Introduction
Inflammation is a double-edged sword in cancer.
Inflammation was initially believed to be a host response against
tumors, resulting in tumor suppression and a favorable prognosis
(1). However, it has been reported
that inflammation, particularly chronic inflammation, is associated
with an unfavorable clinical prognosis of cancer patients (2,3), and
inflammation is now suggested as the seventh hallmark for cancer
establishment and progression (4).
There are abundant data suggesting that inflammation and hypoxia in
the tumor microenvironment are critical components that are
necessary for tumor progression and the metastatic cascade
(5). Indeed, such an environment
is more permissive for tumor cell proliferation and motility than
are normal conditions. Moreover, several studies have indicated
that tumor cell signaling and extracellular signaling affect cancer
cell migration and therefore, metastasis in vivo and in
vitro (reviewed in ref. 6).
However, the innate pathways or mechanisms controlling the
inflammatory response in the tumor microenvironment are not yet
fully understood.
Pro-inflammatory cytokines, such as interleukin
(IL)-1β and IL-18, are detected at high levels in cancer patients,
and are suggested to promote an immune-suppressive tumor
microenvironment (4,7, 8).
The inflammasome is an important innate immune pathway responsible
for the production of mature IL-1β. Inflammasome sensors are
classified according to their structural features into
nucleotide-binding domain-like receptors (NLRs), absent in melanoma
2-like receptors (ALRs), and the recently identified pyrin. These
receptors can assemble the inflammasome and activate the cysteine
protease, caspase-1. Active caspase-1 cleaves the precursor
pro-inflammatory cytokines, pro-IL-1β and pro-IL-18, into their
mature secreted forms, and these cytokines can ultimately be
released (9). In particular, IL-1β
is abundant in tumor tissue and enhances tumor growth, invasion,
carcinogenesis and host-tumor interactions (10,11),
and increased concentrations of IL-1β in tumor tissues are
associated with a poor prognosis in cancer patients (12-14),
suggesting that IL-1β is one of the essential components that
mediate inflammation-associated tumor progression.
Of note, the inflammasome has been reported to be
activated by adenosine triphosphate (ATP) (15). Various cellular stimuli trigger the
secretion of ATP (16,17) and subsequently induce the
activation of purinergic receptors present on the cell surface
and/or on adjacent cells. Under pathological conditions, ATP is
released passively from damaged cells at high levels, acts as a
pro-inflammatory danger signal, and activates the NLRP3
inflammasome through bonding to the P2 purinergic receptor, P2Y
purinergic receptor 2 (P2X7R) (15). Recent studies have reported that
ATP is released from both damaged cells and tumor cells and
accumulates in the tumor microenvironment, which can be related to
tumor progression (18,19). Among the purinergic receptors that
are activated by ATP, P2Y2R is expressed (or
overexpressed) in cancer cells or solid tumors and performs various
functions; it regulates proliferation in various tumors, such as
lung, bladder, and prostate cancer and melanoma (20-23).
In our previous studies, we reported that highly metastatic
MDA-MB-231 breast cancer cells released higher levels of ATP and
exhibited a higher P2Y2R activity than the MCF7 breast
cancer cells with a low metastatic potential (24). In addition, ATP-activated
P2Y2R played an important role in tumor progression,
particularly in invasion and metastasis, by regulating
hypoxia-inducible factor-1α (HIF-1α) (24,25).
In general, cancer patients are treated based on a
combinatorial approach that consists of surgery, chemotherapy and
radiotherapy. However, each therapy has inherent limitations that
lead to therapeutic resistance and disease recurrence, ultimately
resulting in therapeutic failure. Radiotherapy is a crucial
treatment option in modern cancer therapy in addition to surgery
and systemic therapy; currently, >60% of all cancer patients
receive radiotherapy. Radiotherapy has been shown to improve
overall survival (26-28), to help avoid surgical amputation
and to preserve bodily beauty, and it can be used in palliative
settings (29,30). Although benefits are achievable
with radiotherapy, tumor recurrence following radiotherapy is
common; particularly for ductal carcinoma and early invasive
cancer, advanced invasive tumors can exhibit radiotherapy
resistance, and the related molecular mechanisms are poorly
understood. Thus, in this study, we established radiotherapy-
resistant (RT-R)-breast cancer cells and investigated the
association between P2Y2R and the inflammasome in RT-R-
breast cancer cell progression and invasiveness.
Materials and methods
Cell culture
The human breast cancer cell lines, MDA-MB-231, MCF7
and T47D, were obtained from the Korea Cell Line Bank and grown in
RPMI-1640 supplemented with 10% FBS (HyClone, Logan, UT, USA), 100
U/ml penicillin and 10 µg/ml streptomycin (HyClone). The
human umbilical endothelial cell line, EA.hy926, was obtained from
ATCC (Manassas, VA, USA) and grown in DMEM supplemented with 10%
FBS, 100 U/ml penicillin and 10 µg/ml streptomycin.
Establishment of RT-R MDA-MB-231 MCF7 and
T47D cells (termed RT-R-MDA-MB-231, RT-R-MCF7 and RT-R- T47D cells,
respectively)
Isogenic models of radiotherapy resistance can be
generated by the exposure of cancer cells to various schedules with
the total concentrations within a 40-60 Gy range (31,32).
Moreover, the fractionated irradiation is clinically universal.
Clinical total body irradiation is generally fractionated with
smaller doses delivered in several sessions, rather than delivering
the entire doses at once, due to lower toxicity and better outcomes
(33). Thus, in this study,
RT-R-MDA-MB-231, RT-R-MCF7 and RT-R-T47D cells were generated by
treating the cells with fractionated X-ray irradiation until a
final concentration of 50 Gy was attained. In detail, the cells
that had grown to 70% confluence in a cell culture flask were
irradiated with 2 Gy (radiation dose rate, 1.0 Gy/min) using a 6-MV
photon beam produced by a linear accelerator (Clinac 21EX; Varian
Medical Systems, Palo Alto, CA, USA). Following irradiation, the
cells were incubated with fresh complete medium immediately. When
the cell confluence reached approximately 90%, the cells were
trypsinized and subcultured into new flasks. The cells were
irradiated again when they grew to approximately 70% confluence.
Until the total irradiation dose attained 50 Gy, the fractionated
irradiations were continued. The parental cells were subjected to
identical trypsinization, subculture and incubation conditions, but
were not subjected to irradiation. The RT-R-MDA-MB-231, RT-R-MCF7
and RT-R-T47D cells were used through 5 passages. The radiation
output was regularly checked by medical physicists in the
Department of Radiation Oncology using an ionization detector.
Extracellular ATP release
measurements
The extracellular release of ATP was measured
according to previously described methods (24). Briefly, the cells were incubated
with HEPES buffer containing adenosine-5′-O-(α,β-methylene)-
diphosphonate (AOPCP), an ectonucleotidase inhibitor
(Sigma-Aldrich, St. Louis, MO, USA) for 15 min at 37°C. The cells
were treated with 10 ng/ml tumor necrosis factor (TNF)-α (R&D
Systems, Minneapolis, MN, USA) or PBS as a vehicle for an
additional 5 min, and the supernatants were then collected. ATP
release was measured with the ENLITEN ATP assay system kit
(Promega, Madison, WI, USA), and the ATP levels were calculated
based on an ATP standard curve.
Measurement of intracellular calcium ion
concentration ([Ca2+]i)
[Ca2+]i measurements were made
according to previously described methods (34). Briefly, the cells were seeded on a
coverslip mounted onto a self-designed perfusion chamber, and then
incubated for 45 min with 5 µM fluo-3-AM (Invitrogen,
Carlsbad, CA, USA) in culture medium. The stained cells were washed
with a physiological solution and then treated with ATP.
Fluorescent images were scanned every 5 sec using a confocal laser
scanning microscope (IX70 Fluoview, Olympus, Tokyo, Japan). Every
scanned image was processed to analyze changes in
[Ca2+]i. The basal fluorescence intensity
(F0), fluorescence intensity (F), and the maximum level of
fluorescence intensity (Fmax) were recorded.
Matrigel invasion assay
For invasion assays, the upper chambers of inserts
were coated with 100 µl of Matrigel (1 mg/ml; BD
Biosciences, Franklin Lakes, NJ, USA), and endothelial cells
(2×105 cells) were added to the Matrigel-coated insert
wells. The breast cancer cells were pre-treated with 20 µM
Ac-YVAD-CMK (Sigma-Aldrich), a selective and irreversible inhibitor
of caspase-1, for 1 h, and then stimulated with 10 ng/ml TNF-α or
10 µM ATP. After 6 h, the cells were harvested, and
2×105 cells per insert were added to the upper chambers
in serum-free medium, and 500 µl of RPMI medium was added to
the lower chambers. The invasion chambers were incubated for 24 h
in a 37°C cell culture incubator. The non-invaded cells that
remained on the upper surface of the insert membranes were removed
by scrubbing. The cells that had invaded across the insert well
membrane were stained with 4′,6-diamidine-2′-phenylindole
dihydrochloride (DAPI, Sigma-Aldrich), and the cells were counted
under a fluorescence microscope (Eclipse Ti-U, Nikon, Tokyo,
Japan).
Caspase-1 activity assay
Caspase-1 activity was measured using Caspase-1/ICE
Colorimetric Assay kit (R&D Systems) following the
manufacturer’s instructions. Briefly, the cells were treated with
10 ng/ml TNF-α or 10 µM ATP for indicated times. As shown in
Fig. 3, the cells were transfected
with indicated siRNA (100 nM) as described below, and then
pre-treated with 10 U/ml apyrase (Sigma-Aldrich), an enzyme that
rapidly hydrolyzes extracellular nucleotides. After 1 h, the cells
were stimulated with 10 ng/ml TNF-α or 10 µM ATP for 3 h,
and the total proteins were then extracted from the cells using
lysis buffer. A volume of 50 µl of protein sample from cells
was added to 50 µl of 2X caspase-1 reaction buffer
containing 10 mM dithiothreitol (DTT) in a 96-well plate. Five
microliters of 4 mM caspase-1 colorimetric substrate (YVAD-pNA)
were added to each sample and then incubated at 37°C for 1-2 h. The
colorimetric intensity was measured at a wavelength of 405 nm using
a microplate reader (Tecan, Männedorf, Switzerland).
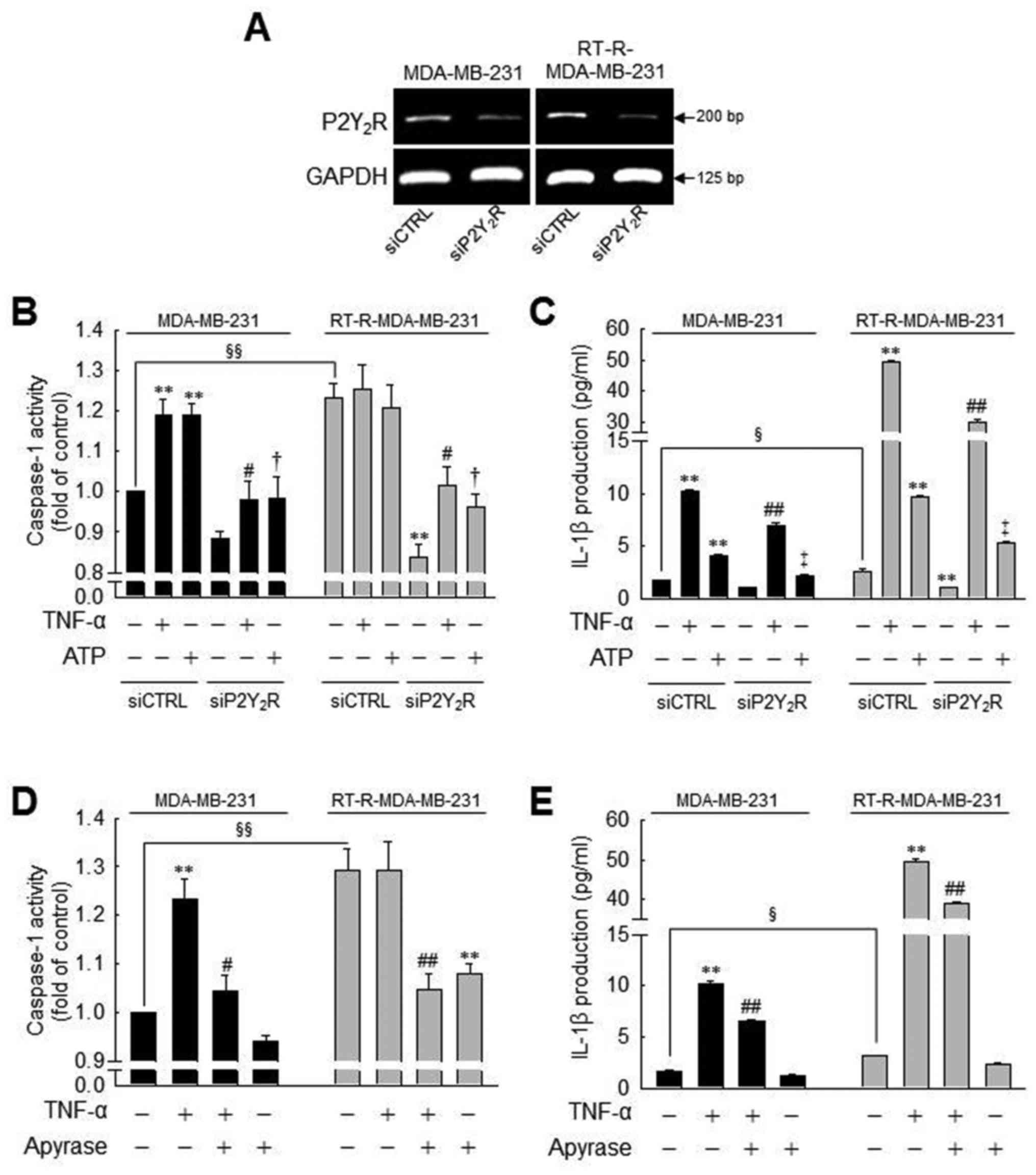 | Figure 3Caspase-1 activity or IL-1β secretion
induced by TNF-α or ATP is significantly suppressed by
P2Y2R knockdown or apyrase (an enzyme that rapidly
hydrolyzes extracellular nucleotides) in MDA-MB-231 or
RT-R-MDA-MB-231 cells. (A) P2Y2R mRNA levels were
analyzed by RT-PCR to confirm the efficiency of the knockdown in
control siRNA (siCTRL)- or P2Y2R siRNA
(siP2Y2R)-transfected cells. (B and C) siCTRL or
siP2Y2R-transfected cells were stimulated with TNF-α (10
ng/ml) or ATP (10 µM). (B) Caspase-1 activity and (C) IL-1β
secretion were measured 3 h and 24 h after treatment, respectively,
as described in the Materials and methods. (D and E) The cells were
pre-treated with 10 U/ml apyrase for 1 h and stimulated with TNF-α
(10 ng/ml). (D) Caspase-1 activity and (E) IL-1β secretion were
measured as described in (A). The values represent the means ± SEM
of 3 independent experiments. **P<0.01, compared to
the control (CTRL) of each cells; #P<0.05 and
##P<0.05, compared to the TNF-α treatment of each of
the cells; †P<0.05 and ‡P<0.01,
compared to the ATP treatment of each of the cells;
§P<0.05 and §§P<0.01, significance
between the MDA-MB-231 and RT-R-MDA-MB-231 cells. ATP, adenosine
triphosphate; RT-R, radiotherapy-resistant; IL-1β, interleukin-1β;
TNF-α, tumor necrosis factor-α. |
Quantification of IL-1β secretion
To quantify the amounts of secreted cytokines, cell
culture supernatants or animal serum samples were assayed using the
Human IL-1β/IL-1F2 Quantikine ELISA kit (R&D Systems),
according to the manufacturer’s instructions. Briefly, the cells
were treated as described above in the ‘Caspase-1 activity assay’.
Following stimulation with 10 ng/ml TNF-α or 10 µM ATP for
24 h, cell culture supernatants were collected. Mouse serum was
obtained by heart puncture before sacrifice and centrifugation was
used for the measurement of the IL-1β levels. A volume of 200
µl of sample was added to a microplate and sequentially
mixed with Conjugate, Substrate Solution and Stop Solution. The
optical density of each well was measured at a wavelength of 450 nm
using a microplate reader (Tecan).
Gene silencing with siRNA
Breast cancer cells and RT-R breast cancer cells
were transfected with 100 nM negative control siRNA (siCTRL) or
P2Y2R siRNA (siP2Y2R) (Bioneer, Daejeon,
Korea) in serum-containing medium using Turbofect®
(Thermo Fisher Scientific, Waltham, MA, USA). The sequences of the
siRNAs were as follows: siCTRL forward, 5′-CCUACGCCACCAAUUUCGU-3′
and reverse, 5′-ACGAA AUUGGUGGCGUAGG-3′; siP2Y2R
forward, 5′-GAGGAAGGUGGCUUACCAA-3′ and reverse,
5′-UUGGUAAGCCACCUUCCUC-3′. Following 24 h of incubation at 37°C,
the transfection medium was replaced with fresh serum-free medium
for starvation. Following serum starvation for 16 h, the cells were
treated with the indicated reagents. Gene silencing efficiency was
determined by reverse transcription-polymerase chain reaction
(RT-PCR).
RT-PCR
Total RNA was extracted from the cells using TRIzol
reagent (Thermo Fisher Scientific), and RT-PCR was performed using
TOPscript One-step RT PCR Drymix (Enzynomics, Daejeon, Korea),
according to the manufacturer’s instructions. The primer sets used
were as follows: hP2Y2R forward, 5′-GTG CTC TAC TTC CTG
GCT-3′ and reverse, 5′-CTG AAG TGT TCT GCT CCT AC-3; ′ and hGAPDH
forward, 5′- TCA ACA GCG ACA CCC ACT CC-3′ and reverse, 5′-TGA GGT
CCA CCA CCC TGT TG-3′. Thirty cycles of amplification were
performed under the following conditions: Melting at 95°C for 30
sec, annealing at 56°C for 30 sec and extension at 72°C for 30
sec.
Colony formation assay
The MDA-MB-231 or RT-R- MDA-MB-231 cells
(1×103) were seeded in 6-well plates. Following serum
starvation for 16 h, the cells were pre-treated with 20 µM
Ac-YVAD-CMK for 1 h, and then stimulated with 10 ng/ml TNF-α or 10
µM ATP at 37°C. The culture medium was discarded following
treatment, and changed with complete medium every 2-3 days. After
10 days, the medium was discarded and each well was carefully
washed with PBS. The colonies were fixed in methanol for 10 min at
room temperature and then stained with 0.1% Giemsa staining
solution, and the number of visible colonies was counted.
Gelatin zymography
The cells which were transfected with the indicated
siRNA (100 nM or the untransfected cells were pre-treated with 20
µM Ac-YVAD-CMK or 10 µM AR-C 118925XX (Tocris
Bioscience, Bristol, UK), a specific P2Y2R antagonist.
After 1 h, the cells were stimulated with 10 ng/ml TNF-α or 10
µM ATP for 6 h, and the same volume of each conditioned
medium was then concentrated 20-fold using protein concentrators
(9K MWCO; Thermo Fisher Scientific) at a fixed angle (35 degrees)
and centrifugation at 6,000 × g for 25 min at 4°C. The concentrated
samples were mixed with 2X loading dye, and the proteins were
separated on 8% SDS-polyacrylamide gels containing 1 mg/ml gelatin.
Following electrophoresis, the gels were washed in 2.5% Triton
X-100 twice for 30 min to remove the remaining SDS. The gels were
then incubated in developing buffer (50 mM Tris, 20 mM NaCl, 5 mM
CaCl2, 0.02% Brij35) at 37°C overnight. Following
incubation, the gels were stained with coomassie blue solution
(0.2% coomassie brilliant blue R, 50% methanol, 10% acetic acid)
for 30 min at room temperature, and destained with destaining
buffer (50% methanol, 10% acetic acid). Enzyme-digested regions
which represent matrix metalloproteinase (MMP)-9 activity were
identified as white bands on a blue background.
Animal experiments
RT-R-MDA-MB-231 cells were transfected with the
P2Y2R shRNA plasmid (Santa Cruz Biotechnology Inc.,
Dallas, TX, USA) which contains a puromycin resistance gene for the
selection of cells stably expressing targeted shRNA in serum-free
medium using Lipofectamine 2000 (Thermo Fisher Scientific).
Following 4 h of incubation at 37°C, the transfection medium was
replaced with fresh medium containing 5 µg/ml puromycin
(Sigma-Aldrich). The culture medium containing puromycin was
changed every 2-3 days. At 30 days following transfection, the
stably treasfected subclone was designated
RT-R-MDA-MB-231-P2Y2R shRNA. This subclone and a CTRL
subclone transfected with an empty vector (designated as
MDA-MB-231-EV) were grown in serum-containing culture medium until
the cell density was ~70-80%. The cells were then trypsinized, and
the pellets were resuspended in serum-free RPMI at
5-6×106 cells/100 µl of cell suspension. A total
of 10 female NU-Foxn1nu athymic nude mice at 7-8 weeks of age
(weighing 20-22 g) were purchased from OrientBio (Gyeonggi-do,
Korea). The animals were maintained under the following
environmental conditions: 22-26°C; 40-60% of humidity, 12
h-light/dark cycle; with free access to sterilized feed and water.
The mice were injected subcutaneously with
RT-R-MDA-MB-231-P2Y2R shRNA or RT-R-MDA-MB-231-EV. Body
weights and tumor volumes were measured every 3 days, starting at 7
days after the injection. At the end of 60 days, the mice were
sacrificed, and the tumor tissues were fixed in 4% formaldehyde at
room temperature, followed by paraffin infiltration and embedding.
Sections of 5 µm thickness were mounted onto ProbeOn Plus
microscope slides (Thermo Fisher Scientific). Immunohistochemical
analysis was performed using a Lab Vision™ UltraVision™ LP
Detection System: HRP Polymer/DAB Plus Chromogen and an anti-MMP-9
antibody (ab38898, Abcam, Cambridge, UK). Briefly, the tissue
sections were deparaffinized and rehydrated, and incubated in
Hydrogen Peroxide Block solution for 10 min and then incubated in
Ultra V Block solution for 5 min at room temperature to reduce
non-specific background staining. After the washing step, the
slides were incubated with MMP-9 primary antibody (1:100) for 1 h
at room temperature, and then sequentially applied with Primary
Antibody Enhancer (10 min), HRP Polymer (15 min), and DAB Plus
Chromogen and DAB Plus Substrate mixture (5 min) at room
temperature. Following DAB staining, the sections were
counterstained with Mayer’s Hematoxylin solution (Sigma-Aldrich)
for 3 min at room temperature. Immunohistochemical analysis was
performed under a light microscope (CKX41, Olympus). Mouse serum
was obtained by heart puncture before sacrifice and centrifugation,
and the IL-1β levels were measured from the serum of mice using
Quantikine ELISA kits for IL-1β as described above. The animal
experimental protocol was approved by the Institutional Animal Care
and Use Committee at Gyeongsang National University (approval
number: GLA-120208-M004), and all experiments were performed in
compliance with the institutional guidelines set.
Statistical analysis
All statistical analysis was carried out using
SigmaPlot (version 7.0 for windows, SPSS Inc.). The data are
represented as the means ± standard error of the mean (SEM) of the
results obtained from the number of replicate treatments. Treatment
groups were compared using one-way analysis of variance (ANOVA)
with the Newman-Keuls post-hoc test. A P-value <0.05 was
considered to indicate a statistically significant difference.
Results
RT-R-MDA-MB-231 cells derived from highly
metastatic MDA-MB-231 breast cancer cells exhibit much higher
levels of released ATP, P2Y2R activity and invasiveness
than other RT-R breast cancer cells
First, we observed the amount of released ATP from
various breast cancer cells (MDA-MB-231, MCF7 and T47D cells) and
RT-R breast cancer cells (RT-R-MDA-MB-231, RT-R-MCF7 and RT-R-T47D
cells). As previously reported (24), we confirmed that the amounts of ATP
released into the extra- cellular medium of MDA-MB-231 cells were
higher than those released by MCF7 and T47D cells. Of note, in this
study, we found that the RT-R-MDA-MB-231 cells derived from highly
metastatic MDA-MB-231 breast cancer cells released much higher
levels of ATP than did the MDA-MB-231 and other RT-R breast cancer
cells, and that effect was enhanced by TNF-α (Fig. 1A), an essential factor in tumor
progression and metastasis and released highly in tumor
microenvironment (35,36). To further compare the
P2Y2R activity between the breast cancer cells and RT-R
breast cancer cells, [Ca2+]i was measured in
response to ATP, an agonist of P2Y2R. Although 10
µM ATP evoked a rapid and prompt augmentation in
[Ca2+]i in all the breast cancer cells, the
activities of P2Y2R in the MCF7/RT-R-MCF7 and the
T47D/RT-R-T47D cells were significantly lower than those observed
in the MDA-MB-231 or RT-R-MDA-MB231 cells (Fig. 1B). Moreover, the RT-R-MDA-MB-231
cells also exhibited a higher invasiveness than the other RT-R
breast cancer cells and markedly increased invasiveness following
treatment with ATP (Fig. 1C).
These results suggest that RT-R-MDA-MB-231 cells derived from
highly metastatic MDA-MB-231 breast cancer cells exhibit much
higher levels of released ATP, P2Y2R activity and
invasiveness than other RT-R breast cancer cells derived from
breast cancer cells with low metastatic potential. From these
results, we considered it more suitable to investigate the
association between P2Y2R and the inflammasome in
MDA-MB-231 and RT-R-MDA-MB-231 cells which exhibited high levels of
released ATP, P2Y2R activity and invasiveness, than the
MCF7 and T47D cells, breast cancer cells with low metastatic
potential. Thus, the following experiments were performed with the
MDA-MB-231 and RT-R-MDA-MB-231 cells.
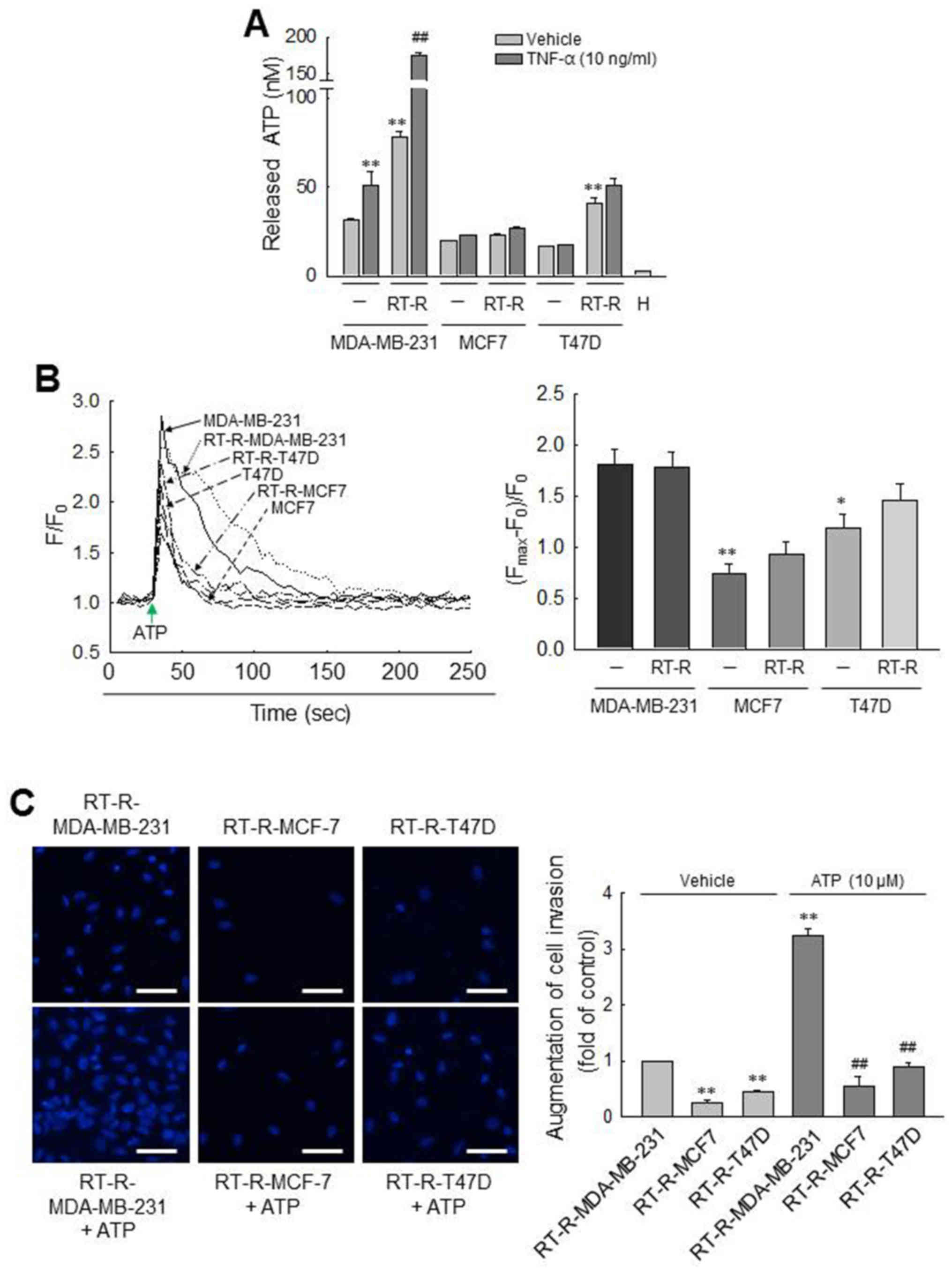 | Figure 1Comparisons of ATP release,
P2Y2R activity and invasiveness between breast cancer
cells and RT-R breast cancer cells. (A) ATP released into the
extracellular medium was measured using the ENLITEN ATP assay
system kit, as described in the Materials and methods. The values
represent the means ± SEM of 3 independent experiments (H, HEPES
buffer only). **P<0.01, compared to the control
(CTRL) of each parent breast cancer cell; ##P<0.01,
compared to the CTRL of each RT-R breast cancer cells. (B)
[Ca2+]i levels were determined in breast
cancer cells and RT-R breast cancer cells to measure
P2Y2R activities. Arrows indicate the points at which
ATP (10 µM) was added. The values represent the means ± SEM
from 3 independent determinations. *P<0.05 and
**P<0.01, compared to the CTRL of MDA-MB-231 cells.
(C) RT-R-breast cancer cells were treated with ATP for 6 h, and
Matrigel invasion assay was performed as described in the Materials
and methods. The values represent the means ± SEM of 3 independent
experiments. **P<0.01, compared to the CTRL of
RT-R-MDA-MB-231 cells; ##P<0.01, compared to
ATP-treated RT-R-MDA-MB-231 cells. Scale bar, 50 µm. ATP,
adenosine triphosphate; RT-R, radiotherapy-resistant. |
MDA-MB-231 cells exhibit an increased
caspase-1 activity and IL-1β secretion induced by TNF-α and ATP
treatment, and these effects are enhanced in RT-R-MDA-MB-231,
through P2Y2R activation
As described above in the Introduction, the
inflammasome, a multiprotein complex, regulates the activation of
caspase-1, which promotes the secretion of the pro-inflammatory
cytokine, IL-1β (37,38), and IL-1β is abundant in tumor
tissue and enhances tumor growth and invasion (10). Therefore, in this study, we
investigated whether TNF-α- or ATP-mediated P2Y2R
activation increases inflammasome activity by examining the
activities of caspase-1 and the levels of IL-1β production in the
MDA-MB-231 and RT-R-MDA-MB-231 cells. As previously mentioned
(35,36), TNF-α is highly released in the
tumor microenvironment and as shown in Fig. 1A, the release of ATP from breast
cancer cells was promoted in response to TNF-α treatment. Thus, we
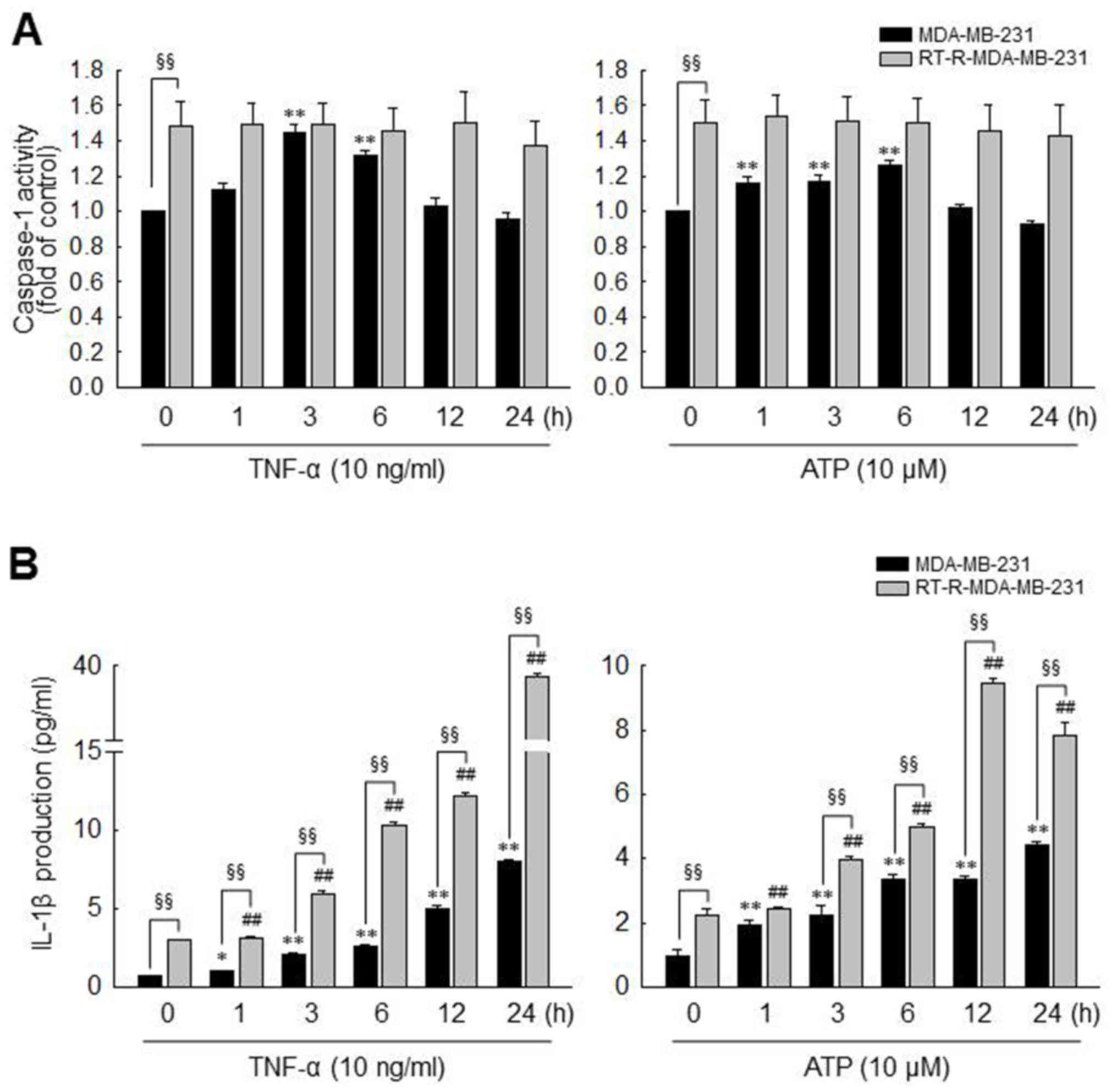
experimented with ATP, as well as TNF-α. As shown in Fig. 2A, caspase-1 activity was
significantly increased and reached a maximum level at 3 or 6 h in
response to TNF-α (10 ng/ml) or ATP (10 µM), respectively,
in the MDA-MB-231 cells. By contrast, caspase-1 activity was not
altered by TNF-α or ATP in the RT-R-MDA-MB-231 cells. However, the
basal activity of caspase-1 in the RT-R-MDA-MB-231 cells was higher
than that observed in the MDA-MB-231 cells, and the maximum
activity level of caspase-1 induced by TNF-α or ATP in the
MDA-MB-231 cells did not exceed the basal activity in the
RT-R-MDA-MB-231 cells. Of note, the IL-1β production levels were
markedly increased following stimulation with TNF-α or ATP in both
types of cells, and the RT-R-MDA-MB-231 cells produced higher
levels of IL-1β than the MDA-MB-231 cells, not only in terms of
basal levels but also following stimulation (Fig. 2B).
Moreover, to clarify whether ATP-activated
P2Y2R is involved in inflammasome activation in the
MDA-MB-231 and RT-R-MDA-MB-231 cells, we examined caspase-1
activity and IL-1β production using P2Y2R siRNA or
apyrase, an enzyme that rapidly hydrolyzes extracellular
nucleotides. First, we confirmed the efficiency of the
P2Y2R siRNA by determining the mRNA levels in the
MDA-MB-231 and RT-R-MDA-MB-231 cells (Fig. 3A). The increased caspase-1 activity
and IL-1β secretion induced by TNF-α or ATP were significantly
reduced by P2Y2R knockdown (Fig. 3B and C) or in the presence of
apyrase (Fig. 3D andE) in both the
MDA-MB-231 and RT-R-MDA-MB-231 cells. Although treatment with TNF-α
or ATP did not alter the activity of caspase-1 in the
RT-R-MDA-MB-231, the knockdown of P2Y2R or the
hydrolyzation of ATP significantly reduced the activity of
caspase-1 in these (Fig. 3B and
D). Furthermore, apyrase treatment alone decreased caspase-1
activity in RT-R-MDA-MB-231 cells, and apyrase also suppressed the
TNF-α-induced increase in caspase-1 activity and IL-1β secretion in
both the MDA-MB-231 and RT-R-MDA-MB-231 cells. These results
suggest that P2Y2R activation by ATP released from
RT-R-MDA-MB-231 cells and MDA-MB-231 cells may regulate
inflammasome activation.
TNF-α and ATP increase the invasive and
colony-forming ability of MDA-MB-31 cells, with enhanced effects in
the RT-R-MDA-MB-231 cells, in a caspase-1-dependent
manner
Subsequently, we investigated whether the
P2Y2R- mediated activation of the inflammasome is linked
to the invasive and colony-forming ability of the MDA-MB-231 and
RT-R-MDA-MB-231 cells. The invasive ability of the MDA-MB-231 and
RT-R-MDA-MB-231 cells was increased by treatment with TNF-α (10
ng/ml) or ATP (10 µM), and this induction was abolished by
inhibiting the activity of the inflammasome with 20 µM
Ac-YVAD-CMK, a selective and irreversible inhibitor of caspase-1
(Fig. 4A). Moreover, the
colony-forming ability of the cells was also promoted following
stimulation with TNF-α or ATP and abolished by the suppression of
caspase-1 activity in both the MDA-MB-231 and RT-R-MDA-MB-231 cells
(Fig. 4B). Of note, the invasive
and colony-forming ability of the RT-R-MDA-MB-231 cells was
significantly enhanced compared to that of the MDA-MB-231 cells
following TNF-α- or ATP treatment and in non-stimulated conditions
(Fig. 4).
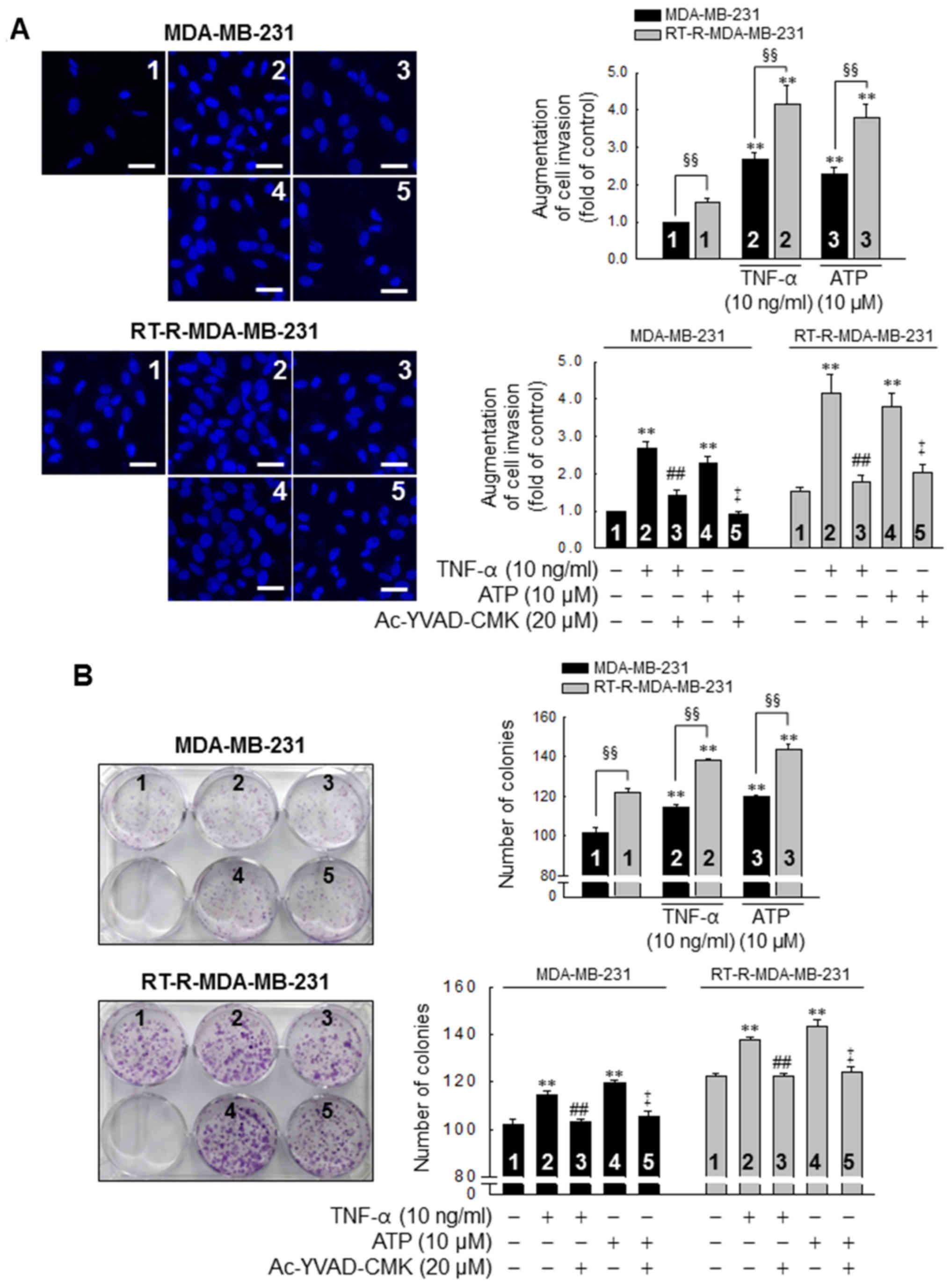 | Figure 4TNF-α and ATP increase the invasive
and colony-forming ability of MDA-MB-231 cells, with an enhanced
effect in the RT-R-MDA-MB-231 cells, in a caspase-1-dependent
manner. (A) Cells were pre-treated with Ac-YVAD-CMK, an
irreversible caspase-1 inhibitor and stimulated with TNF-α or ATP
for 6 h. Matrigel invasion assay was then performed as described in
the Materials and methods. The values represent the means ± SEM of
3 independent experiments. **P<0.01, compared to the
control (CTRL) of each of the cells; ##P<0.01,
compared to the TNF-α treatment of each of the cells;
‡P<0.01, compared to the ATP treatment of each of the
cells; §§P<0.01, significance between the MDA-MB-231
and RT-R-MDA-MB-231 cells. Scale bar, 100 µm. (B) Cells
(1,000 cells/well) were seeded in 6-well plates. The cells were
pre-treated with Ac-YVAD-CMK and stimulated with TNF-α or ATP for 6
h. Following treatment, colony formation assay was performed as
described in the Materials and methods, and quantified by counting
the colonies. The values represent the means ± SEM of 3 independent
experiments. **P<0.01, compared to the CTRL of each
of the cells; ##P<0.01, compared to the TNF-α
treatment of each of the cells; ‡P<0.01, compared to
the ATP treatment of each of the cells; §§P<0.01,
between the MDA-MB-231 and RT-R-MDA-MB-231 cells. ATP, adenosine
triphosphate; RT-R, radiotherapy-resistant; IL-1β, interleukin-1β;
TNF-α, tumor necrosis factor-α. |
MMP-9 activity is modulated by caspase-1
in a P2Y2R-dependent manner, in the MDA-MB-231 or
RT-R-MDA-MB-231 cells
In addition, we examined the effects of the
inflammasome and P2Y2R activation on MMP activity, which
is involved in tumor invasion and metastasis (39). Treatment with TNF-α or ATP
increased MMP-1 activity in the MDA-MB-231 cells, which was
diminished by a caspase-1 inhibitor. These phenomena were more
prominent in the RT-R-MDA-MB-231 cells (Fig. 5A). The prominent induction of MMP-9
activity by ATP in the RT-R-MDA-MB-231 cells was diminished by a
P2Y2R antagonist and P2Y2R siRNA, suggesting
that TNF-α- or ATP-mediated MMP-9 activity is dependent on
caspase-1 and P2Y2R. Even though MMP-9 activity in the
MDA-MB-231 and RT-R-MDA-MB-231 cells exhibited a similar tendency,
the real difference between the MDA-MB-231 and RT-R-MDA-MB-231
cells was that the RT-R-MDA-MB-231 cells exhibited relatively
higher MMP-9 activity than the MDA-MB-231 cells in the control
level and much increased MMP-9 activity following treatment with
TNF-α or ATP. Coherent with this finding, the RT-R-MDA-MB-231 cells
exhibited higher MMP-9 activity than the MDA-MB-231 cells, as shown
in the colony formation assay.
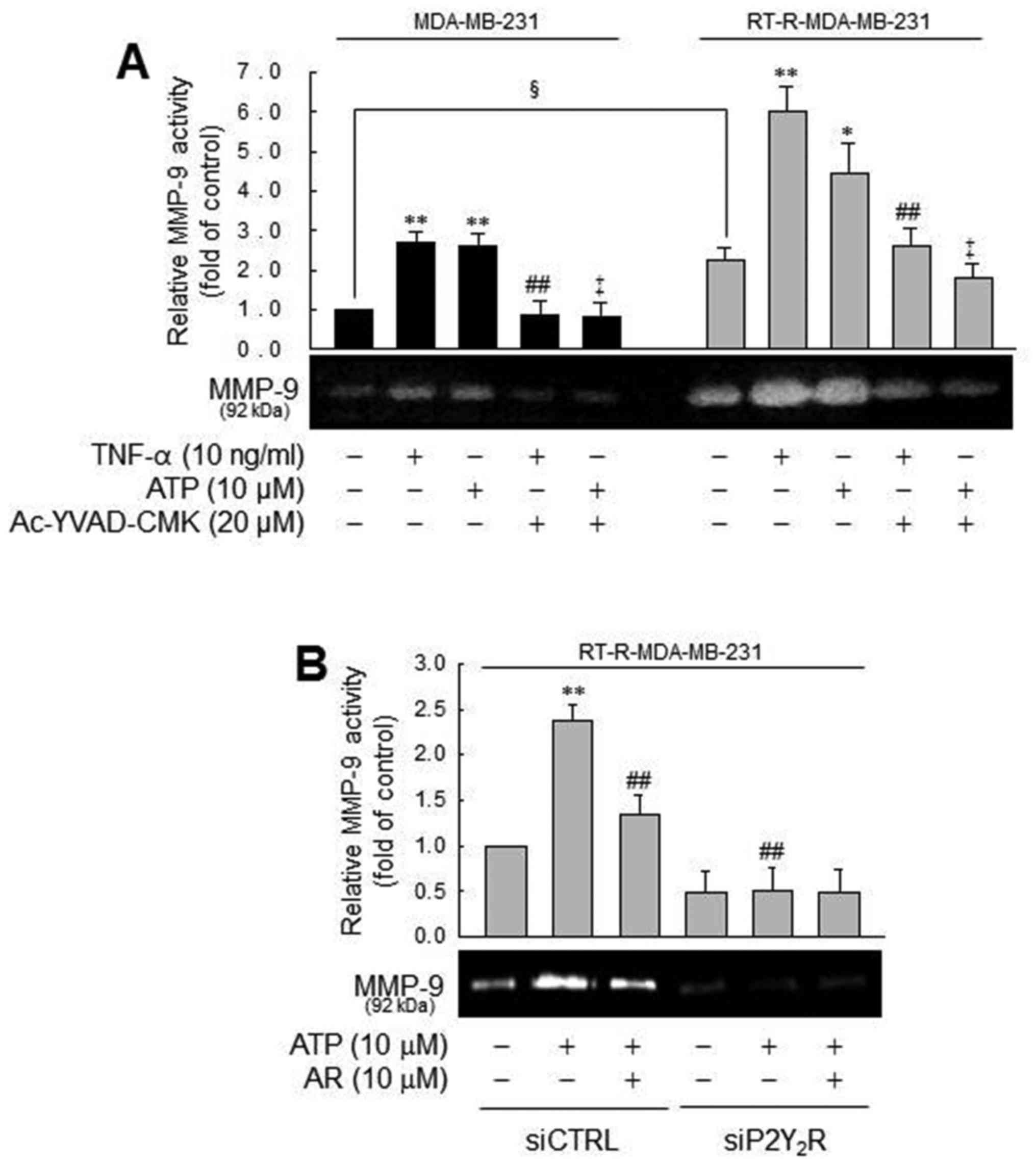 | Figure 5MMP-9 activity is modulated by
caspase-1 in a P2Y2R-dependent manner, in MDA-MB-231 or
RT-R-MDA-MB-231 cells. (A) Cells were pre-treated with Ac-YVAD-CMK
and then stimulated with TNF-α or ATP for 6 h. MMP-9 gelatinase
activity was determined as described in the Materials and methods
and quantified. The values represent the means ± SEM of 3
independent experiments. *P<0.05 and
**P<0.01, compared to the control (CTRL) of each of
the cells; ##P<0.05, compared to the TNF-α treatment
of each of the cells; ‡P<0.05, compared to the ATP
treatment of each of the cells; §P<0.05, comparison
between the MDA-MB-231 and RT-R-MDA-MB-231 cells. (B) siCTRL- or
siP2Y2R-transfected RT-R-MDA-MB-231 cells were
pre-treated with AR-C 118925XX (AR), a specific P2Y2R
antagonist or not. The cells were then stimulated with ATP, and
MMP-9 gelatinase activity was determined as described in the
Materials and methods. The values represent the means ± SEM of 3
independent experiments. **P<0.05, compared to the
CTRL; ##P<0.01, compared to ATP treatment. ATP,
adenosine triphosphate; RT-R, radiotherapy-resistant; IL-1β,
interleukin-1β; TNF-α, tumor necrosis factor-α; MMP-9, matrix
metalloproteinase-9. |
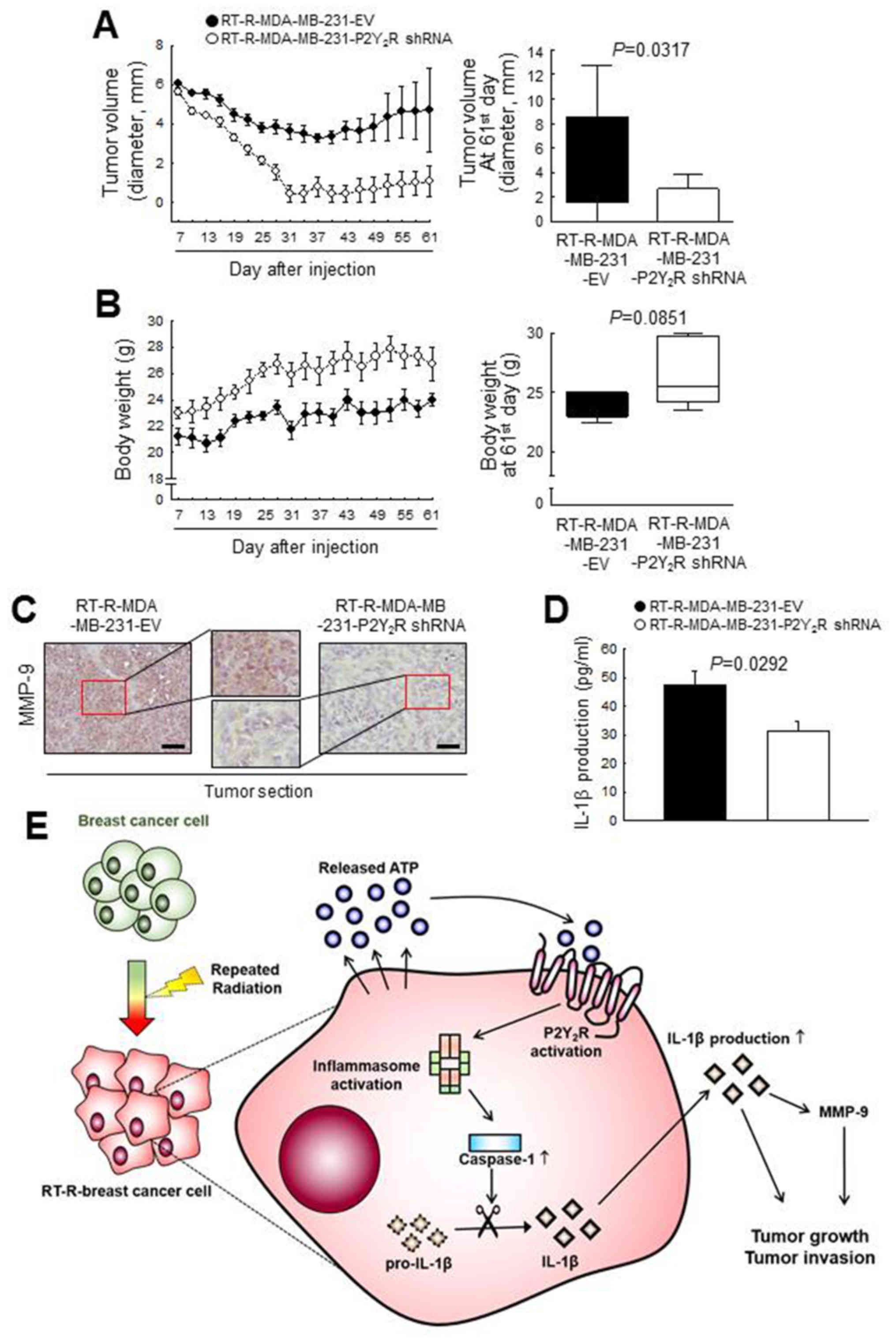
P2Y2R is related to the tumor growth
and invasion of RT-R- breast cancer cells in an in vivo mouse model
by regulating MMP-9 and inflammasome activation Finally, to
confirm the involvement of the P2Y2R in RT-R-MDA-MB-231
cell progression in vivo, athymic nude mice were injected
subcutaneously with control-shRNA-transfected RT-R-MDA-MB-231 cells
(RT-R-MDA-MB-231-EV) or P2Y2R shRNA-transfected
RT-R-MDA-MB-231 cells (RT-R-MDA-MB-231-P2Y2R shRNA). The
tumor sizes and body weights of the mice were measured every 3 days
from 7 days until 60 days after the injection. The tumor volume in
the mice injected with RT-R-MDA-MB-231-P2Y2R shRNA cells
was markedly decreased and the body weight was slightly augmented
compared with that of the RT-R-MDA-MB-231-EV cell- injected mice
(Fig. 6A and B). Furthermore, a
higher MMP-9 expression was detected in the tumor sections from the
mice injected with RT-R-MDA-MB-231-EV cells than in the sections
from mice injected with the RT-R-MDA-MB-231- P2Y2R shRNA
cells (Fig. 6C). In addition, we
observed that the IL-1β levels in the serum were significantly
lower in the mice injected with RT-R-MDA-MB-231-P2Y2R
shRNA cells than in the mice injected with RT-R-MDA-MB-231-EV cells
(Fig. 6D). These results suggest
that P2Y2R plays an important role in RT-R-breast tumor
progression via inflammasome regulation, as shown by the schematic
representation in Fig. 6E.
Discussion
Radiation is an essential component of breast cancer
therapy for the majority of breast cancer patients at all stages of
disease following lumpectomy (40). However, it has been reported that a
small population of breast cancer cells that exhibits resistance
against radiotherapy promotes tumor recurrence and metastasis and
leads to a poor prognosis (41,42).
Our previous study demonstrated that RT-R breast cancer cells were
established by repeated irradiation and exhibited enhanced
invasiveness (43). Moreover, we
have previously reported that breast cancer growth and metastasis
are involved in the activation of P2Y2R by ATP released
from breast cancer cells (24,25).
Therefore, we hypothesized that P2Y2R may also play a
role in the enhanced invasiveness of RT-R breast cancer cells.
Recent studies have reported that inflammasome
signaling is closely associated with several human cancers.
However, the function of the inflammasome in tumor growth and
metastasis remains controversial. The activation of the
inflammasome is beneficial in colitis-associated colorectal cancer,
mostly due to the positive epithelial effects of the IL-18
signaling pathway, the control of cellular proliferation, the
maturation and maintenance of a healthy gut microbiota and cell
death (44,45). In addition, inflammasome-modulated
IL-1 signaling induces the suppression of anticancer immune
responses in natural killer (NK) cells and T cells, which is
disadvantageous for the progression and development of
fibrocarcinoma, gastric cancer, melanoma and breast cancer
(46-49). Thus, in this study, we wished to
determine whether RT-R breast cancer cells release higher levels of
ATP than breast cancer cells, and whether P2Y2R
activation caused by ATP released from RT-R breast cancer cells
enhances invasiveness through inflammasome activation. We
demonstrated that RT-R-MDA-MB-231 cells derived from highly
metastatic MDA-MB-231 breast cancer cells released much higher
levels of ATP, and a exhibited stronger P2Y2R activity
and invasiveness than MDA-MB-231 cells and other RT-R breast cancer
cells. In addition, RT-R-MDA-MB-231 cells exhibited an increased
caspase-1 activity and IL-1β secretion in response to TNF-α and ATP
treatment through P2Y2R activation, resulting in
enhanced invasiveness and colony forming ability.
The extracellular concentration of ATP is known to
be higher in the tumor microenvironment than in normal conditions
(50,51), and extracellular ATP can activate
purinergic receptors, especially P2Y2R (52). In this study, the released ATP
concentration of RT-R breast cancer cells was higher than that of
breast cancer cells; in particular, the RT-R-MDA-MB-231 cells
exhibited the highest levels of ATP released. Although the activity
of P2Y2R induced by the same dose of ATP did not differ
between the MDA-MB-231 and RT-R-MDA-MB-231 cells, the higher ATP
levels released from the RT-R-MDA-MB-231 cells compared to the
MDA-MB-231 cells may mediate P2Y2R activation and the
P2Y2R-mediated signaling cascade. Actually, the
concentration of ATP in the extracellular space and tumor
microenvironment is the net amount of release and degradation.
Thus, the actual ATP concentration released from cancer cells may
be higher than the extracellular concentration of ATP observed in
the tumor microenvironment as previously reported by Jin et
al (24). Moreover, we
previously demonstrated that ATP and UTP increased the cancer cell
proliferation, adhesion molecules expression, and MMP activity at
very low concentration (0.1-1 µM) through the activation of
P2Y2R (24). Based on
these reports, it is suggested that the concentration of ATP in the
tumor microenvironment is sufficient to activate P2Y2R
in breast cancer cells.
Inflammasomes are key signaling platforms that
detect pathogenic microorganisms and sterile stressors. The
sentinel receptor of the inflammasome recognizes these pathogens
and is then activated via assembling apoptosis-associated
speck-like protein containing a carboxy-terminal CARD (ASC) and
caspase-1, which cleaves immature, pro-IL-1β into its mature
secreted form (53). Through these
sequential processes, mature released IL-1β performs diverse roles
in tumor progression (10-14). Of note, in this study, we found
that TNF-α and ATP significantly increased caspase-1 activity in
the MDA-MB-231 cells, but not in the RT-R-MDA-MB-231 cells. The
basal caspase-1 activity in the RT-R-MDA-MB-231 cells was much
higher level than the basal levels, and TNF-α- or ATP-stimulated
levels in MDA-MB-231 cells. This phenomenon may be caused by the
high level of ATP released from RT-R-MDA-MB-231 cells in the basal
state, which is supported by the finding that the basal level of
caspase-1 in RT-R-MDA-MB-231 cells was significantly decreased by
the knockdown of P2Y2R and in the presence of apyrase,
an enzyme that rapidly hydrolyzes extracellular nucleotides. In
addition, we found that the production of IL-1β in response to
stimulation with TNF-α or ATP exhibited a steady increase until 24
h, while caspase-1 activity peaked at 3-6 h following stimulation
and then decreased. The synthesis and secretion of IL-1β are
stimulated by pathogen-associated molecular pattern molecules
(PAMPs) or damage-associated molecular pattern molecules (DAMPs)
and involve several steps (37,54).
IL-1β is first synthesized as inactive pro-IL-1β and then processed
into active IL-1β by caspase-1 and sequentially secreted into the
extracellular space (55).
According to previous reports, extracellular nucleotides also
induce pro-IL-1β production by activating nuclear factor-κB (NF-κB)
or mitogen-activated protein kinase (MAPK) (56-58).
In addition, released mature IL-1β can promote the production of
pro-IL-1β by binding to the IL-1 receptor, which is known to be
expressed in various breast cancer cells including MDA-MB-231 cells
(13). Based on these studies, we
considered that IL-1β production would increase constantly until
late time points after stimulation with extracellular nucleotides
and autocrine mechanism of IL-1β, despite caspase-1 activity was
peaked at a relatively early phase. Moreover, it was determined
that the inflammasome activation induced by TNF-α or ATP, as
determined by caspase-1 activity and the resulting IL-1β
production, was regulated via the activation of P2Y2R,
which was proven by knocking down P2Y2R or hydrolyzing
ATP. This finding suggests that among the purinergic receptors, not
only P2X7R but also P2Y2R, a G
protein-coupled receptor, are involved in the activation of the
inflammasome. Interestingly, some evidence indicated that
extracellular ATP released from cells is finally converted to
adenosine which also activates inflammasome through binding to
adenosine receptors (59,60). Therefore, it is possible that
adenosine, as well as ATP could increase of IL-1β production until
late time through adenosine receptor activation. Accordingly, we
plan to further study the role of adenosine and adenosine receptor
on cancer cell progression and involvement with inflammasome
activation. In addition, several studies have indicated that
microRNAs, particularly, micro-RNA-144 regulates breast cancer cell
proliferation, invasion and migration (61,62).
Moreover, Yu et al (63)
described the role of micro-RNA-144 in the regulation of
radiotherapy sensitivity, migration and invasion of breast cancer
cells. Thus, it is expected that micro-RNA-144 may be sufficient to
affect P2Y2R-mediated inflammasome activation in RT-R
breast cancer cells, even though it has not been examined in this
study.
Breast cancer metastasis is a complex process
determined by a number of factors and pathways. Metastasis begins
with the local invasion of the surrounding host tissue by tumor
cells that are located in the primary tumor and continues until the
tumor cells invade and intravasate into the blood or lymphatic
vessels (64,65). The tumor cells are spread through
the blood stream or lymphatic vessels to distant organs and then
undergo cell cycle arrest and adhere to capillary beds within the
target organ. Consequently, the tumor cells extravasate into the
organ parenchyma and proliferate within the organ (64). Our previous study demonstrated that
RT-R-MDA-MB-231 cells derived from highly metastatic MDA-MB-231
breast cancer cells have more aggressive properties in invasion and
adhesion to endothelial cells due to upregulated the
epithelial-mesenchymal transition (EMT)- or adhesion-involved
proteins (43). In this study, we
determined that RT-R-MDA-MB-231 cells derived from highly
metastatic MDA-MB-231 breast cancer cells exhibited an increased
invasiveness and colony-forming ability compared to MDA-MB-231 or
other breast cancer cells and RT-R breast cancer cells.
Furthermore, the activation of P2Y2R by ATP enhanced the
invasive and colony-forming ability of the RT-R-MDA-MB-231 cells,
which was reduced by an inflammasome inhibitor.
The IL-1β stimulation of tumor cells activates
multiple signaling pathways involving protein kinase B, MAPK and
NF-κB (65). The activation of
these signaling molecules is required for IL-1β-mediated production
of MMP-9, a matrix degrading enzyme that is regarded as a critical
regulator for IL-1β-induced tumor invasion (66-68).
Recently, it was reported that MMP-9 overexpression in the serum is
associated with poor patient prognosis in breast cancer (69). It has also been demonstrated that
HIF-1α response elements are present in the human and mouse IL-1β
promoter (60,70), and this finding led us to
hypothesize that the activation of HIF-1α may be an important step
in increasing pro-IL-1β production. In addition, IL-1β activates
the hypoxia-angiogenesis program by upregulating HIF-1α, the
pivotal mediator of cellular responses to hypoxia (71). HIF-1α expression in cancers is
associated with clinical aggressiveness and poor outcomes (72). HIF-1α rapidly accumulates and
transactivates hundreds of genes under hypoxic conditions,
including angiogenic and growth factors and receptors and
extracellular proteases, such as MMPs (73,74).
In our results, the expression levels of HIF-1α were notably
increased in the RT-R-MDA-MB-231 cells following stimulation with
P2Y2R with ATP, and this effect was markedly decreased
in the cells treated with P2Y2R antagonist or subjected
to P2Y2R knockdown (data not shown).
Finally, we aimed to confirm whether
P2Y2R plays important roles in radiotherapy-resistant
tumor progression, including tumor growth and invasion in an in
vivo animal model; Mice injected with RT-R-MDA-MB-231-EV cells
exhibited a marked increase in tumor size. Of note, the tumors in
both mice injected with RT-R-MDA-MB-231-EV or
RT-R-MDA-MB-231-P2Y2R shRNA cells exhibited a similar
increase in size until the 7th day (data not shown), but seemed to
decline during the early phase. The tumors in the mice then
exhibited growth again and the tumors in the mice injected with
RT-R-MDA-MB-231-P2Y2R shRNA cells grew more rapidly than
those in the mice injected with RT-R-MDA-MB-231-EV cells. In this
study, we used athymic nude mice (absent of T cell function, high
functional activity of macrophages and natural killer cells) which
are acceptable for use in xenograft and allograft transplantation
experiments. Thus, we hypothesized that NK cells may be functional
to the xenograft tumor cells during in the early phase when tumor
cells could not compose the tumor microenvironment yet. In
addition, in tumor tissue sections from mice injected with
RT-R-MDA-MB-231- P2Y2R shRNA cells, we observed higher
levels of MMP-9 compared with the levels observed in the mice
injected with RT-R-MDA-MB-231-EV cells. The concentration of IL-1β
was slightly reduced in serum from mice injected with
RT-R-MDA-MB-231-P2Y2R shRNA cells. Thus, we hypothesized
that P2Y2R is related to the tumor growth and invasion
of RT-R breast cancer cells in vivo, and the regulation of
this purinergic receptor in RT-R-tumor cells may be helpful for
controlling tumor progression in patients.
In conclusion, in this study, we demonstrate that
RT-R breast cancer cells, particularly RT-R-MDA-MB-231 cells,
release higher levels of ATP than do breast cancer cells, and
extracellular ATP promotes invasion and tumor growth through the
activation of P2Y2R. Moreover, inflammasome activation
is more prominent in RT-R breast cancer cells and is
P2Y2R dependent, ultimately resulting in increased tumor
invasion and progression. Our results suggest the involvement of
the P2Y2 purinergic receptor in inflammasome activation
in breast cancer cells and RT-R breast cancer cells for the first
time, at least to the best of our knowledge and highlight the
importance of controlling P2Y2R activity to achieve a
good prognosis in patients with RT-R tumors (Fig. 6E).
Funding
This study was supported by Basic Science Research
Program through the National Research Foundation of Korea (NRF)
funded by the Ministry of Education, Science and Technology
(NRF-2015R1A1A3A04001029) and by the Ministry of Science, ICT and
Future Planning (NRF-2015R1A5A2008833).
Availability of data and materials
All the data generated and analyzed during the
study are available from the corresponding author on reasonable
request.
Author’s contributions
HJ performed the experiments and wrote the
manuscript. YSK performed data analysis. HJK conceived the
hypothesis, directed the project and wrote the manuscript. All
authors have read and approved the final manuscript.
Ethics approval and consent to
participate
The animal experimental protocol was approved by
the Institutional Animal Care and Use Committee at Gyeongsang
National University (approval number: GLA-120208-M004), and all
experiments were performed in compliance with the institutional
guidelines set.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Abbreviations:
|
ALR
|
absent in melanoma 2-like
receptors
|
|
ASC
|
apoptosis-associated speck-like
protein containing a carboxy-terminal CARD
|
|
ATP
|
adenosine triphosphate
|
|
EMT
|
epithelial-mesenchymal transition
|
|
IL-1β
|
interleukin-1β
|
|
MAPK
|
mitogen-activated protein kinase
|
|
MMP-9
|
matrix metalloproteinase-9
|
|
NF-κB
|
nuclear factor-κB
|
|
NLRs
|
nucleotide-binding domain-like
receptors
|
|
P2X7R
|
P2X purinergic receptor 7
|
|
P2Y2R
|
P2Y purinergic receptor 2
|
|
RT-R
|
radiotherapy-resistant
|
|
TNF-α
|
tumor necrosis factor-α
|
References
|
1
|
Jinushi M: The role of innate immune
signals in antitumor immunity. OncoImmunology. 1:189–194. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Grivennikov SI, Greten FR and Karin M:
Immunity, inflammation, and cancer. Cell. 140:883–899. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Baniyash M, Sade-Feldman M and Kanterman
J: Chronic inflammation and cancer: Suppressing the suppressors.
Cancer Immunol Immunother. 63:11–20. 2014. View Article : Google Scholar
|
|
4
|
Zitvogel L, Kepp O, Galluzzi L and Kroemer
G: Inflammasomes in carcinogenesis and anticancer immune responses.
Nat Immunol. 13:343–351. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Coussens LM and Werb Z: Inflammation and
cancer. Nature. 420:860–867. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Clark AG and Vignjevic DM: Modes of cancer
cell invasion and the role of the microenvironment. Curr Opin Cell
Biol. 36:13–22. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dinarello CA: Immunological and
inflammatory functions of the interleukin-1 family. Annu Rev
Immunol. 27:519–550. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Novick D, Kim S, Kaplanski G and Dinarello
CA: Interleukin-18, more than a Th1 cytokine. Semin Immunol.
25:439–448. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lamkanfi M: Emerging inflammasome effector
mechanisms. Nat Rev Immunol. 11:213–220. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Apte RN, Dotan S, Elkabets M, White MR,
Reich E, Carmi Y, Song X, Dvozkin T, Krelin Y and Voronov E: The
involvement of IL-1 in tumorigenesis, tumor invasiveness,
metastasis and tumor- host interactions. Cancer Metastasis Rev.
25:387–408. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Apte RN, Krelin Y, Song X, Dotan S, Recih
E, Elkabets M, Carmi Y, Dvorkin T, White RM, Gayvoronsky L, et al:
Effects of micro-environment- and malignant cell-derived
interleukin-1 in carcinogenesis, tumour invasiveness and
tumour-host interactions. Eur J Cancer. 42:751–759. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jin L, Yuan RQ, Fuchs A, Yao Y, Joseph A,
Schwall R, Schnitt SJ, Guida A, Hastings HM, Andres J, et al:
Expression of interleukin- 1beta in human breast carcinoma. Cancer.
80:421–434. 1997. View Article : Google Scholar
|
|
13
|
Pantschenko AG, Pushkar I, Anderson KH,
Wang Y, Miller LJ, Kurtzman SH, Barrows G and Kreutzer DL: The
interleukin-1 family of cytokines and receptors in human breast
cancer: Implications for tumor progression. Int J Oncol.
23:269–284. 2003.PubMed/NCBI
|
|
14
|
Perrier S, Caldefie-Chézet F and Vasson
MP: IL-1 family in breast cancer: Potential interplay with leptin
and other adipocytokines. FEBS Lett. 583:259–265. 2009. View Article : Google Scholar
|
|
15
|
Iyer SS, Pulskens WP, Sadler JJ, Butter
LM, Teske GJ, Ulland TK, Eisenbarth SC, Florquin S, Flavell RA,
Leemans JC, et al: Necrotic cells trigger a sterile inflammatory
response through the Nlrp3 inflammasome. Proc Natl Acad Sci USA.
106:20388–20393. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bergfeld GR and Forrester T: Release of
ATP from human erythrocytes in response to a brief period of
hypoxia and hypercapnia. Cardiovasc Res. 26:40–47. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bodin P and Burnstock G: Purinergic
signalling: ATP release. Neurochem Res. 26:959–969. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Vénéreau E, Ceriotti C and Bianchi ME:
DAMPs from cell death to new life. Front Immuno l. 6:4222015.
|
|
19
|
Pellegatti P, Raffaghello L, Bianchi G,
Piccardi F, Pistoia V and Di Virgilio F: Increased level of
extracellular ATP at tumor sites: In vivo imaging with plasma
membrane luciferase. PLoS One. 3:e25992008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Schafer R, Sedehizade F, Welte T and
Reiser G: ATP- and UTP-activated P2Y receptors differently regulate
proliferation of human lung epithelial tumor cells. Am J Physiol
Lung Cell Mol Physiol. 285:L376–L385. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shabbir M, Ryten M, Thompson C,
Mikhailidis D and Burnstock G: Purinergic receptor-mediated effects
of ATP in high-grade bladder cancer. BJU Int. 101:106–112.
2008.
|
|
22
|
Janssens R and Boeynaems JM: Effects of
extracellular nucleotides and nucleosides on prostate carcinoma
cells. Br J Pharmacol. 132:536–546. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
White N, Butler PE and Burnstock G: Human
melanomas express functional P2 X(7) receptors. Cell Tissue Res.
321:411–418. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jin H, Eun SY, Lee JS, Park SW, Lee JH,
Chang KC and Kim HJ: P2Y2 receptor activation by
nucleotides released from highly metastatic breast cancer cells
increases tumor growth and invasion via crosstalk with endothelial
cells. Breast Cancer Res. 16:R772014. View Article : Google Scholar
|
|
25
|
Joo YN, Jin H, Eun SY, Park SW, Chang KC
and Kim HJ: P2Y2R activation by nucleotides released
from the highly metastatic breast cancer cell MDA-MB-231
contributes to pre- metastatic niche formation by mediating lysyl
oxidase secretion, collagen crosslinking, and monocyte recruitment.
Oncotarget. 5:9322–9334. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Clarke M, Collins R, Darby S, Davies C,
Elphinstone P, Evans V, Godwin J, Gray R, Hicks C, James S, et al:
Early Breast Cancer Trialists’ Collaborative Group (EBCTCG):
Effects of radiotherapy and of differences in the extent of surgery
for early breast cancer on local recurrence and 15-year survival:
An overview of the randomised trials. Lancet. 366:2087–2106. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Veronesi U, Boyle P, Goldhirsch A,
Orecchia R and Viale G: Breast cancer. Lancet. 365:1727–1741. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gebski V, Lagleva M, Keech A, Simes J and
Langlands AO: Survival effects of postmastectomy adjuvant radiation
therapy using biologically equivalent doses: A clinical
perspective. J Natl Cancer Inst. 98:26–38. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ringborg U, Bergqvist D, Brorsson B,
Cavallin-Ståhl E, Ceberg J, Einhorn N, Frödin JE, Järhult J,
Lamnevik G, Lindholm C, et al: The Swedish Council on Technology
Assessment in Health Care (SBU) systematic overview of radiotherapy
for cancer including a prospective survey of radiotherapy practice
in Sweden 2001 - summary and conclusions. Acta Oncol. 42:357–365.
2003. View Article : Google Scholar
|
|
30
|
Delaney G, Jacob S, Featherstone C and
Barton M: The role of radiotherapy in cancer treatment: Estimating
optimal utilization from a review of evidence-based clinical
guidelines. Cancer. 104:1129–1137. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Fukuda K, Sakakura C, Miyagawa K, Kuriu Y,
Kin S, Nakase Y, Hagiwara A, Mitsufuji S, Okazaki Y, Hayashizaki Y,
et al: Differential gene expression profiles of radioresistant
oesophageal cancer cell lines established by continuous
fractionated irradiation. Br J Cancer. 91:1543–1550. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Henness S, Davey MW, Harvie RM, Banyer J,
Wasinger V, Corthals G and Davey RA: Changes in gene expression
associated with stable drug and radiation resistance in small cell
lung cancer cells are similar to those caused by a single X-ray
dose. Radiat Res. 161:495–503. 2004. View
Article : Google Scholar
|
|
33
|
Thomas ED, Clift RA, Hersman J, Sanders
JE, Stewart P, Buckner CD, Fefer A, McGuffin R, Smith JW and Storb
R: Marrow transplantation for acute nonlymphoblastic leukemic in
first remission using fractionated or single-dose irradiation. Int
J Radiat Oncol Biol Phys. 8:817–821. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Jin H, Ham SA, Kim MY, Woo IS, Kang ES,
Hwang JS, Lee KW, Kim HJ, Roh GS, Lim DS, et al: Activation of
peroxisome proliferator-activated receptor-δ attenuates
glutamate-induced neurotoxicity in HT22 mouse hippocampal cells. J
Neurosci Res. 90:1646–1653. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Suganuma M, Okabe S, Marino MW, Sakai A,
Sueoka E and Fujiki H: Essential role of tumor necrosis factor
alpha (TNF-alpha) in tumor promotion as revealed by TNF-alpha-
deficient mice. Cancer Res. 59:4516–4518. 1999.PubMed/NCBI
|
|
36
|
Egberts JH, Cloosters V, Noack A,
Schniewind B, Thon L, Klose S, Kettler B, von Forstner C, Kneitz C,
Tepel J, et al: Anti-tumor necrosis factor therapy inhibits
pancreatic tumor growth and metastasis. Cancer Res. 68:1443–1450.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Schroder K and Tschopp J: The
inflammasomes. Cell. 140:821–832. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Fink SL and Cookson BT: Apoptosis,
pyroptosis, and necrosis: Mechanistic description of dead and dying
eukaryotic cells. Infect Immun. 73:1907–1916. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Radisky ES and Radisky DC: Matrix
metalloproteinase-induced epithelial-mesenchymal transition in
breast cancer. J Mammary Gland Biol Neoplasia. 15:201–212. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Carlson RW and McCormick B: Update: NCCN
breast cancer clinical practice guidelines. J Natl Compr Canc Netw.
3(Suppl 1): S7–S11. 2005.PubMed/NCBI
|
|
41
|
Lee SY, Jeong EK, Ju MK, Jeon HM, Kim MY,
Kim CH, Park HG, Han SI and Kang HS: Induction of metastasis,
cancer stem cell phenotype, and oncogenic metabolism in cancer
cells by ionizing radiation. Mol Cancer. 16:102017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
von Essen CF: Radiation enhancement of
metastasis: A review. Clin Exp Metastasis. 9:77–104. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ko YS, Jin H, Joo Y, Lee JS, Park SW,
Chang KC, Kang KM, Jeong BK and Kim HJ: Radio-resistant breast
cancer cells derived from highly metastatic breast cancer cells
exhibit increased resistance to chemotherapy, enhanced invasive
properties and premetastatic niche formation due to cancer stem
cells. Oncol Rep. In press.
|
|
44
|
Allen IC, TeKippe EM, Woodford RM, Uronis
JM, Holl EK, Rogers AB, Herfarth HH, Jobin C and Ting JP: The NLRP3
inflammasome functions as a negative regulator of tumorigenesis
during colitis-associated cancer. J Exp Med. 207:1045–1056. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Dupaul-Chicoine J, Yeretssian G, Doiron K,
Bergstrom KS, McIntire CR, LeBlanc PM, Meunier C, Turbide C, Gros
P, Beauchemin N, et al: Control of intestinal homeostasis, colitis,
and colitis-associated colorectal cancer by the inflammatory
caspases. Immunity. 32:367–378. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Chow MT, Sceneay J, Paget C, Wong CS,
Duret H, Tschopp J, Möller A and Smyth MJ: NLRP3 suppresses NK
cell-mediated responses to carcinogen-induced tumors and
metastases. Cancer Res. 72:5721–5732. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Bhagat Tu, Cui GS, Takaishi G, Kurt-Jones
S, Rickman EA, Betz B, Penz-Oesterreicher KS, Bjorkdahl M, Fox OJG,
et al: Overexpression of interleukin-1beta induces gastric
inflammation and cancer and mobilizes myeloid-derived suppressor
cells in mice. Cancer Cell. 14:408–419. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Liu W, Luo Y, Dunn JH, Norris DA,
Dinarello CA and Fujita M: Dual role of apoptosis-associated
speck-like protein containing a CARD (ASC) in tumorigenesis of
human melanoma. J Invest Dermatol. 133:518–527. 2013. View Article : Google Scholar
|
|
49
|
Kolb R, Phan L, Borcherding N, Liu Y, Yuan
F, Janowski AM, Xie Q, Markan KR, Li W, Potthoff MJ, et al:
Obesity-associated NLRC4 inflammasome activation drives breast
cancer progression. Nat Commun. 7:130072016. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Corriden R and Insel PA: Basal release of
ATP: An autocrine- paracrine mechanism for cell regulation. Sci
Signal. 3. pp. re12010, View Article : Google Scholar
|
|
51
|
Lazarowski E: Regulated release of
nucleotides and UDP sugars from astrocytoma cells. Novartis Found
Symp. 276:73–84; discussion 84–90, 107–112, 275–281.
2006.PubMed/NCBI
|
|
52
|
Jacobson KA, Ivanov AA, de Castro S,
Harden TK and Ko H: Development of selective agonists and
antagonists of P2Y receptors. Purinergic Signal. 5:75–89. 2009.
View Article : Google Scholar :
|
|
53
|
Martinon F, Burns K and Tschopp J: The
inflammasome: A molecular platform triggering activation of
inflammatory caspases and processing of proIL-beta. Mol Cell.
10:417–426. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Takeuchi O and Akira S: Pattern
recognition receptors and inflammation. Cell. 140:805–820. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Brough D and Rothwell NJ:
Caspase-1-dependent processing of pro-interleukin-1beta is
cytosolic and precedes cell death. J Cell Sci. 120:772–781. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Korcok J, Raimundo LN, Ke HZ, Sims SM and
Dixon SJ: Extracellular nucleotides act through P2X7 receptors to
activate NF-kappaB in osteoclasts. J Bone Miner Res. 19:642–651.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Ferrari D, Wesselborg S, Bauer MK and
Schulze-Osthoff K: Extracellular ATP activates transcription factor
NF-kappaB through the P2Z purinoreceptor by selectively targeting
NF-kappaB p65. J Cell Biol. 139:1635–1643. 1997. View Article : Google Scholar
|
|
58
|
Wilden PA, Agazie YM, Kaufman R and
Halenda SP: ATP-stimulated smooth muscle cell proliferation
requires independent ERK and PI3K signaling pathways. Am J Physiol.
275:H1209–H1215. 1998.PubMed/NCBI
|
|
59
|
Baron L, Gombault A, Fanny M, Villeret B,
Savigny F, Guillou N, Panek C, Le Bert M, Lagente V, Rassendren F,
et al: The NLRP3 inflammasome is activated by nanoparticles through
ATP, ADP and adenosine. Cell Death Dis. 6:e16292015. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Ouyang X, Ghani A, Malik A, Wilder T,
Colegio OR, Flavell RA, Cronstein BN and Mehal WZ: Adenosine is
required for sustained inflammasome activation via the
A2A receptor and the HIF-1α pathway. Nat Commun.
4:29092013. View Article : Google Scholar
|
|
61
|
Yin Y, Cai J, Meng F, Sui C and Jiang Y:
MiR-144 suppresses proliferation, invasion, and migration of breast
cancer cells through inhibiting CEP55. Cancer Biol Ther.
19:306–315. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Pan Y, Zhang J, Fu H and Shen L: miR-144
functions as a tumor suppressor in breast cancer through inhibiting
ZEB1/2-mediated epithelial mesenchymal transition process.
OncoTargets Ther. 9:6247–6255. 2016. View Article : Google Scholar
|
|
63
|
Yu L, Yang Y, Hou J, Zhai C, Song Y, Zhang
Z, Qiu L and Jia X: MicroRNA-144 affects radiotherapy sensitivity
by promoting proliferation, migration and invasion of breast cancer
cells. Oncol Rep. 34:1845–1852. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Hunter KW, Crawford NP and Alsarraj J:
Mechanisms of metastasis. Breast Cancer Res. 10(Suppl 1): S22008.
View Article : Google Scholar :
|
|
65
|
Talmadge JE and Fidler IJ: AACR centennial
series: the biology of cancer metastasis: historical perspective.
Cancer Res. 70:5649–5669. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Yokoo T and Kitamura M: Dual regulation of
IL-1 beta-mediated matrix metalloproteinase-9 expression in
mesangial cells by NF-kappa B and AP-1. Am J Physiol.
270:F123–F130. 1996.PubMed/NCBI
|
|
67
|
Ruhul Amin AR, Senga T, Oo ML, Thant AA
and Hamaguchi M: Secretion of matrix metalloproteinase-9 by the
proinflammatory cytokine, IL-1beta: A role for the dual signalling
pathways, Akt and Erk. Genes Cells. 8:515–523. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Bauvois B: New facets of matrix
metalloproteinases MMP-2 and MMP-9 as cell surface transducers:
Outside-in signaling and relationship to tumor progression. Biochim
Biophys Acta. 1825.29–36. 2012.
|
|
69
|
Ren F, Tang R, Zhang X, Madushi WM, Luo D,
Dang Y, Li Z, Wei K and Chen G: Overexpression of MMP family
members functions as prognostic biomarker for breast cancer
patients: A systematic review and meta-analysis. PLoS One.
10:e01355442015. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Zhang W, Petrovic JM, Callaghan D, Jones
A, Cui H, Howlett C and Stanimirovic D: Evidence that
hypoxia-inducible factor-1 (HIF-1) mediates transcriptional
activation of interleukin-1beta (IL-1beta) in astrocyte cultures. J
Neuroimmunol. 174:63–73. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Jung YJ, Isaacs JS, Lee S, Trepel J and
Neckers L: IL-1beta- mediated up-regulation of HIF-1alpha via an
NFkappaB/COX-2 pathway identifies HIF-1 as a critical link between
inflammation and oncogenesis. FASEB J. 17:2115–2117. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Semenza GL: Molecular mechanisms mediating
metastasis of hypoxic breast cancer cells. Trends Mol Med.
18:534–543. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Semenza GL: Surviving ischemia: Adaptive
responses mediated by hypoxia-inducible factor 1. J Clin Invest.
106:809–812. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Semenza GL: HIF-1 and tumor progression:
pathophysiology and therapeutics. Trends Mol Med. 8(Suppl 4):
S62–S67. 2002. View Article : Google Scholar : PubMed/NCBI
|