Introduction
Liver cancer is estimated to be the sixth most
common cancer and the fourth leading cause of cancer-associated
morality worldwide in 2018 (1).
Hepatocellular carcinoma (HCC) is the major histological subtype of
primary liver cancer. Certain factors have been reported to
contribute to HCC, including aflatoxin, chronic hepatitis B virus
infection, obesity, type 2 diabetes, heavy alcohol consumption and
smoking, among others (2). Despite
the advances in current therapeutic approaches, the prognosis of
HCC is extremely poor and the recurrence rate is high.
Consequently, it is crucial to elucidate the molecular mechanisms
underlying metastasis and progression and identify novel
therapeutic targets for HCC.
MicroRNAs (miRNAs/miRs) are a class of highly
conserved, small, non-coding RNAs, ~22 nucleotides in length, which
negatively regulate gene expression by targeting the 3′untranslated
region (3′UTR) of mRNAs (3-5). A
number of studies have confirmed that miRNAs are closely associated
with the HCC cell proliferation, metastasis and cell cycle
distribution, and the prognosis of HCC patients (6-9). A
recent study reported that the expression of miR-124 was
significantly decreased in HCC tissues and cell lines, whereas
miR-124 overexpression inhibited cell proliferation and migration
by targeting baculoviral IAP repeat containing 3 and regulating the
nuclear factor-κB signaling pathway (10). Another study confirmed that
increased miR-142-3p expression suppressed the proliferation,
migration and invasion and reversed epithelial-to-mesenchymal
transition (EMT) of HCC cells by targeting zinc finger E-box
binding homeobox 1 (11).
Recently, attention has focused on miR-300, a novel microRNA that
has been reported to be involved in cell proliferation, migration
and invasion in certain types of cancer, including colorectal
(12), lung (13) and pancreatic (14) cancer. Although there is only one
study reporting that miR-300 may regulate the migration and
invasion of HCC cells by targeting the focal adhesion
kinase/phosphoinositide-3-kinase/protein kinase B signaling pathway
and is associated with poor prognosis (15), the role and mechanism of action of
miR-300 in the development of HCC has not been fully
elucidated.
In the present study, the function of miR-300 in HCC
was found to be associated with lymphoid enhancer-binding factor 1
(LEF-1), a member of the LEF/T-cell-specific factor family, which
has been shown to be associated with the development of tumors and
serves as a prognostic marker for certain types of cancer. Shang
et al (16) demonstrated
that the expression of LEF-1 was increased in stage III/IV and
grade 3 human renal cell carcinoma (RCC) compared with that in
early-stage, low-grade RCC and normal kidney tissues, and further
demonstrated that LEF-1 overexpression increased cell proliferation
by reversing G2/M arrest in HCC cells. In addition, Xu et al
(17) reported that increased
levels of LEF-1 were correlated with poor prognosis of BRAF and
NRAS mutation-negative acral melanoma. A recent study confirmed
that LEF-1 overexpression promoted cell proliferation and
metastasis through the miR-371a-5p/SRC kinase signalling inhibitor
1 (SRCIN1)/pleiotrophin/Slug pathway in HCC cells (18); however, to the best of our
knowledge, whether miR-300 is involved in the regulation of cell
proliferation and metastasis induced LEF-1 in HCC has not been
reported to date.
The aim of the present study was to measure miR-300
expression in HCC and determine whether it is involved in the
proliferation, migration and invasion of HCC cells. It was also
aimed to investigate whether the effects of miR-300 on HCC cells
are mediated through regulation of LEF-1, and their association
with the prognosis of patients with HCC.
Materials and methods
Patient tissue
A total of 86 samples, including 62 HCC tissues
(male 41 and female 21; age range 26-74 years old; mean 52.3±9.8)
and 24 non-tumor liver tissues (male 15 and female 9; age range
26-68 years old; mean 52.0±10.9), were collected from patients with
HCC that underwent surgery at the First Affiliated Hospital of
Bengbu Medical College (Bengbu, China) between September 2011 and
December 2015. The specimens were stored at −80°C immediately after
harvesting for reverse transcription-quantitative polymerase chain
reaction (RT-qPCR) analysis. None of the patients received any
preoperative chemotherapy or radiotherapy prior to surgery.
Informed consent was obtained from each patient, and all the
protocols of this study were approved by the Ethics Committee of
Bengbu Medical College.
Cell culture
Human HCC cell lines (Huh-7, Li-7, Hep3B and
SNU-449) and the normal hepatocyte cell line L02 were purchased
from Cellcook Cell Biotechnology Co., Ltd. (Guangzhou, China) and
cultured in Dulbecco’s modified Eagle’s medium (DMEM; HyClone; GE
Healthcare Life Sciences, Logan, UT, USA) with 10% fetal bovine
serum (FBS; Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) and 1% penicillin/streptomycin (Beyotime Institute of
Biotechnology, Haimen, China). All cell lines were cultured at 37°C
in 5% CO2.
RT-qPCR analysis
Total RNA was purified using TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) following the
manufacturer’s instructions. RT was performed with 2 µg
total RNA using the Revert Aid First-Strand cDNA Synthesis kit
(Thermo Fisher Scientific, Inc.). The incubation conditions were
25°C for 5 min, 42°C for 60 min, 72°C for 5 min and 4°C until the
end of the reaction. Subsequently, PCR was performed using SYBR
Premix Ex Taq II (Takara Biotechnology Co., Ltd., Dalian, China)
with primers against target genes on the ABI 7500 Real-Time PCR
System (Thermo Fisher Scientific, Inc.). The thermocycling
conditions were as follows: Holding stage, 50°C for 2 min, 95°C for
30 sec; PCR stage (40 cycles), 95°C for 15 sec, 53.6°C for 30 sec,
72°C for 34 sec. For miRNA, RT and PCR were performed using
TransScript Green miRNA Two-Step RT-qPCR SuperMix (Beijing Transgen
Biotech Co., Ltd., Beijing, China) with miR-specific primers on the
ABI 7500 Real-Time PCR System. The incubation conditions for RT
were 37°C for 60 min and 85°C for 5 sec. The thermocycling
conditions for PCR were: Holding stage, 50°C for 2 min and 94°C for
30 sec; PCR stage (40 cycles) 94°C for 5 sec, 55°C for 34 sec and
72°C for 34 sec. All primers were purchased from Sangon Biotech
Co., Ltd. (Shanghai, China). GAPDH and U6 were used as endogenous
control for normalizing mRNA and miRNA, respectively.
Quantification was conducted to analyze mRNA or microRNA expression
relative to the endogenous control using the 2−ΔΔCq
method (19). ΔΔCq = [(Cq gene of
interest - Cq internal control) sample A - (Cq gene of interest -
Cq internal control) sample B]; sample A is the treated sample and
sample B, the untreated control. Each sample was run in triplicate.
The primers used in the present study were as follows: LEF-1,
forward 5′-GCAGCACTTAGAAGGGGCTT-3′ and reverse
5′-GCAGCACTTAGAAGGGGCTT-3′; E-cadherin, forward
5′-GAAGTGTCCGAGGACTTTGG-3′ and reverse 5′-CAGTGTCTCTCCAAA
TCCGATA-3′; vimentin, forward 5′-TGTCCAAATCGATGT GGATGTTTC-3′ and
reverse 5′-TTGTACCATTCTTCTGCC TCCTG-3′; miR-300, forward,
5′-CGTATACAAGGGCAGACTCTCTCT-3′; GAPDH, forward
5′-CAGCCTCAAGATCATCAGCA-3′ and reverse 5′-TGTGGTCATGAGTCCTTCCA-3′;
and U6, forward, 5′-AGAGAAGATTAGCATGGCCCCT-3′.
Vectors and cell transfections
LEF-1 overexpression and knockdown lentiviral
vectors, and their negative controls, were purchased from Shanghai
GeneChem Co., Ltd. (Shanghai, China). Huh-7 and Hep3B cells were
infected with lentivirus-transducing units plus 10 µg/ml
polybrene (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany). When
infected, the cell paving density remained around 30-40%. Huh-7 and
Hep3B cells were infected with recombinant lentiviruses at a
multiplicity of infection of 10. LEF-1 overexpression vector and
short hairpin RNA (shRNA) were designed and co-transfected with
miR-300 mimic and inhibitor. LEF-1 coding sequence was inserted
into pcDNA 3.1 vector. Huh-7 and Hep3B cells transfected by LEF-1
overexpression vector were set as LEF-1 group. Furthermore, pcDNA
3.1 empty vector was used to transfect Huh-7 and Hep3B cells, and
these cells were named as control group. miRNAs and their
corresponding negative controls were synthesized by Shanghai
GeneChem Co., Ltd. The sequences were as follows: LEF-1 shRNA1,
5′-GCAGCTATCAACCAGATTCTT-3′; LEF-1 shRNA2,
5′-CCATCAGATGTCAACTCCAAA-3′; LEF-1 shRNA3, 5′-
GCACGGAAAGAAAGACAGCTA-3′; sh R NA non-targeting sequences for
negative control, 5′-TTCTCCGAA CGTGTCACGT-3′; miR-300 mimic, sense
5′-UAUACAAGGGCAGACUCUCUCU-3′ and anti-sense
5′-AGAGAGAGUCUGCCCUUGUAUA-3′; miR negative control, sense
5′-UUCUCCGAACGUGUCACGUTT-3′ and anti-sense 5′-ACGUGACAC
GUUCGGAGAATT-3′; miR-300 inhibitor, sense 5′-GAGAGA
GUCUGCCCUUGUAU-3′; miR-300-inhibitor NC, sense
5′-CAGUACUUUUGUGUAGUACAA-3′. Target cells were transiently
transfected with miR-300 mimic and miR-300 inhibitor using
Lipofectamine® 2000 (Thermo Fisher Scientific, Inc.)
according to the manufacturer’s protocol. A total of
3×105 cells per well were seeded on 6-well-plates and
transfected with 100 pmol miR-300 mimic/inhibitor or 4 µg
LEF-1 overexpression vector. Puromycin (Beijing Solarbio Science
& Technology Co., Ltd., Beijing, China) was used to select
lentivirus stably transfected clones in accordance with the
manufacturer’s protocol, and validation was performed by western
blot and RT-PCR assays. At 24 h after transfection, total RNA from
cells was isolated. At 48 h after transfection, HCC cells were
collected for western blot, cell proliferation and apoptosis, cell
cycle, migration and invasion, colony formation assay, wound
healing, Transwell assay and flow cytometry. Wild-type or mutant
3′UTR sequences of LEF-1 were inserted into the pGL3-promoter
vector (Promega Corporation, Madison, WI, USA) to generate LEF-1
expression vectors. The luciferase-mutant vector, in which the
nucleotides complementary to the miR-300 seed-region were mutated
by site-directed mutagenesis, was constructed as a mutant control.
Luciferase and control signals were measured at 48 h after
transfection.
Western blot analysis
HCC cells were lysed in radioimmunoprecipitation
assay buffer (Beyotime Institute of Biotechnology) supplemented
with protease and phosphatase inhibitor cocktail (Beyotime
Institute of Biotechnology) and placed on ice for 40 min. The cell
lysates were centrifuged at 15,294 × g at 4°C for 20 min. Protein
concentration was quantified using the Bicinchoninic Acid Protein
Assay kit (Beyotime Institute of Biotechnology). The cell extracts
were boiled with loading dye for 10 min. The protein samples (40
µg) were separated by SDS-PAGE on 10% gels and then
transferred onto polyvinylidene difluoride membranes (EMD
Millipore, Billerica, MA, USA). The membranes were blocked in
Western Blocking Buffer at room temperature (Beyotime Institute of
Biotechnology) for 2 h, followed by incubation with primary
antibodies against LEF-1 (cat. no. 76010S; 1:1,000; Cell Signaling
Technology, Inc., Danvers, MA, USA), E-cadherin (cat. no. ab1416;
1:1,000; Abcam, Cambridge, MA, USA), vimentin (cat. no. ab20346;
1:1,000; Abcam) and GAPDH (cat. no. AC002; 1:1,000; ABclonal
Biotech Co., Ltd., Woburn, MA, USA) according to the manufacturer’s
recommendations overnight at 4°C. Following incubation with
goat-anti-rabbit IgG (cat. no. BA1054) and goat-anti-mouse IgG
(cat. no. BA1050) antibody (1:5,000; horseradish peroxidase
conjugates; Beyotime Institute of Biotechnology) at 37°C for 2 h,
chemiluminescence was detected with enhanced chemiluminescence (EMD
Millipore) to analyze the protein levels. GAPDH (cat. no. AC002;
1:1,000; ABclonal Biotech Co., Ltd.) was used as the loading
control. The western blot experiment was analyzed three times, and
representative images are shown in the figures.
miRNA target prediction and luciferase
reporter assay
miR-300 target prediction was performed using the
miRanda and miRDB (mirdb.org/) algorithms. Cells (5×104)
were seeded in 24-well plates and transiently transfected with 0.2
µg reporter plasmid and 20 pmol miRNA using
Lipofectamine® 2000. After 48 h, the cells were
harvested and lysed, and luciferase activity was measured using a
GloMax 20/20 Luminometer (Promega Corporation). PmirGLO vector
Renilla luciferase (hRluc-neo) was used for
normalization.
Cell proliferation and colony formation
assays
Cell proliferation was measured using MTT and colony
formation assays. To evaluate cell viability, 3×103
cells were plated in 96-well plates and incubated for 24 h.
Subsequently, 20 µl MTT dye solution (5 mg/ml; Beijing
Solarbio Science & Technology Co., Ltd.) was added to each well
and incubation was continued for 3-4 h. The supernatant was then
removed, 200 µl DMSO was added to stop the reaction and the
mixture was oscillated for 20 min. Finally, the optical density was
determined using an Epoch microplate spectrophotometer (BioTek
Instruments, Inc., Winooski, VT, USA) at a wavelength of 490 nm. A
cell viability curve was generated based on absorbance and time.
For the colony formation assay, 3,000 cells were seeded into each
well of a 6-well plate and cultured for 7-14 days. The colonies
(>50 cells) were fixed with 20% methanol for 15 min at room
temperature, stained with 0.1% crystal violet solution for 20 min
at room temperature. Visible cell colonies were counted.
Wound healing assay
Huh-7 and Hep3B cells were seeded in 6-well plates
(4-6×105 cells per well). Longitudinal scratch wounds
were created with a sterile 10 µl pipette tip 48 h after
transfection and floating cell debris was removed by washing three
times with PBS. Subsequently, the cells were cultured in serum-free
medium. Typical wound healing images were observed and captured at
0 and 24 h under a light microscope.
Transwell assay
Cell migration and invasion ability was assessed by
24-well Transwell chambers (Costar; Corning Inc., Corning, NY,
USA). At 48 h after transfection, the cells were trypsinized and
counted. Approximately 2-3×104 cells resuspended in 100
µl serum-free DMEM were seeded into the upper chambers with
1:10 diluted Matrigel-coated (BD Biosciences; Becton, Dickinson and
Company, Franklin, Lakes, NJ, USA) for the invasion assay; uncoated
inserts were used for migration assay, whereas the bottom chamber
was filled with 800 µl 10% FBS-supplemented medium. After 24
h, non-migrating/non-invading cells were wiped off with a cotton
bud, and migrating/invading cells underneath the chamber were fixed
with 4% polyoxymethylene for 5 min at room temperature and stained
with 0.1% crystal violet solution for 10 min at room temperature.
The cells were counted in five randomly selected fields under a
light microscope.
Apoptosis detection and cell cycle
distribution
For cell apoptosis detection, 5 µl Annexin
V-phycoerythrin and 5 µl 7-aminoactinomycin D (BD
Biosciences; Becton, Dickinson and Company) were added to the
cells, followed by incubation for 20 min at room temperature. Cell
apoptosis was evaluated by flow cytometer (BD Biosciences; Becton,
Dickinson and Company) and apoptosis rates were analyzed with
FlowJo software (FlowJo LLC, Ashland, OR, USA).
To synchronize cell cultures, cells were cultured in
medium supplemented with 10% FBS overnight, rinsed with PBS, and
then transferred to serum-free medium for 24 h. To test the cell
cycle distribution, transfected cells were digested using trypsin,
centrifuged at 425 × g (cell density, 1×106 cells/ml),
washed twice with PBS and fixed in −20°C ethanol for 2 h. For cell
cycle distribution detection, 1 µl DAPI was added per 100
µl cell suspension, and the cells were incubated for 40 min
in the dark. All samples were evaluated by flow cytometry (BD
Biosciences; Becton, Dickinson and Company).
Statistical analysis
All data were analyzed using GraphPad Prism 6.0
(GraphPad Software, Inc., San Diego, CA, USA). The results are
presented as mean ± standard deviation of at least three
independent experiments. The data were statistically analyzed by
Student’s t-test for comparing two groups and one-way analysis of
variance for comparing multiple groups. Dunnett and Tukey methods
were used as post-hoc tests. Survival curves were plotted by the
Kaplan-Meier method and compared using the log-rank test.
Correlations between two indices were analyzed using Spearman’s
rank correlation analysis. P<0.05 was considered to indicate a
statistically significant difference.
Results
miR-300 suppresses cell proliferation and
induces apoptosis and cell cycle arrest in HCC cells
The expression of miR-300 was examined in the normal
hepatic cell line L02 and the HCC cell lines, Huh-7, Li-7, Hep3B
and SNU-449, by RT-qPCR. The results demonstrated that miR-300
expression was decreased in all four HCC cell lines compared with
in L02 cells (Fig. 1A). Based on
these results, the effects of miR-300 on the proliferation and
apoptosis of HCC cells were investigated by up- and downregulating
miR-300 expression using miR-300 mimic and inhibitor, respectively
(Fig. 1B). The MTT and colony
formation assays were performed to analyze the effect of miR-300 on
cell proliferation. The results demonstrated that cell viability
was suppressed in miR-300 mimic-transfected Huh-7 cells and
enhanced in miR-300 inhibitor-transfected Hep3B cells compared with
that in negative control cells on days 2, 3, 4 and 5 (Fig. 1C). Furthermore, the colony
formation was significantly decreased in Huh-7 cells transfected
with miR-300 mimic, whereas it was increased in Hep3B cells
transfected with miR-300 inhibitor compared with the respective
controls (Fig. 1D).
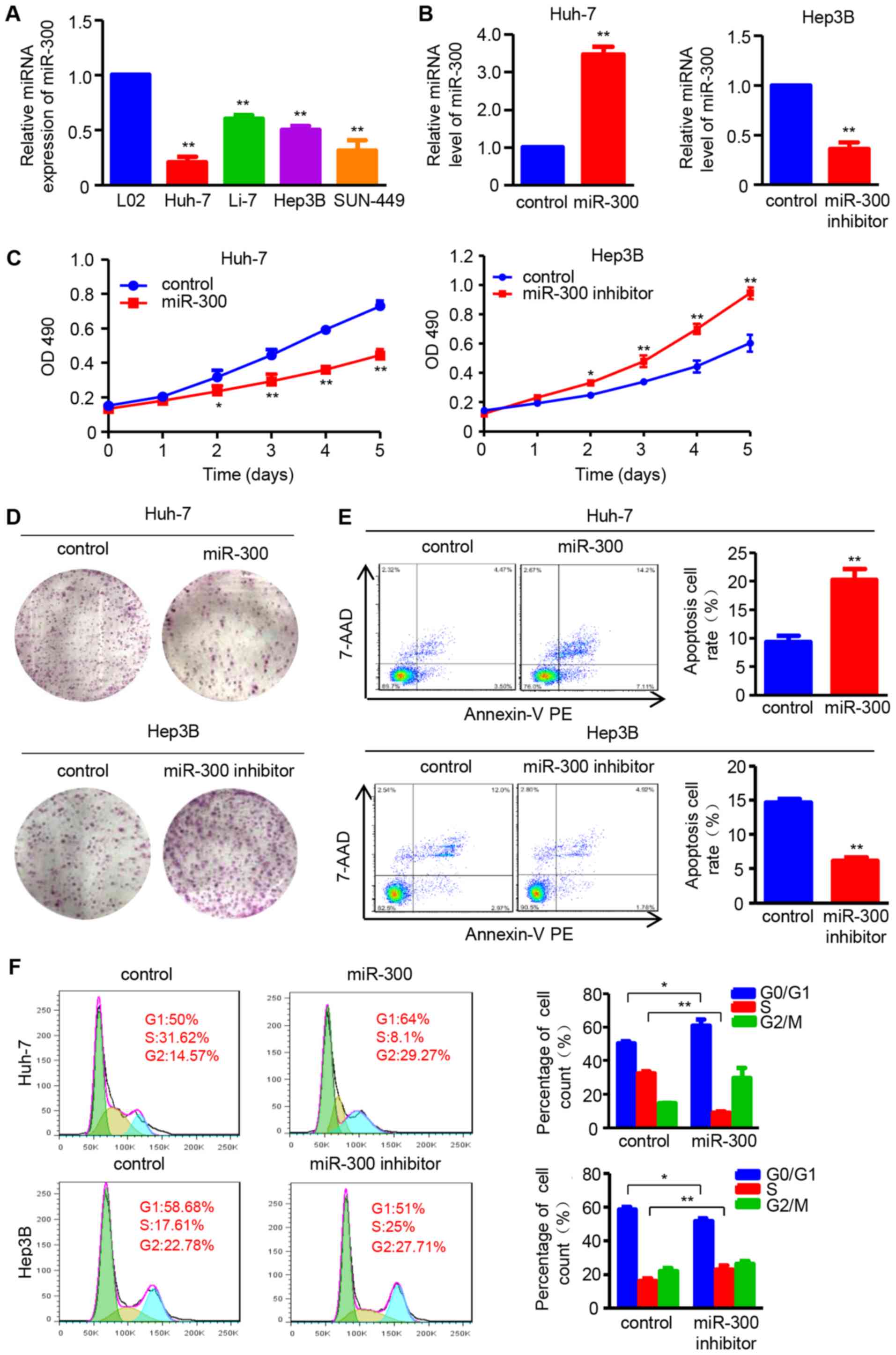 | Figure 1miR-300 suppresses cell proliferation,
and induces apoptosis and cell cycle arrest in HCC cells. (A)
RT-qPCR analysis was used to detect the miR-300 mRNA expression in
the HCC cell lines Huh-7, Li-7, Hep3B and SNU-449, and the normal
hepatic cell line L02 (n=3; **P<0.01 vs. L02). (B)
Expression of miR-300 mRNA was analyzed by RT-qPCR after
transfection with miR-300 mimic (miR-300) or miR-300 inhibitor in
Huh-7 or Hep3B cells, respectively (n=3; **P<0.01 vs.
control). (C) MTT assay was performed to detect the proliferation
of Huh-7 and Hep3B cells transfected with miR-300 mimic (miR-300)
or miR-300 inhibitor, respectively (n=3; *P<0.05,
**P<0.01 vs. control). (D) Colony formation ability
was assessed using colony formation assay; representative images
are shown. (E) Apoptosis rate of Huh-7 and Hep3B cell lines
following transfection with miR-300 mimic (miR-300) or miR-300
inhibitor was examined by flow cytometric analysis (n=3;
**P<0.01 vs. control). (F) DNA content was analyzed
in Huh-7 and Hep3B cells transfected with miR-300 mimic (miR-300)
or miR-300 inhibitor using flow cytometry, and the percentage of
cells in the G0/G1, S and G2/M phases of the cell cycle was
calculated (n=3; **P<0.01 vs. control). HCC,
hepatocellular carcinoma; RT-qPCR, reverse
transcription-quantitative polymerase chain reaction; miR,
microRNA; OD, optical density; 7-AAD, 7-aminoactinomycin D; PE,
phycoerythrin. |
Furthermore, the effect of miR-300 on HCC cell
apoptosis was investigated. The number of apoptotic cells was
increased in miR-300 mimic-transfected Huh-7 cells, whereas it was
decreased in miR-300 inhibitor-transfected Hep3B cells (Fig. 1E). In addition, flow cytometry was
applied to evaluate cell cycle distribution. The results
demonstrated that miR-300 overexpression induced G1/S cell cycle
arrest, as reflected by an increased percentage of cells in the
G0/G1 phase and a decreased percentage of cells in the S phase in
miR-300-overexpressing Huh-7 cells. By contrast, number of cells in
the S phase was increased by miR-300 downregulation in Hep3B cells
transfected with miR-300 inhibitor (Fig. 1F). These data indicate that miR-300
suppresses cell proliferation, and promotes apoptosis and G1/S cell
cycle arrest in HCC cells.
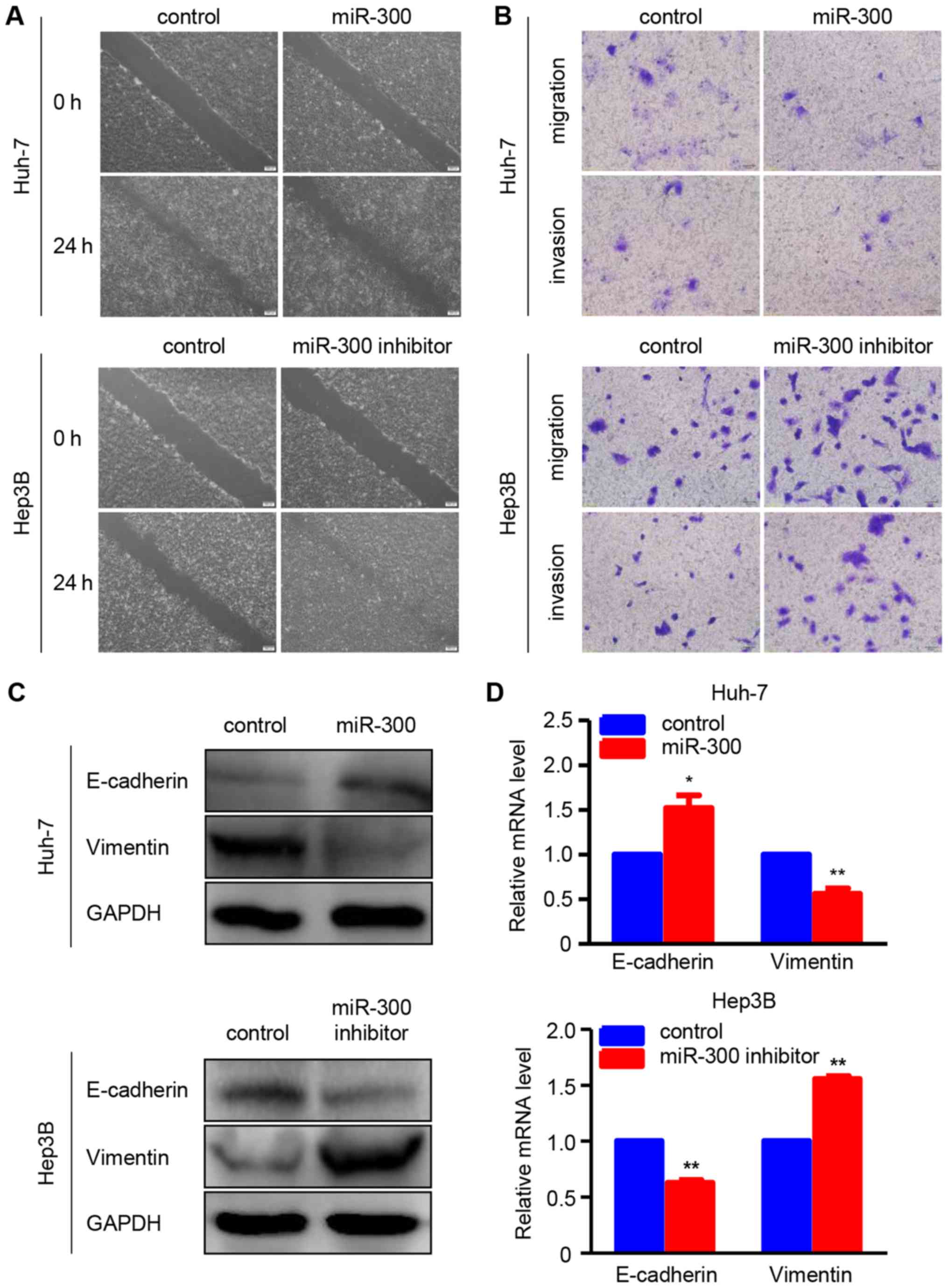
miR-300 suppresses migration, invasion
and EMT in HCC cells
To determine whether miR-300 is associated with HCC
metastasis, wound healing and Transwell assays were performed to
examine the effects of miR-300 on the migration and invasion
abilities of HCC cells. As shown in the wound healing assay, the
migration ability of Huh-7 cells was suppressed by miR-300
overexpression, whereas miR-300 downregulation increased the
migratory ability of Hep3B cells (Fig.
2A). In addition, the results of the Transwell migration assay
were consistent with those of the wound healing assay in Huh-7 and
Hep3B cells. Furthermore, the Transwell Matrigel invasion assay
demonstrated that the invasion ability was suppressed in
miR-300-overexpressing Huh-7 cells and enhanced in miR-300-silenced
Hep3B cells compared wit the respective controls (Fig. 2B). Additionally, the protein and
mRNA expressions of EMT-associated markers, including E-cadherin
and vimentin, were investigated using western blot and RT-qPCR
analyses, respectively. The data revealed that miR-300
overexpression markedly increased the protein and mRNA levels of
the epithelial marker E-cadherin, whereas the protein and mRNA
levels of the mesenchymal marker vimentin were significantly
reduced in Huh-7 cells. As expected, the protein and mRNA
expressions of E-cadherin and vimentin exhibited the opposite trend
in Hep3B cells transfected with miR-300 inhibitor (Fig. 2C and D). All these data confirmed
that miR-300 inhibits the migration, invasion and EMT of HCC
cells.
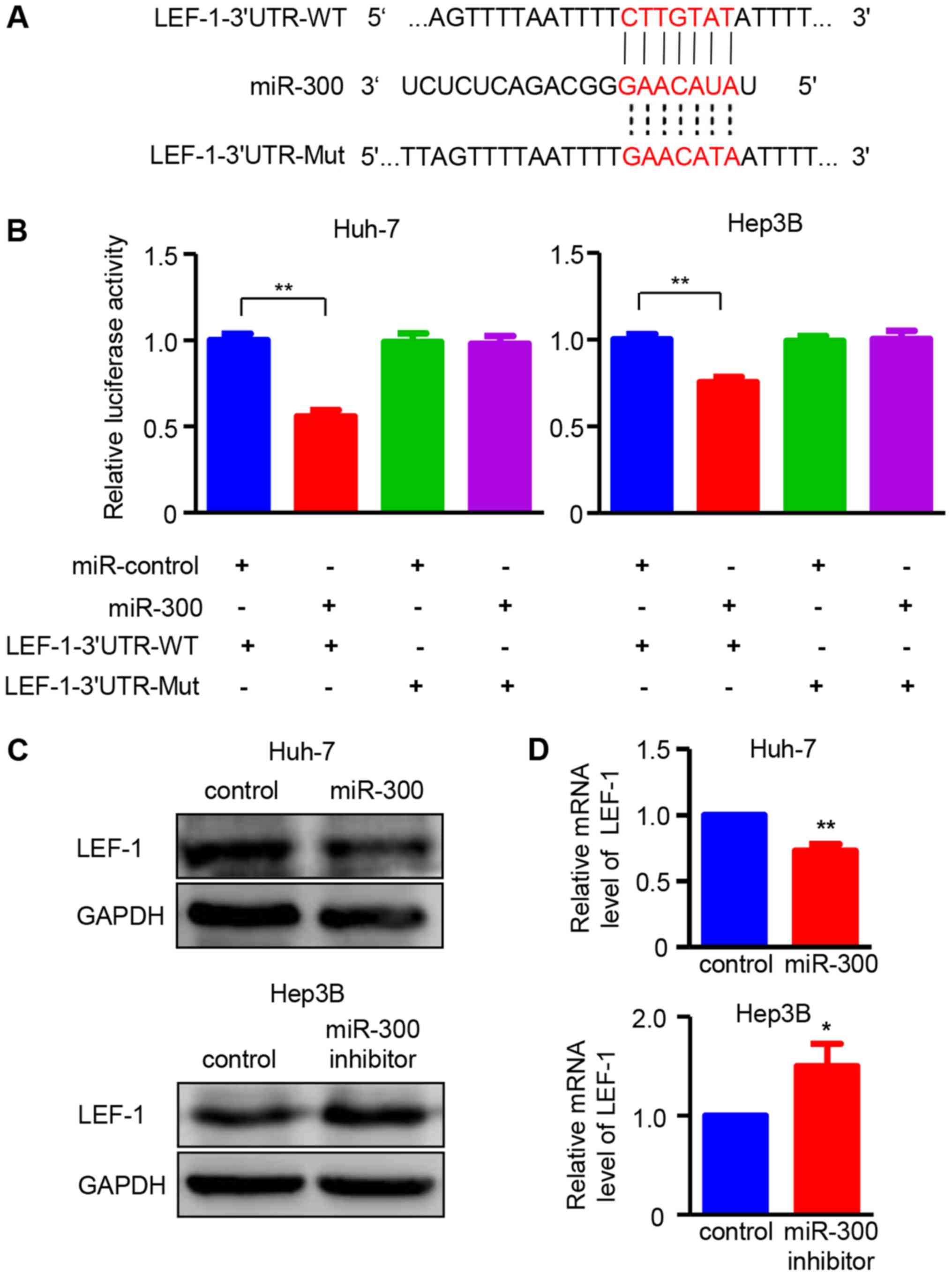
LEF-1 is directly targeted by miR-300 in
HCC
To elucidate the molecular mechanism underlying the
effects of miR-300 on HCC cells, miR-300-targeted transcription
factors were predicted using target prediction programs, including
miRanda and miRDB. LEF-1 was identified as a potential target gene
of miR-300, as the 3′UTR region of LEF-1 contained a complementary
site for the seed region of miR-300 (Fig. 3A). To confirm this result, LEF-1
3′UTR and the mutant binding sequences were designed for use in a
luciferase reporter assay. As demonstrated in Fig. 3B, miR-300 significantly reduced the
relative luciferase activity in Huh-7 and Hep3B cells transfected
with the wild-type LEF-1 3′UTR reported, indicating that miR-300
directly binds to the 3′UTR of LEF-1. Western blot and RT-PCR
analyses were performed to verify the effect of miR-300 on LEF-1.
The results demonstrated that overexpression of miR-300
significantly decreased the protein and mRNA expressions of LEF-1
in Huh-7 cells, whereas miR-300 inhibitor-transfected Hep3B cells
exhibited increased LEF-1 expression at both the protein and mRNA
level (Fig. 3C and D). Taken
together, these results confirm that LEF-1 is a direct target gene
of miR-300 in HCC cells.
LEF-1 promotes the proliferation,
migration and invasion of HCC cells
The role of LEF-1 in HCC cell proliferation,
apoptosis and cell cycle arrest was investigated prior to
determining whether LEF-1 mediated the regulatory effect of miR-300
on HCC cells. Contrary to miR-300 expression, the data demonstrated
that LEF-1 protein and mRNA expression was increased in HCC cell
lines (Huh-7, Hep3B and SNU-449) compared with that in the L02 cell
line (Fig. 4A and B). Based on
these data, the LV-LEF-1 vector and LEF-1 shRNA (shRNA1, shRNA2 and
shRNA3) were designed and then transfected into Huh-7 or Hep3B
cells to overexpress or silence LEF-1, respectively. Western
blotting and RT-qPCR analysis demonstrated that LEF-1 mRNA and
protein expression was upregulated by LV-LEF-1 in Huh-7 cells,
whereas LEF-1 shRNA significantly decreased LEF-1 expression in
Hep3B cells (Fig. 4C and D). MTT
and colony formation assays were then performed, and the results
suggested that cell viability and proliferation ability were
enhanced by LEF-1 overexpression in Huh-7 cells and suppressed by
LEF-1 silencing in Hep3B cells (Fig.
4E and F). Furthermore, the apoptosis rate was significantly
increased in LEF-1 shRNA-transfected Hep3B cells, whereas
Huh-7-LV-LEF-1 resulted in decreased cell apoptosis rate (Fig. 4G). In addition, as shown in
Fig. 4H, the knockdown of LEF-1
significantly decreased the percentage of cells in the S phase and
increased the percentage of cells in the G0/G1 phase of the cell
cycle. By contrast, increased LEF-1 overexpression increased the
percentage of cells in the S phase and decreased that in the G0/G1
phase.
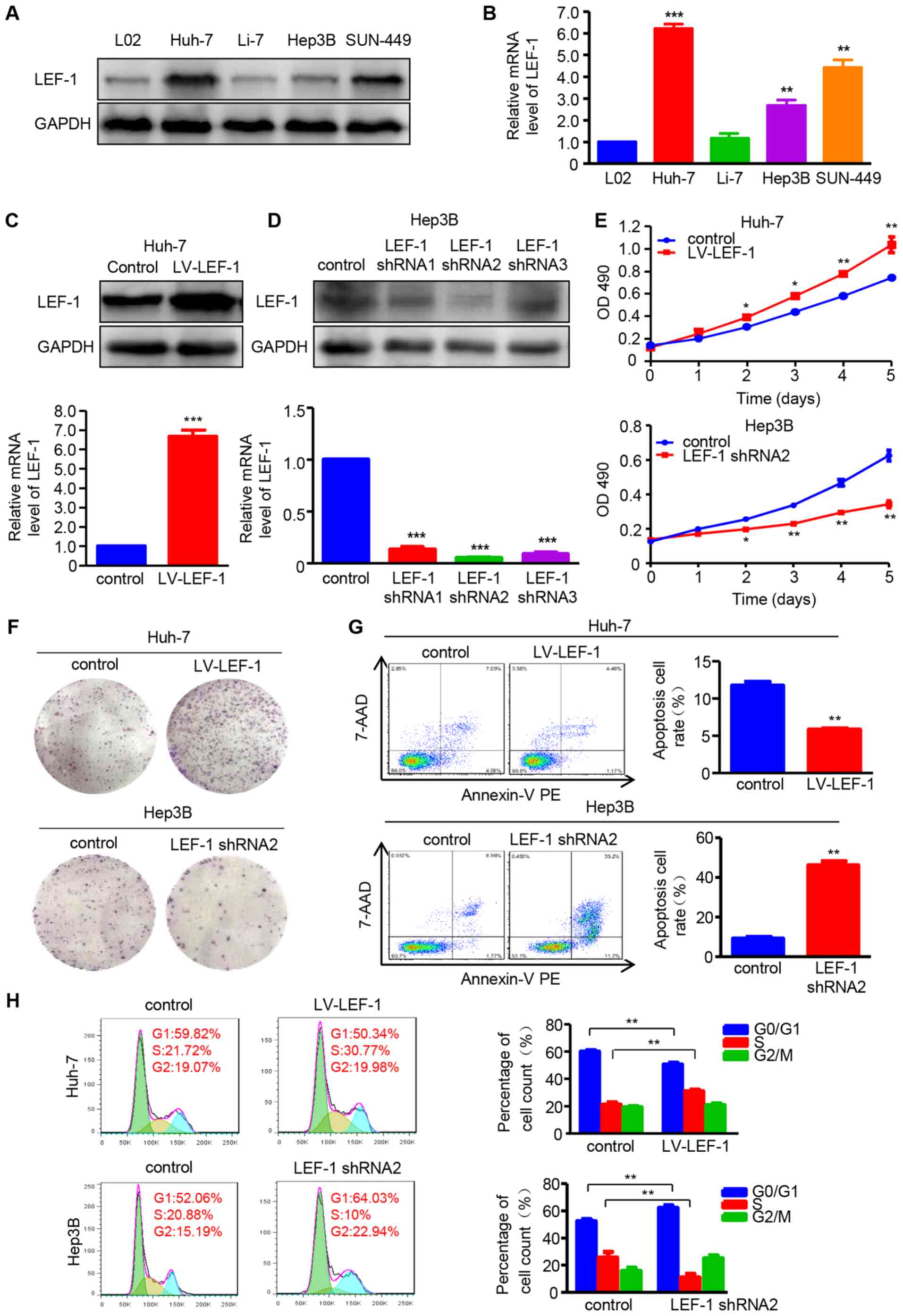 | Figure 4LEF-1 enhances cell proliferation and
inhibits apoptosis and cell cycle arrest in HCC cells. (A) Western
blotting was used to detect the LEF-1 protein expression in the HCC
cell lines Huh-7, Li-7, Hep3B and SNU-449, and the normal hepatic
cell line L02. (B) RT-qPCR was used to detect the LEF-1 mRNA
expression in the HCC cell lines Huh-7, Li-7, Hep3B and SUN-449,
and the normal hepatic cell line L02 (n=3; **P<0.01,
***P<0.001 vs. L02). (C) The protein and mRNA
expression of LEF-1 was evaluated in the Huh-7 cells transfected
with LV-LEF-1 using western blotting and RT-qPCR analysis,
respectively (n=3; ***P<0.001 vs. control). (D) The
protein and mRNA expression of LEF-1 was detected in the Hep3B
cells transfected with three LEF-1 shRNAs using western blotting
and RT-qPCR analysis, respectively (n=3; ***P<0.001
vs. control). (E) MTT assay was performed to detect the
proliferation of Huh-7 cells transfected with LV-LEF-1 and Hep3B
cells transfected with LEF-1 shRNA (n=3; *P<0.05,
**P<0.01 vs. control). (F) The colony-forming ability
was assessed using the colony formation assay. Representative
images are shown. (G) Flow cytometric analysis was used to observe
the apoptosis rate of Huh-7 and Hep3B cells following transfection
with LV-LEF-1 or LEF-1 shRNA, respectively (n=3;
**P<0.01 vs. control). (H) The DNA content was
analyzed in Huh-7 and Hep3B cells transfected with LV-LEF-1 or
LEF-1 shRNA using flow cytometry, and the percentage of cells in
the G0/G1, S and G2/M phases of the cell cycle was calculated (n=3;
**P<0.01). HCC, hepatocellular carcinoma; RT-qPCR,
reverse transcription-quantitative polymerase chain reaction;
LEF-1, lymphoid enhancer-binding factor 1; LV, lentivirus; shRNA,
short hairpin RNA: 7-AAD, 7-aminoactinomycin D; PE,
phycoerythrin. |
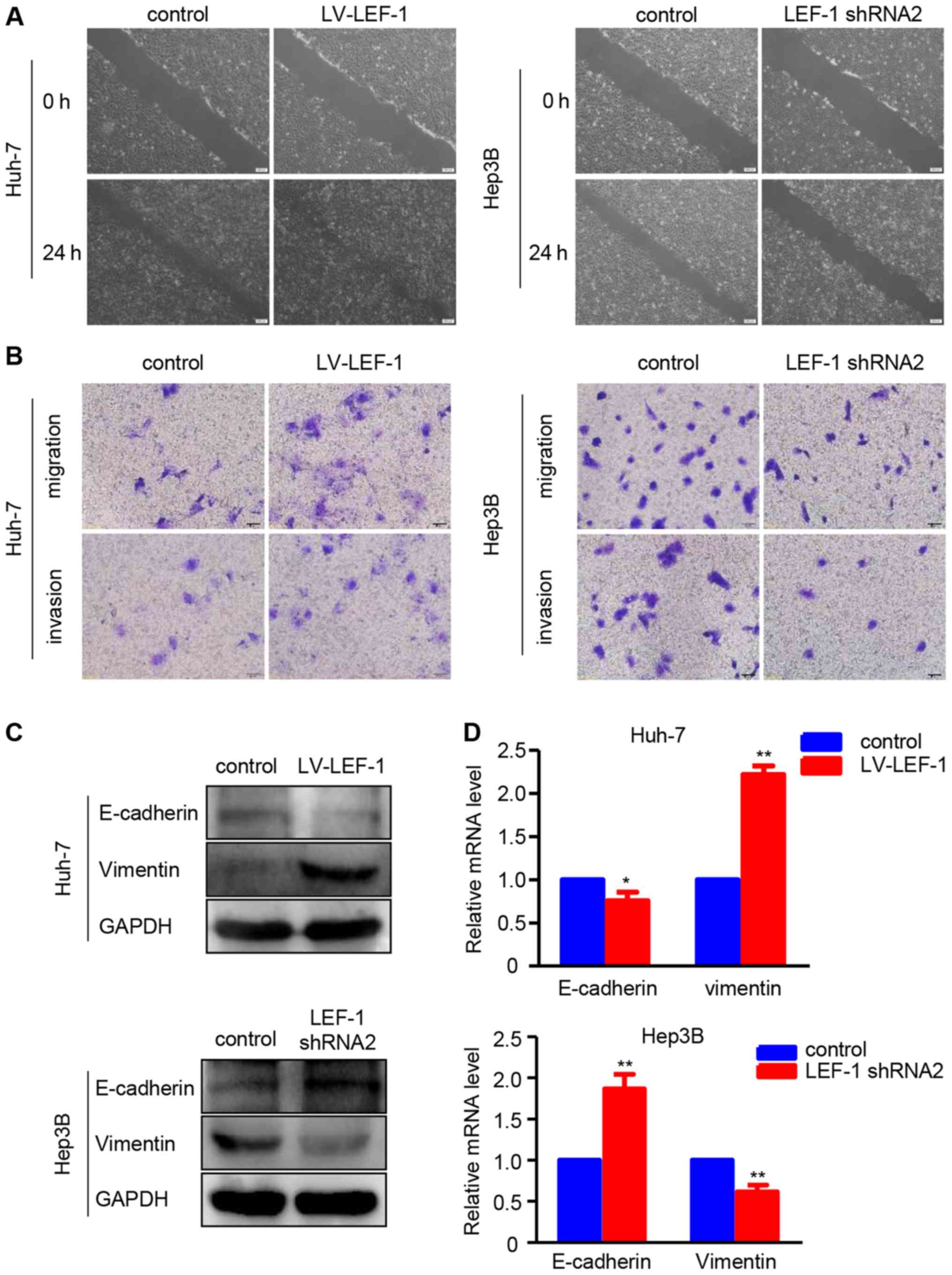
Furthermore, the association of LEF-1 with the
regulation of migration and invasion in HCC cells was investigated.
The wound healing assay revealed that the LEF-1 shRNA-transfected
Hep3B cells exhibited a slower wound closure rate compared with the
control group, whereas the LV-LEF-1-transfected Huh-7 cells
exhibited an increased migratory ability compared with the control
group (Fig. 5A). The migration and
invasion assays demonstrated that LEF-1 overexpression increased
the migration and invasion abilities of Huh7 cells, whereas LEF-1
knockdown decreased the migration and invasion abilities of Hep3B
cells (Fig. 5B). Western blotting
and RT-PCR analysis revealed that LEF-1 knockdown decreased the
protein and mRNA levels of vimentin, but increased the protein and
mRNA levels of E-cadherin in Hep3B cells. Conversely, LEF-1
overexpression increased the protein and mRNA levels of vimentin
but decreased those of E-cadherin in Huh-7 (Fig. 5C and D).
miR-300 affects proliferation, migration
and invasion by regulating LEF-1 expression in HCC cells
To determine whether miR-300 regulates cell
proliferation and metastasis by targeting LEF-1, the LEF-1 vector
and miR-300 mimic/LEF-1 shRNA and miR-300 inhibitor were
co-transfected into Huh-7 or Hep3B cells, followed by evaluation of
cell proliferation, migration and invasion abilities. The data
demonstrated that the inhibitory effect of miR-300 mimic on cell
viability and proliferation was partially rescued by the LEF-1
vector. In addition, LEF-1 shRNA suppressed the miR-300
inhibitor-induced cell growth (Fig. 6A
and B). Similar to these results, LEF-1 and LEF-1 shRNA
reversed the effects of miR-300 mimic and inhibitor on cell
apoptosis and the cell cycle, respectively (Fig. 6C and D).
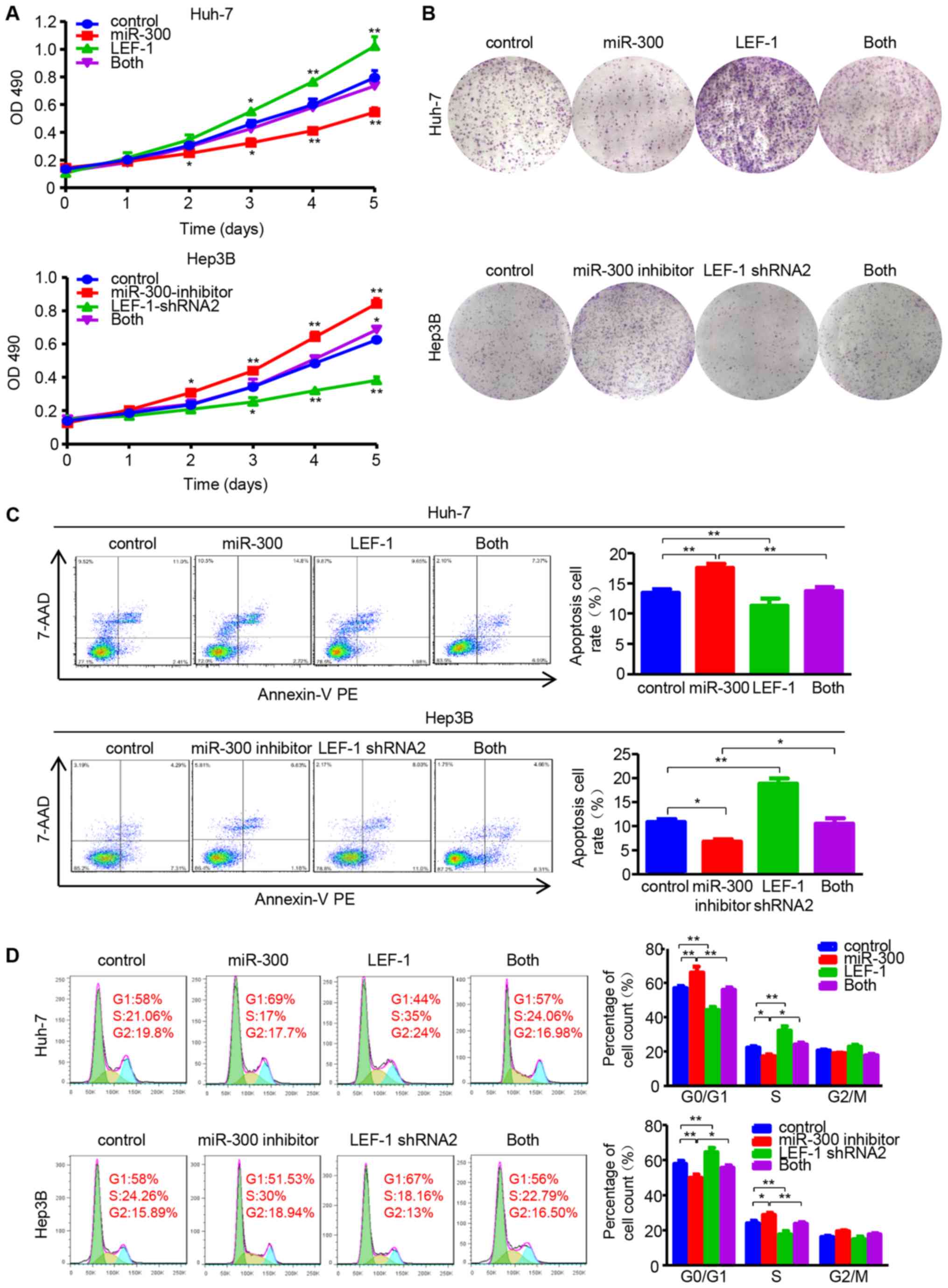 | Figure 6miR-300 impacts on proliferation,
apoptosis and cell cycle via regulating LEF-1 expression in
hepatocellular carcinoma cells. (A) MTT assay was performed to
detect the proliferation in Huh-7 cells co-transfected with LEF-1
vector (LEF-1) and miR-300 mimic (miR-300), as well as Hep3B cells
co-transfected with LEF-1 shRNA and miR-300 inhibitor (n=3;
*P<0.05, **P<0.01 vs. control). (B) The
colony-forming ability was assessed using the colony formation
assay. Representative images are shown. (C) The apoptosis rate was
evaluated in Huh-7 and Hep3B cells by flow cytometric analysis
following co-transfection with LEF-1 vector and miR-300/LEF-1 shRNA
and miR-300 inhibitor (n=3; *P<0.05,
**P<0.01). (D) The DNA content was analyzed using
flow cytometry, and the percentage of cells in the G0/G1, S and
G2/M phases of the cell cycle was calculated (n=3;
*P<0.05, **P<0.01). OD, optical
density; miR, microRNA; LEF-1, lymphoid enhancer-binding factor 1;
shRNA, short hairpin RNA; 7-AAD, 7-aminoactinomycin D; PE,
phycoerythrin. |
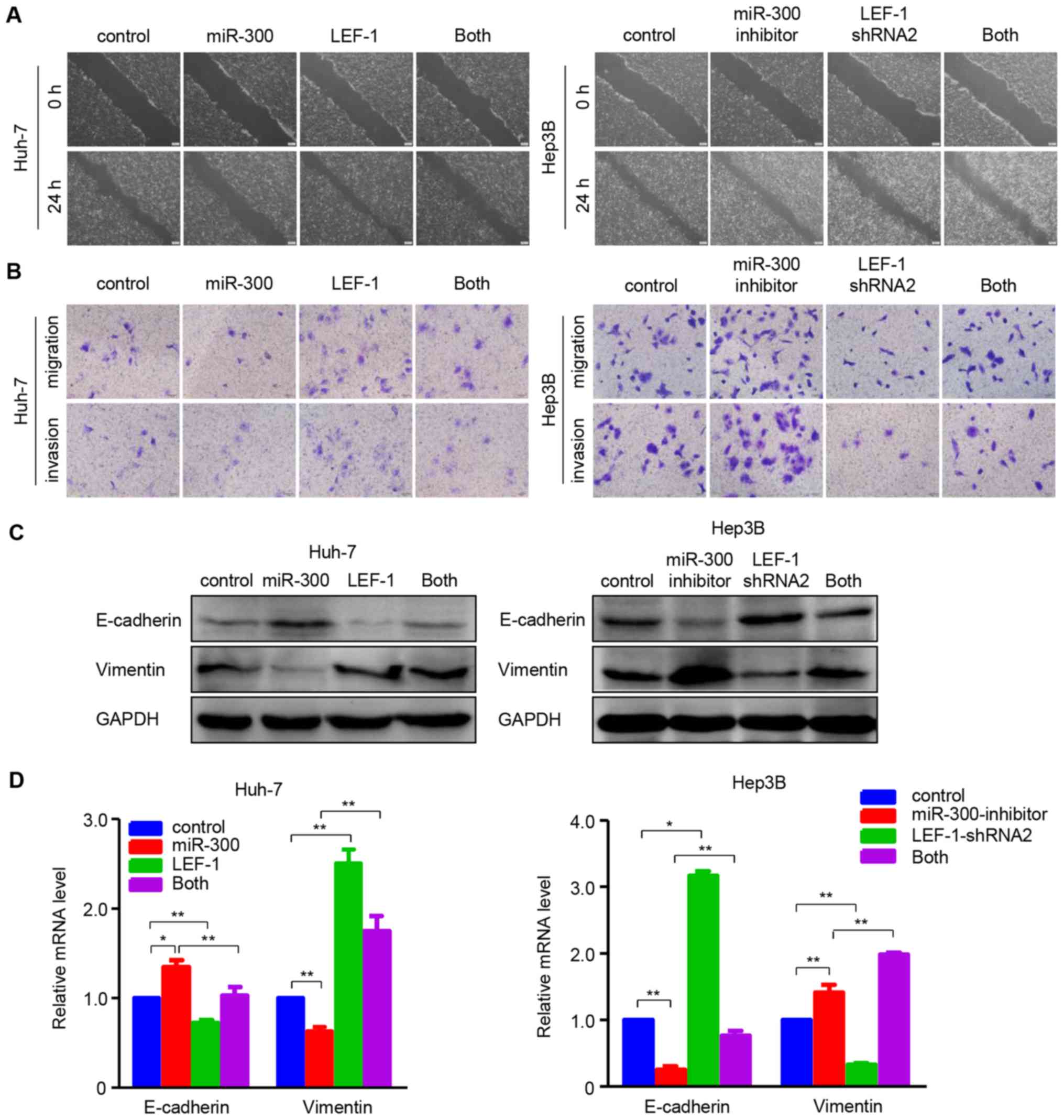
Furthermore, functional experiments were performed
to determine whether LEF-1 mediated the effects of miR-300 on the
migration and invasion in HCC cells. The wound healing and
Transwell assays indicated that migration and invasion were
inhibited in Huh-7 cells overexpressing miR-300, whereas they were
restored when miR-300 mimic and LEF-1 vector were co-transfected
into the cells. In the Hep3B cell line, miR-300 silencing-induced
migration and invasion were inhibited by transfecting LEF-1 shRNA
(Fig. 7A and B). In addition, the
effects of miR-300 mimic/inhibitor on the mRNA and protein
expression of EMT-associated factors were partially reversed by up-
or downregulating LEF-1 expression using LEF-1 vector or LEF-1
shRNA, as shown by western blotting and RT-qPCR analysis (Fig. 7C and D). These data indicate that
overexpressing/silencing miR-300 may suppress/promote cell
proliferation, migration and invasion through down-/upregu-lating
LEF-1 expression in HCC cells.
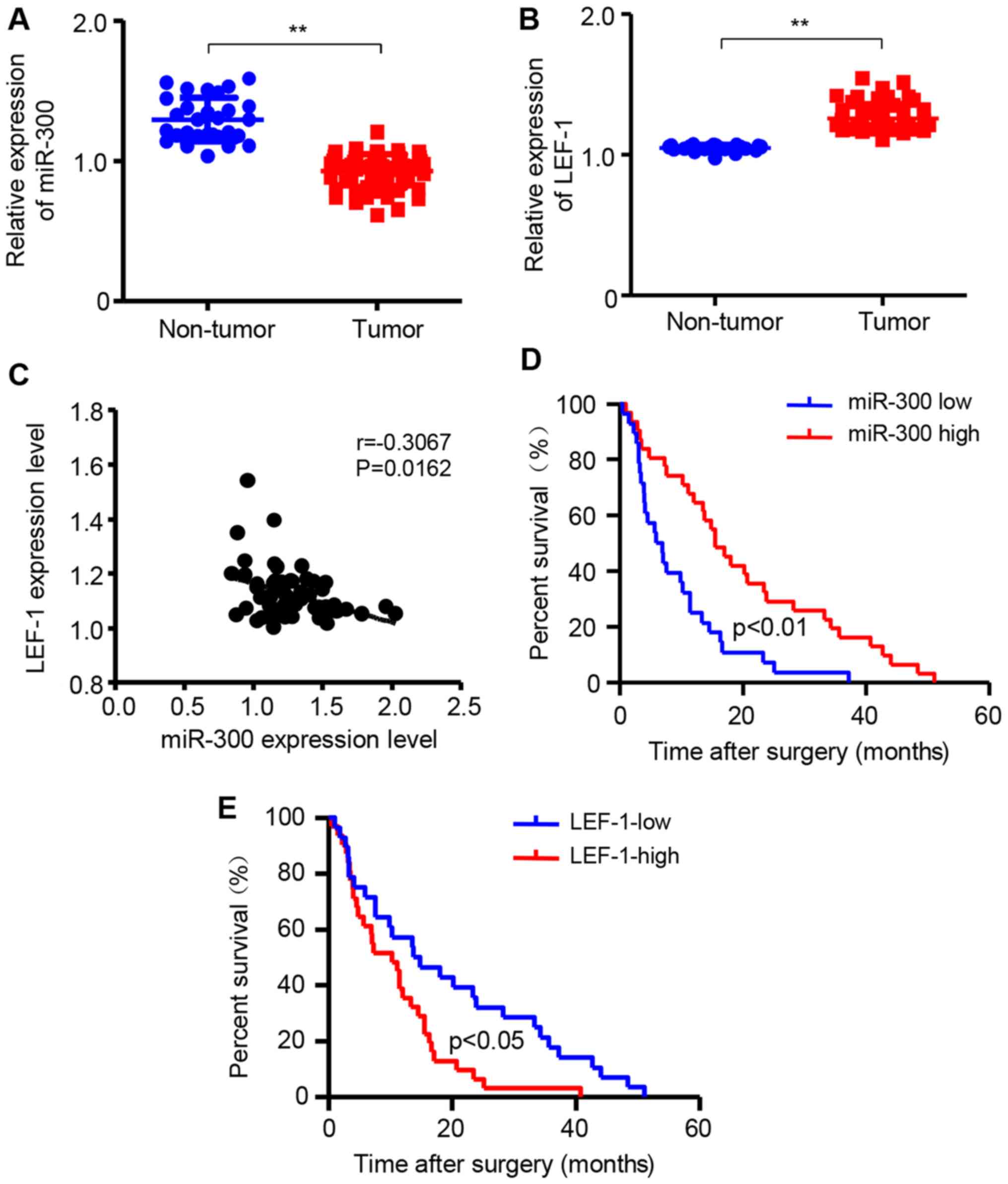
miR-300 is downregulated while LEF-1 is
upregulated in HCC tissues
To further confirm that miR-300 and LEF-1 are
associated with HCC development, the mRNA expression of miR-300 and
LEF-1 was detected in HCC tissues and non-tumor liver tissues using
RT-qPCR assay. The 62 patients with HCC were divided into two
groups according to the median value of miR-300 or LEF-1
expression. The results demonstrated that miR-300 expression in HCC
tissues was significantly decreased compared with that in non-tumor
liver tissues (Fig. 8A). However,
the mRNA expression of LEF-1 in HCC tissues was increased compared
with that in non-tumor liver tissues (Fig. 8B). Additionally, the association
between LEF-1 and miR-300 was investigated in patients with HCC and
the expression level of LEF-1 was negatively correlated with the
level of miR-300 mRNA (Fig. 8C).
In addition, the association of miR-300 and LEF-1 with the overall
survival time was investigated by Kaplan-Meier survival analysis.
As shown in Fig. 8D and E,
patients with HCC with low miR-300 levels had significantly shorter
overall survival compared with those with high miR-300 levels. By
contrast, the LEF-1 expression level was negatively associated with
overall survival in patients with HCC. Based on those data, it may
be concluded that low expression of miR-300 and high expression of
LEF-1 in HCC tissues are associated with poor prognosis of patients
with HCC, and that LEF-1 is a downstream target of miR-300 in
HCC.
Discussion
miR-300 was recently reported to be involved in the
development and metastasis of different types of cancer (20,21).
However, the effects of miR-300 on tumor cells and the underlying
mechanism(s) remain unclear, particularly in HCC. The aim of the
present study was to elucidate whether miR-300 affected the
biological behavior of cancer cells and the possible mechanism of
action. As demonstrated by the findings, miR-300 expression was low
in HCC tissues and cell lines. Furthermore, miR-300 suppressed the
proliferation, migration and invasion of HCC cells. In addition,
LEF-1, which promotes the malignant behavior of HCC cells, was
confirmed to mediate the effects of miR-300. Clinically, it was
also observed that low miR-300 expression and high LEF-1 expression
in HCC tissues were both associated with poor prognosis of patients
with HCC. These finding suggest that miR-300 may be a potential
treatment target and prognostic factor for HCC.
As mentioned above, accumulating evidence indicates
that miR-300 is involved in the development and metastasis of
several types of tumors; however, miR-300 may exert opposing
effects on distinct types of human cancer. Zhou et al
(22) demonstrated that miR-300
was significantly downregulated in glioblastoma tissues and cells
(U87 and U251), and that the overexpression of miR-300 could
suppress cell development and progression in vitro and in vivo,
which was partially rescued by inhibiting Rho-associated protein
kinase 1 expression. Similar to these results, Yu et al
(23) confirmed that miR-300
inhibited cell invasion and metastasis by downregulating
Twist-mediated EMT in human epithelial cancers. However, other
studies demonstrated that miR-300 could promote cell growth in
certain cancers. A previous study indicated that miR-300
upregulation in human gastric cancer tissues and cells promoted
gastric cancer cell proliferation and invasion by targeting p53
(21). Xue et al (24) revealed that miR-300 acted as an
oncogene in osteosarcoma, and demonstrated that increased
expression of miR-300 promoted cell proliferation, invasion and EMT
by suppressing bromodomain-containing protein 7; this discrepancy
was attributed to differences in the tumor microenvironment. Only
few studies on miR-300 have been reported in HCC, and only one
study indicated that miR-300 was decreased in HCC and that this
decrease was significantly associated with poor prognosis of
patients with HCC (15). Similar
to these results, miR-300 was downregulated in HCC tissues and
cells, whereas miR-300 upregulation suppressed HCC cell growth,
migration and invasion. These findings confirm that miR-300 is
closely associated with the development and progression of HCC.
Most importantly, to the best of our knowledge, this
study was the first to demonstrate that LEF-1 is a target gene of
miR-300 and that it mediates the effects of miR-300 on HCC cell
proliferation and metastasis. miR-300 has been confirmed to
regulate cancer cell behavior by targeting downstream genes.
Several target genes of miR-300 have been confirmed to date. For
example, miR-300 was able to inhibit lung squamous cell carcinoma
cell proliferation and invasion by targeting ROS1 (25). Furthermore, through targeting p53,
miR-300 promotes the proliferation and EMT-mediated colorectal
cancer cell migration and invasion, and desensitizes lung cancer
cells to ionizing radiation by suppressing p53-dependent G2 cell
cycle arrest, apoptosis and senescence (12,13).
Notably, the present study identified a novel miR-300-targeted
transcription factor, LEF-1, which has been confirmed to be a
direct target gene of miR-300 by bioinformatics prediction and
luciferase reporter assay. Furthermore, LEF-1 was revealed to be
involved in the process of miR-300-mediated regulation of HCC cell
behavior. These data provide a new direction for treatment of
patients with HCC.
LEF-1 has been identified to be involved in the
progression of HCC. A recent study demonstrated that Oct 4 promoted
EMT by upregulating LEF-1 expression, which induced activation of
β-catenin-dependent Wnt signaling (26). Another study demonstrated that
LEF-1 binds the promoter of miR-HCC1 and activates its expression,
thus contributing to the proliferation, migration and invasion of
HCC cells (27). Similar results
were observed in our study, with LEF-1 downregulated by miR-300,
affecting the ability of proliferation and metastasis of HCC cells.
The factors affecting the LEF-1 level in patients with HCC remain
unclear. Hepatitis B virus (HBV) may induce LEF-1-mediated HCC cell
growth. Tian et al (28)
reported that the expression of LEF-1 was increased in
HBsAg-positive HCC tissues, and hypothesized that HBsAg may
stimulate the proliferation and functional modification of
hepatocytes via LEF-1. A later study also revealed that HBV
promoted the proliferation and metastasis of HCC cells through the
LEF-1/miR-371a-5p/SRCIN1/pleiotrophin/Slug pathway (18). However, whether miR-300 and LEF-1
regulation of HCC progression is associated with HBV requires
further investigation.
In summary, miR-300 was found to be frequently
downregulated in HCC tissues and cells, which was significantly
associated with poor patient prognosis. In addition, miR-300
overexpression inhibited the proliferation, migration and invasion
of HCC cells. Furthermore, LEF-1 was identified as a direct target
of miR-300, and mediated the effects of miR-300 on HCC cells.
Collectively, these results suggest that miR-300 may represent a
novel biomarker and promising therapeutic target for patients with
HCC.
Funding
The present study was supported by grants from the
National Natural Science Foundation of China (grant no. 81572458),
the University Outstanding Youth Talent Support Program of Anhui
Province (grant no. gxyqZD2017065) and the Program for Graduate
Research of Bengbu Medical College (grant no. Byycx1753).
Availability of data and materials
The datasets generated and analyzed during the
present study are available from the corresponding author upon
reasonable request.
Authors’ contributions
RW and QW participated in the design of the study,
data interpretation and drafting of the manuscript. YC and JY
performed the experiments. YG drafted the manuscript, and
participated in analysis and interpretation of data. YL and JL
participated in the clinical sample collection. All authors have
read and approved the manuscript and agree to be accountable for
all aspects of the research in ensuring that the accuracy or
integrity of any part of the work are appropriately investigated
and resolved.
Ethics approval and consent to
participate
The human tissue protocol utilized in this study was
approved by the Ethics Committee of Bengbu Medical College (Bengbu,
China). Informed consent was obtained from each patient registered
in the study, in accordance with the institutional guidelines.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mileo AM, Mattarocci S, Matarrese P,
Anticoli S, Abbruzzese C, Catone S, Sacco R, Paggi MG and Ruggieri
A: Hepatitis C virus core protein modulates pRb2/p130 expression in
human hepatocellular carcinoma cell lines through promoter
methylation. J Exp Clin Cancer Res. 34:1402015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lombardi G, Perego S, Sansoni V and Banfi
G: Circulating miRNA as fine regulators of the physiological
responses to physical activity: Pre-analytical warnings for a novel
class of biomarkers. Clin Biochem. 49:1331–1339. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zamore PD and Haley B: Ribo-gnome: The big
world of small RNAs. Science. 309:1519–1524. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhou SL, Hu ZQ, Zhou ZJ, Dai Z, Wang Z,
Cao Y, Fan J, Huang XW and Zhou J: miR-28-5p-IL-34-macrophage
feedback loop modulates hepatocellular carcinoma metastasis.
Hepatology. 63:1560–1575. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kumar A: MicroRNA in HCV infection and
liver cancer. Biochim Biophys Acta. 1809.694–699. 2011.
|
|
8
|
Ji J, Shi J, Budhu A, Yu Z, Forgues M,
Roessler S, Ambs S, Chen Y, Meltzer PS, Croce CM, et al: MicroRNA
expression, survival, and response to interferon in liver cancer. N
Engl J Med. 361:1437–1447. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tang H, Zhang J, Yu Z, Ye L, Li K, Ding F,
Feng X and Meng W: Mir-452-3p: A Potential Tumor Promoter That
Targets the CPEB3/EGFR Axis in Human Hepatocellular Carcinoma.
Technol Cancer Res Treat. 16:1136–1149. 2017. View Article : Google Scholar
|
|
10
|
Cao J, Qiu J, Wang X, Lu Z, Wang D, Feng
H, Li X, Liu Q, Pan H, Han X, et al: Identification of microRNA-124
in regulation of Hepatocellular carcinoma through BIRC3 and the
NF-κB pathway. J Cancer. 9:3006–3015. 2018. View Article : Google Scholar :
|
|
11
|
He C, Liu Z, Jin L, Zhang F, Peng X, Xiao
Y, Wang X, Lyu Q and Cai X: lncRNA TUG1-Mediated Mir-142-3p
Downregulation Contributes to Metastasis and the
Epithelial-to-Mesenchymal Transition of Hepatocellular Carcinoma by
Targeting ZEB1. Cell Physiol Biochem. 48:1928–1941. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang L and Yu P: miR-300 promotes
proliferation and EMT-mediated colorectal cancer migration and
invasion by targeting p53. Oncol Rep. 36:3225–3232. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
He J, Feng X, Hua J, Wei L, Lu Z, Wei W,
Cai H, Wang B, Shi W, Ding N, et al: miR-300 regulates cellular
radiosensitivity through targeting p53 and apaf1 in human lung
cancer cells. Cell Cycle. 16:1943–1953. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang JQ, Chen S, Gu JN, Zhu Y, Zhan Q,
Cheng DF, Chen H, Deng XX, Shen BY and Peng CH: MicroRNA-300
promotes apoptosis and inhibits proliferation, migration, invasion
and epithelial-mesenchymal transition via the Wnt/β-catenin
signaling pathway by targeting CUL4B in pancreatic cancer cells. J
Cell Biochem. 119:1027–1040. 2018. View Article : Google Scholar
|
|
15
|
Wang R, Yu Z, Chen F, Xu H, Shen S, Chen
W, Chen L, Su Q, Zhang L, Bi J, et al: miR-300 regulates the
epithelial-mesenchymal transition and invasion of hepatocellular
carcinoma by targeting the FAK/PI3K/AKT signaling pathway. Biomed
Pharmacother. 103:1632–1642. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shang D, Bi R, Han T, Wang D, Tian Y and
Liu Y: Expression and proliferation-promoting role of lymphoid
enhancer-binding factor 1 in human clear cell renal carcinoma.
Cancer Invest. 32:368–374. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xu S, Yang Z, Zhang J, Jiang Y, Chen Y, Li
H, Liu X, Xu D, Chen Y, Yang Y, et al: Increased levels of
β-catenin, LEF-1, and HPA-1 correlate with poor prognosis for acral
melanoma with negative BRAF and NRAS mutation in BRAF exons 11 and
15 and NRAS exons 1–2. DNA Cell Biol. 34:69–77. 2015. View Article : Google Scholar :
|
|
18
|
Bai PS, Hou P and Kong Y: Hepatitis B
virus promotes proliferation and metastasis in male Chinese
hepatocellular carcinoma patients through the
LEF-1/miR-371a-5p/SRCIN1/pleiotrophin/Slug pathway. Exp Cell Res.
370:174–188. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
20
|
Chen Z, Zhang W, Jiang K, Chen B, Wang K,
Lao L, Hou C, Wang F, Zhang C and Shen H: MicroRNA-300 regulates
the ubiquitination of PTEN through the CRL4BDCAF13 E3 ligase in
osteosarcoma cells. Mol Ther Nucleic Acids. 10:254–268. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shen Z, Li C, Zhang K, Yu W, Xiao H, Li B
and Liu T: The up-regulation of miR-300 in gastric cancer and its
effects on cells malignancy. Int J Clin Exp Med. 8:6773–6783.
2015.PubMed/NCBI
|
|
22
|
Zhou F, Li Y, Hao Z, Liu X, Chen L, Cao Y,
Liang Z, Yuan F, Liu J, Wang J, et al: MicroRNA-300 inhibited
glioblastoma progression through ROCK1. Oncotarget. 7:36529–36538.
2016.PubMed/NCBI
|
|
23
|
Yu J, Xie F, Bao X, Chen W and Xu Q:
miR-300 inhibits epithelial to mesenchymal transition and
metastasis by targeting Twist in human epithelial cancer. Mol
Cancer. 13:1212014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xue Z, Zhao J, Niu L, An G, Guo Y and Ni
L: Up-Regulation of MiR-300 promotes Proliferation and Invasion of
Osteosarcoma by Targeting BRD7. PLoS One. 10:e01276822015.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ge W, Han C, Wang J and Zhang Y: MiR-300
suppresses laryngeal squamous cell carcinoma proliferation and
metastasis by targeting ROS1. Am J Transl Res. 8:3903–3911.
2016.PubMed/NCBI
|
|
26
|
Sun L, Liu T, Zhang S, Guo K and Liu Y:
Oct4 induces EMT through LEF1/β-catenin dependent WNT signaling
pathway in hepatocellular carcinoma. Oncol Lett. 13:2599–2606.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hu Y, Guo X, Wang J, Liu Y, Gao H, Fan H,
Nong X, Yang X, Liu M, Li S, et al: A novel microRNA identified in
hepatocellular carcinomas is responsive to LEF1 and facilitates
proliferation and epithelial-mesenchymal transition via targeting
of NFIX. Oncogenesis. 7:222018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tian X, Li J, Ma ZM, Zhao C, Wan DF and
Wen YM: Role of hepatitis B surface antigen in the development of
hepatocellular carcinoma: Regulation of lymphoid enhancer-binding
factor 1. J Exp Clin Cancer Res. 28:582009. View Article : Google Scholar : PubMed/NCBI
|