Introduction
1α,25-dihydroxyvitamin D3 [1α,25(OH)2D3],
or vitamin D, is a lipid-soluble secosteroid produced by skin
subjected to ultraviolet (UV)B radiation (1-3).
Apart from its widely known beneficial role in the regulation of
calcium homeostasis, vitamin D exerts pleiotropic effects,
including regulation of the cell cycle, proliferation,
differentiation and apoptosis (4-6). The
active forms of vitamin D are important in the protection against
DNA damage (7-9) and UVB-induced carcinogenesis in the
skin (10-15). An inverse correlation between the
concentration of vitamin D in serum and total cancer incidence and
mortality has recently been described (16), implying, that vitamin D deficiency
is a serious cancer risk factor (13,17).
An inverse correlation has also been demonstrated between the
expression of the vitamin D receptor (VDR) and 25-hydroxyvitamin D3
1-α-hydroxylase (CYP27B1) with melanoma progression and disease
outcome (15,18,19).
Therefore, active forms of vitamin D are now considered for
therapeutic use in cancer prevention and treatment, supported by
numerous epidemiological and preclinical studies (20-30).
It should be emphasized that active forms of vitamin D used in
combined therapy enhance the effectiveness of a number of
anticancer drugs, including cisplatin (31,32),
doxorubicin (33) and proton
therapy (34). A previous study
indicated that vitamin D analogs enhance the antiproliferative
activity of cisplatin on keratinocytes (35). Furthermore, vitamin D and its
analogs are currently being tested in clinical trials on various
types of cancer, including melanoma (36,37).
Melanoma, while accounting for only 4% of skin
cancers, is linked to 80% of mortalities due to skin tumors, and
therefore represents a significant public health problem (30,38-42).
This tumor is aggressive, but potentially curable by surgical
excision if it is diagnosed at the early stages of development,
including melanoma in situ or at the radial growth phase.
However, with progression of the disease to the vertical growth
phase, melanoma cells become resistant to the majority of forms of
treatment, and acquire the ability to metastasize (38,39,43).
Furthermore, the incidence of melanoma has been rising in the
Caucasian population worldwide over recent decades (38,39).
In 2017, melanoma was expected to be the fifth most common cancer
in males and sixth most common in females in the USA (44). In recent years, major progress has
been made with respect to our understanding of the molecular nature
of melanoma and the interaction of melanoma cells with the immune
system. Unfortunately, despite the marked expansion of advanced
treatment options, primary or acquired resistance develops in
patients, emphasizing the requirement for additional effort to
develop effective melanoma therapy (42,45).
The aim of the present study was to investigate the
modulation of the anticancer properties of selected anti-melanoma
chemotherapy agents by vitamin D and its non- or low-calcemic
analogs 20S-hydroxyvitamin D3 [20(OH)D3],
21-hydroxypregnacalciferol [21(OH)pD] and calcipotriol (46-50),
since the use of the hormonally active form of vitamin D,
1α,25(OH)2D3, at high doses is limited due to the risk
of toxic effects, including hypercalcemia (51,52).
Notably, 20(OH)D3 is a natural product synthesized in the human
body and detectable in human serum (53-55).
It was hypothesized that vitamin D analogs would sensitize melanoma
cells to classic chemotherapeutic drugs, based on a recent study
documenting the association between vitamin D and oxidative stress
in keratinocytes with a high proliferative potential, and the
effect of vitamin D analogs on the sensitivity of these cells to
cisplatin (35). Even though it is
known that cisplatin induces DNA damage (56), it should be noted that the
mechanism of action of cisplatin partially relies on the generation
of reactive oxygen species (ROS) (57). Therefore the effects of
dacarbazine, still used in melanoma therapy and also known to
produce ROS in cells (58), and
cisplatin, used in combination with vitamin D or its low calcemic
analogs, were tested on the human malignant melanoma A375 cell
line.
Materials and methods
Chemicals
1,25(OH)2D3, hydrogen peroxide (30%),
cisplatin and dacarbazine were Sigma-Aldrich products (Merck KGaA,
Darmstadt, Germany). 21(OH)pD was synthesized according to
Zmijewski et al (50) by ProChimia
Surfaces Sp. z o. o. (Sopot, Poland). 20(OH)D3 was synthesized and
purified as described previously (59). Calcipotriol was a gift from the
Pharmaceutical Research Institute (Warsaw, Poland).
Cell culture
Human melanoma A375 cells (CRL-1619) were purchased
from the American Type Culture Collection (Manassas, VA, USA). The
cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM)
supplemented with 10% fetal bovine serum (FBS) (both Sigma-Aldrich;
Merck KGaA) and 1% penicillin/streptomycin in an incubator with 5%
CO2 at 37°C. DMEM medium supplemented with 2%
charcoal-stripped FBS was used for all experimental procedures
where the effects of vitamin D derivatives were examined.
Proliferation assay
The sulphorhodamine B (SRB) assay was performed as
previously described (35).
Briefly, the human melanoma A375 cells were seeded in 96-well
plates (7,000 cells per well), cultured overnight and then treated
with serial dilutions of the compounds (vitamin D,
10−12-10−6 M; hydrogen peroxide, 0.004-0.250
mM; cisplatin, 0.19-300 µM; and dacarbazine, 0.15-10
µM) being tested for an additional 24 or 48 h. Following
cell fixation with 10% trichloroacetic acid for 1 h at 4°C, the
plates were washed 5 times with distilled water and air-dried.
Staining solution comprising of 0.4% SRB (Sigma-Aldrich; Merck
KGaA) in acetic acid was added to each well for 15 min, followed by
washing with 1% acetic acid. The SRB dye was solubilized using a
solution of 10 mM buffered Tris Base (pH 10.5) and the absorbance
was measured at 570 nm using an Epoch™ microplate spectrophotometer
(BioTek Instruments, Inc., Winooski, VT, USA). The relative
IC50 value was calculated as the concentration at which
half the maximum inhibition was observed, i.e., the mid-point
between no inhibition and the maximum observed decrease in
proliferation (60,61).
Cell cycle analysis
The cell cycle was analyzed by quantification of DNA
content using flow cytometry. Trypsinized cells and cells from
culture medium were fixed in 70% ethanol for 24-48 h at 4°C,
treated with ribonuclease in order to remove any contaminating RNA,
and the DNA was stained with propidium iodide (PI; Sigma-Aldrich;
Merck KGaA) for 30 min at 37°C. The fluorescence of the PI-stained
cells was measured by flow cytometry (excitation, 536 nm; emission,
617 nm; FACSCalibur™; Becton, Dickinson and Company, Franklin,
Lakes, NJ, USA). The results were analyzed using the CellQuest™ Pro
Software version 6.0 (Becton, Dickinson and Company) and expressed
as a percentage of cells with DNA content corresponding to
apoptotic/necrotic cells (subG1 fraction) or cells in
G1, S and G2/M phases of the cycle.
Measurement of changes in the
mitochondrial membrane potential (Δψm)
The detection of changes in the inner
electrochemical Δψm in living cells was performed as
described previously (35), using
the cationic, lipophilic JC-1 dye (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). Carbonyl cyanide 3-chlorophenylhydrazone (CCCP;
Sigma-Aldrich; Merck KGaA), a mitochondrial potential disrupter,
was used as a control. The melanoma A375 cells were pre-treated
with secosteroids at a concentration of 100 nM and then exposed to
2.4 and 12 µM cisplatin or 2.0 and 10 µM dacarbazine
for an additional 3 h, or to 75 nM hydrogen peroxide for 1-3 h.
Following the treatment with the selected compounds, the cells were
harvested and suspended in 1 ml PBS at room temperature. CCCP
solution in dimethylsulfoxide (DMSO) was added to the positive
control tube only (2 µM final concentration) and the cells
incubated at 37°C for 5 min. JC-1 solution (2 µM in DMSO)
was added to all tubes and the cells were incubated at 37°C for 15
min, then centrifuged at 1,000 × g for 10 min at room temperature,
and resuspended in 500 µl PBS. The samples were kept on ice
until they were analyzed on the FACSCalibur flow cytometer using
the CellQuest Pro analysis software.
Detection of intracellular ROS
production
The intracellular production of ROS was measured
using H2DCFDA (Thermo Fisher Scientific, Inc.). Cells
were incubated with 100 nM 1,25(OH)2D3 for 24 h followed
by exposure to 24 µM cisplatin or 6 µM dacarbazine
for 1 or 24 h. H2DCFDA was added to a final
concentration of 10 µM 30 min before the end of the
incubation. The cells were washed and suspended in cold PBS. The
samples were kept on ice until they were analyzed using the
FACSCalibur flow cytometer using CellQuest Pro analysis
software.
Measurement of mRNA levels
The relative mRNA levels of particular genes were
determined by a reverse transcription-quantitative polymerase chain
reaction (qPCR) assay. Total RNA was isolated using the Total RNA
Mini kit (A&A Biotechnology, Gdynia, Poland), according to the
manufacturer’s instructions. The concentration and quality of RNA
samples were determined using the Epoch spectrophotometer. A total
of 1 μg RNA was used for reverse transcription using the
RevertAid™ First Strand cDNA Synthesis kit (Thermo Fisher
Scientific, Inc.) by incubating at 42°C for 1 h. The qPCR reaction
comprised 1 µl cDNA and 150 nM of each primer (Table I), and was performed using the
SensiFAST™ SYBR No-ROX kit (Bioline Reagents Limited, London, UK)
in a total volume of 20 µl on the StepOnePlus Real-Time PCR
System (Thermo Fisher Scientific, Inc.). The thermocycling
conditions were as follows: Initial denaturation at 95°C for 2 min,
followed by 40 cycles of 95°C for 5 sec, 55-63°C for 10 sec, 72°C
for 15 sec and 79°C for 10 sec. The melting curve analysis of the
PCR products was performed following the qPCR reaction and
consisted of 95°C for 15 sec, 60°C for 1 min and 95°C for 15 sec.
The reactions were run in duplicate and the resulting data were
averaged prior to analysis with the StepOnePlus version 2.2.2.
software (Thermo Fisher Scientific, Inc.). The RPL37 gene was used
as a control to normalize the values by the 2−ΔΔCq
quantification method (62).
 | Table IPrimer sequences. |
Table I
Primer sequences.
| Gene | Forward primer
(3′-5′) | Reverse primer
(3′-5′) |
|---|
| RPL37A |
TTCTGATGGCGGACTTTACC |
CACTTGCTCTTTCTGTGGCA |
| SOD1 |
CCACACCTTCACTGGTCCAT |
CTAGCGAGTTATGGCGACG |
| SOD2 |
TAGGGCTGAGGTTTGTCCAG |
CACCGAGGAGAAGTACCAGG |
| CAT |
ACGGGGCCCTACTGTAATAA |
AGATGCAGCACTGGAAGGAG |
| VDR |
CCAGTTCGTGTGAATGATGG |
GTCGTCCATGGTGAAGGA |
| PDIA3 |
CTCCGACGTGCTAGAACTCA |
CAGGTGTTAGTGTTGGCAGT |
| CYP2R1 |
AGAGACCCAGAAGTGTTCCAT |
GTCTTTCAGCACAGATGAGGTA |
| CYP3A4 |
AAGGCACCACCCACCTATGATACT |
TACTTTGGGTCACGGTGAAGAGCA |
| CYP27B1 |
TGTTTGCATTTGCTCAGA |
CCGGGAGAGCTCATACAG |
| CYP24A1 |
GCAGCCTAGTGCAGATTT |
ATTCACCCAGAACTGTTG |
| CYP11A1 |
TGGGTCGCCTATCACCAGTAT |
CCACCCGGTCTTTCTTCCA |
Statistical analysis
Statistical analysis was performed using Microsoft
Excel 2007 (Microsoft Corporation, Redmond, WA, USA) or GraphPad
Prism version 6.03 (GraphPad Software, Inc., La Jolla, CA, USA).
The data were subjected to Student’s t-test for the comparison of
two groups, one-way analysis of variance followed by Dunnett’s or
Tukey’s multiple comparison post hoc tests. The data are expressed
as the mean ± standard deviation (n=3-6). Differences were
considered statistically significant when P<0.05.
Results
Vitamin D analogs modulate the cytotoxic
effects of hydrogen peroxide in human malignant melanoma A375
cells
In agreement with previous studies (35,49,50,63,64)
vitamin D analogs 1,25(OH)2D3, 20(OH)D3, 21(OH)pD and
calcipotriol, effectively inhibited the proliferation of human
melanoma A375 cells, as demonstrated by the SRB assay (Fig. 1A–D). A decrease of ≤20% in cell
proliferation was observed, significant at the highest tested
concentration of vitamin D analogs (10−6 M). The
relative IC50 values ranged from 5.3 nM for 20(OH)D3 to
~0.274 nM for 1,25(OH)2D3 and 0.038 nM for calcipotriol
(Fig. 1A–D).
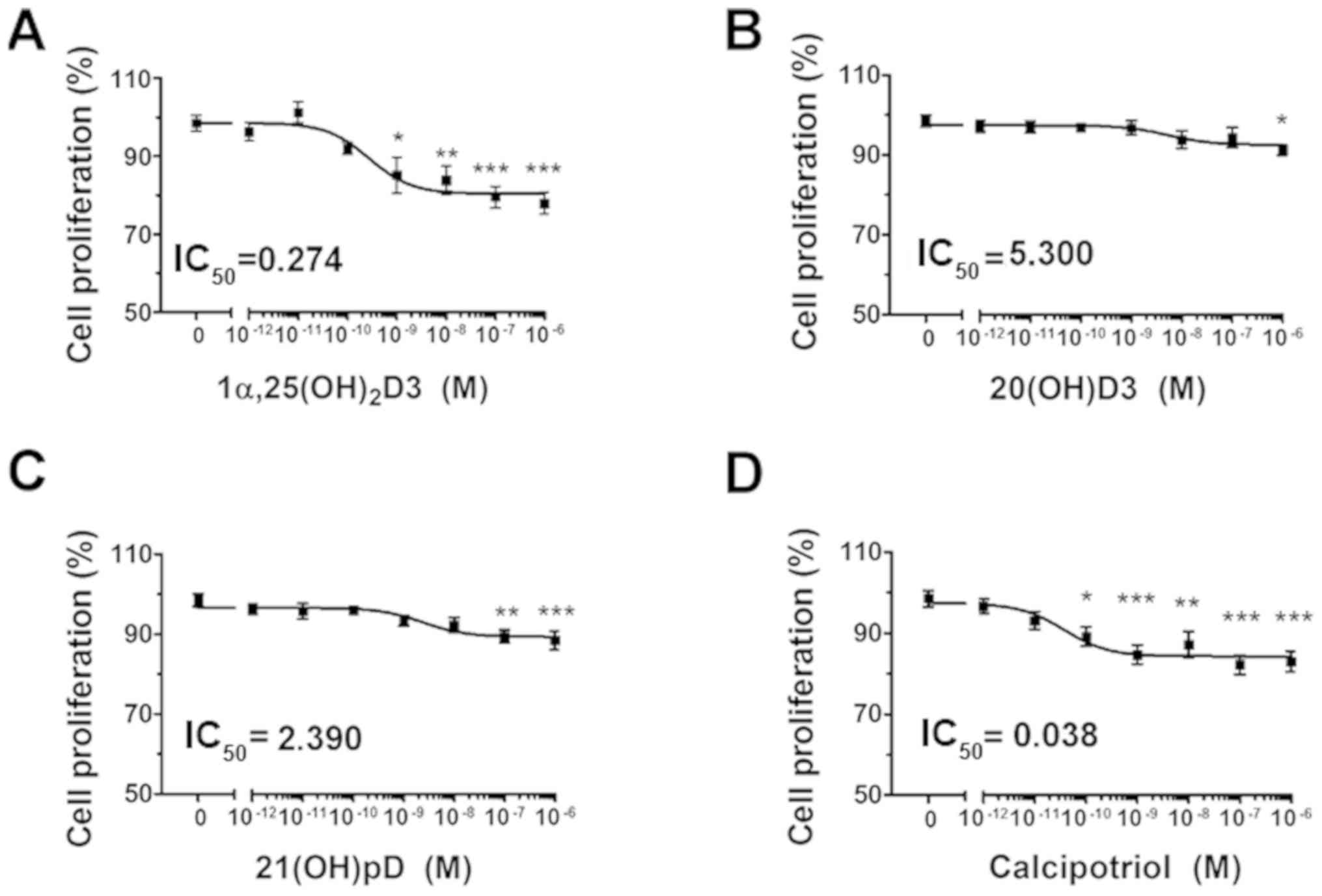 | Figure 1The effect of vitamin D derivatives
on the proliferation of human melanoma A375 cells treated with
H2O2. The cells were treated with serial
dilutions (10−12-10−6 M) of (A)
1,25(OH)2D3, (B) 20(OH)D3, (C) 21(OH)pD or (D)
calcipotriol. *P<0.05, **P<0.005 and
***P<0.0005 versus control using one-way analysis of
variance. The cells were treated with serial dilutions of
H2O2 (0.0039-0.25 mM) alone or in combination
with (E) 10 nM 1,25(OH)2D3, (F) 20(OH)D3, (G) 21(OH)pD
or (H) calcipotriol for 24 h. The results presented are
representative of three experiments (n=6). *P<0.05,
**P<0.01 and ***P<0.001 between the two
treatments at each H2O2 concentration, using
one-way analysis of variance followed by Tukey’s multiple
comparison test. The same control data is plotted in each graph. In
order to investigate the effect of secosteroid pre-treatment on
mitochondrial transmembrane potential, human melanoma A375 cells
were treated with (I) 100 nM 1,25(OH)2D3 or calcipotriol
for 24 h, and subsequently exposed to 7.5 µM
H2O2 for 1 or 3 h, then stained with JC-1 and
analyzed by flow cytometry. The data are presented as mean ±
standard deviation of 3 independent experiments.
***P<0.001 versus untreated control or between the
two groups indicated by the bracket using one-way analysis of
variance followed by Tukey’s multiple comparison test. The positive
control was exposed to CCCP for 5 min prior to staining with JC-1.
IC50, half maximal inhibitory concentration;
1α,25(OH)2D3, 1α, 25-dihydroxyvitamin D3; 20(OH)D3,
20S-hydroxyvitamin D3; 21(OH)pD, 21-hydroxypregnacalciferol;
H2O2, hydrogen peroxide; CCCP, carbonyl
cyanide 3-chlorophenylhydrazone. |
The effects of vitamin D derivatives on the
sensitivity of A375 cells to ROS were also tested. Hydrogen
peroxide, an oxidative stress-generating compound, inhibited the
proliferation of the cells with a relative IC50 of 17
µM (Fig. 1E–H).
Simultaneous treatment with hydrogen peroxide and
1α,25(OH)2D3, 20(OH)D3 or calcipotriol at a
concentration of 10 nM (Fig. 1E, F and
H) for 24 h resulted in a further decrease in the proliferation
of the melanoma cells. The effect was more pronounced for
1α,25(OH)2D3 (Fig. 1E)
and calcipotriol (Fig. 1H),
however we did not observe any significant decrease between the
calculated IC50 values (Table II). It has been suggested that
altered mitochondrial activity may be a signature of certain
melanoma cells (65). In the
present study, changes in Δψm were monitored using the
JC-1 dual-emission potential-sensitive probe, by flow cytometry.
The results revealed that the pre-incubation of the A375 cells with
calcipotriol, but not 1α,25(OH)2D3, for 24 h modulated
the effect of hydrogen peroxide on the Δψm (Fig. 1I). Notably, the pre-treatment with
calcipotriol resulted in a protective effect on Δψm in
melanoma cells treated with hydrogen peroxide for 1 h (Fig. 1I). Prolonged exposure to hydrogen
peroxide (3 h) in combination with pre-treatment of melanoma cells
with either 1,25(OH)2D3 or calcipotriol triggered a
decrease in Δψm (Fig.
1I), although the observed differences were not
significant.
| Table IISummary of the relative
IC50 values for inhibition of proliferation of human
melanoma A375 cells by H2O2 (0.004-0.250 mM),
cisplatin (0.19-300 µM) or dacarbazine (0.15-10 µM)
in the presence or absence of the tested secosteroids. |
Table II
Summary of the relative
IC50 values for inhibition of proliferation of human
melanoma A375 cells by H2O2 (0.004-0.250 mM),
cisplatin (0.19-300 µM) or dacarbazine (0.15-10 µM)
in the presence or absence of the tested secosteroids.
| Incubation time,
h | Tested
compound | Relative IC 50
|
|---|
| Monotreatment | +10 nM
1α,25(OH)2D3 | +10 nM
20(OH)D3 | +10 nM
21(OH)pD | +10 nM
calcipotriol |
|---|
| 24 |
H2O2 | 0.017±0.07 | 0.011±0.001 | 0.013±0.0006 | 0.017±0.002 | 0.012±0.0006 |
| 24 | Cisplatin | 4.81±2.2 | 11.61±0.98a | 14.08±3.29b | 9.37±1.64 | 15.23±6.15b |
| 48 | Cisplatin | 2.57±0.19 | 1.97±0.22 | 3.47±1.04 | 3.71±1.90 | 2.13±1.01 |
| 48 | Dacarbazine | 1.07±0.31 | 0.45±0.35a | 1.17±0.40 | 1.04±0.36 | 0.85±0.39 |
Vitamin D analogs modulate the cytotoxic
effects of cisplatin and dacarbazine on human malignant melanoma
A375 cells
It is well established that oxidative stress and the
resulting cell damage is one of the mechanisms of cell death
induced by anticancer drugs. Thus, based on the aforementioned
results with hydrogen peroxide (Fig.
1E–I), the effect of the treatment of A375 human melanoma cells
with 1α,25(OH)2D3, 20(OH) D3, 21(OH)pD or calcipotriol,
on the ability of cisplatin or dacarbazine to inhibit
proliferation, was investigated. These two drugs are widely used in
melanoma treatment and their activity, at least partially, relies
on ROS generation (35,57,58,66).
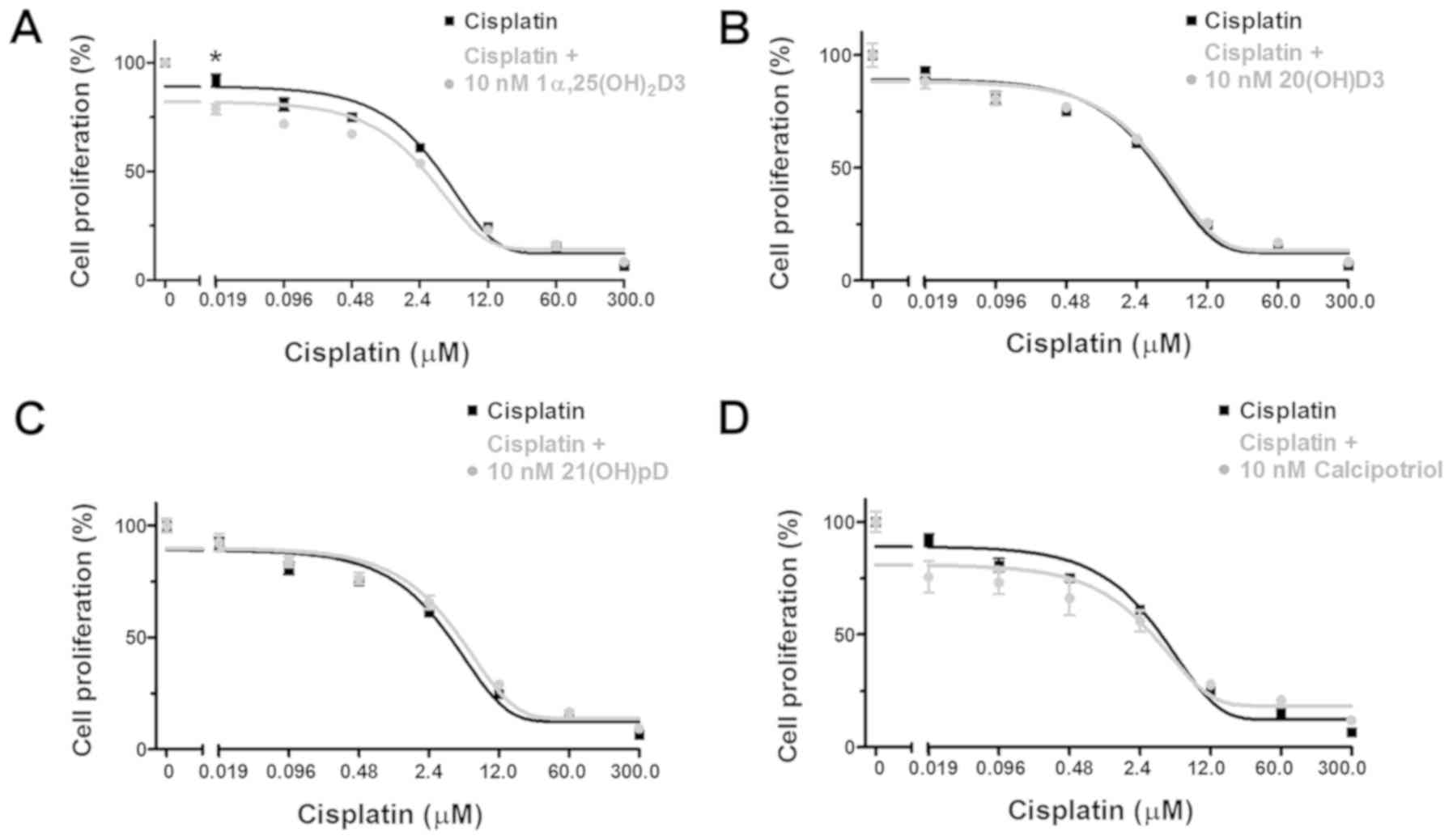
The anti-melanoma effects of cisplatin (Fig. 2A–D) or dacarbazine (Fig. 3A–D) alone or with 10 nM
1α,25(OH)2D3, 20(OH)D3, 21(OH)pD or calcipotriol were
investigated in A375 cells using the SRB assay. Simultaneous
treatment with vitamin D analogs and cisplatin for 24 h resulted in
an unexpected increase in the cisplatin relative IC50,
suggesting protective effects of the secosteroids (Table II). However, during prolonged
incubation with cisplatin (48 h), the addition of
1,25(OH)2D3, but not 20(OH)D3, 21(OH)pD or calcipotriol,
resulted in a decreasing trend in the relative IC50 in
comparison to cisplatin alone (Fig.
2; Table II), however the
observed differences were not significant.
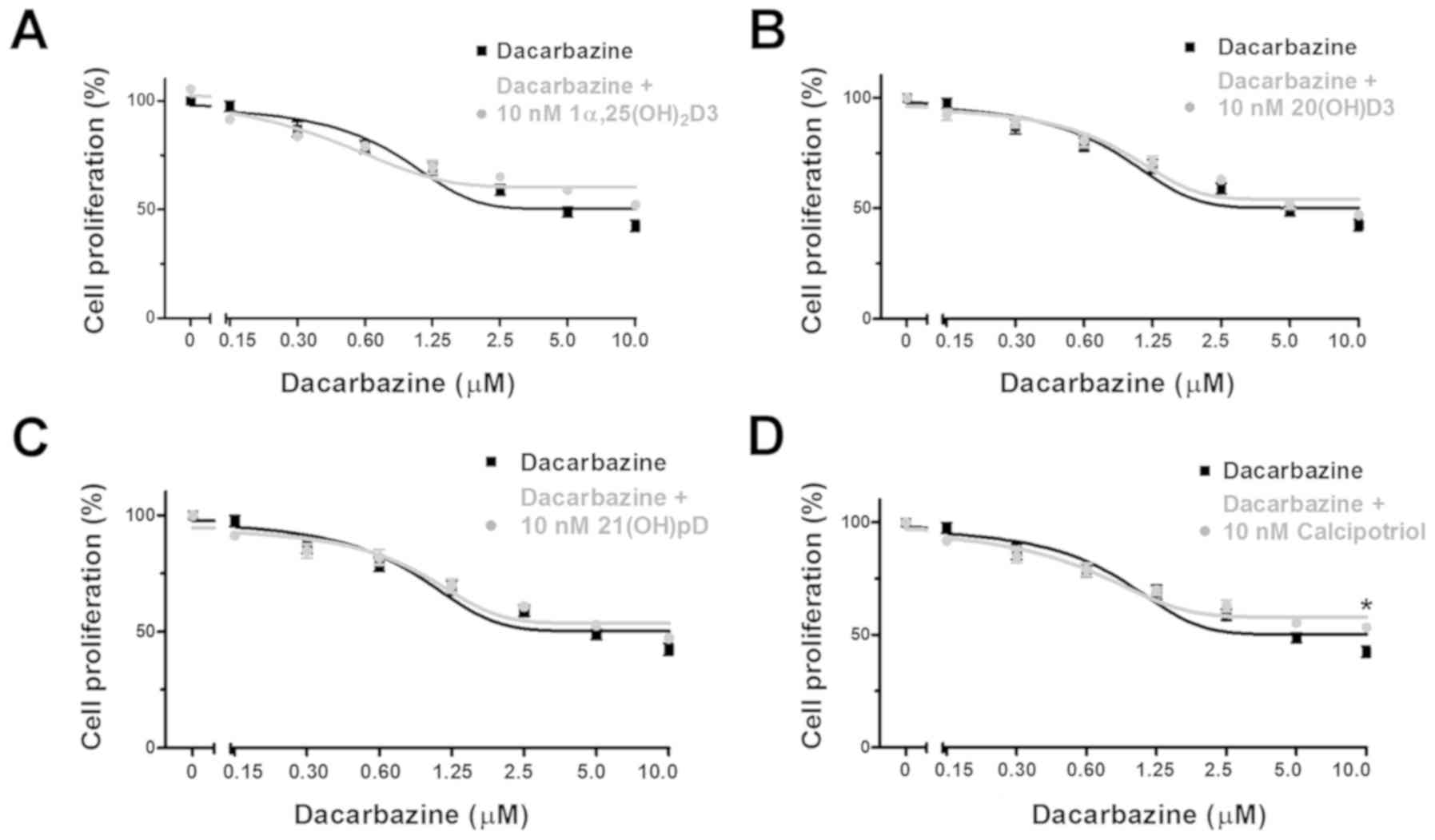
Dacarbazine inhibited the proliferation of human
melanoma A375 cells during a 48 h incubation with a relative
IC50 of 1.07 µM (Table II). The results from the combined
treatment with the vitamin D analogs revealed that
1α,25(OH)2D3, but not 20(OH)D3, 21(OH)pD or
calcipotriol, decreased the relative IC50 observed with
dacarbazine alone by 2.3-fold (Table
II).
Pre-treatment of human malignant melanoma
A375 cells with vitamin D derivatives alters the distribution of
the cells in the cell cycle phases following treatment with
dacarbazine, but not cisplatin
To investigate the mechanism of proliferation
inhibition of melanoma A375 cell by the combination of vitamin D
analogs and the tested drugs, changes in the distribution of the
cells in the cell cycle phases were investigated by flow cytometry.
The cells were pre-treated with the vitamin D analogs for 24 h and
then incubated with cisplatin or dacarbazine for an additional 24
or 48 h. The initial experiments revealed no significant effects of
pre-treatment of melanoma cells for 24 h with 10 nM secosteroids in
combination with additional incubation with cisplatin for 24 h on
the cell cycle distribution (data not shown). Since the results of
the aforementioned SRB tests (Fig.
1A–D) demonstrated a plateau in the inhibition of cell
proliferation at 10 and 100 nM concentrations, and taking into
consideration that vitamin D is widely used at higher
concentrations (100-1,000 nM) in in vitro studies (67-69),
the concentration of vitamin D analogs was raised to 100 nM for the
present assay. Additionally, the time of incubation with cisplatin
or dacarbazine was increased to 48 h, similar to the conditions
used during proliferation tests, and their concentrations were
increased to 24 and 6 µM, respectively, to maximize the
observed effect.
The treatment of melanoma A375 cells with 24
µM cisplatin alone for 48 h resulted in an increase in the
number of SubG1 cells (P<0.001), indicating induction
of apoptosis with a concomitant decrease in the number of cells in
the G0/G1 (P<0.001), S (P<0.001) and
G2/M (P<0.001) phases (Fig. 4A–D). No impact of the vitamin D
pre-treatment was observed on the distribution of cisplatin-treated
melanoma cells in the cell cycle. The effect of pre-treatment of
melanoma A375 cells with vitamin D analogs prior to incubation with
dacarbazine was also tested (Fig.
4E–H). The treatment with 6 µM dacarbazine alone for 48
h resulted in an increase in the fraction of cells in
G0/G1 compared with untreated cells
(P<0.01), with a minor effect on the SubG1 fraction
(P<0.001) in comparison with cells treated with cisplatin alone
(<10 vs. >60% of all cells analyzed at SubG1
following treatment with dacarbazine or cisplatin, respectively;
P<0.001 cisplatin versus untreated cells; P<0.001 dacarbazine
versus untreated cells; Fig. 4A and
E). In addition, 24 h pre-treatment with 100 nM
1α,25(OH)2D3 or calcipotriol prior to dacarbazine
treatment resulted in an increase in the percentage of cells in the
G0/G1 phase compared with that observed with
dacarbazine alone (P<0.001; Fig. 4E
and H). The effect was accompanied by a decrease in the
percentage of cells in the G2/M phase for
1α,25(OH)2D3 (P<0.05; Fig. 4E), and in S and G2/M
phases for calcipotriol (P<0.001 and P<0.05, respectively;
Fig. 4H).
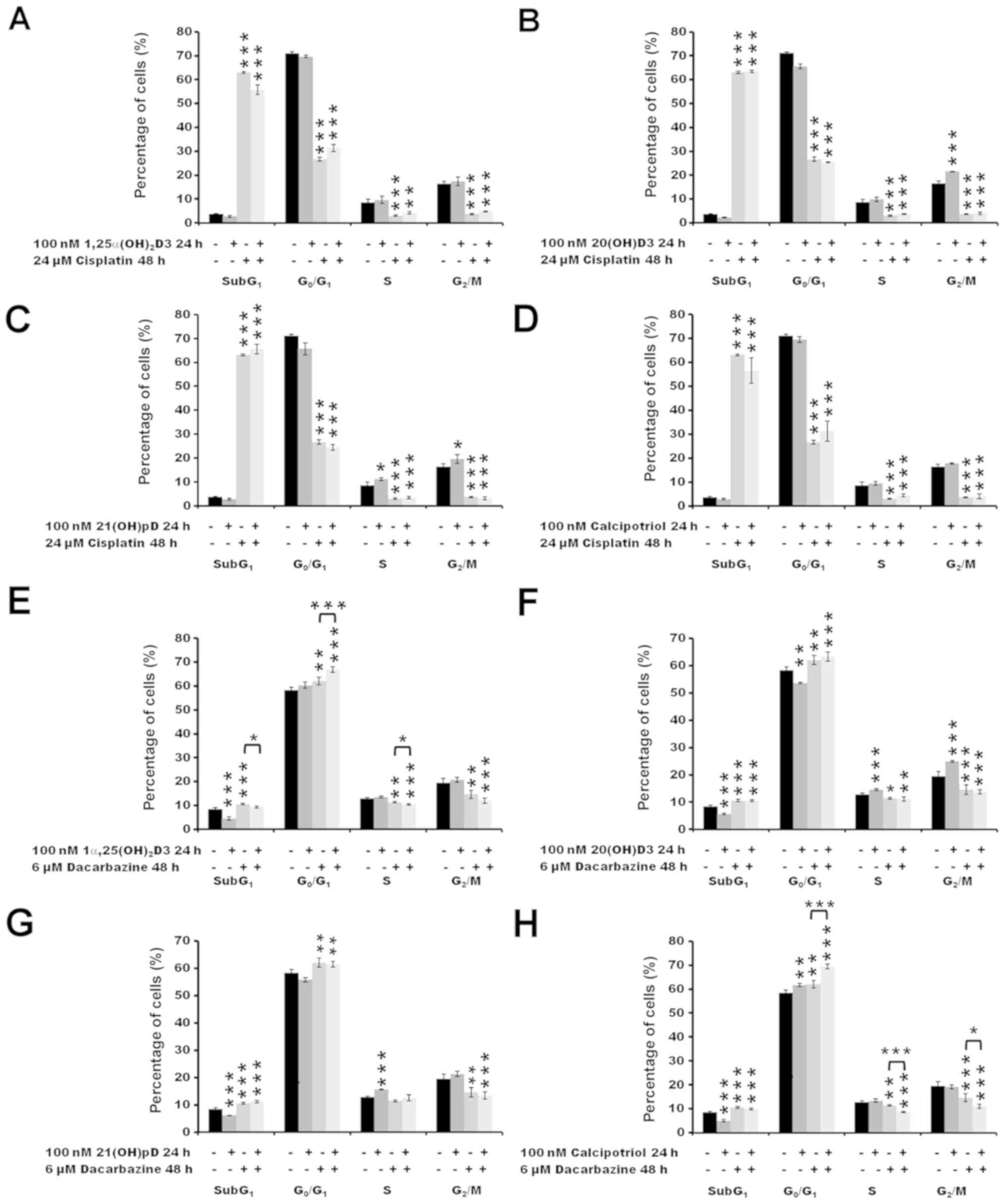 | Figure 4The effect of secosteroids and
cisplatin or dacarbazine on the distribution of human melanoma A375
cells through the cell cycle. Cells that were treated with 24
µM cisplatin for 48 h had been pre-treated with (A) 100 nM
1α,25(OH)2D3, (B) 20(OH)D3, (C) 21(OH)pD or (D)
calcipotriol for 24 h. Similarly, cells that were treated with 6
µM dacarbazine 48 h had been pre-treated with (E) 100 nM
1α,25(OH)2D3, (F) 20(OH)D3, (G) 21(OH)pD or (H)
calcipotriol for 24 h. The cells were harvested, stained with
propidium iodide and analyzed by flow cytometry. The data are
presented as the mean ± standard deviation (n=3).
*P<0.05, **P<0.01 and
***P<0.001, calculated using one-way analysis of
variance followed by Tukey’s multiple comparison test versus
untreated control or between the two groups indicated by the
bracket. SubG1, apoptotic/necrotic cells; G1, growth; S, DNA
synthesis; G2/M, preparation for mitosis/mitosis;
1α,25(OH)2D3, 1α, 25-dihydroxyvitamin D3; 20(OH)D3,
20S-hydroxyvitamin D3; 21(OH)pD, 21-hydroxypregnacalciferol. |
Pre-treatment with vitamin D derivatives
changes the Δψm in human melanoma A375 cells and alters
the cisplatin- or dacarbazine-induced production of ROS
The effects of the anti-cancer drugs on the
Δψm of the melanoma A375 cells were analyzed by
measuring JC-1 fluorescence by flow cytometry (Figs. 5 and 6). Treatment with cisplatin alone for 3 h
did not influence the Δψm, at either of the two
concentrations tested (2.4 and 12 µM; Fig. 5A–D). A 24 h pre-treatment with
21(OH) pD (Fig. 5C) or
calcipotriol (Fig. 5D) resulted in
a decrease in Δψm following treatment with cisplatin,
compared to the cisplatin effect observed without pre-treatment.
Notably, pre-treatment of the melanoma cells with 20(OH)D3 resulted
in an increase in Δψm (Fig.
5B) following exposure to 2.4 µM cisplatin (P<0.001).
However, this effect was not observed at higher concentration of
the drug, or without the drug treatment.
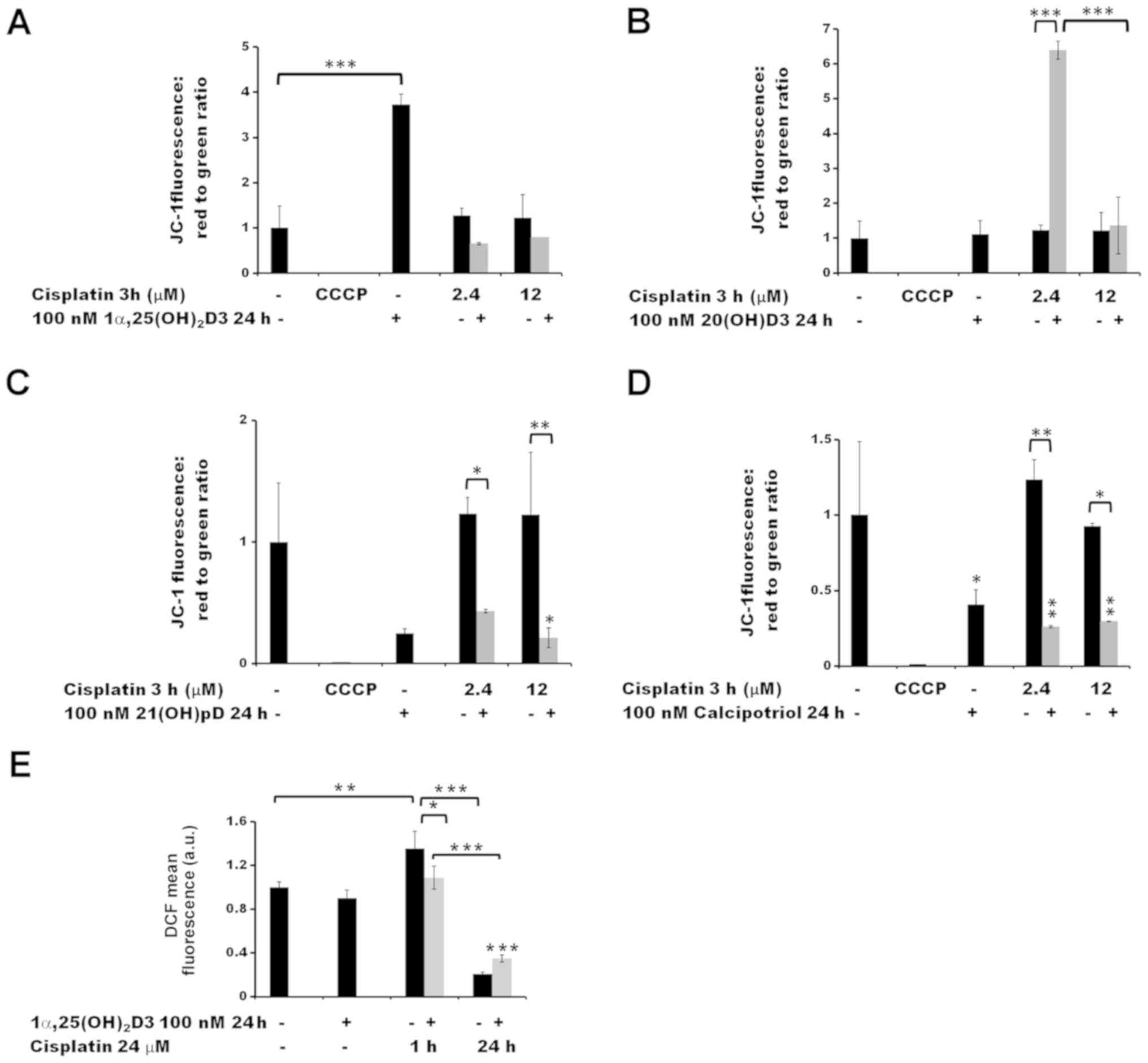 | Figure 5The effect of pre-treatment of human
melanoma A375 cells with vitamin D derivatives on the
cisplatin-induced changes in the mitochondrial membrane potential
and ROS levels. A375 cells were treated with (A) 100 nM
1α,25(OH)2D3, (B) 20(OH)D3, (C) 21(OH)pD or (D)
calcipotriol for 24 h, and subsequently exposed to 2.4 or 12
µM cisplatin for 3 h. The cells were stained with JC-1 and
analyzed by flow cytometry. The positive control was exposed to
CCCP for 5 min prior to staining with JC-1. (E) The effect of
1α,25(OH)2D3 on ROS levels. The cells were treated with
100 nM 1,25(OH)2D3 for 24 h and subsequently exposed to
24 µM cisplatin for 1 or 24 h. The cells were stained with
H2DCFDA and analyzed by flow cytometry. The data are
presented as the mean ± standard deviation (n=3).
*P<0.05; **P<0.01; and
***P<0.001, calculated using one way analysis of
variance followed by Tukey’s multiple comparison test between the
two groups indicated by the bracket or compared with the untreated
control. 1α,25(OH)2D3, 1α, 25-dihydroxyvitamin D3;
20(OH)D3, 20S-hydroxyvitamin D3; 21(OH)pD,
21-hydroxypregnacalciferol; CCCP, carbonyl cyanide
3-chlorophenylhydrazone; ROS, reactive oxygen species. |
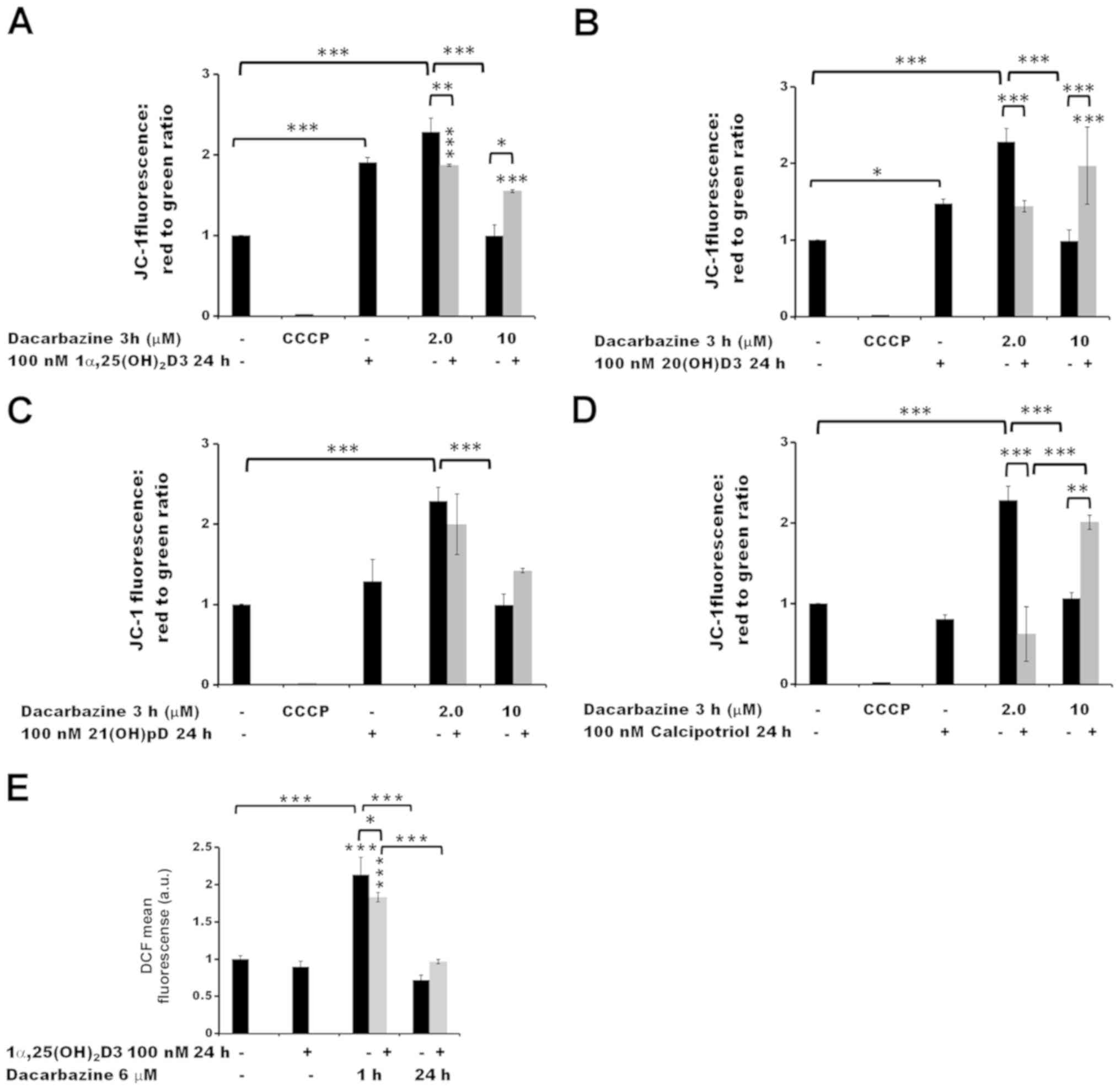 | Figure 6The effect of pre-treatment of human
melanoma A375 cells with vitamin D derivatives on the
dacarbazine-induced changes in the mitochondrial membrane potential
and ROS levels. A375 cells were treated with (A) 100 nM
1α,25(OH)2D3, (B) 20(OH)D3, (C) 21(OH)pD or (D)
calcipotriol for 24 h, and subsequently exposed to 2.0 or 10
µM dacarbazine for 3 h The cells were stained with JC-1 and
analyzed by flow cytometry. The positive control was exposed to
CCCP for 5 min prior to staining with JC-1. (E) The effect of
1α,25(OH)2D3 on ROS levels. The cells were treated with
100 nM 1,25(OH)2D3 for 24 h and subsequently exposed to
6 µM dacarbazine for 1 or 24 h. The cells were stained with
H2DCFDA and analyzed by flow cytometry. The data are
presented as the mean ± standard deviation (n=3).
*P<0.05, **P<0.01 and
***P<0.001, calculated using one way analysis of
variance followed by Tukey’s multiple comparison test between the
two groups indicated by the bracket or compared with the untreated
control. 1α,25(OH)2D3, 1α,25-dihydroxyvitamin D3;
20(OH)D3, 20S-hydroxyvitamin D3; 21(OH)pD,
21-hydroxypregnacalciferol; CCCP, carbonyl cyanide
3-chlorophenylhydrazone; ROS, reactive oxygen species. |
A 3 h treatment with 2.0 µM dacarbazine alone
led to an increase in the Δψm of melanoma A375 cells
(P<0.001) but this was not the case at the higher concentration
(10 µM) (Fig. 6A–D). The 24
h pre-treatment of the cells with 1α,25(OH)2D3 (Fig. 6A), 20(OH)D3 (Fig. 6B) or calcipotriol (Fig. 6D) resulted in a decrease in
Δψm following exposure to 2.0 µM dacarbazine
(P<0.01 for 1α,25(OH)2D3 and P<0.001 for 20(OH)D3
and calcipotriol versus dacarbazine alone). In the case of 21(OH)pD
(Fig. 6C), the effect was not
statistically significant. In contrast, at the higher concentration
of dacarbazine (10 µM), the pre-treatment of the cells with
1,25(OH)2D3, 20(OH)D3 or calcipotriol resulted in an
increase in Δψm (P<0.05, P<0.001 and P<0.01,
respectively, versus 10 µM dacarbazine alone). No
significant difference was observed in the case of pre-treatment
with 21(OH)pD.
A pre-treatment of malignant melanoma A375 cells
with 100 nM 1,25(OH)2D3 for 24 h did not influence the
production of ROS in comparison with untreated cells, as determined
by the H2DCFDA assay (Figs.
5E and 6E). However, this
pre-treatment affected the ROS production following treatment with
either cisplatin (Fig. 5E) or
dacarbazine (Fig. 6E). The
observed effect was time-dependent. Exposure of the cells to
cisplatin or dacarbazine alone for 1 h, without vitamin D
pre-treatment, led to a significant increase in the ROS levels
(P<0.01 for cisplatin and P<0.001 for dacarbazine; Figs. 5E and 6E, respectively). However, 24 h
pre-treatment of melanoma cells with 1α,25(OH)2D3
decreased the effect that the 1 h cisplatin or dacarbazine
treatment had on the ROS levels (P<0.05 versus no pre-treatment;
Fig. 5E). In contrast, prolonged
exposure (24 h) to cisplatin or dacarbazine alone tended towards a
decrease in the ROS levels in the melanoma cells, whereas the
1α,25(OH)2D3 pre-treatment alleviated the effect of the
24 h cisplatin or dacarbazine treatment on the ROS levels, although
the observed differences were not significant.
Modulation of the expression of selected
genes by cisplatin or dacarbazine in the presence or absence of
1α,25(OH)2D3
In order to verify the aforementioned changes in ROS
generation and the Δψm, the impact of
1α,25(OH)2D3 pre-treatment on the expression of the
selected ROS-associated genes was tested in melanoma A375 cells
treated with cisplatin or dacarbazine (Fig. 7). No significant effect was
observed in the expression of superoxide dismutases 1 and 2
(SOD1 and SOD2) or catalase (CAT) by
1α,25(OH)2D3 under the experimental conditions used
(Fig. 7A–C). Treatment of the
cells with the anticancer drugs had a limited effect on the mRNA
levels of the selected ROS-associated genes. A decrease in
SOD2 gene expression was observed under the influence of
cisplatin alone (P<0.05 vs. no treatment control; Fig. 7B), as well as in SOD1 and
CAT gene expression following treatment with dacarbazine
alone (both P<0.05 vs. no treatment control; Fig. 7A and C, respectively).
Pre-treatment of the cells with 1α,25(OH)2D3 prior to
incubation with dacarbazine resulted in an increase of CAT
mRNA compared with cells treated solely with dacarbazine
(P<0.05; Fig. 7C).
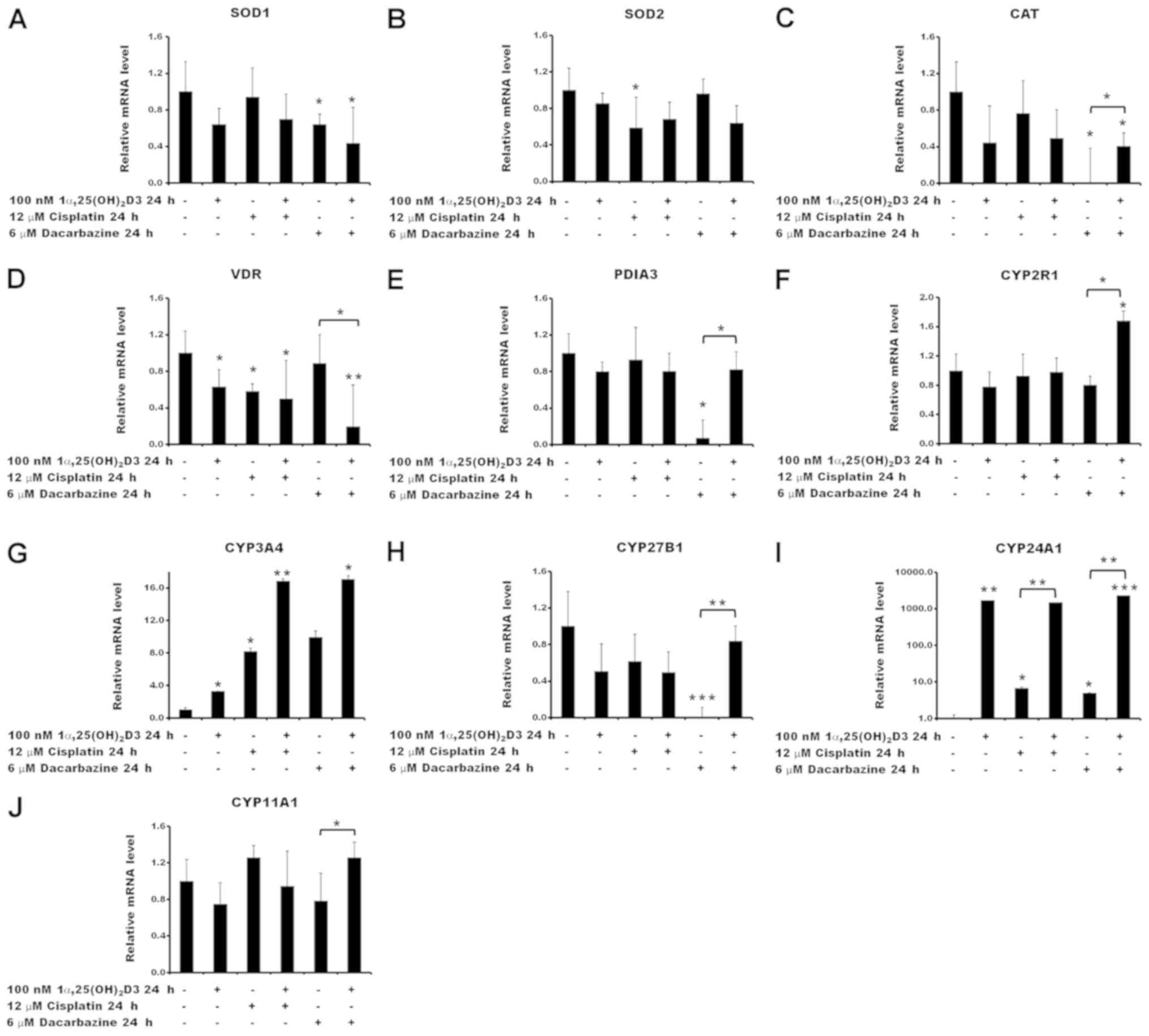 | Figure 7Relative mRNA quantification of
reactive oxygen species- and vitamin D-associated genes. Effects of
cisplatin or dacarbazine treatment on the mRNA levels of (A)
SOD1, (B) SOD2, (C) CAT, (D) VDR, (E)
PDIA3, (F) CYP2R1, (G) CYP3A4, (H)
CYP27B1, (I) CYP24A1 and (J) CYP11A1 gene
expression in human melanoma A375 cells pre-treated with
1,25(OH)2D3. The cells were incubated with 100 nM
1,25(OH)2D3 for 24 h, followed by exposure to 12
µM cisplatin or 6 µM dacarbazine for an additional 24
h. The mRNA levels were measured by reverse
transcription-quantitative polymerase chain reactions. The data are
presented as the mean ± standard deviation of 3 independent
experiments carried out in duplicate. *P<0.05,
**P<0.01 and ***P<0.001, calculated
using Student’s t-test vs. untreated control or between the two
groups indicated by the bracket. 1α,25(OH)2D3, 1α,
25-dihydroxyvitamin D3; SOD, superoxide dismutase; CAT, catalase;
VDR, vitamin D receptor; PDIA3, protein disulfide-isomerase A3;
CYP2R1, vitamin D 25-hydroxylase; CYP3A4, cytochrome P450 3A4;
CYP27B1, 25-hydroxyvitamin D3 1-α-hydroxylase; CYP24A1, vitamin D
24-hydroxylase; CYP11A1, cholesterol side-chain cleavage
enzyme. |
Subsequently, the effect of cisplatin or dacarbazine
on the expression of vitamin D-associated genes, including ones
encoding vitamin D receptors VDR and protein
disulfide-isomerase A3 (PDIA3), and vitamin D metabolizing
hydroxylases that belong to the cytochrome P450 (CYP) family,
CYP2R1, CYP3A4, CYP27B1, CYP24A1 and
CYP11A1, was investigated in melanoma A375 cells, as well as
the consequences of pre-treatment with 1α,25(OH)2D3. The
results revealed that 1α,25(OH)2D3 and cisplatin, used
alone, decreased VDR mRNA levels in the A375 cells
(P<0.05; Fig. 7D). The effect
of dacarbazine was statistically significant only in the case of
the 1α,25(OH)2D3 pre-treatment (P<0.05; Fig. 7D). In contrast,
1α,25(OH)2D3 and cisplatin had no effect on PDIA3
mRNA levels (Fig. 7E), whereas
dacarbazine alone led to a significant decrease (P<0.05).
Notably, the effect of dacarbazine alone was reversed by the
1α,25(OH)2D3 pre-treatment (P<0.05).
Although the transcription of CYP2R1 was not
affected by 1α,25(OH)2D3, cisplatin or dacarbazine
alone, pre-treatment of the A375 cells with 1α,25(OH)2D3
with subsequent exposure to dacarbazine resulted in an increase in
its mRNA (P<0.05; Fig. 7F)
compared with that in the cells treated with dacarbazine alone.
Stimulation of CYP3A4 expression was observed with all
combinations of the drugs tested. The effect was further
exacerbated by a 24 h 1α,25(OH)2D3 pre-treatment
(Fig. 7G). Treatment with
1α,25(OH)2D3 or cisplatin alone or in combination had no
statistically significant effect on the CYP27B1 mRNA levels
(Fig. 7H). However, treatment with
dacarbazine alone resulted in a decrease (P<0.001) and this
effect was reversed by prior administration of
1α,25(OH)2D3 (P<0.01). As expected, pre-treatment of
the cells with 1α,25(OH)2D3 resulted in a strong
stimulation of CYP24A1, which encodes the vitamin D
deactivation enzyme, 24-hydroxylase (Fig. 7I). An increase in CYP24A1 mRNA
levels was also observed for cisplatin or dacarbazine alone,
although to a lesser extent [7- and 5-fold increase, respectively,
versus a 1,700-fold increase for 1α,25(OH)2D3].
Furthermore, cisplatin or dacarbazine had no effect on the level of
CYP24A1 mRNA following pre- treatment with
1α,25(OH)2D3, compared with cells treated solely with
1α,25(OH)2D3 (Fig. 7I).
Treatment of the A375 cells with 1α,25(OH)2D3, cisplatin
or dacarbazine alone did not affect the transcription levels of the
CYP11A1 gene (Fig. 7J).
Finally, the pre-treatment with 1α,25(OH)2D3 followed by
treatment with dacarbazine resulted in a small, but significant,
increase in the CYP11A1 mRNA level (P<0.05 vs.
dacarbazine alone).
Discussion
It is well established that UV radiation is a major
skin carcinogen that serves an important role in melanomagenesis
(14,70,71).
However, UVB is also indispensable for the production of vitamin D
in the skin (1-3). Considering the antiproliferative and
differentiation-promoting function of vitamin D and its analogs, it
seemed advantageous to explore their efficacy as anticancer drugs
and their potential for positive interactions with other
antimelanoma drugs or therapeutic approaches (34). The effects of the active forms of
vitamin D require VDR activation, which results in the modulation
of the expression in ~3,000 target genes in humans (72), including those involved in DNA
repair and the oxidative stress response (73). Vitamin D deficiency is considered
to contribute to carcinogenesis, and notably, to poor prognosis due
to multidrug resistance (74,75).
Recently published data suggest an inverse correlation between the
vitamin D serum level and the relative risk of melanoma and
non-melanoma skin cancer, as well as melanoma thickness at
diagnosis (30,75,76).
Wyatt et al (77) also
suggested that vitamin D deficiency at the time of melanoma
diagnosis is not only associated with a higher Breslow thickness
but also with a poorer prognosis. Ogbah et al (78) reported that even in patients living
in the sunny Mediterranean area, 1α,25(OH)2D3 levels
were sub-optimal at the time of melanoma diagnosis. Patients with
metastatic melanoma, who were initially vitamin D deficient, had
significantly poorer outcomes in comparison to individuals who,
being initially deficient, exhibited a >20 ng/ml increase in
their 25-hydroxyvitamin D3 [25(OH) D3] concentration during the
therapy period (75). Vitamin D
deficient patients with stage IV metastatic melanoma also had
significantly poorer prognosis (75). Therefore, the administration of
vitamin D is potentially beneficial in cancer therapy.
A previous study has revealed that melanoma A375
cells are ≥10 times more sensitive to hydrogen peroxide than human
immortalized HaCaT keratinocytes. The interaction between hydrogen
peroxide, as a model oxidative stress inducer, and vitamin D
analogs were investigated (35).
First, as reported for HaCaT keratinocytes (35), the incubation of melanoma A375
cells with vitamin D analogs resulted in higher sensitivity of the
cells to hydrogen peroxide treatment (Fig. 1). It should be emphasized that
HaCaT keratinocytes represent a cellular model of epithelial cells,
whereas melanocytes are derived from neural crest cells (79) and therefore represent a different
cellular model. Hence, the present study focused on human malignant
melanoma cells. Hydrogen peroxide treatment was used to investigate
the association between ROS levels and vitamin D analogs, and
subsequently the interaction between vitamin D analogs and
anticancer drugs was explored.
Secondly, similar effects to those discussed above
for hydrogen peroxide were observed for dacarbazine, but not
cisplatin, following treatment with 1α,25(OH)2D3, since
sensitization of 1α,25(OH)2D3-treated melanoma cells to
this drugs was observed. Notably, the highest concentration of
cisplatin (300 µM) resulted in a decrease in cell
proliferation as measured using the SRB assay, by >90%, whereas
treatment with 10 μM dacarbazine decreased proliferation by
50%. Incubation of the melanoma A375 cells with
1α,25(OH)2D3, 20(OH)D3, 21(OH)pD or calcipotriol for 24
h resulted in up to a 20% decrease in cell proliferation at the
highest concentrations tested. This inhibitory effect of the
vitamin D analogs, with the exception of 20(OH)D3, is consistent
with previous studies (80,81),
however certain differences were noted in the relative
IC50 values [i.e., 1α,25(OH)2D3 relative
IC50, 0.274 vs. 6.4 nM reported by Wasiewicz et
al (81)]. The variation among
the relative IC50 values could be explained by variable
experimental conditions, including a shorter incubation time with
vitamin D analogs (24 vs. 48 h), as well as a lower FBS
concentration in the medium. It is already known that vitamin D
inhibits cell proliferation and promotes their differentiation
(80,82,83).
Therefore, the inhibition of melanoma cell proliferation by vitamin
D should not be considered as a direct cytotoxic effect, but rather
reveals its antiproliferative potential.
Thirdly, it appears that these two drugs inhibit
melanoma cell proliferation via distinct mechanisms (58,66).
It should be noted that even though cisplatin and dacarbazine
function primarily based on the induction of DNA damage (56,84),
it is apparent that these drugs also lead to the generation of ROS
inside treated cells (57,58). The current study design was based
on a 24 h pre-treatment with vitamin D analogs at a low
concentration (100 nM). This corresponds to the optimal level of
25(OH)D3 in the serum (75-125 nM) (28), since, according to Timerman et
al (75), vitamin D deficiency
is associated with a poorer prognosis in metastatic melanoma. As
demonstrated by the cell cycle analyses, induction of apoptosis
(increase in SubG1 cell fraction) was observed for the
cells treated with cisplatin, consistent with other studies
(85,86). On the other hand, the inhibition of
melanoma cell proliferation by dacarbazine probably results from
cell cycle arrest, as observed from an increase in the number of
cells in the G0/G1 fraction (P<0.01) and
decreases in the S and G2/M phases (both P<0.01). As
expected based on previous studies (15,17,80,82,83),
pre-treatment with active forms of vitamin D resulted in an
increase in the number of cells in G0/G1,
with this effect being observed in cells treated with dacarbazine,
but not cisplatin. Notably, the two anticancer drugs exhibited
similar effects on oxidative stress. Treatment of the melanoma
cells with the two drugs resulted in an initial significant
increase in oxidative stress (at 1 h), whereas prolonged incubation
(24 h) resulted in a downward trend of 2′,7′-dichlorofluorescein
fluorescence in cells treated with cisplatin or dacarbazine
compared with the untreated control. The pre-incubation with
vitamin D analogs, however, resulted in a drug-specific effect on
Δψm. In the case of cisplatin, a significant decrease in
Δψm was only observed in cells pre-treated with vitamin
D derivatives, 21(OH)pD and calcipotriol (P<0.05 and P<0.01,
respectively). It has been reported that cisplatin-resistant lung
cancer cells exhibit increased Δψm in comparison with
cisplatin-sensitive counterparts (87). Therefore, a decrease in
Δψm in cisplatin-treated cells elicited by vitamin D
analogs possibly reflects their drug-sensitization potential. The
effect of the incubation of melanoma cells with the secosteoids,
with the exception of 21(OH)pD, prior to treatment with dacarbazine
was dose-dependent, with a decrease in Δψm in cells treated with a
low concentration of the drug (2 μM), and an increase at the
high concentration (10 μM).
The analyses of the expression levels of selected
genes involved in the response to ROS or the modulation of vitamin
D activity, revealed potential regulatory properties of
dacarbazine. This drug resulted in significant inhibition of the
expression of CAT, the alternative vitamin D binding protein
encoded by PDIA3, and CYP27B1, with these effects
being reversed by pre-treatment with 1α,25(OH)2D3. All
the tested analogs efficiently induced the expression of
CYP3A4, with the effect of cisplatin or dacarbazine
treatment being enhanced by secosteroid pre-treatment. This
observation indicates the induction of an anti-xenobiotic response
in the melanoma A375 cells. Similar results for cisplatin and other
anticancer compounds were reported in hepatocyte-derived HepG2
cells, in which chemotherapeutic agents activated cellular tumor
antigen p53 protein to induce the expression of the main enzymes
involved in the systemic clearance of these drugs (88). On the other hand, dacarbazine, a
prodrug, requires activation by the cytochrome P450 (CYP450) enzyme
family, to which the product of CYP3A4 belongs (89). Therefore, the observed induction of
CYP3A4 expression may also suggest more efficient oxidation
of dacarbazine to its active metabolite in melanoma cells
pre-treated with vitamin D. Cells overexpressing another CYP450
family member, CYP450 2E1 (CYP2E1), were revealed to be more
sensitive to cisplatin treatment with respect to cell viability and
ROS production, compared with cells lacking CYP2E1
expression (90).
CYP24A1 is pivotal for vitamin D homeostasis, since
it regulates the serum and tissue levels of 25(OH)D3 and
1α,25(OH)2D3, being the major vitamin D inactivating
enzyme (91). A strong induction
of CYP24A1 expression was observed with 100 nM
1α,25(OH)2D3 (Fig. 7I).
This observation is consistent with previous reports for melanoma
A375 cells (81) and HaCaT
keratinocytes (35). Notably, an
increase in CYP24A1 expression was also observed in cells
treated with 12 µM cisplatin or 6 μM dacarbazine
alone (Fig. 7I). A similar
induction of CYP24A1 expression by cisplatin has been
observed in HepG2 cells in a p53-dependent manner (88). Furthermore, dacarbazine is a well
known powerful alkylating agent that activates p53 (92). Therefore, the induction of
CYP24A1 expression in A375 melanoma cells by this
chemotherapeutic agent may involve a p53-dependent mechanism.
However, this hypothesis requires further investigation.
Similarities and differences were noted in the
phenotypic effects between 1α,25(OH)2D3 and calcipotriol
versus non-calcemic 20(OH)D3 and 21(OH)pD. These can be explained
by the different receptors targeted by each of these molecules.
Although the VDR is the primary target for 1α,25(OH)2D3
and calcipotriol, 20(OH)D3 acts only as a biased agonist on the VDR
and can act as a reverse agonist on retinoic acid orphan receptors
(46,93-96),
whereas its downstream metabolite, 20,23(OH)2D3, acts as
an agonist on the aryl hydrocarbon receptor (97). In the case of 21(OH)pD, its nuclear
receptor remains to be identified, since it has low or no affinity
for the VDR (98). Defining the
precise mechanism of action for each secosteroid is a future
goal.
In vitro studies require further validation
by in vivo animal studies prior to the use of vitamin D in
combination with cisplatin or dacarbazine in melanoma treatment.
However, pre-clinical models of human melanoma, including cell
line-transplantable mouse models, genetically engineered mouse
models or immunodeficient mice with patient-derived xenografts
(PDOX), do not reflect the true nature of the primary tumor, being
controversial in their ability to translate the effectiveness of
immunotherapeutic strategies in clinical trials (99,100). Nevertheless, animal models,
including PDOX, are the next logical step to discovering novel
targets for more efficient combinatorial therapy and approaches to
overcome emerging resistance of melanoma cells to any form of
treatment (101-103).
Despite not observing pronounced enhancement of
anti-melanoma activity by the tested chemotherapeutics under the
described experimental conditions, the results of the present study
have demonstrated that vitamin D analogs modulate the response of
melanoma cells to dacarbazine. In conclusion, low- and non-calcemic
vitamin D analogs may serve as beneficial adjuvant agents in
chemotherapy, particularly in patients suffering from vitamin D
deficiency.
Funding
This study was supported by grant no. MN
01-0250/08/280 from the Medical University of Gdansk (Gdansk,
Poland) to AP, and in part by NIH grants (nos. 1R01AR073004-01A1
and 1RO1AR071189-01A1) and a VA merit grant (no. 1I01BX004293-01A1)
to ATS.
Availability of data and materials
The datasets used and/or analyzed during the
current study are available from the corresponding author on
reasonable request.
Authors’ contributions
MAZ and AP conceived and designed and supervised
the study; AP, JW and AR performed the experiments; RCT provided
vitamin D analogs; MAZ, AP, JW, RCT and ATS analyzed the data; AP,
MAZ, RCT and ATS wrote the manuscript. All authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Abbreviations:
|
1α,25(OH)2D3
|
1α,25-dihydroxyvitamin D3
|
|
20(OH) D3
|
20S-hydroxyvitamin D3
|
|
21(OH)pD
|
21-hydroxypregnacalciferol
|
|
25(OH)D3
|
25-hydroxyvitamin D3
|
|
CAT
|
catalase
|
|
CCCP
|
carbonyl cyanide
3-chlorophenylhydrazone
|
|
PDIA3
|
protein disulfide-isomerase A3
|
|
PDOX
|
patient-derived orthotropic
xenograft
|
|
ROS
|
reactive oxygen species
|
|
SOD1
|
superoxide dismutase 1
|
|
SOD2
|
superoxide dismutase 2
|
|
SRB
|
sulphorhodamine B
|
|
UV
|
ultraviolet
|
|
VDR
|
vitamin D receptor
|
|
Δψm
|
mitochondrial membrane potential
|
References
|
1
|
Piotrowska A, Wierzbicka J and Żmijewski
MA: Vitamin D in the skin physiology and pathology. Acta Biochim
Pol. 63:17–29. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Holick MF: Vitamin D: Evolutionary,
physiological and health perspectives. Curr Drug Targets. 12:4–18.
2011. View Article : Google Scholar
|
|
3
|
Bikle DD: Vitamin D and the skin:
Physiology and pathophysiology. Rev Endocr Metab Disord. 13:3–19.
2012. View Article : Google Scholar
|
|
4
|
Samuel S and Sitrin MD: Vitamin D’s role
in cell proliferation and differentiation. Nutr Rev. 66(Suppl 2):
S116–S124. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bikle DD: Vitamin D regulated keratinocyte
differentiation. J Cell Biochem. 92:436–444. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kubis AM and Piwowar A: The new insight on
the regulatory role of the vitamin D3 in metabolic pathways
characteristic for cancerogenesis and neurodegenerative diseases.
Ageing Res Rev. 24(Pt B): 126–137. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Slominski AT, Janjetovic Z, Kim TK,
Wasilewski P, Rosas S, Hanna S, Sayre RM, Dowdy JC, Li W and Tuckey
RC: Novel non-calcemic secosteroids that are produced by human
epidermal keratinocytes protect against solar radiation. J Steroid
Biochem Mol Biol. 148:52–63. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gordon-Thomson C, Gupta R, Tongkao-on W,
Ryan A, Halliday GM and Mason RS: 1α,25 dihydroxyvitamin D3
enhances cellular defences against UV-induced oxidative and other
forms of DNA damage in skin. Photochem Photobiol Sci. 11:1837–1847.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fedirko V, Bostick RM, Long Q, Flanders
WD, McCullough ML, Sidelnikov E, Daniel CR, Rutherford RE and
Shaukat A: Effects of supplemental vitamin D and calcium on
oxidative DNA damage marker in normal colorectal mucosa: A
randomized clinical trial. Cancer Epidemiol Biomarkers Prev.
19:280–291. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jiang YJ, Teichert AE, Fong F, Oda Y and
Bikle DD: 1α,25(OH)2-dihydroxyvitamin D3/VDR protects the skin from
UVB-induced tumor formation by interacting with the β-catenin
pathway. J Steroid Biochem Mol Biol. 136:229–232. 2013. View Article : Google Scholar
|
|
11
|
Dixon KM, Deo SS, Wong G, Slater M, Norman
AW, Bishop JE, Posner GH, Ishizuka S, Halliday GM, Reeve VE, et al:
Skin cancer prevention: A possible role of 1,25dihydroxyvitamin D3
and its analogs. J Steroid Biochem Mol Biol. 97:137–143. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wong G, Gupta R, Dixon KM, Deo SS, Choong
SM, Halliday GM, Bishop JE, Ishizuka S, Norman AW, Posner GH, et
al: 1,25-Dihydroxyvitamin D and three low-calcemic analogs decrease
UV-induced DNA damage via the rapid response pathway. J Steroid
Biochem Mol Biol. 89–90:567–570. 2004. View Article : Google Scholar
|
|
13
|
Bikle DD, Jiang Y, Nguyen T, Oda Y and Tu
CL: Disruption of Vitamin D and Calcium Signaling in Keratinocytes
Predisposes to Skin Cancer. Front Physiol. 7:2962016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Holick MF: Sunlight, ultraviolet
radiation, vitamin D and skin cancer: How much sunlight do we need?
Adv Exp Med Biol. 810:1–16. 2014.PubMed/NCBI
|
|
15
|
Slominski AT, Brożyna AA, Skobowiat C,
Zmijewski MA, Kim TK, Janjetovic Z, Oak AS, Jozwicki W, Jetten AM,
Mason RS, et al: On the role of classical and novel forms of
vitamin D in melanoma progression and management. J Steroid Biochem
Mol Biol. 177:159–170. 2018. View Article : Google Scholar :
|
|
16
|
Yin L, Ordóñez-Mena JM, Chen T, Schöttker
B, Arndt V and Brenner H: Circulating 25-hydroxyvitamin D serum
concentration and total cancer incidence and mortality: A
systematic review and meta-analysis. Prev Med. 57:753–764. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Reichrath J, Zouboulis CC, Vogt T and
Holick MF: Targeting the vitamin D endocrine system (VDES) for the
management of inflammatory and malignant skin diseases: An
historical view and outlook. Rev Endocr Metab Disord. 17:405–417.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Brożyna AA, Jóźwicki W, Janjetovic Z and
Slominski AT: Expression of the vitamin D-activating enzyme
1α-hydroxylase (CYP27B1) decreases during melanoma progression. Hum
Pathol. 44:374–387. 2013. View Article : Google Scholar
|
|
19
|
Brożyna AA, Jóźwicki W and Slominski AT:
Decreased VDR expression in cutaneous melanomas as marker of tumor
progression: New data and analyses. Anticancer Res. 34:2735–2743.
2014.
|
|
20
|
Ma Y, Trump DL and Johnson CS: Vitamin D
in combination cancer treatment. J Cancer. 1:101–107. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Krishnan AV, Swami S and Feldman D:
Equivalent anticancer activities of dietary vitamin D and
calcitriol in an animal model of breast cancer: Importance of
mammary CYP27B1 for treatment and prevention. J Steroid Biochem Mol
Biol. 136:289–295. 2013. View Article : Google Scholar :
|
|
22
|
Morris HA: Vitamin D activities for health
outcomes. Ann Lab Med. 34:181–186. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mitchell D: The relationship between
vitamin D and cancer. Clin J Oncol Nurs. 15:557–560. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Welsh J, Wietzke JA, Zinser GM, Byrne B,
Smith K and Narvaez CJ: Vitamin D-3 receptor as a target for breast
cancer prevention. J Nutr. 133(Suppl): 2425S–2433S. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lamprecht SA and Lipkin M: Chemoprevention
of colon cancer by calcium, vitamin D and folate: Molecular
mechanisms. Nat Rev Cancer. 3:601–614. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bolland MJ, Grey A, Gamble GD and Reid IR:
Calcium and vitamin D supplements and health outcomes: A reanalysis
of the Women’s Health Initiative (WHI) limited-access data set. Am
J Clin Nutr. 94:1144–1149. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Grant WB: 25-hydroxyvitamin D and breast
cancer, colorectal cancer, and colorectal adenomas: Case-control
versus nested case-control studies. Anticancer Res. 35:1153–1160.
2015.PubMed/NCBI
|
|
28
|
Płudowski P, Karczmarewicz E, Bayer M,
Carter G, Chlebna-Sokół D, Czech-Kowalska J, Dębski R, Decsi T,
Dobrzańska A, Franek E, et al: Practical guidelines for the
supplementation of vitamin D and the treatment of deficits in
Central Europe - recommended vitamin D intakes in the general
population and groups at risk of vitamin D deficiency. Endokrynol
Pol. 64:319–327. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Skobowiat C, Oak AS, Kim TK, Yang CH,
Pfeffer LM, Tuckey RC and Slominski AT: Noncalcemic
20-hydroxyvitamin D3 inhibits human melanoma growth in in vitro and
in vivo models. Oncotarget. 8:9823–9834. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Slominski AT, Brożyna AA, Zmijewski MA,
Jóźwicki W, Jetten AM, Mason RS, Tuckey RC and Elmets CA: Vitamin D
signaling and melanoma: Role of vitamin D and its receptors in
melanoma progression and management. Lab Invest. 97:706–724. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ma Y, Yu WD, Trump DL and Johnson CS:
1,25D3 enhances antitumor activity of gemcitabine and cisplatin in
human bladder cancer models. Cancer. 116:3294–3303. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Rassnick KM, Muindi JR, Johnson CS,
Balkman CE, Ramnath N, Yu WD, Engler KL, Page RL and Trump DL: In
vitro and in vivo evaluation of combined calcitriol and cisplatin
in dogs with spontaneously occurring tumors. Cancer Chemother
Pharmacol. 62:881–891. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wietrzyk J, Nevozhay D, Filip B, Milczarek
M and Kutner A: The antitumor effect of lowered doses of
cytostatics combined with new analogs of vitamin D in mice.
Anticancer Res. 27(5A): 3387–3398. 2007.PubMed/NCBI
|
|
34
|
Podgorska E, Drzal A, Matuszak Z, Swakon
J, Slominski A, Elas M and Urbanska K: Calcitriol and Calcidiol Can
Sensitize Melanoma Cells to Low(−)LET Proton Beam Irradiation. Int
J Mol Sci. 19:192018. View Article : Google Scholar
|
|
35
|
Piotrowska A, Wierzbicka J, Ślebioda T,
Woźniak M, Tuckey RC, Slominski AT and Żmijewski MA: Vitamin D
derivatives enhance cytotoxic effects of H2O2 or cisplatin on human
keratinocytes. Steroids. 110:49–61. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Saw RP, Armstrong BK, Mason RS, Morton RL,
Shannon KF, Spillane AJ, Stretch JR and Thompson JF: Adjuvant
therapy with high dose vitamin D following primary treatment of
melanoma at high risk of recurrence: A placebo controlled
randomised phase II trial (ANZMTG 02.09 Mel-D). BMC Cancer.
14:7802014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Pettijohn E, Martone B, Rademaker A and
Kuzel T: A phase I study of high-dose calcitriol in combination
with temozolomide for patients with metastatic melanoma. J Pers
Med. 4:448–458. 2014. View Article : Google Scholar
|
|
38
|
Batus M, Waheed S, Ruby C, Petersen L,
Bines SD and Kaufman HL: Optimal management of metastatic melanoma:
Current strategies and future directions. Am J Clin Dermatol.
14:179–194. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Shah DJ and Dronca RS: Latest advances in
chemotherapeutic, targeted, and immune approaches in the treatment
of metastatic melanoma. Mayo Clin Proc. 89:504–519. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Arnold M, de Vries E, Whiteman DC, Jemal
A, Bray F, Parkin DM and Soerjomataram I: Global burden of
cutaneous melanoma attributable to ultraviolet radiation in 2012.
Int J Cancer. 143:1305–1314. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Mattia G, Puglisi R, Ascione B, Malorni W,
Carè A and Matarrese P: Cell death-based treatments of
melanoma:conventional treatments and new therapeutic strategies.
Cell Death Dis. 9:1122018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Slominski AT and Carlson JA: Melanoma
resistance: A bright future for academicians and a challenge for
patient advocates. Mayo Clin Proc. 89:429–433. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Russo A, Ficili B, Candido S, Pezzino FM,
Guarneri C, Biondi A, Travali S, McCubrey JA, Spandidos DA and
Libra M: Emerging targeted therapies for melanoma treatment
(Review). Int J Oncol. 45:516–524. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Siegel RL, Miller KD and Jemal A: Cancer
Statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Johnson DB, Pollack MH and Sosman JA:
Emerging targeted therapies for melanoma. Expert Opin Emerg Drugs.
21:195–207. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Slominski AT, Kim TK, Li W, Yi AK,
Postlethwaite A and Tuckey RC: The role of CYP11A1 in the
production of vitamin D metabolites and their role in the
regulation of epidermal functions. J Steroid Biochem Mol Biol.
144(Pt A): 28–39. 2014. View Article : Google Scholar
|
|
47
|
Slominski AT, Janjetovic Z, Fuller BE,
Zmijewski MA, Tuckey RC, Nguyen MN, Sweatman T, Li W, Zjawiony J,
Miller D, et al: Products of vitamin D3 or 7-dehydrocholesterol
metabolism by cytochrome P450scc show anti-leukemia effects, having
low or absent calcemic activity. PLoS One. 5:e99072010. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Wang J, Slominski A, Tuckey RC, Janjetovic
Z, Kulkarni A, Chen J, Postlethwaite AE, Miller D and Li W:
20-hydroxyvitamin D3 inhibits proliferation of cancer
cells with high efficacy while being non-toxic. Anticancer Res.
32:739–746. 2012.PubMed/NCBI
|
|
49
|
Wasiewicz T, Szyszka P, Cichorek M,
Janjetovic Z, Tuckey RC, Slominski AT and Zmijewski MA: Antitumor
effects of vitamin d analogs on hamster and mouse melanoma cell
lines in relation to melanin pigmentation. Int J Mol Sci.
16:6645–6667. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Zmijewski MA, Li W, Chen J, Kim TK,
Zjawiony JK, Sweatman TW, Miller DD and Slominski AT: Synthesis and
photochemical transformation of 3β,21-dihydroxypregna-5,7-die
n-20-one to novel secosteroids that show anti-melanoma activity.
Steroids. 76:193–203. 2011. View Article : Google Scholar
|
|
51
|
Koul PA, Ahmad SH, Ahmad F, Jan RA, Shah
SU and Khan UH: Vitamin d toxicity in adults: A case series from an
area with endemic hypovitaminosis d. Oman Med J. 26:201–204. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Podgorska E, Sniegocka M, Mycinska M,
Trybus W, Trybus E, Kopacz-Bednarska A, Wiechec O, Krzykawska-Serda
M, Elas M, Krol T, et al: Acute hepatologic and nephrologic effects
of calcitriol in Syrian golden hamster (Mesocricetus auratus). Acta
Biochim Pol. 65:351–358. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Slominski AT, Kim TK, Shehabi HZ, Semak I,
Tang EK, Nguyen MN, Benson HA, Korik E, Janjetovic Z, Chen J, et
al: In vivo evidence for a novel pathway of vitamin D3
metabolism initiated by P450scc and modified by CYP27B1. FASEB J.
26:3901–3915. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Slominski AT, Kim TK, Li W, Postlethwaite
A, Tieu EW, Tang EK and Tuckey RC: Detection of novel
CYP11A1-derived secosteroids in the human epidermis and serum and
pig adrenal gland. Sci Rep. 5:148752015. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Slominski AT, Li W, Kim TK, Semak I, Wang
J, Zjawiony JK and Tuckey RC: Novel activities of CYP11A1 and their
potential physiological significance. J Steroid Biochem Mol Biol.
151:25–37. 2015. View Article : Google Scholar
|
|
56
|
Dasari S and Tchounwou PB: Cisplatin in
cancer therapy: Molecular mechanisms of action. Eur J Pharmacol.
740:364–378. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Marullo R, Werner E, Degtyareva N, Moore
B, Altavilla G, Ramalingam SS and Doetsch PW: Cisplatin induces a
mitochondrial-ROS response that contributes to cytotoxicity
depending on mitochondrial redox status and bioenergetic functions.
PLoS One. 8:e811622013. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Pourahmad J, Amirmostofian M, Kobarfard F
and Shahraki J: Biological reactive intermediates that mediate
dacarbazine cytotoxicity. Cancer Chemother Pharmacol. 65:89–96.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Slominski A, Semak I, Zjawiony J, Wortsman
J, Li W, Szczesniewski A and Tuckey RC: The cytochrome P450scc
system opens an alternate pathway of vitamin D3 metabolism. FEBS J.
272:4080–4090. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Aykul S and Martinez-Hackert E:
Determination of half-maximal inhibitory concentration using
biosensor-based protein interaction analysis. Anal Biochem.
508:97–103. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Kalliokoski T, Kramer C, Vulpetti A and
Gedeck P: Comparability of mixed IC50 data - a
statistical analysis. PLoS One. 8:e610072013. View Article : Google Scholar
|
|
62
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
63
|
Slominski AT, Janjetovic Z, Kim TK, Wright
AC, Grese LN, Riney SJ, Nguyen MN and Tuckey RC: Novel vitamin D
hydroxy-derivatives inhibit melanoma growth and show differential
effects on normal melanocytes. Anticancer Res. 32:3733–3742.
2012.PubMed/NCBI
|
|
64
|
Janjetovic Z, Brozyna AA, Tuckey RC, Kim
TK, Nguyen MN, Jozwicki W, Pfeffer SR, Pfeffer LM and Slominski AT:
High basal NF-κB activity in nonpigmented melanoma cells is
associated with an enhanced sensitivity to vitamin D3 derivatives.
Br J Cancer. 105:1874–1884. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Corazao-Rozas P, Guerreschi P, Jendoubi M,
André F, Jonneaux A, Scalbert C, Garçon G, Malet-Martino M,
Balayssac S, Rocchi S, et al: Mitochondrial oxidative stress is the
Achille’s heel of melanoma cells resistant to Braf-mutant
inhibitor. Oncotarget. 4:1986–1998. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Florea AM and Büsselberg D: Cisplatin as
an anti-tumor drug: Cellular mechanisms of activity, drug
resistance and induced side effects. Cancers (Basel). 3:1351–1371.
2011. View Article : Google Scholar
|
|
67
|
Le TYL, Ogawa M, Kizana E, Gunton JE and
Chong JJH: Vitamin D Improves Cardiac Function After Myocardial
Infarction Through Modulation of Resident Cardiac Progenitor Cells.
Heart Lung Circ. 27:967–975. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Cai L, Luo L, Tang Z and Meng X: Combined
antitumor effects of 1,25 dihydroxy vitamin D3 and Notch inhibitor
in liver cancer. Oncol Rep. 40:1515–1524. 2018.PubMed/NCBI
|
|
69
|
Corachan A, Ferrero H, Aguilar A, Garcia
N, Monleon J, Faus A, Cervelló I and Pellicer A: Inhibition of
tumor cell proliferation in human uterine leiomyomas by vitamin D
via Wnt/β-catenin pathway. Fertil Steril. 111:397–407. 2019.
View Article : Google Scholar
|
|
70
|
Linos E, Swetter SM, Cockburn MG, Colditz
GA and Clarke CA: Increasing burden of melanoma in the United
States. J Invest Dermatol. 129:1666–1674. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Slominski AT, Zmijewski MA, Plonka PM,
Szaflarski JP and Paus R: How UV Light Touches the Brain and
Endocrine System Through Skin, and Why. Endocrinology.
159:1992–2007. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Haussler MR, Jurutka PW, Mizwicki M and
Norman AW: Vitamin D receptor (VDR)-mediated actions of
1α,25(OH)2 vitamin D3: genomic and
non-genomic mechanisms. Best Pract Res Clin Endocrinol Metab.
25:543–559. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Moukayed M and Grant WB: Molecular link
between vitamin D and cancer prevention. Nutrients. 5:3993–4021.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Wacker M and Holick MF: Sunlight and
Vitamin D: A global perspective for health. Dermatoendocrinol.
5:51–108. 2013. View Article : Google Scholar
|
|
75
|
Timerman D, McEnery-Stonelake M, Joyce CJ,
Nambudiri VE, Hodi FS, Claus EB, Ibrahim N and Lin JY: Vitamin D
deficiency is associated with a worse prognosis in metastatic
melanoma. Oncotarget. 8:6873–6882. 2017. View Article : Google Scholar :
|
|
76
|
Caini S, Boniol M, Tosti G, Magi S, Medri
M, Stanganelli I, Palli D, Assedi M, Marmol VD and Gandini S:
Vitamin D and melanoma and non-melanoma skin cancer risk and
prognosis: A comprehensive review and meta-analysis. Eur J Cancer.
50:2649–2658. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Wyatt C, Lucas RM, Hurst C and Kimlin MG:
Vitamin D deficiency at melanoma diagnosis is associated with
higher Breslow thickness. PLoS One. 10:e01263942015. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Ogbah Z, Visa L, Badenas C, Ríos J,
Puig-Butille JA, Bonifaci N, Guino E, Augé JM, Kolm I, Carrera C,
et al: Serum 25-hydroxyvitamin D3 levels and vitamin D receptor
variants in melanoma patients from the Mediterranean area of
Barcelona. BMC Med Genet. 14:262013. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Sinnberg T, Levesque MP, Krochmann J,
Cheng PF, Ikenberg K, Meraz-Torres F, Niessner H, Garbe C and Busch
C: Wnt-signaling enhances neural crest migration of melanoma cells
and induces an invasive phenotype. Mol Cancer. 17:592018.
View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Piotrowska A, Wierzbicka J, Nadkarni S,
Brown G, Kutner A and Żmijewski MA: Antiproliferative Activity of
Double Point Modified Analogs of 1,25-Dihydroxyvitamin
D2 Against Human Malignant Melanoma Cell Lines. Int J
Mol Sci. 17:172016. View Article : Google Scholar
|
|
81
|
Wasiewicz T, Piotrowska A, Wierzbicka J,
Slominski AT and Zmijewski MA: Antiproliferative Activity of
Non-Calcemic Vitamin D Analogs on Human Melanoma Lines in Relation
to VDR and PDIA3 Receptors. Int J Mol Sci. 19:192018. View Article : Google Scholar
|
|
82
|
Field S and Newton-Bishop JA: Melanoma and
vitamin D. Mol Oncol. 5:197–214. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Szyszka P, Zmijewski MA and Slominski AT:
New vitamin D analogs as potential therapeutics in melanoma. Expert
Rev Anticancer Ther. 12:585–599. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Al-Qatati A and Aliwaini S: Combined
pitavastatin and dacarbazine treatment activates apoptosis and
autophagy resulting in synergistic cytotoxicity in melanoma cells.
Oncol Lett. 14:7993–7999. 2017.
|
|
85
|
Del Bello B, Toscano M, Moretti D and
Maellaro E: Cisplatin-induced apoptosis inhibits autophagy, which
acts as a pro-survival mechanism in human melanoma cells. PLoS One.
8:e572362013. View Article : Google Scholar
|
|
86
|
Kissel CK, Schadendorf D and Röckmann H:
The altered apoptotic pathways in cisplatin and etoposide-resistant
melanoma cells are drug specific. Melanoma Res. 16:527–535. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Wangpaichitr M, Wu C, Li YY, Nguyen DJM,
Kandemir H, Shah S, Chen S, Feun LG, Prince JS, Kuo MT, et al:
Exploiting ROS and metabolic differences to kill cisplatin
resistant lung cancer. Oncotarget. 8:49275–49292. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Goldstein I, Rivlin N, Shoshana OY, Ezra
O, Madar S, Goldfinger N and Rotter V: Chemotherapeutic agents
induce the expression and activity of their clearing enzyme CYP3A4
by activating p53. Carcinogenesis. 34:190–198. 2013. View Article : Google Scholar
|
|
89
|
Ortiz de Montellano PR: Cytochrome
P450-activated prodrugs. Future Med Chem. 5:213–228. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Lu Y and Cederbaum AI: Cisplatin-induced
hepatotoxicity is enhanced by elevated expression of cytochrome
P450 2E1. Toxicol Sci. 89:515–523. 2006. View Article : Google Scholar
|
|
91
|
Wierzbicka J, Piotrowska A and Zmijewski
MA: The renaissance of vitamin D. Acta Biochim Pol. 61:679–686.
2014. View Article : Google Scholar
|
|
92
|
Box NF, Vukmer TO and Terzian T: Targeting
p53 in melanoma. Pigment Cell Melanoma Res. 27:8–10. 2014.
View Article : Google Scholar :
|
|
93
|
Slominski AT, Kim TK, Hobrath JV, Oak ASW,
Tang EKY, Tieu EW, Li W, Tuckey RC and Jetten AM: Endogenously
produced nonclassical vitamin D hydroxy-metabolites act as ‘biased’
agonists on VDR and inverse agonists on RORα and RORγ. J Steroid
Biochem Mol Biol. 173:42–56. 2017. View Article : Google Scholar
|
|
94
|
Slominski AT, Kim TK, Takeda Y, Janjetovic
Z, Brozyna AA, Skobowiat C, Wang J, Postlethwaite A, Li W, Tuckey
RC, et al: RORα and ROR γ are expressed in human skin and serve as
receptors for endogenously produced noncalcemic 20-hydroxy- and
20,23-dihydroxyvitamin D. FASEB J. 28:2775–2789. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Lin Z, Marepally SR, Goh ESY, Cheng CYS,
Janjetovic Z, Kim TK, Miller DD, Postlethwaite AE, Slominski AT,
Tuckey RC, et al: Investigation of 20S-hydroxyvitamin D3 analogs
and their 1α-OH derivatives as potent vitamin D receptor agonists
with anti-inflammatory activities. Sci Rep. 8:14782018. View Article : Google Scholar
|
|
96
|
Lin Z, Chen H, Belorusova AY, Bollinger
JC, Tang EKY, Janjetovic Z, Kim TK, Wu Z, Miller DD, Slominski AT,
et al: 1α,20S-Dihydroxyvitamin D3 Interacts with Vitamin D
Receptor: Crystal Structure and Route of Chemical Synthesis. Sci
Rep. 7:101932017. View Article : Google Scholar
|
|
97
|
Slominski AT, Kim TK, Janjetovic Z,
Brożyna AA, Żmijewski MA, Xu H, Sutter TR, Tuckey RC, Jetten AM and
Crossman DK: Differential and Overlapping Effects of
20,23(OH)2 D3 Epidermal Keratinocytes:
Identification of AhR as an Alternative and
1,25(OH)2D3 on Gene Expression in Human
Receptor for 20,23(OH)2D3. Int J Mol Sci. 19:192018.
View Article : Google Scholar
|
|
98
|
Kim TK, Wang J, Janjetovic Z, Chen J,
Tuckey RC, Nguyen MN, Tang EK, Miller D, Li W and Slominski AT:
Correlation between secosteroid-induced vitamin D receptor activity
in melanoma cells and computer-modeled receptor binding strength.
Mol Cell Endocrinol. 361:143–152. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Barutello G, Rolih V, Arigoni M, Tarone L,
Conti L, Quaglino E, Buracco P, Cavallo F and Riccardo F: Strengths
and Weaknesses of Pre-Clinical Models for Human Melanoma Treatment:
Dawn of Dogs’ Revolution for Immunotherapy. Int J Mol Sci.
19:192018. View Article : Google Scholar
|
|
100
|
Kuzu OF, Nguyen FD, Noory MA and Sharma A:
Current State of Animal (Mouse) Modeling in Melanoma Research.
Cancer Growth Metastasis. 8(Suppl 1): 81–94. 2015.PubMed/NCBI
|
|
101
|
Hoffman RM: Patient-Derived Orthotopic
Xenograft (PDOX) Models of Melanoma. Int J Mol Sci. 18:182017.
View Article : Google Scholar
|
|
102
|
Kawaguchi K, Han Q, Li S, Tan Y, Igarashi
K, Miyake K, Kiyuna T, Miyake M, Chemielwski B, Nelson SD, et al:
Intra-tumor L-methionine level highly correlates with tumor size in
both pancreatic cancer and melanoma patient-derived orthotopic
xenograft (PDOX) nude-mouse models. Oncotarget. 9:11119–11125.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Kawaguchi K, Igarashi K, Li S, Han Q, Tan
Y, Miyake K, Kiyuna T, Miyake M, Murakami T, Chmielowski B, et al:
Recombinant methioninase (rMETase) is an effective therapeutic for
BRAF-V600E-negative as well as -positive melanoma in
patient-derived orthotopic xenograft (PDOX) mouse models.
Oncotarget. 9:915–923. 2017.
|