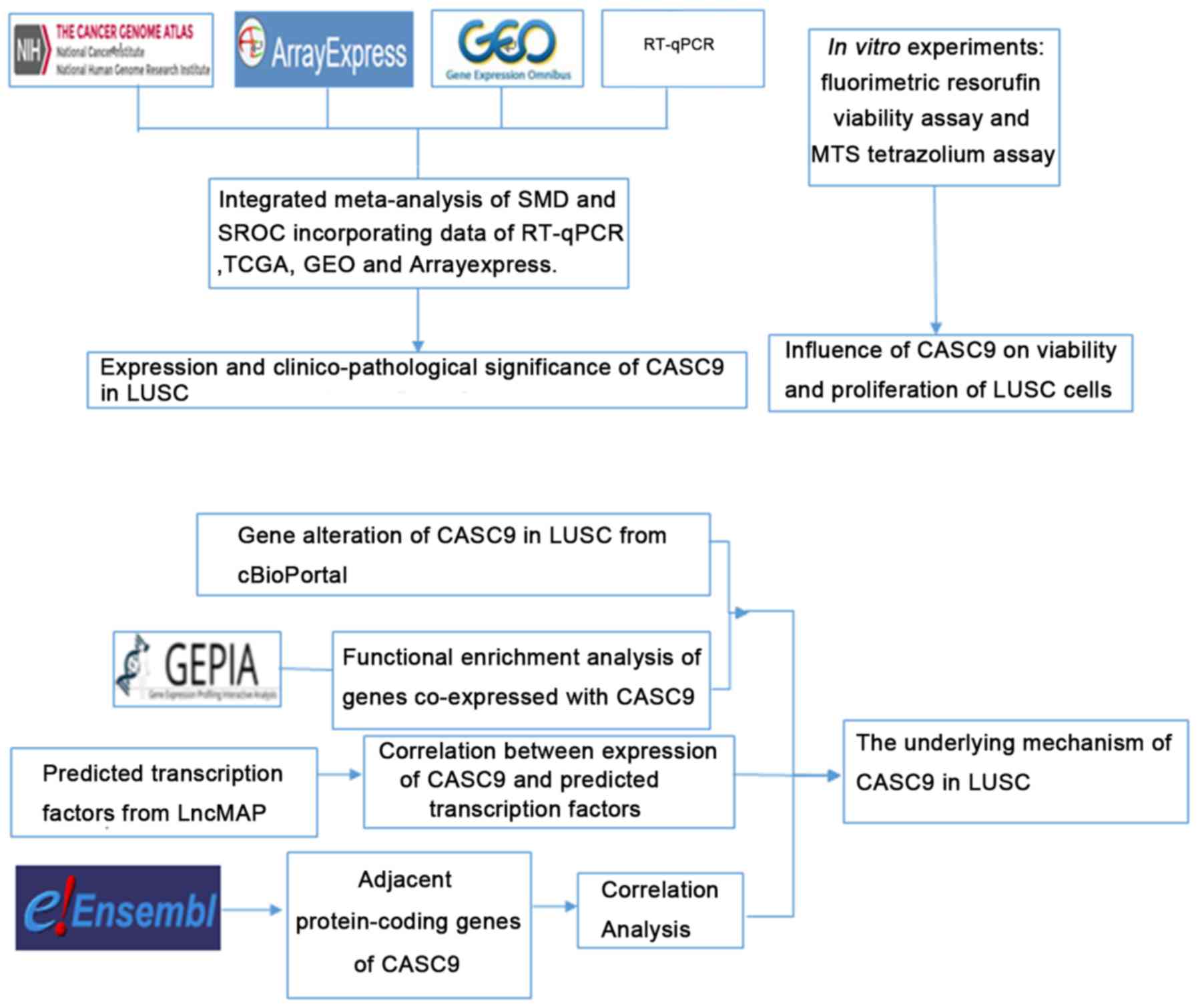
Introduction
Lung cancer (LC) is one of the most common malignant
tumors in the world (1,2). According to data compiled by the
American Cancer Society, there were 234,030 estimated new LC cases
and 154,050 estimated LC-associated mortalities in the USA in 2018
(3). In terms of histology, there
are two types of LC: Non-small cell LC (NSCLC) and small cell LC,
of which the former makes up 85% of all cases (4). NSCLC is comprised of two subtypes:
Lung squamous cell carcinoma (LUSC) and lung adenocarcinoma (LUAD)
(3,5). Drugs targeting mutated versions of
the epidermal growth factor receptor (EGFR), GTPase KRas and ALK
tyrosine kinase receptor proteins mutations have already
demonstrated beneficial effects in patients with LUAD (6). However, these drugs are not
applicable to LUSC, which is associated with poor survival rates
(7), highlighting the urgent
requirement for effective diagnostic and therapeutic targets for
LUSC.
Long non-coding RNAs (lncRNAs) are a class of
non-protein-coding RNAs that control gene expression in complex
ways (8,9). Numerous studies have revealed that
the ectopic expression of lncRNAs is implicated in human cancer
(10-13). As a member of the lncRNA family,
the cancer susceptibility candidate 9 (CASC9) gene, located on
chromosome 8q21.11, was originally observed to be upregulated in
esophageal squamous cell carcinoma (ESCC) (14). In addition, CASC9 was reported to
constitute a crucial component in the tumorigenesis of other types
of cancer, including gastric and nasopharyngeal cancer (15,16).
With regard to NSCLC, recent studies have discovered that CASC9 was
associated with the resistance of NSCLC cells to EGFR tyrosine
kinase inhibitors, and that it served important roles in the
proliferation and metastasis of LUAD cells (17,18).
Although studies on CASC9 in NSCLC have reported promising results,
they have mainly focused on LUAD, and studies on the role of CASC9
in LUSC are lacking.
In the present study, the clinicopathological effect
of CASC9 in LUSC was investigated using reverse
transcription-quantitative polymerase chain reaction (RT-qPCR) and
data mining of public databases, including The Cancer Genome Atlas
(TCGA), the Gene Expression Omnibus (GEO), ArrayExpress, and the
Cancer Cell Line Encyclopedia (CCLE). Furthermore, the functional
influence of CASC9 on LUSC, and the underlying mechanism, were
explored through in vitro experiments, investigation of the
mutation status of CASC9 in LUSC from cBioPortal, functional
enrichment analysis of co-expressed genes using Gene Expression
Profiling Interactive Analysis (GEPIA), prediction of potential
transcription factors, and inspection of adjacent protein-coding
genes. The design of the current study is illustrated in Fig. 1.
Materials and methods
Tissue samples
A total of 20 patients first diagnosed with LUSC
(age range, 35-68 years; mean, 51 years; 12 male and 8 female)
attending the First Affiliated Hospital of Guangxi Medical
University (Nanning, China) were included in the present study. The
patients were receiving no medication and underwent radical
resection of lung cancer between August 2017 and March 2018. Tumor
and matched non-cancer lung tissues were obtained from these
patients during the radical resection surgery. The samples were
fixed in 10% buffered formalin under ambient temperature for 16 h
and paraffin-embedded. All patients provided signed informed
consent and approval of this study was granted by the Ethics
Committee of the First Affiliated Hospital of Guangxi Medical
University.
RT-qPCR
The isolation and relative quantification of RNA was
conducted based on methods described previously (19). RT and qPCR kits were used to
examine the CASC9 expression levels in LUSC and adjacent non-cancer
tissues, following the manufacturers' protocols. qPCR was performed
on an ABI 7900 Real-time PCR System (Applied Biosystems; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) using the
LightCycler® FastStart DNA Master plus SYBR Green kit
(Roche Diagnostics, Basel, Switzerland). The denaturation,
annealing and extension steps of the 40 PCR cycles were set at 95°C
for 10 sec, 60°C for 5 sec and 72°C for 5 sec, following a 10-min
hot start at 95°C. The primers for CASC9 and the reference gene
GAPDH were as follows: CASC9 forward, 5′-AAAACCAGGTGGGACCCAGA-3′;
reverse, 5′-TGATCAGAAGAAGAGGGGCA-3′; GAPDH forward,
5′-ACCCACTCCTCCACCTTTG-3′; and reverse, 5′-CTCTTGTGCTCTTGCTGGG-3′.
CASC9 expression was calculated according to the formula:
2−ΔCq= 2−(CqCASC9-CqGAPDH), where the Cq
value is the quantification cycle number (20).
Evaluation of the clinicopathological
associations of CASC9 in LUSC, using TCGA data
TCGA (http://cancergenome.nih.gov/), an extensive reservoir
of DNA methylation, exome sequencing, single nucleotide
polymorphism array, RNA-seq and microRNA-seq data (21,22),
assists researchers in conducting studies using complicated cancer
genomics profiles. In the present study,
log2(x+1)-transformed level 3 transcripts per million
reads (TPM) RNA-seq data of CASC9 expression in 501 LUSC and 49
adjacent normal tissues as well as the clinicopathological data of
the 501 patients from whom the samples were obtained (TCGA-LUSC),
were collected from TCGA data portal (https://portal.gdc.cancer.gov/) and analyzed using
SPSS version 22.0 (IBM Corp., Armonk, NY, USA).
Integrated meta-analysis of CASC9
expression in LUSC and non-cancer tissues
To comprehensively appraise the differential
expression of CASC9 in LUSC and non-cancer tissues, expression data
of CASC9 in LUSC and non-cancer tissues published before September
19, 2018 were searched in the GEO (https://www.ncbi.nlm.nih.gov/gds/), ArrayExpress
(https://www.ebi.ac.uk/arrayexpress/)
and literature databases, including Pubmed (https://www.ncbi.nlm.nih.gov/pubmed), Google Scholar
(https://scholar.google.com/), Wiley
Online Library (https://onlinelibrary.wiley.com/), Cochrane Library
(https://www.cochranelibrary.com/library), Web of
Science (http://apps.webofknowledge.com), Embase (https://www.embase.com), Ebsco (https://www.ebsco.com/), Chinese VIP (http://www.cqvip.com/), China National Knowledge
Infrastructure (http://www.cnki.net/), Sinomed
(http://www.sinomed.ac.cn/) and Wang Fang
(http://www.wanfangdata.com.cn/index.html). The search
terms used in the GEO were: ('Lung neoplasms'[Mesh]) AND
('microarray analysis'[Mesh] OR 'tissue array analysis'[Mesh] OR
'transcriptome'[Mesh] OR 'sequence analysis, RNA'[Mesh] OR
'high-throughput nucleotide sequencing'[Mesh]). The key words for
searching in ArrayExpress were: ('Cancer' OR 'carcinoma' OR
'adenocarcinoma' OR 'tumour' OR 'tumor' OR 'malignanc*' OR
'neoplas*') AND ('lung' OR 'pulmonary' OR 'respiratory' OR
'respiration' OR 'aspiration' OR 'bronchi' OR 'bronchioles' OR
'alveoli' OR 'pneumocytes' OR 'air way'). Regarding the literature
survey, the search terms used were as follows: ('CASC9' OR 'cancer
susceptibility 9' OR 'ESSCAL1' OR 'ESCCAL-1' OR 'LINC0098') AND
('lung' OR 'pulmonary' OR 'respiratory' OR 'respiration' OR
'aspiration' OR 'bronchi' OR 'bronchioles' OR 'alveoli' OR
'pneumocytes' OR 'air way') AND ('cancer' OR 'carcinoma' OR
'adenocarcinoma' OR 'tumour' OR 'tumor' OR 'malignanc*' OR
'neoplas*'). Studies using human-derived samples offering
sufficient CASC9 expression data (where the numbers of LUSC and
non-cancer cases exceeded 3) in LUSC and non-cancer samples for the
calculation of a standardized mean difference (SMD) were included.
Basic information, as well as expression and diagnostic data,
including accession ID, first author, publication year, country,
experiment type, sample type, platform, number of cases in the
cancer or non-cancer groups, mean ± SD of CASC9 expression in the
cancer or non-cancer groups, true positives, false positives, false
negatives and true negatives, were extracted from the included
studies according to methods described previously (23). An SMD with the 95% confidence
interval (CI) and summarized receiver operating characteristic
(SROC) curves were produced for the integrated meta-analysis,
consisting of the in-house RT-qPCR data, TCGA data and microarray
chip data from ArrayExpress, as described previously (23).
Cell line data of CASC9 expression from
CCLE
The expression data of CASC9 in all available LC
cell lines was downloaded from CCLE (https://portals.broadinstitute.org/ccle/about) on
October 5, 2018. The original data included 192 LC cell lines. Due
to contamination or misidentification of the PC-14 cell line being
reported by the International Cell Line Authentication Committee,
Database of Cross-Contaminated or Misidentified Cell Lines
(http://iclac.org/databases/cross-contaminations/),
it was excluded from the present expression analysis. Finally, the
expression data of CASC9 from a total of 191 cell lines were merged
into a heat map using GraphPad Prism 7 (GraphPad Software, Inc., La
Jolla, CA, USA).
Cell transfection and in vitro
experiments
The human LUSC H226 cells were purchased from the
American Type Culture Collection (Manassas, VA, USA), cultured in
Dulbecco's modified Eagle's medium (Gibco; Thermo Fisher
Scientific, Inc.) and supplemented with 10% fetal bovine serum
(Biological Industries, Kibbutz Beit Haemek, Israel) and
penicillin-streptomycin at 37°C under a humidified atmosphere of 5%
CO2. Each in vitro experiment was performed in
triplicate. The H226 cells were seeded into 96-well plates, at
2.5×103 cells per well, and incubated at 37°C for 24 h
prior to transfection. Transfections of mock control, scrambled
small interfering (si)RNAs, and CASC9 siRNAs (Ambion; Thermo Fisher
Scientific, Inc.) were performed in H226 cells at a concentration
of 200 nM for 96 h using the CombiMag Magnetofection™ transfection
kit (OZBiosciences SAS, Marseille, France), according to the
manufacturer's protocol. The siRNAs were designed for the best
efficiency to knockdown CASAC9 using the online tool InvivoGen
siRNA Wizard Software version 3.1 (https://www.invivogen.com/sirnawizard/index.php), and
4 siRNAs were finally selected. Scrambled siRNAs were included to
serve as negative controls for each of these. The siRNAs sequences
are listed in Table SI. The 4
selected CASC9 siRNAs were pooled for use in subsequent
experiments, as were the scrambled siRNAs. To investigate the
influence of CASC9 on the viability and proliferation of H226
cells, fluorimetric resorufin viability and MTS tetrazolium assays
were conducted as described previously (24-27).
cBioPortal gene alteration of CASC9 in
LUSC tissue
The gene alteration status of CASC9 in LUSC (dataset
TCGA, provisional) was acquired from cBioPortal version 2.0.1
(http://www.cbioportal.org) (28). The distribution of the CASC9
alteration in 179 sequenced patients with LUSC was visualized using
the OncoPrint module of cBioPortal.
Functional enrichment analysis of
co-expressed genes
Genes co-expressed with CASC9 were identified from
the GEPIA. The biological functions of the co-expressed genes were
investigated via the enrichment annotation modules in Metascape
(http://metascape.org) (Kyoto Encyclopedia of
Genes and Genomes Pathway, GO Biological Processes, Reactome
Pathway Database, Canonical Pathways, and CORUM). Terms with
P<0.01 and the number of enriched genes >3 were considered to
be significantly associated with the genes.
Prediction of potential transcription
factors for CASC9
Potential transcription factors associated with
CASC9 were identified by referring to a series of databases,
including starBase (http://starbase.sysu.edu.cn/), TransmiR (http://www.cuilab.cn/transmir), TRED (http://rulai.cshl.edu/TRED), ITFP (https://omictools.com/itfp-tool), TFe (http://www.cisreg.ca/tfe), AnimalTFDB (http://bioinfo.life.hust.edu.cn/AnimalTFDB/) and
LncMAP (http://bio-bigdata.hrbmu.edu.cn/LncMAP/). HT-Seq TPM
RNA-seq data of predicted transcription factors in LUSC were
downloaded from TCGA, and Pearson's correlation analysis was
performed in GraphPad Prism 7 to evaluate the correlation between
them and CASC9 expression.
Correlations between CASC9 and adjacent
protein-coding genes
Protein-coding genes adjacent to CASC9 were searched
using the Ensemble Genome Browser (http://grch37.ensembl.org/index.html) and the HT-Seq
TPM RNA-seq data of these genes in LUSC were downloaded from TCGA.
The association between CASC9 expression and that of the identified
adjacent protein-coding genes was analyzed by Pearson's correlation
tests in GraphPad Prism 7.
Statistical analysis
The statistical analyses for the RT-qPCR and TCGA
data were performed in SPSS version 22.0. The expression values of
CASC9 in LUSC and non-cancer tissues are presented as the mean ±
SD. Paired t-tests were conducted to compare the expression of
CASC9 between LUSC and non-cancer tissues, as derived from the
RT-qPCR assays. Regarding the clinicopathological effect of CASC9
expression in LUSC from TCGA data, the Mann-Whitney test was
employed to evaluate the differential expression of CASC9 in two
subgroups of clinicopathological variables. When a
clinicopathological variable contained >3 subgroups, the
Kruskal-Wallis test was performed. To assess the ability of CASC9
to distinguish LUSC from non-cancer tissue, ROC curves were
created. The discernment capacity of CASC9 for LUSC increased with
the area under the curves (AUC) varying from 0.5-1.0. The impact of
high versus low CASC9 expression (divided by the average CASC9
expression value) on the survival rate of patients with LUSC was
evaluated using the Kaplan-Meier survival curves. Multivariate Cox
regression analysis was performed to judge whether CASC9 expression
could serve as a prognostic indicator for LUSC independent of
clinical variables. Two-way analysis of variance and Bonferroni
post-tests were performed for the comparison of the groups in the
fluorimetric resorufin viability and MTS tetrazolium assays.
P<0.05 was considered to indicate statistically significant
differences.
Results
RT-qPCR
Fig. 2 illustrates
the finding that the expression level of CASC9 in LUSC tissues
(relative expression, 2.771±0.974) was significantly higher than
that in paired, non-cancer tissues (relative expression,
1.397±0.857) (P<0.001; Fig.
2A). ROC curves with an AUC value of 0.855 indicated the
diagnostic ability of CASC9 expression levels to distinguish LUSC
tissues from non-cancer tissues (P<0.001; Fig. 2A, right panel).
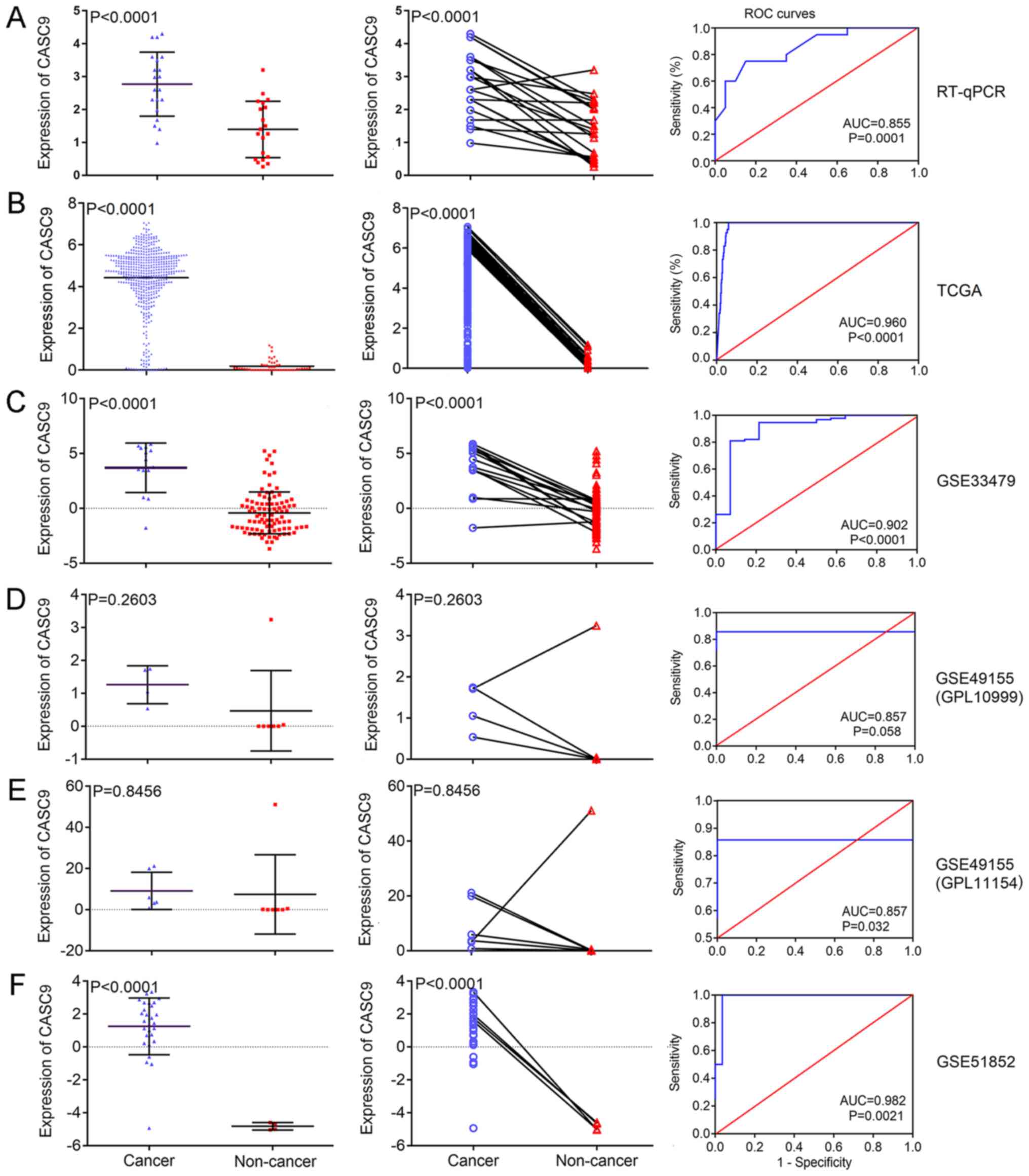 | Figure 2CASC9 expression and its diagnostic
ability in LUSC for all the investigated datasets. The scatter
plots (left panels) and wiring diagrams (middle panels) present the
differential expression levels of CASC9 in LUSC samples and
non-cancer samples for (A) RT-qPCR, (B) TCGA and GEO microarray (C)
GSE33479, (D) GSE49155 (GPL10999), (E) GSE49155 (GPL11154) and (F)
GSE51852 data. The ROC curves (right panels) demonstrate the
capacity of CASC9 to identify LUSC from normal samples. CASC9,
cancer susceptibility candidate 9; LUSC, lung squamous cell
carcinoma; RT-qPCR, reverse transcription-quantitative polymerase
chain reaction; TCGA, The Cancer Genome Atlas; GEO, Gene Expression
Omnibus; ROC, receiver operating characteristic; AUC, area under
the curve. |
Evaluation of the clinicopathological
influence of CASC9 in LUSC from TCGA data
Compared with adjacent normal lung tissue, CASC9 was
significantly upregulated in LUSC tissue (relative expression,
0.190±0.282 vs. 4.420±1.528; P<0.001; Fig. 2B). Apart from the differential
expression between LUSC and normal tissue, overexpression of CASC9
in LUSC was observed to be significantly associated with several
clinicopathological parameters of LUSC, including sex (P=0.029),
clinical stage (P<0.001), distant metastasis (P=0.004) and lymph
node metastasis (P=0.039) (Table
I). Male patients and patients with advanced clinical stage
(III-IV), distant metastasis and lymph node metastasis exhibited
notably higher expression of CASC9. No significant associations
were observed between the expression levels of CASC9 and other
clinicopathological variables. The ROC curves in Fig. 2 suggest that the measurement of
CASC9 performed well in discriminating LUSC from normal tissue
(AUC=0.960; P<0.001). There was no significant difference
between the survival rates of patients divided into high- and
low-expression groups according to the average CASC9 expression
value (P=0.189; Fig. S1) and the
multivariate Cox regression analysis revealed that age (HR, 0.480;
95% CI, 0.269-0.855; P=0.013) and tumor location (HR, 1.850; 95%
CI, 1.080-3.169; P=0.025) were independent prognostic factors for
LUSC (Table II).
 | Table IClinicopathological variables and
CASC9 expression in lung squamous cell carcinoma data from The
Cancer Genome Atlas. |
Table I
Clinicopathological variables and
CASC9 expression in lung squamous cell carcinoma data from The
Cancer Genome Atlas.
| Clinicopathological
feature | n | CASC9 expression
| P-value |
|---|
| Mean ± SD | z-score |
|---|
| Tissue type | | | 10.921 | <0.001 |
| Cancer | 501 | 4.420±1.528 | | |
| Normal | 49 | 0.190±0.282 | | |
| Sex | | | −2.181 | 0.029 |
| Male | 370 | 4.503±1.472 | | |
| Female | 131 | 4.187±1.627 | | |
| Age, yearsa | | | −0.187 | 0.852 |
| >60 | 405 | 4.072±1.864 | | |
| <60 | 87 | 4.045±1.923 | | |
| Tumor location | | | −0.929 | 0.353 |
| Central | 145 | 4.179±1.845 | | |
| Peripheral | 94 | 3.911±1.964 | | |
| Stage | | | −4.040 | <0.001 |
| I-II | 412 | 3.987±1.885 | | |
| III-IV | 85 | 4.795±1.396 | | |
| Distant
metastasis | | | −2.843 | 0.004 |
| M0-MX | 489 | 4.043±1.889 | | |
| M1-M1b | 7 | 5.691±0.770 | | |
| Lymph node
metastasis | | | −2.064 | 0.039 |
| N0-NX | 325 | 4.328±1.546 | | |
| N1-N3 | 176 | 4.592±1.457 | | |
| Primary tumor
stage | | | −1.077 | 0.281 |
| T1-T2 | 407 | 4.397±1.521 | | |
| T3-T4 | 94 | 4.524±1.517 | | |
| Anatomical
classification | | | 3.253 | 0.661 |
| Bronchial | 10 | 4.387±1.586 | | |
| L-lower | 77 | 4.356±1.772 | | |
| L-upper | 134 | 4.292±1.628 | | |
| R-lower | 107 | 4.118±1.834 | | |
| R-middle | 16 | 3.636±2.277 | | |
| R-upper | 139 | 3.834±2.048 | | |
| Table IIMultivariate Cox regression analysis
of clinical parameters and CASC9 expression in TCGA cohort of
patients with lung squamous cell carcinoma. |
Table II
Multivariate Cox regression analysis
of clinical parameters and CASC9 expression in TCGA cohort of
patients with lung squamous cell carcinoma.
| Variablea | HR (95% CI) | P-value |
|---|
| CASC9 expression
(high vs. low) | 1.308
(0.790-2.168) | 0.297 |
| Sex | 1.177
(0.681-2.034) | 0.560 |
| Age, years | 0.480
(0.269-0.855) | 0.013 |
| Tumor location | 1.850
(1.080-3.169) | 0.025 |
| Tumor stage | 1.886
(0.991-3.590) | 0.053 |
| Distant
metastasis | 2.382
(0.321-17.689) | 0.396 |
| Lymph node
metastasis | 0.842
(0.492-1.441) | 0.529 |
| Primary tumor
stage | 1.582
(0.843-2.971) | 0.153 |
| Anatomical
subdivision | 1.024
(0.874-1.201) | 0.768 |
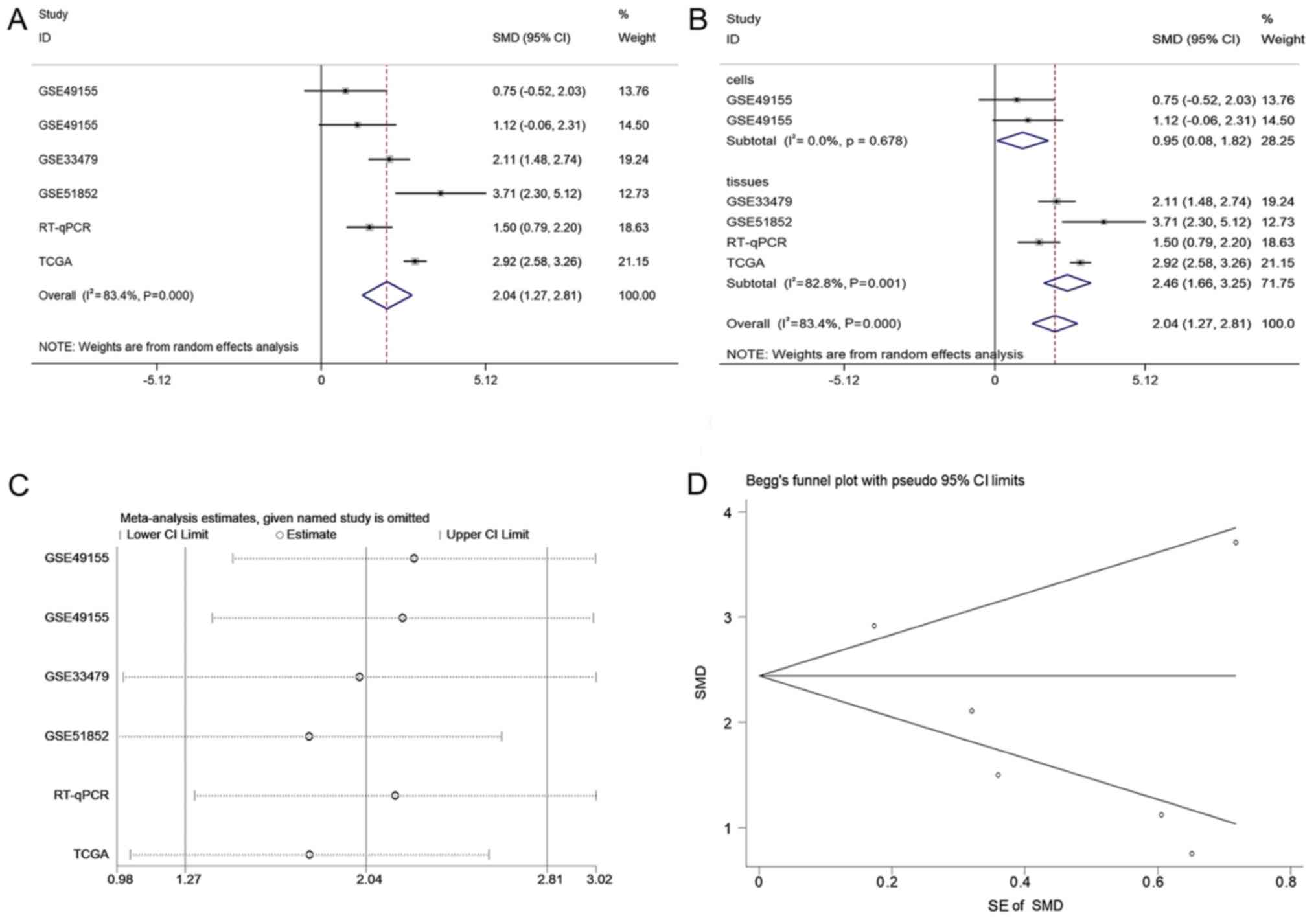
Integrated meta-analysis for the
expression difference of CASC9 between LUSC and non-cancer
tissue
A total of 2,921 and 1,860 studies were identified
in the initial searches in GEO and ArrayExpress, respectively. Of
these, 85 GEO microarray chips and 9 ArrayExpress chips were
included following the scanning of titles and abstracts. Finally, 3
GSE datasets, GSE49155, GSE33479 and GSE51852, were included
(29,30). The distribution of CASC9 expression
and its ability to discriminate between LUSC and non-cancer tissue
in each GSE dataset are displayed in Fig. 2. A flowchart of the selection
process for appropriate microarray chips is illustrated in Fig. 3. No study from the literature
survey was included in the final selection. The in-house RT-qPCR,
TCGA and the included GEO microarray data were merged into a large
pool containing 574 LUSC cases and 182 non-cancer cases for the
integrated meta-analysis. Information on the three cohorts is
listed in Table III. As
demonstrated in Fig. 4A, the SMD
generated from all the cohorts verified the overexpression of CASC9
in LUSC tissues, albeit with significant heterogeneity (SMD, 2.04;
95% CI, 1.27-2.81; I2=83.4%; P<0.001). A subgroup
analysis based on sample type and a sensitivity analysis failed to
trace the source of the heterogeneity (Fig. 4B and C). No heterogeneity was
observed in two studies on patient LUSC cells (I2=0.0%;
P=0.678), in contrast with the large heterogeneity from the tissue
data (I2=82.8%; P=0.001). The reason for this is that
the two studies with different platforms in the cells subgroup were
from the same GSE dataset (GSE49155). Begg's funnel plot indicated
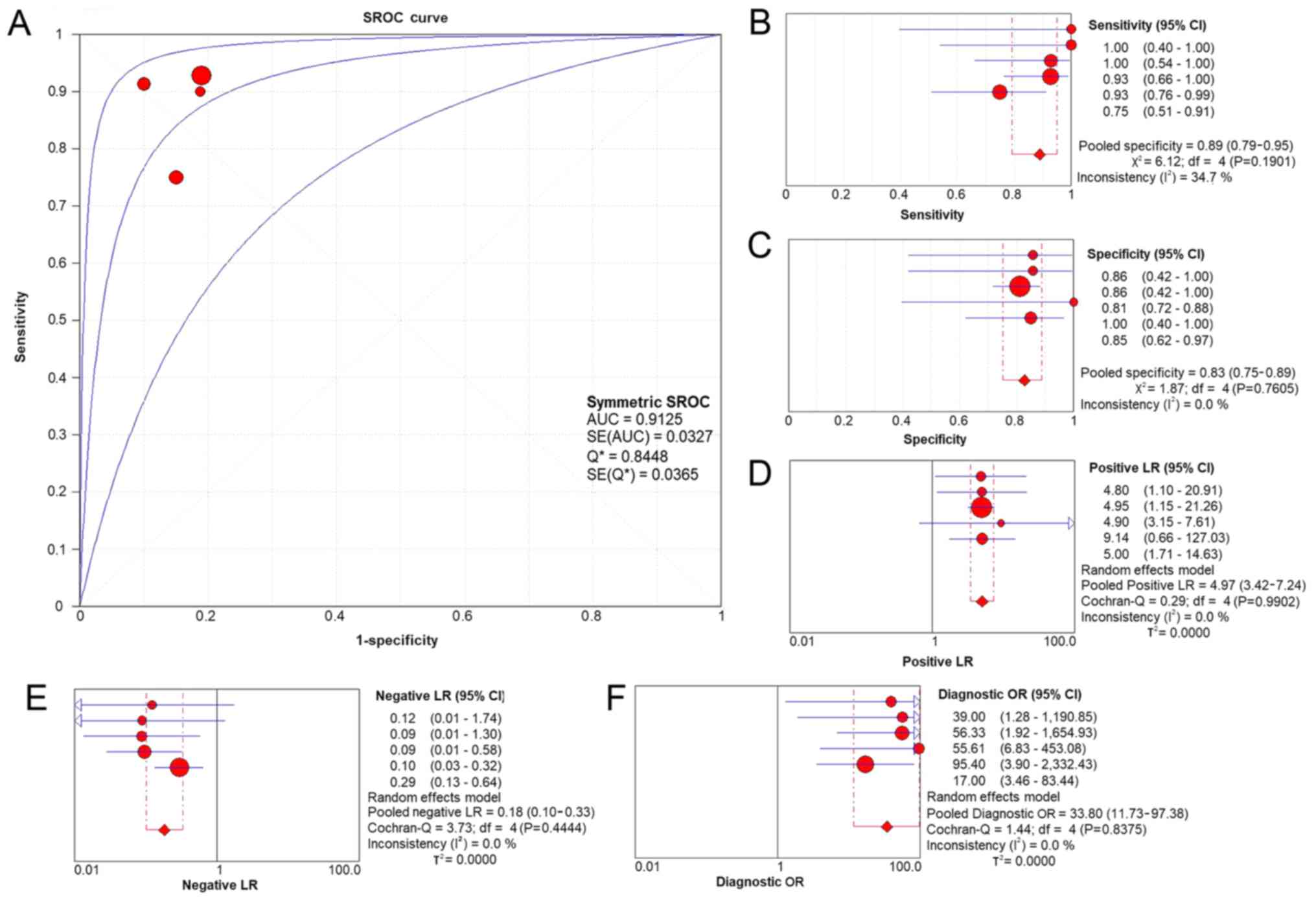
no publication bias (P=0.348). According to the evaluation with the
SROC curves in Fig. 5, a high AUC
(0.9125) revealed the marked ability of CASC9 in distinguishing
LUSC from non-cancer tissues. The aggregated sensitivity,
specificity, positive likelihood ratio, negative likelihood ratio
and diagnostic odds ratio were 0.89, 0.83, 4.97, 0.18 and 33.80,
respectively (Fig. 5B-F).
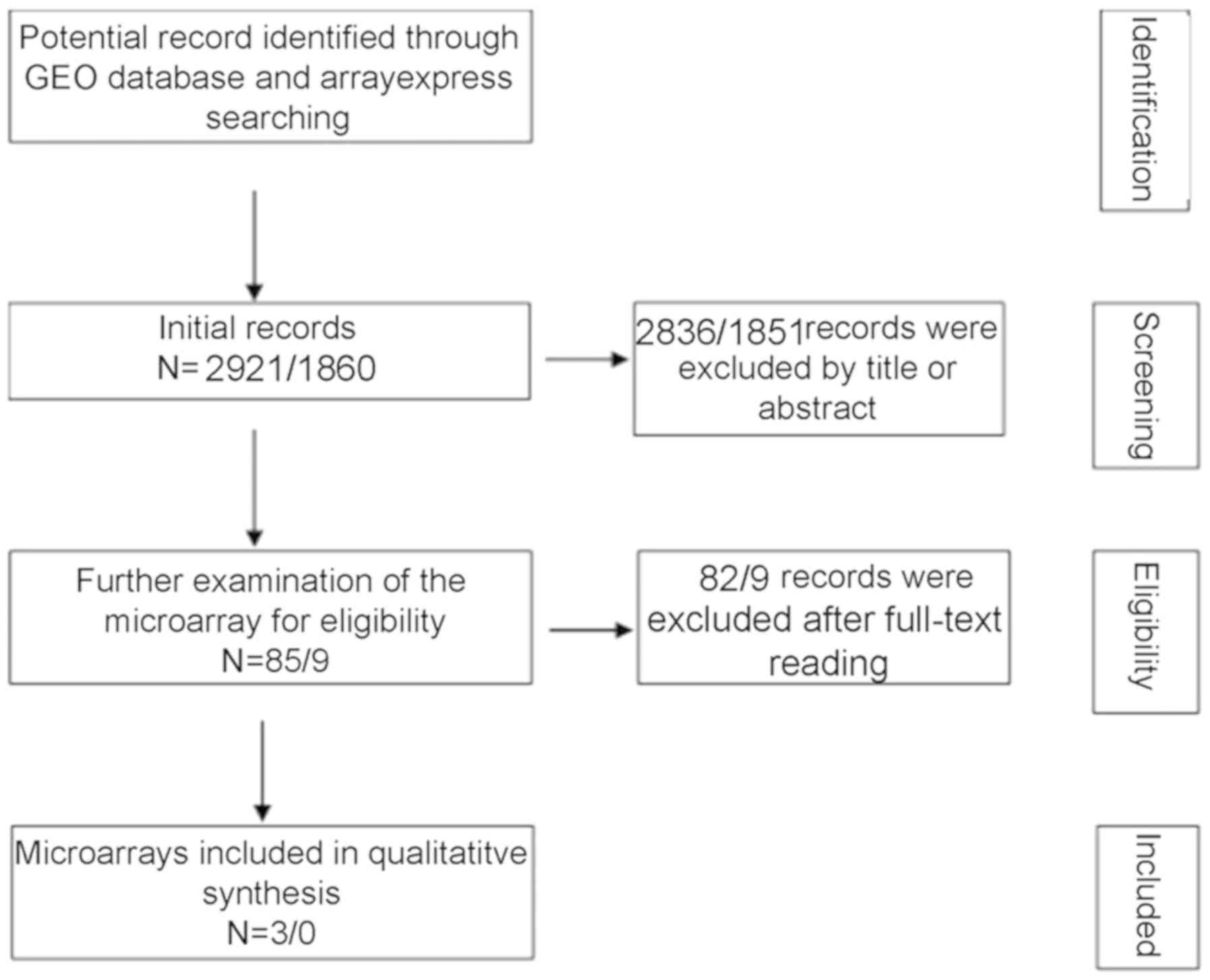 | Figure 3Flowchart of the integrated
meta-analysis data selection. A total of 2,921 and 1,860 studies
appeared as the initial records from the GEO and ArrayExpress,
respectively. Following the screening of titles and abstracts,
2,836 and 1,851 studies from GEO and ArrayExpress, respectively,
were excluded. Finally, 3 GSE datasets were enrolled for the
meta-analysis following the process of full-text reviewing. GEO,
Gene Expression Omnibus. |
| Table IIIBasic, expression and diagnostic
information of all datasets. |
Table III
Basic, expression and diagnostic
information of all datasets.
| | | | Cancer
| Normal
| | | | | |
|---|
| Dataset | Sample type | Study | Sequencing or
microarray platform | n | Mean ± SD CASC9
expression | n | Mean ± SD CASC9
expression | TP | FP | FN | TN | Refs. |
|---|
| GSE49155 | Cells | Ooi et al,
2014 | GPL10999 | 4 | 1.263±0.578 | 7 | 0.470±1.223 | 4 | 1 | 0 | 6 | (31) |
| GSE49155 | Cells | Ooi et al,
2014 | GPL11154 | 6 | 5.976±7.863 | 7 | 0.028±0.070 | 6 | 1 | 0 | 6 | (31) |
| GSE33479 | Tissues | N/A | GPL6480 | 14 | 3.706±2.256 | 95 | −0.415±1.908 | 13 | 18 | 1 | 77 | |
| GSE51852 | Tissues | Arima et al,
2014 | GPL6480 | 28 | 1.255±1.725 | 4 | −4.824±0.231 | 26 | 0 | 2 | 4 | (32) |
| RT-qPCR | Tissues | Present study | N/A | 20 | 2.771±0.974 | 20 | 1.397±0.857 | 15 | 3 | 5 | 17 | - |
| TCGA | Tissues | N/A | Seq-TPM | 502 | 4.422±1.518 | 49 | 0.185±0.282 | 465 | 0 | 37 | 49 | - |
Cell line data of CASC9 expression from
the CCLE
The expression of CASC9 in each of the 191 LC cell
lines was obtained and plotted in a heat map. The spectrum of
colors ranged from red to green, reflecting a wide range of CASC9
expression from high to low (Fig.
6A).
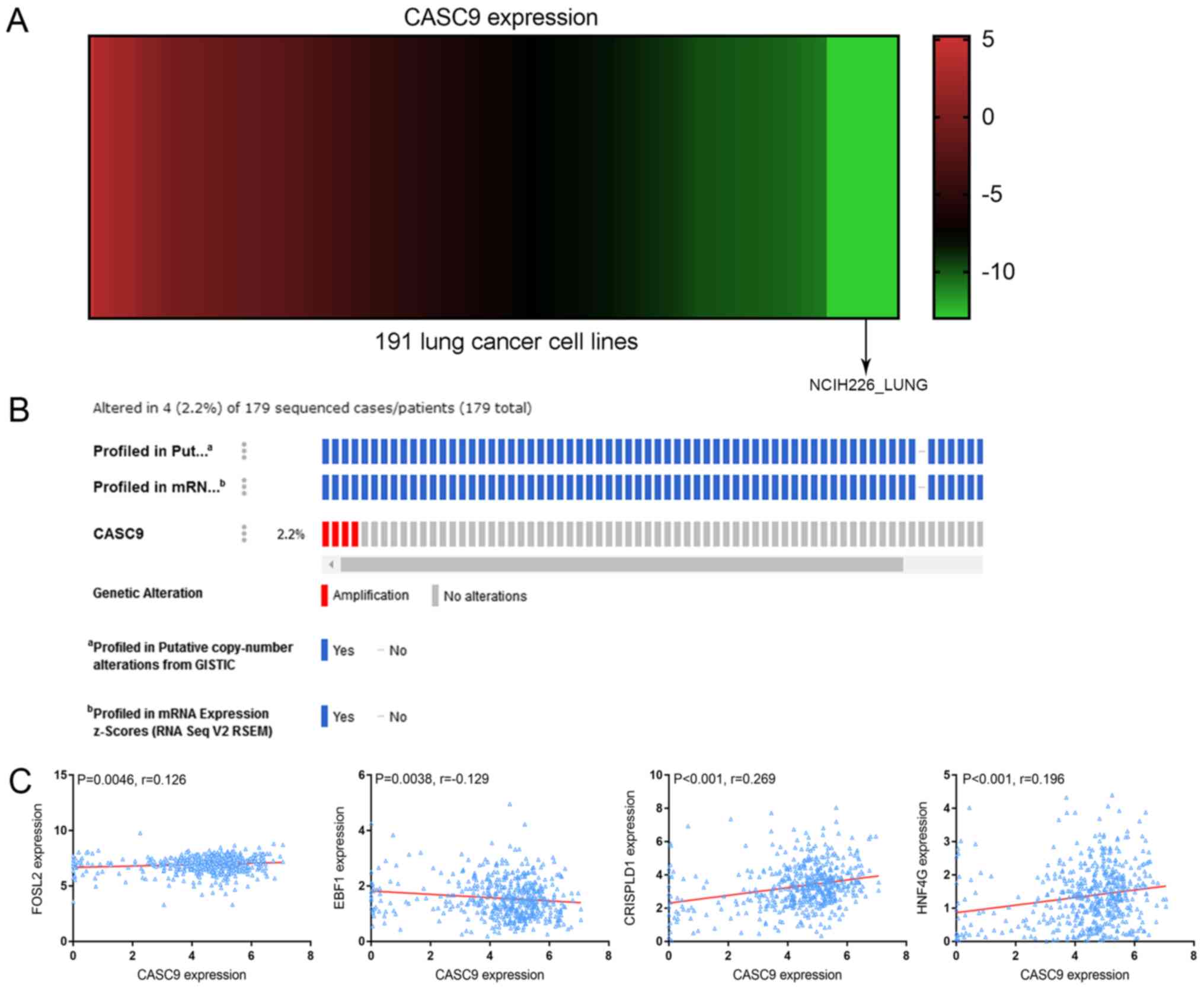 | Figure 6CASC9 expression heat-map, gene
alteration prediction and correlation analysis. (A) Heat-map of
CASC9 expression in 191 lung cancer cell lines arranged from high
to low, corresponding to the spectrum of colors from red to green.
(B) A total of 4 (2.2%) incidences of CASC9 amplification were
revealed in 179 sequenced cases. (C) Correlation between the
expression of CASC9 and predicted transcription factors or adjacent
protein-coding genes. CASC9 was positively correlated with FOSL2
(r=0.126; P=0.0046), CRISPLD1 (r=0.269; P<0.001) and HNF4G
(r=0.196; P<0.001), and negatively correlated with EBF1
(r=−0.129; P=0.0038). CASC9, cancer susceptibility candidate 9;
GISTIC, genomic identification of significant targets in cancer;
FOSL2, Fos-related antigen 2; EBF1, transcription factor COE1;
CRISPLD1, cysteine-rich secretory protein LCCL domain-containing 1;
HNF4G, hepatocyte nuclear factor 4-γ. |
Gene alteration of CASC9 in LUSC tissue
from cBioPortal
The alteration profiles of CASC9 from OncoPrint
revealed that the CASC9 sequence was altered in 4 (2.2%) out of the
179 sequenced cases. The 4 cases of alteration all belonged to the
category of amplification, i.e. multiple copies of the complete
CASC9 gene were naturally occurring in 4 of the 179 sequenced cases
(Fig. 6B).
Prediction of potential transcription
factors for CASC9
Based on the prediction results from LncMAP, 3
transcription factors, including Fos-related antigen 2 (FOSL2),
SWI/SNF complex subunit SMARCC2, and transcription factor COE1
(EBF1), are likely to be involved in the regulatory effect of CASC9
on downstream molecules, including diacylglycerol kinase a,
transient receptor potential cation channel subfamily V member 4,
ankyrin repeat domain-containing protein SOWAHC, semaphorin-3G and
DNA-binding protein Ikaros (Table
IV). No prediction results of transcription factors associated
with CASC9 were produced by other online programs. The correlation
analyses demonstrated a positive correlation between FOSL2 and
CASC9 expression (r=0.126; P=0.0046) and a negative correlation
with EBF1 (r=−0.129; P=0.0038) (Fig.
6C). No significant correlations between SMARCC2 and CASC9 were
observed (data not shown).
| Table IVPotential transcription factors and
genes associated with CASC9 (lncRNA ID, ENSG00000249395) from
LncMAP. |
Table IV
Potential transcription factors and
genes associated with CASC9 (lncRNA ID, ENSG00000249395) from
LncMAP.
| TF ID | TF name | Gene ID | Gene name | Correlation
coefficient (r) according to CASC9 expression
| Score | P-value | FDR |
|---|
| Low expression | High
expression |
|---|
| EN S
G00000075426 | FOSL2 |
ENSG00000065357 | DGKA | 0.513 | −0.002 | 0.995 | <0.001 | <0.001 |
|
ENSG00000075426 | FOSL2 |
ENSG00000111199 | TRPV4 | 0.600 | −0.098 | 1.000 | <0.001 | <0.001 |
|
ENSG00000075426 | FOSL2 |
ENSG00000198142 | SOWAHC | 0.607 | −0.082 | 1.000 | <0.001 | <0.001 |
|
ENSG00000139613 | SMARCC2 |
ENSG00000010319 | SEMA3G | 0.079 | 0.588 | 0.997 | <0.001 | <0.001 |
|
ENSG00000164330 | EBF1 |
ENSG00000185811 | IKZF1 | 0.608 | 0.135 | 0.995 | <0.001 | <0.001 |
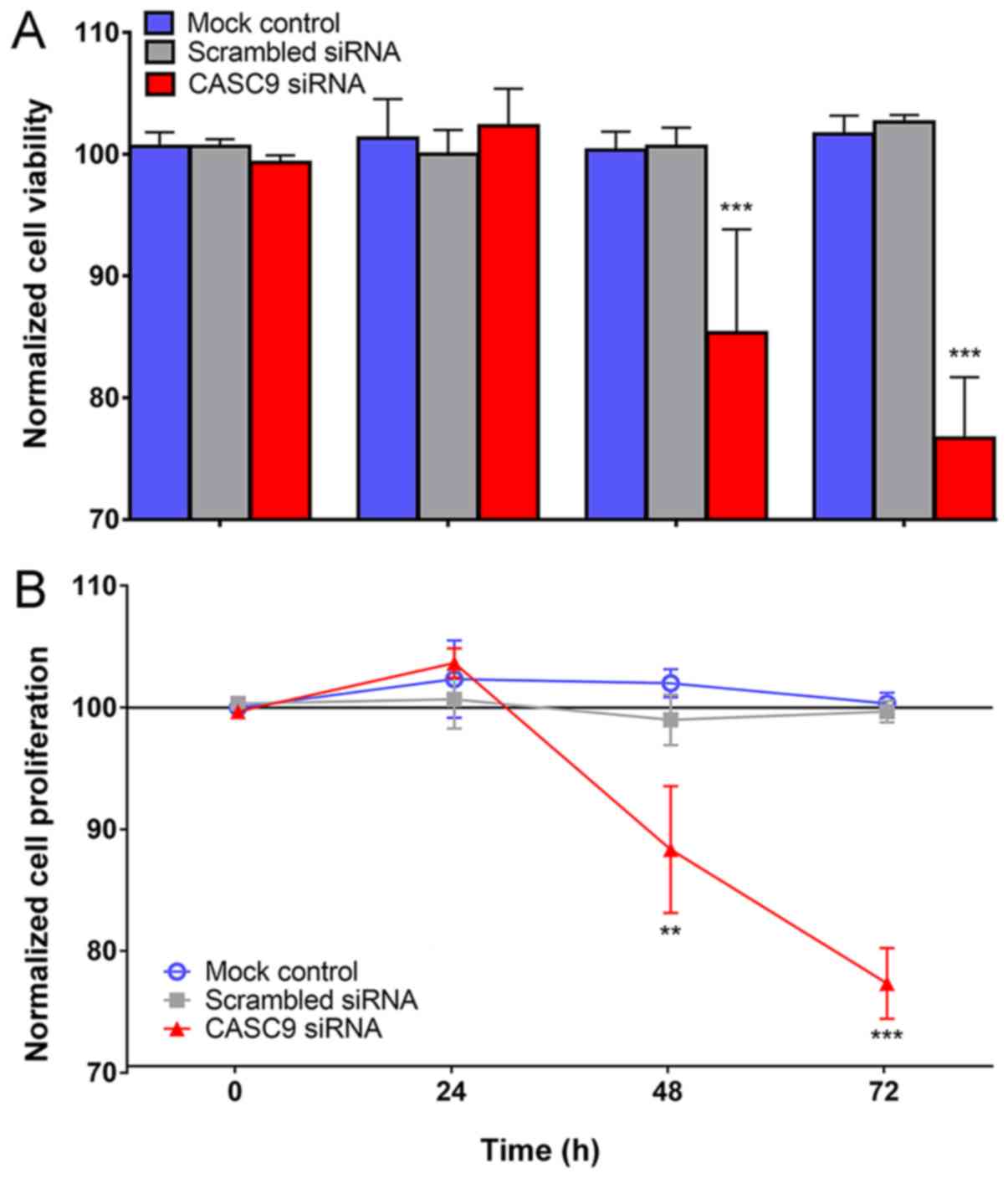
In vitro experiments
The transfection efficiency of the 4 selected CASC9
siRNAs was >80% at 96 h, as determined by RT-qPCR. It can be
observed that CASC9 mRNA expression was obviously lower in CASC9
siRNA group than in scrambled siRNA group and blank control at 48,
72 and 96 h (Fig. S2). Fig. 7 illustrates the behavioral changes
in LUSC cells caused by the siRNA knockdown of CASC9 in terms of
cell viability and proliferation. The viability of the H226 cells
in the CASC9 siRNA group decreased significantly after 48 and 72 h
(both P<0.001), while no significant change of cell viability
occurred in H226 cells among the mock control (P=0.9821) and
scrambled siRNA group (P=0.8790) at 72 h (Fig. 7A). Similarly, compared with the
mock and scrambled siRNA controls, a substantial decrease in the
cell proliferation of H226 cells of CASC9 siRNAs group was recorded
using the MTS tetrazolium assay at 48 (P<0.01) and 72 h
(P<0.001) (Fig. 7B).
Functional enrichment analysis of genes
co-expressed with CASC9
With the aid of GEPIA, 200 genes were identified to
be co-expressed with CASC9. An enrichment analysis of the 200 genes
indicated 16 terms of biological processes and pathways
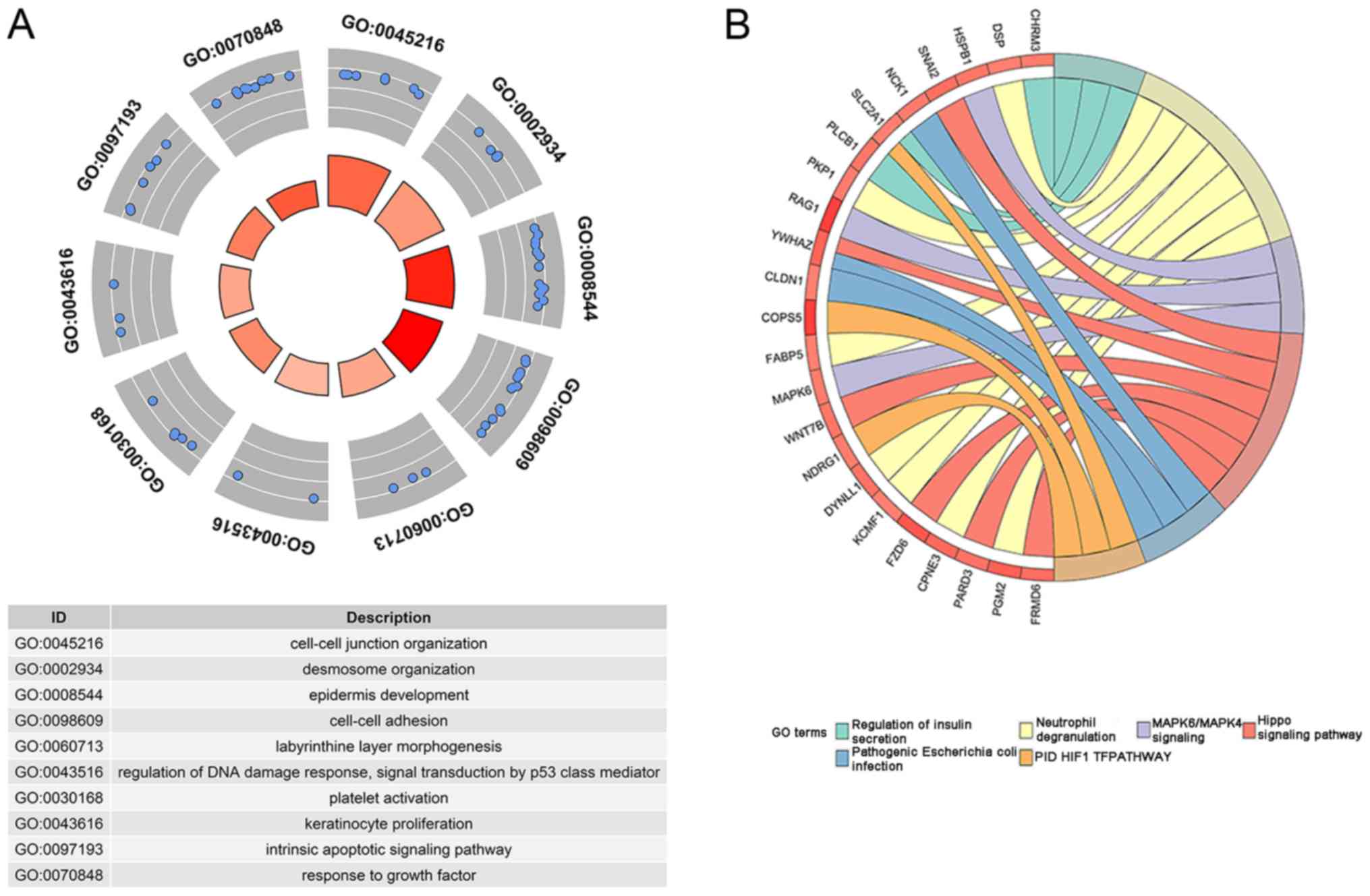
significantly associated with them (Table V; Fig.
8). Among the 16 significant terms, the top 3 biological
processes were 'cell-cell junction organization', 'desmosome
organization' and 'epidermis development', and the top 3 pathways
were 'Hippo signaling pathway', 'pathogenic Escherichia coli
infection', and 'PID HIF1 TF pathway' (all P<0.01).
| Table VMetascape enrichment analysis for the
co-expressed genes of cancer susceptibility candidate 9. |
Table V
Metascape enrichment analysis for the
co-expressed genes of cancer susceptibility candidate 9.
| GO ID | Category | Term | Count of enriched
genes |
Log10(P) |
|---|
| GO:0045216 | GO Biological
Processes | 'Cell-cell junction
organization' | 9 | −7.340 |
| GO:0002934 | GO Biological
Processes | 'Desmosome
organization' | 4 | −7.010 |
| GO:0008544 | GO Biological
Processes | 'Epidermis
development' | 13 | −6.630 |
| GO:0098609 | GO Biological
Processes | 'Cell-cell
adhesion' | 15 | −5.420 |
| hsa04390 | KEGG Pathway | 'Hippo signaling
pathway' | 6 | −4.030 |
| GO:0060713 | GO Biological
Processes | 'Labyrinthine layer
morphogenesis' | 3 | −3.960 |
| GO:0043516 | GO Biological
Processes | 'Regulation of DNA
damage response, signal transduction by p53 class mediator' | 3 | −3.420 |
| GO:0030168 | GO Biological
Processes | 'Platelet
activation' | 5 | −3.100 |
| GO:0043616 | GO Biological
Processes | 'Keratinocyte
proliferation' | 3 | −3.020 |
| hsa05130 | KEGG Pathway | 'Pathogenic
Escherichia coli infection' | 3 | −2.650 |
| GO:0097193 | GO Biological
Processes | 'Intrinsic
apoptotic signaling pathway' | 6 | −2.610 |
| M255 | Canonical
Pathways | 'PID HIF1 TF
pathway' | 3 | −2.420 |
| R-HSA-422356 | Reactome Gene
Sets | Regulation of
insulin secretion | 3 | −2.190 |
| GO:0070848 | GO Biological
Processes | Response to growth
factor | 9 | −2.130 |
| R-HSA-6798695 | Reactome Gene
Sets | Neutrophil
degranulation | 7 | −2.110 |
| R-HSA-5687128 | Reactome Gene
Sets | MAPK6/MAPK4
signaling | 3 | −2.060 |
Associations between CASC9 and
adjacentprotein-coding genes
Location-based displays (chromosome 8,
75,686,308-76,686,308) in the Ensemble Genome Browser unfolded the
positional associations between CASC9 and adjacent protein-coding
genes. Two such genes, cysteine-rich secretory protein LCCL
domain-containing 1 (CRISPLD1; chromosome 8, 75,896,750-75,946,793)
and hepatocyte nuclear factor 4-γ (HNF4G; chromosome 8,
76,320,271-76,476,562), were located 188,846 bp downstream and
129,575 bp upstream of CASC9 (chromosome 8, 76,135,639-76,190,696),
respectively (Fig. 9). Correlation
analyses indicated positive relationships between CASC9 and
CRISPLD1 (r=0.269, P<0.001) or HNF4G expression (r=0.196;
P<0.001) (Fig. 6C).
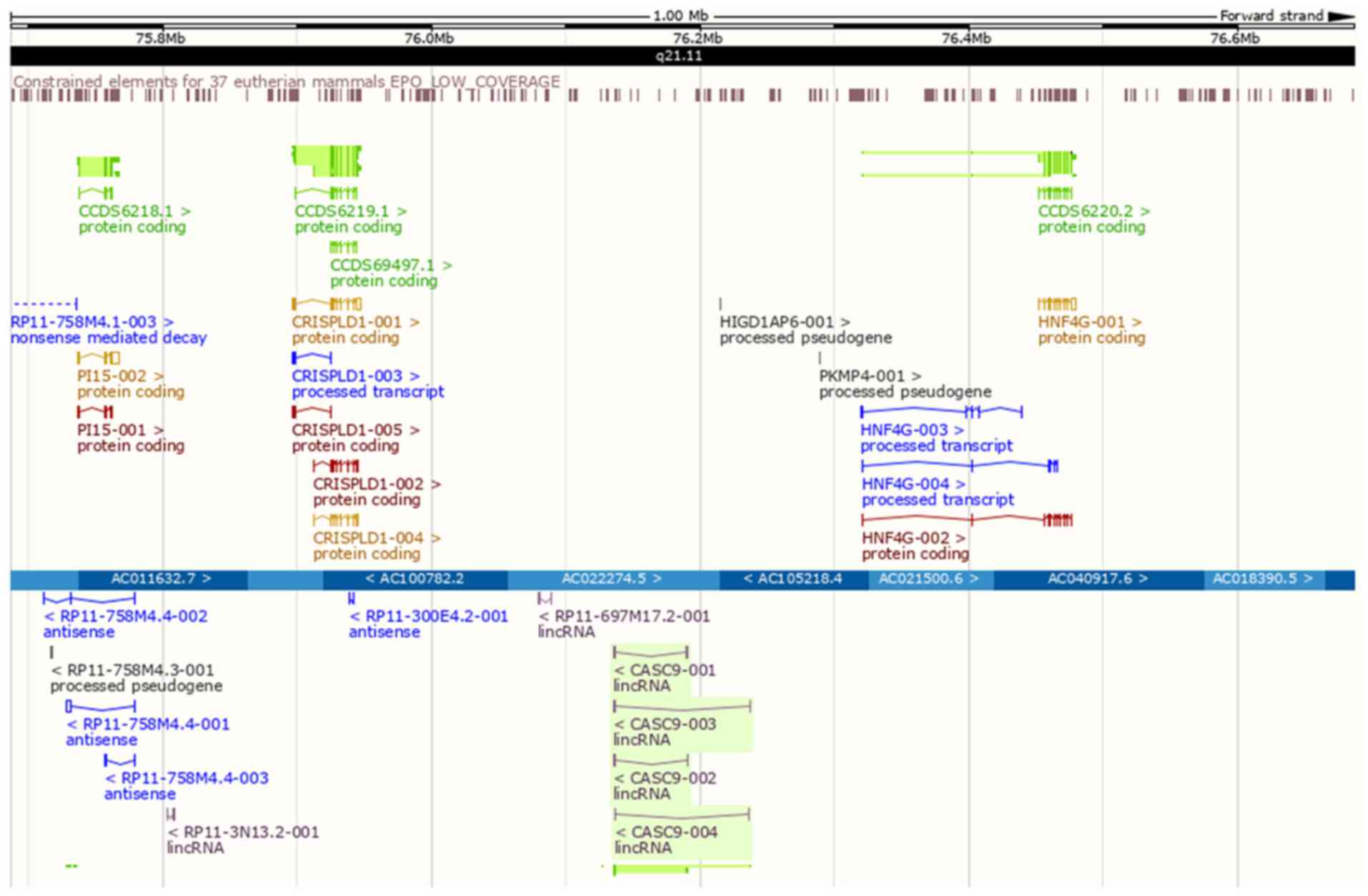 | Figure 9Genomic locations of CASC9 and
adjacent protein-coding genes. CRISPLD1 (chromosome 8,
75,896,750-75,946,793) and HNF4G (chromosome 8,
76,320,271-76,476,562) were 188,846 bp downstream and 129,575 bp
upstream from CASC9 (chromosome 8, 76,135,639-76,190,696),
respectively. CASC9, cancer susceptibility candidate 9; CRISPLD1,
cysteine-rich secretory protein LCCL domain-containing 1; HNF4G,
hepatocyte nuclear factor 4-γ. |
Discussion
There is a large amount of evidence that lncRNAs are
involved in the development of human cancer, with their regulatory
effect on gene expression. Therefore, identifying tumor-associated
lncRNAs and investigating the role of these molecules in the onset
and progression of human cancer may facilitate the discovery of
novel diagnostic and therapeutic biomarkers for LUSC.
The present study concentrated on CASC9, a
cancer-associated lncRNA with carcinogenic function in several
types of human cancer that has not been studied in LUSC. The
expression level of CASC9 between LUSC and non-cancer tissue was
first examined through RT-qPCR. The results demonstrated that CASC9
is overexpressed in LUSC tissue compared with non-cancer tissues.
The statistical analysis of TCGA data and the integrated
meta-analysis also reported significantly elevated CASC9 expression
in LUSC tissues, supporting the RT-qPCR data. One of the highlights
of the present study lies in the integrated meta-analysis
incorporating in-house RT-qPCR, TCGA and GEO data. Evaluating the
expression of CASC9 between LUSC and non-cancer tissues with the
combined methods of data excavation from various public databases
balanced the limited sample size of the clinical specimens obtained
for the present study to a certain extent. The results of the
analysis of the association between CASC9 expression and the
clinicopathological parameters of LUSC imply that upregulated CASC9
may promote the malignant development of LUSC. The ROC and SROC
curves for TCGA data reflected the capacity of CASC9 to distinguish
LUSC from non-cancer tissues. These findings suggest that CASC9 may
be applied as a therapeutic target for LUSC patients.
To gain deeper insights into the oncogenic influence
of CASC9 on LUSC, in vitro experiments were performed
investigating its functional role of CASC9 in this disease.
Knockdown of CASC9 significantly diminished the viability and
proliferation of H226 cells. This provided indirect evidence
reinforcing the effect of CASC9 on the viability and growth of LUSC
cells. Previous studies have probed into the influence of CASC9 on
the cell growth in other cancer types, including ESCC and LUAD
(19,31). The study conducted by Wu et
al (31) demonstrated the
negative correlation between CASC9 and programmed cell death
protein 4 (PDCD4) in ESCC. PDCD4 is a tumor suppressor gene that
participates in the regulation of apoptosis, proliferation and the
cell cycle (32-34). Wu et al (31) further proposed that CASC9 may
downregulate PDCD4 expression by recruiting histone-lysine
N-methyltransferase EZH2 to augment the proliferative ability of
ESCC. It was hypothesized that the regulatory association between
CASC9 and PDCD4 is be a possible explanation for the effect of
CASC9 on the viability and proliferation of LUSC cells.
Following the in vitro experiments, the
molecular mechanism of CASC9 in LUSC was investigated. The 2.2%
incidence of naturally occurring gene duplication of CASC9 in LUSC
samples was consistent with the upregulation of this lncRNA,
providing a possible explanation for its overexpression in this
cancer type. Since the functions of lncRNAs are dependent on
binding to proteins (35,36), a functional enrichment analysis of
genes co-expressed with CASC9 is conducive to comprehending the
molecular basis of CASC9-associated carcinogenesis of LUSC. From
the annotation results, the top biological processes significantly
clustered with the genes co-expressed with CASC9 were mainly
associated with the epithelial-mesenchymal transition (EMT). Gao
et al (37) reported that
CASC9 promotes ESCC proliferation and metastasis by modulating the
EMT signaling pathway. The combination of the functional annotation
results from the present study and the literature search suggest
that CASC9 is involved in the dysregulation of the EMT process in
LUSC. Apart from the aforementioned biological processes, the
functional enrichment analysis revealed significantly assembled
pathways, including the Hippo signaling and mitogen-activated
protein kinase (MAPK)6/MAPK4 signaling pathways, which serve roles
in the promotion or suppression of tumors (38,39).
It is speculated that CASC9 contributes to the initiation and
progression of LUSC by participating in these biological processes
and pathways. Other noteworthy aspects of the CASC9-centered
tumorigenesis of LUSC are its upstream mediators and adjacent
protein-coding genes. Potential factors that regulated the
transcriptional activity of CASC9 were searched in a number of
online programs. However, only lncRNAMap provided prediction
results of CASC9-associated downstream transcription factors and
mRNAs. Two of the predicted transcription factors were linked to
NSCLC. The abnormal expression of SMARCC2 was discovered in
squamous NSCLC tissue, and FOSL2 is required for transforming
growth factor p1-induced migration in NSCLC (40,41).
The other transcription factor, EBF1, is a B-lineage
transcriptional regulator involved in B-cell acute lymphoblastic
leukemia (42). Although only EBF1
expression was negatively correlated with CASC9 expression in LUSC,
and none of the predicted transcription factors or mRNAs were
confirmed in the literature search to be targeted by CASC9, these
predicted downstream molecules hold important implications for the
pathogenesis of LUSC. The association between CASC9 and adjacent
protein-coding genes was also analyzed and CASC9 expression was
positively correlated with two such genes, CRISPLD1 and HNF4G.
Notably, HNF4G was reported in a previous study to serve an
oncogenic role in LC by promoting cell proliferation (43). Therefore, it was hypothesized that
CASC9 modulates the expression of HNF4G or interacts with it to
affect the occurrence and progression of LUSC. Additionally, CASC9
has been reported to enhance the malignant potential of human
cancer types, including breast cancer, hepatocellular carcinoma and
ESCC, by interacting with numerous target genes, such as EZH2,
heterogeneous nuclear ribonucleoprotein L and laminin subunit γ-2
(44-46). Although these target genes were not
revealed in the bioinformatics analysis of the present study, the
association between them and CASC9 in LUSC is also worth exploring
in future studies.
Although certain notable findings have been
revealed, the limitations of the present study lie in the following
aspects: Only the discernment capacity of CASC9 to distinguish LUSC
from non-cancer tissues was assessed, and there was not sufficient
evidence to support its diagnostic value in this disease. The serum
CASC9 levels in patients should be investigated in future
experiments for the evaluation of the diagnostic significance.
Additionally, in the in vitro experiments, only CASC9
knockout was conducted. To comprehensively investigate the
influence of CASC9 on the biological function of LUSC cells,
overexpression experiment should also be performed. Furthermore,
using >3 types of cells lines in the in vitro experiment
would be more conclusive than a single cell line. Further in
vitro and in vivo experiments are required to verify the
influence of CASC9 on the aforementioned biological processes and
pathways, as well as the targeting regulatory association between
CASC9 and the predicted transcription factors or adjacent
protein-coding genes.
In summary, the present study revealed the
overexpression and clinicopathological significance of CASC9 in
LUSC for the first time. The role of CASC9 as a cancer-promoting
factor in LUSC may be accomplished by strengthening the viability
and proliferation capacity of the tumor cells. A functional
enrichment analysis of co-expressed genes and the correlation
between the expression of CASC9 and predicted transcription factors
or adjacent protein-coding genes inferred a potential molecular
mechanism of CASC9-associated LUSC tumorigenesis.
Supplementary Materials
Funding
The present study was supported by the Natural
Science Foundation of Guangxi, China (grant nos. 2017GXNSFAA
198016, 2015GXNSFCA139009 and 2017GXNSFAA198067), the Fund of
National Natural Science Foundation of China (grant no. NSFC
81560469), the Guangxi Zhuang Autonomous Region Health and Family
Planning Commission Self-financed Scientific Research Project
(grant no. Z20180979), a Guangxi Medical University Training
Program for Distinguished Young Scholars, a Medical Excellence
Award Funded by the Creative Research Development Grant from the
First Affiliated Hospital of Guangxi Medical University, and a
Future Academic Stars Project from Guangxi Medical University
(grant no. WLXSZX18068).
Availability of data and materials
The datasets generated and/or analyzed during the
current study were TCGA-LUSC (https://portal.gdc.cancer.gov/) GSE49155, GSE33479 and
GSE51852 (https://www.ncbi.nlm.nih.gov/gds/). The datasets used
and analyzed during the current study are available from the
corresponding author on reasonable request.
Authors' contributions
GC, KLW and LG designed the study. LG, JHZ, FCM, JL,
HWZ and SX performed the analyses and calculations. LG and YNG
reviewed the microarray and RNA-sequencing sources in the
meta-analysis, and contributed to the writing of the manuscript.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The patients involved in the present study provided
signed informed consent and approval was granted by the Ethics
Committee of the First Affiliated Hospital of Guangxi Medical
University, Nanning, China (approval no. 2015-KY-NSFC-059).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Perez-Moreno P, Brambilla E, Thomas R and
Soria JC: Squamous cell carcinoma of the lung: molecular subtypes
and therapeutic opportunities. Clin Cancer Res. 18:2443–2451. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Guo NL, Dowlati A, Raese RA, Dong C, Chen
G, Beer DG, Shaffer J, Singh S, Bokhary U, Liu L, et al: A
predictive 7-gene assay and prognostic protein biomarkers for
non-small cell lung cancer. EBioMedicine. 32:102–110. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2018. CA Cancer J Clin. 68:7–30. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yan X, Shen H, Jiang H, Hu D, Wang J and
Wu X: YXQ-EQ induces apoptosis and inhibits signaling pathways
important for metastasis in non-small cell lung carcinoma cells.
Cell Physiol Biochem. 49:911–919. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wu C, Li X, Zhang D, Xu B, Hu W, Zheng X,
Zhu D, Zhou Q, Jiang J and Wu C: IL-1beta-mediated up-regulation of
WT1D via miR-144-3p and their synergistic effect with
NF-kappaB/COX-2/HIF-1alpha pathway on cell proliferation in LUAD.
Cell Physiol Biochem. 48:2493–2502. 2018. View Article : Google Scholar
|
|
6
|
Lee B, Lee T, Lee SH, Choi YL and Han J:
Clinicopathologic characteristics of EGFR, KRAS, and ALK
alterations in 6,595 lung cancers. Oncotarget. 7:23874–23884.
2016.PubMed/NCBI
|
|
7
|
Hou Z, Xu C, Xie H, Xu H, Zhan P, Yu L and
Fang X: Long noncoding RNAs expression patterns associated with
chemo response to cisplatin based chemotherapy in lung squamous
cell carcinoma patients. PLoS One. 9:e1081332014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhao J, Liu Y, Zhang W, Zhou Z, Wu J, Cui
P, Zhang Y and Huang G: Long non-coding RNA Linc00152 is involved
in cell cycle arrest, apoptosis, epithelial to mesenchymal
transition, cell migration and invasion in gastric cancer. Cell
Cycle. 14:3112–3123. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fan CN, Ma L and Liu N: Systematic
analysis of lncRNA-miRNA-mRNA competing endogenous RNA network
identifies four-lncRNA signature as a prognostic biomarker for
breast cancer. J Transl Med. 16:2642018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Huang YK and Yu JC: Circulating microRNAs
and long non-coding RNAs in gastric cancer diagnosis: An update and
review. World J Gastroenterol. 21:9863–9886. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang X, Sehgal L, Jain N, Khashab T,
Mathur R and Samaniego F: LncRNA MALAT1 promotes development of
mantle cell lymphoma by associating with EZH2. J Transl Med.
14:3462016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dong L, Ding H, Li Y, Xue D and Liu Y:
LncRNA TINCR is associated with clinical progression and serves as
tumor suppressive role in prostate cancer. Cancer Manag Res.
10:2799–2807. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liang X, Qi M, Wu R, Liu A, Chen D, Tang
L, Chen J, Hu X, Li W, Zhan L, et al: Long non-coding RNA CUDR
promotes malignant phenotypes in pancreatic ductal adenocarcinoma
via activating AKT and ERK signaling pathways. Int J Oncol.
53:2671–2682. 2018.PubMed/NCBI
|
|
14
|
Cao W, Wu W, Shi F, Chen X, Wu L, Yang K,
Tian F, Zhu M, Chen G, Wang W, et al: Integrated analysis of long
noncoding RNA and coding RNA expression in esophageal squamous cell
carcinoma. Int J Genomics. 2013:4805342013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Su X, Li G and Liu W: The long noncoding
RNA cancer susceptibility candidate 9 promotes nasopharyngeal
carcinogenesis via stabilizing HIF1a. DNA Cell Biol. 36:394–400.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shang C, Sun L, Zhang J, Zhao B, Chen X,
Xu H and Huang B: Silence of cancer susceptibility candidate 9
inhibits gastric cancer and reverses chemoresistance. Oncotarget.
8:15393–15398. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ma P, Zhang M, Nie F, Huang Z, He J, Li W
and Han L: Transcriptome analysis of EGFR tyrosine kinase
inhibitors resistance associated long noncoding RNA in non-small
cell lung cancer. Biomed Pharmacother. 87:20–26. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhou J, Xiao H, Yang X, Tian H, Xu Z,
Zhong Y, Ma L, Zhang W, Qiao G and Liang J: Long noncoding RNA
CASC9.5 promotes the proliferation and metastasis of lung
adenocarcinoma. Sci Rep. 8:372018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xiong DD, Li ZY, Liang L, He RQ, Ma FC,
Luo DZ, Hu XH and Chen G: The lncRNA NEAT1 accelerates lung
adenocarcinoma deterioration and binds to mir-193a-3pas a
competitive endogenous RNA. Cell Physiol Biochem. 48:905–918. 2018.
View Article : Google Scholar
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Δ Δ C(T)) Method. Methods. 25:402–408. 2001. View Article : Google Scholar
|
|
21
|
Tomczak K, Czerwinska P and Wiznerowicz M:
The Cancer Genome Atlas (TCGA): An immeasurable source of
knowledge. Contemp Oncol (Pozn). 19A:A68–A77. 2015.
|
|
22
|
Hutter C and Zenklusen JC: The Cancer
Genome Atlas: Creating lasting value beyond its data. Cell.
173:283–285. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
He RQ, Gao L, Ma J, Li ZY, Hu XH and Chen
G: Oncogenic role of miR-183-5p in lung adenocarcinoma: A
comprehensive study of qPCR, in vitro experiments and bioinformatic
analysis. Oncol Rep. 40:83–100. 2018.PubMed/NCBI
|
|
24
|
Dang Y, Luo D, Rong M and Chen G:
Underexpression of miR-34a in hepatocellular carcinoma and its
contribution towards enhancement of proliferating inhibitory
effects of agents targeting c-MET. PLoS One. 8:e610542013.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Rong M, Chen G and Dang Y: Increased
miR-221 expression in hepatocellular carcinoma tissues and its role
in enhancing cell growth and inhibiting apoptosis in vitro. BMC
Cancer. 13:212013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Huang S and He R: Synergistic effect of
MiR-146a mimic and cetuximab on hepatocellular carcinoma cells.
BioMed Research International. 2014:3841212014.PubMed/NCBI
|
|
27
|
Dang YW, Zeng J, He RQ, Rong MH, Luo DZ
and Chen G: Effects of miR-152 on cell growth inhibition, motility
suppression and apoptosis induction in hepatocellular carcinoma
cells. Asian Pac J Cancer Prev. 15:4969–4976. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gao J, Aksoy BA, Dogrusoz U, Dresdner G,
Gross B, Sumer SO, Sun Y, Jacobsen A, Sinha R, Larsson E, et al:
Integrative analysis of complex cancer genomics and clinical
profiles using the cBio- Portal. Sci Signal. 6:pl 12013. View Article : Google Scholar
|
|
29
|
Ooi AT, Gower AC, Zhang KX, Vick JL, Hong
L, Nagao B, Wallace WD, Elashoff DA, Walser TC, Dubinett SM, et al:
Molecular profiling of premalignant lesions in lung squamous cell
carcinomas identifies mechanisms involved in stepwise
carcinogenesis. Cancer Prev Res (Phila). 7:487–495. 2014.
View Article : Google Scholar
|
|
30
|
Arima C, Kajino T, Tamada Y, Imoto S,
Shimada Y, Nakatochi M, Suzuki M, Isomura H, Yatabe Y, Yamaguchi T,
et al: Lung adenocarcinoma subtypes definable by lung
development-related miRNA expression profiles in association with
clinicopathologic features. Carcinogenesis. 35:2224–2231. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wu Y, Hu L, Liang Y, Li J, Wang K, Chen X,
Meng H, Guan X, Yang K and Bai Y: Up-regulation of lncRNA CASC9
promotes esophageal squamous cell carcinoma growth by negatively
regulating PDCD4 expression through EZH2. Mol Cancer. 16:1502017.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lankat-Buttgereit B and Göke R: The tumour
suppressor Pdcd4: Recent advances in the elucidation of function
and regulation. Biol Cell. 101:309–317. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Allgayer H: Pdcd4, a colon cancer
prognostic that is regulated by a microRNA. Crit Rev Oncol Hematol.
73:185–191. 2010. View Article : Google Scholar
|
|
34
|
Hwang SK, Minai-Tehrani A, Lim HT, Shin
JY, An GH, Lee KH, Park KR, Kim YS, Beck GR Jr, Yang HS, et al:
Decreased level of PDCD4 (programmed cell death 4) protein
activated cell proliferation in the lung of A/J mouse. J Aerosol
Med Pulm Drug Deliv. 23:285–293. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kaneko S, Li G, Son J, Xu CF, Margueron R,
Neubert TA and Reinberg D: Phosphorylation of the PRC2 component
Ezh2 is cell cycle-regulated and up-regulates its binding to ncRNA.
Genes Dev. 24:2615–2620. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang KC, Yang YW, Liu B, Sanyal A,
Corces-Zimmerman R, Chen Y, Lajoie BR, Protacio A, Flynn RA, Gupta
RA, et al: A long noncoding RNA maintains active chromatin to
coordinate homeotic gene expression. Nature. 472:120–124. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gao GD, Liu XY, Lin Y, Liu HF and Zhang
GJ: LncRNA CASC9 promotes tumorigenesis by affecting EMT and
predicts poor prognosis in esophageal squamous cell cancer. Eur Rev
Med Pharmacol Sci. 22:422–429. 2018.PubMed/NCBI
|
|
38
|
Kostenko S, Dumitriu G and Moens U: Tumour
promoting and suppressing roles of the atypical MAP kinase
signalling pathway ERK3/4-MK5. J Mol Signal. 7:92012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hu Y, Yang C, Yang S, Cheng F, Rao J and
Wang X: miR-665 promotes hepatocellular carcinoma cell migration,
invasion, and proliferation by decreasing Hippo signaling through
targeting PTPRB. Cell Death Dis. 9:9542018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wang J, Sun D, Wang Y, Ren F, Pang S, Wang
D and Xu S: FOSL2 positively regulates TGF-ß1 signalling in
non-small cell lung cancer. PLoS One. 9:e1121502014. View Article : Google Scholar
|
|
41
|
Remmelink M, Mijatovic T, Gustin A,
Mathieu A, Rombaut K, Kiss R, Salmon I and Decaestecker C:
Identification by means of cDNA microarray analyses of gene
expression modifications in squamous non-small cell lung cancers as
compared to normal bronchial epithelial tissue. Int J Oncol.
26:247–258. 2005.
|
|
42
|
Georgopoulos K: Ebf1 in DNA repair and
leukemogenesis. Blood. 125:3969–3971. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wang J, Zhang J, Xu L, Zheng Y, Ling D and
Yang Z: Expression of HNF4G and its potential functions in lung
cancer. Oncotarget. 9:18018–18028. 2017.
|
|
44
|
Jiang B, Li Y, Qu X, Zhu H, Tan Y, Fan Q,
Jiang Y, Liao M and Wu X: Long noncoding RNA cancer susceptibility
candidate 9 promotes doxorubicin-resistant breast cancer by binding
to enhancer of zeste homolog 2. Int J Mol Med. 42:2801–2810.
2018.PubMed/NCBI
|
|
45
|
Klingenberg M, Groß M, Goyal A,
Polycarpou-Schwarz M, Miersch T, Ernst AS, Leupold J, Patil N,
Warnken U, Allgayer H, et al: The long noncoding RNA cancer
susceptibility 9 and RNA binding protein heterogeneous nuclear
ribonucleoprotein L form a complex and coregulate genes linked to
AKT signaling. Hepatology. 68:1817–1832. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Liang Y, Chen X, Wu Y, Li J, Zhang S, Wang
K, Guan X, Yang K and Bai Y: LncRNA CASC9 promotes esophageal
squamous cell carcinoma metastasis through upregulating LAMC2
expression by interacting with the CREB-binding protein. Cell Death
Differ. 25:1980–1995. 2018. View Article : Google Scholar : PubMed/NCBI
|