Introduction
Colorectal cancer (CRC) is one of the most common
malignancies and is the leading cause of cancer-related mortality
worldwide (1). Although advanced
therapies are increasingly being developed, the 5-year survival
rate for CRC remains <50% (2).
In addition, it is notable that 90% of cancer mortalities are
attributed to metastasis and not the primary tumor in general
(3). Therefore, clarification of
the molecular mechanisms underlying metastasis and the
identification of key molecules involved in the process are of
great clinical value.
Metastasis requires cells to acquire migratory and
invasive capabilities in order to travel from the primary tumor to
a secondary site (4).
Epithelial-mesenchymal transition (EMT) serves a critical role
during this process, and primarily involves the following steps: i)
Dissociation of points of adhesion between epithelial cells; ii)
loss of the apical-basolateral polarity; iii) reorganization of the
actin cytoskeleton; and iv) an increase in cell motility (5). A recent study suggested that
dysregulation of the actin cytoskeleton may be a hallmark of
subsequent metastatic dissemination (6), which may indicate cytoskeletal
regulators as potential targets for CRC therapies based on
metastasis inhibition.
Profilin 2 (PFN2) belongs to a class of small
G-actin-binding proteins and is a well-characterized regulator of
actin polymerization (7). PFN2 was
previously considered to be expressed mainly in the neurons of
vertebrates (8). However, altered
expression of PFN2 has been reported in different types of cancer;
for example, PFN2 expression was downregulated in oral squamous
cell carcinoma (9). These data
indicated that PFN2 may serve a role in cancer progression and
metastasis; however, its expression and function are largely
unknown in CRC.
In the present study, the expression levels and the
effects of PFN2 were investigated in the development of human CRC,
particularly in regard to metastatic capability. PFN2 expression
was observed to be downregulated in patients with CRC with
metastasis, as well as in metastatic CRC cell lines. In addition,
alterations in the expression of PFN2 affected the EMT process and
the metastasis of CRC. Furthermore, the results indicated that
cytoskeletal reorganization may be involved in PFN2-regulated
metastasis. Finally, clinical data suggested that tumors with low
PFN2 expression were significantly associated with poor prognosis.
Results from the present study indicated that PFN2 may function as
a negative regulator of CRC metastasis and, thus, may represent a
potential target for CRC therapy.
Materials and methods
Patients and specimens
The present study was approved by the Biomedical
Research Ethics Committee of Jiading District Central Hospital
Affiliated Shanghai University of Medicine & Health Sciences
(Shanghai, China). Informed consent was obtained from all study
subjects before sample collection and these samples were used
according to ethical standards. CRC and normal paracancerous tissue
samples were collected from 70 patients (38 male, 32 female; age,
48–72) that underwent surgery at the Department of Surgery, Jiading
District Central Hospital Affiliated Shanghai University of
Medicine & Health Sciences between February 2008 and October
2011; none of the patients received any preoperative treatment.
Patients with familial adenomatous polyposis, inflammatory bowel
diseases and primary tumors in other tissues were excluded. The
histological sections were reviewed by two expert pathologists to
verify the histological diagnosis. For reverse
transcription-quantitative polymerase chain reaction (RT-qPCR) and
western blot analysis, 3 groups of human colorectal tissues
(n=10/group; 21 male, 9 female; age 49–81), including normal
colorectal tissues, non-metastatic CRC tissues and metastatic CRC
tissues) were freshly collected at the Department of Surgery,
Jiading District Central Hospital Affiliated Shanghai University of
Medicine & Health Sciences between January 2015 and February
2017. Tumors were staged according to the American Joint Committee
on Cancer pathological tumor-node-metastasis classification
(10).
Cell culture
The human CRC cell lines SW620 and HCT116 were
purchased from the American Type Culture Collection (ATCC,
Manassas, VA, USA). Preliminary western blotting experiments were
conducted to examine the expression of PFN2 in several CRC cell
lines, and it was determined that PFN2 protein expression levels
were highest in HCT116 cells and lowest in SW620; subsequently,
HCT116 and SW620 were selected as representative CRC cell lines for
use in the present study. These cell lines were routinely
maintained in the laboratory according to the instructions from
ATCC. Briefly, cells were cultured in 6- or 12-well plates (BD
Biosciences, Franklin Lakes, NJ, USA) and grown as a monolayer.
HCT116 cells were cultured at 37°C in a humidified incubator with
5% CO2 in McCoy's 5A (Modified) medium (Gibco; Thermo
Fisher Scientific, Inc., Waltham, MA, USA). SW620 cells were
cultured at 37°C in an incubator without CO2 in
Leibovitz's L 15 Medium (Gibco; Thermo Fisher Scientific, Inc.).
Both of the media were supplemented with 10% fetal bovine serum
(FBS; Thermo Fisher Scientific, Inc.) and 100 U/ml penicillin and
100 µg/ml streptomycin (Thermo Fisher Scientific, Inc.).
In certain experiments, the cell-permeable small
molecule Rho-associated kinase inhibitor Y27632 (3 µmol/l;
Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) was added into the
cell culture medium for 24 h to inhibit myosin light chain (MLC)
phosphorylation.
Cell line transfection and
transduction
The PFN2 overexpression vector was constructed as
described previously (9).
Full-length human PFN2 cDNA was amplified by RT-PCR using the
forward primer 5′-GCGGCCGCATGGCCGGTTGGCAGAGCTACG-3′ and the reverse
primer 5′-GGATCCTTACACATCAGACCTCCTCAG-3′. To construct the
overexpression vector, the PFN2 coding sequence was excised from
the pCR-Blunt plasmid (Invitrogen; Thermo Fisher Scientific, Inc.)
by NotI and BamHI digestion, and subcloned into the
pQCXIH expression vector (Clontech Laboratories, Inc., Mountain
View, CA, USA). To characterize the PFN2 expression vector,
pQCXIH-PFN2 (PFN2-OE) or empty vector (vector) was transfected into
SW620 cells using Lipofectamine® 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.), according to the manufacturer's
protocol. Briefly, SW620 cells were seeded (2×105
cells/well) in a 6-well plate 24 h prior to transfection and
cultured to ~80% confluency, at 37°C in an incubator without
CO2. Subsequently, SW620 cells were transfected with 1.5
µg of plasmid and 2.5 µl of Lipofectamine 2000 in
6-well plates, and then cultured at 37°C in an incubator without
CO2 for 24 h, then harvested for further
experiments.
Liver and lung metastasis of CRC model
mice
Animal experiments were approved by the Animal
Experiment Administration Committee of the Soochow University
(Suzhou, China), and experiments were conducted in accordance with
The National Institutes of Health guidelines for animal care during
the study. Male BALB/c nude mice (age, 8–10 weeks; average weight,
24.0 g) were obtained from the Shanghai Laboratory Animal Center at
the Chinese Academy of Sciences (Shanghai, China). They were
maintained in a specific pathogen free room with a constant
temperature (20–26°C), relative humidity (40–70%), normal
atmosphere (with 21% O2 and 0.03% CO2; 15
times per hour ventilation), 12-h light-dark cycle and free access
to food and water.
To establish the liver cancer metastasis model, mice
were anesthetized, a transverse incision was made in the left flank
to expose the spleen and 1×106 PFN2-OE-transfected or
empty vector-transfected SW620 cells in 100 µl
phosphate-buffered saline (PBS; Thermo Fisher Scientific, Inc.)
were injected intrasplenically (n=5 mice/group). For the lung
cancer metastasis model, the mice were anesthetized and
1×106 PFN2-OE-transfected or empty vector-transfected
SW620 cells in 100 µl of PBS were injected via the tail vein
(5 mice/group). During the 6 weeks following injections, body
weight and survival of the mice was monitored; the body weight of
each mouse was measured once per week. The volume of the largest
metastatic tumor nodules was calculated using the formula: Volume =
(width2 × length) / 2. All mice were euthanized 6 weeks
post-injection.
RT-qPCR
Total RNA was extracted from tissues or cells using
TRIzol (Invitrogen; Thermo Fisher Scientific, Inc.) and 1 µg
RNA was subsequently reverse transcribed using the AMV Reverse
Transcription System (Takara Bio Inc., Otsu, Japan). qPCR was
performed using the FastStart Universal SYBR Green Master with Rox
(Roche Diagnostics, Basel, Switzerland) and an ABI PRISM 7900HT
system (Applied Biosystems; Thermo Fisher Scientific, Inc.), with
the following thermocycling conditions: an initial 2-min incubation
at 50°C, followed by 10 min at 95°C and 40 cycles of 95°C for 15
sec and 60°C for 60 sec. The following primers were used: PFN2,
forward 5′-ATGATTGTAGGAAAAGACCGGGA-3′, reverse
5′-GCAGTCACCATCGACGTATAGAC-3′; epithelial (E)-cadherin, forward
5′-CGAGAGCTACACGTTCACGG-3′, reverse 5′-GGGTGTCGAGGGAAAAATAGG-3′;
neural (N)-cadherin, forward 5′-AGCCAACCTTAACTGAGGAGT-3′, reverse
5′-GGCAAGTTGATTGGAGGGATG-3′; vimentin, forward
5′-GACGCCATCAACACCGAGTT-3′, reverse 5′-CTTTGTCGTTGGTTAGCTGGT-3′;
snail, forward 5′-ACTGCGACAAGGAGTACACC-3′, reverse
5′-GAGTGCGTTTGCAGATGGG-3′; slug, forward
5′-TGACCTGTCTGCAAATGCTC-3′, reverse 5′-TCGGACCCACACATTACCTT-3′; and
GAPDH, forward 5′-GGAGCGAGATCCCTCCAAAAT-3′, reverse
5′-GGCTGTTGTCATACTTCTCATGG-3′. mRNA levels in each group were
calculated by the 2−ΔΔCt method and normalized to the
internal reference gene GAPDH (11). Independent experiments were
repeated three times.
Western blotting
Tissues (~5mg) were homogenized with an electric
homogenizer and lysed with 100 µl ice cold T-PER Tissue
Protein Extraction Reagent (Pierce; Thermo Fisher Scientific,
Inc.). Cells (5×105) were washed twice with PBS and
lysed with 100 µl ice cold radioimmunoprecipitation assay
lysis and extraction buffer (Pierce; Thermo Fisher Scientific,
Inc.). The samples were centrifuged at 10,000 × g for 5 min at 4°C
to pellet tissue/cell debris and the supernatant containing protein
was collected. Protein concentrations were determined using a
Bicinchoninic Acid Protein kit (Pierce; Thermo Fisher scientific,
Inc.). Total protein (20 µg) was separated by 10% SDS-PAGE
and proteins were transferred to polyvinylidene difluoride
membranes. The membranes were blocked with 5% fat-free milk in PBS
with 0.1% Tween-20 (Sinopharm Chemical Reagent Co., Ltd., Shanghai,
China) for 2 h at room temperature and subsequently incubated with
the following primary antibodies (1:2,000) in PBS with 0.1%
Tween-20 at 4°C overnight: Anti-PFN2 (cat. no. 60094-2-Ig;
ProteinTech Group Inc., Chicago, IL, USA), anti-slug (cat. no.
ab51772; Abcam, Cambridge, MA, USA), anti-snail (cat. no. ab82846;
Abcam), anti-E-cadherin (cat. no. ab1416; Abcam), anti-N-cadherin
(cat. no. ab18203; Abcam), anti-vimentin (cat. no. ab20346; Abcam),
anti-GAPDH (cat. no. ab8245; Abcam), anti-phosphorylated (p)-MLC
(cat. no. 3675; Cell Signaling Technology, Inc., Danvers, MA, USA),
anti-MLC (cat. no. 3672, Cell Signaling Technology, Inc.) and
anti-β-tubulin (cat. no. ab6046; Abcam). Following washing with
PBS, the membranes were incubated with horseradish peroxidase
(HRP)-conjugated goat anti-mouse immunoglobulin (Ig)G (cat. no.
32430; Thermo Fisher Scientific Inc.) or HRP-conjugated goat
anti-rabbit IgG (cat. no. 31460; Thermo Fisher Scientific Inc.)
secondary antibodies (1:5,000) in PBS with 0.1% Tween-20 for 1 h at
room temperature. Following subsequent washes with PBS, the
immunoreactive bands were visualized with Enhanced
Chemiluminescence Plus Western Blotting Detection reagents (cat.
no. 2650; Merck KGaA). Densitometric analysis was performed to
quantify protein expression using ImageJ software (version 1.48;
National Institutes of Health, Bethesda, MA, USA); protein
expression levels were normalized to the loading controls GAPDH or
β-tubulin. Independent experiments were repeated three times.
Immunohistochemistry (IHC)
Tissues were fixed with 4% paraformaldehyde in PBS
for 48 h at room temperature and dehydrated through a series of
graded ethanol baths at room temperature (70% for 1 h; 95% for 1 h;
100% for 1 h; 100% for 1.5 h; 100% for 1.5 h; 100% for 2 h),
followed by clearing in xylene for 1 h at room temperature and
infiltrated with paraffin for 1 h at 60°C. The paraffin embedded
tissues were cut into 5 µm sections. The tissue sections
were subjected to routine deparaffinization and rehydration at room
temperature: 100% Xylene (20 min); 100% xylene (20 min); 100%
ethanol (10 min); 95% ethanol (10 min); 90% ethanol (10 min); 80%
ethanol (10 min). Antigen retrieval was achieved by microwaving the
sections in 0.01 mol/l citrate buffer for 10 min and cooling for 30
min to reach the room temperature. Endogenous peroxidase activity
was inhibited by incubating the sections with 3% hydrogen peroxide
in methanol for 20 min, and non-specific binding was blocked by
incubation with 5% bovine serum albumin (Sigma-Aldrich; Merck KGaA)
in PBS at room temperature. Sections were washed three times with
PBS and incubated overnight at 4°C with anti-PFN2 primary antibody
(cat. no. 60094-2-Ig; ProteinTech Group Inc.). Subsequently,
sections were incubated with HRP-conjugated rat anti-mouse-IgG2b
(cat. no. ab157293, Abcam, Inc.), and the signal was developed with
3,3′-diaminobenzidine tetrahydrochloride in Tris-HCl buffer (pH
7.6) containing 0.02% hydrogen peroxide. The sections were
counterstained with hematoxylin and mounted. Negative controls were
obtained by incubating sections with PBS as opposed to the specific
primary antibodies. IHC staining was independently examined by two
clinical pathologists blinded to the study. For each sample, five
high-power fields (magnification, ×100) were randomly selected
under a Nikon Eclipse Ti-S light microscope (Nikon Corporation,
Tokyo, Japan) and analyzed using NIS-Elements version 4.0 (Nikon
Corporation) software. The intensity of cytoplasmic staining was
determined semi-quantitatively on a scale of 0–3 as follows: 0
(negative), 1 (weakly positive), 2 (moderately positive) and 3
(strongly positive). A consensus score was assigned for each
section following discussion and careful review of all slides by
the two pathologists. Tissues with a final staining score of 0 or 1
were grouped into the 'low' expression group, and tissues with a
score of 2 were grouped into the 'moderate' expression group.
Wound-healing assay
Cells (5×105 cells/well) were seeded into
a 12-well plate and grown until they reached 80% confluency, ~24 h.
HCT116 cells were cultured at 37°C in a humidified incubator with
5% CO2 in McCoy's 5A (Modified) medium (Thermo Fisher
Scientific, Inc.). SW620 cells were cultured at 37°C in an
incubator without CO2 in Leibovitz's L 15 Medium (Thermo
Fisher Scientific, Inc.). Both of the media were supplemented with
10% FBS (Thermo Fisher Scientific, Inc.) and 100 U/ml penicillin
and 100 µg/ml streptomycin (Thermo Fisher Scientific, Inc.).
The cell monolayers were manually wounded by scraping with a 200
µl pipette tip. Cells were washed with PBS to remove the
debris and were cultured for 24 h to allow for wound healing.
Within each well, five sites of a unique regular wound were
selected for analysis using a live cell imaging microscope (Zeiss
AG, Oberkochen, Germany); images of the wounds were captured
immediately following wounding (0 h) and at the end of the
experiment (24 h), and the distance between the two edges of the
injury were quantified using Adobe Photoshop CS3 (Adobe Systems,
Inc., San Jose, CA, USA). The migration distances were recorded as
the percentage of cell migration, and the data are presented as the
mean ± sandard error of the mean (SEM) of triplicate assays for
each cell line; at least three independent experiments were
performed.
Transwell migration assay
24-well Transwell insert chambers (Merck KGaA) were
used for cell migration assay. Cells (1×105) were added
to the top chambers in serum-free medium (200 ml; McCoy's 5A for
HCT116 and L 15 for SW620), and the bottom chambers were filled
with medium (McCoy's 5A for HCT116 and L 15 for SW620) containing
10% FBS. Cells were cultured for 24 h at 37°C in a 5%
CO2-humidified incubator (HCT116) or
non-CO2-humidified incubator (SW620). To quantify
migration, cells were removed from the upper membrane by gently
scrapping this side with a wet cotton swab. The migrated cells
attached to the lower membrane were fixed with 4% paraformaldehyde
for 20 min, stained with 0.5% crystal violet solution for 30 min,
and washed two times with PBS; all at room temperature. The number
of migrated cells was subsequently counted in five randomly
selected fields under a light microscope (Zeiss AG), and the
migrated cell numbers were quantified using Adobe Photoshop CS3
(Adobe Systems, Inc.). Independent experiments were repeated three
times.
Statistical analysis
Statistical differences were evaluated using
Statistical Package for Social Science software (version 17.0;
SPSS, Inc., Chicago, IL, USA). The association between staining
intensity and clinicopathological patterns was assessed using the
χ2-test and two-sided Fisher's exact test to determine
the significance of the difference between the covariates. All
measurement data are presented as mean ± SEM. Statistical
significance between two groups was evaluated by the Student's
t-test and unpaired nonparametric Mann-Whitney U test. Comparison
of multiple groups was performed using analysis of variance and
Bonferroni post hoc tests. P<0.05 was considered to indicate a
statistically significant difference.
Results
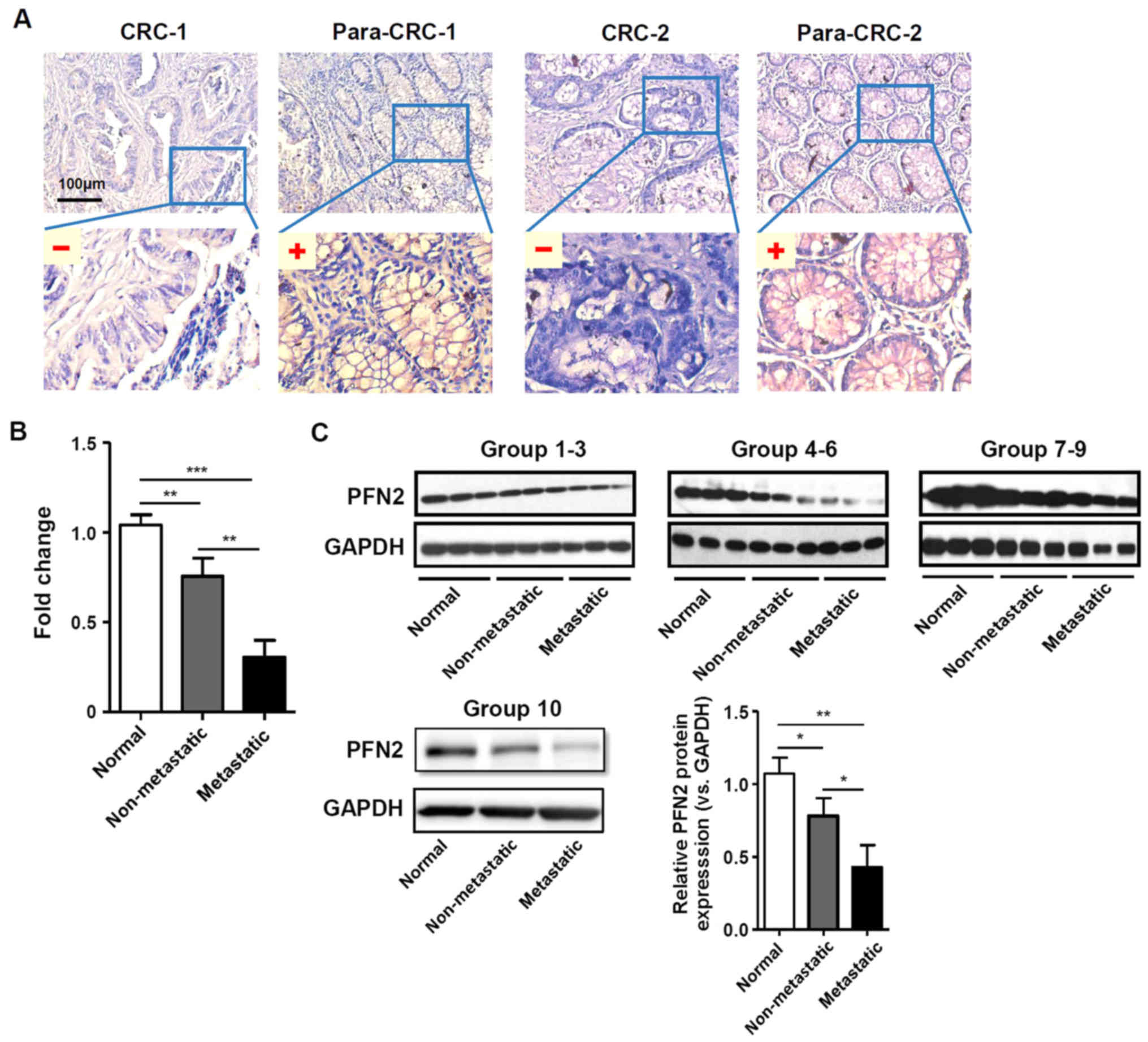
Downregulation of PFN2 expression in CRC
patients with metastasis
PFN2 expression was examined in CRC and normal
paracancerous colon mucosa tissues by IHC staining. The clinical
features of the patients and the results of the IHC staining
statistical analyses are provided in Table I and Fig. 1A, respectively. The results
indicated that PFN2 expression was significantly lower in CRC
tissues compared with expression levels in the normal adjacent
colorectal tissues. Furthermore, the expression level of PFN2 was
significantly lower in CRC patients with lymph node and distant
metastasis compared with CRC patients without metastasis (Table I). In addition, RT-qPCR and western
blot analyses on freshly collected colorectal tissue samples
demonstrated that PFN2 expression was significantly decreased in
metastatic tumors compared with either non-metastatic tumor or
normal colorectal tissues (Fig. 1B and
C, respectively), which suggested that there may be a clinical
association between PFN2 expression and CRC progression.
 | Table IAssociation between PFN2 expression
and clinicopathological features in patients with CRC. |
Table I
Association between PFN2 expression
and clinicopathological features in patients with CRC.
| Clinicopathological
parameters | n | PFN2 expression
| P-value |
|---|
| Low | Medium |
|---|
| Total cases | 70 | 40 | 30 | |
| Age (years) | | | | NS |
| ≤60 | 28 | 16 | 12 | |
| >60 | 42 | 24 | 18 | |
| Tissue type | | | | <0.001 |
| Normal colorectal
tissue | 70 | 0 | 70 | |
| CRC tissue | 70 | 40 | 30 | |
| Sex | | | | NS |
| Male | 38 | 22 | 16 | |
| Female | 32 | 18 | 14 | |
| Tumor size
(cm) | | | | NS |
| ≤5 | 41 | 20 | 21 | |
| >5 | 29 | 20 | 9 | |
| TNM stage | | | | <0.001 |
| I | 8 | 0 | 8 | |
| II | 21 | 6 | 15 | |
| III | 26 | 20 | 6 | |
| IV | 15 | 14 | 1 | |
| Lymph node
metastasis | | | | <0.001 |
| Negative | 29 | 6 | 23 | |
| Positive | 41 | 34 | 7 | |
| Distance
metastasis | | | | <0.001 |
| Negative | 55 | 26 | 29 | |
| Positive | 15 | 14 | 1 | |
Low PFN2 expression affects CRC cell
migration
Given the aforementioned differential expression of
PFN2, the function of PFN2 in human CRC development was
subsequently investigated. CRC metastasis was previously reported
to be closely associated with the EMT (12,13).
Therefore, the present study examined the mRNA expression levels of
EMT-related molecules, including E-cadherin, N-cadherin, vimentin,
slug and snail in normal colon, non-metastatic and metastatic CRC
tissues. The results demonstrated that the mRNA expression levels
of vimentin, slug and snail were significantly higher in metastatic
CRC tissues compared with normal colon tissues (Fig. 2A). In addition, PFN2 expression was
also determined in the non-metastatic CRC cell line HCT116 and in
the SW620 cell line, which was originally derived from a metastatic
site of a patient with CRC. PFN2 mRNA was expressed at
significantly lower levels in the SW620 cell line compared with
expression levels in HCT116 cells. The mRNA expression level of
E-cadherin was also lower in SW620 cells, whereas the expression
levels of N-cadherin, vimentin, snail and slug were significantly
higher in SW620 cells compared with the respective expression
levels in HCT116 cells (Fig. 2B).
Western blotting results revealed that the epithelial marker
E-cadherin and PFN2 protein expression levels were notably
decreased, whereas expression levels of the mesenchymal marker
proteins vimentin, N-cadherin and related molecules, such as snail
and slug, were increased in SW620 cells compare with the respective
expression levels in HCT116 cells (Fig. 2C). These results suggested that
PFN2 may have negative effects on the EMT process. To confirm the
relationship between PFN2 and cancer metastasis, wound-healing and
Transwell migration assays were performed in HCT116 and SW620
cells. The migratory ability of SW620 cells was significantly
higher compared with that of HCT116 cells (Fig. 2D and E). These data suggested that
the reduced level of PFN2 expression in SW620 CRC cells may be
associated with higher migratory capability.
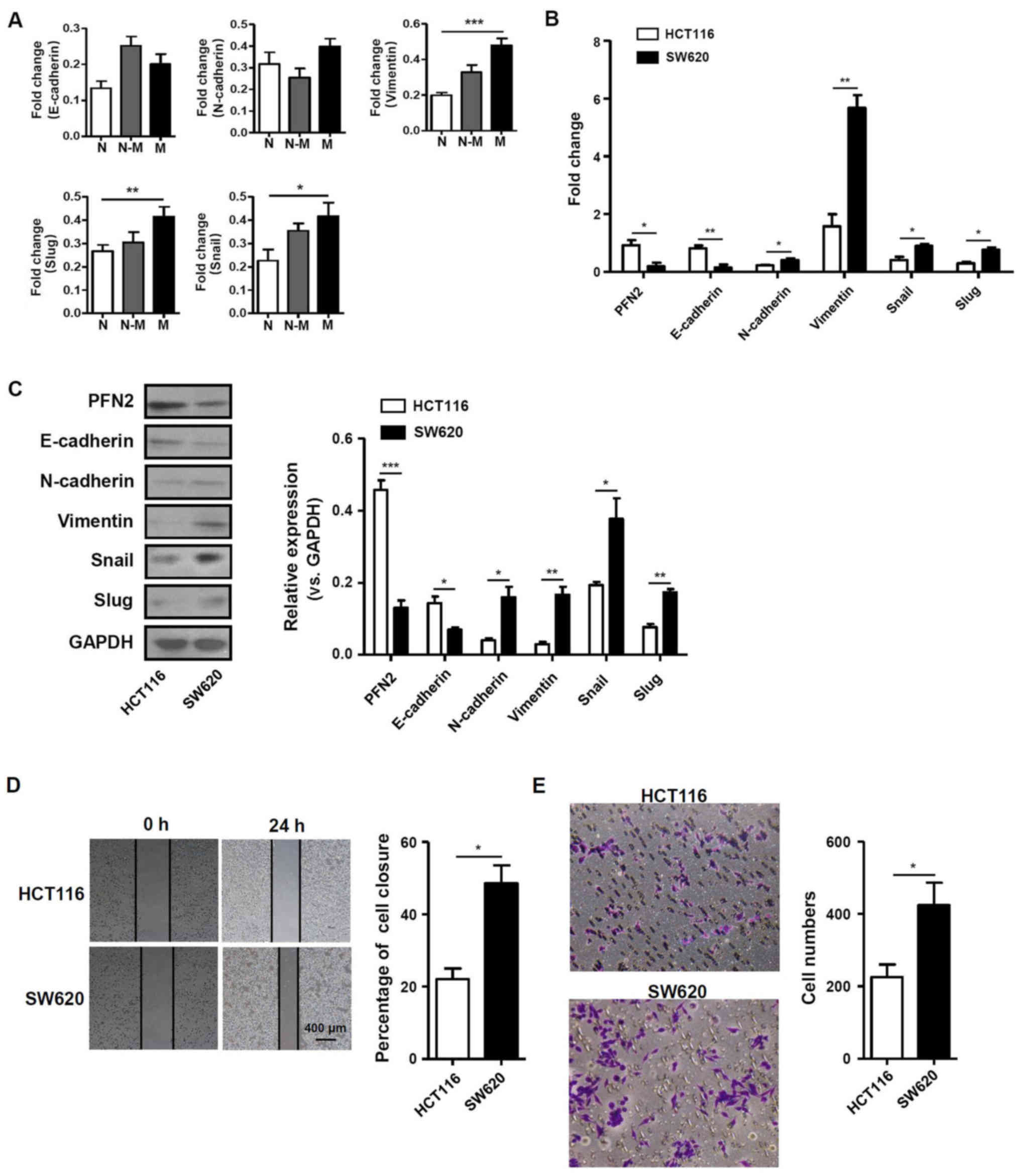 | Figure 2PFN2 expression level is associated
with EMT-related molecules and migratory activity in CRC cells. (A)
The expression of EMT-related molecules in human normal colorectal,
non-metastatic CRC and metastatic CRC tissues. (B and C) mRNA and
protein expression levels of PFN2 and EMT related molecules,
E-cadherin, N-cadherin, vimentin, slug and snail, were measured in
two CRC cell lines, HCT116 and SW620, by (B) reverse
transcription-quantitative polymerase chain reaction and (C)
western blotting, respectively; GAPDH served as the internal
reference. (D and E) The migratory activity of CRC cell lines was
analyzed by (D) wound-healing and (E) Transwell migration assays at
24 h. Data are presented as the mean ± sandard error of the mean of
three independent experiments. *P<0.05,
**P<0.01 and ***P<0.001. CRC,
colorectal cancer; EMT, epithelial-mesenchymal transition; N,
normal; N-M, non-metastatic; M, metastatic; PFN2, profilin 2. |
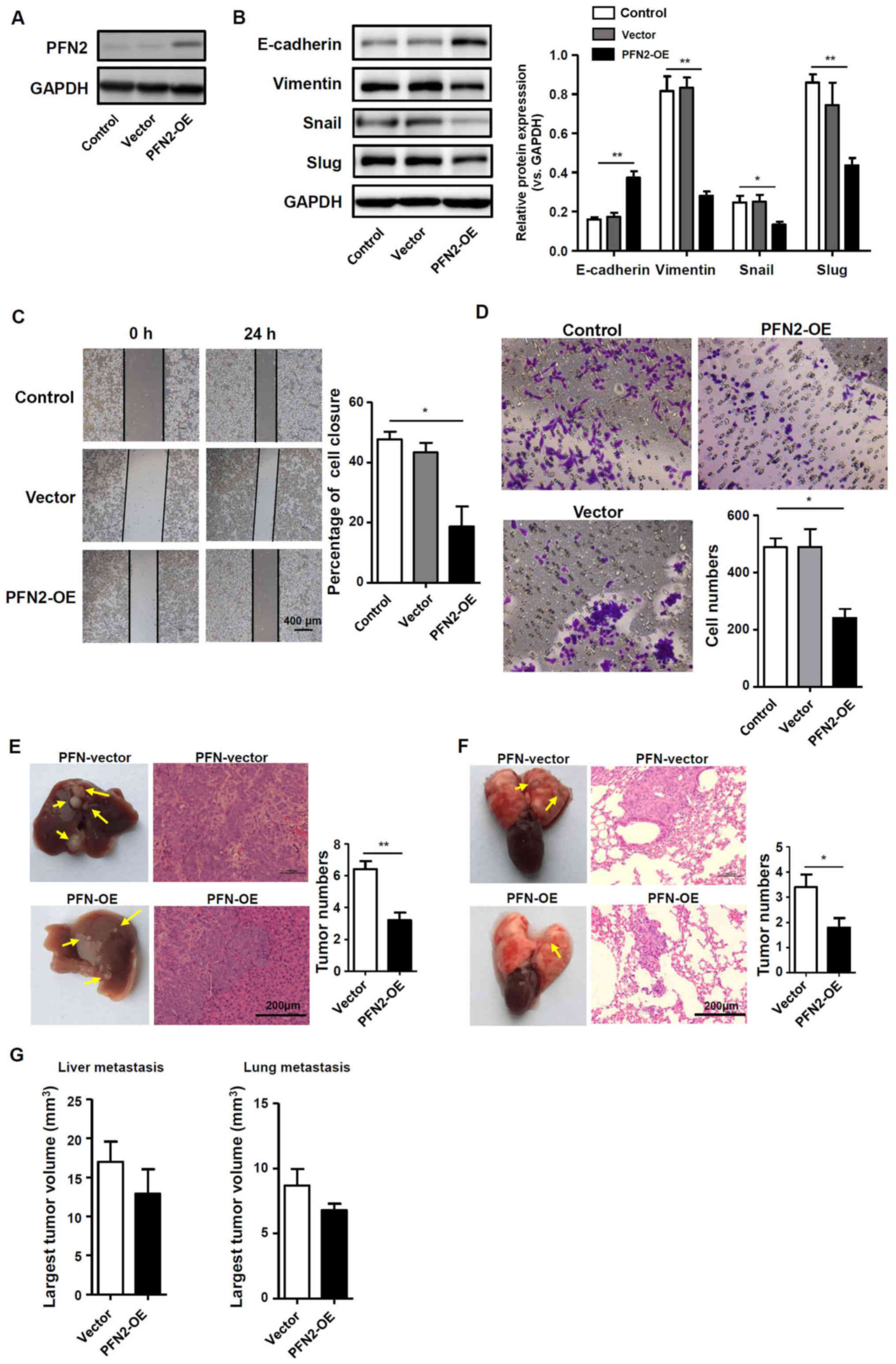
PFN2 suppresses CRC cell migration with
reduced EMT
To further determine if PFN2 serves a role in EMT
and CRC cell migration, the pQCXIH vector was used to transiently
over-express PFN2 in SW620 cells, which express a low level of
endogenous PFN2, as confirmed by western blotting (Fig. 3A); untransfected and empty
vector-transfected SW620 cells were used as the controls. The
potential role of PFN2 as a negative regulator of EMT was further
explored by examining the expression of EMT markers and known EMT
regulators, slug and snail. Western blotting demonstrated that
E-cadherin protein expression was increased, whereas vimentin, slug
and snail expression levels were decreased in SW620 cells
overexpressing PFN2, which suggested that the inhibition of EMT may
be regulated by PFN2 overexpression in SW620 cells (Fig. 3B). Correspondingly, SW620 cell
migration was significantly reduced by PFN2 overexpression compared
with control cells (Fig. 3C and
D). In vivo liver and lung metastasis mouse models were
used to evaluate the potential roles of PFN2 in regulating CRC
metastasis. The results revealed that PFN2-overexpressing SW620
cells exhibited reduced metastatic potential compared with
non-PFN2-overexpressing SW620 cells in liver and lung (Fig. 3E and F, respectively); however, no
significant difference in the largest tumor nodule volume was
identified between these two groups (Fig. 3G). These results suggested a
negative role of PFN2 in CRC cell migration.
PFN2 inhibits CRC EMT by regulating
cytoskeletal reorganization
PFN2 triggers various cellular pathways to exert
disparate functions. As an actin-binding protein, one of these
functions is to regulate cytoskeletal reorganization (14). Notably, contractile actin bundles
are thought to be suppressors of cancer protrusive activity,
migration and invasion (15). As
MLC phosphorylation is a marker of myosin motor contractions
(16), the present study examined
the level of pMLC in CRC tissues and cell lines. In normal colon
tissues, pMLC was expressed at notably higher levels and pMLC
expression was significantly lower in metastatic CRC tissues
compared with non-metastatic CRC tissues (Fig. 4A). In addition, in PFN2-OE SW620
cells, pMLC expression was significantly increased compared with
the untransfected and vector-trans-fected control groups (Fig. 4B), which indicated that PFN2
expression in CRC cells may regulate the contractile actin bundles.
To determine the relationship between the generation of contractile
actin bundles and PFN2-regulated cancer metastasis, the present
study assessed the effects of altering myosin activity on the
suppressive effects of PFN2. Therefore, the pharmacological
inhibitor of MLC phosphorylation, Y27632, was used in subsequent
experiments. Inhibition of MLC phosphorylation attenuated the
inhibitory effects of PFN2-OE on EMT, as the expression of
E-cadherin decreased whereas the expression of vimentin increased
in PFN2-OE SW620 cells treated with Y27632 compared with those
cells without Y27632 treatment for 24 h (Fig. 4C). The results of the wound-healing
assay demonstrated that with the migratory ability of PFN2-OE SW620
cells treated with Y27632 significantly increased compared with
those cells without Y27632 (Fig.
4D). Therefore, it was hypothesized that the suppressive role
of PFN2 on CRC metastasis may have resulted from PFN2-regulated
cytoskeletal reorganization.
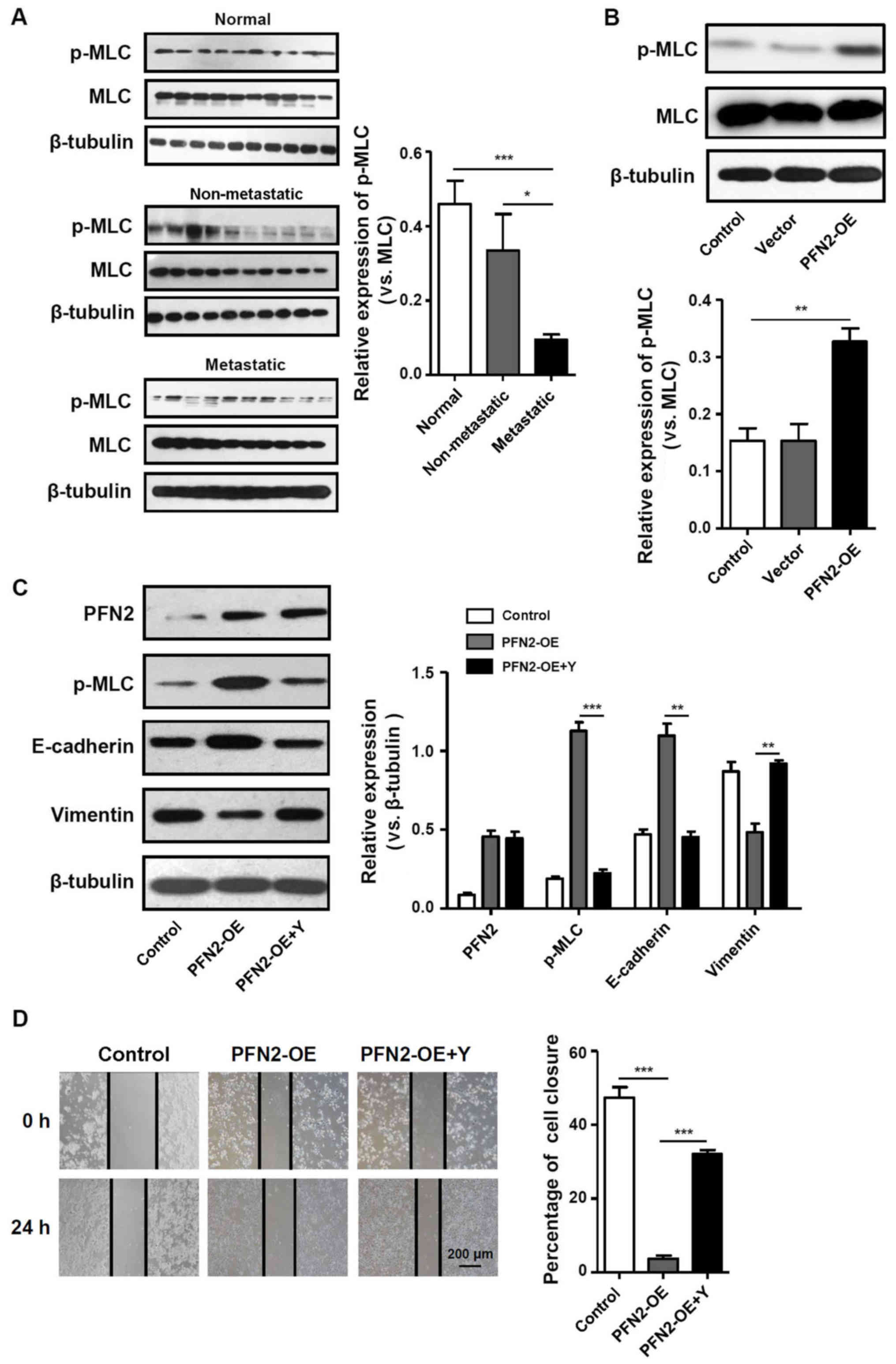 | Figure 4Cytoskeletal remodeling may be
involved in PFN2-regulated EMT. (A and B) p-MLC expression was
measured by western blotting in (A) normal colon, non-metastatic
CRC and metastatic CRC tissues, and (B) in untransfected, vector-
and PFN2-OE-transfected SW620 cells. (C) Protein expression levels
of PFN2, p-MLC, E-cadherin and vimentin were examined by western
blotting in PFN2-OE-transfected SW620 cells with or without
co-treatment with Y27632; β-tubulin served as the internal
reference. (D) Wound-healing assays were performed to determine the
migratory ability of PFN2-OE-transfected SW620 with or without
co-treatment with Y27632. Data are presented as the mean ± sandard
error of the mean of three independent experiments.
*P<0.05, **P<0.01 and
***P<0.001. CRC, colorectal cancer; EMT,
epithelial-mesenchymal transition; MLC, myosin light chain; OE,
overexpression; p, phosphorylated; PFN2, profilin 2; Y,
Rho-associated kinase inhibitor Y27632. |
Discussion
The actin cytoskeleton is a highly dynamic
structure, which undergoes constant remodeling in a living cell
(5). In general, the regulation of
such a structure is a prerequisite for such processes as
endocytosis, cell motility and cancer cell invasion. As reported
previously, the actin cytoskeleton pathway serves important roles
in the development of metastatic CRC (17). Therefore, molecules involved in
this pathway may be candidate regulators of the migratory potential
of CRC. The aim of the present study was to investigate whether the
actin cytoskeleton regulator PFN2 serves specific roles in the
progression of human CRC, particularly in the process of
metastasis. PFN2 expression was demonstrated to be significantly
lower in metastatic CRC tissues, which may function by negatively
affecting EMT through the regulation of the actin cytoskeleton.
Although the roles and underlying mechanisms of PFN2
in cytoskeletal regulation has been demonstrated in a number of
previous studies (15,18), its relationship with cancer has not
been as extensively studied until recently. For example in head and
neck squamous cell carcinoma, high PFN2 expression independently
predicted poor overall survival; microRNA-30a-5p suppresses EMT by
targeting PFN2 in high invasive non-small cell lung cancer cell
lines; however, further mechanistic studies are still expected
(19–21). However, previous studies have
focused on another member of the profilin family in cancer
development, PFN1 (22). As a
ubiquitously expressed actin polymerization regulator, PFN1 has
been studied in different types of cancer, including gastric,
bladder, renal and breast cancer (23–26).
Considering its inhibitory roles in the progression of carcinomas,
it is considered to be a tumor suppressor and may be a novel
biomarker for predicting the response to anticancer therapy.
However, there may be some limitations in its application owing to
its non-specific expression in various tissues: PFN1 is expressed
during all embryonic stages, and a previous study reported that it
is expressed in nearly all cell and tissue types; by contrast, PFN2
is expressed primarily in the developing nervous system and in
adult differentiated neurons (27). Therefore, the neuronal
tissue-specific isoform PFN2 may be better candidate for anticancer
therapy with fewer side effects on other tissues.
To date, PFN2 has been linked with oral squamous
cell carcinoma, breast cancer and inflammatory bowel
disease-associated CRC (9,15,17).
However, it has not been investigated in depth for its
cancer-related functions. To demonstrate the role of PFN2 in CRC,
the present study analyzed the expression levels of PFN2 in
patients with CRC and in CRC cell lines. The results demonstrated
that PFN2 expression was higher in non-metastatic CRC tissues and
cell lines, whereas expression in metastatic CRC tissues and cells
was comparatively low. These data suggested a suppressive role for
PFN2 in cancer metastasis, which was consistent with a previous
report (15). Although cancer cell
growth has also been suggested to be regulated by PFN2 (28), the present study primarily focused
on cell migration, as significant differences in expression were
observed between CRC with and without metastasis.
EMT is a key process during cancer metastasis,
characterized by a loss of cell-cell adhesion and an increase in
cell motility (29). In the
present study, the expression levels of several EMT markers were
significantly altered by the overexpression of PFN2 in CRC cell
lines. In CRC cells expressing low levels of PFN2, higher
expression of the mesenchymal markers, including vimentin, snail
and slug, was observed, whereas overexpressing PFN2 suppressed the
EMT process and, thus, the migration of CRC cells. EMT is regulated
by complex signaling pathways, such as transforming growth
factor-β, phosphoinositide-3 kinase, mitogen-activated protein
kinase, integrin-linked kinase and jagged1 (30). As none of these signaling pathways
have been reported in relation to PFN2 functions, the present study
detected the expression of the EMT-induced transcription factors,
snail and slug (31). Although the
signaling of EMT in CRC has been studied previously, whether
PFN2-regulated EMT shares similar mechanisms requires further
investigation.
As aforementioned, PFN2 is a cytoskeleton regulator.
The actin cytoskeleton serves a central role in many carcinogenesis
and cancer cell migration processes (32–34).
Therefore, it is reasonable to suggest that the actin cytoskeleton
may be associated with PFN2-regulated EMT and subsequent cancer
migration. MLC phosphorylation is known to activate actomyosin
contractility, which is crucial for cytoskeleton remodeling
(16). A previous study
demonstrated that PFN2 knockdown in breast cancer cells reduced the
expression of total p-MLC (15),
which was accompanied by reduced cortical actin bundling. The
present study overexpressed PFN2 in the CRC cell line SW620 and
observed similar effects of PFN2 on MLC phosphorylation. Altered
expression levels of the EMT markers indicated that PFN2 may reduce
the metastatic ability of CRC by inducing MLC phosphorylation.
Furthermore, inhibiting the phosphorylation of MLC using a
pharmacological inhibitor reversed the effect of PFN2 on EMT.
In conclusion, results from the present study
revealed that PFN2 expression was reduced in metastatic CRC. PFN2
was observed to exert a suppressive effect on EMT, and subsequently
on the migratory abilities of CRC cells, and this may be through
regulating cytoskeletal reorganization. Therefore, decreased PFN2
expression may have important clinicopathological implications for
CRC metastasis and may be a potential target for future CRC
therapies.
Abbreviations:
|
CRC
|
colorectal cancer
|
|
EMT
|
epithelial-mesenchymal transition
|
|
IHC
|
immunohistochemistry
|
|
MLC
|
myosin light chain
|
|
PFN2
|
profilin 2
|
Acknowledgments
Not applicable.
References
|
1
|
Jemal A, Siegel R, Xu J and Ward E: Cancer
statistics, 2010. CA Cancer J Clin. 60:277–300. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Iseki Y, Shibutani M, Maeda K, Nagahara H,
Ohtani H, Sugano K, Ikeya T, Muguruma K, Tanaka H, Toyokawa T, et
al: Impact of the preoperative controlling nutritional status
(CONUT) score on the survival after curative surgery for colorectal
cancer. PLoS One. 10:e01324882015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Liu H, Ren G, Wang T, Chen Y, Gong C, Bai
Y, Wang B, Qi H, Shen J, Zhu L, et al: Aberrantly expressed Fra-1
by IL-6/STAT3 transactivation promotes colorectal cancer
aggressiveness through epithelial-mesenchymal transition.
Carcinogenesis. 36:459–468. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Grünert S, Jechlinger M and Beug H:
Diverse cellular and molecular mechanisms contribute to epithelial
plasticity and metastasis. Nat Rev Mol Cell Biol. 4:657–665. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yilmaz M and Christofori G: EMT, the
cytoskeleton, and cancer cell invasion. Cancer Metastasis Rev.
28:15–33. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
König K, Meder L, Kröger C, Diehl L,
Florin A, Rommerscheidt-Fuss U, Kahl P, Wardelmann E, Magin TM,
Buettner R, et al: Loss of the keratin cytoskeleton is not
sufficient to induce epithelial mesenchymal transition in a novel
KRAS driven sporadic lung cancer mouse model. PLoS One.
8:e579962013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jockusch BM, Murk K and Rothkegel M: The
profile of profilins. Rev Physiol Biochem Pharmacol. 159:131–149.
2007.PubMed/NCBI
|
|
8
|
Witke W, Podtelejnikov AV, Di Nardo A,
Sutherland JD, Gurniak CB, Dotti C and Mann M: In mouse brain
profilin I and profilin II associate with regulators of the
endocytic pathway and actin assembly; EMBO J. 17:967–976. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ma CY, Zhang CP, Zhong LP, Pan HY, Chen
WT, Wang LZ, Andrew OW, Ji T and Han W: Decreased expression of
profilin 2 in oral squamous cell carcinoma and its
clinicopathological implications. Oncol Rep. 26:813–823.
2011.PubMed/NCBI
|
|
10
|
Greene FL: Current TNM staging of
colorectal cancer. Lancet Oncol. 8:572–573. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
12
|
Castosa R, Martinez-Iglesias O, Roca-Lema
D, Casas-Pais A, Díaz-Díaz A, Iglesias P, Santamarina I, Graña B,
Calvo L, Valladares-Ayerbes M, et al: Hakai overexpression
effectively induces tumour progression and metastasis in vivo. Sci
Rep. 8:3466–3475. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Findlay VJ, Wang C, Watson DK and Camp ER:
Epithelial-to-mesenchymal transition and the cancer stem cell
phenotype: Insights from cancer biology with therapeutic
implications for colorectal cancer. Cancer Gene Ther. 21:181–187.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ferron F, Rebowski G, Lee SH and Dominguez
R: Structural basis for the recruitment of profilin-actin complexes
during filament elongation by Ena/VASP. EMBO J. 26:4597–4606. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mouneimne G, Hansen SD, Selfors LM, Petrak
L, Hickey MM, Gallegos LL, Simpson KJ, Lim J, Gertler FB, Hartwig
JH, et al: Differential remodeling of actin cytoskeleton
architecture by profilin isoforms leads to distinct effects on cell
migration and invasion. Cancer Cell. 22:615–630. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Takebe T, Enomura M, Yoshizawa E, Kimura
M, Koike H, Ueno Y, Matsuzaki T, Yamazaki T, Toyohara T, Osafune K,
et al: Vascularized and complex organ buds from diverse tissues via
mesenchymal cell-driven condensation. Cell Stem Cell. 16:556–565.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kanaan Z, Qadan M, Eichenberger MR and
Galandiuk S: The actin-cytoskeleton pathway and its potential role
in inflammatory bowel disease-associated human colorectal cancer.
Genet Test Mol Biomarkers. 14:347–353. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Schlüter K, Jockusch BM and Rothkegel M:
Profilins as regulators of actin dynamics. Biochim Biophys Acta.
1359:97–109. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu J, Wu Y, Wang Q, Liu X, Liao X and Pan
J: Bioinformatic analysis of PFN2 dysregulation and its prognostic
value in head and neck squamous carcinoma. Future Oncol.
14:449–459. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cui XB, Zhang SM, Xu YX, Dang HW, Liu CX,
Wang LH, Yang L, Hu JM, Liang WH, Jiang JF, et al: PFN2, a novel
marker of unfavorable prognosis, is a potential therapeutic target
involved in esophageal squamous cell carcinoma. J Transl Med.
14:137–152. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yan J, Ma C and Gao Y: MicroRNA-30a-5p
suppresses epithelial-mesenchymal transition by targeting
profilin-2 in high invasive non-small cell lung cancer cell lines.
Oncol Rep. 37:3146–3154. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ding Z and Roy P: Profilin-1 versus
profilin-2: Two faces of the same coin. Breast Cancer Res.
15:311–312. 2013. View
Article : Google Scholar
|
|
23
|
Cheng YJ, Zhu ZX, Zhou JS, Hu ZQ, Zhang
JP, Cai QP and Wang LH: Silencing profilin-1 inhibits gastric
cancer progression via integrin β1/focal adhesion kinase pathway
modulation. World J Gastroenterol. 21:2323–2335. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Karamchandani JR, Gabril MY, Ibrahim R,
Scorilas A, Filter E, Finelli A, Lee JY, Ordon M, Pasic M,
Romaschin AD, et al: Profilin-1 expression is associated with high
grade and stage and decreased disease-free survival in renal cell
carcinoma. Hum Pathol. 46:673–680. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Coumans JV, Gau D, Poljak A, Wasinger V,
Roy P and Moens PD: Profilin-1 overexpression in MDA-MB-231 breast
cancer cells is associated with alterations in proteomics
biomarkers of cell proliferation, survival, and motility as
revealed by global proteomics analyses. OMICS. 18:778–791. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liang JW, Shi ZZ, Shen TY, Che X, Wang Z,
Shi SS, Xu X, Cai Y, Zhao P, Wang CF, et al: Identification of
genomic alterations in pancreatic cancer using array-based
comparative genomic hybridization. PLoS One. 9:e1146162014.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Alkam D, Feldman EZ, Singh A and Kiaei M:
Profilin1 biology and its mutation, actin(g) in disease. Cell Mol
Life Sci. 74:967–981. 2017. View Article : Google Scholar :
|
|
28
|
Tang YN, Ding WQ, Guo XJ, Yuan XW, Wang DM
and Song JG: Epigenetic regulation of Smad2 and Smad3 by profilin-2
promotes lung cancer growth and metastasis. Nat Commun.
6:8230–8239. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tsai JH and Yang J: Epithelial-mesenchymal
plasticity in carcinoma metastasis. Genes Dev. 27:2192–2206. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Savagner P: Epithelial-mesenchymal
transitions: From cell plasticity to concept elasticity. Curr Top
Dev Biol. 112:273–300. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Medici D, Hay ED and Olsen BR: Snail and
Slug promote epithelial-mesenchymal transition through
beta-catenin-T-cell factor-4-dependent expression of transforming
growth factor-beta3. Mol Biol Cell. 19:4875–4887. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Qiao Y, Chen J, Lim YB, Finch-Edmondson
ML, Seshachalam VP, Qin L, Jiang T, Low BC, Singh H, Lim CT, et al:
YAP regulates actin dynamics through ARHGAP29 and promotes
metastasis. Cell Reports. 19:1495–1502. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wieczorek K, Wiktorska M, Sacewicz-Hofman
I, Boncela J, Lewiński A, Kowalska MA and Niewiarowska J: Filamin A
upregulation correlates with Snail-induced epithelial to
mesenchymal transition (EMT) and cell adhesion but its inhibition
increases the migration of colon adenocarcinoma HT29 cells. Exp
Cell Res. 359:163–170. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Izdebska M, Gagat M and Grzanka A:
Overexpression of lamin B1 induces mitotic catastrophe in colon
cancer LoVo cells and is associated with worse clinical outcomes.
Int J Oncol. 52:89–102. 2018.
|