Introduction
Despite improvements in patient care over the past
years, breast cancer remains a leading cause of mortality among
women worldwide, accounting for >500 000 deaths in 2012
(1). Breast tumors are most often
angiogenic, i.e., they induce the development of neo-vessels to
their profit. Angiogenesis is associated with lymph node metastasis
and a poor prognosis for breast cancer patients (2). Anti-angiogenic agents have thus
emerged as promising therapies (3). Nevertheless, such treatments have
thus far failed to significantly improve the overall survival of
breast cancer patients (4). A
better knowledge of the mechanisms underpinning the complex
interactions between cancers and vessels may aid in resolving this
issue and in improving the therapeutic strategies.
Sprouting angiogenesis occurs through a branching
morphogenesis process and requires endothelial cell proliferation,
migration and maturation to form new capillaries. The latter step
of differentiation is often lacking during tumoral angiogenesis,
which leads to more permeant vessels that are hardly effective, but
may facilitate the access of tumor cells to the systemic
circulation (5). A better
understanding of the mechanisms that induce cancer cells to trigger
the angiogenic response and modulate the vessel features should
provide crucial information with which to target this critical
phenomenon in cancer progression.
Ets-1 proto-oncogene overexpression in breast
cancers is associated with a poor prognosis (6). Ets-1 is a critical regulator of
invasion induced by dialogues between the epithelium and mesenchyme
(7). We previously demonstrated
that Ets-1 overexpression stimulated breast cancer cell scattering
through the coordination of several classes of invasion-related
molecules (8). Among these, Ets-1
regulates several soluble factors potentially involved in
angiogenesis, such as growth factors or extracellular matrix
proteases, which prompted us to determine whether Ets-1
overexpression in breast cancer cells can trigger a paracrine
angiogenic environment.
To address this issue, we manipulated Ets-1 activity
in cancer cells, either by overexpressing Ets-1 or by expressing a
dominant negative mutant, as previously described (8,9). In
this study, we investigated the effects of such manipulations upon
the interactions between cancer and endothelial cells. We performed
direct heterotypic co-cultures of breast cancer cells with normal
endothelial cells, or administered conditioned media collected from
one cell type on the other. We used three-dimensional matrix
models, which recreate a physiological environment (10), which allowed us to mimic several
physiological aspects of capillary formation. Finally, we examined
the effects of Ets-1 on the angiogenic pattern of experimental
tumors in vivo. This allowed us to highlight an interesting
and complex role of Ets-1 expression by breast cancer cells in
controlling the angiogenic induction.
Materials and methods
Cell culture
Mouse mammary tumor epithelial cells (MMT CCL-51
cells; ATCC, Manassas, VA, USA) were routinely cultured in DMEM
(BioWhittaker; Lonza Walkersville, Inc., Walkersville, MD, USA)
supplemented with penicillin, streptomycin, non-essential amino
acids, L-glutamine and 10% fetal calf serum (Gibco/Thermo Fisher
Scientific, Waltham, MA, USA). They were infected in parallel by
supernatants from virus-producing GP+E86 cells (kind gift from Dr
Arthur Bank, Columbia University, New York, NY, USA) transfected
with MFG retroviral constructs to overexpress Ets-1 or a dominant
negative mutant composed of the DNA binding domain of Ets-1, as
previously described (8,9). Briefly, the cDNA sequence encoding
either the full sequence or the DNA-binding domain of murine Ets1
was cloned upstream of the internal ribosome entry site (IRES) and
the neomycin-resistance gene then inserted into a retroviral MFG
vector in replacement of the Gag, Pol and Env genes, as previously
described in detail (11).
Murine endothelial mouse spleen stroma (MSS-31)
cells were obtained from Dr N. Yanai (12). They were grown in α-MEM
(BioWhittaker; Lonza Walkersville, Inc.) supplemented with
penicillin-streptomycin, glutamine and 10% fetal calf serum.
All cell lines used were checked with the list of
known misidentified cell lines available from the International
Cell Line Authentication Committee and ExPASy Cellosaurus, and they
are not misidentified nor contaminated.
Co-culture models
For two-dimensional and three-dimensional matrix
models, we used a matrix substrate composed of type I collagen and
Matrigel™, as previously described (8,9).
Conditioned media were prepared by respectively seeding
3×106 MMT or 1×106 MSS-31 cells suspended in
3 ml appropriate medium upon 100 mm-diameter Petri dishes coated
with this matrix. Twenty-four hours later, the cell culture
supernatants were harvested, filtered through Millex 0.22 µm
filters, and stored at −80°C in Sorenson™ pre-lubricated tubes
prior to their use.
For two-dimensional MSS-31 morphogenesis assays, 150
µl of matrix was poured in 48-well plates and incubated for
30 min at 37°C. The MSS-31 cells were then seeded at a density of
20,000 cells per well, in 5% FCS α-MEM. An equal volume of
conditioned medium from the MMT cells, or DMEM as a control, was
then added. The media were refreshed every other day. At the end of
the experiment, cellular structures were stained with neutral red
(0.1% in PBS) then fixed with 4% paraformaldehyde in PBS. Images
were acquired using a Leica DMIRB inverted microscope (Leica
Microsystems GmbH, Wetzlar, Germany).
For three-dimensional MSS-31 morphogenesis assays, a
first acellular layer of matrix was deposited in 48-well plates.
The MSS-31 cells were then resuspended in that matrix solution and
seeded at a density of 25,000 cells per well. Serum-free
conditioned media from the MMT cells or DMEM were added and
refreshed every 3 days. Images were acquired under a phase contrast
light or epifluorescence using a Leica DMIRB inverted microscope
(Leica Microsystems GmbH)
For direct co-culture assays, the MMT and MSS-31
cells were respectively labeled by diI and diO Vybrant™
Cell-Labeling solutions (V-22885 and V-22886; Molecular
Probes/Thermo Fisher Scientific), according to the manufacturer’s
instructions.
For three-dimensional co-cultures, 4,000 MMT cells
suspended in 300 µl of matrix were seeded in 24-well plates
above a first acellular layer matrix. A suspension of 2,000 MSS-31
cells was then added upon these gels.
Image analysis
Primary branch thickness, fluorescence intensity and
capillary lumen areas were quantified with ImageJ measurement tools
(National Institutes of Health, Bethesda, Maryland, USA, https://imagej.nih.gov/ij/).
Cell proliferation
The MSS-31 cells were seeded in 96-well plates at a
density of 500 cells per well, and 30% of their culture medium was
replaced by conditioned media from MMT cells, or complete DMEM as a
control. Reciprocally, the MMT cells were seeded at a density of
2,000 cells per well in 12-well plates, and 30% of their culture
medium was replaced by conditioned media from MSS-31 cells or
complete α-MEM as a control. The media were refreshed every day.
The cell number was assessed by MTT assay (M-2128; Sigma-Aldrich,
St. Louis, MO, USA). Briefly, the cells were incubated at 37°C for
2 h in presence of 1 mM MTT, and the purple formazan crystals were
then dissolved in a solution of 5 mM HCl in isopropanol. The plates
were shaken for 5 min and the optical density was then measured at
a wavelength of 570 nm with an MRX Revelation plate reader (Dynex
Technologies GmBH, Denkendorf, Germany).
Wound assays
The MMT cells were seeded in 60 mm diameter Petri
dishes and grown until sub-confluence. A wound was created on the
cell layer using a scraper. The dishes were rinsed 3 times with
fresh medium, and equal volumes of DMEM and conditioned medium from
the MSS-31 cells, or 5% FCS α-MEM as a control, were then added.
Cell migration in the denuded area was recorded 24 h later. Images
were acquired using a Leica DMIRB inverted microscope (Leica
Microsystems GmbH).
Transwell assays
The MMT cells were seeded at a density of 10,000
cells per well in 24-well Falcon™Companion plates (Thermo Fisher
Scientific). Twenty hours later, the medium culture was refreshed
and 15,000 MSS-31 cells per well were seeded under serum-free
conditions upon Transwell tissue culture inserts with 8.0 µm
pore size (Falcon™), at a density of 25,000 cells per well.
Alternately, MSS-31 cells were seeded at a density of 20,000 cells
per well in 24-well Companion plates (Falcon™). Twenty hours later,
the medium was replaced and MMT cells were seeded in Transwell
tissue culture filters with 8.0 µm pore size (Falcon™), at a
density of 25,000 cells per well. The cells were allowed to migrate
for 30 h, then non-migratory cells were removed from the top of the
filter by swiping with humidified cotton swabs. Cells that had
migrated through the filter pores to the lower face of the inserts
were fixed in 4% paraformaldehyde in PBS, their nuclei were stained
with Hoechst at 5 mg/l in PBS, and counted. Values from the control
conditions were set as the reference value of 1. Data are expressed
as means of 3 independent experiments.
Cell adhesion
The MMT cells were fluorescently labeled as
described above in ‘Co-culture models’ section, and seeded in
12-well plates at a density of 50,000 cells per well, upon MSS-31
cell confluent monolayers. Following 45 min of incubation at 37°C,
wells were thoroughly rinsed 3 times by fresh medium, and the
plates were further incubated for 4 additional hours at 37°C so
that the cells could complete their adhesion. The cells were then
fixed with paraformaldehyde and observed under epifluorescence to
count the fluorescent MMT cells, using a Leica DMIRB inverted
microscope (Leica Microsystems GmbH). Values from the control
conditions were set as the reference value of 1. The results are
means of 3 independent experiments.
Zymography
Conditioned media or membrane-enriched fractions
were analyzed for MMP-2 and MMP-9 gelatinolytic activities.
Briefly, SDS-PAGE was performed under non-reducing conditions in
7.5% polycrylamide gels containing 0.1% gelatin. The gels were then
rinsed for 30 min in 10 mM Tris-HCl/2.5% Triton X-100 (pH 8.0), and
further incubated at 37°C for 20 h in 50 mM Tris-HCl, 0.5 mM
CaCl2, 1 µM ZnCl2 (pH 8.0) to restore
gelatinases activity. Gels were stained in 0.25% Coomassie blue
R250 and destained in 7% acetic acid plus 25% methanol. Gelatin
degradation is detected by the appearance of translucent bands in
the dark blue-stained gelatin background.
Western blot analysis
The cells were subjected to hypotonic and thermic
lysis as previously described (8).
Protein amounts were measured according to the Bradford method
(13). Proteins from
membrane-enriched fractions or cell conditioned media were
separated by polyacrylamide gel electrophoresis, then transferred
onto nitrocellulose membranes. Blocking was performed with 5%
non-fat dry milk in PBS Tween. Primary anti-VE-cadherin antibody
was purchased from Bender MedSystems™ (BMS158) and used at 1
µg/ml. For the loading control, the primary anti-GAPDH (6C5)
sc-32233 was purchased from Santa Cruz Biotechnologies (Santa Cruz,
CA, USA) and used at used at 0.1 µg/ml. Secondary
peroxidase-conjugated antibodies (anti-mouse 115-035-146 for GAPDH,
and anti-rabbit 711-035-152 for VE-cadherin) were purchased from
Jackson ImmunoResearch Laboratories, Inc (West Grove, PA USA) and
used at 0.08 µg/ml. Peroxidase activity was recorded by
chemoluminescent detection using the ECL™ kit and Hyperfilm™ MP
from Amersham Biosciences/GE Healthcare (Little Chalfont, UK).
In vivo experiments
In vivo experiments were performed according
to approved institutional guidelines. Specific authorization no.
59-00994 was granted by the institutional veterinary
authorities.
Subcutaneous injections
MMT cells were subcutaneously injected into female
nu/nu BALB/c mice, in Growth Factor-Reduced Matrigel ™, at a
density of 300,000 cells per 100 µl plug. Mice were housed
in a sterile atmosphere in isolators (25°C; humidity range
maintained between 40 and 70%) with food and water ad
libitum, and 8-week-old mice, weighing approximately 20 g, were
used in the experiments. A total of 4-5 mice were used for each
condition. Ten days later, the anesthetized animals received
injections of 100 ml PBS with 2% FITC-dextran (MW 250 000,
Sigma-Aldrich, St. Louis, MO, USA) in the tail vein. An excision
was then performed in the skin of the flanks to unveil the
underlying tumor vasculature. Mice were then sacrificed and the
tumors were fixed in PBS with 4% paraformaldehyde and processed for
paraffin embedding.
Ex vivo tumor explant angiogenesis
assay
Tumors obtained in the above-mentioned experiment
were partially cut in small sections prior to the fixation step.
Fragments of equivalent size were seeded upon matrix gels in which
10,000 diI-labelled MSS-31 cells had been embedded.
Metastasis assays
For lung colonization assays, the MMT cells were
injected in the tail vein of female nu/nu BALB/c mice (100,000
cells per mouse in sterile PBS). Four mice, different from those
used for subcutaneous injections, were used for each condition. The
mice were sacrificed 12 days later and tumors were dissected. The
lungs were fixed with 4% paraformaldehyde in PBS and
paraffin-embedded.
Immunohistology
Immunohistodetection of the endothelial markers was
performed as previously described (14). Briefly, 7-µm-thick tissue
sections were dewaxed and incubated for 30 min in PBS with 2.5
mg/ml trypsin to restore antigen accessibility. Following
endogenous peroxidase inhibition, blocking was carried out with
0.5% casein in PBS. The sections were incubated overnight at 4°C
with anti-CD31/PECAM (01951A, 50 µg/ml) or anti-CD34
(553731, 50 µg/ml) antibodies (Pharmingen/BD Biosciences,
San Jose, CA, USA), then sequentially incubated with biotinylated
anti-rat IgG secondary antibodies (65-153, Jackson ImmunoResearch
Laboratories) diluted 1:5,000 and avidin peroxidase (A-2004, Vector
Laboratories, Burlingame, CA, USA) diluted 1:1,000. Peroxidase
activity was revealed by DAB chromogen (Dako, Glostrup, Denmark)
and tissues were counterstained with Mayer’s hematoxylin.
Statistical analysis
Each experiment was repeated at least 3 times and
the results are expressed as the mean ± standard error. Statistical
analysis was carried out with RStudio (RStudio, Inc., Boston, MA,
USA) open source data analysis software. ANOVA with the post hoc
Tukey range test provided adjusted P-values. Statistical
significance was reached with values of P<0.05, or P<0.01;
otherwise differences were declared non-significant (NS).
Results
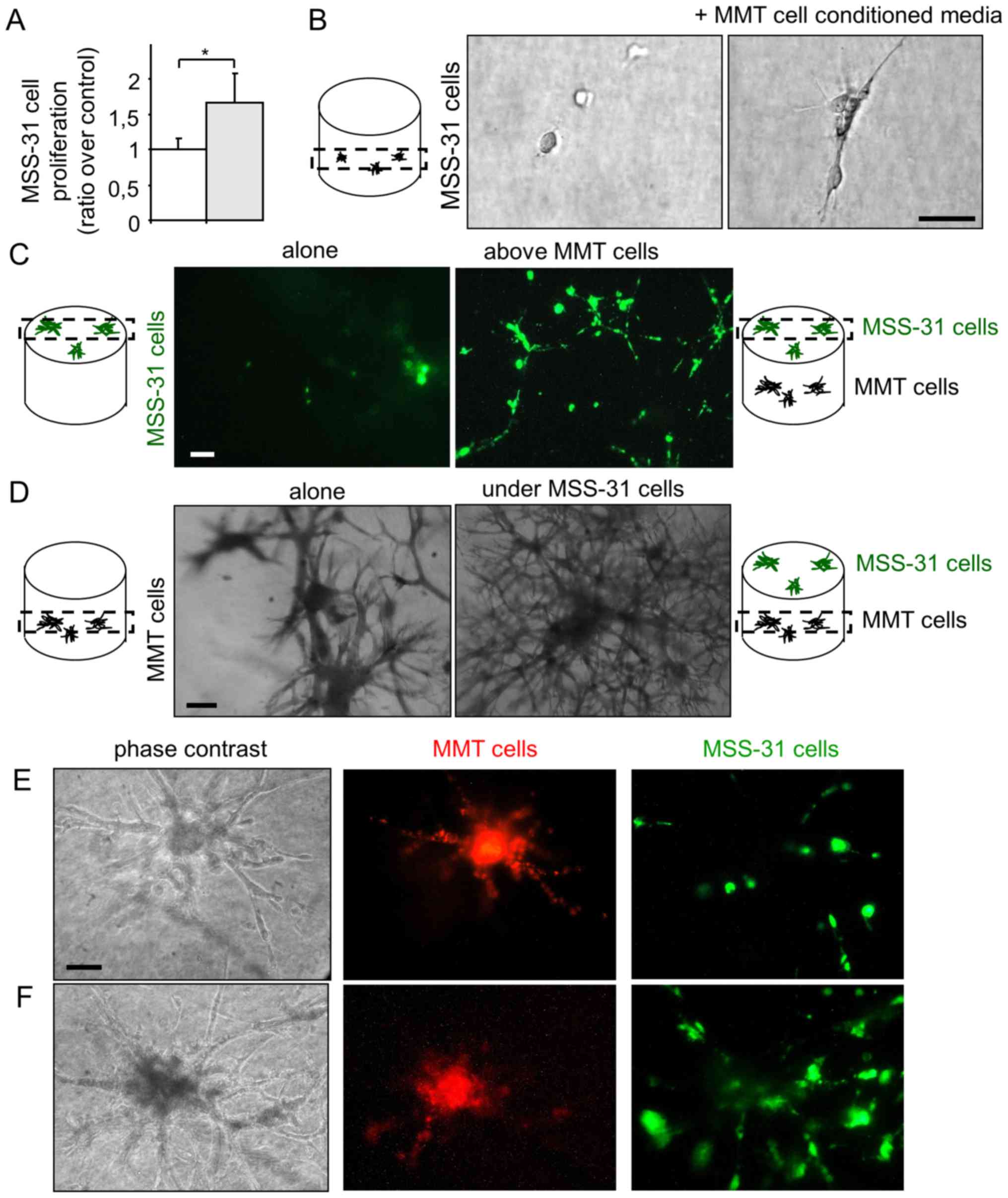
Breast cancer cells and endothelial cells
establish a bidirectional dialogue in co-culture models
In order to examine angiogenic induction by breast
cancer cells, we set up co-culture models of MMT cancer cells with
normal endothelial MSS-31 cells. We first collected conditioned
media from MMT control cancer cells, and administered them to
endothelial cell cultures. Endothelial cells were responsive to
cancer cell soluble stimuli and displayed an increased
proliferation (Fig. 1A), when
compared to the condition control in which they received the
corresponding vehicle alone. To more closely mimic physiological
angiogenic induction, we placed the endothelial cells in 3D matrix
gels composed by type I collagen and reconstituted basal
membrane-like compounds, namely Matrigel™. In the absence of
additional stimuli, endothelial cells failed to divide in this
environment. On the contrary, when they received conditioned media
from the cancer cells, the endothelial cells began to grow,
elongate and organize (Fig.
1B).
Finally, we took our experiment a step further and
co-cultured both the cancer and endothelial cells in such matrix
gels. We used the intravital fluorochromes, diI and diO, to label
each cell type prior to its seeding. Endothelial cells were grown
on top of the gels containing (or not) cancer cells. Within 1 week,
the endothelial cells formed a layer under the control conditions,
and became organized as a network when the cancer cells were
cultured in the gels (Fig. 1C,
highlighting diO-labeled endothelial cells). At the same time, the
MMT cancer cells grew within the gel and lost most of their
fluorescent signal during cell divisions. The presence of
endothelial cells upon the gel stimulated the morphogenesis of
underlying cancer cells, with more branching points and thinner
primary branches when compared to the control condition (Fig. 1D), thus providing evidence of a
reciprocal dialogue between cancer and endothelial cells in
co-culture.
When the MMT cancer cells grew in the vicinity of
the gel surface, it was possible to observe the formation of
connected branched structures between cancer and endothelial cells
(Fig. 1E and F). These hybrid
structures developed over time (8 days of co-culture for Fig. 1F vs. 3 days for Fig. 1E); however, cell fluorescence
weakening did not allow an unambiguous observation of heterotypic
cell contacts. The increased presence of endothelial cells after 8
days of co-culture suggests a possible recruitment of endothelial
cells by cancer cells, although we cannot completely rule out the
contribution of endothelial cell proliferation.
Ets-1 enhances breast cancer cell ability
to induce endothelial cell capillary-like morphogenesis, but not
proliferation
Ets-1 regulates the expression of invasion-related
molecules by breast cancer cells. In order to determine whether its
expression by cancer cells can influence their ability to induce
angiogenesis, the MMT cells were infected by MFG retroviral vectors
to overexpress Ets-1 or the dominant negative mutant DB (for DNA
binding domain), as previously described (8). These cells will be respectively
termed MMT Ets-1 and MMT DB cells hereinafter. MMT cells infected
with the empty vector carrying neomycin resistance were used as
control MMT neo cells. The validation of construct expression has
been previously provided (8).
To assess the impact of these manipulations on
cancer cell angiogenic ability, MSS-31 murine endothelial cells
received conditioned media (CM) from our different MMT sublines.
The administration of these conditioned media for 3 days stimulated
MSS-31 cell proliferation (+67% with MMT neo CM, Fig. 2A). Neither Ets-1 overexpression nor
the dominant negative expression statistically altered this induced
proliferation (+75% with MMT Ets-1 CM, and +74% with MMT DB CM,
Fig. 2A; NS).
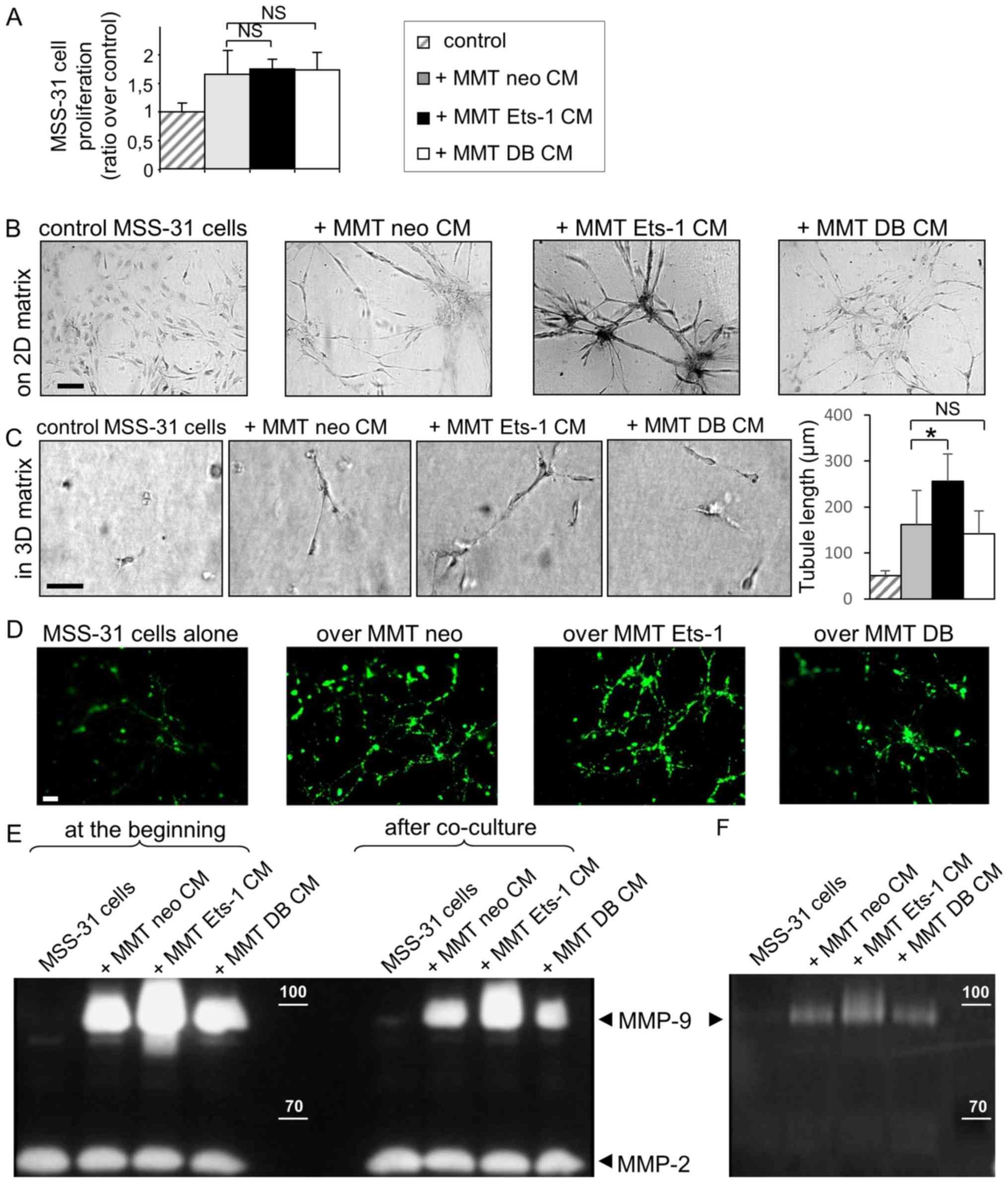 | Figure 2Ets-1 enhances breast cancer cell
ability to induce endothelial cell morphogenesis but not
proliferation. (A) MSS-31 cell proliferation was measured after a
3-day culture in 70% complete medium + 30% MMT cancer cell
conditioned media or corresponding vehicle. Values are
representative of 3 independent experiments. NS, non-significant.
(B) MSS-31 cell morphogenesis on top of matrix gels is increased in
response to MMT cell conditioned media, in an Ets-1-dependent
manner. Images were acquired after neutral red staining of a 6-day
culture. (C) MMT cell conditioned media enhance MSS-31 cell
morphogenesis within 3D matrix gels in serum-free conditions, in an
Ets-1-dependent manner. Images were acquired after a 10-day
culture. *P<0.05; NS, non-significant (D) DiO-labeled
MSS-31 cells were cultured upon matrix gels containing MMT cells.
Endothelial cell networks were visualized under epifluorescence.
(E) Gelatin zymography was performed with MSS-31 culture
supernatants, just after (left columns) or 24 h after (right
columns) the addition of conditioned media from MMT neo, MMT Ets-1
or MMT DB cells, or their control non-conditioned medium. MMP-2 and
MMP-9 activities are detected as translucent bands amid the
blue-stained gelatin substrate, and can be discriminated thanks to
their different molecular weight. The position of 100 and 70 kDa
known from the molecular ladder is indicated by the white dashes.
Zymography evidences the decrease in MMP-9 activity in supernatants
after incubation with endothelial cells. (F) Gelatin zymography of
MSS-31 cell membrane extracts following a 5-min incubation with
conditioned media from MMT cells, or their medium in control,
highlights the recruitment of cancer-derived MMP-9 to the
endothelial cell membrane. Scale bars, 50 µm. |
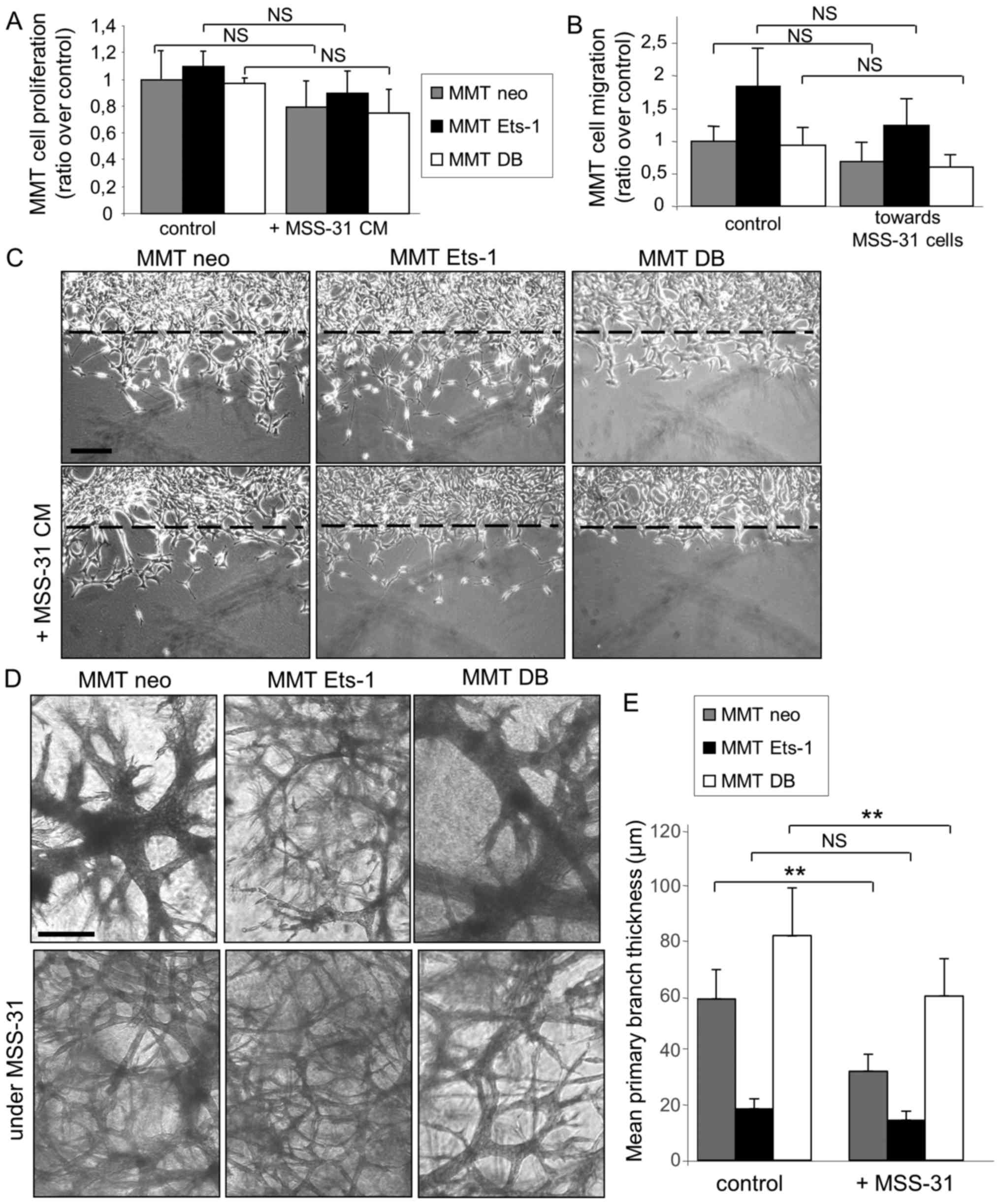
Proliferation is required for angiogenesis, but
invasion and cell organization represent other key features of this
process. Endothelial cell branching morphogenesis in matrix gels
recapitulates several aspects of physiological capillary formation
(15). In this study, we thus
investigated the role of Ets-1 in MMT cell angiogenic potential in
matrix models. When cultured upon these matrix gels in the absence
of proper stimulation, the MSS-31 cells formed a layer (Fig. 2B). The administration of MMT cell
conditioned medium stimulated endothelial capillary-like
morphogenesis (3.3+/−1.2 branching points per field of view) and
this effect was exacerbated when medium was conditioned from
Ets-1-overexpressing MMT cells (Fig.
2B) (5+/−1.7 branching points per field of view). By contrast,
dominant negative DB mutant expression reduced MMT cell ability to
induce endothelial cell morphogenesis, with the failure to trigger
any tubular structure, further highlighting the critical role of
Ets-1 in MMT cell angiogenic potential.
We then investigated angiogenic induction within the
three-dimensional matrix gels under serum-free conditions, in order
to remove the contribution of growth factors and proteases present
in the serum. Endothelial cells embedded alone in these 3D matrix
gels failed to form any cellular structure. The administration of
MMT cell conditioned medium induced the alignment of elongated
MSS-31 cells (with a mean structure length of 161 µm)
(Fig. 2C). Ets-1 overexpression in
MMT cells increased the mean length of MSS-31 structures (251
µm; P=0.04), and DB expression did not alter it in a
significant manner (142 µm; P=0.9) (Fig. 2C). Cell conditioned media were,
however, not sufficient to trigger the formation of hollow lumen
endothelial tubules within these gels.
As already explained in Fig. 1C, we set up 3D co-culture models of
breast cancer and endothelial cells to further investigate the
interactions between these cells. Endothelial cells, labeled with
the intravital fluorescent diO probe prior to their seeding, were
seeded upon 3D matrix gels containing MMT cancer cells. This model
allowed us to follow endothelial capillary-like morphogenesis in
the vicinity of breast cancer cells. The analysis of cell
structures by epifluorescence revealed the implementation of an
endothelial network with branched tubules in presence of MMT cells.
Ets-1 overexpression in the MMT cells stimulated the architecture
of endothelial tubules, whereas DB dominant negative mutant
expression hindered their formation (Fig. 2D).
We also wished to determine which soluble factors
may participate in this angiogenic induction. Ets-1 orchestrates
matrix metalloproteinase (MMP) expression/activation by breast
cancer cells (8) and MMPs are
important for endothelial cell morphogenesis. In this study, we
thus investigated whether MMPs produced by breast cancer cells
could be recruited by endothelial cells. To do that, we analyzed
the gelatinolytic (MMP-2/-9) activity of MMT conditioned media just
after their administration to MSS-31 cells, or 24 h later. At both
time-points, MMP-9 activity was mainly derived from MMT conditioned
media, as there was almost no activity in the control MSS-31 cells
condition (with the appropriate vehicle), and MMP-9 activity was
associated with the Ets-1 status in MMT cells (Fig. 2E), in agreement with our previous
findings (8). Nevertheless, MMP-9
activities in the media were decreased following a 24-h incubation
with endothelial cells (right columns compared to left columns,
Fig. 2E). MMP-2, which is mainly
derived from the control medium, served as a loading control. In
order to determine whether MMP-9 can be recruited by MSS-31 cells,
we incubated the MSS-31 cells for a short period of time (5 min)
with MMT conditioned media, then thoroughly washed the cells 3
times. While the MSS-31 cell membrane extracts displayed no MMP-9
activity under the control conditions, those from the MSS-31 cells
incubated with MMT cell conditioned media exhibited MMP-9
activities varying accordingly to MMT cell productions, whereas no
MMP-2 activity was detected (Fig.
2F). Endothelial cells can thus recruit MMP-9 released by
cancer cells.
Endothelial cells slightly decrease
breast cancer cell proliferation and migration on plastic, but
increase their invasive morphogenetic properties in matrix
gels
After having ascertained MMT breast cancer cell
angiogenic potential and having highlighted Ets-1 critical role in
this process, we investigated the mechanisms through which
endothelial cells can reciprocally modify breast cancer cell
behaviour, as suggested in Fig.
1D.
We first assessed the ability of endothelial cell
conditioned medium to influence MMT cell proliferation.. The
proliferation of the three MMT cell sublines was reduced in
presence of MSS-31 cell conditioned medium, although statistical
significance was not reached for any of them (Fig. 3A). The chemotactic capacity of
endothelial MSS-31 cells for MMT cells was then assessed with
Transwell assays. We noted that the presence of MSS-31 cells
induced a slight (but non-statistically significant) decrease in
the migration of the 3 MMT cell sublines, although
Ets-1-overexpressing cells still migrated to a greater extent than
their counterparts (Fig. 3B).
MSS-31 cell conditioned medium was also administered to wounded MMT
cell cultures, and similarly slightly decreased their migration, as
visualized 24 h later (Fig.
3C).
We further examined the response of the breast
cancer cells to the endothelial cell presence in direct co-culture
systems, as described above; i.e., with MSS-31 cells cultured upon
matrix gels containing MMT cells. The branching morphogenesis of
MMT cell sublines under control conditions, i.e., without
endothelial cells above them, was characteristic of previously
described features (8,9). Ets-1 overexpression favoured
individual invasion, while dominant negative DB expression induced
the formation of thicker tubules when compared to MMT neo cells
(Fig. 3D, top panels). Of note,
MSS-31 culture above MMT-containing gels altered the morphogenesis
of our 3 MMT cell sublines (Fig.
3D, bottom panels), with a decrease in the primary branch
thickness (Fig. 3E) of the MMT neo
and MMTDB cells. These findings demonstrate that endothelial cells
per se can favour the expression of aggressive traits by
cancer cells without providing them with any blood supply.
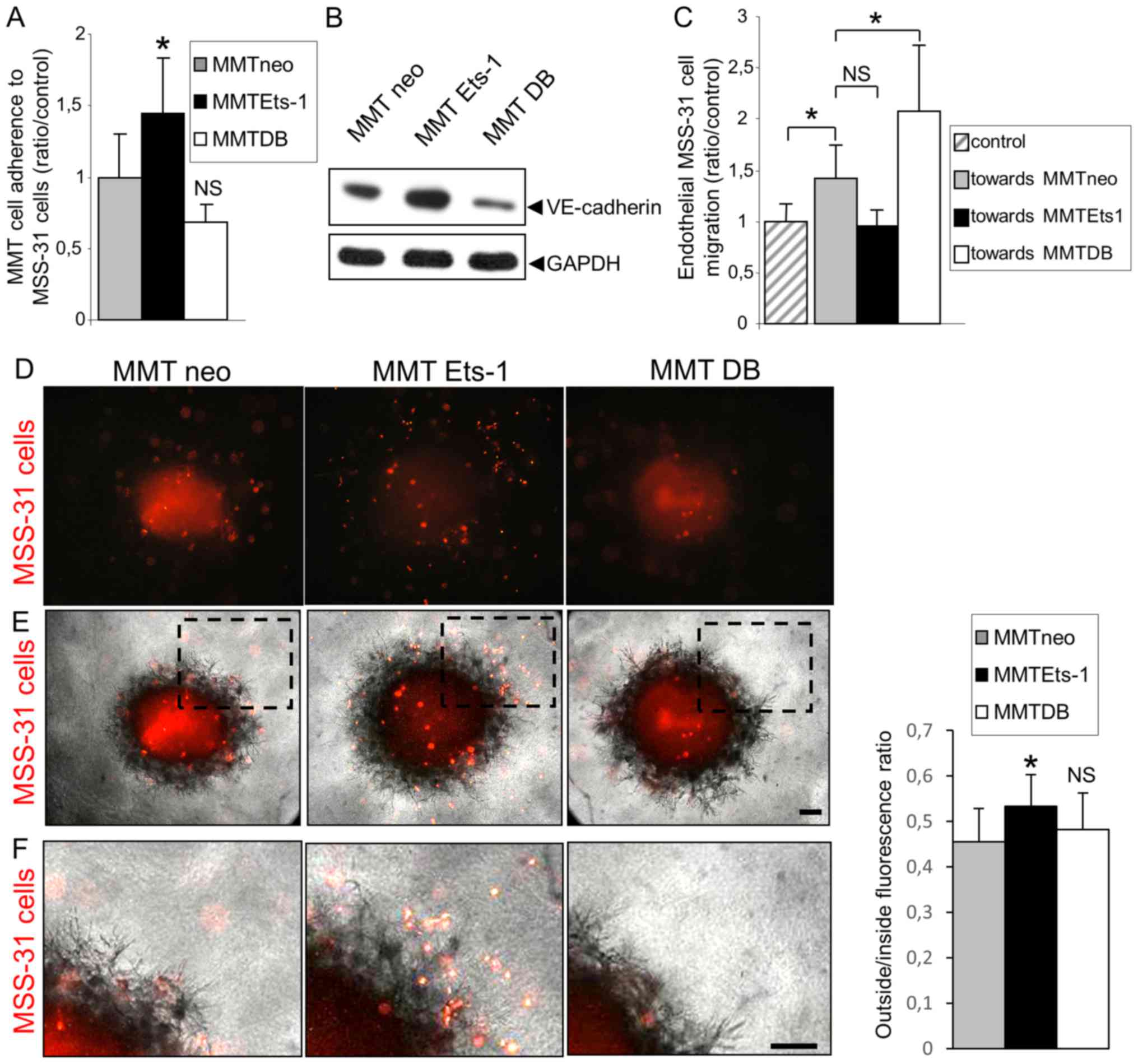
Ets-1 overexpression promotes breast
cancer cell adhesion to endothelial cells, while decreasing their
chemo-attractive potential for endothelial cells
Another key component of cancer cell interactions
with endothelial cells in vivo is their ability to physically
interact with the latter, which may physiologically affect their
metastatic potential. Such interactions depend on two main
parameters: Intercellular adhesion and chemoattraction.
To evaluate whether Ets-1 regulates the processes of
adhesion between cancer and endothelial cells, we examined whether
the modulation of Ets-1 in cancer cells can alter their adherence
to endothelial cells. MMT cell sublines were fluorescently labeled
prior to their seeding on a confluent MSS-31 cell monolayer.
Following 30 min of incubation, non-adherent cells were removed by
3 washes and epifluorescence analysis was performed to quantify the
number of cancer cells attached to the endothelial layer. Of note,
there were 41.2% (P=0.04) more MMT Ets-1 cells adherent to
endothelial cells, and 24.8% (P=0.056) less MMT DB cells adherent
when compared with the MMT neo cells (Fig. 4A). We found that Ets-1
overexpression favored VE-cadherin expression in the MMT cells and
DB mutant decreased it (Fig. 4B),
highlighting a potential factor involved in these heterotypic
interactions.
Physical interactions between cancer and endothelial
cells also depend on their probability to meet each other. We
previously reported that Ets-1 favored breast cancer cell migration
(8), and we wished to determine
whether it can also affect endothelial cell recruitment. For this
purpose, we co-cultured the MSS-31 cells and MMT cells in Boyden
chambers. Cancer cells were grown at the bottom of the wells, and
endothelial cells were seeded upon inserts in these wells. The
presence of MMT neo cells increased MSS-31 cell migration by 35.2%
(P=0.03) when compared to the control condition without underlying
cancer cells (Fig. 4C).
Unexpectedly, Ets-1 overexpression in MMT cells led to a decrease
in MSS-31 cell migration by 28.5% (P=0.18; NS), while DB mutant
expression tended to increase it (+35.8%; P=0.04), in comparison
with the presence of MMT neo cells. We also evaluated the ability
of ex vivo MMT tumor fragments retrieved from in vivo
grafts in mice to recruit endothelial cells. These fragments were
dropped on 3D matrix gels containing fluorescently labeled and
homogenously scattered MSS-31 endothelial cells. MSS-31 cell
distribution in these gels was followed over time by
epifluorescence. Following a 3-day culture, control MMT neo and MMT
DB fragments had recruited most endothelial cells in their core or
their vicinity, whereas endothelial cells were still scattered
around MMT Ets-1 tumor fragments (Fig.
4D and E, and enlargements in Fig.
4F). Fluorescence distribution was quantified inside and
outside the fragment zone, and confirmed that endothelial cells
were less recruited by MMT Ets-1 fragments (outside/inside ratio of
53.4% vs. 45.5% for MMT neo, P=0.02, and 48.2% for MMT DB, P=0.85,
NS when compared to MMT neo).
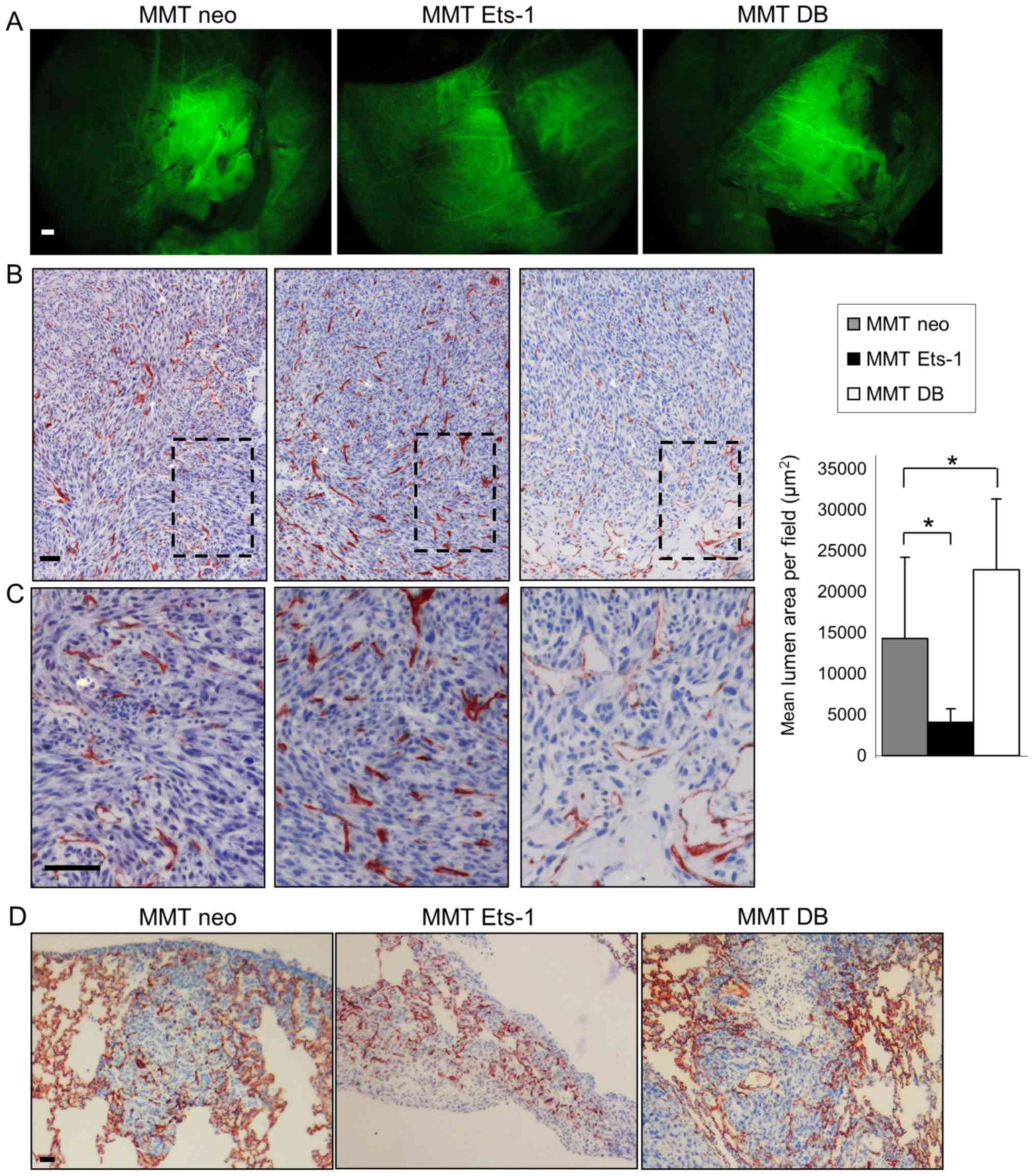
Ets-1 qualitatively alters MMT cell tumor
vascularization in vivo
In order to understand the in vivo relevance
of our afore-mentioned in vitro observations, we
investigated tumor angiogenesis induction in MMT cell tumor
xenografts using nude mice.
MMT cells subcutaneously injected with growth factor
reduced Matrigel™ gave rise to tumors, whose angiogenesis was
analyzed 10 days later. Mice were intravenously injected with
FITC-dextran to visualize blood vessels and then euthanized and
dissected to unveil the tumors from the covering skin. All MMT
tumors were perfused with the systemic circulation independently of
Ets-1 modulation (Fig. 5A). These
experiments revealed that although Ets-1 overexpression hindered
isolated endothelial cell recruitment in vitro, it did not
impair the co-option in vivo by the tumor of surrounding
blood vessels and its perfusion.
The tumors were then processed for immunostaining
with a CD34 endothelial marker, which highlighted the presence of
both large vessels and capillaries. This staining evidenced
Ets-1-driven differences in the tumor vasculature features. While
Ets-1 overexpression favored the presence of numerous capillaries
in MMT tumors, DB mutant expressing tumors displayed only few
intratumoral capillaries, but instead presented bigger vessels,
particularly at the tumor periphery (Fig. 5B and C). This effect was quantified
by measuring the capillary lumen areas on the tumor sections. The
MMT neo tumor sections displayed a mean cumulative lumen surface of
14,301 µm2 per field of view; Ets-1
overexpression by MMT tumors decreased this value to 4,075
µm2 (P=0.03), whereas the dominant negative
increased it to 22,685 µm2 (P=0.048) (Fig. 5B).
Since interactions between tumors and blood vessels
play a key role in the metastatic process, we wished to determine
whether the Ets-1-mediated modulation of these interactions results
in the modification of the tumor metastatic potential. The rapid
growth of MMT subcutaneous tumors however, prevented the
observation of metastasis when mice were dissected. To circumvent
this issue, we directly injected MMT cells into the tail vein of
nude mice, which is a classical experimental metastasis assay. In
this experimental frame, Ets-1 or its dominant negative mutant
overexpression did not alter the number of lung metastases nor
their vascularization, probably derived from the lung dense
capillary network (Fig. 5D). MMT
cell tropism for the lung, which is a highly vascularised organ,
was not favourable to understand a putative angiogenic modulation
by Ets-1 following cancer cell dissemination. These data merely
indicate that Ets-1 modulation does not affect cancer cell
entrapment in the lung capillaries.
Discussion
Cancer cell invasion and sprouting angiogenesis
share some common molecular circuitry. Since Ets-1 orchestrates
breast cancer cell invasion via different soluble factors, we
reasoned that it may affect their angiogenic potential, which
proved to be the case. Among such soluble factors, tumor-secreted
MMP-9 has been reported to induce angiogenesis (16). In this study, we demonstrate that
endothelial cells actually recruit cancer cell-derived MMP-9, and
that the amount of recruited MMP-9 depends on Ets-1
expression/activity in cancer cells. Future experiments are
warranted to unveil the importance of this mechanism in endothelial
cell morphogenesis. We also demonstrate that Ets-1 overexpression
in cancer cells promotes their adhesion to endothelial cells.
VE-cadherin classically links adjacent endothelial cells, but can
be expressed by aggressive cancer cells in what is known as
vasculogenic mimicry (17). Ets-1
has been proposed to drive together with HIF-2α the expression of
the VE-cadherin gene by tumor cells (18). We actually found that Ets-1 favored
VE-cadherin expression in MMT cells. In 3D co-culture models, we
observed hybrid tubules made of cancer and endothelial cells.
However, the rapid loss of cancer cell fluorescent labeling
impaired us to quantify these structures and to investigate the
putative impact of Ets-1 on vascular mimicry. With regard to cell
adhesion, Ets-1 also controls β3 subunit integrin expression
(8), which can contribute to the
adhesion of cancer cells to endothelial cells (19). To sum up, Ets-1 controls the
expression of many molecules associated with adhesion and invasion.
Therefore, it probably regulates angiogenic interactions via this
network but not via a single molecule. Identifying the contribution
of these individual actors and their combinations may be complex,
since each actor can influence the others (e.g., MMPs can release
or activate growth factors, which in turn control MMP expression…).
Moreover, both cell partners can express the same molecules and
reciprocally influence their secretion, further complicating such a
type of study. In this study, we chose to rather focus on the
phenotypic results of cellular interactions and dialog.
Importantly, this study highlights the reciprocal
communication existing between cancer and endothelial cells. The
in vitro signals elicited by endothelial cells demonstrate
that these cells are far from being ‘inert’ cells whose only role
is to form blood vessels in order to supply blood. This stands in
agreement with other studies showing the importance of angiocrine
factors in both normal and tumor settings. For example, it has been
shown that during embryogenesis, primitive endothelial cells can
control the developing endoderm (20) and that vessels contribute to
pancreatic differentiation via inductive interactions (21).
In the field of tumorigenesis, much emphasis has
been placed on the nutrient and oxygen supply since tumor
angiogenesis was highlighted by Judah Folkman (22). The production by endothelial cells
of paracrine factors potentially acting on tumor cells is yet a not
so recent concept (23), but it
has received little attention in the decade following its
discovery. To the best of our knowledge, few studies have focused
on reciprocal paracrine interactions between cancer and endothelial
cells in co-culture. Bi-directional interactions have been reported
in vitro between prostate cancer cells and endothelial cells
(24). The induction by cancer
cells of an autocrine endothelial activation was also evidenced in
another co-culture model (25).
With regard to in vivo relevance, it has beem described that
endothelial cells can promote the early survival of tumor cells
injected in mice, in a paracrine manner, prior to the onset of
angiogenesis (26). More recently,
it was shown that endothelial cords promote experimental melanoma
growth in mice prior to their blood perfusion (27). Our in vitro data support
this angiocrine notion and suggest that breast cancer cells may
also be sensitive to such signals elicited by their endothelial
partner in the tumor microenvironment.
In this study, we demonstrate that Ets-1
qualitatively controls tumor angiogenesis, rather than being a mere
on/off switch. It is interesting to note that MMT DB tumors display
larger blood vessels, particularly on their periphery. It is
unlikely that such a vasculature is efficient enough to satisfy
tumors with their needs, as evidenced by frequent necrotic areas.
Conversely, Ets-1 expression in MMT tumors rather favors the
formation of capillaries in the whole tumor. This network may
sustain the release of angiocrine factors by activated endothelium,
but is probably not a better guarantee of perfusion since tumor
capillaries are often permeant and lack organization. These
opposite angiogenic features may be reconciled in a same natural
tumor if we consider that Ets-1 overexpression is classically not
widespread in human breast tumors, but is rather induced in the
tumor-stroma interaction zone (28). While the tumor bulk may recruit
blood vessels, Ets-1-overexpressing cells can disseminate owing to
their exacerbated motility capacities, which confer to them a high
probability to meet partners in the nearby stroma. Moreover, their
higher capacity to adhere to endothelial cells may help them to
find a way in the systemic circulation via intravasation. Since the
experimental metastasis assay used bypasses the early steps in the
metastatic process, it does not help to elucidate whether Ets-1
putatively favors intravasation via pro-migratory and pro-adhesive
activities. In the future, more sophisticated systems with for
example, the surgical removal of the primary tumor, may help to
highlight and unveil Ets-1 role in these early steps in
vivo.
Of note, it has recently been shown that
epithelial-to-mesenchymal transition (EMT) phenotypes in tumor
cells are associated with the release of soluble factors able to
induce angiogenesis and recruit myeloid cells (29). In that study, Snail transcription
factor was shown to mediate the induction of such factors.
Targeting transcription factors such as Ets-1, Twist or Snail may
thus represent a therapeutic approach to limit at the same time
invasion and microenvironment activation including
angiogenesis.
Many improvements have been achieved over the past
years in understanding cancer angiogenesis, and have led to the
development of promising therapeutic strategies for breast cancer
patients (30). Nevertheless,
several clinical trials with anti-angiogenic compounds have failed
due to the adaptation of tumors, prompting researchers to obtain
more insight into the tumor vasculature and propose new therapies
(3). Vascular mimicry is
associated with metastasis (31)
and represents a clinical issue since it is a priori refractory to
anti-angiogenic compounds. Identification of compounds blocking
this process should be useful to fight cancers. We have also
illustrated the role that angiocrine factors play in tumor
development. In brief, a better understanding of angiogenesis
modulation by tumors and of the complex interactions between cancer
and endothelial cells may help us to tackle more efficiently this
cancer hallmark and may lead to the development of effective
treatment strategies for patients. Many opportunities are open and
at the same time, much has still to be done to decipher the
underlying mechanisms.
Acknowledgments
The authors are particularly grateful to Dr Fabrice
Soncin and all members of our laboratories for providing tools,
helpful discussions and comments.
Funding
This study was supported by CNRS, Association pour
la Recherche contre le Cancer (ARC), Ligue Nationale contre le
Cancer and Société Française du Cancer (SFC). AF was supported by
Ministère de l’Education Nationale, Ligue Nationale contre le
Cancer and Région Hauts de France (CPER Photonics 4 Society and
1-PRIMER).
Availability of data and materials
Data sharing is not applicable to this article, as
no datasets were generated or analyzed during the current
study.
Authors’ contributions
AF, XD and AP conceived the study. AF and AP
designed experiments. AF, CV, NW and AP performed experiments. AF,
CV, NW, LH, XD and AP analysed and interpreted the data. AF, LH and
AP wrote the manuscript. All authors have read and approved the
final manuscript.
Ethics approval and consent to
participate
In vivo experiments were performed according
to approved institutional guidelines. Specific authorization no.
59-00994 was granted by the institutional veterinary
authorities.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN. 2012.Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar
|
|
2
|
Hansen S, Grabau DA, Sørensen FB, Bak M,
Vach W and Rose C: The prognostic value of angiogenesis by Chalkley
counting in a confirmatory study design on 836 breast cancer
patients. Clin Cancer Res. 6:139–146. 2000.PubMed/NCBI
|
|
3
|
Carmeliet P and Jain RK: Molecular
mechanisms and clinical applications of angiogenesis. Nature.
473:298–307. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Aalders KC, Tryfonidis K, Senkus E and
Cardoso F: Anti-angiogenic treatment in breast cancer: Facts,
successes, failures and future perspectives. Cancer Treat Rev.
53:98–110. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sivridis E, Giatromanolaki A and
Koukourakis MI: The vascular network of tumours - what is it not
for? J Pathol. 201:173–180. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Span PN, Manders P, Heuvel JJ, Thomas CMG,
Bosch RR, Beex LVAM and Sweep CGJ: Expression of the transcription
factor Ets-1 is an independent prognostic marker for relapse-free
survival in breast cancer. Oncogene. 21:8506–8509. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kola I, Brookes S, Green AR, Garber R,
Tymms M, Papas TS and Seth A: The Ets1 transcription factor is
widely expressed during murine embryo development and is associated
with mesodermal cells involved in morphogenetic processes such as
organ formation. Proc Natl Acad Sci USA. 90:7588–7592. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Furlan A, Vercamer C, Desbiens X and
Pourtier A: Ets-1 triggers and orchestrates the malignant phenotype
of mammary cancer cells within their matrix environment. J Cell
Physiol. 215:782–793. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Furlan A, Vercamer C, Bouali F, Damour I,
Chotteau-Lelievre A, Wernert N, Desbiens X and Pourtier A: Ets-1
controls breast cancer cell balance between invasion and growth.
Int J Cancer. 135:2317–2328. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yamada KM and Cukierman E: Modeling tissue
morphogenesis and cancer in 3D. Cell. 130:601–610. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mattot V, Vercamer C, Soncin F, Calmels T,
Huguet C, Fafeur V and Vandenbunder B: Constitutive expression of
the DNA-binding domain of Ets1 increases endothelial cell adhesion
and stimulates their organization into capillary-like structures.
Oncogene. 19:762–772. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yanai N, Satoh T and Obinata M:
Endothelial cells create a hematopoietic inductive microenvironment
preferential to erythropoiesis in the mouse spleen. Cell Struct
Funct. 16:87–93. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bradford MM: A rapid and sensitive method
for the quantitation of microgram quantities of protein utilizing
the principle of protein-dye binding. Anal Biochem. 72:248–254.
1976. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pourtier-Manzanedo A, Vercamer C, Van
Belle E, Mattot V, Mouquet F and Vandenbunder B: Expression of an
Ets-1 dominant-negative mutant perturbs normal and tumor
angiogenesis in a mouse ear model. Oncogene. 22:1795–1806. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Arnaoutova I, George J, Kleinman HK and
Benton G: The endothelial cell tube formation assay on basement
membrane turns 20: State of the science and the art. Angiogenesis.
12:267–274. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mira E, Lacalle RA, Buesa JM, de Buitrago
GG, Jiménez-Baranda S, Gómez-Moutón C, Martínez-A C and Mañes S:
Secreted MMP9 promotes angiogenesis more efficiently than
constitutive active MMP9 bound to the tumor cell surface. J Cell
Sci. 117:1847–1857. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hendrix MJC, Seftor EA, Hess AR and Seftor
REB: Vasculogenic mimicry and tumour-cell plasticity: Lessons from
melanoma. Nat Rev Cancer. 3:411–421. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Le Bras A, Lionneton F, Mattot V, Lelièvre
E, Caetano B, Spruyt N and Soncin F: HIF-2alpha specifically
activates the VE-cadherin promoter independently of hypoxia and in
synergy with Ets-1 through two essential ETS-binding sites.
Oncogene. 26:7480–7489. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Leroy-Dudal J, Demeilliers C, Gallet O,
Pauthe E, Dutoit S, Agniel R, Gauduchon P and Carreiras F:
Transmigration of human ovarian adenocarcinoma cells through
endothelial extracellular matrix involves alphav integrins and the
participation of MMP2. Int J Cancer. 114:531–543. 2005. View Article : Google Scholar
|
|
20
|
Bahary N and Zon LI: Development.
Endothelium - chicken soup for the endoderm. Science. 294:530–531.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lammert E, Cleaver O and Melton D:
Induction of pancreatic differentiation by signals from blood
vessels. Science. 294:564–567. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Folkman J: Tumor angiogenesis: Therapeutic
implications. N Engl J Med. 285:1182–1186. 1971. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Rak J, Filmus J and Kerbel RS: Reciprocal
paracrine interactions between tumour cells and endothelial cells:
The ‘angiogenesis progression’ hypothesis. Eur J Cancer.
32A:2438–2450. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Barrett JM, Mangold KA, Jilling T and Kaul
KL: Bi-directional interactions of prostate cancer cells and bone
marrow endothelial cells in three-dimensional culture. Prostate.
64:75–82. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Khodarev NN, Yu J, Labay E, Darga T, Brown
CK, Mauceri HJ, Yassari R, Gupta N and Weichselbaum RR:
Tumour-endothelium interactions in co-culture: Coordinated changes
of gene expression profiles and phenotypic properties of
endothelial cells. J Cell Sci. 116:1013–1022. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shan S, Robson ND, Cao Y, Qiao T, Li CY,
Kontos CD, Garcia-Blanco M and Dewhirst MW: Responses of vascular
endothelial cells to angiogenic signaling are important for tumor
cell survival. FASEB J. 18:326–328. 2004. View Article : Google Scholar
|
|
27
|
Zhao C, Zhang W, Zhao Y, Yang Y, Luo H, Ji
G, Dong E, Deng H, Lin S, Wei Y, et al: Endothelial cords promote
tumor initial growth prior to vascular function through a paracrine
mechanism. Sci Rep. 6:194042016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Behrens P, Rothe M, Wellmann A, Krischler
J and Wernert N: The Ets-1 transcription factor is up-regulated
together with MMP 1 and MMP 9 in the stroma of pre-invasive breast
cancer. J Pathol. 194:43–50. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Suarez-Carmona M, Bourcy M, Lesage J,
Leroi N, Syne L, Blacher S, Hubert P, Erpicum C, Foidart JM,
Delvenne P, et al: Soluble factors regulated by
epithelial-mesenchymal transition mediate tumour angiogenesis and
myeloid cell recruitment. J Pathol. 236:491–504. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Earl HM, Hiller L, Dunn JA, Blenkinsop C,
Grybowicz L, Vallier AL, Abraham J, Thomas J, Provenzano E,
Hughes-Davies L, et al ARTemis Investigators: Efficacy of
neoadjuvant bevacizumab added to docetaxel followed by
fluorouracil, epirubicin, and cyclophosphamide, for women with
HER2-negative early breast cancer (ARTemis): An open-label,
randomised, phase 3 trial. Lancet Oncol. 16:656–666. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wagenblast E, Soto M, Gutiérrez-Ángel S,
Hartl CA, Gable AL, Maceli AR, Erard N, Williams AM, Kim SY,
Dickopf S, et al: A model of breast cancer heterogeneity reveals
vascular mimicry as a driver of metastasis. Nature. 520:358–362.
2015. View Article : Google Scholar : PubMed/NCBI
|