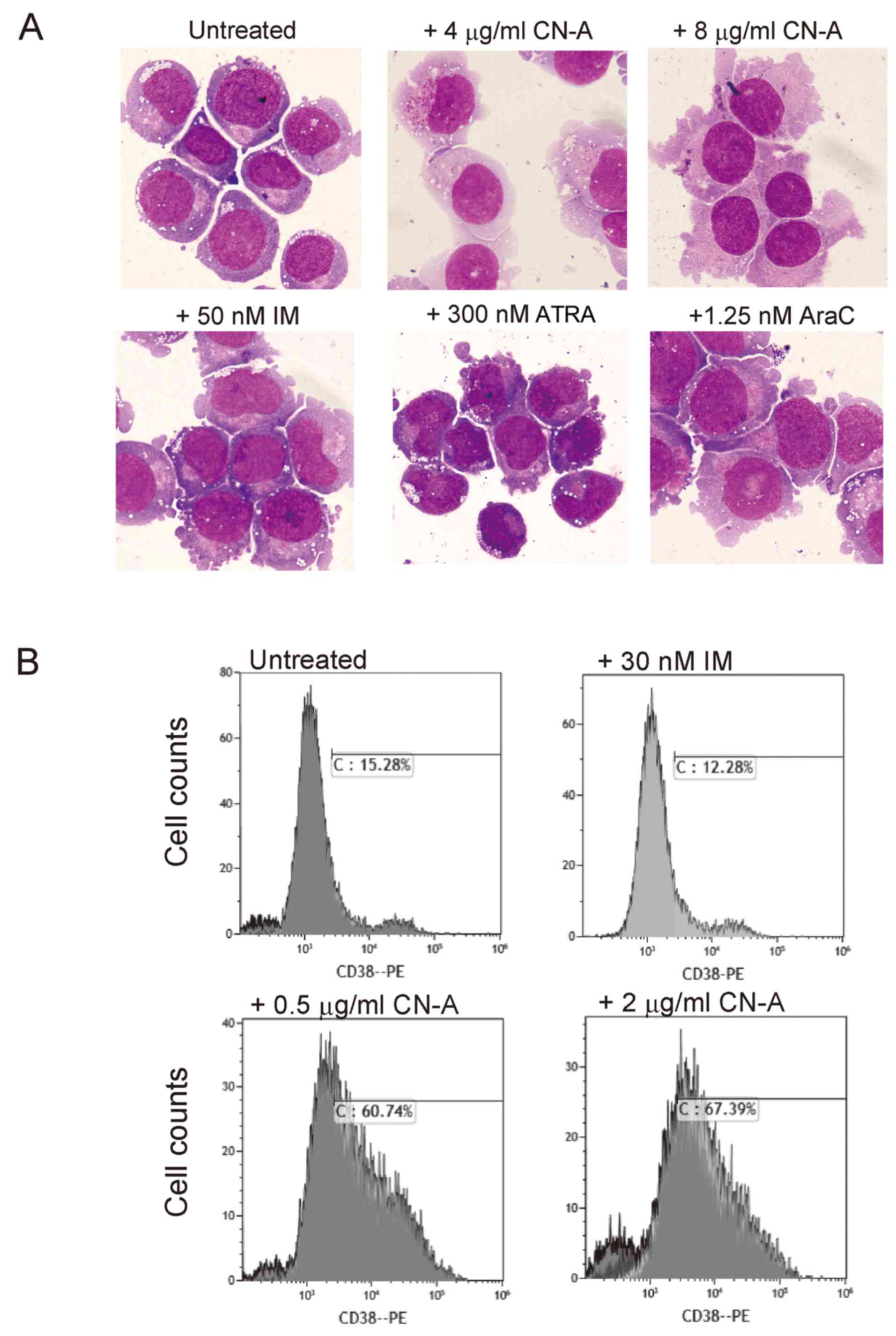
Modulation of the phenotypes of KU812
cells by CN-A
Since CN-A is a potent inducer of the
differentiation of AML cells (6,13),
in this study, we examined the effects of CN-A on the
differentiation-associated phenotypes of the K562 and KU812 cells.
CN-A induced morphological changes in these cells, whereas ATRA,
AraC and IM alone did not (Fig.
3A). The enlargement of the cytoplasm, decreased cytoplasmic
basophilicity and compact nuclei were observed in the CN-A-treated
cells. In the KU812 cells, morphological changes induced by CN-A
were not affected by IM. CN-A did not induce CD11b expression,
α-naphthyl acetate esterase activity or the reduction of nitroblue
tetrazolium (markers of myelomonocytic differentiation) in either
cell line even in the presence of IM (data not shown), although
CN-A has been shown to effectively induce these
differentiation-associated phenotypes in AML cell lines and AML
cells in primary culture (13).
Similar results were obtained in the CN-A-treated K562 cells (data
not shown).
CD38 is an important marker of human hematopoietic
stem cells. CD34+CD38− bone marrow cells are
highly enriched for long-term repopulating hematopoietic stem
cells, while CD34+CD38+ cells are more
committed progenitor cells. These same markers also apply to CML
cells (19). Therefore, in this
study, we examined the expression of CD38 in the CN-A-treated
cells. CN-A at concentrations as low as 0.5 μg/ml
efficiently converted the KU812 cells from CD38− to
CD38+ (Fig. 3B),
consistent with its role in promoting myeloid differentiation to an
intermediate stage, but not to mature stages. IM did not affect
CD38 expression even in the presence of CN-A.
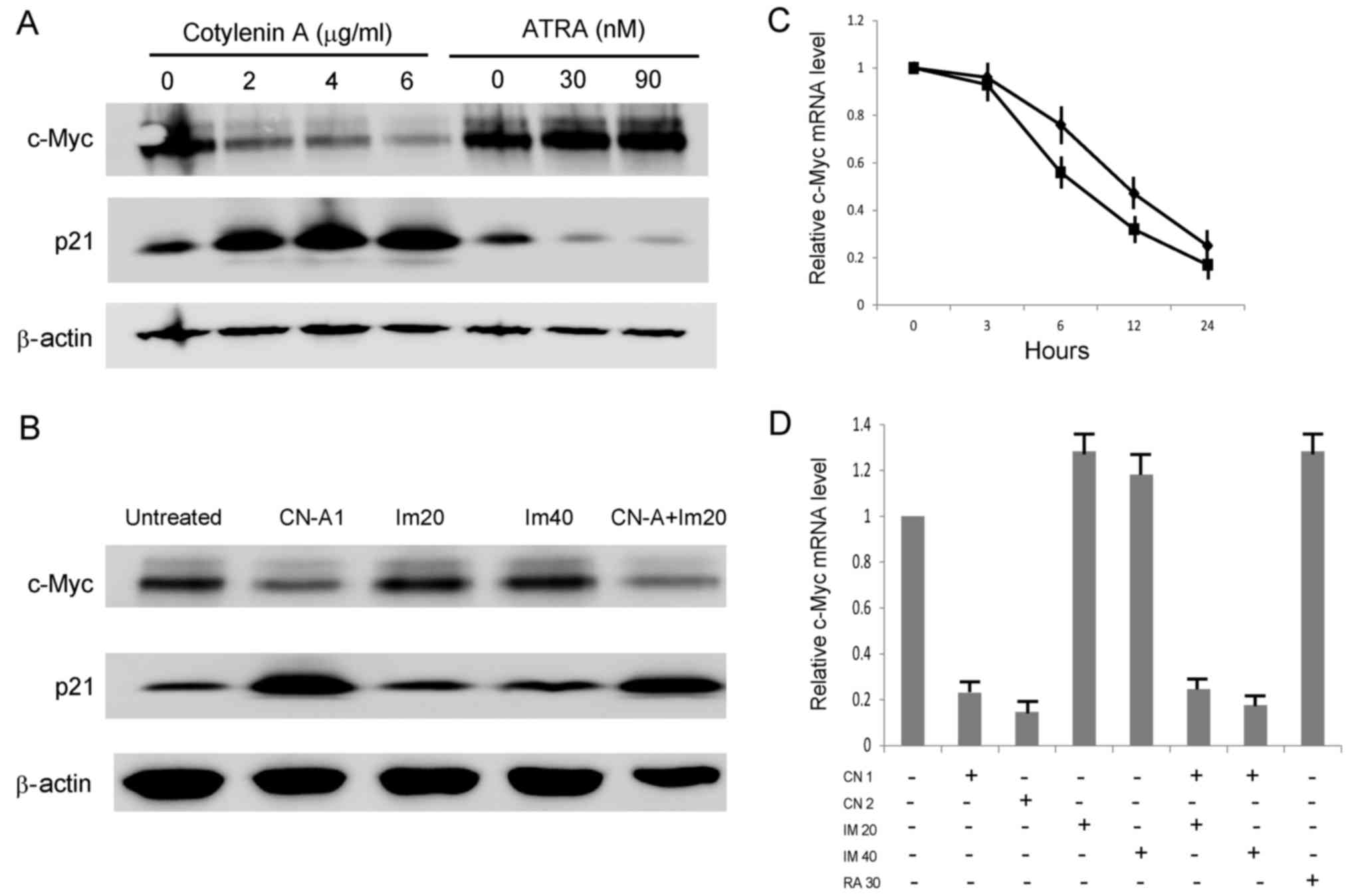
Effect of CN-A on the expression of c-Myc
and p21 in KU812 cells
A previous study indicated that the dual targeting
of p53 and c-Myc selectively eliminated CML stem cells (4). Since p21 is a target gene of p53, we
examined p21 expression as a marker of the p53 signal transduction
pathway. CN-A effectively decreased the c-Myc protein level and
increased the p21 protein level in the KU812 cells, while ATRA did
not affect the c-Myc protein level and decreased the p21 protein
level (Fig. 4A). IM did not affect
the protein levels of p21 or c-Myc in the KU812 cells, even in the
presence of CN-A (Fig. 4B). We
then examined the effect of CN-A on c-Myc mRNA expression in the
KU812 cells. The downregulation of c-Myc mRNA by CN-A was observed
within 6 h (Fig. 4C), suggesting
that it is an early event in the action of CN-A. Neither IM nor
ATRA affected mRNA expression (Fig.
4D).
Discussion
In the present study, the combination of TKIs and
CN-A was found to be more effective than either drug used alone at
reducing the CML cell bulk, and TKIs did not interfere with the
detrimental effect of CN-A on stem cell potential or clonogenic
activity.
Differentiation-inducing agents can alter the
phenotype of cancer cells. A previous study indicated that ATRA
converted KCL-22 CML cells from CD38− to
CD38+ cells (20), and
the results of the present study demonstrated that ATRA effectively
inhibited the growth of KU812 cells (Table I). However, ATRA did not affect the
growth of the K562 cells and did not effectively convert the KU812
cells from CD38− to CD38+ cells. CN-A altered
the phenotype of CML cells more effectively than ATRA. In a
previous study, CN-A affected the differentiation of leukemic cells
that had been freshly isolated from patients with AML in primary
culture. It significantly stimulated both the functional and
morphological differentiation of leukemia cells in 9 of 12 cases.
Its differentiation-inducing activity was shown to be more potent
than those of ATRA and VD3 (13).
However, CN-A did not induce CD11b or NBT reduction in CML cells,
although it induced CD38 expression. These results indicated that
CN-A induces the differentiation of CML cells to an intermediate
stage. Previous studies have indicated that CN-A and related
compounds inhibit clonogenic activity, cell-surface expression of
cancer stem cell markers and stemness-associated gene expression in
some malignant cells (21–23). CN-A and related compounds have been
shown to significantly inhibit the growth of malignant cells as
xenografts without apparent adverse effects (22,24).
Combined treatment with CN-A and TKIs had a selective cytotoxic
effect on more primitive LSCs at clinically tolerable doses.
A receptor of fusicoccin, which is closely related
to CN-A, has been reported to be a member of a family of 14-3-3
proteins that are commonly found in a wide variety of signaling and
regulatory pathways (25). The
14-3-3 proteins bind to discrete phosphoserine-containing motifs
present in many signaling molecules. The 14-3-3 proteins are
associated with dynamic nucleo-cytoplasmic shuttling. A number of
nuclear proteins can become phosphorylated, bind to 14-3-3 proteins
and accumulate in the cytoplasm. The 14-3-3 proteins negatively
regulate histone deacetylase 4 (26) and cyclin-dependent kinase inhibitor
p27 (27) by preventing their
nuclear localization. A special subfamily of 14-3-3 proteins may
bind CN-A and affect its interaction with some signaling molecules.
This modification may lead to downregulation of the c-Myc gene
(Fig. 4), which is an early gene
that is suppressed by CN-A in CML cells. c-Myc upregulates BCR/ABL
expression and is necessary for BCR/ABL-induced transformation and
LSC maintenance (28). However,
further studies are warranted in order to elucidate the mechanism
through which the downregulation of c-Myc is related to the effects
of CN-A on the 14-3-3 signaling pathway.
Discontinuation trials with second generation TKIs
have shown that 40–50% of patients with CML maintain the treatment
response without continued therapy. However, cardiopulmonary
adverse effects are now impacting treatment choice (29,30).
Therefore, more effective TKI therapy combined with targeted
therapy against pathways involved in CML-LSC survival should be
explored. CML-LSCs have been extensively characterized, with the
aim of developing novel curative approaches based on the
eradication of LSCs. A number of potential molecular targets have
been identified to eradicate LSCs in CML. Several studies have
presented other possible targets to eliminate LSCs in CML therapy
in combination with TKIs. These include interleukin-1 signaling,
hypoxia-inducible factor and mitochondrial oxidative
phosphorylation (31–34). At present, however, no drug is
available for clinical use against these targets. A number of
pathways and mechanisms may promote the survival of LSCs in CML.
Therefore, the dual or triple inhibition of these targets may be
more effective in the treatment of CML with TKIs.
Acknowledgments
Not applicable.
Funding
The present study was supported in part by the
SUIGAN Project, Shimane University, Japan.
Availability of data and materials
The analyzed datasets generated during the study are
available from the corresponding author on reasonable request.
Authors’ contributions
YH, TO and JS designed the study. FI and YH
performed the experiments. YH, TU and JS wrote, edited and revised
the manuscript critically for important intellectual content. FI
and YH analyzed the results. All authors have read and approved the
final manuscript.
Ethics approval and consent to
participate
Leukemic bone marrow specimens were collected at
diagnosis, after the patients provided written informed consent for
sample collection in accordance with institutional policy. The
Shimane University Institutional Committee on Ethics (Shimane,
Japan) approved the present study.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Graham SM, Jørgensen HG, Allan E, Pearson
C, Alcorn MJ, Richmond L and Holyoake TL: Primitive, quiescent,
Philadelphia-positive stem cells from patients with chronic myeloid
leukemia are insensitive to STI571 in vitro. Blood. 99:319–325.
2002. View Article : Google Scholar
|
|
2
|
Mahon FX, Réa D, Guilhot J, Guilhot F,
Huguet F, Nicolini F, Legros L, Charbonnier A, Guerci A, Varet B,
et al Intergroupe Français des Leucémies Myéloïdes Chroniques:
Discontinuation of imatinib in patients with chronic myeloid
leukaemia who have maintained complete molecular remission for at
least 2 years: The prospective, multicentre Stop Imatinib (STIM)
trial. Lancet Oncol. 11:1029–1035. 2010. View Article : Google Scholar
|
|
3
|
Corbin AS, Agarwal A, Loriaux M, Cortes J,
Deininger MW and Druker BJ: Human chronic myeloid leukemia stem
cells are insensitive to imatinib despite inhibition of BCR-ABL
activity. J Clin Invest. 121:396–409. 2011. View Article : Google Scholar
|
|
4
|
Abraham SA, Hopcroft LE, Carrick E, Drotar
ME, Dunn K, Williamson AJ, Korfi K, Baquero P, Park LE, Scott MT,
et al: Dual targeting of p53 and c-MYC selectively eliminates
leukaemic stem cells. Nature. 534:341–346. 2016. View Article : Google Scholar
|
|
5
|
Collins SJ, Bodner A, Ting R and Gallo RC:
Induction of morphological and functional differentiation of human
promyelocytic leukemia cells (HL-60) by componuds which induce
differentiation of murine leukemia cells. Int J Cancer. 25:213–218.
1980. View Article : Google Scholar
|
|
6
|
Honma Y: Cotylenin A - a plant growth
regulator as a differentiation-inducing agent against myeloid
leukemia. Leuk Lymphoma. 43:1169–1178. 2002. View Article : Google Scholar
|
|
7
|
Ishii Y, Kasukabe T and Honma Y: Immediate
up-regulation of the calcium-binding protein S100P and its
involvement in the cytokinin-induced differentiation of human
myeloid leukemia cells. Biochim Biophys Acta. 1745:156–165. 2005.
View Article : Google Scholar
|
|
8
|
Ishii Y, Kasukabe T and Honma Y: Induction
of CCAAT/enhancer binding protein-delta by cytokinins, but not by
retinoic acid, during granulocytic differentiation of human myeloid
leukaemia cells. Br J Haematol. 128:540–547. 2005. View Article : Google Scholar
|
|
9
|
Tsumura H, Akimoto M, Kiyota H, Ishii Y,
Ishikura H and Honma Y: Gene expression profiles in differentiating
leukemia cells induced by methyl jasmonate are similar to those of
cytokinins and methyl jasmonate analogs induce the differentiation
of human leukemia cells in primary culture. Leukemia. 23:753–760.
2009. View Article : Google Scholar
|
|
10
|
Liebermann DA and Hoffman B:
Differentiation primary response genes and proto-oncogenes as
positive and negative regulators of terminal hematopoietic cell
differentiation. Stem Cells. 12:352–369. 1994. View Article : Google Scholar
|
|
11
|
Dimberg A, Bahram F, Karlberg I, Larsson
LG, Nilsson K and Oberg F: Retinoic acid-induced cell cycle arrest
of human myeloid cell lines is associated with sequential
down-regulation of c-Myc and cyclin E and post-transcriptional
up-regulation of p27(Kip1). Blood. 99:2199–2206. 2002. View Article : Google Scholar
|
|
12
|
Hu XT and Zuckerman KS: Role of cell cycle
regulatory molecules in retinoic acid- and vitamin D3-induced
differentiation of acute myeloid leukaemia cells. Cell Prolif.
47:200–210. 2014. View Article : Google Scholar
|
|
13
|
Yamada K, Honma Y, Asahi KI, Sassa T, Hino
KI and Tomoyasu S: Differentiation of human acute myeloid leukaemia
cells in primary culture in response to cotylenin A, a plant growth
regulator. Br J Haematol. 114:814–821. 2001. View Article : Google Scholar
|
|
14
|
Honma Y, Ishii Y, Sassa T and Asahi K:
Treatment of human promyelocytic leukemia in the SCID mouse model
with cotylenin A, an inducer of myelomonocytic differentiation of
leukemia cells. Leuk Res. 27:1019–1025. 2003. View Article : Google Scholar
|
|
15
|
Lozzio BB and Lozzio CB: Properties of the
K562 cell line derived from a patient with chronic myeloid
leukemia. Int J Cancer. 19:1361977. View Article : Google Scholar
|
|
16
|
Kishi K: A new leukemia cell line with
Philadelphia chromosome characterized as basophil precursors. Leuk
Res. 9:381–390. 1985. View Article : Google Scholar
|
|
17
|
Maniwa Y, Kasukabe T and Kumakura S:
Vitamin K2 and cotylenin A synergistically induce monocytic
differentiation and growth arrest along with the suppression of
c-MYC expression and induction of cyclin G2 expression in human
leukemia HL-60 cells. Int J Oncol. 47:473–480. 2015. View Article : Google Scholar
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−ΔΔC(T)) method. Methods. 25:402–408. 2001. View Article : Google Scholar
|
|
19
|
Terstappen LW, Huang S, Safford M,
Lansdorp PM and Loken MR: Sequential generations of hematopoietic
colonies derived from single nonlineage-committed
CD34+CD38− progenitor cells. Blood.
77:1218–1227. 1991.
|
|
20
|
Wang Z, Liu Z, Wu X, Chu S, Wang J, Yuan
H, Roth M, Yuan YC, Bhatia R and Chen W: ATRA-induced cellular
differentiation and CD38 expression inhibits acquisition of BCR-ABL
mutations for CML acquired resistance. PLoS Genet. 10:e10044142014.
View Article : Google Scholar
|
|
21
|
Takahashi T, Honma Y, Miyake T, Adachi K,
Takami S, Okada M, Kumanomidou S, Ikejiri F, Jo Y, Onishi C, et al:
Synergistic combination therapy with cotylenin A and vincristine in
multiple myeloma models. Int J Oncol. 46:1801–1809. 2015.
View Article : Google Scholar
|
|
22
|
Kawakami K, Hattori M, Inoue T, Maruyama
Y, Ohkanda J, Kato N, Tongu M, Yamada T, Akimoto M, Takenaga K, et
al: A novel fusicoccin derivative preferentially targets hypoxic
tumor cells and inhibits tumor growth in xenografts. Anticancer
Agents Med Chem. 12:791–800. 2012. View Article : Google Scholar
|
|
23
|
Miyake T, Honma Y, Urano T, Kato N and
Suzumiya J: Combined treatment with tamoxifen and a fusicoccin
derivative (ISIR-042) to overcome resistance to therapy and to
enhance the antitumor activity of 5-fluorouracil and gemcitabine in
pancreatic cancer cells. Int J Oncol. 47:315–324. 2015. View Article : Google Scholar
|
|
24
|
Honma Y, Ishii Y, Yamamoto-Yamaguchi Y,
Sassa T and Asahi K: Cotylenin A, a differentiation-inducing agent,
and IFN-alpha cooperatively induce apoptosis and have an antitumor
effect on human non-small cell lung carcinoma cells in nude mice.
Cancer Res. 63:3659–3666. 2003.
|
|
25
|
Oecking C, Eckerskorn C and Weiler EW: The
fusicoccin receptor of plants is a member of the 14-3-3 superfamily
of eukaryotic regulatory proteins. FEBS Lett. 352:163–166. 1994.
View Article : Google Scholar
|
|
26
|
Rittinger K, Budman J, Xu J, Volinia S,
Cantley LC, Smerdon SJ, Gamblin SJ and Yaffe MB: Structural
analysis of 14-3-3 phosphopeptide complexes identifies a dual role
for the nuclear export signal of 14-3-3 in ligand binding. Mol
Cell. 4:153–166. 1999. View Article : Google Scholar
|
|
27
|
Fujita N, Sato S and Tsuruo T:
Phosphorylation of p27Kip1 at threonine 198 by p90 ribosomal
protein S6 kinases promotes its binding to 14-3-3 and cytoplasmic
localization. J Biol Chem. 278:49254–49260. 2003. View Article : Google Scholar
|
|
28
|
el-Deiry WS, Tokino T, Velculescu VE, Levy
DB, Parsons R, Trent JM, Lin D, Mercer WE, Kinzler KW and
Vogelstein B: WAF1, a potential mediator of p53 tumor suppression.
Cell. 75:817–825. 1993. View Article : Google Scholar
|
|
29
|
Moslehi JJ and Deininger M: Tyrosine
kinase inhibitor-associated cardiovascular toxicity in chronic
myeloid leukemia. J Clin Oncol. 33:4210–4218. 2015. View Article : Google Scholar
|
|
30
|
Caldemeyer L, Dugan M, Edwards J and Akard
L: Long-term side effects of tyrosine kinase inhibitors in chronic
myeloid leukemia. Curr Hematol Malig Rep. 11:71–79. 2016.
View Article : Google Scholar
|
|
31
|
Ågerstam H, Hansen N, von Palffy S, Sandén
C, Reckzeh K, Karlsson C, Lilljebjörn H, Landberg N, Askmyr M,
Högberg C, et al: IL1RAP antibodies block IL-1-induced expansion of
candidate CML stem cells and mediate cell killing in xenograft
models. Blood. 128:2683–2693. 2016. View Article : Google Scholar
|
|
32
|
Zhang B, Chu S, Agarwal P, Campbell VL,
Hopcroft L, Jørgensen HG, Lin A, Gaal K, Holyoake TL and Bhatia R:
Inhibition of interleukin-1 signaling enhances elimination of
tyrosine kinase inhibitor-treated CML stem cells. Blood.
128:2671–2682. 2016. View Article : Google Scholar
|
|
33
|
Cheloni G, Tanturli M, Tusa I, Ho DeSouza
N, Shan Y, Gozzini A, Mazurier F, Rovida E, Li S and Dello Sbarba
P: Targeting chronic myeloid leukemia stem cells with the
hypoxia-inducible factor inhibitor acriflavine. Blood. 130:655–665.
2017. View Article : Google Scholar
|
|
34
|
Kuntz EM, Baquero P, Michie AM, Dunn K,
Tardito S, Holyoake TL, Helgason GV and Gottlieb E: Targeting
mitochondrial oxidative phosphorylation eradicates
therapy-resistant chronic myeloid leukemia stem cells. Nat Med.
23:1234–1240. 2017. View
Article : Google Scholar
|