Introduction
Breast cancer, one of the most common malignancies
affecting women, has attracted increasing attention by the
international community and has evoked tremendous interest
paritcularly in the medical and academic fields (1). An increasing number of studies have
been performed to explore potential biomarkers which may be
involved in the initiation and progression of breast cancer.
Estrogen receptor (ER), progesterone receptor (PR), c-erbB-2, p53
and Ki-67 have been investigated in the conventional
histopathological setting (2,3).
However, numerous other genes and proteins are also involved in the
abnormal regulation of signaling pathways and the progression of
beast cancer.
Zinc finger of the cerebellum 1 (ZIC1) is located on
chromosome 3q25.1 and belongs to the ZIC family. ZIC1 encodes a
zinc finger transcription factor and is involved in the mediation
of growth and physiological metabolism (4,5).
Studies have reported that ZIC1 is a putative suppressor gene in
various carcinomas, including digestive system cancers and thyroid
cancer (6–8). Moreover, its stimulation inhibits the
growth of cancer cells by reducing Akt and Erk phosphorylation
(6–8). The methylation frequency of ZIC1 in a
variety of tumors has been shown to be significantly higher than
that in the corresponding non-cancerous tissues (7–9). In
addition, a high expression of ZIC1 has become a potential
biomarker for a good prognosis (10). In breast cancer, the expression of
ZIC1 has been shown to be elevated in BT-549 cells following the
knockdown PIGX, RCN1, or RCN2, along with the growth of transfected
cells being markedly inhibited (11). However, a further mechanistic
analysis of ZIC1 in breast cancer still needs to be conducted.
Survivin, known as baculoviral IAP repeat containing
5 (BIRC5), is located on chromosome 17q25.3. As an inhibitor of
apoptotic proteins, survivin plays an important role in controlling
cell mitosis, and enhancing the resistance to anticancer agents
(12,13). Moreover, it is frequently
overexpressed in multifarious tumors, including breast cancer,
while it is rarely expressed in normal human tissues (12–15).
These findings suggest that elevated survivin is involved in
carcinogenesis. In addition, previous studies have demonstrated
that survivin suppresses the apoptosis of cancer cells through the
inactivation of caspase-9 and caspase-3 (16,17).
In addition, the phosphorylation of Akt and mammalian target of
rapamycin (mTOR) is considered to be responsible for the
upregulation of surviving (16,17).
Therefore, this study was designed to identify the role of ZIC1 in
breast cancer in vitro and in vivo, and to examine
the association of ZIC1 with surviving, elucidating the underlying
molecular mechanisms.
Materials and methods
Patients and tissue specimens
Specimens from a total of 120 patients with invasive
breast cancer who underwent radical mastectomy at Shanghai Changhai
Hospital (Shanghai, China) from January, 2014 to December, 2015
were collected, with a mean age of 52.19±12.38 years. Each case
consisted of a breast tumor and its corresponding adjacent normal
tissue. None of the patients had received radiotherapy or
chemotherapy prior to surgery. In addition, 20 pairs of
fresh-frozen invasive breast tumors and matched normal tissues were
also collected from Kunshan First People's Hospital Affiliated to
Jiangsu University (Kunshan, China) and were used for total protein
extraction and western blot analysis, following the approval of
Kunshan First People's Hospital Ethics Committee. Each patient
signed the informed consent form.
A database (https://www.proteinatlas.org/), the Human Protein
Atlas (HPA), which is a Swedish-based program, containing 1,075
TCGA RNA breast cancer samples with overall survival information
and human genome-wide expression analyses, was also used. We
selected this database to compare the RNA level of ZIC1 with that
of survivin and to explore the prognostic roles of ZIC1 and
survivin in patients with breast cancer. More detailed information
of these two genes can be found in the following websites: For
ZIC1, https://www.proteinatlas.org/ENSG00000152977-ZIC1/pathology/tissue/breast+cancer;
and for survivin, https://www.proteinatlas.org/ENSG00000089685-BIRC5/pathology/tissue/breast+cancer.
Cell lines and cell culture
Human breast cancer cell lines (MCF-7, MDA-MB-231,
MDA-MB-453, SK-BR3, BT-474 and BT-549) and the human mammary
epithelial cell line, MCF-10A, were obtained from the Cell Center
at the Institutes of Biomedical Sciences (IBS), Fudan University,
Fudan, China. All cancer cell lines were cultured in appropriate
medium supplemented with 10% fetal bovine serum (FBS; Gibco Life
Technologies/Thermo Fisher Scientific, Waltham, MA, USA) and 1%
antibiotic/antimycotic solution (Sigma-Aldrich, St. Louis, MO, USA)
at 37°C in a humidified atmosphere containing 5% CO2
with Dulbecco's modified Eagle's medium (DMEM) for the MCF-7 and
SK-BR3 cells; RPMI-1640 for the MDA-MB-231, BT-549 and BT-474
cells; and Leibovitz's L-15 (all from Gibco Life
Technologies/Thermo Fisher Scientific) for the MDA-MB-453 cells.
The MCF-10A cells were grown in specific medium with DF-12 + 5% HS
+ EGF (20 ng/ml) + hydrocortisone (0.5 µg/ml) + choleratoxin (100
µg/ml) + insulin (10 µg/ml).
Cell transfection
The MDA-MB-231 and SK-BR3 cells were cultured for 24
h in a 24-well plate and transfected with rLV-Zic1-PGK-Puro
lentivirus and rLV-ZsGreen-PGK-Puro lentivirus (Wuhan Biobuffer
Biotechnology Service Co., Ltd., Wuhan, China) when the cells
reached a confluence of 30–60%. The medium for the transfected
cells was supplemented with polybrene (8 mg/ml; Sigma-Aldrich) to
elevate the transfection efficiency. After 24 h, the transfected
cells were selected in a new cell culture medium with 0.5–1.0 µg/ml
puromycin (TargetMol, Shanghai, China) for 10–12 days. The cells
trans-fected with rLV-ZsGreen-PGK-Puro were classified as the
'Vector' group and the cells transfected with rLV-Zic1-PGK-Puro
were classified as the 'ZIC1' group. Green fluorescent protein
(GFP) in the 'Vector' group was observed using a fluorescence
microscope (IX73; Olympus, Shanghai, China) to examine the
efficiency of transfection. In addition, when we constructed the
lentiviral expression vector of the ZIC1 gene, the locus of GFP
expression was cut off. As a result, the cells transfected with
rLV-Zic1-PGK-Puro failed to produce GFP, and only the cells
transfected with rLV-ZsGreen-PGK-Puro were observed through a
fluorescence microscope. To resolve this issue, we also performed
western blot analysis and examined the efficiency of transfection
in the 'ZIC1' and 'Vector' groups. The time period between
transfection and subsequent experimentation was <4 weeks.
Reverse transcription-quantitative PCR
(RT-qPCR)
The transfected cells were collected to isolate
total RNA using TRIzol regent (Thermo Fisher Scientific). A total
of 2 µg RNA of each sample was reverse transcribed using the
SuperScript II RNase-Reverse Transcriptase system (Thermo Fisher
Scientific). cDNA was subjected to qPCR using primers specific for
ZIC1, survivin and GAPDH. The PCR primers were designed as follows:
ZIC1 forward, 5′-GCGTCCTTTTGTGGATCTTTAA-3′ and reverse,
5′-AGTAATCACATCTGCTTCTGGG-3′ (178 bp); survivin forward,
5′-ATGTCCATTTTTCAGGTTCTCTAAG-3′ and reverse,
5′-GCACTGCTGTCTCTACTTTCC-3′ (142 bp); and GAPDH (internal control)
forward, 5′-GAAGGTGAAGG TCGGAGT-3′ and reverse,
5′-GAAGATGGTGATGGGAT TTC-3′ (226 bp). The PCR cycling conditions
were as follows: i) 94°C for 4 min; ii) 40 cycles of 95°C for 1
min; iii) 60°C for 1 min; and iv) 72°C for 1 min. Amplified DNA was
measured using the SYBR Premix Ex Taq™ kit (Takara Bio, Tokyo,
Japan), and qPCR was performed using an iQ5 real-time PCR detection
system (Bio-Rad, Hercules, CA, USA). We then used the
2−ΔΔCq value to calculate the relative expression and
ΔΔCq = (CqrLV-Zic1-PGK-Puro-ZIC1/survivin −
CqrLV-Zic1-PGK-Puro-GAPDH) −
(CqVector-ZIC1/survivin- CqVector-GAPDH) (18).
Cell viability assay
To assess cell proliferation, the stably
trans-fected cells seeded in 96-well plates for 1–5 days, were
treated with MTT (Sigma-Aldrich),
3-(4,5-Dimethylthigal-2-yl)-2,5-(diphenyltetragalium) bromide for 4
h at the incubator. The supernatant was replaced with DMSO, and
then the 96-well plates were oscillated for 10 min. The absorbance
of each group was read at 490 nm of the microplate reader (OD
value) (MD/F5; Shanghai Magvalley Technology Co., Ltd., Shanghai,
China).
Colony formation assay
Stably transfected cells were seeded in 6-well
plates at a density of 200 cells and cultured for about 10 days,
when the clones were visible (≥50 cells). The colonies were fixed
for 15 min with methanol, and stained with crystal violet for 15
min after airing. Then, the numbers of colonies were counted.
Cell cycle analysis
Stably transfected cells were treated with 0.25%
trypsin without EDTA (Gibco Life Technologies/Thermo Fisher
Scientific) at 37°C for approximately 5 min. Subsequently, after
being neutralized by medium, the cells were centrifuged at 1,000 ×
g for 5 min. The centrifuged cells were washed 2 times with
pre-cooling phosphate-buffered saline (PBS), and were fixed in 80%
ethanol for 12 h at 4°C. The cells were then washed twice again
with pre-cooling PBS prior to incubation for 30 min at 37°C in the
dark with propidium iodide (PI) and RNAse A (KeyGen Biotech,
Nanjing, China). Fluorescence-activated cell sorting analysis was
performed using a flow cytometer (FACScalibur, E97600114; BD
Biosciences, New Jersey, NY, USA).
Analysis of cell apoptosis
Due to the expression of GFP in the 'Vector' group
only, we selected red the fluorescent proteins, Annexin V-APC and
7-AAD as the fluorescent markers in cell apoptosis assay. Stably
transfected cells were treated with 0.25% trypsin without EDTA
(Gibco Life Technologies/Thermo Fisher Scientific) at 37°C for
approximately 5 min. Subsequently, after being neutralized by
medium, the cells were centrifuged at 1,000 × g for 5 min. The
centrifuged cells were washed 2 times with PBS, and stained with
Annexin V-APC and 7-AAD for 15 min at room temperature in the dark
(KeyGen Biotech). After being diluted in moderate PBS, the stained
cells were analyzed using a flow cytometer.
Western blot analysis
Proteins from tissues or cells were extracted using
RIPA extraction buffer with PMSF (Beyotime Institute of
Biotechnology, Shanghai, China) and phosphatase inhibitor (CWBio,
Beijing, China). BCA was used for protein quantification and the
proteins were then processed for western blot analysis by means of
8–16% pre-cast protein gel electrophoresis (Thermo Fisher
Scientific) at 20 µg per lane and were transferred onto PVDF
membranes (Beyotime Institute of Biotechnology). ZIC1 (1:500;
bs-11609R; Bioss Biotechnology, Beijing, China), survivin
(RLM3419), Akt (RLT0178), p-Akt (RLP0006), mTOR (RLT2915), p-mTOR
(RLP0176), cleaved caspase-9 (RLC0012), cleaved caspase-3 (RLC0004)
(all 1:1,000; Suzhou Ruying Biotechnology, Suzhou, China), cyclin
D1 (bs-20596R, 1:1,000; Bioss Biotechnology), p21 (YT3497), p27
(YT3501) (both 1:500; ImmunoWay Biotechnology, Piano, TX, USA),
P70S6K (2708S), p-P70S6K (9204S), Bcl-2 (15071S), Bad (9268S),
p-Bad (9291S) and Bcl-xL (2764S) (all 1:1,000) were used as the
primary antibodies, and GAPDH (5174S, 1:1,000) (both from Cell
Signaling Technology, Danvers, MA, USA) was used as the reference
protein. The secondary antibodies were HRP-labeled goat anti-rabbit
IgG (A0208) and HRP-labeled goat anti-mouse IgG (A0216) (both
1:1,000; Beyotime Institute of Biotechnology). The enhanced
chemiluminescent (ECL) kit (Beyotime Institute of Biotechnology)
was used to detect the results of western blot analysis. The
relative level of each protein was deduced from the ratio of the
mean value of each band to that of GAPDH. The relative densities
were quantified with a digital imaging analyzer, ImageJ 1.4.1.
Mitochondrial membrane potential
assay
In order to detect cytochrome c (Cyto-c)
expression in the mitochondria and cytoplasm, we first extracted
mitochondrial cytosolic proteins sing the Minute™ Mitochondria
Isolation kit (Invent Biotechnologies, Inc., Plymouth, MN, USA).
Cyto-c (A13430, 1:1,000; ABclonal Technology, Wuhan, China) in the
mitochondria or cytosol was detected by by western blot analysis as
described above. In addition, COX-IV (A6564) and tubulin (A0482)
(1:1,000; ABclonal Technology) were used as the reference proteins
of the mitochondria and cytosol, respectively. The JC-1 kit
(Beyotime Institute of Biotechnology) was used to investigate the
level of red fluorescence and green fluorescence in mitochondrial
proteins using a microplate reader (Shanghai Magvalley Technology
Co., Ltd.). The ratio of red to green fluorescence was used to
measure the level of depolarization of the mitochondria.
Construction of xenograft mouse
models
A total of 6 female BALB/C-nu/nu nude mice (5–6
weeks old, mean weight upon purchase, 19.02±0.29 g) were purchased
from Wuhan Biobuffer Biotechnology Service Co., Ltd. (Wuhan,
China). The aniamls were kept in an environment at 28°C and 50%
humidity under pathogen-free conditions with access to aseptic food
and water. We randomly divided the nude mice into 2 groups as
follows: The 'Vector' group (rLV-ZsGreen-PGK-Puro) and the 'ZIC1'
group (rLV-Zic1-PGK-Puro). Stably transfected MDA-MB-231 cells were
collected in the logarithmic growth phase, and were suspended in
serum-free medium at a concentration of 2.5×107
cells/ml. Subsequently, we injected 0.2 ml subcutaneously into the
right lateral back area of the BALB/C-nu/nu nude mice. The mice
were observed for tumor growth from 5 to 39 days, and the tumor
volume (mm3) was measured as follows: Tumor volume
(mm3) = length (mm) × width2
(mm2)/2. After 39 days, the nude mice were euthanized
(mean weight upon sacrifice, 19.37±0.60 g). The tumors were
excised, cut into paraffin sections and used for
immumohistochemical staining. All animal research was carried out
following the approval of the Jiangsu University Animal Ethics
Committee (Jiangsu, China).
Immunohistochemistry
Paraffin-embedded consecutive sections were
subjected to immunohistochemical staining for the expression of
ZIC1 and survivin, with the same primary antibodies as those used
in western blot analysis, diluted at 1:100 in PBS, with a SP Rabbit
& Mouse HRP Kit (CWBio). PBS without primary antibodies was
used as a negative control. Two pathologists independently
evaluated the scores of ZIC1 and survivin expression through a
semi-quantitative assessment system, with an immunoreactivity score
(IRS), which was combined by a score of the percentage of cells
('0–100%' = '0–10'), and a score of the staining intensity (0, no
staining of cells; 1, mild staining; 2, moderate staining; and 3,
marked staining). When different intensities were detected in the
cytoplasm and nucleus, we used an average score of the cytoplasm
and nucleus. The total score ranged from 0 to 30, and any
disagreement was resolved by discussion.
Statistical analysis
All results were analyzed using a t-test with SPSS
20.0 software, and a value of P<0.05 was considered to indicate
a statistically significant difference. The differences of
protein/gene expression between the breast tumors and matched
normal tissues or the differences between ZIC1 and survivin
expression in tissues were analyzed with a paired t-test, and the
differences in protein expression or biological behaviors between
the 'Vector' group and 'ZIC1' group were analyzed by an unpaired
t-test. An ANOVA followed by Tukey's multiple comparisons test was
used to compare ZIC1/survivin expression levels in the 7 cell
lines. Continuous variables are expressed as the means ± SD and
Spearman's correlation analysis was also performed to assess the
correlation between ZIC1 expression and survivin in 120 cases. In
addition, Pearson's Chi square test were used to analyze the
associations of ZIC protein expression with the patient
clinicopathological characteristics. Kaplan-Meier analysis and the
log rank test were also conducted to analyze the overall survival
of the 1,075 patients from HPA. All graphs were generated using
GraphPad Prism 6.0 software, and all figures were generated using
Adobe Photoshop CS5.
Results
ZIC1 expression inversely correlates with
surviving expression in breast tumors
We investigated the expression of ZIC1 and survivin
in invasive breast cancer by immunohistochemistry and western blot
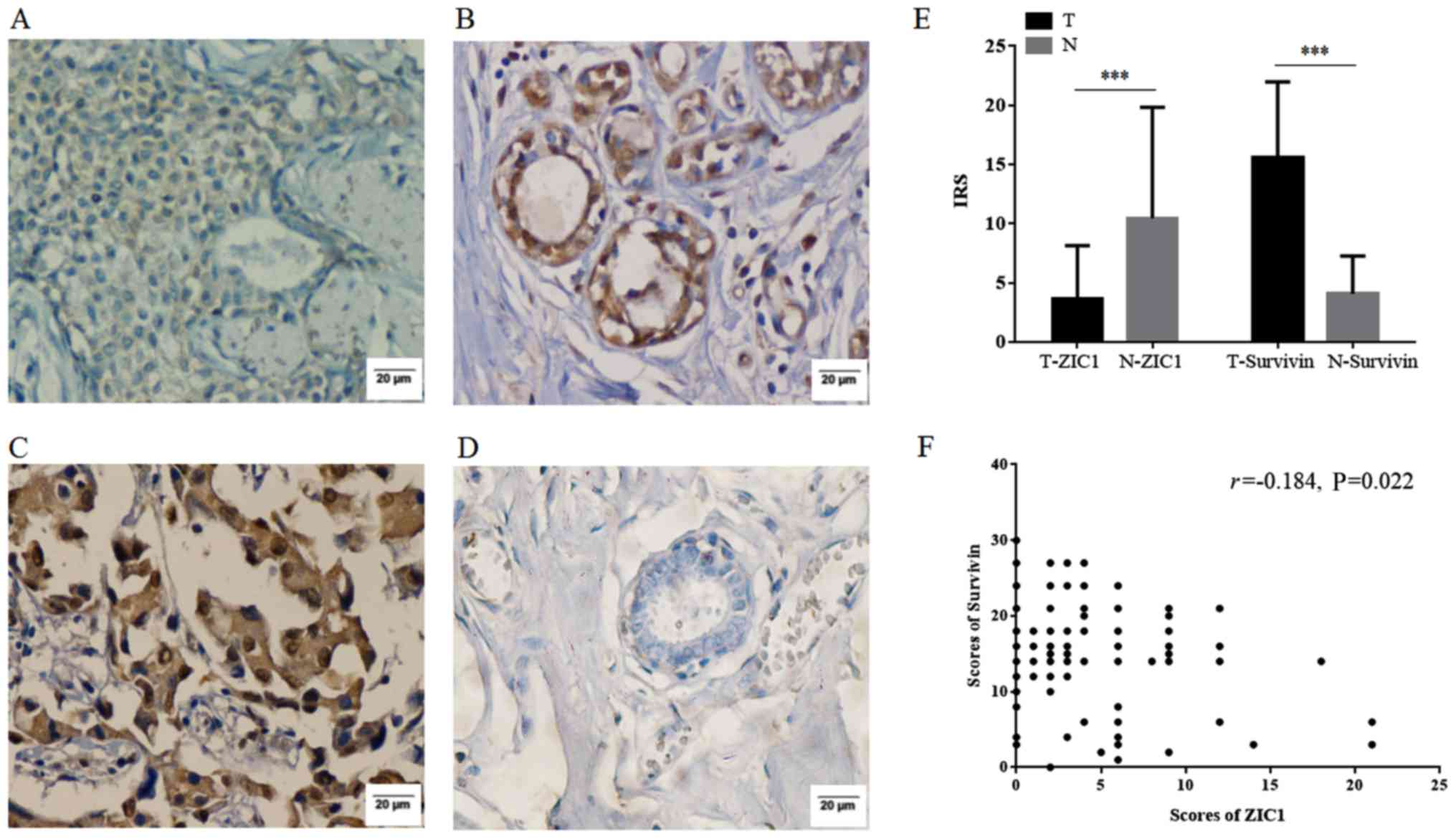
analysis. As shown in Fig. 1A–D,
ZIC1 was expressed mainly in the cytoplasm, while survivin were
expressed both in the cytoplasm and nucleus. The mean score of ZIC1
in the tumors was 3.70±4.44, and was significantly lower than that
in normal tissues (10.46±9.37, P<0.001). The mean score of
survivin in the tumors was markedly higher than that in normal
tissues (15.63±6.35 vs. 4.08±3.20, P<0.001). In addition, the
mean score of ZIC1 in the tumors was also significantly lower than
that of survivin in the tumors (P<0.001) (Fig. 1E). A negative correlation was also
observed between the scores of ZIC1 and survivin (P=0.022, Fig. 1F), which indicated that ZIC1
expression inversely correlated with survivin expression in the
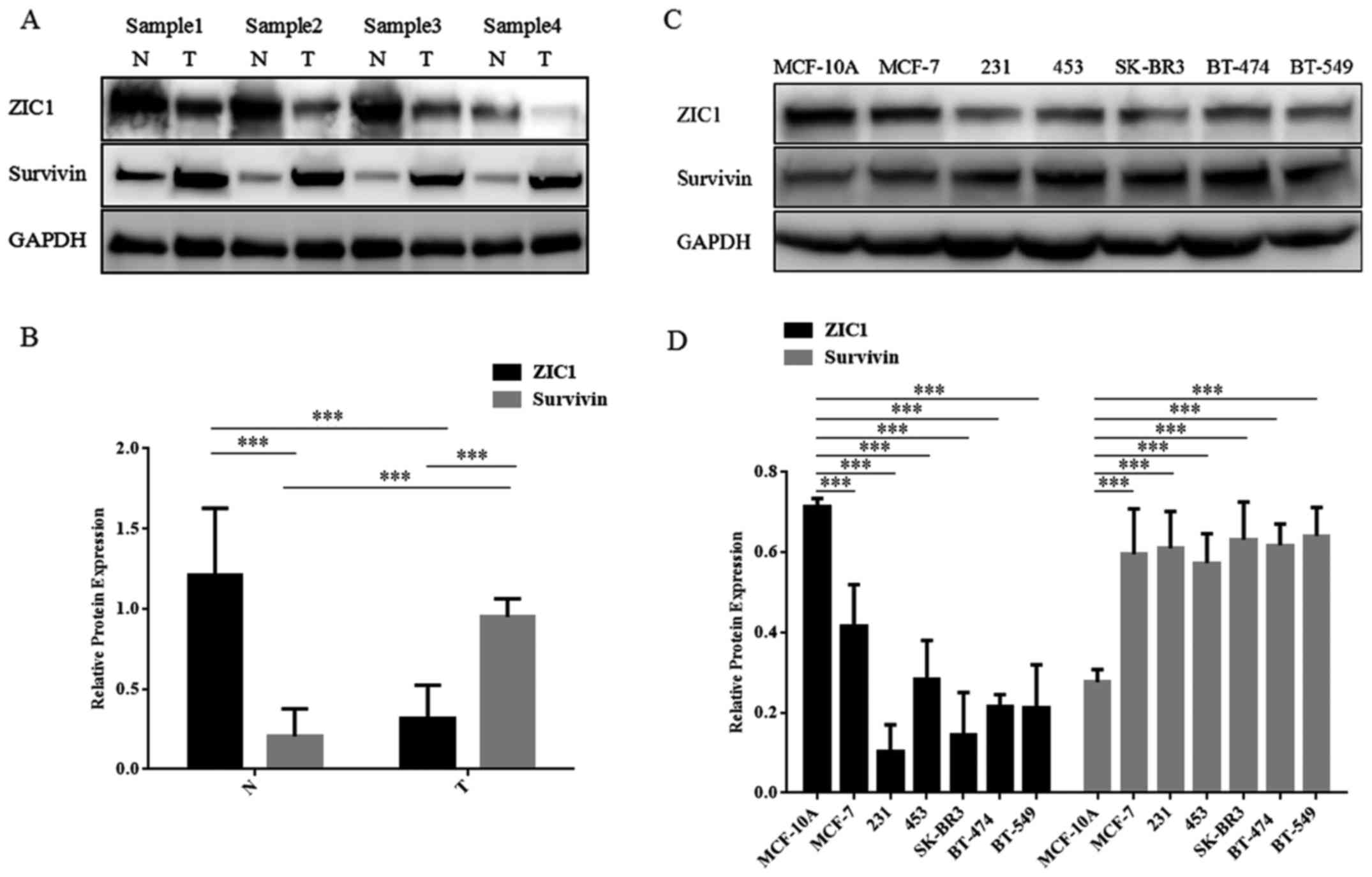
breast tumors. The results of western blot analysis of 20 pairs of
fresh-frozen tissues also yielded a similar result (Fig. 2A and B).
Associations of ZIC1 and survivin
expression with the patient clinicopathological
characteristics
Subsequently, in order to evaluate the association
between ZIC1/survivin and the patient clinicopathological
characteristics, we respectively divided the 120 cases into 2
groups according to the cut-off value (the cut-off of ZIC1 was 4;
and the cut-off of Survivin was 16; Table I). ZIC1 expression was negatively
associated with tumor differentiation (P= 0.040) and TNM staging
(P=0.016), while survivin expression was positively associated with
tumor differentiation (P<0.001), lymph node metastasis (P=0.047)
and TNM staging (P<0.001).
 | Table IAssociations of ZIC1/survivin
expression with various clinicopathological characteristics of the
120 patients with invasive breast cancer. |
Table I
Associations of ZIC1/survivin
expression with various clinicopathological characteristics of the
120 patients with invasive breast cancer.
| Parameters | n | ZIC1 expression
(cut-off=4)
| χ2
value | P-value | Survivin expression
(cut-off=16)
| χ2
value | P-value |
|---|
| (n, score
>4) | (n, score ≤4) | (n, score
>16) | (n, score ≤16) |
|---|
| Total | 120 | 36 | 84 | | | 50 | 70 | | |
| Age, years | | | | | | | | | |
| ≤50 | 67 | 18 | 49 | 0.710 | NS | 31 | 36 | 1.322 | NS |
| >50 | 53 | 18 | 35 | | | 19 | 34 | | |
| Tumor size | | | | | | | | | |
| ≤5 cm | 82 | 23 | 59 | 0.469 | NS | 36 | 46 | 0.533 | NS |
| >5 cm | 38 | 13 | 25 | | | 14 | 24 | | |
| Tumor
differentiation | | | | | | | | | |
| Well | 40 | 18 | 22 | 6.452 | 0.040 | 11 | 29 | 17.167 | <0.001 |
| Moderate | 52 | 12 | 40 | | | 18 | 34 | | |
| Poor | 28 | 6 | 22 | | | 21 | 7 | | |
| Lymph node
metastasis | | | | | | | | | |
| Positive | 79 | 20 | 59 | 2.415 | NS | 38 | 41 | 3.939 | 0.047 |
| Negative | 41 | 16 | 25 | | | 12 | 29 | | |
| TNM staging | | | | | | | | | |
| I | 27 | 13 | 14 | 8.258 | 0.016 | 14 | 13 | 19.305 | <0.001 |
| II | 54 | 17 | 37 | | | 11 | 43 | | |
| III | 39 | 6 | 33 | | | 25 | 14 | | |
ZIC1 expression is inversely associated
with surviving expression in breast cell and breast cancer cell
lines
The relative expression levels of ZIC1 and survivin
in 6 cancer cell lines and 1 normal breast cell line are presented
in Fig. 2C and D. All the breast
cancer cell lines that were examined (MCF-7, MDA-MB-231,
MDA-MB-453, SK-BR3, BT-474 and BT-549) exhibited consistently lower
levels of ZIC1 expression (F=31.30, P<0.001) and higher levels
of survivin expression (F=13.14, P<0.001) than the MCF-10A
normal breast epithelial cells. In addition, we found that the
MDA-MB-231 and SK-BR3 cells exhibited lower expression levels of
ZIC1 than the other breast cancer cells. Thus, these two cell lines
were selected for transfection with lentivirus and for use in in
vitro assays using the stably transfected cells.
Analysis of ZIC1 and survivin in the
HPA
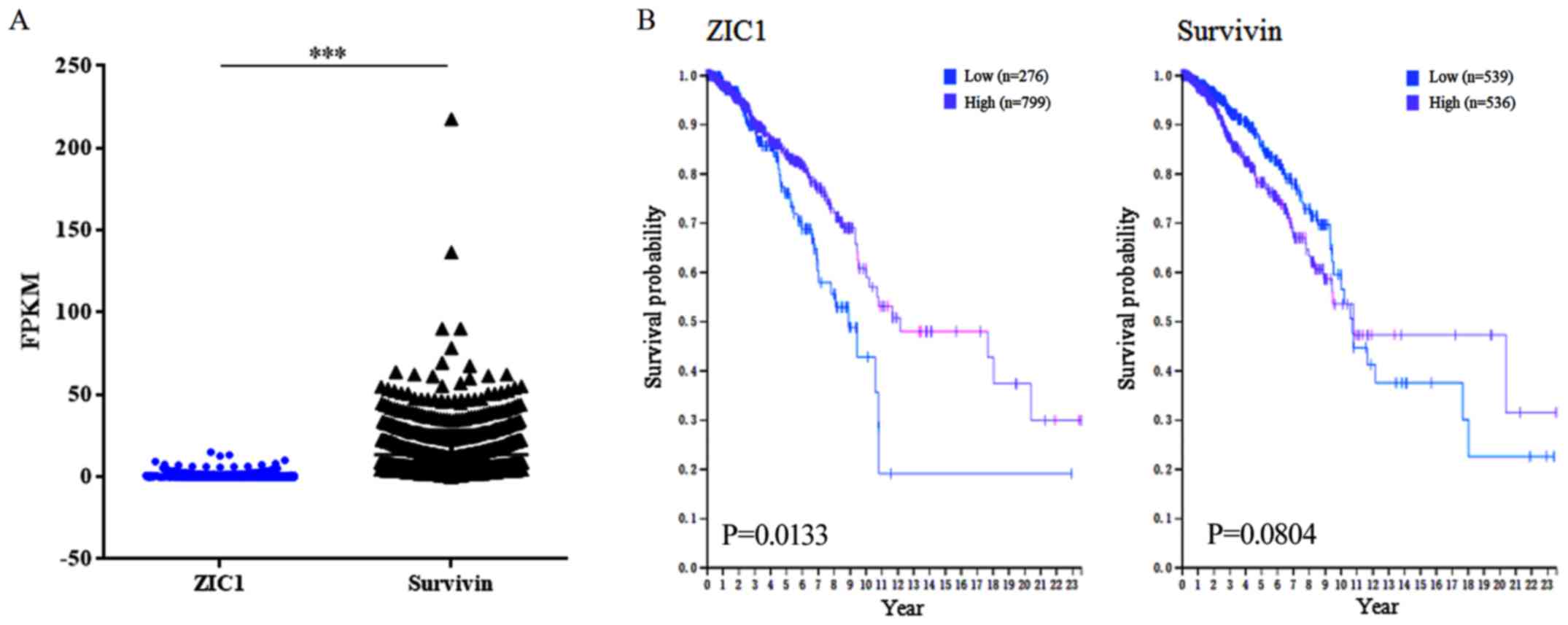
From the HPA, the RNA levels of ZIC1 and survivin of
1,075 TCGA RNA breast cancer samples were available. The average
RNA level of ZIC1 was significantly lower than that of surviving
(0.64±1.44 vs. 13.56±14.52, P<0.0001; Fig. 3A). In survival analyses, 0.0
fragments per kilobase million (FPKM) and 9.2 FPKM were used as the
expression cut-off values for ZIC1 and survivin, respectively. As a
result, 799 samples were included in the 'ZIC1 high' group, with a
higher 5-year survival rate [high vs. low, 84 vs. 76% (671/799 vs.
210/276 from HPA database), P=0.0133; Fig. 3B] while 536 samples were included
in the 'survivin high' group, with a lower survival rate [high vs.
low, 78 vs. 85% (418/536 vs. 458/539 from HPA database), P=0.0804;
Fig. 3B].
ZIC1 inhibits the proliferation and
promotes the apoptosis of breast cancer cells
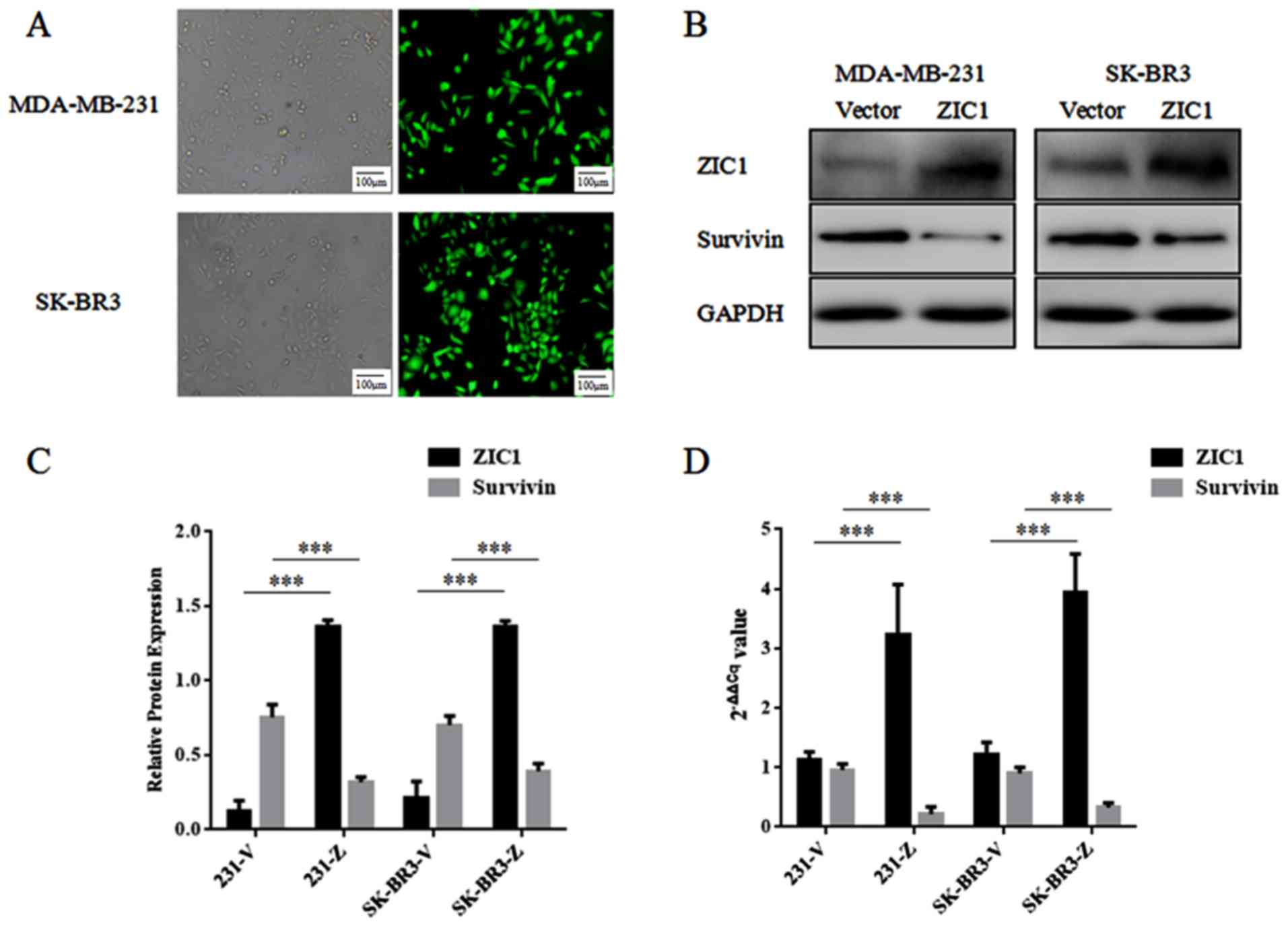
Following transfection with rLV-ZsGreen-PGK-Puro or
rLV-Zic1-PGK-Puro and selection by puromycin, the stably
transfected MDA-MB-231 and SK-BR3 cells were cultured
conventionally. The distributions of green fluorescence in the two
cell lines transfected with rLV-ZsGreen-PGK-Puro were both >90%,
and in western blot analysis, the relative expression levels of
ZIC1 in the 'ZIC1' group were markedly higher than those in the
'Vector' group. Thus, we confirmed that high levels of ZIC1
expression were only observed in the 'ZIC1' group, and not in the
'Vector' group (Fig. 4). In
addition, the expression levels of survivin in the 'ZIC1' group
were significantly decreased, in comparison with those in the
'Vector' group (Fig. 4), which
indicated that an elevated ZIC1 expression may downregulate the
expression level of survivin in breast cancer cells.
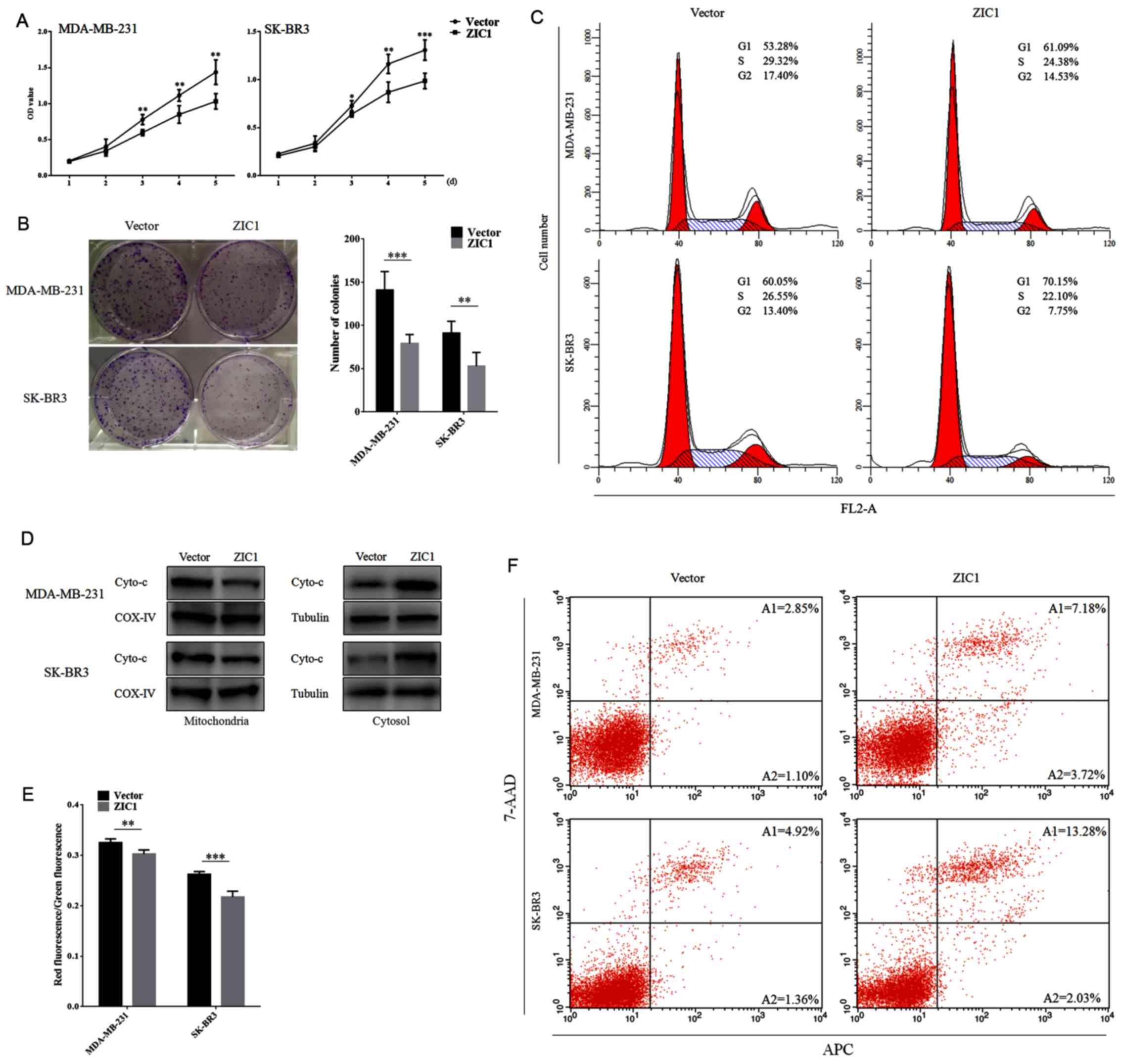
We then performed cell viability assay, colony
formation assay and cell cycle analysis in the stably transfected
cells to examine the effect of ZIC1 on cell proliferation in breast
cancer. As shown in Fig. 5A, the
enhanced expression of ZIC1 markedly suppressed the viability of
the MDA-MB-231 and SK-BR3 cells (P<0.05). We also observed that
the number of colonies formed on 6-well plates in the 'ZIC' group
was markedly lower less than that in the 'Vector' group (P<0.01;
Fig. 5B). In addition, we obtained
a higher proportion of MDA-MB-231 and SK-BR3 cells in the G1 phase
following transfection with rLV-Zic1-PGK-Puro (Fig. 5C). These results confirmed the
suppressive role of ZIC1 on the proliferation of breast cancer
cells.
After extracting mitochondrial proteins and
obtaining the corresponding cytosol, we detected the relative
expression levels of Cyto-c in the mitochondria and cytosol by
western blot analysis. In the mitochondria, the expression level of
Cyto-c was markedly lower in the 'ZIC1' group than in the 'Vector'
group, while in the cytosol, the level of Cyto-c was significantly
enhanced in the 'ZIC1' group in comparison with the 'Vector' group,
which suggested that Cyto-c was released from the mitochondria into
the cytosol following the overexpression of ZIC1 (Fig. 5D). In addition, the ratios of red
to green fluorescence were markedly decreased in the 'ZIC1' group
(Fig. 5E). The rates of cell
apoptosis were also markedly increased in the cells transfected
with the ZIC1 overexpression vector (Fig. 5F). These results suggested that an
elevated ZIC1 expression reduces the mitochondrial membrane
potential of breast cancer cells and induces apoptosis.
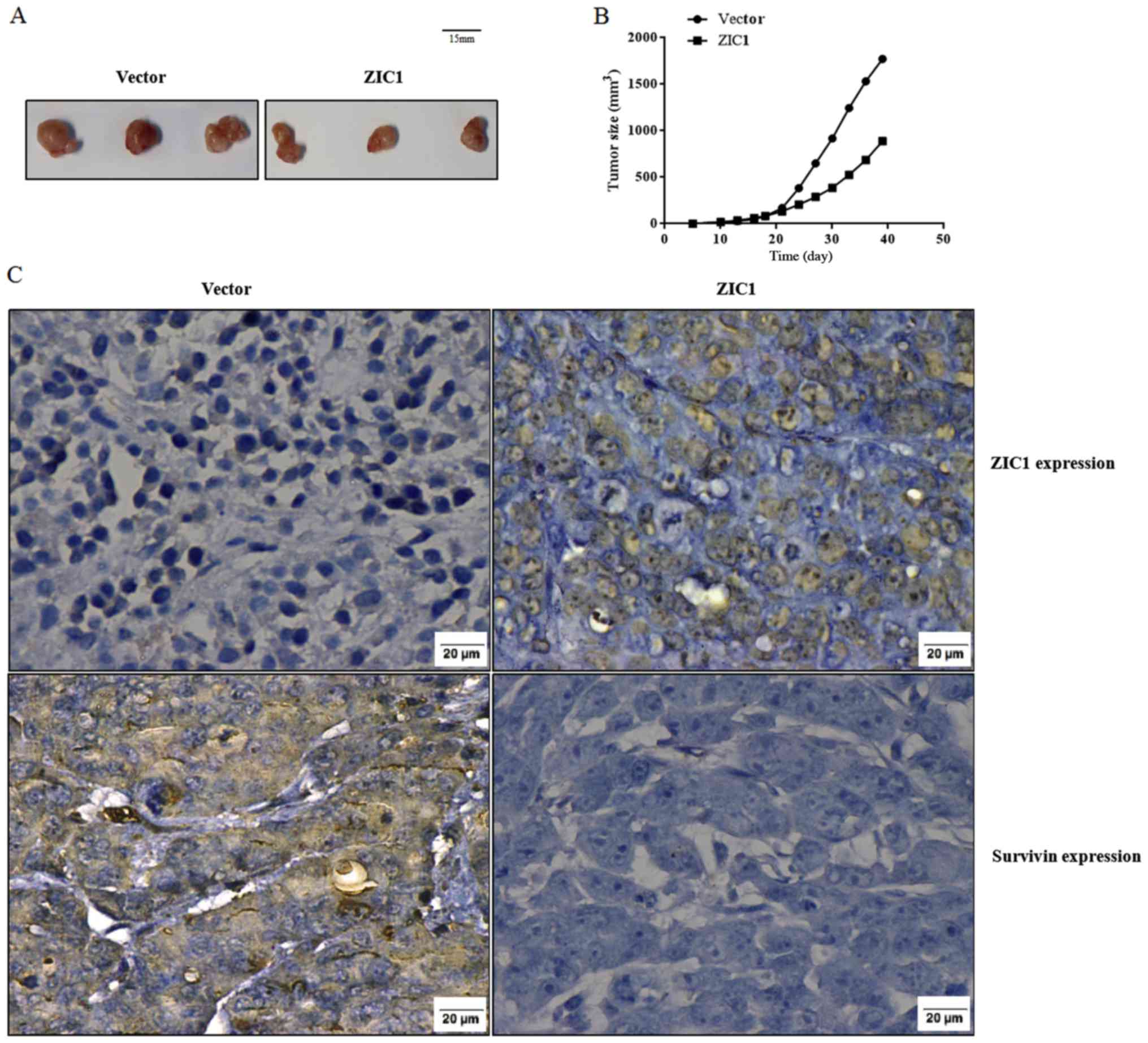
ZIC1 suppresses tumor growth in vivo
We subcutaneously transplanted rLV-ZsGreen-PGK-Puro-
or rLV-Zic1-PGK-Puro-transfected MDA-MB-231 cells into nude mice.
Each mouse had only one transplanted tumor. After 39 days of
transplantation, the tumors of the mice in the 'ZIC1' group were
consistently smaller than those of the mice in the 'Vector' group
(the maximum tumor diameter in the 'ZIC1' group was 1.26 cm, while
the maximum tumor diameter in the 'Vector' group was 1.60 cm;
Fig. 6A). In addition, the sizes
of the tumors during the period of 39 days were markedly lower in
the 'ZIC1' group, in comparison with the 'Vector' group (Fig. 6B). We also validated that high
levels of ZIC1 expression and low levels of survivin expression
were both observed in the 'ZIC1' group. Opposite results were
observed in the 'Vector' group (Fig.
6C).
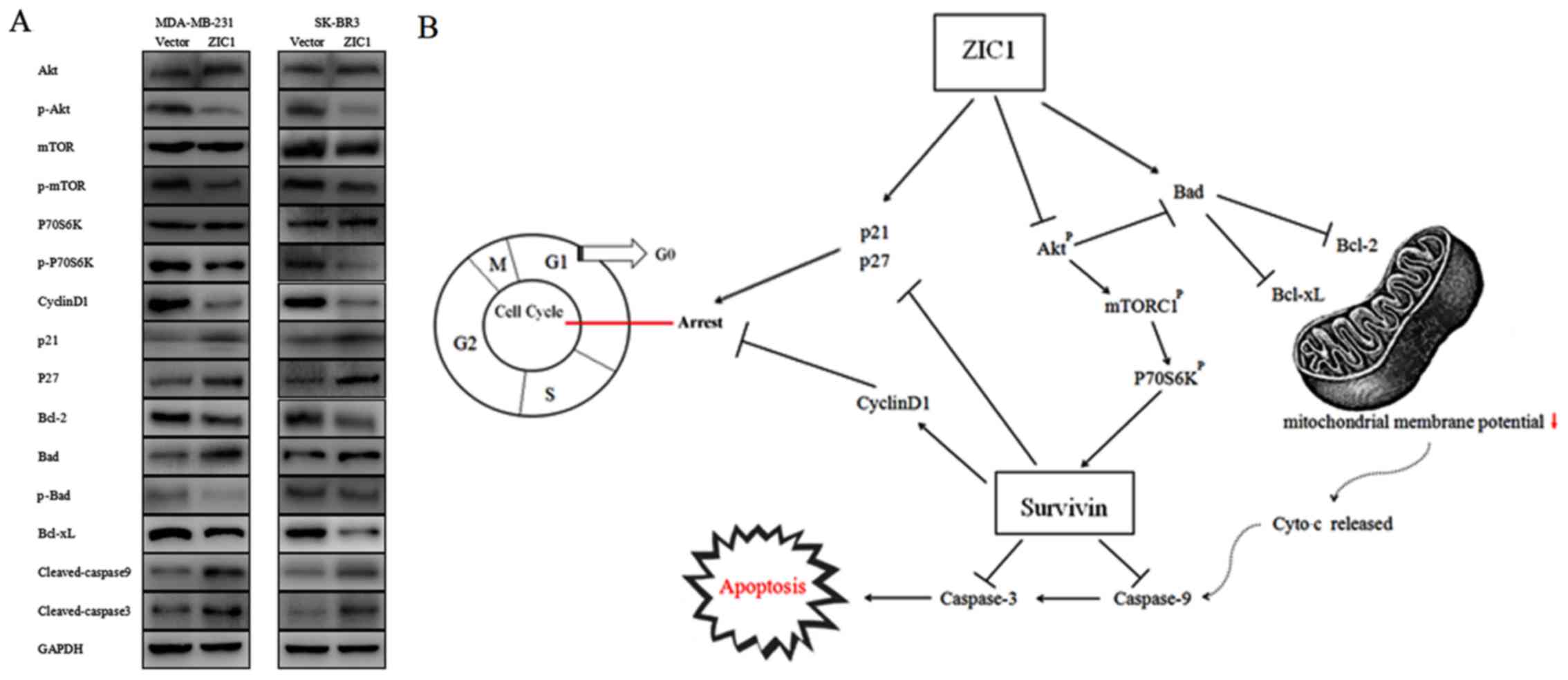
ZIC1 inhibits survivin expression by
inactivating the Akt/mTOR/P70S6K signaling pathway
To explore potential mechanisms underlying the
suppressive role of ZIC1 in breast cancer and its regulatory effect
on survivin expression, we investigated critical proteins of the
Akt signaling pathway, which play a pivotal role in cell
proliferation and survival in breast cancer (19,20).
As shown in Fig. 7A, the
overexpression of ZIC1 significantly reduced phosphorylation levels
of Akt, mTOR and P70S6K, and suppressed the activities of this
pathway both in the MDA-MB-231 and SK-BR3 breast cancer cells. In
addition, the ectopic expression of ZIC1 down-regulated cyclin D1
and upregulated p21 and p27 expression, which indicated that ZIC1
arrested the breast cancer cells at the G1 phase via modulating the
expression levels of these proteins. Our data also indicated that
the upregulation of ZIC1 led to the decreased expression of Bcl-2
and Bcl-xL, the dephosphorylation of Bad and the activation of
cleaved caspase-9 and cleaved caspase-3 (Fig. 7A). These results suggested that
ZIC1 reduces mitochondrial membrane potential and promotes the
apoptosis of breast cancer cells through the mitochondrial
apoptotic pathway. Combined with the molecular modulatory
mechanisms of survivin found in previous studies (17,21–23),
a schematic model of the potential mechanisms of ZIC1 protein is
presented in Fig. 7B.
Discussion
Due to changes in the environment and in spite of
advances in detection techniques, breast cancer has become a
leading threat to the lives of women worldwide. Numerous oncogenes
and anti-oncogenes in breast carcinogenesis are constantly
identified. ZIC1 is a zinc finger transcription factor involved in
neural development and serves as a tumor suppressor gene in solid
tumors (6–8). In this study, we first found that
ZIC1 expression in breast tumors was significantly lower than that
in normal tissues, both through immunohistochemistry and western
blot analysis, and ZIC1 expression was negatively associated with
tumor differentiation and TNM staging. In addition, the expression
levels of ZIC1 in 6 breast cancer cell lines were also lower than
those in the MCF-10A cells, a normal breast cell line. These
results suggest that ZIC1 may be a potential tumor suppressor in
breast cancer. Furthermore, a low ZIC1 expression predicted a poor
prognosis from the HPA.
To verify the tumor suppressive role of ZIC1 in
breast cancer cells, we transfected rLV-ZsGreen-PGK-Puro or
rLV-Zic1-PGK-Puro into two breast cell lines, MDA-MB-231 and
SK-BR3, in which the expression levels of ZIC1 were lower than in
the other cell lines. In vitro, the stably transfected cells
overexpressing ZIC1 had lower cell proliferative abilities and
elevated apoptotic rates. In addition, compared with the 'Vector'
group, a markedly smaller tumor growth was observed the 'ZIC1'
group, following the transplantation of stably transfected
MDA-MB-231 cells into nude mice. All the results revealed that ZIC1
may function as a tumor suppressor in breast carcinogenesis;
however, this should be confirmed in further studies. As reported
in a previous study, ZIC1 is a putative tumor suppressor gene and
is upregulated in breast cancer cells when phosphatidylinositol
glycan anchor biosynthesis, class X containing complex (PIGX)
inhibits the growth of transfected BT-549 cells (11). To the best of our knowledge, this
study was the first to identify the tumor suppressive function of
ZIC1 by using stable transfection technology in breast cancer.
In the mechanistic analysis, the expression levels
of p-Akt, p-mTOR, p-P70S6K, Cyclin D1, Bcl-2 and Bcl-xL were
markedly decreased in the 'ZIC1' group, while the expression levels
of p21, p27, Bad, cleaved caspase-9 and cleaved caspase-3 were
significantly increased. In addition, this study identified that
the mitochondrial membrane potential of the breast cancer cells,
transfected with rLV-Zic1-PGK-Puro, was markedly decreased, due to
the dysregulated expression of Bcl-2 family proteins. In addition,
Cyto-c in the mitochondria was released into the cytosol, which
regulated the activation of caspase-9 and caspase-3. Thus, we
confirmed that ZIC1 was a potential inhibitor of the
Akt/mTOR/P70S6K pathway, and a promoter of the activation of the
mitochondrial apoptotic pathway. Previous studies have also
demonstrated that ZIC1 is a regulator of the PI3K/Akt pathway, and
can lead to decreased phosphorylation levels of Akt (6–8).
However, these studies only found the regulatory effect of ZIC1 on
the expression of Bcl-2, Bax and Bad. None of these analyzed the
changes in the levels of mitochondrial membrane potential in cancer
cells. The findings of this study indicated that ZIC1
overexpression markedly suppressed tumor growth by modulating these
pathways.
As a member of the family of the inhibitors of
apoptosis proteins (IAPs), survivin forcefully inhibits cell
apoptosis and facilitates the activation and proliferation of
breast cancer cells (24).
Admittedly, survivin is a downstream target of NF-κB, which in turn
is activated by mTOR (25).
Furthermore, survivin is a potent inactivator of caspase-9 and
caspase-3 and is regulated via the Akt/mTOR signaling pathway
(16,17). In this study, in 120 female
patients with breast cancer, compared with ZIC1, the expression
level of survivin in the tumors was markedly higher, and a negative
correlation was observed between ZIC1 and surviving expression. In
addition, the role of survivin in the clinicopathological
characteristics was opposite to that of ZIC1. Both in breast cancer
cell lines and the breast cell line, the expression level of
survivin was inversely associated with that of ZIC1. In addition,
following transfection with rLV-Zic1-PGK-Puro, survivin expression
was markedly decreased in the stably transfected cells, as well as
in xenograft tumors. Therefore, we demonstrated that an elevated
ZIC1 expression suppressed the expression of survivin through the
Akt/mTOR/P70S6K pathway, and then activate caspase-9 and caspase-3,
so as to induce the apoptosis of breast cancer cells.
The promoter of survivin contains 3 cell
cycle-dependent element (CDE) and a cell-cycle gene homology region
(CHR), which induce survivin expression intensively at the G2/M
phase, and determine its significance in cell cycle control
(26). The silencing the
expression of survivin through RNAi has been shown to affect the
expression levels of Cyclin D1, p21 and p27, so that the cell cycle
is arrested at the G1 phase, which eventually induces the apoptosis
of hepatocellular carcinoma cells (27). In this study, the overexpression of
ZIC1 suppressed the expression of survivin via the inactivation of
the Akt/mTOR/P70S6K pathway. Consequently, decreased survivin
expression induced the expression of p21 and p27, and led to the
downregulation of Cyclin D1. Therefore, it can be hypothesized that
ZIC1 overexpression induces a G1 phase block directly via
modulating cell cycle related proteins, or indirectly by reducing
survivin expression.
Although Brill et al (28) revealed that ZIC1 contributed to the
pathogenesis of liposarcoma, recent evidence suggested that ZIC1
was downregulated due to hypermethylation in its promoter region in
a variety of carcinomas, including gastric cancer, thyroid cancer,
ovarian cancer and cervical cancer (6,29–31).
In cervical cancer, ZIC1 may become a promising methylation marker,
which is associated with a 3q gain for the detection of cervical
pre-cancer and cervical carcinoma (30). In ovarian cancer, the
hypermethylation of ZIC1 is associated with cisplatin resistance
and it may serve as a potential therapeutic target (31,32).
Therefore, the methylation of the ZIC1 promoter causes its low
expression in malignant cells, and is a key reason for
carcinogenesis. In addition, a lower expression of ZIC1 in gastric
carcinoma has been confirmed to be associated with a worse
prognosis (10). Based on its
molecular mechanisms of action, the DNA methylation of ZIC1 has
been used for clinicopathological diagnosis, targeted clinical
treatment, and prognostic evaluation in various solid neoplasms
(7).
In this study, we explored the main functions of
ZIC1 in cell proliferation and apoptosis and relevant downstream
genes, which suggested that ZIC1 may be a potential tumor
suppressor gene. However, the upstream regulatory mechanisms of
ZIC1 in inhibiting the growth of cancer cells or targeted drugs are
still at a preliminary stage. Following the transfection of BT-549
cells with siRNAs targeting the PIGX containing complex, Nakakido
et al found a strong suppression of the growth of breast
cancer cells in vitro, along with a significant upregulation
of ZIC1 expression through cDNA microarray analysis (11). In addition, SGI-110, a dinucleotide
combining 5-aza-dC and deoxyguanosine, was found to cause the
powerful hypomethylation of ZIC1 in female nude, athymic,
BALB/c-nu/nu mice through pyrosequencing analyses (32). The biweekly treatment regimen of
SGI-110 also markedly decreased xenograft tumor growth (32).
In this study, the MDA-MB-231 cell line was a
triple-negative breast cancer, while the SK-BR-3 cell line was a
human epidermal growth factor receptor 2 (HER2) overexpressing cell
line. We selected 6 different breast cancer cell lines and found
that the MDA-MB-231 and SK-BR3 expressed lower levels of ZIC1 than
the other cell lines. We used these two cell lines in order to
assess the role of ZIC1 more clearly. In addition, due to totally
different genomic background of these two cell lines, we
demonstrated that ZIC1 acted a tumor suppressor in breast cancer
without the influence of HER2 expression. However, the
overexpression had its limitation as it was not a physiological
event and further mechanistic effect remained to be proven. Due to
insufficient follow-ups, we did not evaluate the prognostic role of
ZIC1. However, from the HPA, 799 samples were included in the 'ZIC1
high' group, with a higher 5-year survival rate than the 'ZIC1 low'
group. Thus, we aim to conduct overall survival analysis with
larger samples in the future.
In conclusion, we confirmed that the expression
level of ZIC1 was decreased in breast cancer samples, as well as in
breast cancer cell lines, and an elevated ZIC1 expression
suppressed the growth of breast cancer cells and xenograft tumors,
along with a significant downregulation in surviving expression via
the Akt/mTOR/P70S6K pathway. Although further investigations are
required, our findings illustrate the tumor suppressive role of
ZIC1 in breast carcinoma and a negative association between ZIC1
and survivin.
Acknowledgments
Not applicable.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ding L, Zhang Z, Xu Y and Zhang Y:
Comparative study of Her-2, p53, Ki-67 expression and
clinicopathological characteristics of breast cancer in a cohort of
northern China female patients. Bioengineered. 8:383–392. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Parsa Y, Mirmalek SA, Kani FE, Aidun A,
Salimi-Tabatabaee SA, Yadollah-Damavandi S, Jangholi E, Parsa T and
Shahverdi E: A review of the clinical implications of breast cancer
biology. Electron Physician. 8:2416–2424. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ali RG, Bellchambers HM and Arkell RM:
Zinc fingers of the cerebellum (Zic): Transcription factors and
co-factors. Int J Biochem Cell Biol. 44:2065–2068. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sankar S, Yellajoshyula D, Zhang B, Teets
B, Rockweiler N and Kroll KL: Gene regulatory networks in neural
cell fate acquisition from genome-wide chromatin association of
Geminin and Zic1. Sci Rep. 6:374122016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhong J, Chen S, Xue M, Du Q, Cai J, Jin
H, Si J and Wang L: ZIC1 modulates cell-cycle distributions and
cell migration through regulation of sonic hedgehog, PI(3)K and
MAPK signaling pathways in gastric cancer. BMC Cancer. 12:2902012.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gan L, Chen S, Zhong J, Wang X, Lam EK,
Liu X, Zhang J, Zhou T, Yu J, Si J, et al: ZIC1 is downregulated
through promoter hypermethylation, and functions as a tumor
suppressor gene in colorectal cancer. PLoS One. 6:e169162011.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Qiang W, Zhao Y, Yang Q, Liu W, Guan H, Lv
S, Ji M, Shi B and Hou P: ZIC1 is a putative tumor suppressor in
thyroid cancer by modulating major signaling pathways and
transcription factor FOXO3a. J Clin Endocrinol Metab.
99:E1163–E1172. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang YY, Jiang JX, Ma H, Han J, Sun ZY,
Liu ZM and Xu ZG: Role of ZIC1 methylation in hepatocellular
carcinoma and its clinical significance. Tumour Biol. 35:7429–7433.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ma G, Dai W, Sang A, Yang X and Li Q:
Roles of ZIC family genes in human gastric cancer. Int J Mol Med.
38:259–266. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nakakido M, Tamura K, Chung S, Ueda K,
Fujii R, Kiyotani K and Nakamura Y: Phosphatidylinositol glycan
anchor biosynthesis, class X containing complex promotes cancer
cell proliferation through suppression of EHD2 and ZIC1, putative
tumor suppressors. Int J Oncol. 49:868–876. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tamm I, Wang Y, Sausville E, Scudiero DA,
Vigna N, Oltersdorf T and Reed JC: IAP-family protein survivin
inhibits caspase activity and apoptosis induced by Fas (CD95), Bax,
caspases, and anticancer drugs. Cancer Res. 58:5315–5320.
1998.PubMed/NCBI
|
|
13
|
Colnaghi R, Connell CM, Barrett RM and
Wheatley SP: Separating the anti-apoptotic and mitotic roles of
survivin. J Biol Chem. 281:33450–33456. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hamy AS, Bieche I, Lehmann-Che J, Scott V,
Bertheau P, Guinebretière JM, Matthieu MC, Sigal-Zafrani B, Tembo
O, Marty M, et al: BIRC5 (survivin): A pejorative prognostic marker
in stage II/III breast cancer with no response to neoadjuvant
chemotherapy. Breast Cancer Res Treat. 159:499–511. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Véquaud E, Desplanques G, Jézéquel P, Juin
P and Barillé-Nion S: Survivin contributes to DNA repair by
homologous recombination in breast cancer cells. Breast Cancer Res
Treat. 155:53–63. 2016. View Article : Google Scholar :
|
|
16
|
Wilson JM, Kunnimalaiyaan S,
Kunnimalaiyaan M and Gamblin TC: Inhibition of the AKT pathway in
cholangiocarcinoma by MK2206 reduces cellular viability via
induction of apoptosis. Cancer Cell Int. 15:132015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jin Q, Feng L, Behrens C, Bekele BN,
Wistuba II, Hong WK and Lee HY: Implication of AMP-activated
protein kinase and Akt-regulated survivin in lung cancer
chemopreventive activities of deguelin. Cancer Res. 67:11630–11639.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
19
|
Li Q, Liu J, Meng X, Pang R and Li J:
MicroRNA-454 may function as an oncogene via targeting AKT in
triple negative breast cancer. J Biol Res (Thessalon). 24:102017.
View Article : Google Scholar
|
|
20
|
Sauer SJ, Tarpley M, Shah I, Save AV,
Lyerly HK, Patierno SR, Williams KP and Devi GR: Bisphenol A
activates EGFR and ERK promoting proliferation, tumor spheroid
formation and resistance to EGFR pathway inhibition in estrogen
receptor-negative inflammatory breast cancer cells. Carcinogenesis.
38:252–260. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Du G, Cao D and Meng L: miR-21 inhibitor
suppresses cell proliferation and colony formation through
regulating the PTEN/AKT pathway and improves paclitaxel sensitivity
in cervical cancer cells. Mol Med Rep. 15:2713–2719. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ding JH, Yuan LY and Chen GA: Aspirin
enhances the cytotoxic activity of bortezomib against myeloma cells
via suppression of Bcl-2, survivin and phosphorylation of AKT.
Oncol Lett. 13:647–654. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wu ZH, Lin C, Liu MM, Zhang J, Tao ZH and
Hu XC: Src inhibition can synergize with gemcitabine and reverse
resistance in triple negative breast cancer cells via the AKT/c-Jun
pathway. PLoS One. 11:e01692302016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Khan Z, Khan AA, Yadav H, Prasad GBKS and
Bisen PS: Survivin, a molecular target for therapeutic
interventions in squamous cell carcinoma. Cell Mol Biol Lett.
22:82017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kawakami H, Tomita M, Matsuda T, Ohta T,
Tanaka Y, Fujii M, Hatano M, Tokuhisa T and Mori N: Transcriptional
activation of survivin through the NF-kappaB pathway by human
T-cell leukemia virus type I tax. Int J Cancer. 115:967–974. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lens SM, Vader G and Medema RH: The case
for survivin as mitotic regulator. Curr Opin Cell Biol. 18:616–622.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu W, Zhu F, Jiang Y, Sun D, Yang B and
Yan H: siRNA targeting survivin inhibits the growth and enhances
the chemosensitivity of hepatocellular carcinoma cells. Oncol Rep.
29:1183–1188. 2013. View Article : Google Scholar
|
|
28
|
Brill E, Gobble R, Angeles C,
Lagos-Quintana M, Crago A, Laxa B, Decarolis P, Zhang L, Antonescu
C, Socci ND, et al: ZIC1 overexpression is oncogenic in
liposarcoma. Cancer Res. 70:6891–6901. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Rodríguez-Rodero S, Fernández AF,
Fernández-Morera JL, Castro-Santos P, Bayon GF, Ferrero C,
Urdinguio RG, Gonzalez-Marquez R, Suarez C, Fernández-Vega I, et
al: DNA methylation signatures identify biologically distinct
thyroid cancer subtypes. J Clin Endocrinol Metab. 98:2811–2821.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Verlaat W, Snijders PJ, Novianti PW,
Wilting SM, De Strooper LM, Trooskens G, Vandersmissen J, Van
Criekinge W, Wisman GB, Meijer CJ, et al: Genome-wide DNA
methylation profiling reveals methylation markers associated with
3q gain for detection of cervical pre-cancer and cancer. Clin
Cancer Res. 23:3813–3822. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Du L, Qian X, Dai C, Wang L, Huang D, Wang
S and Shen X: Screening the molecular targets of ovarian cancer
based on bioinformatics analysis. Tumori. 101:384–389. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Fang F, Munck J, Tang J, Taverna P, Wang
Y, Miller DF, Pilrose J, Choy G, Azab M, Pawelczak KS, et al: The
novel, small-molecule DNA methylation inhibitor SGI-110 as an
ovarian cancer chemo-sensitizer. Clin Cancer Res. 20:6504–6516.
2014. View Article : Google Scholar : PubMed/NCBI
|