Introduction
Cancer represents one of the leading causes of
morbidity and mortality globally, with ~14 million new cases and
8.2 million cancer-associated mortalities in 2012. It has been
predicted that the number of cancer cases will increase from the
recorded 14 million in 2012 to 22 million per year within the next
2 decades (1). Colorectal cancer
(CRC) is the third most frequent cancer diagnosis in men and the
second most frequent in women worldwide; however, the incidence of
CRC varies significantly among different countries (2). While surgical resection is a
successful treatment modality in the early stages of this
malignancy, the treatment options for the advanced stages of
colorectal cancer remain limited in terms of their efficiency due
to the generalized spread of tumor cells and their acquired
chemoresistance.
Novel strategies to improve cancer treatment have
been intensively investigated, including the search for substances
with anticancer effects. Among the natural extracts with possible
anticancer activity, inositol hexaphosphate (IP6), also known as
phytic acid, represents a promising option. IP6 has been studied
in vivo and in vitro in different types of cancer
tissues and cells, including those of the colon (3), breast (4), liver (5), prostate (6), skin (7) and bladder (8). In these studies, IP6 was reported to
arrest cellular proliferation in the G0/G1 phase (9), inhibit iron-mediated oxidative
reactions (10), enhance
differentiation and stimulate apoptosis (10), and regulate the process of cellular
differentiation (11–13). Furthermore, published studies have
suggested that IP6 may also affect the invasion and migration of
cancer cells in vitro and in vivo. In one previous
study, IP6 prevented the colorectal cancer development and
metastatic progression to the liver in BALB/c mice by altering the
expression of several extracellular matrix proteins, the
proteolytic enzyme matrix metalloproteinase (MMP)-9, and other
angiogenic and growth factors (14). The expression of the matrix
metalloproteinases and tissue inhibitors of MMPs (TIMPs) was also
studied in an unstimulated and IL-1β-stimulated colon cancer Caco-2
cell line exposed to IP6. The results showed that IP6 exerts its
inhibitory activity through modulation of MMP and TIMP gene
expression to prevent the migration and invasion of cancer cells
(15). In the breast cancer
MDA-MB231 cell line, treatment with IP6 resulted in a 65% reduction
of cell adhesion to fibronectin and a 37% reduction to collagen,
while also decreasing the number of migrating cells. Additionally,
IP6 significantly inhibited MMP-9 secretion (16) and modulated integrin dimerization,
cell surface expression and the integrin-associated signaling
pathway (17). The present study
was designed to evaluate the effect of IP6 on cell migration and on
the expression of adhesion and invasion markers in the colorectal
cancer SW620 cell line.
Materials and methods
Cell culture and treatment
The continuous SW620 cell line (catalog no. CCL-227;
American Type Culture Collection, Manassas, VA, USA) was routinely
maintained in a humidified 5% CO2 atmosphere at 37°C in
Dulbecco's modified Eagle's medium (DMEM) (catalog no. D5796;
Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) with 10% fetal
bovine serum (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) and penicillin/streptomycin (100 U/ml; Gibco; Thermo Fisher
Scientific, Inc.). Stock solutions of IP6 (catalog no. P5681;
Sigma-Aldrich; Merck KGaA) were prepared in PBS without calcium and
magnesium ions and stored in a refrigerator until use. Since IP6
shows poor solubility in the standard DMEM with 10% fetal bovine
serum, Nutrient Mixture F-12 Ham (catalog no. N-4888) with 1% FBS
and 1% L-glutamine (catalog no. G7513) (both from Sigma-Aldrich;
Merck KGaA) was used in all experiments. Experiments were repeated
at least three times. Cultures were regularly checked for the
presence of mycoplasmas.
Cell migration assay
A cell migration assay was performed using the
xCELLigence Real-Time Cell Analyzer Dual Purpose (RTCA-DP) system
(Roche Diagnostics, Basel, Switzerland). The upper chamber of
CIM-plate 16 were filled with 1% bovine serum albumin-supplemented
Ham's F-12 medium and the lower chamber was filled with Ham's F-12
medium supplemented with 10% FBS. The plate was placed in an
incubator for 1 h (37°C, 5% CO2) for equilibration of a
background. Defined amounts of cells (30,000 cells/100 µl)
with the test substance (0.2 or 1 mM IP6) were added into the upper
chamber, the plates were inserted into the instrument and the cells
were allowed to migrate to the lower chambers for 24 h. The
impedance (corresponding to cell migration) was measured every 10
min.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Cells were cultivated in the 6-well plates at a
density of 150 000 cells/ml (in 2 ml total volume) for 24 h (37°C,
5% CO2). Next, the cells were treated for 12, 24 and 48
h with 0.2 and 1 mM concentrations of IP6. RNA was isolated using
TriReagent (Sigma-Aldrich; Merck KGaA) according to the
manufacturer's recommendations. The purity of the extracted RNA was
measured using a NanoDrop ND-2000 UV-VIS spectrophotometer (Thermo
Fisher Scientific, Inc.).
cDNA was synthetized from 0.5 µg extracted
RNA using random hexamers primer, dNTP mix, RiboLock RNase
inhibitor and M-MULV reverse transcriptase according to the kit
protocol (RevertAid First Strand cDNA Synthesis kit; catalog no.
K1622; Thermo Fisher Scientific, Inc.). cDNA synthesis started with
initial denaturation of 0.5 µg RNA and random hexamers
primer (70°C for 5 min) and continued with other reagents (whole
volume 20 µl) for 5 min at 25°C, for 60 min at 37°C and then
for 5 min at 70°C. Obtained cDNA was stored at −20°C until the qPCR
assay.
RT-qPCR of target genes [β-catenin, E-cadherin,
N-cadherin, epithelial cell adhesion molecule (EpCam),
inter-cellular adhesion molecule-1 (ICAM-1), MMP-2, MMP-9] and a
reference housekeeping gene (GAPDH) was performed in a LightCycler
1.5 Real-Time PCR Detection system (Roche Diagnostics). cDNA was
diluted five times. The assay was performed using Master mix (whole
volume 40 µl; Roche Diagnostics) containing SYBR-Green (25
µl), forward primer (1 µl, 10 mM), reverse primer (1
µl, 10 mM) and diethyl pyrocarbonate water (13 µl).
The PCR was initialized with a denaturation step of 10 min at 95°C,
followed by 40 cycles of amplification as follows: Denaturation for
10 sec at 95°C, annealing for 15 sec at 60°C and extension for 20
sec at 72°C. A dissociation protocol with a gradient (0.5°C every 2
sec) from 65 to 95°C was used to investigate the specificity of the
RT-qPCR and the presence of primer dimers. The amounts of mRNA of
target genes were normalized to the GAPDH reference gene.
Calculations were based on the 2−ΔΔCq method (18). The data are expressed as the fold
increase of the untreated cells (=1). Statistical analysis was
performed using a one-way analysis of variance (ANOVA) with
Bonferroni's modification.
Western blot analysis
Cells were cultivated in 6-well plates at a density
of 150,000 cells/ml (in 2 ml total volume) for 24 h (37°C, 5%
CO2). Next, the cells were treated with 0.2 or 1 mM
concentrations of IP6 for 12, 24 and 48 h. The cells were washed
with PBS and harvested in ice-cold lysis buffer (50 mM Tris-HCl,
150 mM NaCl, 10% glycerol, 1% Triton X-100, 2 mM EDTA, 2 mM EGTA,
β-glycerophosphate, 50 mM NaF, 10 mM sodium pyrophosphate, 200
µM sodiumorthovanadate and 2 mM DTT), and the amount of
protein in the lysates was measured by bicinchoninic acid assay.
Protein lysates were diluted with SDS to a final concentration of 1
µg/µl. Lysates (30 µl) were loaded onto the
10% SDS-polyacrylamide gel (0.1% SDS) and transferred to a
polyvinylidene difluoride membrane (100 V, 90 min), and then
blocked at 25°C for 1.5 h with a solution containing 5% skimmed dry
milk, 10 mM Tris-HCl (pH 8.0), 150 mM sodium chloride and 0.1%
Tween-20 (TBST). Membranes were then incubated at 4°C overnight
with the following primary antibodies: Polyclonal rabbit
anti-cluster of differentiation 54/ICAM-1 (catalog no. 4915;
1:5,000 dilution) polyclonal rabbit anti-β-catenin (catalog no.
8480; 1:5,000 dilution), polyclonal rabbit anti-E-cadherin (catalog
no. 1395; 1:1,500 dilution), monoclonal mouse anti-EpCAM (catalog
no. 2929; 1:2,500 dilution) (all Cell Signaling Technology, Inc.,
Danvers, MA, USA) and monoclonal mouse anti-β-actin (catalog no.
A5441; 1:10,000 dilution; Sigma-Aldrich; Merck KGaA). This was
followed by washing in TBST (6 times for 5 min each). The membranes
were then incubated with peroxidase-conjugated secondary antibodies
[1:20,000 dilution; polyclonal swine anti-rabbit
immunoglobulin/horseradish peroxidase (HRP)-conjugated; catalog no.
P039901-2; polyclonal goat anti-mouse
immunoglobulin/HRP-conjugated; catalog no. P044701-2; Agilent
Technologies, Inc., Santa Clara, CA, USA] for 2 h at 25°C, followed
by washing with TBST. Finally, the signal was developed with an
Enhanced Chemiluminescence Prime Western Blotting Detection Reagent
(Amersham; GE Healthcare Life Sciences, Shanghai, China). The
quantity of chemiluminescence was detected using Imaging System Gel
Logic 2200 PRO (Molecular Imaging, Inc., Ann Arbor, MI, USA).
Immunofluorescence microscopy
Changes in the expression of E-cadherin in the cells
were detected by indirect immunofluorescence. IP6 was tested at
concentrations of 0.2 and 1 mM at three time intervals (24, 48 and
72 h). Treated and control cells grown in cytospin chambers were
rinsed with PBS and fixed in 2% paraformaldehyde (20 min at room
temperature). Following rinsing in PBS, samples were incubated with
skimmed milk (5% solution in PBS; 30 min, room temperature) and
primary mouse anti-E-cadherin antibody (catalog no. 14472; 1:100
dilution; Cell Signaling Technology, Inc.) was added for 2 h at 4°C
in the dark. Cells were then rinsed three times in PBS and Alexa
Fluor 488-labeled goat anti-mouse immunoglobulin G secondary
antibody (catalog no. 150113; 1:250 dilution; Abcam, Cambridge, UK)
was added for 1 h at room temperature. At the end of incubation,
cells were post-labeled with 4′,6-diamidino-2-phenylindole for 10
min at 4°C, and then washed in PBS and mounted into Prolong Gold
anti-fade medium (catalog no. 8961; Cell Signaling Technology,
Inc.). Slides were examined using an epifluorescence microscope
(Nikon Eclipse E400; Nikon Corporation, Tokyo, Japan) and the
expression of E-cadherin was evaluated using image analysis
software (LUCIA DI Image Analysis System LIM; Laboratory Imaging
Ltd., Prague, Czech Republic) in at least 2,000 cells per
sample.
Statistical analysis
Statistical analysis was performed using one-way
ANOVA with Bonferroni's modification using GraphPad Prism 7
(GraphPad Software, Inc., La Jolla, CA, USA). Results were compared
with control samples (0 mM IP6) and presented as the mean ±
standard deviation. P<0.05 was used to indicate a statistically
significant difference. All experiments were completed in at least
two independent replicates (n≥2).
Results
Effect of IP6 on migration of SW620
cells
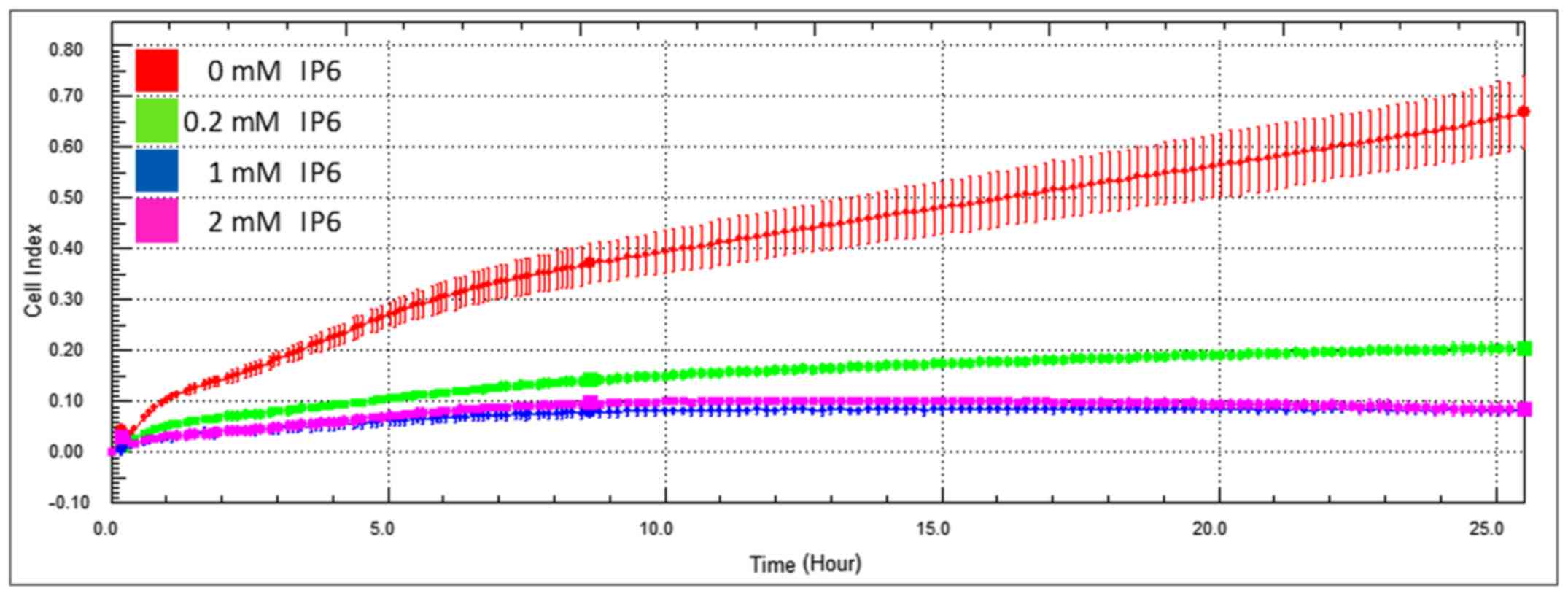
Firstly, the effect of various IP6 concentrations on
the migration of colorectal cancer SW620 cells was evaluated. The
three tested non-toxic IP6 concentrations (0.2, 1 and 2 mM) were
chosen based on our previous experiments (19) and cytotoxicity testing (half
maximal inhibitory concentration, 3.63 mM IP6). All tested
concentrations significantly decreased the cell migration of the
SW620 cells in a dose-dependent manner (Fig. 1). A significantly pronounced effect
was already observable using 0.2 and 1 mM concentrations (the
effect of a 2 mM concentration of IP6 was comparable with that of a
1 mM concentration of IP6); therefore these two concentrations were
chosen for further experiments.
Expression of MMP markers
A significant suppression of the MMPs involved in
cancer progression (MMP-2 and MMP-9) at the mRNA levels was
observed at the 24 and 48 h tested time intervals following
treatment with 0.2 and 1 mM concentrations of IP6 (Fig. 2).
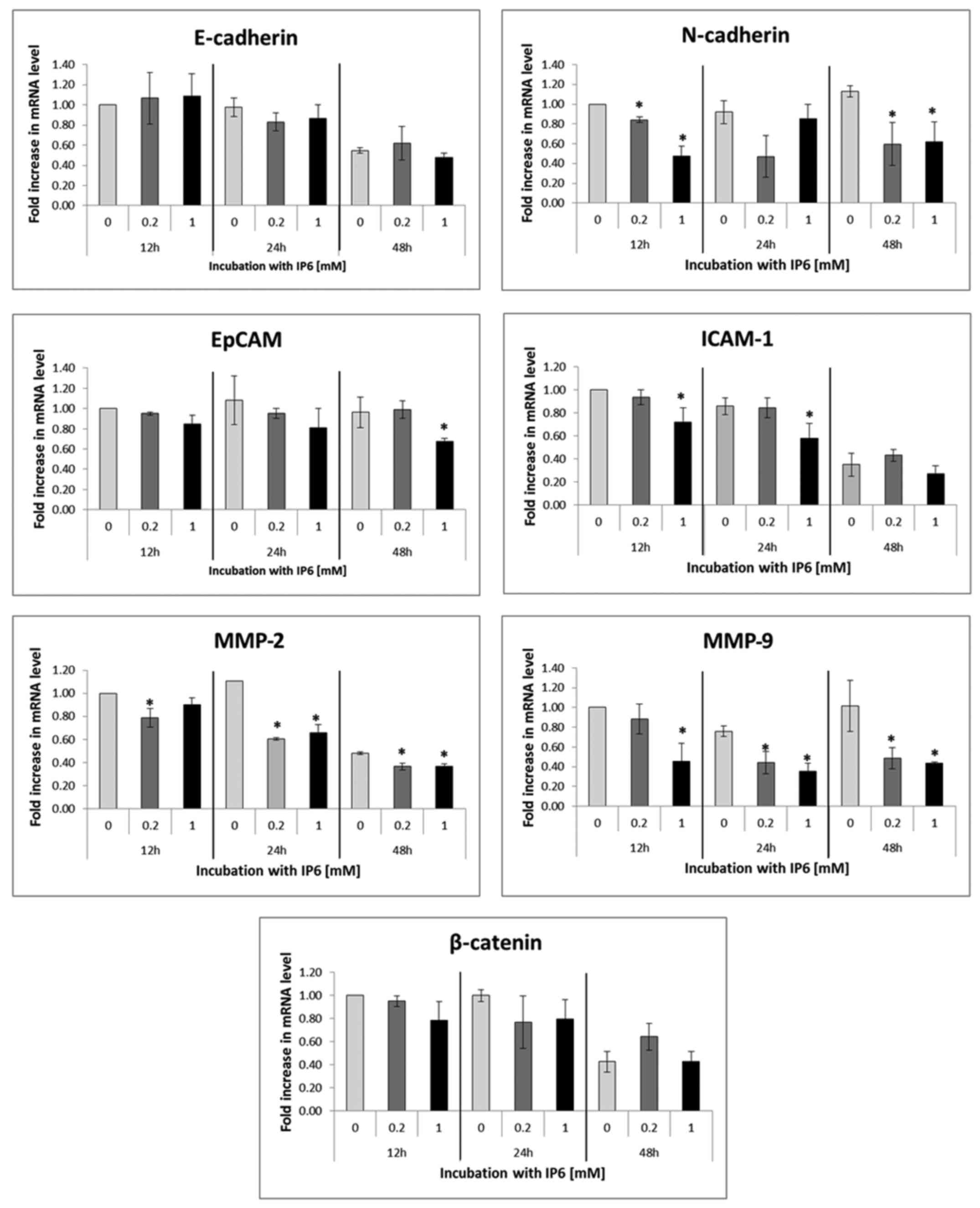 | Figure 2Reverse transcription-quantitative
polymerase chain reaction. The changes in relative mRNA expression
of molecules involved in cancer progression (MMP2, MMP9, EpCAM,
ICAM, E-cadherin, N-cadherin and β-catenin) following treatment
with 0, 0.2 and 1 mM concentrations of IP6 for 12, 24 and 48 h. The
amounts of mRNA of the target genes was normalized to the GAPDH
reference gene. The data are expressed as the fold increase of the
untreated cells (=1). *Significant change of means from
three independent experiments performed in duplicates (P<0.05)
identified by one-way analysis of variance with Bonferroni's
modification. IP6, inositol hexaphosphate; MMP, matrix
metalloproteinase; EpCAM, epithelial cell adhesion molecule;
ICAM-1, intercellular adhesion molecule 1. |
Expression of adhesion and migration
markers
In the next step, the study focused on the
expression of selected adhesion and migration markers at the mRNA
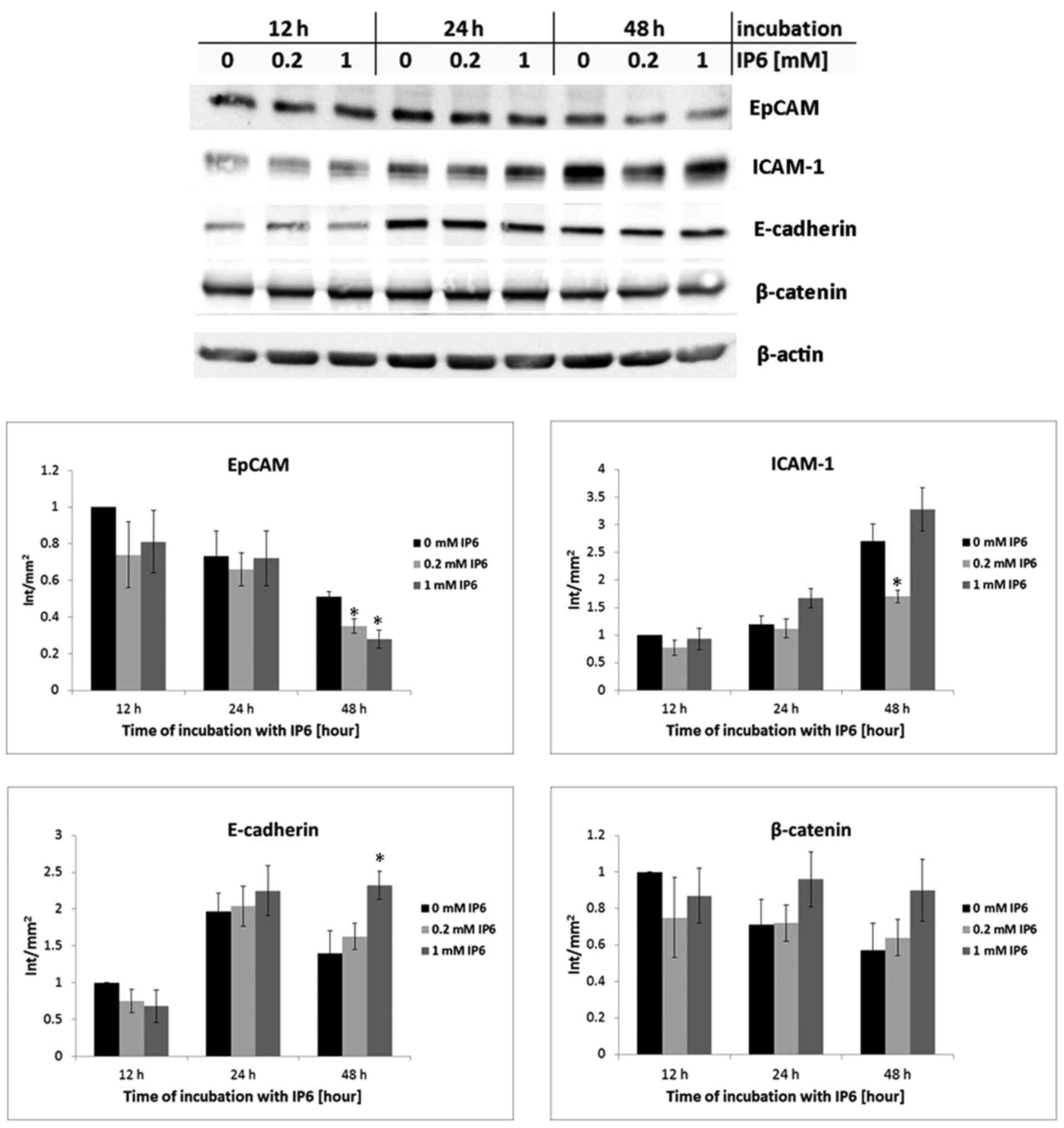
(Fig. 2) and/or protein levels
(Fig. 3). IP6 treatment led to
significant decrease in the N-cadherin levels. Changes in the mRNA
levels of EpCAM and ICAM could be observed only in isolated time
intervals and concentrations (in EpCAM following treatment with 1
mM IP6 at 48 h and in ICAM-1 following treatment with 1 mM IP6 at
12 and 24 h), while the mRNA levels of E-cadherin and β-catenin
were not significantly altered by IP6 at all (Fig. 2). In addition, the protein levels
of β-catenin remained unchanged in IP6-treated SW620 cells in all
time intervals and concentrations compared with the untreated (0 mM
IP6) controls (P>0.05). A significant decrease in EpCAM mRNA
expression was observed only at the highest concentration and time
interval tested, while ICAM-1 expression decreased transiently at
12 and 24 h post-treatment with 1 mM IP6. A significant decrease in
protein levels in comparison to the control group (0 mM IP6) was
observed in EpCAM at the 48-h time interval and at the two tested
concentrations, while in ICAM-1, only at the concentration of 0.2
mM IP6 after 48 h. By contrast, a statistically significant
increase in E-cadherin at the protein level was found after 48 h of
treatment (1 mM concentration of IP6).
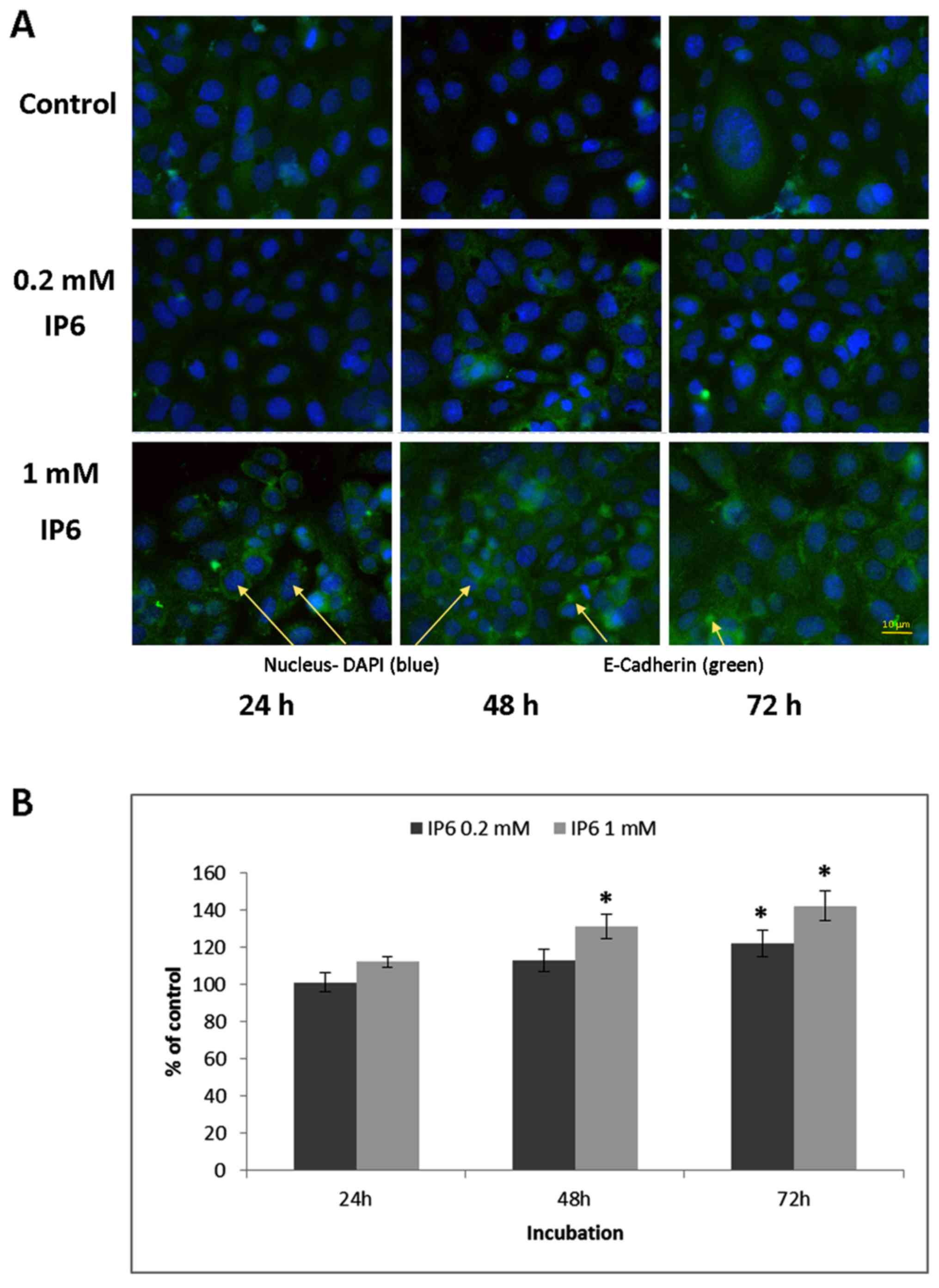
The study also investigated the subcellular
localization of E-cadherin following IP6 treatment.
Immunofluorescence analysis of E-cadherin expression in IP6-treated
SW620 cells demonstrated significantly increased membrane and
cortical localization of this molecule (Fig. 4), in particular after 48 and 72 h
of treatment.
Discussion
It has been suggested that IP6 has a number of
biological functions, including a role in signal transduction, cell
proliferation and differentiation (20). The anticancer effect of IP6 has
been demonstrated in cells isolated from colorectal cancer
(3), liver (14), Barrett's oesophagus (21), prostate (22), breast (16), pancreatic (23), skin (7) and rhabdomyosarcoma (24) tissues, where IP6 acted chiefly via
suppressed proliferation with concomitantly induced apoptosis.
IP6 has also been shown to exert inhibitory effects
on invasiveness and metastasis. A study on the mouse metastatic
FSA-1 cell line showed that injecting IP6 intraperitoneally reduced
subcutaneously transplanted fibrosarcoma growth in mice and reduced
the number of pulmonary metastases (25). Another study confirmed the
anti-metastatic effect of IP6 on the invasive breast MDA-MB231 cell
line via reduced migration and the inhibition of the secretion of
MMP-9 (16). The follow-up study
indicated that the inhibition of cancer cell adhesion, migration
and invasion induced by IP6 may be mediated by modulating integrin
dimerization, cell surface expression and the integrin-associated
signaling pathway (17).
In the present study, it was found that the
concentrations of 0.2 and 1 mM IP6 significantly inhibited the
migration of SW620 cells during 24 h of treatment, as measured
using the xCELLigence RTCA-DP system. Furthermore, functioning in a
time and concentration manner, IP6 affected the expression of
several key molecules involved in cellular adhesion and epithelial
to mesenchymal transition. E-cadherin is a transmembrane protein
mediating cell adhesion via the E-cadherin-β-catenin-α-catenin
complex. A defect in or loss of E-cadherin expression enables the
uncontrolled transcriptional activity of β-catenin in the affected
cells, which is associated with the development of invasive and
metastatic potential (26,27). To this end, a recently published
study on rats with colorectal cancer showed the inhibitory effect
of IP6 on β-catenin activity (28). However, in the present study using
the SW620 cell line, no significant changes were found in β-catenin
expression following IP6 treatment at the mRNA and protein levels.
By contrast, IP6 enabled the increased expression and membrane
localization of E-cadherin in the treated cells, which would
indicate elevated cell adhesion.
N-cadherin belongs to the transmembrane adhesion
proteins whose expression is required for collective cell
migration. Increased N-cadherin levels are correlated with
epithelial to mesenchymal transition and tumor invasion (29). In the present study, the expression
of N-cadherin decreased at all time intervals upon treatment with
0.2 and 1 mM IP6 concentrations. These changes were further
associated with its subcellular localization (data not shown).
Transmembrane glycoprotein EpCAM is involved in
cellular proliferation, migration and differentiation. It is also
known to be highly expressed in epithelial carcinomas (30). ICAM-1 is expressed in endothelial
cells (31). While normal colonic
cells lack ICAM-1, tumor cells are known to exhibit increased
expression of this molecule. A previous study determined that
increased expression was present in colon cancer cells and that
well-differentiated tumors exhibited the highest levels (32). IP6 treatment changed the expression
of the two aforementioned markers, although these changes did not
show a decisively linear trend, suggesting that these molecules are
not a primary marker of IP6 in this model.
MMPs serve an important role in the local and
systemic spread of malignant tumors. MMP-2 and MMP-9 degrade the
extracellular matrix to enable invasion and metastasis of cells.
IP6 has already demonstrated an inhibitory effect on MMP-9
secretion in breast cancer MDA-MB231 cells (16). Another study in Caco-2 cells
demonstrated an increase in the expression of MMP-2 mRNA following
treatment with 1 mM IP6 for 1 h, followed by a decrease in the
expression at longer time intervals, while the expression of MMP-9
was neither constitutively expressed nor induced by IP6 (33). The results of the present study
suggest a significant decrease in MMP-2 and MMP-9 mRNA levels in
SW620 cells exposed to IP6 at all treatment intervals and used
doses.
In conclusion, the present results demonstrate the
ability of IP6 to alter the migration, adhesion and invasion in the
SW620 cell line. Thus, IP6 is a promising anticancer agent that
exerts its effects on multiple aspects of CRC progression. The
reduction in N-cadherin levels and increase in E-cadherin levels
indicate that one of the IP6 targets could be the reversion of the
epithelial-mesenchymal transition; however, levels of other EMT
markers following IP6 treatment should be investigated in future
studies.
Acknowledgments
Not applicable.
Abbreviations:
|
CRC
|
colorectal cancer
|
|
DMEM
|
Dulbecco's modified Eagle's medium
|
|
EpCAM
|
epithelial cell adhesion molecule
|
|
FBS
|
fetal bovine serum
|
|
ICAM-1
|
intercellular adhesion molecule 1
|
|
IP6
|
inositol hexaphosphate
|
|
MMP
|
matrix metalloproteinase
|
|
PBS
|
phosphate-buffered saline
|
|
RT-qPCR
|
reverse transcription-quantitative
polymerase chain reaction
|
|
TBST
|
Tris-buffered saline (TBS) plus
Tween-20
|
Funding
This study was supported by the grant PROGRESS Q40
01 of Charles University, Faculty of Medicine in Hradec
Králové.
Availability of data and materials
The authors declare that the datasets used and/or
analysed during the current study are available from the
corresponding author on reasonable request
Authors' contributions
LS and VH designed the study and experiments; LS,
AJ, ER, KC and VH performed the experiments; LS and VH conducted
the statistical analysis; and LS, VK, VH, ER and KC wrote the
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
McGuire S: World Cancer Report 2014.
Geneva, Switzerland: World Health Organization, International
Agency for Research on Cancer, WHO Press; 2015
Adv Nutr. 7:418–419. 2016. View Article : Google Scholar
|
|
2
|
Arnold M, Sierra MS, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global patterns and trends in
colorectal cancer incidence and mortality. Gut. 66:683–691. 2017.
View Article : Google Scholar
|
|
3
|
Liu G, Song Y, Cui L, Wen Z and Lu X:
Inositol hexaphosphate suppresses growth and induces apoptosis in
HT-29 colorectal cancer cells in culture: PI3K/Akt pathway as a
potential target. Int J Clin Exp Pathol. 8:1402–1410.
2015.PubMed/NCBI
|
|
4
|
Vucenik I, Ramakrishna G, Tantivejkul K,
Anderson LM and Ramljak D: Inositol hexaphosphate (IP6) blocks
proliferation of human breast cancer cells through a
PKCdelta-dependent increase in p27Kip1 and decrease in
retinoblastoma protein (pRb) phosphorylation. Breast Cancer Res
Treat. 91:35–45. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Vucenik I, Tantivejkul K, Zhang ZS, Cole
KE, Saied I and Shamsuddin AM: IP6 in treatment of liver cancer. I
IP6 inhibits growth and reverses transformed phenotype in HepG2
human liver cancer cell line. Anticancer Res. 18:4083–4090.
1998.
|
|
6
|
Gu M, Roy S, Raina K, Agarwal C and
Agarwal R: Inositol hexaphosphate suppresses growth and induces
apoptosis in prostate carcinoma cells in culture and nude mouse
xenograft: PI3K-Akt pathway as potential target. Cancer Res.
69:9465–9472. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wawszczyk J, Kapral M, Lodowska J, Jesse
K, Hollek A and Węglarz L: Antiproliferative effect of inositol
hexaphosphate on human skin melanoma cells in vitro. Acta Pol
Pharm. 72:895–900. 2015.PubMed/NCBI
|
|
8
|
Wei J, Cheang T, Tang B, Xia H, Xing Z,
Chen Z, Fang Y, Chen W, Xu A, Wang S, et al: The inhibition of
human bladder cancer growth by calcium carbonate/CaIP6
nanocomposite particles delivering AIB1 siRNA. Biomaterials.
34:1246–1254. 2013. View Article : Google Scholar
|
|
9
|
El-Sherbiny YM, Cox MC, Ismail ZA,
Shamsuddin AM and Vucenik I: G0/G1 arrest and S phase inhibition of
human cancer cell lines by inositol hexaphosphate (IP6). Anticancer
Res. 21:2393–2403. 2001.PubMed/NCBI
|
|
10
|
Owen RW, Weisgerber UM, Spiegelhalder B
and Bartsch H: Faecal phytic acid and its relation to other
putative markers of risk for colorectal cancer. Gut. 38:591–597.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bozsik A, Kökény S and Olah E: Molecular
mechanisms for the antitumor activity of inositol hexakisphosphate
(IP6). Cancer Genomics Proteomics. 4:43–51. 2007.PubMed/NCBI
|
|
12
|
Cholewa K, Parfiniewicz B, Bednarek I,
Swiatkowska L, Jezienicka E, Kierot J and Weglarz L: The influence
of phytic acid on TNF-alpha and its receptors genes' expression in
colon cancer Caco-2 cells. Acta Pol Pharm. 65:75–79.
2008.PubMed/NCBI
|
|
13
|
Sakamoto K, Venkatraman G and Shamsuddin
AM: Growth inhibition and differentiation of HT-29 cells in vitro
by inositol hexaphosphate (phytic acid). Carcinogenesis.
14:1815–1819. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Fu M, Song Y, Wen Z, Lu X and Cui L:
Inositol hexaphosphate and inositol inhibit colorectal cancer
metastasis to the liver in BALB/c mice. Nutrients. 8:82016.
View Article : Google Scholar
|
|
15
|
Kapral M, Wawszczyk J, Jurzak M, Hollek A
and Węglarz L: The effect of inositol hexaphosphate on the
expression of selected metalloproteinases and their tissue
inhibitors in IL-1β-stimulated colon cancer cells. Int J Colorectal
Dis. 27:1419–1428. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tantivejkul K, Vucenik I and Shamsuddin
AM: Inositol hexaphosphate (IP6) inhibits key events of cancer
metastasis: I. In vitro studies of adhesion, migration and invasion
of MDA-MB231 human breast cancer cells. Anticancer Res.
23:3671–3679. 2003.PubMed/NCBI
|
|
17
|
Tantivejkul K, Vucenik I and Shamsuddin
AM: Inositol hexaphosphate (IP6) inhibits key events of cancer
metastasis: II. Effects on integrins and focal adhesions.
Anticancer Res. 23:3681–3689. 2003.PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
19
|
Schroterova L, Haskova P, Rudolf E and
Cervinka M: Effect of phytic acid and inositol on the proliferation
and apoptosis of cells derived from colorectal carcinoma. Oncol
Rep. 23:787–793. 2010.PubMed/NCBI
|
|
20
|
Shamsuddin AM, Vucenik I and Cole KE: IP6:
A novel anticancer agent. Life Sci. 61:343–354. 1997. View Article : Google Scholar
|
|
21
|
McFadden DW, Riggs DR, Jackson BJ and
Cunningham C: Corn-derived carbohydrate inositol hexaphosphate
inhibits Barrett's adenocarcinoma growth by pro-apoptotic
mechanisms. Oncol Rep. 19:563–566. 2008.PubMed/NCBI
|
|
22
|
Sharma G, Singh RP and Agarwal R: Growth
inhibitory and apoptotic effects of inositol hexaphosphate in
transgenic adenocarcinoma of mouse prostate (TRAMP-C1) cells. Int J
Oncol. 23:1413–1418. 2003.PubMed/NCBI
|
|
23
|
Somasundar P, Riggs DR, Jackson BJ,
Cunningham C, Vona-Davis L and McFadden DW: Inositol hexaphosphate
(IP6): A novel treatment for pancreatic cancer. J Surg Res.
126:199–203. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Vucenik I, Kalebic T, Tantivejkul K and
Shamsuddin AM: Novel anticancer function of inositol hexaphosphate:
Inhibition of human rhabdomyosarcoma in vitro and in vivo.
Anticancer Res. 18:1377–1384. 1998.PubMed/NCBI
|
|
25
|
Vucenik I, Tomazic VJ, Fabian D and
Shamsuddin AM: Antitumor activity of phytic acid (inositol
hexaphosphate) in murine transplanted and metastatic fibrosarcoma,
a pilot study. Cancer Lett. 65:9–13. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Oloumi A, McPhee T and Dedhar S:
Regulation of E-cadherin expression and beta-catenin/Tcf
transcriptional activity by the integrin-linked kinase. Biochim
Biophys Acta. 1691:1–15. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Stockinger A, Eger A, Wolf J, Beug H and
Foisner R: E-cadherin regulates cell growth by modulating
proliferation-dependent beta-catenin transcriptional activity. J
Cell Biol. 154:1185–1196. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Saad N, Esa NM and Ithnin H: Suppression
of β-catenin and cyclooxygenase-2 expression and cell proliferation
in azoxymethane-induced colonic cancer in rats by rice bran phytic
acid (PA). Asian Pac J Cancer Prev. 14:3093–3099. 2013. View Article : Google Scholar
|
|
29
|
Todosi AM, Gavrilescu MM, Aniţei GM, Filip
B and Scripcariu V: Colon cancer at the molecular level -
usefulness of epithelial-mesenchymal transition analysis. Rev Med
Chir Soc Med Nat Iasi. 116:1106–1111. 2012.
|
|
30
|
Dai M, Yuan F, Fu C and Shen G, Hu S and
Shen G: Relationship between epithelial cell adhesion molecule
(EpCAM) overexpression and gastric cancer patients: A systematic
review and meta-analysis. PLoS One. 12:e01753572017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yang Y, Jun CD, Liu JH, Zhang R,
Joachimiak A, Springer TA and Wang JH: Structural basis for
dimerization of ICAM-1 on the cell surface. Mol Cell. 14:269–276.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Paschos KA, Canovas D and Bird NC: The
role of cell adhesion molecules in the progression of colorectal
cancer and the development of liver metastasis. Cell Signal.
21:665–674. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kapral M, Wawszczyk J, Jurzak M, Dymitruk
D and Weglarz L: Evaluation of the expression of metalloproteinases
2 and 9 and their tissue inhibitors in colon cancer cells treated
with phytic acid. Acta Pol Pharm. 67:625–629. 2010.
|