Introduction
Hepatocellular carcinoma (HCC) is one of the most
common malignancies and is the second leading cause of
cancer-related mortalities worldwide (1–3). HCC
prognosis remains poor because tumors are frequently diagnosed at a
late stage that may not treatable by surgical resection (4). Owing to the high rates of metastasis
and recurrence following curative resection, understanding the
underlying mechanisms of tumor metastasis is crucial for the
development of effective therapeutic targets against HCC that may
be used as new targets for treatment and as potential diagnostic
markers (5). Membrane proteins are
abundant biomolecules, a number of which have been used as tumor
markers or drug targets; approximately one-third of protein tumor
markers are membrane proteins (6).
Membrane proteins serve an important role in cell survival, growth,
division and differentiation (7,8).
Signal transduction, material transport, adhesion and
immunogenicity of cells may be altered by membrane proteins, such
as receptor proteins, transporter proteins, adhesion protein
expression and antigens (6,9). It
has been reported previously that membrane proteins are closely
related to malignant tumor cell invasion and metastasis, such as
epithelial membrane protein 1 and lysosome-associated membrane
protein1 (10–12). Therefore, the present study used a
proteomics method to identify differentially expressed membrane
proteins in two metastatic HCC cell lines. The profiling of
differentially expressed proteins in different tumoral states is
essential to understanding the mechanisms of tumor progression, and
may aid future studies in identifying new therapeutic targets.
Advancements in proteomics have permitted the
quantitative detection and accurate monitoring of changes in
protein expression levels (13).
Isobaric tags for relative and absolute quantitation (iTRAQ)
technology is a powerful tool and is used for its selectivity,
sensitivity and specificity to identify proteins that are
differentially expressed in tumors (14). In the present study, iTRAQ was used
to identify differentially expressed membrane proteins between two
human HCC cell lines with differing metastatic potentials. The
lowly metastatic MHCC97-L and the highly metastatic MHCC97-H cell
lines were established from the metastatic hepatocellular carcinoma
cell line MHCC97 by Li et al (15). These two cell lines have been
widely used to investigate the mechanisms of human HCC metastasis,
to identify the membrane proteins that may be associated with HCC
cell metastasis and to provide clues for early diagnosis and
treatment (16–19).
Materials and methods
Cell lines and culture conditions
The human HCC cell lines MHCC97-Hand MHCC97-L were
purchased from the Liver Cancer Institute of Fudan University
(Shanghai, China). Cells were cultured in complete Dulbecco's
modified Eagle's medium (DMEM; HyClone; GE Healthcare Life
Sciences, Logan, UT, USA) containing 10% fetal bovine serum (FBS;
Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) and 100
U/ml penicillin (Invitrogen; Thermo Fisher Scientific, Inc.), and
incubated at 37°C in a humidified atmosphere containing 5% carbon
dioxide.
Cell membrane protein collection
Membrane proteins were extracted using a CelLytic
MEM Protein Extraction kit (Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany), following the manufacturer's protocol. A total between
1×106 and 1×107 cells were collected and
resuspended in 600 µl ice-cold Lysis and Separation Working
Solution, provided in the kit, containing 6 µl protease
inhibitor cocktail. Cell suspensions were incubated on ice for 10
min and subsequently centrifuged in a pre-cooled (4°C)
microcentrifuge at 10,000 × g for 5 min. The clarified lysates were
transferred to a sterile microcentrifuge tube, incubated at 30°C
for 3–5 min and centrifuged at room temperature (>20°C) at 3,000
× g for 3 min. The lower, hydrophobic phase, which is enriched with
hydrophobic and lipid raft-associated proteins, was transferred to
a new microcentrifuge tube and stored at −20°C until use.
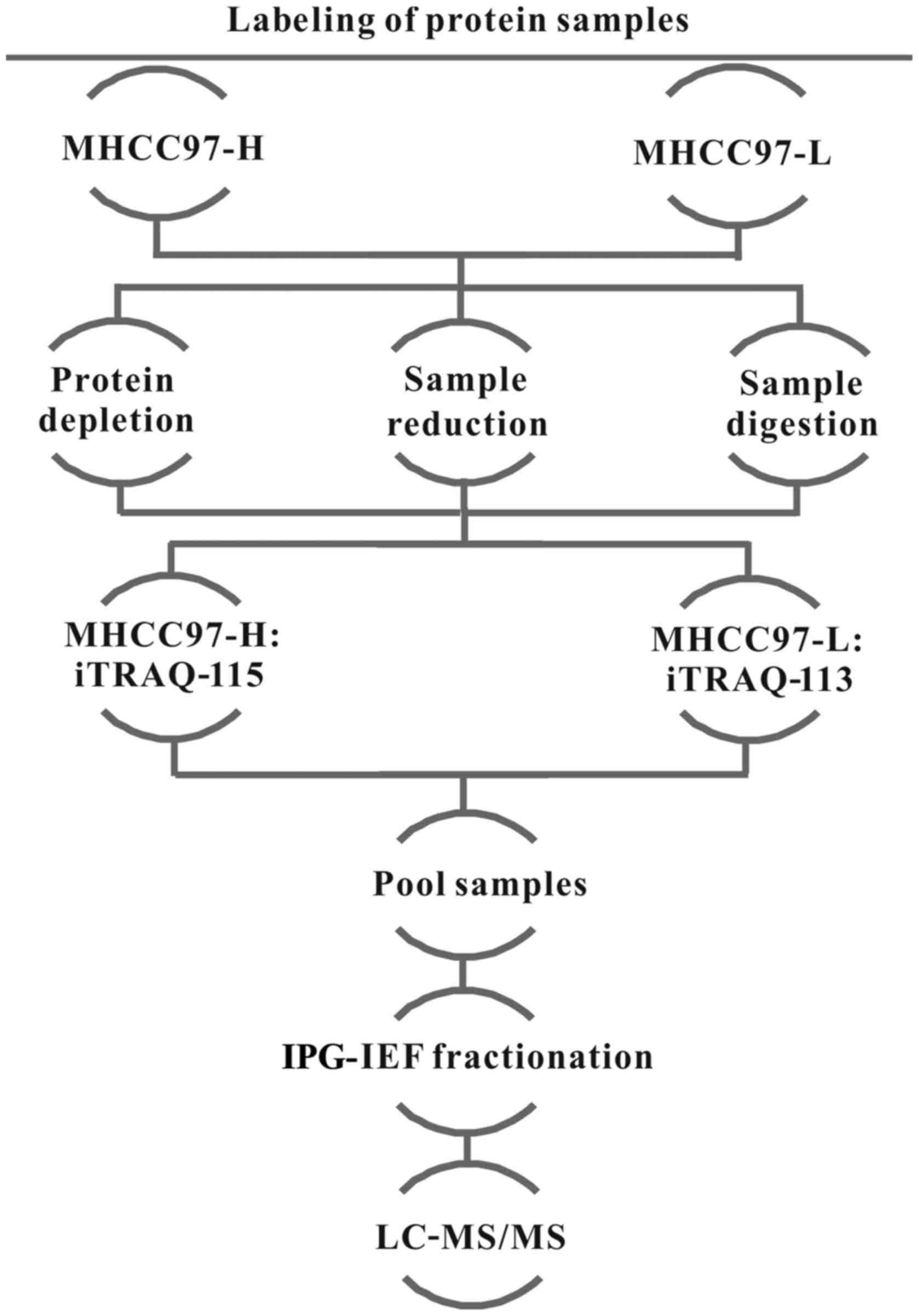
iTRAQ labeling
The 8-plex iTRAQ kits were obtained from Applied
Biosystems (Thermo Fisher Scientific, Inc.), and experiments were
conducted according to the manufacturer's protocol. Briefly,
protein concentrations were quantified by Bicinchoninic Acid assay
(BCA; Qcbio Science & Technologies Co., Ltd., Shanghai, China)
and ~100 µg of prepared proteins from each group were
precipitated, dissolved in dissolution buffer, denatured, cysteine
blocked, digested with 2 µg of sequencing grade modified
trypsin, and labeled with the iTRAQ tags; MHCC97-L proteins were
labeled with the iTRAQ-113 tag and MHCC97-H proteins were labeled
with the iTRAQ-115 tags. A schematic view of the iTRAQ methodology
is shown in Fig. 1; the
experiments were repeated twice.
Peptide fractionation
iTRAQ-labeled peptides were fractionated by
immobilized pH gradient-isoelectric focusing (IPG-IEF). Briefly,
the pooled iTRAQ-labeled peptide samples were solubilized in 300
µl of 8 M urea and 1% Pharmalyte solution, and rehydrated on
an 18 cm-long IPG gel strips (pH 3–10) (both from Amersham; GE
Healthcare Life Sciences, Little Chalfont, UK) for 14 h at 30 V.
The peptides were subjected to IEF at 68 kV/h with an IPGphor IEF
System (Amersham; GE Healthcare Life Sciences). Peptides were
extracted by incubating the gel fractions in 2% acetonitrile and
0.1% formic acid for 1 h at room temperature. Fractions were
purified and concentrated on a C-18 Discovery DSC-18 SPE column
(Sigma-Aldrich; Merck KGaA). The desalted fractions were
lyophilized and stored at −20°C until used in liquid
chromatograph-tandem mass spectrometry (LC-MS/MS) analysis.
LC-MS/MS and bioinformatics
Mass spectrometric analysis was performed on a QStar
Elite mass spectrometer (Applied Biosystems; Thermo Fisher
Scientific, Inc.) coupled to an online capillary LC Dionex Ultimate
3000 system (Thermo Fisher Scientific, Inc.), as previously
described (20,21). The peptide separation was performed
on a PepMap C-18RP capillary column (Thermo Fisher Scientific,
Inc.) with a flow rate of 0.3 µl/min using a linear gradient
ranging between 2 and 100% Buffer B (0.1% formic acid; 98%
acetonitrile) for 90 min, 2.1 kV, and flow rate of 300 nl/min. Data
acquisition was performed in the positive ionization mode with a
selected mass range of 300–1,800 m/z. The two most intensely
charged peptides with >20 counts were chosen for MS/MS with a
dynamic exclusion of 30 sec (22).
Protein identification and quantification were
performed using ProteinPilot software v2.0 (Applied Biosystems/MDS
Sciex Joint Venture; Thermo Fisher Scientific, Inc.; Nordion, Inc.,
Ottawa, ON, Canada). MS/MS data were analyzed with the UniProt
database (http://www.uniprot.org). Differentially
expressed proteins were indicated as having a protein score >1.3
(1.3/1) or <0.77 (1/1.3) (Table
I), and >2 unique peptides with 95% confidence and P<0.05
were required for quantitation.
 | Table IList of differentially expressed
proteins between MHCC97-H and MHCC97-L cell lines identified by
iTRAQ proteomics analysis. |
Table I
List of differentially expressed
proteins between MHCC97-H and MHCC97-L cell lines identified by
iTRAQ proteomics analysis.
| n | Unused | Accession no. | Gene | Protein name | Peptides (95%
CI) | iTRAQ ratios
(115:113) | P-value
(115:113) |
|---|
| Downregulated
proteins |
| 1 | 6.2 |
sp|P68402|PA1B2_HUMAN | PAFAH1B2 | Platelet-activating
factor acetylhydrolase IB subunit β | 3 | 0.5688 | 0.0205 |
| 2 | 13 |
sp|Q9UHL4|DPP2_HUMAN | DPP2 | Dipeptidyl
peptidase 2 | 7 | 0.6872 | 0.0380 |
| 3 | 4.15 |
sp|Q15386|UBE3C_HUMAN | UBE3C | Ubiquitin-protein
ligase E3C | 6 | 0.7098 | 0.0189 |
| 4 | 4.76 |
sp|Q9Y263|PLAP_HUMAN | PLAA | Phospholipase
A-2-activating protein | 7 | 0.7228 | 0.0083 |
| 5 | 23.38 |
tr|Q53FG3|Q53FG3_HUMAN | IFL2 | Interleukin
enhancer binding factor 2 variant | 20 | 0.7278 | 0.0004 |
| 6 | 6.97 |
sp|Q9NV70|EXOC1_HUMAN | EXOC1 | Exocyst complex
component 1 | 4 | 0.7508 | 0.0435 |
| Upregulated
proteins |
| 1 | 37.28 |
sp|Q9ULC5|ACSL5_HUMAN | ACSL5 |
Long-chain-fatty-acid-CoA ligase 5 | 20 | 13.068 | 0.0001 |
| 2 | 54.86 |
sp|P05556|ITB1_HUMAN | ITGB1 | Integrin β-1 | 40 | 13.416 | 10.282 |
| 3 | 20.07 |
sp|P63092|GNAS2_HUMAN | GNAS | Guanine
nucleotide-binding protein G(s) subunit α isoforms short | 14 | 13.416 | 0.0003 |
| 4 | 9.2 |
tr|J3KNF8|J3KNF8_HUMAN | CYB5B | Cytochrome b5 type
B | 13 | 13.451 | 0.0358 |
| 5 | 9.48 |
tr|J3KQ45|J3KQ45_HUMAN | TGOLN2 | Trans-Golgi network
integral membrane protein 2 | 5 | 13.479 | 0.0405 |
| 6 | 5.96 |
sp|Q99871|HAUS7_HUMAN | HAUS7 | HAUS augmin-like
complex subunit 7 | 5 | 13.568 | 0.0277 |
| 7 | 10.91 |
sp|P16422|EPCAM_HUMAN | EPCAM | Epithelial cell
adhesion molecule | 13 | 13.820 | 0.0021 |
| 8 | 282.04 |
sp|Q09666|AHNK_HUMAN | AHNAK | Neuroblast
differentiation-associated protein | 229 | 13.860 | 85.454 |
| 9 | 12.65 |
sp|Q92542|NICA_HUMAN | NCSTN | Nicastrin | 6 | 13.999 | 0.0032 |
| 10 | 45.04 |
tr|F5GZS6|F5GZS6_HUMAN | SLC3A2 | 4F2 cell-surface
antigen heavy chain | 43 | 14.267 | 38.195 |
| 11 | 6.56 |
sp|Q01650|LAT1_HUMAN | SLC7A5 | Large neutral amino
acids transporter small subunit 1 | 7 | 14.293 | 0.0467 |
| 12 | 2.48 |
tr|A6NNI4|A6NNI4_HUMAN | CD9 | Tetraspanin-29 | 3 | 14.478 | 0.0160 |
| 13 | 10.07 |
sp|Q9H3R2|MUC13_HUMAN | MUC13 | Mucin-13 | 10 | 14.557 | 0.0146 |
| 14 | 42.19 |
sp|Q8IVT2|MISP_HUMAN | MISP | Mitotic interactor
and substrate of PLK1 | 26 | 14.639 | 16.779 |
| 15 | 12.61 |
tr|J3KQL8|J3KQL8_HUMAN | APOL2 | Apolipoprotein
L2 | 8 | 15.155 | 0.0184 |
| 16 | 4.23 |
tr|Q53GA9|Q53GA9_HUMAN | CD63 | Tetraspanin-30 | 3 | 17.029 | 0.0457 |
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was isolated from
1×105–1×107 cells using TRIzol (Invitrogen;
Thermo Fisher Scientific, Inc.), according to the manufacturer's
instructions. First strand cDNA was synthesized using 2 µg
total RNA and the A3500 Reverse Transcription System (Promega,
Madison, WI, USA). The resulting cDNA was amplified by PCR using an
ABI 7300 system (Applied Biosystems; Thermo Fisher Scientific,
Inc.), SYBR Premix Ex Taq II (Tli RNaseH Plus) and ROX plus
reference dye (Takara Bio, Inc., Otsu, Japan). Gene-specific primer
sequences are provided in Table
II. PCR thermocycling conditions were as follows: 95°C for 15
sec followed by 35 cycles of 72°C for 30 sec and 60°C for 60 sec.
PCR reactions were evaluated by melting curve analysis, and
relative gene expression levels were calculated according to the
2−ΔΔCq method and normalized to the internal reference
gene GAPDH (23). Experiments were
performed at least three times and similar results were obtained in
each experiment.
| Table IIGene-specific primer sequences used
for polymerase chain reaction. |
Table II
Gene-specific primer sequences used
for polymerase chain reaction.
| Gene (accession
no.) | Primer sequence
(5′→3′) |
|---|
| PAFAH1B2
(NM_002572) | F:
ATCAACACAAGGCAGCCACAGG |
| R:
CAGCGAAACCTTGAGGAGTTGG |
| DPP7
(NM_013379) | F:
TTCGCCAGCAACAATGTGACCG |
| R:
ATGTTGCTGGCGGCTCTGAGAT |
| UBE3C
(NM_014671) | F:
GCTCTATGTGCCAGCATCCAGA |
| R:
AGGCAACTCTGTCAGGACAGCA |
| PLAA
(NM_004253) | F:
AGGCTGGAAGATGTGAGAGGAC |
| R:
TCGCCAGTGATTTGCCACCTTC |
| ILF2
(NM_004515) | F:
AGGACTGTTCCTGCCAGGTTCA |
| R:
GAGGATTCGGACGAGAGTCTGA |
| EXOC1
(NM_018261) | F:
CCTATGTCTGGCAGAACAGGAC |
| R:
GCCGTGATAATGTTCCTCCATCC |
| ACSL5
(NM_016234) | F:
GCTTATGAGCCCACTCCTGATG |
| R:
GGAAGAATCCAACTCTGGCTCC |
| ITGB1
(NM_033667) | F:
GGATTCTCCAGAAGGTGGTTTCG |
| R:
TGCCACCAAGTTTCCCATCTCC |
| GNAS
(NM_001077490) | F:
GCAGACAGATGCGCAAAGAAGC |
| R:
GCTTTTACCAGATTCTCCAGCAC |
| CYB5B
(NM_030579) | F:
AGGTGGCAAAGCGCAACTCCTT |
| R:
GTTCCAGCAGAACCTCTTCTCC |
| TGOLN2
(NM_006464) | F:
GGAGAGCAGCCACTTCTTTGCA |
| R:
CCAAACGTTGGTAGTCACTGGC |
| HAUS7
(NM_207107) | F:
CCTTGGCTCAAGGATTCCGTGA |
| R:
CAGAGGTGTCAGCAACTGCCAT |
| EPCAM
(NM_002354) | F:
GCCAGTGTACTTCAGTTGGTGC |
| R:
CCCTTCAGGTTTTGCTCTTCTCC |
| AHNAK
(NM_001620) | F:
CGTGAAGTCTTCAGCTCCTGCA |
| R:
GAGGTCTCCTTCCACTCCATCT |
| NCSTN
(NM_015331) | F:
GGAGGAACCAACTTCAGCGACA |
| R:
TGCCTGAGGATAGACTGGAACC |
| SLC3A2
(NM_002394) | F:
CCAGAAGGATGATGTCGCTCAG |
| R:
GAGTAAGGTCCAGAATGACACGG |
| SLC7A5
(NM_003486) | F:
GCCACAGAAAGCCTGAGCTTGA |
| R:
ATGGTGAAGCCGATGCCACACT |
| CD9
(NM_001769) | F:
TCGCCATTGAAATAGCTGCGGC |
| R:
CGCATAGTGGATGGCTTTCAGC |
| MUC13
(NM_033049) | F:
TGGCTGTAACCAGACTGCGGAT |
| R:
GCATCAGGACACTTGAGACTGG |
| MISP
(NM_173481) | F:
CACCTACACTCAAACGTGGCGT |
| R:
CCTCTGAGTTGATACCGTCCGA |
| APOL2
(NM_030882) | F:
GAAGCCTGGAATGGATTCGTGG |
| R:
CGTGGCGGTTTTTGTCCTTCATG |
| CD63
(NM_001780) | F:
CAACCACACTGCTTCGATCCTG |
| R:
GACTCGGTTCTTCGACATGGAAG |
Western blotting
Cells were lysed and fractionated using a CelLytic
MEM Protein Extraction Kit (Sigma-Aldrich; Merck KGaA), as
aforementioned. Total protein was quantified by BCA, and equal
amounts of proteins (30 µg protein/lane) were separated by
10% SDS-PAGE and transferred to a polyvinylidene fluoride membrane
(EMD Millipore, Billerica, MA, USA). Membranes were blocked for 1 h
at room temperature in 5% skim milk powder and TBS + 0.5% Tween-20
buffer (pH 7.6) and incubated with primary antibodies at 4°C
overnight. Primary antibodies against PAFAH1B2 (1:1,000; cat. no.
ab157479), DPP2 (1:1,000; cat. no. ab226935), UBE3C (1:1,000; cat.
no. ab226173), PLAA (1:1,000; cat. no. ab32354), ILF2 (1:1,000;
cat. no. ab154169), EXOC1 (1:1,000; cat. no. ab118798), ACSL5
(1:500; cat. no. ab104892), GNAS (1:1,000; cat. no. ab83735),
TGOLN2 (1:500; cat. no. ab2809), HAUS7 (1:500; cat. no. ab192616),
EPCAM (1:1,000; cat. no. ab71916), AHNAK (1:1,000; cat. no.
ab168104), NCSTN (1:500; cat. no. ab45425), SLC3A2 (1:1,000; cat.
no. ab108300), SLC7A5 (1:500; cat. no. ab85226), CD9 (1:1,000; cat.
no. ab92726), MUC13 (1:1,000; cat. no. ab124654), MISP (1:200; cat.
no. ab175101), APOL2 (1:500; cat. no. ab196771), CD63 (1:1,000;
cat. no. ab59479) and ATPase Na+/K+
transporting subunitβ2 (ATP1B2; 1:1,000; cat. no. ab176163) were
purchased from Abcam (Cambridge, MA, USA). ITGB1 (1:1,000; cat. no.
4706) was purchased from Cell Signaling Technology, Inc. (Danvers,
MA, USA) and CYB5B (1:250; cat. no. PA5-52482) was from Invitrogen
(Thermo Fisher Scientific, Inc.). Subsequently, membranes were
incubated with horseradish peroxidase (HRP)-conjugated goat
anti-rabbit (1:10,000; cat. no. ab205718) or goat anti-mouse
(1:10,000; cat. no. ab6789) secondary antibodies (Abcam) for 1 h at
room temperature, and protein bands were visualized with the
Immobilon Western Chemiluminescent HRP substrate (Merck KGaA) on a
ChemiDoc MP imaging system (Bio-Rad Laboratories, Inc., Hercules,
CA, USA). Target protein expression levels were analyzed by
densitometric analysis with Image Lab v4.0 (Bio-Rad Laboratories,
Inc.) and normalized to ATP1B2. Experiments were repeated at least
three times.
CD9 small interfering (si)RNA
transfection
CD9-targeted siRNA oligomer duplexes siRNA1 (cat.
no. 10404) and siRNA2 (cat. no. 145966) and a non-specific,
scrambled-siRNA was used as negative control (cat. no. 4390843)
were purchased from Invitrogen (Thermo Fisher Scientific, Inc.).
MHCC97-H cells (6×105 cells/well) were seeded in 6-well
plates 24 h at 37°C prior to transfection. Cells were transfected
for 6 h at 5% CO2 and 37°C with scrambled-siRNA,
CD9-siRNA1 or CD9-siRNA2 using Lipofectamine® 2000
Transfection Reagent (Invitrogen; Thermo Fisher Scientific, Inc.)
and Opti-MEM (Gibco; Thermo Fisher Scientific, Inc.); untreated
MHCC97-H cells were used as a blank control. Subsequently, the
transfection culture medium was removed and cells were further
incubated with complete culture solution for 48 hat 5%
CO2 and 37°C. Transfected cells were harvested and the
expression levels of CD9 mRNA and protein in each sample were
confirmed by RT-qPCR and western blotting, respectively. All
experiments were repeated at least three times.
Wound-healing migration assay
The wound-healing assay was conducted following
siRNA transfections to determine the effects on the migratory
ability of cells. MHCC97-H cells were seeded in 6-well plates until
they reached 100% confluence at 48 h post-transfection. A sterile
P20 pipette tip was used to scratch a wound into the cell monolayer
and the debris was removed by washing thrice with PBS. The cells
were grown in serum-free medium at 37°C, and digital images of gap
closure were captured under a Nikon TS100/TS100-F inverted light
microscope (Nikon Corporation, Tokyo, Japan) at 0, 24 and 48 h.
Experiments were repeated at least three times.
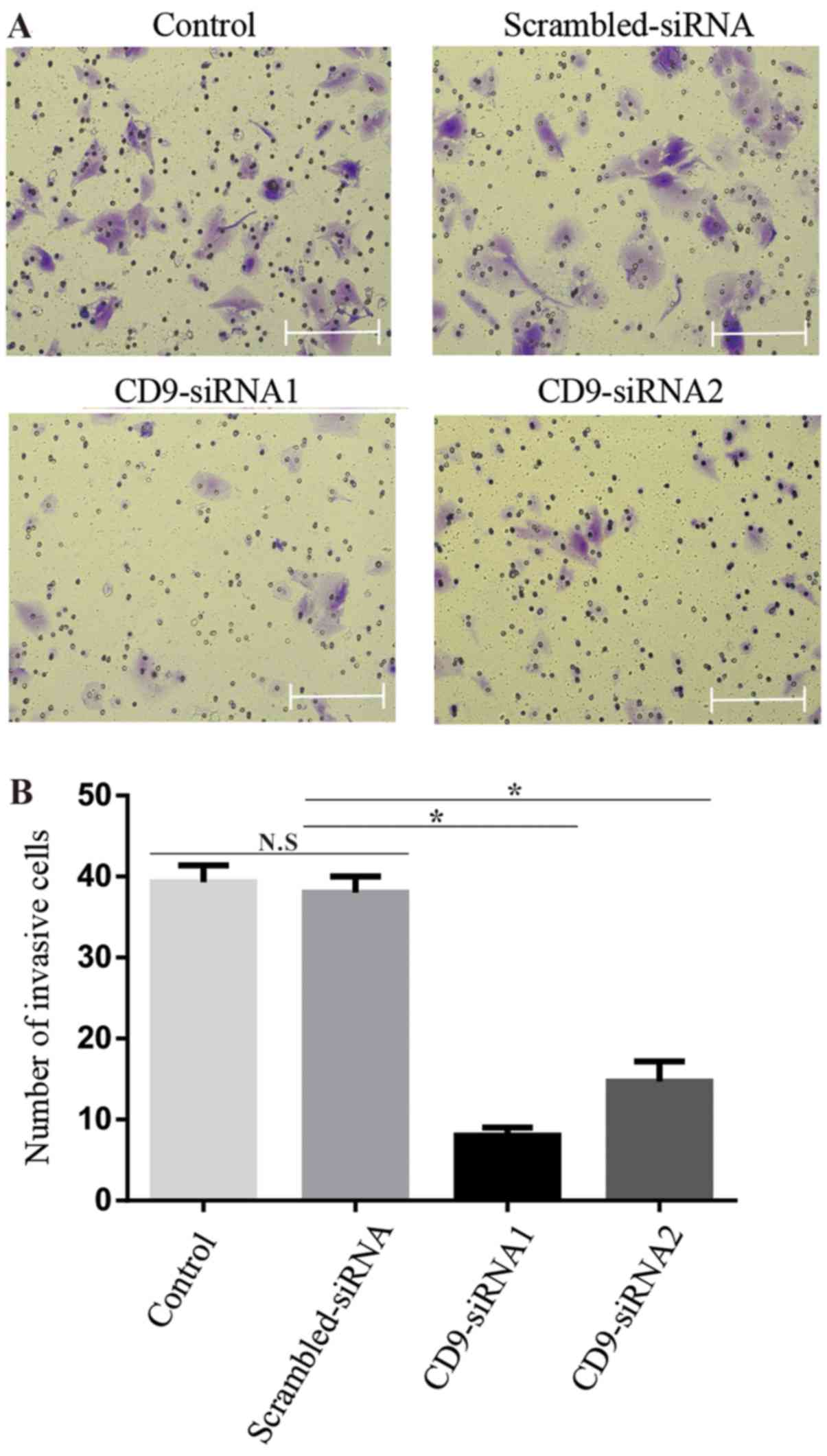
Invasion assay
Invasion assays were conducted using a Cell Invasion
Assay kit (Cell Biolabs, Inc., San Diego, CA, USA). Viable
CD9-siRNA1/2 or scrambled-siRNA transfected MHCC97-H cells
(1×105 cells) were harvested and re-suspended in
serum-free medium, and used to seed the upper chamber of a 24-well
plate; membranes were precoated with Matrigel. A total of 500
µl DMEM + 10% FBS was loaded into the lower chamber. Cells
were incubated for 24 h at 37°C. Subsequently, cells were washed
with PBS and the cells in the upper chamber were wiped off with
cotton swab. Cells in the lower chamber were fixed with 4%
formaldehyde at room temperature for 30 min and stained with 0.1%
crystal violet (Solarbio, Beijing, China) for 20 min at room
temperature. Following three washes with PBS, the number of cells
that invaded through the Matrigel was measured by Image-Pro Plus
6.0. At least three fields of views were randomly selected to count
cells under an inverted light microscope (magnification, ×200).
Experiments were repeated at least three times.
Statistical analysis
Statistical analyses were performed using SPSS
software v16.0 (SPSS Inc., Chicago, IL, USA). Quantitative
variables were presented as the mean ± standard deviation.
Differences between two groups were analyzed by Student's t-test;
multiple comparison between the groups were performed by analysis
of variance followed by Student-Newman-Keuls post hoc test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
iTRAQ comparative proteome analysis
To investigate the mechanisms involved in HCC
metastasis, iTRAQ-based MS was conducted to analyze the proteins
that were isolated from MHCC97-H and MHCC97-L cell membranes. The
ratio of 115-tagged protein expression to 113-tagged protein
expressions in MHCC97-H and MHCC97-L samples was used to determine
differentially expressed proteins. A total of 780 unique proteins
were identified and quantified in each cell line, regardless of
whether the P-value was <0.05 in the iTRAQ ratios. It has been
reported previously that membrane proteins are closely related to
malignant tumor cell invasion and metastasis, thus the membrane
proteins were classified separately in 780 differentially expressed
proteins: ~30 membrane proteins as being differentially expressed
between MHCC97-L and MHCC97-H cells. Based on the 'Unused' column
in Table I (that is, the score of
protein confidence level), ~20 membrane proteins were verified by
RT-qPCR and western blotting, which demonstrated similar mRNA and
protein expression levels (Fig.
2). Therefore, 22 proteins were analyzed in this experiment. Of
these 22 differentially expressed membrane proteins, 16 were
upregulated and 6 were downregulated in the MHCC97-H cell line
compared with MHCC97-L cells (Table
I).
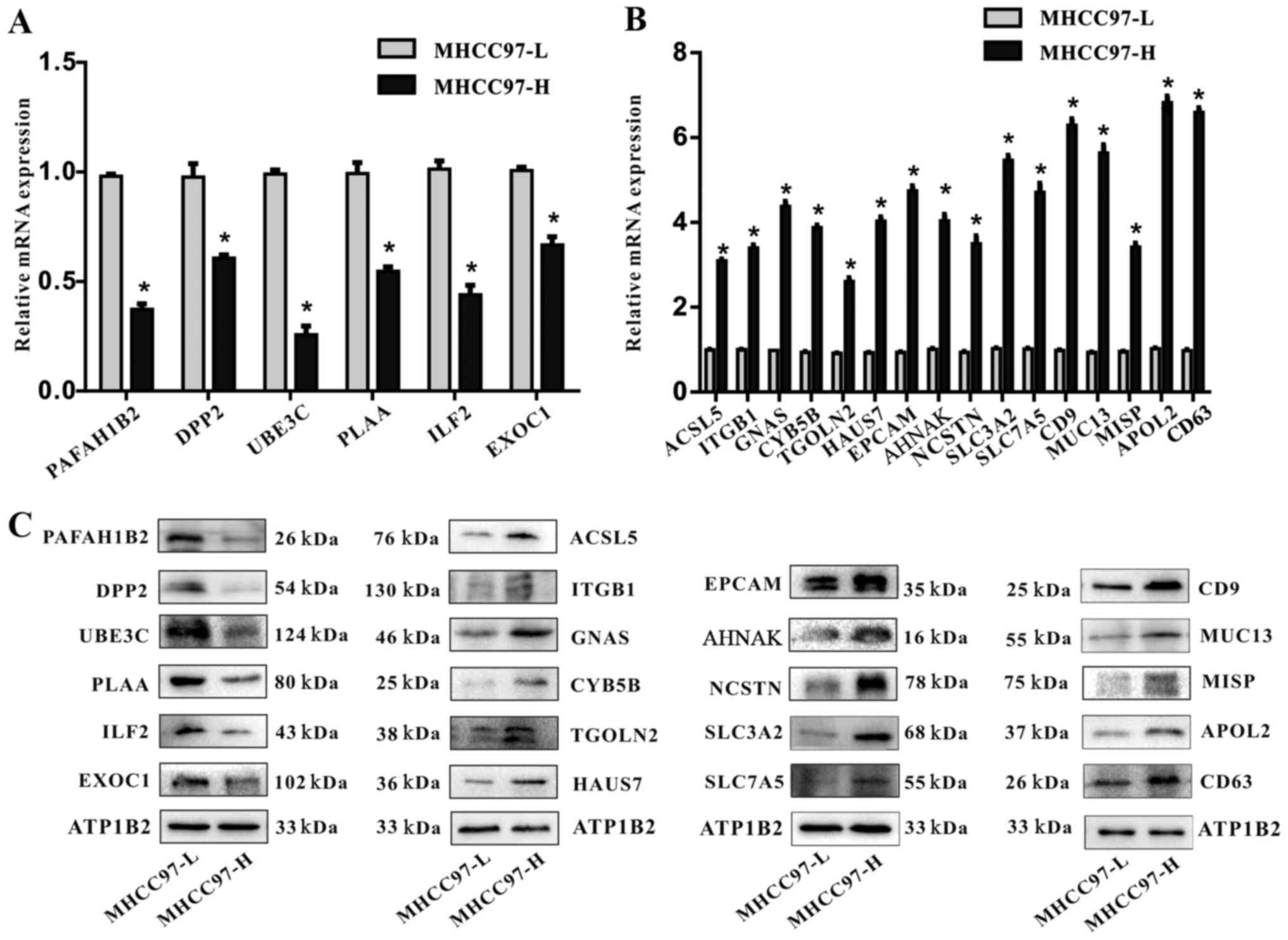 | Figure 2Verification of the differential
expression of mRNA and proteins identified by iTRAQ proteomics
analysis. (A) RT-qPCR detection of the relative mRNA expression
levels of PAFAH1B2, DPP2, UBE3C, PLAA, ILF2 and EXOC1 that were
identified by iTRAQ as downregulated in MHCC97-H cells compared
with MHCC97-L cells; expression levels were normalized to GADPH.
(B) RT-qPCR detection of the relative mRNA expression levels of
ACSL5, ITGB1, GNAS, CYB5B, TGOLN2, HAUS7, EPCAM, AHNAK, NCSTN,
SLC3A2, SLC7A5, CD9, MUC13, MISP, APOL2 and CD63 that were
identified by iTRAQ as upregulated in MHCC97-H cells compared with
MHCC97-L cells; expression levels were normalized to GADPH
(*P<0.05). (C) Representative western blot analysis
of PAFAH1B2, DPP2, UBE3C, PLAA, ILF2, EXOC1, ACSL5, ITGB1, GNAS,
CYB5B, TGOLN2, HAUS7, EPCAM, AHNAK, NCSTN, SLC3A2, SLC7A5, CD9,
MUC13, MISP, APOL2 and CD63 expression levels in MHCC97-H and
MHCC97-L cells; ATP1B2 was used as a loading control. ACSL5,
acyl-CoA synthetase long chain family member 5; AHNAK, AHNAK
nucleoprotein; APOL2, apolipoprotein L2; ATP1B2, ATPase
Na+/K+ transporting subunit β2; CYB5B,
cytochrome B5 type B; DPP2, dipeptidyl peptidase 2; EPCAM,
epithelial cell adhesion molecule; EXOC1, exocyst complex component
1; GNAS, GNAS complex locus; HAUS7, HAUS augmin-like complex
subunit 7; ILF2, interleukin enhancer-binding factor 2; ITGB1,
integrin subunit β1; iTRAQ, isobaric tags for relative and absolute
quantitation; MISP, mitotic spindle positioning; MUC13, mucin 13;
NCSTN, nicastrin; PAFAH1B2, platelet activating factor
acetylhydrolase 1B catalytic subunit 2; PLAA, phospholipase A2
activating protein; RT-qPCR, reverse transcription-quantitative
polymerase chain reaction; SLC3A2, solute carrier family 3 member
2; TGOLN2, trans-Golgi network protein 2; UBE3C, ubiquitin protein
ligase E3C. |
Validation of differentially expressed
proteins
To verify the iTRAQ results, RT-qPCR and western
blot analyses were performed. RT-qPCR analysis demonstrated that,
compared with expression levels in MHCC97-L cells, the mRNA levels
of PAFAH1B2, DPP2, UBE3C, PLAA, IFL2 and EXOC1 were significantly
downregulated in MHCC97-H cells (Fig.
2A), whereas ACSL5, ITGB1, GNAS, CYB5B, TGOLN2, HAUS7, EPCAM,
AHNAK, NCSTN, SLC3A2, SLC7A5, CD9, MUC13, MISP, APOL2 and CD63 mRNA
expression levels were upregulated (Fig. 2B). Western blot analyses were
performed to determine the expression levels of these membrane
proteins, which revealed a similar pattern of down- and upregulated
protein expression levels, as was identified for mRNA in the
MHCC97-H cells compared with the MHCC97-L cell (Fig. 2C). The results of mRNA and protein
expression were consistent with the iTRAQ results.
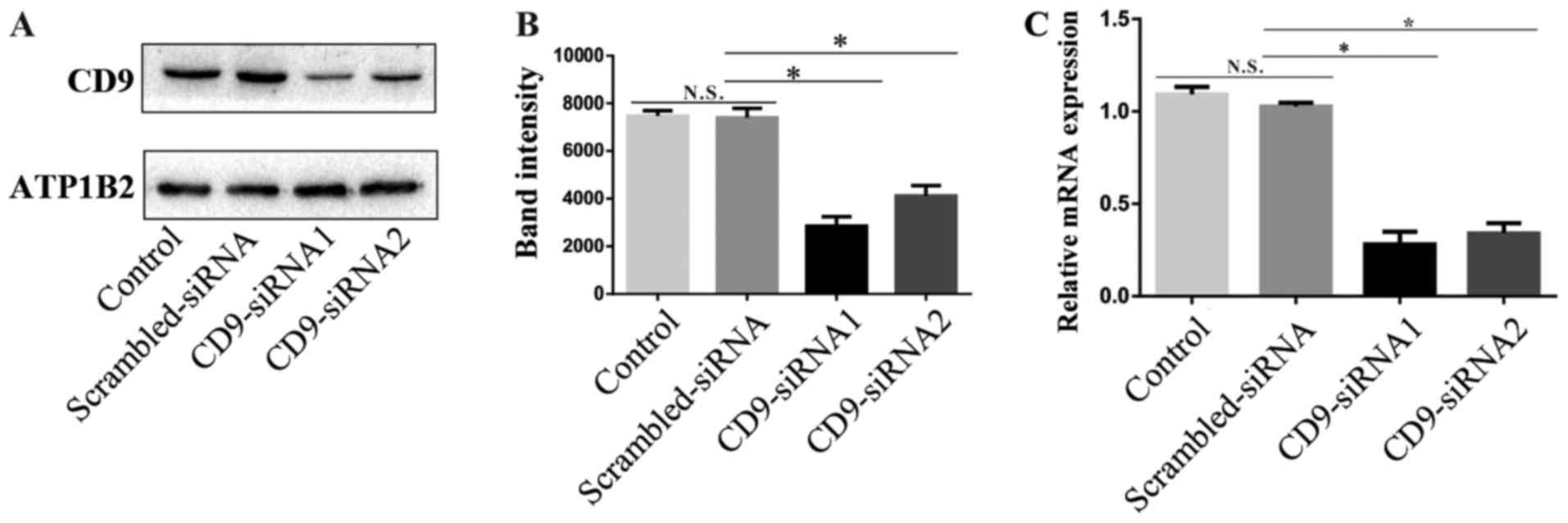
CD9 expression is reduced by siRNA
treatment
The tetraspanin membrane protein CD9 was
demonstrated to be highly expressed in MHCC97-H cells (Fig. 2). CD9 had the lowest score in the
'Unused' column among 22 membrane proteins listed in Table I. The score indicated that CD9 was
less accurate compared with other proteins and, therefore, was used
to verify the accuracy of iTRAQ. CD9-targeted siRNA1 and siRNA2
were transfected into MHCC97-H cells to silence the expression of
CD9. Subsequent western blot analysis demonstrated that CD9 protein
expression was reduced in the CD9-siRNA-treated cells compared with
the untreated control and scrambled-siRNA-treated groups (Fig. 3A and B). A significant decrease of
CD9 mRNA expression levels was also observed intheCD9-siRNA-treated
MHCC97-H groups compared with the untreated control and
scrambled-siRNA-treated groups (Fig.
3C).
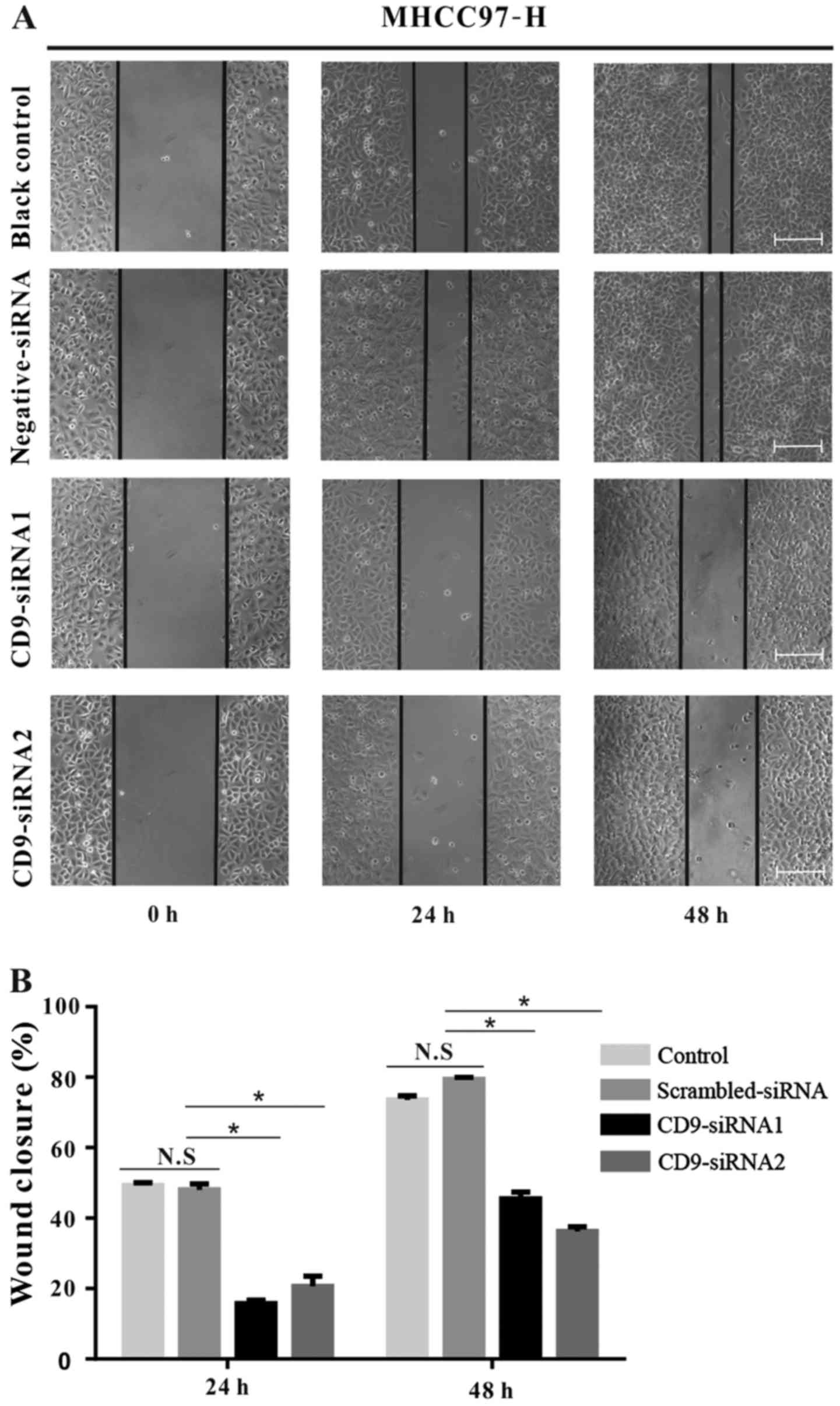
Reduced CD9 expression inhibits MHCC97-H
cell migration and invasion
To further examine the role of CD9 in HCC cell
motility, wound-healing and invasion assays were performed on
untransfected MHCC97-H cells as well as MHCC97-H cells that were
transfected with CD9-siRNA1, CD9-siRNA2 or scrambled-siRNA. Results
from the wound-healing assay demonstrated that CD9 silencing
significantly decreased the migratory ability of MHCC97-H cells
transfected with CD9-siRNA1 and CD9-siRNA2 compared with the
control groups (P<0.05) (Fig.
4). Similarly, the CD9-siRNA transfections resulted in
significantly decreased invasive ability compared with the control
groups (P<0.05) (Fig. 5).
Discussion
Membrane proteins may affect the occurrence,
development and metastasis of tumors in many ways (6,9).
Tumor heterogeneity suggests that the membrane proteins in tumor
cells at differing metastatic potentials may be different (6,24,25).
Examination of cytoplasmic membrane proteins in tumor cells may not
only reveal the mechanisms of tumor progression, but also may
identify reliable tumor markers and drug targets (26–28).
Membrane proteins are difficult to dissolve, isolate and identify
owing to their heterogeneity, hydrophobicity and low abundance
(24), which creates a difficult
problem for membrane protein research (25). iTRAQ analysis has become an
important tool for protein identification and quantitative
analysis. The iTRAQ approach allows for the tagging and detection
of peptides, membrane proteins, hydrophobic or macromolecular
proteins and proteins that are expressed in low abundance (29). Therefore, the present study used
this technique to investigate the membrane proteins in cells with
similar genetic backgrounds, but with different metastatic
potential, to identify the proteins that may be related to tumor
metastasis.
MHCC97-L and MHCC97-H are two subtype cell lines
that were established by cloning of the metastatic liver cancer
cell line MHCC97 (30). A previous
study reported that, although these lines share the same genetic
background, the MHCC97-L cell line is characterized by larger cell
volume and slower growth, invasion and metastasis, compared with
MHCC97-H cells, which exhibit a highly invasive phenotype (31).
In the present study, a total of 16 proteins among
the 22 identified as differentially expressed were upregulated in
MHCC97-H cells, some of which have been previously reported to
serve key roles in tumor metastases, including AHNAK, EPCAM and
ITGB1 (32–34). CD9 was one of the highly expressed
membrane proteins in MHCC97-H cells.
CD9 is a cell surface glycoprotein that belongs to
the tetraspanin superfamily of proteins, which are characterized by
the presence of four hydrophobic transmembrane domains (35). Tetraspanin proteins may affect
tumorigenesis in a number of ways, such as by promoting or
inhibiting tumor growth, invasion and metastasis (35–39).
CD9 is widely expressed on the cell surface, particularly in
malignant tumor cell lines (37,40).
Functional studies have demonstrated that CD9 serves a role in
tumor cell motility and adhesion, and may have an effect on
metastasis (41–43). Previous studies reported that
overexpression of CD9 in breast, prostate, colon, lung,
neuroblastoma and ovarian carcinoma cells appeared to suppress the
motility and metastatic potential of cells (44–49).
However, CD9 expression varies greatly among different organ
tumors, and may either promote or inhibit metastasis (36). Previous studies have demonstrated
that the expression level of CD9 was elevated in certain tumoral
cells, such as cervical cancer, prostate cancer and fibrosarcoma,
and that CD9 may promote tumor progression (50–52).
These data suggested that CD9 may differentially affect particular
aspects of tumor growth and progression.
Currently, there are a limited number of studies on
the effects of CD9 on the malignant biological behavior of HCC. A
previous study suggested that high CD9 expression levels may
inhibit liver cancer metastasis (53), and a different study reported that
CD9 expression levels were high in healthy liver cells and low in
liver parenchymal cells (54). The
role of CD9 in liver cancer remains unclear, and the relationship
between CD9 expression and HCC metastasis, one of the most
metastatic types of tumor, should be further examined in future
studies.
In the present study, CD9 was demonstrated to be
highly expressed in MHCC97-H cell lines compared with MHCC97-L
cells. The functional role of CD9 was characterized using a cell
model, which revealed that the migratory capacity was significantly
inhibited in MHCC97-H cells following the knockdown of CD9
expression, which was consistent with previous studies that
demonstrated a potent role of CD9 in accelerating tumor cell growth
(50–52). But the expression of CD9 was
reported to be reduced when tumor metastasis occurred in pancreatic
cancer and colon cancer (55,56).
Future studies will compare normal hepatocytes and HCC cells to
further analyze the role of CD9 in the liver. In conclusion, iTRAQ
proteomics analysis and cell functional experiments performed in
the present study indicated that CD9 may promote metastasis in HCC
cells. Results from the wound healing and Matrigel assays suggested
that CD9 expression may serve an important role in HCC metastasis
and invasion.
Acknowledgments
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YY and SP conceived and designed the study. QL
performed the experiments and wrote the paper. YY reviewed and
edited the manuscript. All authors read and approved the
manuscript.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Chaffer CL and Weinberg RA: A perspective
on cancer cell metastasis. Science. 331:1559–1564. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Parkin DM, Bray F, Ferlay J and Pisani P:
Global cancer statistics, 2002. CA Cancer J Clin. 55:74–108. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
El-Serag HB and Kanwal F: Epidemiology of
hepatocellular carcinoma in the United States: Where are we? Where
do we go? Hepatology. 60:1767–1775. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bruix J and Sherman M; American
Association for the Study of Liver Diseases: Management of
hepatocellular carcinoma: An update. Hepatology. 53:1020–1022.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Josic D, Clifton JG, Kovac S and Hixson
DC: Membrane proteins as diagnostic biomarkers and targets for new
therapies. Curr Opin Mol Ther. 10:116–123. 2008.PubMed/NCBI
|
|
7
|
Boonstra MC, de Geus SW, Prevoo HA,
Hawinkels LJ, van de Velde CJ, Kuppen PJ, Vahrmeijer AL and Sier
CF: Selecting targets for tumor imaging: An overview of
cancer-associated membrane proteins. Biomark Cancer. 8:119–133.
2016.PubMed/NCBI
|
|
8
|
Mamede AC, Laranjo M, Carvalho MJ,
Abrantes AM, Pires AS, Brito AF, Moura P, Maia CJ and Botelho MF:
Effect of amniotic membrane proteins in human cancer cell lines: An
exploratory study. J Membr Biol. 247:357–360. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Rucevic M, Hixson D and Josic D: Mammalian
plasma membrane proteins as potential biomarkers and drug targets.
Electrophoresis. 32:1549–1564. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li SY, Yu B, An P, Liang ZJ, Yuan SJ and
Cai HY: Effects of cell membrane phospholipid level and protein
kinase C isoenzyme expression on hepatic metastasis of colorectal
carcinoma. Hepatobiliary Pancreat Dis Int. 3:411–416.
2004.PubMed/NCBI
|
|
11
|
Sun GG, Wang YD, Cui DW, Cheng YJ and Hu
WN: Epithelial membrane protein 1 negatively regulates cell growth
and metastasis in colorectal carcinoma. World J Gastroenterol.
20:4001–4010. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Agarwal AK, Srinivasan N, Godbole R, More
SK, Budnar S, Gude RP and Kalraiya RD: Role of tumor cell surface
lysosome-associated membrane protein-1 (LAMP1) and its associated
carbohydrates in lung metastasis. J Cancer Res Clin Oncol.
141:1563–1574. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lee TY, Chang HH, Kuo JJ and Shen JJ:
Changes of hepatic proteome in bile duct ligated rats with hepatic
fibrosis following treatment with Yin-Chen-Hao-Tang. Int J Mol Med.
23:477–484. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ross PL, Huang YN, Marchese JN, Williamson
B, Parker K, Hattan S, Khainovski N, Pillai S, Dey S, Daniels S, et
al: Multiplexed protein quantitation in Saccharomyces cerevisiae
using amine-reactive isobaric tagging reagents. Mol Cell
Proteomics. 3:1154–1169. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li Y, Tang ZY, Ye SL, Liu YK, Chen J, Xue
Q, Chen J, Gao DM and Bao WH: Establishment of cell clones with
different metastatic potential from the metastatic hepatocellular
carcinoma cell line MHCC97. World J Gastroenterol. 7:630–636. 2001.
View Article : Google Scholar
|
|
16
|
Wang C, Zhu ZM, Liu CL, He XJ, Zhang HY
and Dong JH: Knockdown of yes-associated protein inhibits
proliferation and downregulates large tumor suppressor 1 expression
in MHCC97H human hepatocellular carcinoma cells. Mol Med Rep.
11:4101–4108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zou YH, Li XD, Zhang QH and Liu DZ: RACK1
silencing induces cell apoptosis and inhibits cell proliferation in
hepatocellular carcinoma MHCC97-H cells. Pathol Oncol Res.
24:101–107. 2018. View Article : Google Scholar
|
|
18
|
Tian M, Cheng H, Wang Z, Su N, Liu Z, Sun
C, Zhen B, Hong X, Xue Y and Xu P: Phosphoproteomic analysis of the
highly-metastatic hepatocellular carcinoma cell line, MHCC97-H. Int
J Mol Sci. 16:4209–4225. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sun H and Liu GT: Inhibitory effect of
anti-hepatitis drug bicyclol on invasion of human hepatocellular
carcinoma MHCC97-H cells with high metastasis potential and its
relative mechanisms. J Asian Nat Prod Res. 11:576–583. 2009.
View Article : Google Scholar
|
|
20
|
Yang Y, Toy W, Choong LY, Hou P, Ashktorab
H, Smoot DT, Yeoh KG and Lim YP: Discovery of SLC3A2 cell membrane
protein as a potential gastric cancer biomarker: Implications in
molecular imaging. J Proteome Res. 11:5736–5747. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang LN, Tong SW, Hu HD, Ye F, Li SL, Ren
H, Zhang DZ, Xiang R and Yang YX: Quantitative proteome analysis of
ovarian cancer tissues using a iTRAQ approach. J Cell Biochem.
113:3762–3772. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
She S, Xiang Y, Yang M, Ding X, Liu X, Ma
L, Liu Q, Liu B, Lu Z, Li S, et al: C-reactive protein is a
biomarker of AFP-negative HBV-related hepatocellular carcinoma. Int
J Oncol. 47:543–554. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-ΔΔC(T)) method. Methods. 25:402–408. 2001. View Article : Google Scholar
|
|
24
|
Bledi Y, Inberg A and Linial M: PROCEED: A
proteomic method for analysing plasma membrane proteins in living
mammalian cells. Brief Funct Genomics Proteomics. 2:254–265. 2003.
View Article : Google Scholar
|
|
25
|
Josic D and Clifton JG: Mammalian plasma
membrane proteomics. Proteomics. 7:3010–3029. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang H, Tian B, Yu H, Yao H and Gao Z:
LAPTM4B-35 protein as a potential therapeutic target in gastric
cancer. Tumour Biol. 35:12737–12742. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yoshizaki T, Kondo S, Endo K, Nakanishi Y,
Aga M, Kobayashi E, Hirai N, Sugimoto H, Hatano M, Ueno T, et al:
Modulation of the tumor microenvironment by Epstein-Barr virus
latent membrane protein 1 in nasopharyngeal carcinoma. Cancer Sci.
109:272–278. 2018. View Article : Google Scholar
|
|
28
|
Xue Q, Zhou Y, Wan C, Lv L, Chen B, Cao X,
Ju G, Huang Y, Ni R and Mao G: Epithelial membrane protein 3 is
frequently shown as promoter methylation and functions as a tumor
suppressor gene in non-small cell lung cancer. Exp Mol Pathol.
95:313–318. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liang S, Xu Z, Xu X, Zhao X, Huang C and
Wei Y: Quantitative proteomics for cancer biomarker discovery. Comb
Chem High Throughput Screen. 15:221–231. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tian J, Tang Z and Ye S: Establishment of
a human hepatocellular carcinoma (HCC) cell line with high
metastatic potential (MHCC97) and its biological characteristics.
Zhonghua Zhong Liu Za Zhi. 20:405–407. 1998.In Chinese.
|
|
31
|
Tian J, Tang ZY, Ye SL, Liu YK, Lin ZY,
Chen J and Xue Q: New human hepatocellular carcinoma (HCC) cell
line with highly metastatic potential (MHCC97) and its expressions
of the factors associated with metastasis. Br J Cancer. 81:814–821.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen B, Wang J, Dai D, Zhou Q, Guo X, Tian
Z, Huang X, Yang L, Tang H and Xie X: AHNAK suppresses tumour
proliferation and invasion by targeting multiple pathways in
triple-negative breast cancer. J Exp Clin Cancer Res. 36:652017.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hoe SL, Tan LP, Abdul Aziz N, Liew K, Teow
SY, Abdul Razak FR, Chin YM, Mohamed Shahrehan NA, Chu TL, Mohd
Kornain NK, et al: CD24, CD44 and EpCAM enrich for
tumour-initiating cells in a newly established patient-derived
xenograft of nasopharyngeal carcinoma. Sci Rep. 7:123722017.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Qin Q, Wei F, Zhang J and Li B: miR-134
suppresses the migration and invasion of non small cell lung cancer
by targeting ITGB1. Oncol Rep. 37:823–830. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yáñez-Mó M, Barreiro O, Gordon-Alonso M,
Sala-Valdés M and Sánchez-Madrid F: Tetraspanin-enriched
microdomains: A functional unit in cell plasma membranes. Trends
Cell Biol. 19:434–446. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zöller M: Tetraspanins: Push and pull in
suppressing and promoting metastasis. Nat Rev Cancer. 9:40–55.
2009. View
Article : Google Scholar
|
|
37
|
Hemler ME: Tetraspanin proteins mediate
cellular penetration, invasion, and fusion events and define a
novel type of membrane microdomain. Annu Rev Cell Dev Biol.
19:397–422. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zijlstra A, Lewis J, Degryse B, Stuhlmann
H and Quigley JP: The inhibition of tumor cell intravasation and
subsequent metastasis via regulation of in vivo tumor cell motility
by the tetraspanin CD151. Cancer Cell. 13:221–234. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Schröder HM, Hoffmann SC, Hecker M, Korff
T and Ludwig T: The tetraspanin network modulates MT1-MMP cell
surface trafficking. Int J Biochem Cell Biol. 45:1133–1144. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Boucheix C and Rubinstein E: Tetraspanins.
Cell Mol Life Sci. 58:1189–1205. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Boucheix C, Benoit P, Frachet P, Billard
M, Worthington RE, Gagnon J and Uzan G: Molecular cloning of the
CD9 antigen. A new family of cell surface proteins. J Biol Chem.
266:117–122. 1991.PubMed/NCBI
|
|
42
|
Lanza F, Wolf D, Fox CF, Kieffer N, Seyer
JM, Fried VA, Coughlin SR, Phillips DR and Jennings LK: cDNA
cloning and expression of platelet p24/CD9. Evidence for a new
family of multiple membrane-spanning proteins. J Biol Chem.
266:10638–10645. 1991.PubMed/NCBI
|
|
43
|
Maecker HT, Todd SC and Levy S: The
tetraspanin superfamily: Molecular facilitators. FASEB J.
11:428–442. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Miyake M, Nakano K, Itoi SI, Koh T and
Taki T: Motility-related protein-1 (MRP-1/CD9) reduction as a
factor of poor prognosis in breast cancer. Cancer Res.
56:1244–1249. 1996.PubMed/NCBI
|
|
45
|
Copeland BT, Bowman MJ, Boucheix C and
Ashman LK: Knockout of the tetraspanin Cd9 in the TRAMP model of de
novo prostate cancer increases spontaneous metastases in an
organ-specific manner. Int J Cancer. 133:1803–1812. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Hashida H, Takabayashi A, Tokuhara T, Taki
T, Kondo K, Kohno N, Yamaoka Y and Miyake M: Integrin α3 expression
as a prognostic factor in colon cancer: Association with MRP-1/CD9
and KAI1/CD82. Int J Cancer. 97:518–525. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Higashiyama M, Taki T, Ieki Y, Adachi M,
Huang CL, Koh T, Kodama K, Doi O and Miyake M: Reduced motility
related protein-1 (MRP-1/CD9) gene expression as a factor of poor
prognosis in non-small cell lung cancer. Cancer Res. 55:6040–6044.
1995.PubMed/NCBI
|
|
48
|
Fabian J, Opitz D, Althoff K, Lodrini M,
Hero B, Volland R, Beckers A, de Preter K, Decock A, Patil N, et
al: MYCN and HDAC5 transcriptionally repress CD9 to trigger
invasion and metastasis in neuroblastoma. Oncotarget.
7:66344–66359. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Houle CD, Ding XY, Foley JF, Afshari CA,
Barrett JC and Davis BJ: Loss of expression and altered
localization of KAI1 and CD9 protein are associated with epithelial
ovarian cancer progression. Gynecol Oncol. 86:69–78. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Sauer G, Windisch J, Kurzeder C, Heilmann
V, Kreienberg R and Deissler H: Progression of cervical carcinomas
is associated with down-regulation of CD9 but strong local
re-expression at sites of transendothelial invasion. Clin Cancer
Res. 9:6426–6431. 2003.PubMed/NCBI
|
|
51
|
Wang JC, Bégin LR, Bérubé NG, Chevalier S,
Aprikian AG, Gourdeau H and Chevrette M: Down-regulation of CD9
expression during prostate carcinoma progression is associated with
CD9 mRNA modifications. Clin Cancer Res. 13:2354–2361. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Herr MJ, Kotha J, Hagedorn N, Smith B and
Jennings LK: Tetraspanin CD9 promotes the invasive phenotype of
human fibrosarcoma cells via upregulation of matrix
metalloproteinase-9. PLoS One. 8:e677662013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Zheng R, Yano S, Zhang H, Nakataki E,
Tachibana I, Kawase I, Hayashi S and Sone S: CD9 overexpression
suppressed the liver metastasis and malignant ascites via
inhibition of proliferation and motility of small-cell lung cancer
cells in NK cell-depleted SCID mice. Oncol Res. 15:365–372. 2005.
View Article : Google Scholar
|
|
54
|
Uhlén M, Fagerberg L, Hallström BM,
Lindskog C, Oksvold P, Mardinoglu A, Sivertsson Å, Kampf C,
Sjöstedt E, Asplund A, et al: Proteomics. Tissue-based map of the
human proteome. Science. 347:12604192015. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Sho M, Adachi M, Taki T, Hashida H,
Konishi T, Huang CL, Ikeda N, Nakajima Y, Kanehiro H, Hisanaga M,
et al: Transmembrane 4 superfamily as a prognostic factor in
pancreatic cancer. Int J Cancer. 79:509–516. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Mori M, Mimori K, Shiraishi T, Haraguchi
M, Ueo H, Barnard GF and Akiyoshi T: Motility related protein 1
(MRP1/CD9) expression in colon cancer. Clin Cancer Res.
4:1507–1510. 1998.PubMed/NCBI
|