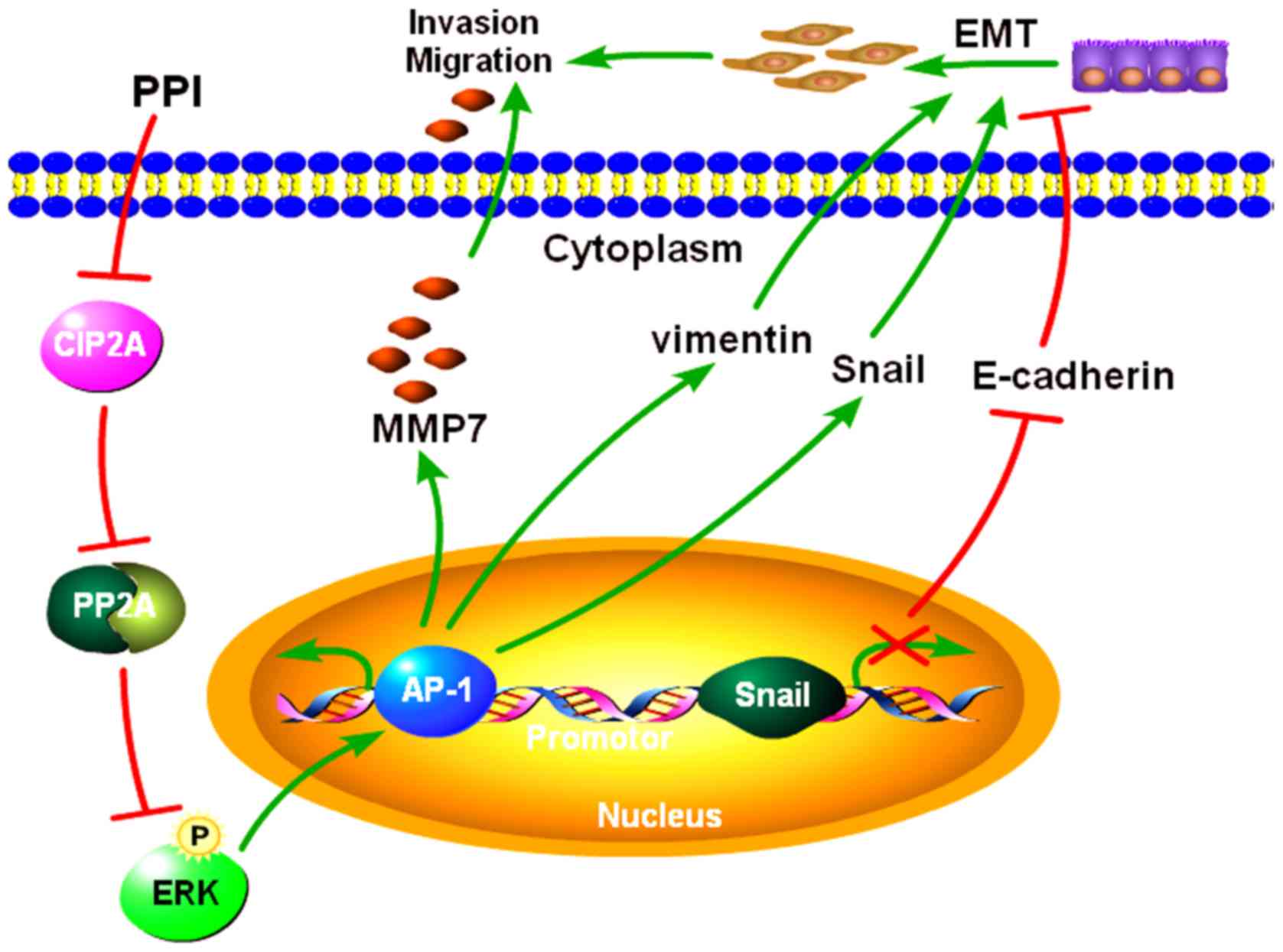
Introduction
Prostate cancer (PC) is a common malignancy
diagnosed in men. In 2012, a total of ~1,1111,700 cases of PC were
newly diagnosed, and 307,500 mortality cases have been estimated
worldwide (1). The morbidity of PC
is increasing in China in recent years (2). Although patients diagnosed with
localized PC exhibit a survival rate of >5 years, the majority
of patients with PC will progress to castration-resistant PC, and
eventually, relapse (3).
Therefore, novel drugs that may suppress the tumor initiation,
progression and metastasis of PC are required.
Oncogenes and anti-oncogenes serve pivotal roles in
the pathogenesis of several cancer types. Cancerous inhibitor of
protein phosphatase 2A (CIP2A) is an oncoprotein that suppresses
protein phosphatase 2A (PP2A) activity in the degradation of c-Myc
protein in cancer cells (4). CIP2A
is overexpressed in numerous types of human cancer, including PC,
gastric, colon, breast, head and neck, lung and ovarian cancer
(4–10). Additionally, CIP2A overexpression
is associated with malignant proliferation and aggressiveness
(11). De et al suggested a
role of oncogenic nexus of CIP2A in carcinogenesis (12). This nexus is created by
CIP2A-mediated inhibition of PP2A, which activates oncogenic signal
factors including Akt, extracellular signaling-related kinase (ERK)
and c-Myc, and thus may serve as a biomarker and therapeutic target
for the treatment of cancer. Recently, Wu et al reported
that CIP2A physically associates with H-Ras and activates the
MEK/ERK signaling pathway, thus promoting epithelial-mesenchymal
transition (EMT) and progression of cervical cancer (13). Therefore, targeting CIP2A may be a
promising therapeutic strategy for cancer.
Increasing evidences has shown that natural products
not only serve key roles in the detection and evolvement of novel
drugs, but they may be used as molecular probes for screening
treatment targets (14,15). Paris polyphyllin is a medicinal
herb in the Shennongjia National Nature Reserve of China, which may
treat fevers, headaches, burns, wounds, and exhibit
immune-regulatory and antitumor abilities (16,17).
Polyphyllin I (PPI), a bioactive phytochemical isolated from the
rhizoma of P. polyphylla, exhibits preclinical antitumor
efficacy in several cancer types, including breast, lung, gastric,
and ovarian cancer, as well as osteosarcoma (16,18,19).
Recently, PPI was found to increase the sensitivity of liver cancer
HepG2 cells to cisplatin, and activate autophagy via the
PI3K/Akt/mTOR signaling pathway (20). The antitumor activity of PPI on PC
cells has not previously been evaluated. The aim of the present
study is to evaluate the antitumor effects and potential molecular
mechanisms of PPI in PC.
Materials and methods
Reagents
PPI with a purity of 98% was purchased from Shanghai
YuanYe Bio-Technology Co., Ltd. (Shanghai, China). PPI was
dissolved in dimethylsulfoxide (DMSO; Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) at a stock solution of 4 mM and stored at
−20°C. Okadaic acid (OA) was purchased from Sigma-Aldrich; Merck
KGaA.
Cell culture
Human PC cancer cell lines PC3 and DU145 were
purchased from American Type Culture Collection (ATCC; Manassas,
VA, USA) and cultured in Dulbecco's modified Eagle's medium (DMEM;
high glucose; Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) containing 10% fetal bovine serum (FBS; HyClone, Logan, UT,
USA), 100 U/ml penicillin and 100 µg/ml streptomycin (both
from Gibco; Thermo Fisher Scientific, Inc.) at 37°C in a humidified
atmosphere containing 5% CO2.
MTT assay
A total of 6×104 cells were seeded into a
96-well plate and pre-cultured with DMEM for 24 h, then treated
with PPI (0.4–3 µM) for 24 h at 37°C in a humidified
atmosphere containing 5% CO2. Cell cytotoxicity was
determined using MTT assay. The absorbance was measured at 490 nm
using an automated microplate reader (BioTek Instruments, Inc.,
Winooski, VT, USA), and the inhibition rate was calculated as
follows: Inhibition rate (%) = (average A490 of the
control group-average A490 of the experimental
group)/(average A490 of the control group-average
A490 of the blank group) ×100%. Cell viability was
estimated using trypan blue dye exclusion.
Soft agar colony formation assay
A total of 1×103 cells were suspended in
1 ml of DMEM containing 0.3% agarose (Amresco, Cleveland, OH, USA)
and 10% FBS, and plated on a bottom layer containing 0.6% agarose
and 10% FBS in each of the triplicate wells of a 6-well plate.
Colonies formed on the soft agar plates were stained with 0.2%
crystal violet and counted under light microscope (IX70; Olympus
Corporation, Tokyo, Japan) after 2 weeks.
Invasion assay
Invasion assays were performed using a 24-well
Transwell chamber (Corning Incorporated, Corning, NY, USA).
Polyvinylpyrrolidone-free polycarbonate filters (8 µm pore
size) (Corning Incorporated) were coated with Matrigel (BD
Biosciences, Franklin Lakes, NJ, USA). The lower chamber contained
medium supplemented with 20% FBS. Cells (1×104
cells/well) were placed into the upper chamber with the coated
membrane. After incubation for 20 h at 37°C, the invaded cells at
the bottom of the membrane were fixed and stained with 2% ethanol
containing 0.2% crystal violet (15 min) at room temperature for 20
min and observed under light microscope (×10 objective). For
quantification, the invaded stained cells on the other side of the
membrane were extracted with 33% acetic acid. The absorbance of the
eluted stain was determined at 570 nm.
Wound healing assay
Cells (4×105 cells/2 ml) were seeded in a
6-well plate and incubated at 37°C until they reach 90–100%
confluence. Confluent cells were scratched using a 200 µl
pipet tip. After washing with PBS, cells were treated with
serum-free medium for 24 h. Then, the cells were fixed and stained
with 2% ethanol containing 0.2% crystal violet powder at room
temperature for 20 min.
Reverse transcription quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from cells using TRIzol
reagent (Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. A total of 2 µg RNA was
reverse-transcribed into cDNA using ReverTra Ace qPCR RT Kit
(Toyobo Life Science, Osaka, Japan), according to the
manufacturer's protocol. The reverse transcription steps as follow:
37°C for 15 min and 98°C for 5 min. The samples were stored at
−20°C. qPCR was performed using the SYBR® Green
Real-Time PCR Master Mix (Toyobo Life Science) and the ABI
StepOnePlus™ Real-Time PCR System (ABI; Thermo Fisher Scientifc,
Inc.), according to the manufacturer's protocol. GAPDH was used as
an endogenous control. The PCR thermocycling conditions were as
follows: Initial denaturation at 95°C for 3 min followed by 40
cycles of 95°C for 15 sec and extension at 60°C for 1 min. The
threshold cycle for each sample was selected from the linear range
and converted to a starting quantity by interpolation from a
standard curve generated on the same plate for each set of primers
(Table I). Epithelial
(E)-cadherin, vimentin, matrix metalloproteinase
7 (MMP7), MMP9 and CIP2A mRNA levels were
normalized for each well to GAPDH mRNA levels using the
2−ΔΔCq method (21).
Each experiment was repeated three times.
 | Table IPrimer sequences for reverse
transcription quantitative polymerase chain reaction. |
Table I
Primer sequences for reverse
transcription quantitative polymerase chain reaction.
| Gene | Primer sequence
(5′-3′) |
|---|
|
E-cadherin | F:
CAGGTCTCCTCATGGCTTTGC |
| R:
CTTCCGAAAAGAAGGCTGTCC |
|
Vimentin | F:
ACTACGTCCACCCGCACCTA |
| R:
CAGCGAGAAGTCCACCGAGT |
| MMP7 | F:
GAGGCATGAGTGAGCTACAGTG |
| R:
ACATCTGGGCTTCTGCATTATT |
| MMP9 | F:
CCGGACCAAGGATACAGTT |
| R:
CGGCACTGAGGAATGATCTA |
| CIP2A | F:
TGCGGCACTTGGAGGTAATTTC |
| R:
AGCTCTACAAGGCAACTCAAGC |
| GAPDH | F:
TGTTGCCATCAATGACCCCTT |
| R:
CTCCACGACGTACTCAGCG |
Western blot analysis
Cells were lysed in radioimmunoprecipitation assay
buffer containing 50 mM Tris, pH 8.0, 150 mM NaCl, 0.1% SDS, 0.5%
sodium deoxycholate, 1% NP-40, 1 mM DTT, 1 mM NaF, 1 mM sodium
orthovanadate, 1 mM phenyl-methylsulfonyl fluoride (PMSF;
Sigma-Aldrich; Merck KGaA) and 1% protease inhibitors cocktail (EMD
Millipore, Billerica, MA, USA). Protein concentration was
determined using the Bradford method. Equal amounts of sample (25
µg) were separated by SDS-PAGE (8–12% gels). Electrophoresed
proteins were then transferred onto polyvinylidene fluoride
membranes (EMD Millipore). The membranes were blocked with 5%
skimmed milk in Tris-buffered saline at room temperature for 1 h.
Following blocking, membranes were incubated overnight at 4°C with
primary antibodies and then rinsed with Tris-buffered saline with
Tween-20. The following primary antibodies were used:
anti-E-cadherin (1:2,000; 20874-1-AP), anti-vimentin (1:5,000;
10366-1-AP), anti-Snail (1:3,000; 13099-1-AP), anti-MMP7 (1:1,000;
10374-2-AP), anti-insulin-like growth factor-binding protein 3
(IGFBP3; 1:1,000; 10189-2-AP), anti-proliferating cell nuclear
antigen (PCNA; 1:5,000; 10205-2-AP) (both from ProteinTech Group,
Inc., Chicago, IL, USA); anti-MMP9 (1:500; sc-6840), anti-CIP2A
(1:500; sc-80662); anti-Fos-related antigen 1 (FRA-1; 1:500;
sc-271657) (Santa Cruz Biotechnology, Santa Cruz, CA, USA),
anti-PP2A (1:1,000; 2038), anti-ERK1/2 (1:1,000; 9102),
anti-phospho-ERK1/2 (Thr202/Tyr204) (1:1,000; 9101), anti-c-Fos
(1:1,000; 2250; Cell Signaling Technology, Inc., Danvers, MA, USA),
and anti-GAPDH (1:5,000; M20006; Abmart, Shanghai, China).
Membranes were then washed, and incubated with horseradish
peroxidase-conjugated secondary antibody (1:1,000; E030120-01 and
E030110-01; EarthOx, LLC, San Francisco, CA, USA) for 1.5 h at room
temperature. The protein bands were visualized using
SuperSignal® West Pico PLUS Chemiluminescent substrate
(34579; Pierce; Thermo Fisher Scientific, Inc.) (22). Densitometric quantification of the
western blots was determined using ImageJ software (version
1.4.3.67; National Institutes of Health, Bethesda, MD, USA).
Luciferase reporter assay
DU145 and PC3 cells were seeded in 12-well culture
plates. When cells reached 70% confluence, they were transfected
with pAP-1-Luc plasmids (Beyotime Institute of Biotechnology,
Haimen, China) using Lipofectamine 3000 (Thermo Fisher Scientifc,
Inc.), according to manufacturer's protocol. Following transfection
for 4 h, the cells were treated with PPI (0.6–1.8 µM) for 20
h. Firefly luciferase activities were assayed using the Luciferase
Assay System (Promega Biotech Co., Ltd., Beijing, China) according
to the manufacturer's protocol.
Chromatin immunoprecipitation (ChIP)
assay
DU145 and PC3 cells (2×105 cells/ml)
treated with PPI for 24 h were used for ChIP assay using a
ChIP-IT® Express Enzymatic Shearing kit (Active Motif,
Beijing, China), according to the manufacturer's protocol.
DNA-bound protein was immunoprecipitated using an anti-c-Fos
(component of AP-1) antibody or rabbit IgG as a control. The AP-1
binding site of MMP7 promoter was detected by RT-qPCR using
the following primers: 5′-CCAGATGTTGCAGAATACTCACTA-3′ (forward) and
5′-GATCCACTGTAATATGCGGTAAGT-3′ (reverse). The AP-1 binding site of
vimentin promoter was detected by RT-qPCR using the
following primers: 5′-CGAGCTGGGCAGTAGAGAGT-3′ (forward) and
5′-CCAAGACATGGAGGGAGTGT-3′ (reverse).
PP2A activity assay
PP2A immunoprecipitation phosphatase assay kit
(Upstate Biotechnology, Inc., Lake Placid, NY, USA) was used
according to the manufacturer's protocol in order to evaluate
phosphate release as an index of phosphatase activity. Briefly, 100
µg protein isolated from cells was incubated with 4
µg anti-PP2A monoclonal antibody overnight. A total of 4
µl protein A agarose beads were added and the mixture was
incubated at 4°C for 2 h. Subsequently, the beads were collected
and washed three times with 700 µl ice-cold TBS and one time
with 500 µl Ser/Thr assay buffer. The beads were further
incubated with 750 mM phosphopeptide in Ser/Thr assay buffer for 10
min at 30°C with constant agitation. A total of 100 µl
Malachite Green Phosphate Detection solution was added and the
absorbance at 650 nm was measured using a microplate reader.
Xenograft studies
Male nude immunodeficient mice (nu/nu) (weighing ~16
g, 5-week-old) were purchased from Hunan SJA Laboratory Animal Co.,
Ltd. (Changsha, China), and maintained under specific pathogen-free
conditions. Mice were given free access to sterile water and to a
standard diet, and maintained under controlled conditions of
temperature (22–24°C), humidity (40–70%), light (12-h light-dark
cycle). The present study was approved by the Hubei University of
Medicine Animal Care and Use Committee (Hubei, China) and performed
in compliance with the Regulations for the Administration of
Affairs Concerning Experimental Animals. The mice were
subcutaneously injected with human PC DU145 cells (6×106
cells which were suspended in 100 µl DMEM media) into the
right flank of each mouse. Treatments started when tumors reached a
palpable size (0.5 cm in diameter). Mice were randomly divided into
the following two groups: control group (vehicle; 0.8% DMSO, 12%
cremophor and 8% ethanol in normal saline; n=8) and PPI-treated
group (intraperitoneal injection of 1 mg/kg PP1; n=8). The mice
were treated 5 times per week for a total of 30 weeks. Caliper
measurements of the longest perpendicular tumor diameters were
performed twice a week to estimate the tumor volume, using the
following formula: 4π/3 × (width/2)2 × (length/2),
representing the 3-dimensional volume of an ellipse. Animals were
sacrificed when tumors reached 1.5 cm or if the mice appeared
moribund to prevent unnecessary morbidity to the mice.
Statistical analysis
All experiments were repeated at least three times
and the data are presented as the mean ± standard deviation (SD).
All statistical analyses were conducted using GraphPad Prism5
(GraphPad Software, Inc., La Jolla, CA, USA) and SPSS 22.0 (IBM
Corp., Armonk, NY, USA). Results were analyzed using unpaired
Student's t-test or one-way analysis of variance followed by
Bonferroni post-test. P<0.05 was considered to indicate a
statistically significant difference. All experiments were repeated
at least three times.
Results
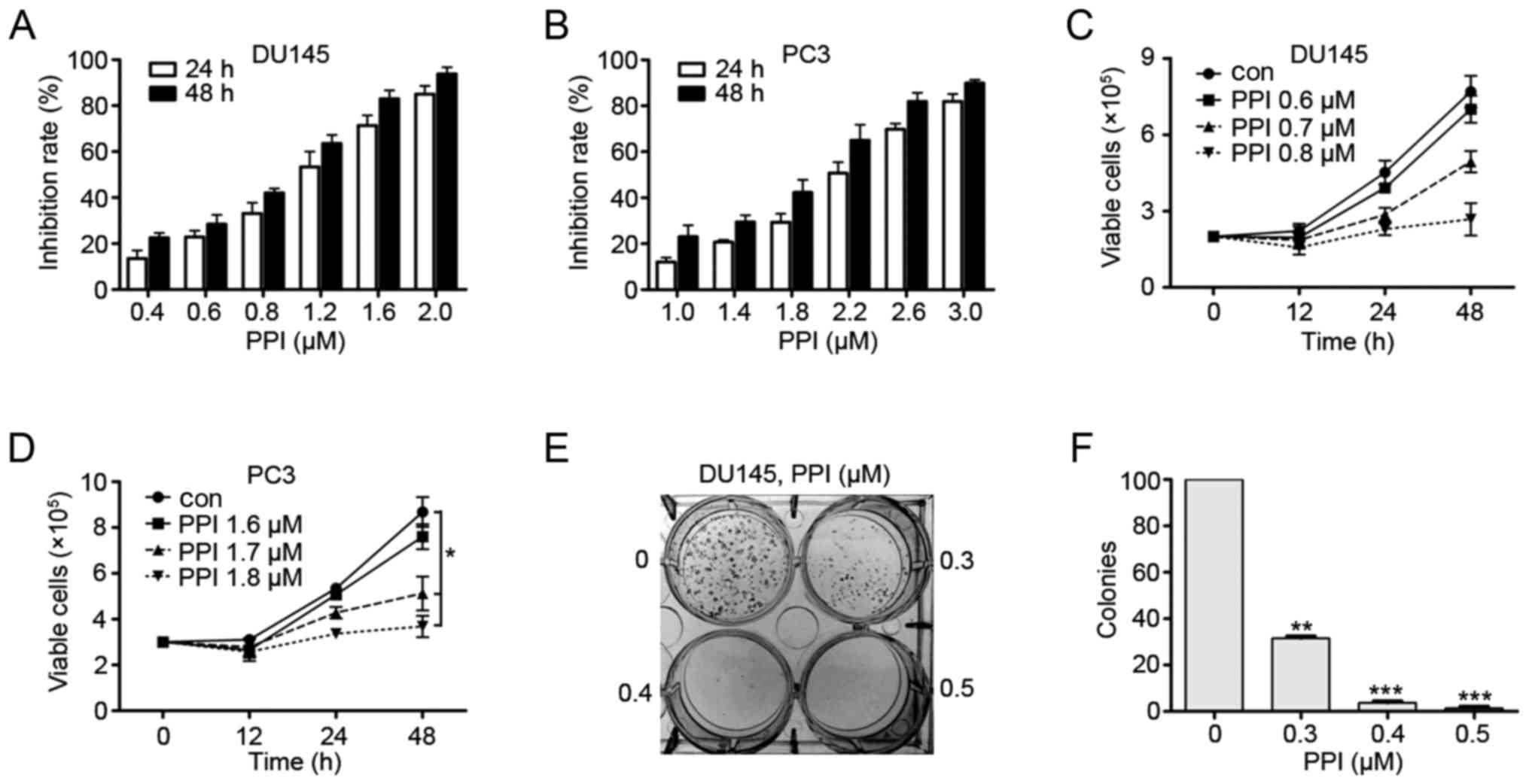
Effects of PPI on the viability of PC
cell lines
DU145 and PC3 cells were seeded into 96-well plates
for 24 h and then treated with PPI (Fig. 1). After 24 or 48 h, the cell
survival rate was detected using the MTT assay. The results
suggested that PPI exhibited moderate cytotoxicity to PC cells. The
IC50 values of PPI against DU145 and PC3 cells were 1.03
µM and 2.13 µM, respectively (Fig. 2A and B). Trypan blue exclusion
assay showed that PPI decreased viable DU145 and PC3 cells in a
dosage- and time-dependent manner (Fig. 2C and D). Next, colony formation
assay showed that PPI markedly suppressed the clonogenic ability of
DU145 cells (Fig. 2E and F). These
results suggest that PPI may decrease anchorage-dependent (cell
viability) and anchorage-independent (colony formation) growth of
PC cells.
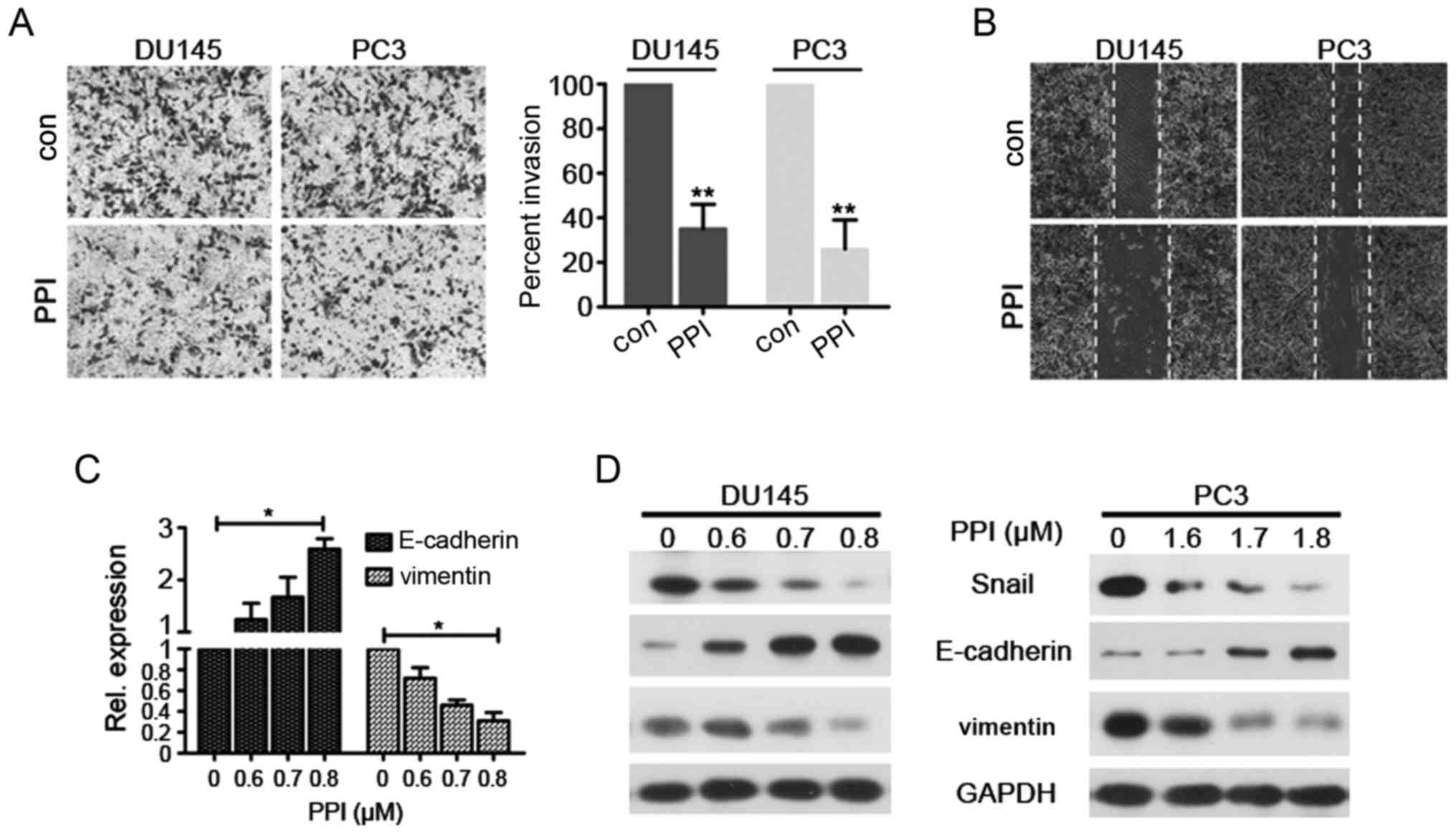
PPI suppresses the invasion and migration
of PC cells as well as reverses EMT
In the present study the effects of PPI on the
invasion and migration of PCs (DU145 and PC3) cells were
investigated. The results demonstrated that PPI suppressed the
invasion of DU145 and PC3 cells (Fig.
3A). Additionally, cell migration was assessed using a wound
healing assay. DU145 and PC3 cells were treated with PPI for 24 h.
The results revealed that PPI decreased DU145 and PC3 cell
migration (Fig. 3B). These results
suggest that PPI may exhibit an anti-invasive effect on PCs at low
concentrations.
EMT is an important process in tumor metastasis.
During EMT, cells lose their polarized immotile epithelial
characteristics and acquire motile mesenchymal features leading to
increased invasion and motility (23). Moreover, the transition is
characterized by a decrease in the expression of epithelial markers
(including E-cadherin) as well as an increase in the expression of
mesenchymal markers (including Snail and vimentin) (23,24).
To determine whether PPI treatment may reverse EMT and inhibit PC
metastasis in vitro, the expression of EMT-associated
markers were assessed in PPI-treated DU145 and PC3 cells. The
results demonstrated an increased expression of E-cadherin and a
decreased expression of vimentin in PPI-treated DU145 and PC3 cells
compared with the control cells (Fig.
3C and D). These results indicate that PPI treatment may
suppress EMT. The effects of PPI treatment on EMT-associated
transcription factors were investigated and it was revealed that
PPI treatment decreased the expression levels of Snail (Fig. 3C and D). Therefore, these results
suggest that PPI upregulates E-cadherin expression, and
downregulates the expression of Snail and vimentin.
PPI inhibits the invasion and EMT in PC
cells by downregulating MMP7, vimentin and Snail via the ERK
signaling pathway
Tumor invasion and migration are associated with
increased expression levels of matrix MMPs. MMP7 and MMP9 are two
critical enzymes which contribute to cell invasion and migration
(25,26). Thus, mRNA and protein expression
levels of MMP7 and MMP9 were assessed (Fig. 4A–D). PPI treatment decreased MMP7
expression but it had no effect on MMP9 expression in DU145 and PC3
cells, suggesting that PPI may inhibit the invasion and migration
of PC cells, by inhibiting MMP7.
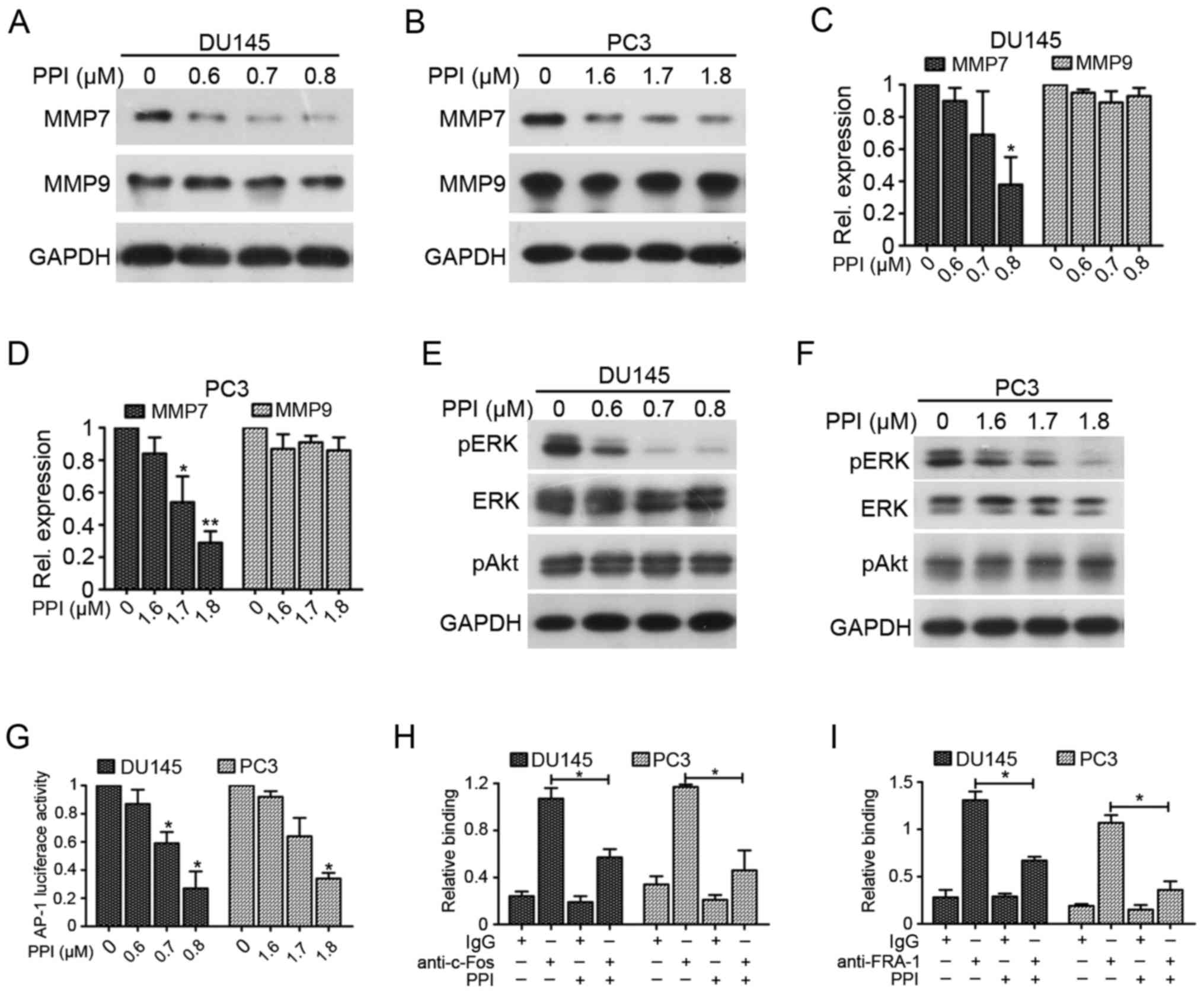 | Figure 4PPI suppresses PC cell invasion
through the downregulation of MMP7 via ERK/AP-1 pathway. Western
blot analysis for MMP7 and MMP9 expression in (A) DU145 and (B) PC3
cells were treated with PPI for 24 h. RT-qPCR analysis of mRNA
levels of MMP7 and MMP9 in (C) DU145 and (D) PC3
cells treated with PPI for 24 h. *P<0.05 and
**P<0.01 vs. 0 µM. Western blot analysis of
pERK, ERK and pAkt expression in (E) DU145 and (F) PC3 cells were
incubated with PPI at the indicated concentrations for 24 h. (G)
DU145 and PC3 cells were transfected with the AP-1 luciferase
reporter construct for 4 h and then treated with PPI. Luciferase
activity was measured using the luciferase assay system.
*P<0.05 vs. 0 µM. (H) PPI inhibited binding of
c-Fos to MMP7 promoter. Chromatins were extracted from DU145
and PC3 cells treated with or without PPI (DU145 cells, 0.7
µM PPI; PC3 cells, 1.7 µM PPI). Chromatin
immunoprecipitation was performed by using anti-c-Fos antibody, and
the amplification was carried out by RT-qPCR.
*P<0.05. (I) PPI inhibited binding of FRA-1 to
vimentin promoter. Chromatins were extracted from DU145 and
PC3 cells treated with or without PPI (DU145 cells, 0.7 µM
PPI; PC3 cells, 1.7 µM PPI). Chromatin immunoprecipitation
was performed by using anti-FRA-1 antibody, and the amplification
was carried out by RT-qPCR. RT-qPCR, reverse transcription
quantitative polymerase chain reaction; PPI, polyphyllin I; ERK,
extracellular signaling-related kinase; p, phosphorylated; MMP,
matrix metal-loproteinase; FRA, Fos-related antigen 1. |
Expression of MMP7, vimentin, and Snail may be
regulated by the ERK signaling pathway, which serves an important
role in invasion and EMT of PC cells (26–28).
Therefore, the activation of ERK in PC cells treated with various
PPI concentrations was assessed. The results demonstrated that PPI
decreased the phosphorylation levels of ERK in a dose-dependent
manner without affecting the total ERK expression and Akt
phosphorylation in DU145 and PC3 cells (Fig. 4E and F). AP-1, a key transcription
factor that promotes MMP7 and vimentin transcription,
is regulated by MAPKs (including ERK, JNK and P38 kinases)
(27,29). Luciferase reporter assay was
performed to investigate the effect of PPI on the transcriptional
activity of AP-1 and the results indicated that PPI decreased AP-1
transcription activity (Fig. 4G).
ChIP assays showed that PPI decreased c-Fos binding to the
MMP7 promoter (Fig. 4H).
Additionally, PPI decreased FRA-1 (component of AP-1) binding to
the vimentin promoter (Fig.
4I). These results demonstrate that PPI may inhibit invasion
and EMT in PC cells through the ERK/AP-1 signaling pathway.
Effects of PPI treatment on the
CIP2A/PP2A/ERK signaling axis
CIP2A is an oncogenic PP2A inhibitor, which is
highly expressed in PC cells and mediates PC progression and
castration-resistance (30).
Western blot analysis was used to evaluate the effects PPI
treatment on CIP2A protein expression. Fig. 5A shows that the expression of CIP2A
was decreased in DU145 cells treated with 0.7 µM PPI and
could not be detected in response to 0.8 µM PPI.
Additionally, the expression of CIP2A was downregulated in response
to treatment with 1.7 µM PPI in PC3 cells, and treatment
with 1.8 µM PPI led to an elimination of CIP2A expression
(Fig. 5B). In addition, the
results indicated that PPI treatment may downregulates CIP2A
expression in a time-dependent manner (Fig. 5C and D). The dephosphorylation of
ERK is frequently regulated by PP2A, which is the downstream factor
of CIP2A, therefore whether PPI may affect PP2A activity was
assessed. Fig. 5E showed that the
activity of PP2A was increased in PPI-treated PC cells, indicating
that PPI may influence the CIP2A/PP2A/ERK signaling axis. Moreover,
a PP2A inhibitor, OA, was used for the in vitro experiments.
The results indicated that OA reversed the cell growth and invasion
that were inhibited by PPI treatment (Fig. 5F and G). Additionally, combined
treatment with OA and PPI increased the expression levels of
phosphorylated ERK and upregulated MMP7, Snail, vimentin expression
compared with those in PPI-treated cells (Fig. 5H and I). In conclusion, these
results demonstrate that reactivation of PP2A may be required for
PPI-inhibited invasion, MMP7, Snail, vimentin expression and ERK
phosphorylation via CIP2A inhibition.
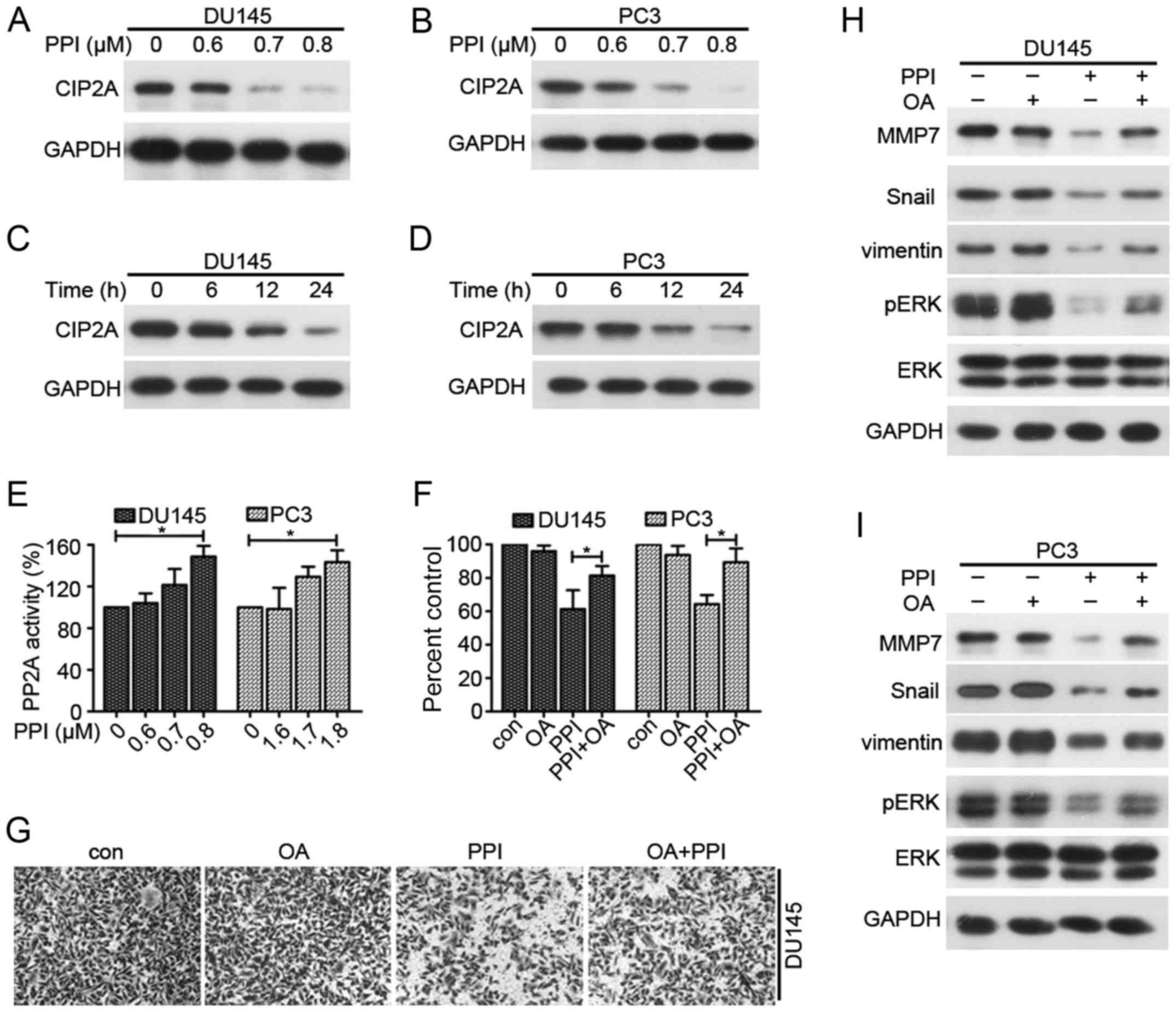 | Figure 5Effects of PPI on CIP2A/PP2A/ERK
signaling axis. Western blot analysis of CIP2A expression in (A)
DU145 and (B) PC3 cells treated with PPI for 24 h. Western blot
analysis of CIP2A expression in PPI-treated (C) DU145 and (D) PC3
cells for the indicated time-points. PP2A activity in (E) DU145 and
PC3 cells treated with indicated concentrations of PPI for 24.
*P<0.05. (F) MMT assay in DU145 and PC3 cells were
treated with PPI (0.7 or 1.7 µM) and/or OA (50 nM) for 24 h.
*P<0.05. (G) DU145 cells were treated with PPI (0.6
µM) and/or OA (50 nM) for 20 h and their invasive ability
was examined. Western blot analysis of MMP7, Snail, vimentin, pERK
and ERK in (H) DU145 and (I) PC3 cells treated with PPI and/or OA
for 24 h. PPI, polyphyllin I; ERK, extracellular signaling-related
kinase; p, phosphorylated; MMP, matrix metalloproteinase; CIP2A,
cancerous inhibitor of protein phosphatase 2A; OA, okadaic acid;
PP2A, protein phosphatase 2A. |
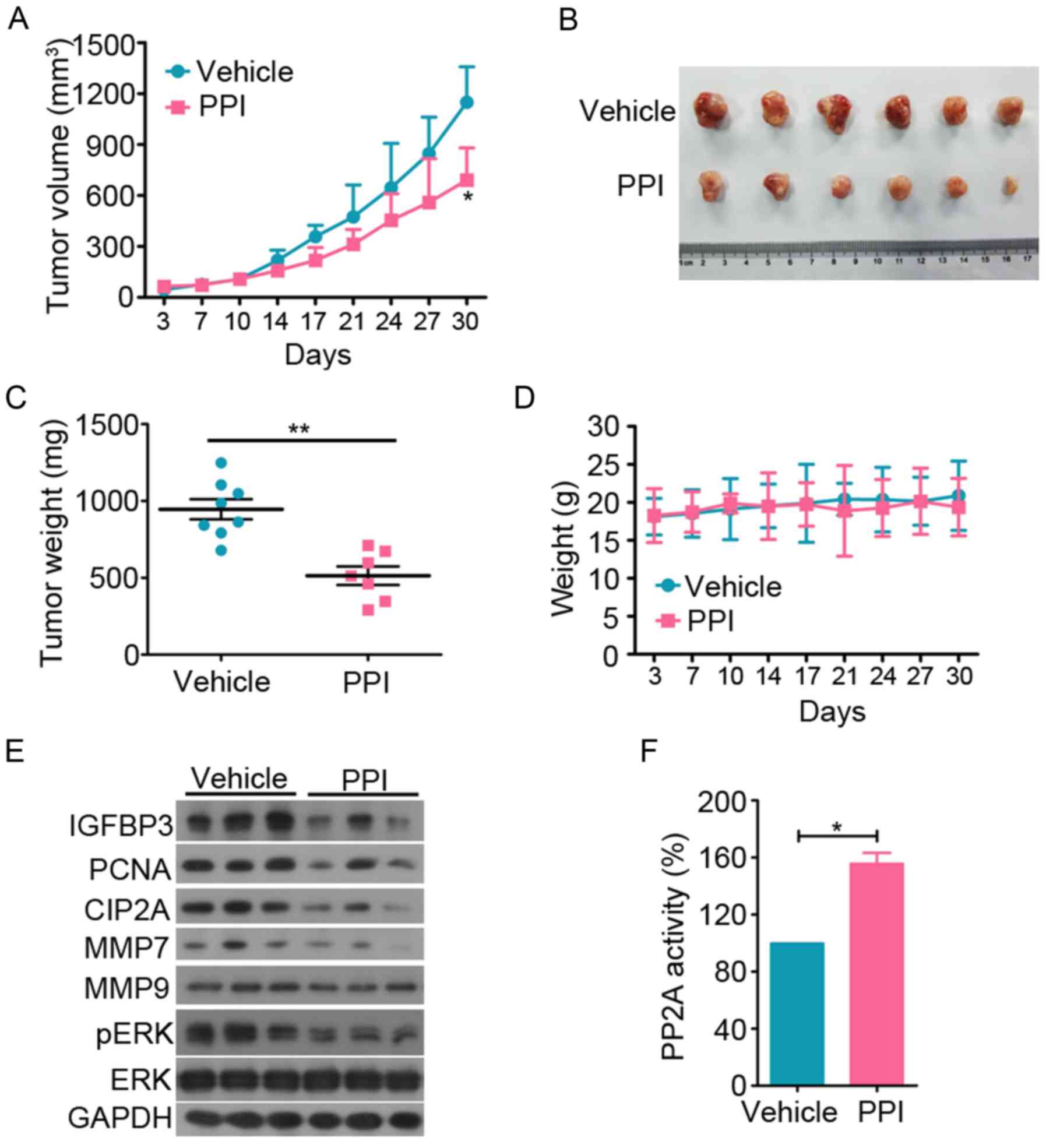
PPI suppresses tumor growth in vivo
To investigate the antitumor effect of PPI on PC
in vivo, 6×106 DU145 cells in 100 µl of
DMEM medium were subcutaneously inoculated in the right flank of
nude mice. The mice were divided into vehicle or PPI-treated
groups. The results indicated that PPI treatment significantly
suppressed tumor growth compared to the vehicle control (P<0.05;
Fig. 6A and B). PPI treatment also
decreased the tumor weight of the mice (Fig. 6C). PPI treatment did not affect the
body weight of the mice (Fig. 6D).
Western blot analysis and PP2A activity assay revealed that the
expression levels of pERK, MMP7, CIP2A and PCNA were decreased in
the PPI-treated group (Fig. 6E).
IGFBP3 is a critical biomarker involved in biological pathways
thought to be important in pathogenesis and progression of PC
(31). IGFBP3 expression was
detected in tumor specimens and it was found that PPI decreased
IGFBP3 expression (Fig. 6E). The
PP2A activity of PPI-treated group was significantly upregulated
(Fig. 6F).
Based on these results, PPI may suppress PC
viability, invasion and EMT by inhibiting the CIP2A/PP2A/ERK
signaling cascade (Fig. 7).
Discussion
PC is the second major cause of cancer mortality in
males worldwide, which is due to late-stage diagnosis and
inefficient therapy, especially for recurrent disease (25,32).
In Chinese traditional medicine, various compounds which are
present in natural herbs may have an anticancer potential due to
their ability to inhibit tumor growth, metastasis and angiogenesis
without having significant side effects. Therefore, TCM may be an
important approach for reducing the tumor incidence and mortality
by preventing, reversing or delaying the process of tumorigenesis
(22,33,34).
PPI is a compound found in Paris polyphylla rhizomes. PPI
has been reported to exhibit antiproliferative and anticancer
activities according to previous studies (16,18,19).
Therefore, the mode of action and potential utility of PPI was
explored in PC cells in search of a novel, effective anticancer
agent.
In the present study, the effects of PPI were
investigated using DU145 and PC3 cell lines. MTT and trypan blue
dye exclusion assays suggested that PPI may suppress the growth of
PC cells. Furthermore, PPI may inhibit the colony formation ability
of PC cells. Therefore, PPI was shown to exert anti-proliferative
effects in human PC cells.
Inhibition of tumor metastasis and invasion is
critical to improving the prognosis of patients with PC (26). Therefore, treatment strategies for
prevention or suppression of tumor metastasis and invasion may
ameliorate the survival of patients with PC. The results of the
present study suggest that PPI may suppress the invasive and
migratory abilities of DU145 and PC3 cells. Tumor metastasis is a
multifactor, multistage and multistep process regulated by multiple
factors and genes. EMT serves a key role in cancer metastasis
(35). Therefore, in the present
study the expression of EMT-associated markers was investigated and
it was found that PPI-mediated mobility inhibition was associated
with EMT reversion.
Another finding of the present study was that PPI
may inhibit the expression of MMP7. Metastasis and invasion of
tumor cells involves the following processes: adhesion, enzymolysis
and movement. MMP7 is a membrane protein in the MMP family
(29). This type of protease
degrades the extracellular matrix and basement membrane, which are
important mediators for the metastasis and invasion of cancer cells
(26). Previous studies have
confirmed that transcription factors, including Snail, Slug and
ZEB1, which act as E-cadherin repressors, are involved in promoting
EMT (36). And MMP7, vimentin and
Snail expression is regulated by the ERK signaling pathway
(27–29). In the present study the effects of
PPI on the phosphorylation level of ERK were examined. The results
demonstrated that PPI may inhibit ERK phosphorylation in DU145 and
PC3 cells. The transcription factor AP-1 may form either a homo- or
hetero- complex which can bind as c-fos/c-fos, c-jun/c-jun or
c-fos/c-jun to activate endogenous or exogenous motivation
(37). AP-1 is also a critical
transcriptional modulator in ERK signal transduction (15). Moreover, the MMP7 and vimentin
promoter region contains a cis-regulatory element of one AP-1
binding site. Thus, the effects of PPI on the AP-1 transcription
activity and binding ability to the MMP7 or vimentin
promoter was assessed. The results revealed that PPI may suppress
AP-1 transcriptional activity and AP-1 binding ability to the
MMP7 and vimentin promoter. In conclusion, these
results suggest that the anti-invasive effect of PPI may be
mediated by suppressing AP-1 transcription activities and the
ERK/MMP7 and ERK/EMT signal.
CIP2A is a oncogene that may facilitate cell
transformation and tumorigenesis via inhibiting PP2A activities, as
it is a protein phosphatase that regulates several critical
oncogenic signal proteins including Akt, c-Myc, ERK and p70S6K
(38–40). The results of the present study
revealed that PPI may downregulate CIP2A expression and upregulate
PP2A activity. It was speculated that PPI may inhibit CIP2A/PP2A
signal to downregulate ERK phosphorylation and the expression of
MMP7, vimentin, and Snail, and then decrease PC cell EMT and
invasion. To further determine the effect of PP2A activation in
PPI-mediated inhibition of invasion and downregulation of MMP7, PCs
were treated with PPI and OA, an inhibitor of PP2A. The results
showed that inactivation of PP2A may antagonize the effect of PPI
in EMT and invasion through the ERK pathway. These results
confirmed that the CIP2A/PP2A signaling axis serves a crucial
function in PC proliferation, invasion, and EMT that is mediated by
inhibiting ERK activity. ERK activation may be induced by PPI. In a
xenograft mouse model, PPI significantly repressed tumor growth,
whereas the body weight of the mice remained unaffected. The
results also demonstrated that PPI suppressed pERK, MMP7, IGFBP3
and CIP2A expression. Therefore, PPI may be considered a novel
anticancer agent in preventing and treating PC.
Acknowledgments
Not applicable.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siegel RL, Miller KD and Jemal A: Cancer
Statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Junttila MR, Puustinen P, Niemelä M, Ahola
R, Arnold H, Böttzauw T, Ala-aho R, Nielsen C, Ivaska J, Taya Y, et
al: CIP2A inhibits PP2A in human malignancies. Cell. 130:51–62.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Khanna A, Böckelman C, Hemmes A, Junttila
MR, Wiksten JP, Lundin M, Junnila S, Murphy DJ, Evan GI, Haglund C,
et al: MYC-dependent regulation and prognostic role of CIP2A in
gastric cancer. J Natl Cancer Inst. 101:793–805. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Teng HW, Yang SH, Lin JK, Chen WS, Lin TC,
Jiang JK, Yen CC, Li AF, Chen PC, Lan YT, et al: CIP2A is a
predictor of poor prognosis in colon cancer. J Gastrointest Surg.
16:1037–1047. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Vaarala MH, Väisänen MR and Ristimäki A:
CIP2A expression is increased in prostate cancer. J Exp Clin Cancer
Res. 29:1362010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Böckelman C, Lassus H, Hemmes A, Leminen
A, Westermarck J, Haglund C, Bützow R and Ristimäki A: Prognostic
role of CIP2A expression in serous ovarian cancer. Br J Cancer.
105:989–995. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tseng LM, Liu CY, Chang KC, Chu PY, Shiau
CW and Chen KF: CIP2A is a target of bortezomib in human triple
negative breast cancer cells. Breast Cancer Res. 14:R682012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu Z, Ma L, Wen ZS, Hu Z, Wu FQ, Li W,
Liu J and Zhou GB: Cancerous inhibitor of PP2A is targeted by
natural compound celastrol for degradation in non-small-cell lung
cancer. Carcinogenesis. 35:905–914. 2014. View Article : Google Scholar
|
|
11
|
Côme C, Laine A, Chanrion M, Edgren H,
Mattila E, Liu X, Jonkers J, Ivaska J, Isola J, Darbon JM, et al:
CIP2A is associated with human breast cancer aggressivity. Clin
Cancer Res. 15:5092–5100. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
De P, Carlson JH, Leyland-Jones B and Dey
N: Role of 'oncogenic nexus' of CIP2A in breast oncogenesis: How
does it work? Am J Cancer Res. 5:2872–2891. 2015.
|
|
13
|
Wu Y, Gu TT and Zheng PS: CIP2A cooperates
with H-Ras to promote epithelial-mesenchymal transition in
cervical-cancer progression. Cancer Lett. 356:646–655. 2015.
View Article : Google Scholar
|
|
14
|
Cai F, Zhang L, Xiao X, Duan C, Huang Q,
Fan C, Li J, Liu X, Li S and Liu Y: Cucurbitacin B reverses
multidrug resistance by targeting CIP2A to reactivate protein
phosphatase 2A in MCF-7/adriamycin cells. Oncol Rep. 36:1180–1186.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Xiao X, He Z, Cao W, Cai F, Zhang L, Huang
Q, Fan C, Duan C, Wang X, Wang J, et al: Oridonin inhibits
gefitinib-resistant lung cancer cells by suppressing
EGFR/ERK/MMP-12 and CIP2A/Akt signaling pathways. Int J Oncol.
48:2608–2618. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li GB, Fu RQ, Shen HM, Zhou J, Hu XY, Liu
YX, Li YN, Zhang HW, Liu X, Zhang YH, et al: Polyphyllin I induces
mitophagic and apoptotic cell death in human breast cancer cells by
increasing mitochondrial PINK1 levels. Oncotarget. 8:10359–10374.
2017.PubMed/NCBI
|
|
17
|
Chen JC, Hsieh MJ, Chen CJ, Lin JT, Lo YS,
Chuang YC, Chien SY and Chen MK: Polyphyllin G induce apoptosis and
autophagy in human nasopharyngeal cancer cells by modulation of AKT
and mitogen-activated protein kinase pathways in vitro and in vivo.
Oncotarget. 7:70276–70289. 2016.PubMed/NCBI
|
|
18
|
He H, Zheng L, Sun YP, Zhang GW and Yue
ZG: Steroidal saponins from Paris polyphylla suppress adhesion,
migration and invasion of human lung cancer A549 cells via
downregulating MMP-2 and MMP-9. Asian Pac J Cancer Prev.
15:10911–10916. 2014. View Article : Google Scholar
|
|
19
|
Yue G, Wei J, Qian X, Yu L, Zou Z, Guan W,
Wang H, Shen J and Liu B: Synergistic anticancer effects of
polyphyllin I and evodiamine on freshly-removed human gastric
tumors. PLoS One. 8:e651642013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shi YM, Yang L, Geng YD, Zhang C and Kong
LY: Polyphyllin I induced-apoptosis is enhanced by inhibition of
autophagy in human hepatocellular carcinoma cells. Phytomedicine.
22:1139–1149. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
22
|
Cao W, Liu Y, Zhang R, Zhang B, Wang T,
Zhu X, Mei L, Chen H, Zhang H, Ming P, et al: Homoharringtonine
induces apoptosis and inhibits STAT3 via IL-6/JAK1/STAT3 signal
pathway in Gefitinib-resistant lung cancer cells. Sci Rep.
5:84772015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bucay N, Bhagirath D, Sekhon K, Yang T,
Fukuhara S, Majid S, Shahryari V, Tabatabai Z, Greene KL, Hashimoto
Y, et al: A novel microRNA regulator of prostate cancer
epithelial-mesen-chymal transition. Cell Death Differ.
24:1263–1274. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Montanari M, Rossetti S, Cavaliere C,
D'Aniello C, Malzone MG, Vanacore D, Di Franco R, La Mantia E,
Iovane G, Piscitelli R, et al: Epithelial-mesenchymal transition in
prostate cancer: An overview. Oncotarget. 8:35376–35389. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen CM, Lin CL, Chiou HL, Hsieh SC, Lin
CL, Cheng CW, Hung CH, Tsai JP and Hsieh YH: Loss of endothelial
cell-specific molecule 1 promotes the tumorigenicity and metastasis
of prostate cancer cells through regulation of the TIMP-1/MMP-9
expression. Oncotarget. 8:13886–13897. 2017.PubMed/NCBI
|
|
26
|
Zhang Q, Liu S, Parajuli KR, Zhang W,
Zhang K, Mo Z, Liu J, Chen Z, Yang S, Wang AR, et al:
Interleukin-17 promotes prostate cancer via MMP7-induced
epithelial-to-mesenchymal transition. Oncogene. 36:687–699. 2017.
View Article : Google Scholar :
|
|
27
|
Andreolas C, Kalogeropoulou M, Voulgari A
and Pintzas A: Fra-1 regulates vimentin during Ha-RAS-induced
epithelial mesenchymal transition in human colon carcinoma cells.
Int J Cancer. 122:1745–1756. 2008. View Article : Google Scholar
|
|
28
|
Zhao J, Ou B, Han D, Wang P, Zong Y, Zhu
C, Liu D, Zheng M, Sun J, Feng H, et al: Tumor-derived CXCL5
promotes human colorectal cancer metastasis through activation of
the ERK/Elk-1/Snail and AKT/GSK3β/β-catenin pathways. Mol Cancer.
16:702017. View Article : Google Scholar
|
|
29
|
Chang MC, Chen CA, Chen PJ, Chiang YC,
Chen YL, Mao TL, Lin HW, Lin Chiang WH and Cheng WF: Mesothelin
enhances invasion of ovarian cancer by inducing MMP-7 through
MAPK/ERK and JNK pathways. Biochem J. 442:293–302. 2012. View Article : Google Scholar
|
|
30
|
Khanna A, Rane JK, Kivinummi KK, Urbanucci
A, Helenius MA, Tolonen TT, Saramäki OR, Latonen L, Manni V,
Pimanda JE, et al: CIP2A is a candidate therapeutic target in
clinically challenging prostate cancer cell populations.
Oncotarget. 6:19661–19670. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Terracciano D, Bruzzese D, Ferro M,
Mazzarella C, Di Lorenzo G, Altieri V, Mariano A, Macchia V and Di
Carlo A: Preoperative insulin-like growth factor-binding protein-3
(IGFBP-3) blood level predicts gleason sum upgrading. Prostate.
72:100–107. 2012. View Article : Google Scholar
|
|
32
|
Terracciano D, Mazzarella C, Di Carlo A,
Mariano A, Ferro M, Di Lorenzo G, Giordano A, Altieri V, De Placido
S and Macchia V: Effects of the ErbB1/ErbB2 kinase inhibitor GW2974
on androgen-independent prostate cancer PC-3 cell line growth and
NSE, chromogranin A and osteopontin content. Oncol Rep. 24:213–217.
2010.PubMed/NCBI
|
|
33
|
Feng T, Cao W, Shen W, Zhang L, Gu X, Guo
Y, Tsai HI, Liu X, Li J, Zhang J, et al: Arctigenin inhibits STAT3
and exhibits anticancer potential in human triple-negative breast
cancer therapy. Oncotarget. 8:329–344. 2017.
|
|
34
|
Liu Y, Dong Y, Zhang B and Cheng YX: Small
compound 6-O-angeloylplenolin induces caspase-dependent apoptosis
in human multiple myeloma cells. Oncol Lett. 6:556–558. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Cao Z, Koochekpour S, Strup SE and
Kyprianou N: Reversion of epithelial-mesenchymal transition by a
novel agent DZ-50 via IGF binding protein-3 in prostate cancer
cells. Oncotarget. 8:78507–78519. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chen HN, Yuan K, Xie N, Wang K, Huang Z,
Chen Y, Dou Q, Wu M, Nice EC, Zhou ZG, et al: PDLIM1 stabilizes the
E-cadherin/β-catenin complex to prevent epithelial-mesenchymal
transition and metastatic potential of colorectal cancer cells.
Cancer Res. 76:1122–1134. 2016. View Article : Google Scholar
|
|
37
|
Wen-Sheng W: ERK signaling pathway is
involved in p15INK4b/p16INK4a expression and HepG2 growth
inhibition triggered by TPA and Saikosaponin a. Oncogene.
22:955–963. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Rincón R, Cristóbal I, Zazo S, Arpí O,
Menéndez S, Manso R, Lluch A, Eroles P, Rovira A, Albanell J, et
al: PP2A inhibition determines poor outcome and doxorubicin
resistance in early breast cancer and its activation shows
promising therapeutic effects. Oncotarget. 6:4299–4314. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Tsukamoto S, Huang Y, Umeda D, Yamada S,
Yamashita S, Kumazoe M, Kim Y, Murata M, Yamada K and Tachibana H:
67-kDa laminin receptor-dependent protein phosphatase 2A (PP2A)
activation elicits melanoma-specific antitumor activity overcoming
drug resistance. J Biol Chem. 289:32671–32681. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wang J, Okkeri J, Pavic K, Wang Z, Kauko
O, Halonen T, Sarek G, Ojala PM, Rao Z, Xu W, et al: Oncoprotein
CIP2A is stabilized via interaction with tumor suppressor PP2A/B56.
EMBO Rep. 18:437–450. 2017. View Article : Google Scholar : PubMed/NCBI
|