Introduction
In the United States, prostate cancer (PCa) is one
of the most common malignant tumors affecting males. Almost 161,360
new cases were diagnosed in 2017, leading to 26,730 deaths within
one year (1). Although early
diagnosis and treatment have reduced the mortality rate of patients
with PCa, it is still the second leading cause of cancer-related
mortality in the United States among males (1). Radiotherapy and radical prostatectomy
are used in organ-confined PCa; however, >40% of patients
develop recurrence or metastasis (2). In fact, the majority of PCa cases are
diagnosed in the advanced stages of the disease. Androgen
deprivation therapy can treat PCa with controlled or improved
conditions in a large number of patients; however, after a median
remission period of 18-36 months, tumors also recur, gaining the
androgen-independent ability to proliferate (3,4). The
occurrence and development of tumors is the result of multi-gene
and multi-factor actions. Thus, the exploration of the molecular
mechanisms and biomarkers of tumorigenesis has attracted much
research attention in oncology.
The Human Genome Project has revealed that there is
a large amount of non-coding RNAs (ncRNAs) in the human genome,
which has been proven by >40-60% of genome transcripts (5). MicroRNAs (miRNAs or miRs) are
endogenous small ncRNA molecules (19-22 nucleotides in length) that
regulate protein coding. They regulate gene expression by binding
to a specific sequence of messenger RNA, thereby inhibiting the
translation or shearing of RNA transcripts (6,7).
Previous bioinformatics analyses have revealed that miRNAs regulate
the expression levels of 60% of all genes (8,9).
Therefore, miRNAs are considered to be a factor widely involved in
the micro-regulation of gene expression and may participate in
almost all biological processes (10,11).
In 2002, it was reported that the expression of miRNAs was
downregulated in the majority of chronic B-lymphoblastic leukemia
tissues, emphasizing for the first time the importance of miRNAs in
human cancer (12). Increasing
evidence has indicated that abnormally expressed miRNAs regulate
tumorigenesis, progression, metastasis and drug resistance by
targeting some tumor-related genes (13,14).
In prostate cells, some miRNAs are differentially expressed, of
which miR-375, miR-200c, miR-143 and miR-145 have been reported to
have a particularly significant effect (15-19).
Metadherin (MTDH) is alternatively known as
astrocyte elevated gene (AEG)-1 (20,21).
A number of studies have found that MTDH is highly expressed in
several tumor cells and is closely related to the proliferation,
apoptosis and migration of tumor cells (22-26).
There is evidence to indicate that several miRNAs are involved in
tumor cell progression by directly targeting MTDH (27-30).
Although, several studies had investigated the
effects of MTDH or other miRNAs targeting MTDH on PCa, an miRNA can
regulate multiple mRNA targets, and similarly, multiple miRNAs can
act on the same mRNA (31,32). Moreover, as a heterogeneous tumor,
on the one hand, PCa exhibits an abnormal miRNA expression; for
instance, miR-183 has been found to be upregulated in PCa, while
miR-miR-145 has been shown to be downregulated in prostate tissues
(17). On the other hand, MTDH may
have different functions in different PCa cell lines. It can be
thus concluded that an individual miRNA may play different roles in
different cell types by targeting different pathways/genes.
Therefore, it is still necessary to study the relations among new
miRNAs, MTDH and PCa, as well as their effects on tumor growth and
metastasis.
Materials and methods
Tissue samples
A total of 46 PCa tissues and adjacent normal
tissues were collected from patients with PCa who underwent
resection surgery and were admitted to the First Affiliated
Hospital and College of Clinical Medicine of Henan University of
Science and Technology (Luoyang, China) from June, 2011 to May,
2012. Tissues were confirmed by pathological analysis. One part of
the paired tissues were used for pathological diagnosis and stored
in 4% formaldehyde solution, while the other part was used in later
experiments and stored in a liquid nitrogen tank. The association
between MTDH expression and the clinicopathological characteristics
of patients with PCa is presented in Table I. The Ethics Committees of the
First Affiliated Hospital and College of Clinical Medicine of Henan
University of Science and Technology approved the study. All
patients signed informed consent forms prior to participation.
 | Table IThe association between MTDH
expression and the clinicopathological characteristics of patients
with prostate cancer. |
Table I
The association between MTDH
expression and the clinicopathological characteristics of patients
with prostate cancer.
|
Characteristics | No. of patients
(n=46) | No. of patients
with high MTDH expression (n=31) | No. of patients
with low MTDH expression (n=15) | P-value |
|---|
| Age (years) | | | |
0.592b |
| <55 | 21 | 15 | 6 | |
| ≥55 | 25 | 16 | 9 | |
| Median
survival | 26.3 | 25.4 | 28.4 | 0.468 |
| Tumor size
(cm) | | | |
0.047a,b |
| <3 | 21 | 11 | 10 | |
| ≥3 | 25 | 20 | 5 | |
| TNM stage | | | |
0.497c |
| I and II | 33 | 21 | 12 | |
| IIIa | 13 | 10 | 3 | |
| Osseous
metastasis | | | |
0.027a,b |
| No | 20 | 10 | 10 | |
| Yes | 26 | 21 | 5 | |
| Lymphatic
metastasis | | | |
0.161c |
| No | 27 | 16 | 11 | |
| Yes | 19 | 15 | 4 | |
Immunohistochemistry (IHC)
IHC was performed to detect the protein expression
levels of MTDH in 2 paired PCa and adjacent tissues by
streptavidin-peroxidase (SP) staining. The fresh PCa and adjacent
tissues were fixed in 4% formaldehyde for >24 h at room
temperature. The samples (5 μm thickness) were
deparaffinized in xylene, dehydrated with gradient ethanol, and 3%
H2O2 was used to block endogenous peroxidase.
The samples were first incubated with MTDH antibody (1:2,000;
ab227981; Abcam, Cambridge, MA, USA) at 4°C overnight and then were
washed by PBS and incubated at room temperature for 30 min with the
secondary antibody, HRP-conjugated goat anti-rabbit IgG (1:1,000;
cat. no. 10285-1-AP; Proteintech Group, Inc., Rosemont, IL, USA).
Diaminobenzidine (DBA) was performed by chromogen and hematoxylin
was used to re-dye the samples. The substitution of PBS for primary
antibody was used as a negative control. Staining patterns were
analyzed by selected representative slices. The immunostains were
observed by 2 independent experienced pathologists. MTDH staining
intensities were categorized as negative, faint yellow, yellow and
brown.
Cells and cell culture
Human prostate epithelial cell lines [PWR-1E
(CRL-11611) and RWPE-1 cells (CRL-11609)] and PCa cell lines [PC-3
(CRL-1435™), DU145 (HTB-81), C4-2 (CRL-3314), 22Rv1 (CRL-2505) and
NCI-H660 cells (CRL-5813)] were obtained from the American Type
Culture Collection (ATCC, Manassas, VA, USA) and LNCaP cells were
purchased from Cell Lines Service (CLS, Eppelheim, Germany). The
cells were cultured at 37°C in 5% CO2 in RPMI-1640
medium (GENOM, Hangzhou, China) with 10% fetal bovine serum (Thermo
Fisher Scientific, Waltham, MA, USA), 100 U/ml penicillin and 100
U/ml streptomycin (Gibco/Thermo Fisher Scientific). The medium was
changed every 8-10 h.
Cell transfection
Cells were seeded in a 6-well plate
(1.0×105) for 24 h prior to transfection. Overexpression
and MTDH siRNA (silencing MTDH, 5′-GGTGAAGATAACTCTACTG-3′),
inhibitors and mimics of miR-145-5p, miR-145-3p, miR-499a-5p and
miR-22-3p, as well as mock and negative control (NC,
5′-GGACGAUGGCUAAUUACAU-3′) plasmids were synthetized by
Invitrogen/Thermo Fisher Scientific. Transient transfection was
performed using Lipofectamine 2000 (Invitrogen/Thermo Fisher
Scientific) according to the manufacturer’s instructions. A total
of (100 nmol/l) overexpression or silencing MTDH/ mimics or
inhibitors of miRNAs, mock or NC and Lipofectamine 2000 were added
to Opti-MEM medium followed by incubation at 25°C for 10 min,
respectively. Lipofectamine 2000 was then mixed into each well and
cultured in Opti-MEM RPMI-1640 medium. Following 6 h of culturing,
the fluid was changed back to RPMI-1640 medium containing 10%
FBS.
Cell viability
3-(4,5-Dimethylthiazol-2-yl)-2,5-dipheny-ltetrazolium bromide (MTT;
Solarbio Life Sciences, Beijing, China) was used to detect cell
viability. The cells (1×104 cells/well) were seeded in
96-well plates and cultured for 24, 48 72 h at 37°C. The cells were
then washed with PBS buffer and 20 μl MTT reagent were added
to each well and cultured for a further 4 h. Subsequently, 150
μl DMSO were added after the MTT was removed. The optical
density was measured at 562 nm using a microplate reader (Thermo
Fisher Scientific).
Cell scratch wound assay
The cells were seeded in 6-well plates and incubated
at 37°C for 24 h. A wound was drawn in the center of the plate
using the sterile 100 μl pipette tip, and PBS was used to
gently wash the cells 3 times and serum-free medium was then added.
Cell migrations were observed using an inverted microscope (CKX53,
Olympus, Tokyo, Japan) per 24 h. The scratch area was measured
using ImageJ software Version 1.49 (NIH, Bethesda, MD, USA).
Cell invasion assay
Cell invasion was performed by Matrigel-coated
Transwell cell culture chambers. Following transfection, the cells
were resuspended in serum-free medium and 1×104 cells
were added to the upper chamber coated with Matrigel. In the lower
24-well chamber, DMEM medium containing 10% fetal bovine serum was
added and the cells were first incubated for 24 h for 37°C and were
then fixed with 1% formaldehyde for 10 min at 25°C and finally
stained with 0.5% crystal violet (Leagene Biotechnology, Beijing,
China) for a further 5 min at room temperature. Invasion cells were
counted at ×200 magnification.
Bioinformatics analysis
Data regarding 501 cases of PCa were downloaded from
the TCGA database. The association between MTDH and overall
survival was detected by the Kaplan-Meier method, followed bythe
log rank test. Differential expression levels of miRNAs in normal
prostate tissues and cancer tissues were screened by DianaTools
(http://diana.imis.athena-innovation.gr/DianaTools/index.php?r=site/page&view=software).
DianaTools, miRTarBase (http://mirtarbase.mbc.nctu.edu.tw/php/search.php),
miRWalk (http://zmf.umm.uni-heidelberg.de/apps/zmf/mirwalk2/)
and TargetScan (http://www.targetscan.org/vert_72/) were used to
predict the miRNAs targeting MTDH.
Luciferase assay
Empty vector or miR-145-5p/miR-145-3p/
miR-499a-5p/miR-22-3p and luciferase reporter comprising 3′-UTR of
MTDH wild-type or mutant fragment (GeneChem, Shanghai, China) were
co-transfected using Lipofectamine 2000 (Invitrogen/Thermo Fisher
Scientific) into 293T cells (ACS-4500; ATCC) cultured in a 96-well
plate. Following 48 h of transfection, the cells were harvested and
luciferase activity was measured the chemiluminescence using the
dual-luciferase reporter assay (normalization with Renilla
luciferase activity, Promega, Madison, WI, USA) according to the
manufacturer’s instructions.
Reverse transcription-quantitative PCR
(RT-qPCR)
According to the manufacturer’s instructions, total
RNA was isolated from the tissues or cells using TRIzol regent
(Invitrogen/Thermo Fisher Scientific). The GoScriptTM Reverse
Transcription kit (Promega) was used for reverse transcription at
37°C for 60 min and at 85°C for 5 min. qPCR was carried out on SYBR
Fast qPCR Mix (Invitrogen/Thermo Fisher Scientific) for MTDH,
hsa-miR-145-5p, hsa-miR-145-3p, hsa-m iR- 499a-5p, hsa-m iR-22-3p,
hsa-m iR-375, hsa-miR-30e-5p, hsa-miR-30a-5p, hsa-miR-200b-3p,
hsa-miR-34b-5p, hsa-miR-217, hsa-miR-378a-5p, hsa-miR-200c-3p,
hsa-miR-136-5p, hsa-miR-30c-5p, hsa-miR-320a and hsa-miR-30b-5p.
The primer sequences are listed in Table II. Samples were run using the
following cycling parameters: 95°C for 5 min, 95°C for 10 sec,
followed by 40 cycles of 60°C for 20 sec and 72°C for 10 sec. After
completion of the PCR amplification, the 2-ΔΔCq
[relative quantity (RQ)] method was used to detect comparative
quantification (33). Primers were
synthetized commercially (Invitrogen/Thermo Fisher Scientific).
| Table IIPrimers used in RT-qPCR. |
Table II
Primers used in RT-qPCR.
| Gene | Primer | Sequence |
|---|
| MTDH | Forward |
5′-TGCCTCCTTCACAGACCAA-3′ |
| Reverse |
5′-TCGGCTGCAGATGAGATAG-3′ |
| hsa-miR-145-5p | Forward |
5′-GTCCAGTTTTCCCAGGAATCCCT-3′ |
| Reverse |
5′-TGGTGTCGTGGAGTCG-3′ |
| hsa-miR-145-3p | Forward |
5′-GCCCTGTAGTGTTTCCTACTT-3′ |
| Reverse |
5′-GTGCAGGGTCCGAGGT-3′ |
|
hsa-miR-499a-5p | Forward |
5′-GACTTGCAGTGATGTTTGTCGT-3′ |
| Reverse |
5′-TGTCGTGGAGTCGGCAATTG-3′ |
| hsa-miR-22-3p | Forward |
5′-AAGCTGCCAGTTGAAGAACTGTA-3′ |
| Reverse |
5′-GCTGTCAACGATACGCTACGTAAC-3′ |
| hsa-miR-375 | Forward |
5′-TTGTTCGTTCGGCTCGCG-3′ |
| Reverse |
5′-TTTGGCACTAGCACATT-3′ |
| hsa-miR-30e-5p | Forward |
5′-GGCGTGTAAACATCCTTGACTG-3′ |
| Reverse |
5′-GTGCAGGGTCCGAGGT-3′ |
| hsa-miR-30a-5p | Forward |
5′-ACATCCTCGACTGGAAGGTC-3′ |
| Reverse |
5′-TGTCGTGGAGTCGGCAATTG-3′ |
|
hsa-miR-200b-3p | Forward |
5′-TGCCTGGTAATGATGAGTCGT-3′ |
| Reverse |
5′-GTGTCGTGGAGTCGGCAATT-3′ |
| hsa-miR-34b-5p | Forward |
5′-TGATTGGTCGTATCCAGTGCAA-3′ |
| Reverse |
5′-GTATCCAGTGCGTGTCGTGG-3′ |
| hsa-miR-217 | Forward |
5′-TCCTAATGCATTGCCTTCAGC-3′ |
| Reverse |
5′-CGGCAATTGCACTGGATACG-3′ |
|
hsa-miR-378a-5p | Forward |
5′-TCCAGGTCCTGTGTGTCGTA-3′ |
| Reverse |
5′-GTATCCAGTGCGTGTCGTGG-3′ |
|
hsa-miR-200c-3p | Forward |
5′-TCGTCTTACCCAGCAGTG-3′ |
| Reverse |
5′-CGGCAGTATTAGAGACTCC-3′ |
| hsa-miR-136-5p | Forward |
5′-TGGAGTCGTATCCAGTGCAA-3′ |
| Reverse |
5′-GTCGTATCCAGTGCGTGTCG-3′ |
| hsa-miR-30c-5p | Forward |
5′-GCTTCGGCAGCACATATACTAAAAT-3′ |
| Reverse |
5′-CGCTTCACGAATTTGCGTGTCAT-3′ |
| hsa-miR-320a | Forward |
5′-ATCCAGTGCAGGGTCCGAGG-3′ |
| Reverse |
5′-CGCGGTTAAAAGCTGGGTTGAGA-3′ |
| hsa-miR-30b-5p | Forward |
5′-AGCTGTCGTATCCAGTGCAA-3′ |
| Reverse |
5′-GTCGTATCCAGTGCGTGTCG-3′ |
| β-actin | Forward |
5′-CATGTACGTTGCTATCCAGGC-3′ |
| Reverse |
5′-CTCCTTAATGTCACGCACGA-3′ |
Western blot analysis
RIPA lysis buffer (Beyotime, Shanghai, China) was
used to extract the protein, which was obtained from the tissues
and cells. The concentration of the proteins was detected using a
BCA protein kit (Beyotime). Aliquots of protein were separated by
12% SDS-PAGE and resolved proteins were transferred onto
polyvinylidene fluoride (PVDF) membranes (Millipore, Billerica, MA,
USA), which were blocked in 5% milk PBS with 0.1% Triton X-100 and
incubated with anti-MTDH primary antibody (1:1,000; ab227981;
Abcam) overnight at 4℃. The membranes were then incubated with the
appropriate HRP-conjugated secondary antibody (1:10,000; cat. no.
10285-1-AP; Proteintech). Protein bands were detected with ECL
(Thermo Fisher Scientific) and visualized using Quantity One
software Version 4.6.2 (Bio-Rad, Hercules, CA, USA).
Statistical analysis
GraphPad Prism software version 6.0 was used to
conduct statistical analysis. All data are presented as the means ±
standard deviation (SD). Differences were analyzed using one-way
analysis of variance (ANOVA) followed by Tukey’s multiple
comparisons test. The Chi-square test or Fisher’s test were carried
out to examine the association between MTDH expression and the
clinicopathological characteristics of the patients with PCa. The
cut-off value of the patients with a high MTDH expression was
>1(RQ) and the cut-off value of patients with a low MTDH
expression was <1(RQ). A value of P<0.05 was considered to
indicate a statistically significant difference.
Results
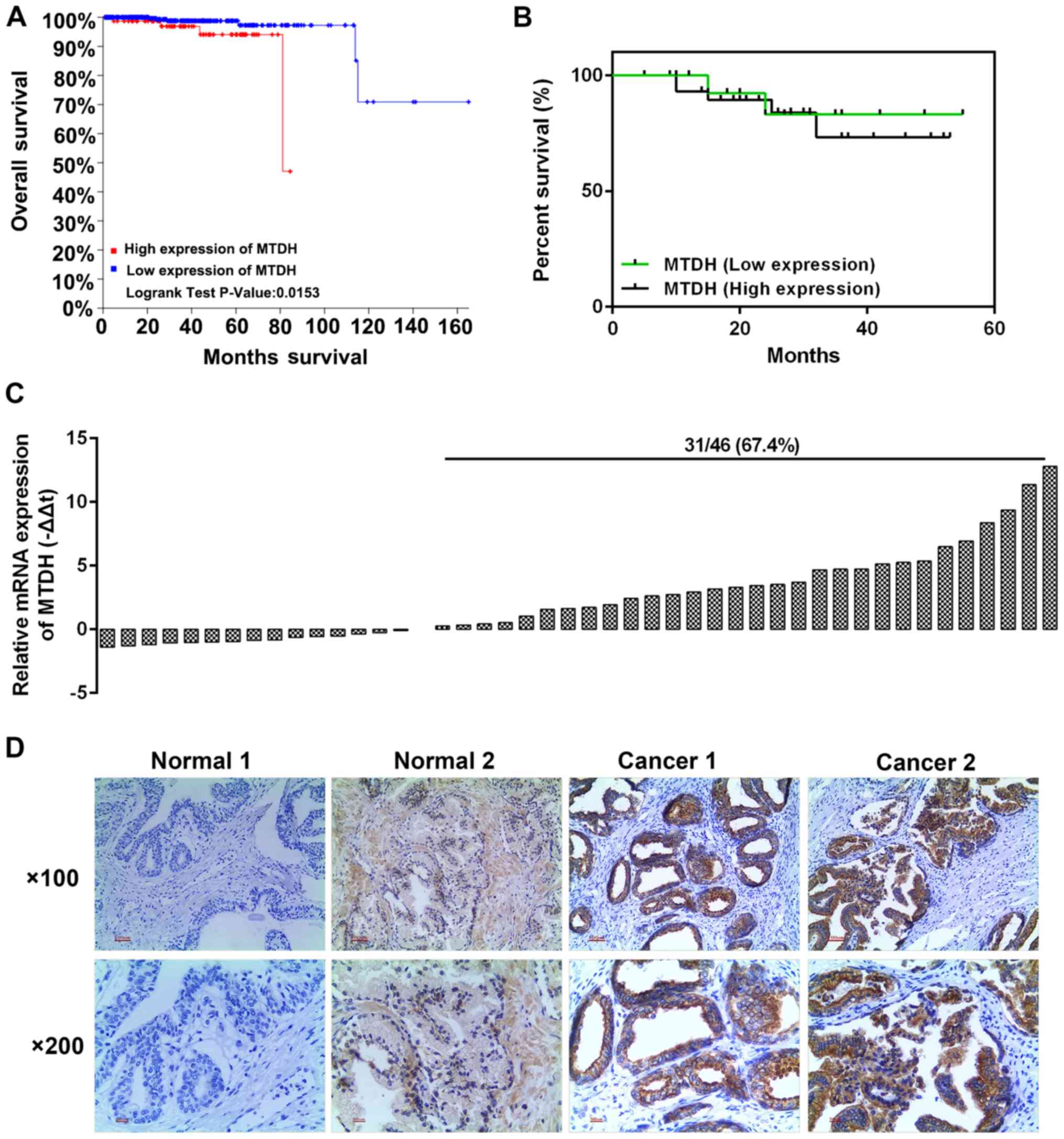
MTDH is upregulated in PCa tissues and is
associated with a poor prognosis of patients with the disease
The analysis of 501 PCa cases from the TCGA database
revealed that a high expression of MTDH was associated with a
relatively low overall survival rate (P=0.0153; Fig. 1A). We selected 46 cases of PCa
tissues and adjacent normal tissues. The results of RT-qPCR
revealed that a high expression of MTDH existed 31 PCa tissues
(Fig. 1C). Kaplan-Meier analysis
revealed that the patients with a high expression of MTDH had a
poor 5-year survival rate (Fig.
1B). Furthermore, immunohistochemical staining was performed to
determine protein expression in two randomly selected patients. As
shown in Fig. 1D, the selected
patient tissues were representative in a way that, MTDH had high
expression in the tumor tissues with a few brown granules located
in the nucleus and the majority of brown granules were confined to
the cytoplasm and cell membrane (Fig.
1D). However, the normal tissues had a negative or weak
expression (resembling the color of popcorn) of MTDH. Furthermore,
the analysis of the association between MTDH expression and the
clinicopathological characteristics of the patients with PCa
revealed that MTDH expression was significantly associated with
tumor size and osseous metastasis (P<0.05; Table I). The median survival in both
groups exhibited no significant difference, and such a phenomenon
may be explained by the fact that the sample size was not large
enough.
Expression of MTDH in normal prostate and
PCa cell lines
Two normal human prostate epithelial cell lines
(PWR-1E and RWPE-1 cells) and 6 PCa cell lines (PC-3, LNCaP, DU145,
C4-2, 22Rv1 and NCI-H660 cells) were examined to determine the
expression levels of MTDH. The 2 normal cells exhibited no obvious
differences in the mRNA and protein expression of MTDH. However,
compared with the 2 normal cells, almost all 6 PCa cell lines
exhibited a significantly increased expression of MTDH at both the
mRNA and protein level (P<0.01; Fig. 2A-C).
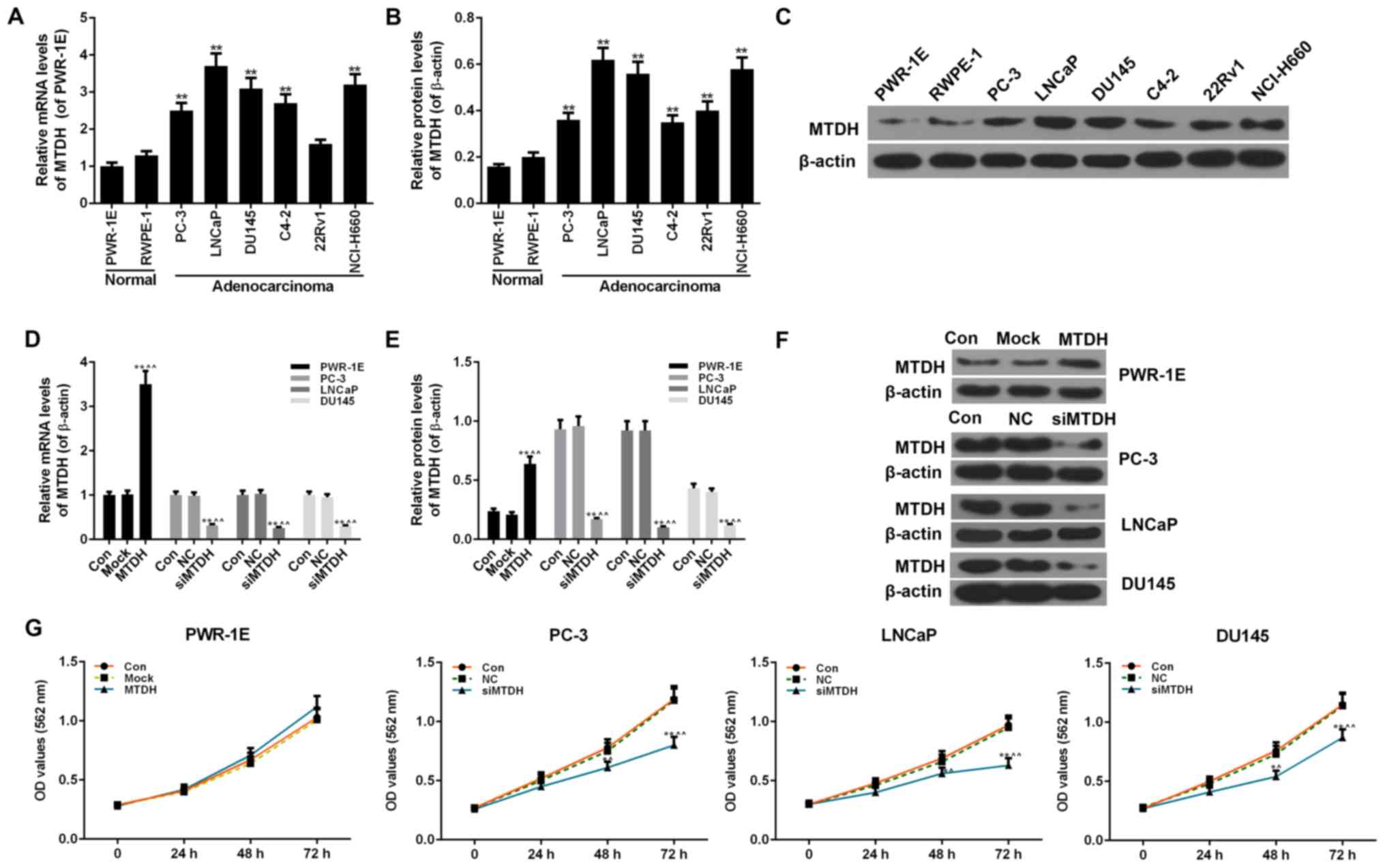 | Figure 2Expression of MTDH in prostate cancer
(PCa) and normal cell lines, and the effect of the overexpression
or silencing MTDH on cell viability. (A) RT-qPCR was used to detect
the mRNA level of MTDH in 6 PCa (PC-3, LNCaP, DU145, C4-2, 22Rv1
and Ncl-H660 cells) and 2 normal cell lines (PWR-1E and RWPE-1
cells). (B and C) Western blot analysis was used to detect the
protein level of MTDH in 6 PCa (PC-3, LNCaP, DU145, C4-2, 22Rv1 and
Ncl-H660 cells) and 2 normal cell lines (PWR-1E and RWPE-1 cells).
Relative protein levels of MTDH are shown as bar diagrams
(**P<0.01, compared with normal cell lines). The
expression of each protein in cells was determined following
normalization with a loading control, β-actin. (D-F) Transfection
efficiency of MTDH in prostate normal and cancer cell lines
following transfection with MTDH overexpression plasmid or MTDH
siRNA, both at the (D) mRNA and (E and F) protein levels determined
by RT-qPCR and western blot analysis. β-actin served as an internal
control. (G) MTT assay was performed to determine the effects of
the overexpression/silencing MTDH on cell viability at 24, 48 and
72 h [*P<0.05 and **P<0.01, compared
with control; ^P<0.05 and ^^P<0.01,
compared with mock/negative control (NC) groups]. Data are shown as
the means ± SD from 3 independent experiments. MTDH,
metadherin. |
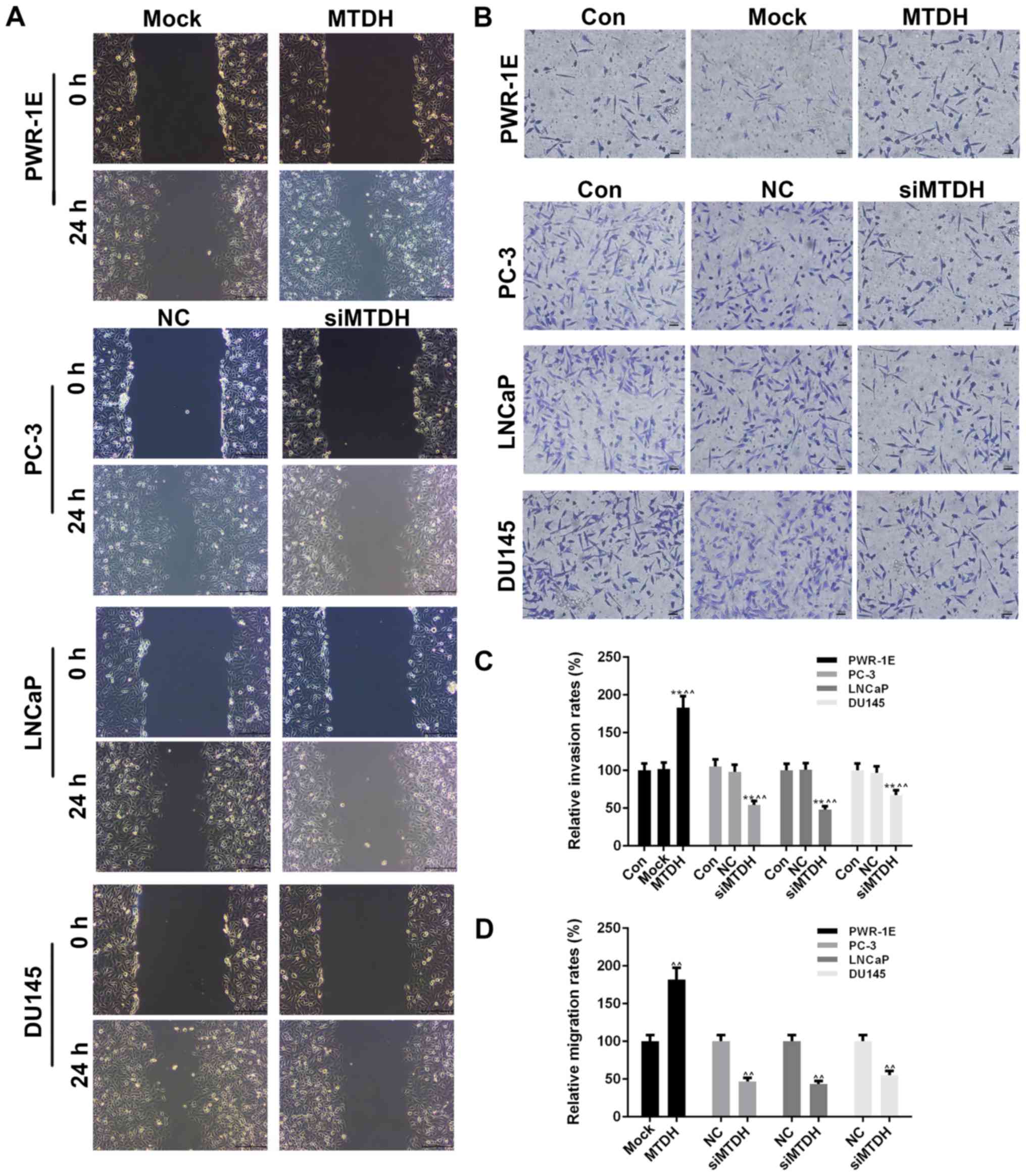
Effects of the overexpression or
silencing of MTDH on cell viability, and on the invasion and
migration of normal and PCa cell lines
The overexpression of MTDH was successfully induced
in the normal human prostate epithelial cell line, PWR-1E, by
transfection with an MTDH overexpression plasmid. The results of
RT-qPCR and western blot analysis revealed that the expression of
MTDH significantly increased in the PWR-1E cells by transfection
with MTDH overexpression plasmid, compared to the control
(P<0.01; Fig. 2D-F). In
addition, 3 PCa cell lines (PC-3, LNCaP and DU145) that had a
higher expression of MTDH or studied widely were transfected with
siRNA against MTDH (siMTDH). No differences were observed between
the control and NC groups in terms of the expression of MTDH.
However, in the siMTDH groups, the mRNA and protein levels of MTDH
exhibited a significant decrease (P<0.01; Fig. 2D-F). As regards cell viability, the
overexpression of MTDH slightly enhanced the viability of the
normal PWR-1E cells in comparison to the control or mock groups
(P>0.05; Fig. 2G). In the PC-3,
LNCaP and DU145 PCa cells, the silencing of MTDH significantly
decreased cell viability starting from 48 h in the siMTDH groups
(48 h, P<0.05; 72 h, P<0.01; Fig. 2G).
Cell migration and invasion were also detected by
scratch wound assay and Transwell assay, respectively (Fig. 3A and B). At 24 h after the scratch
wound was created, the overexpression of MTDH significantly
increased the migration of the PWR-1E cells (P<0.01; Fig. 3D), while the silencing of MTDH
significantly decreased the migration of the PCa cells (P<0.01).
The effects of the overexpression or silencing of MTDH on cell
invasion were similar to those observed on cell migration.
Specifically, the overexpression of MTDH increased the invasion of
the PWR-1E, while the silencing of MTDH decreased the invasion of
the 3 PCa cell lines (P<0.01; Fig.
3C).
Screening of miRNAs associated with MTDH
and the expression of miRNAs in normal prostate normal and PCa cell
lines
By using the DianaTools, TargetScan, miRWalk and
miRTarBase databases, 16 miRNAs were found to be associated with
MTDH in PCa and normal tissues (Fig.
4B). As shown by RT-qPCR, similar results were observed for
hsa-miR-145-5p, hsa-miR-145-3p, hsa-miR-499a-5p and hsa-miR-22-3p,
in that they exhibited high expression levels in the normal PWR-1E
cells, and relatively low expression levels in the PC-3, LNCaP and
DU145 PCa cells (P<0.01; Fig.
4A). Similar results were also observed for hsa-miR-30e-5p,
hsa-miR-375, hsa-miR-30a-5p, hsa-miR-200b-3p, hsa-miR-200c-3p,
hsa-miR-30c-5p and hsa-miR-30b-5p, in that they exhibited high
expression levels in the PC-3, LNCaP and DU145 cells, but low
expression levels in the PWR-1E cells (P<0.01). The expression
levels of other miRNAs did not exhibit clear differences between
the normal and cancer cell lines.
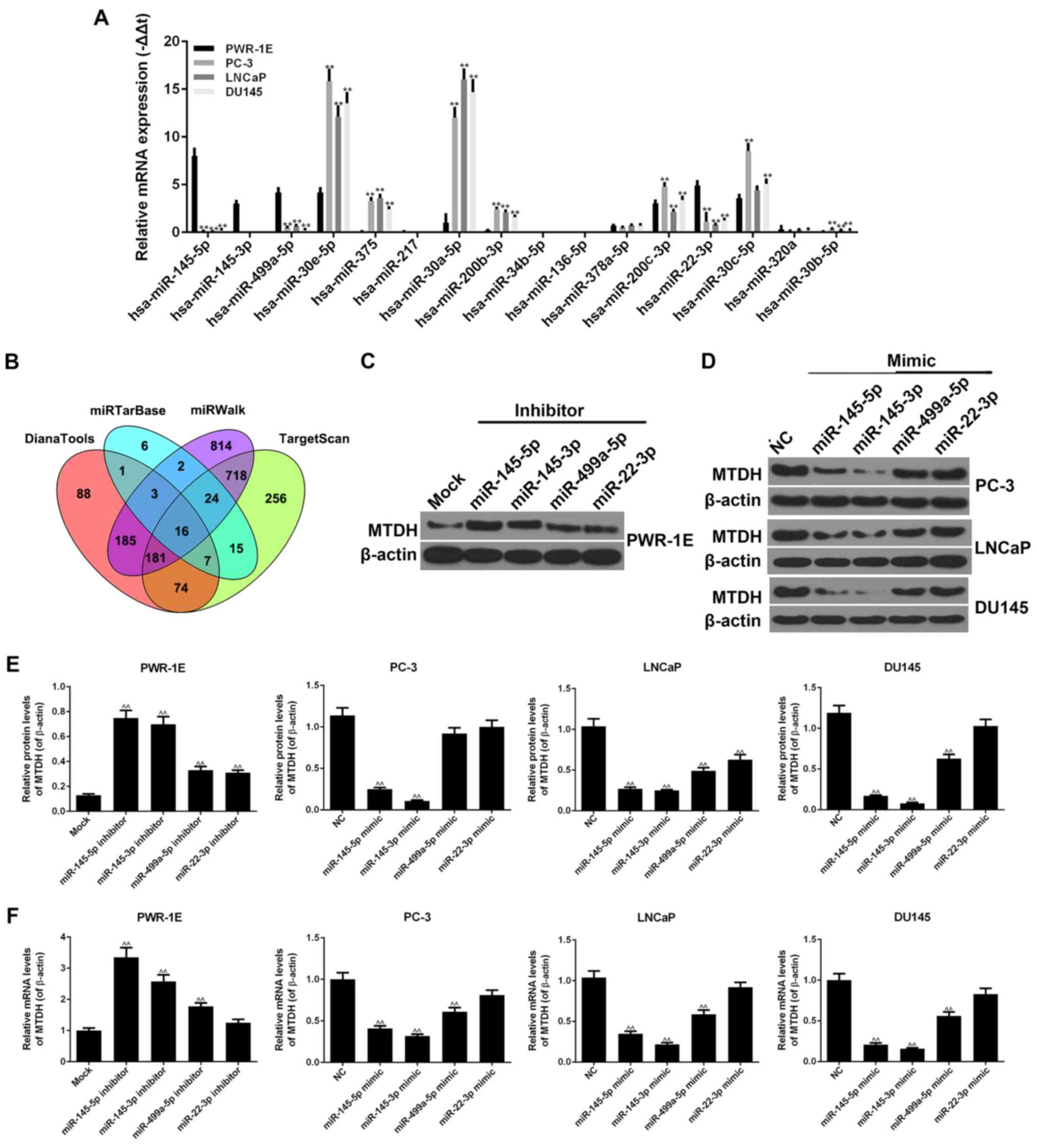 | Figure 4miRNAs are closely associated with
MTDH. (A) RT-qPCR was used to detect the expression levels of 16
miRNAs in PWR-1E, PC-3, LNCaP and DU145 cells. (B) The 16 miRNAs
associated with MTDH were screened by DianaTools, miRTarBase,
miRWalk and TargetScan (**P<0.01, compared to normal
PWR-1E cells). (C) Four representative miRNAs, miR-145-5p,
miR-145-3p, miR-499a-5p and miR-22-3p, were selected to perform the
following experiments. The inhibitors of these 4 miRNAs were used
to determine their effects on the protein expression of MTDH in
PWR-1E cells. β-actin served as an internal control. (D) The mimics
of these 4 miRNAs were used to determine their effects on the
protein expression of MTDH in PC-3, LNCaP and DU145 cells. β-actin
served as an internal control. (E) The effects of the inhibitors or
mimics of the 4 miRNAs at the protein levels of MTDH in PWR-1E or 3
prostate cancer (PCa) cells are shown as bar diagrams. (F) The
effects of the inhibitors or mimics of the 4 miRNAs on the mRNA
levels of MTDH in PWR-1E or 3 PCa cells are shown as bar diagrams.
Data are shown as the means ± SD from 3 independent experiments
[^^P<0.01, compared to mock/negative control (NC)
group]. MTDH, metadherin. |
Effects of inhibitors or mimics of miRNAs
on the expression of MTDH
We selected 4 representative miRNAs, miR-145-5p,
miR-145-3p, miR-499a-5p and miR-22-3p, which had a high expression
in the PWR-1E cells, but a low expression in the PC-3, LNCaP and
DU145 cells. The inhibitors of these 4 miRNAs were transfected into
the PWR-1E cells. The results revealed that all 4 miRNA inhibitors
increased the protein and mRNA expression levels of MTDH (Fig. 4C, E and F). In particular, the
increasing effects of the miR-145-5p and miR-145-3p inhibitors on
the expression of MTDH were more significant than those of the
other inhibitors (P<0.01). miR-22-3p inhibitor exerted a
comparative effect on the mRNA expression of MTDH in comparison
with the mock group. In addition, the mimics of these 4 miRNAs were
transfected into 3 PCa cell lines. The miR-145-5p and miR-145-3p
mimics noticeably decreased the expression of MTDH at both the
protein and mRNA level (P<0.01; Fig. 4D-F). The miR-22-3p mimic hardly
inhibited the expression of MTDH in the PCa cells, apart from the
LNCaP cells. In the PC-3, LNCaP and DU145 cell lines, the
miR-499a-5p mimic decreased the expression of MTDH (P<0.01),
although not at the protein level in the PC-3 cells.
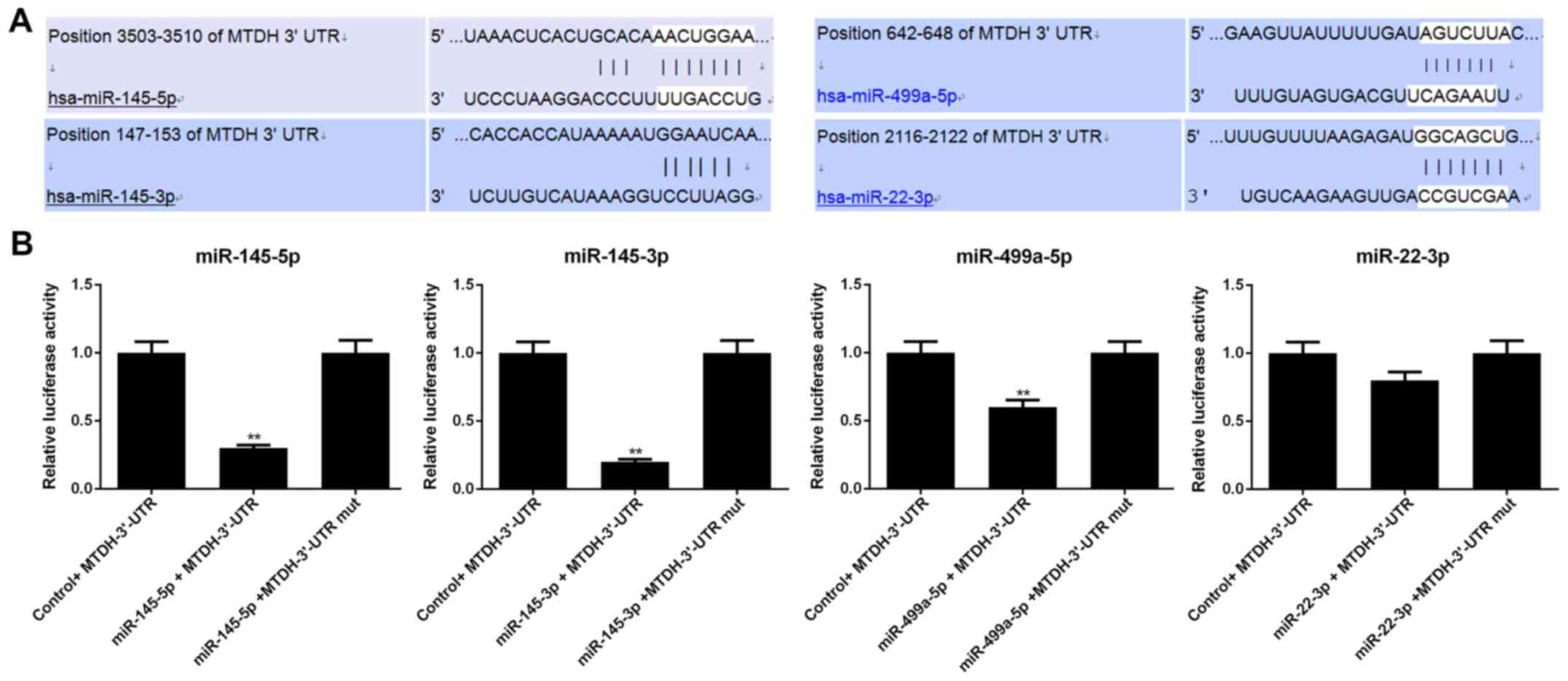
MTDH is a direct target of miR-145-5p,
miR-145-3p and miR-499a-5p
Luciferase reporter assay was conducted to determine
whether MTDH is regulated by miR-145-5p, miR-145-3p, miR-499a-5p
and miR-22-3p. Luciferase reporter was subcloned with the wild-type
or mutant sequence of MTDH and was then co-transfected with
miR-145-5p, miR-145-3p, miR-499a-5p, miR-22-3p or miR-NC into 293T
cells (Fig. 5). The luciferase
activities significantly decreased after miR-145-5p/
miR-145-3p/miR-499a-5p and MTDH 3′-UTR-wt were co-transfected into
the 293T cells (P<0.01); however, the luciferase activities in
the cells co-transfected with MTDH 3′-UTR-mut and
miR-145-5p/miR-145-3p/miR-499a-5p (P>0.05) remained stable. In
addition, neither MTDH 3′-UTR-wt nor MTDH 3′-UTR-mut with miR-22-3p
exerted obvious effects on luciferase activity (P>0.05).
Effects of inhibitors or mimics of
miR-145-5p/miR-145-3p/ miR-499a-5p on cell viability
In order to determine the effects of
miR-145-5p/miR-145-3p/miR-499a-5p on normal prostate or PCa cell
viability, inhibitors or mimics were used to determine the effects
on cell viability. The result revealed that inhibitors of these 3
miRNAs weakly increased PWR-1E cell viability (P>0.05; Fig. 6A). However, the mimics of
miR-145-5p and miR-145-3p exerted a similar effect on PCa cell
viability, which was significantly decreased at 48 h (P<0.05).
miR-499a-5p slightly attenuated the viability of the PC-3, LNCaP
and DU145 cells (P>0.05; Fig.
6A).
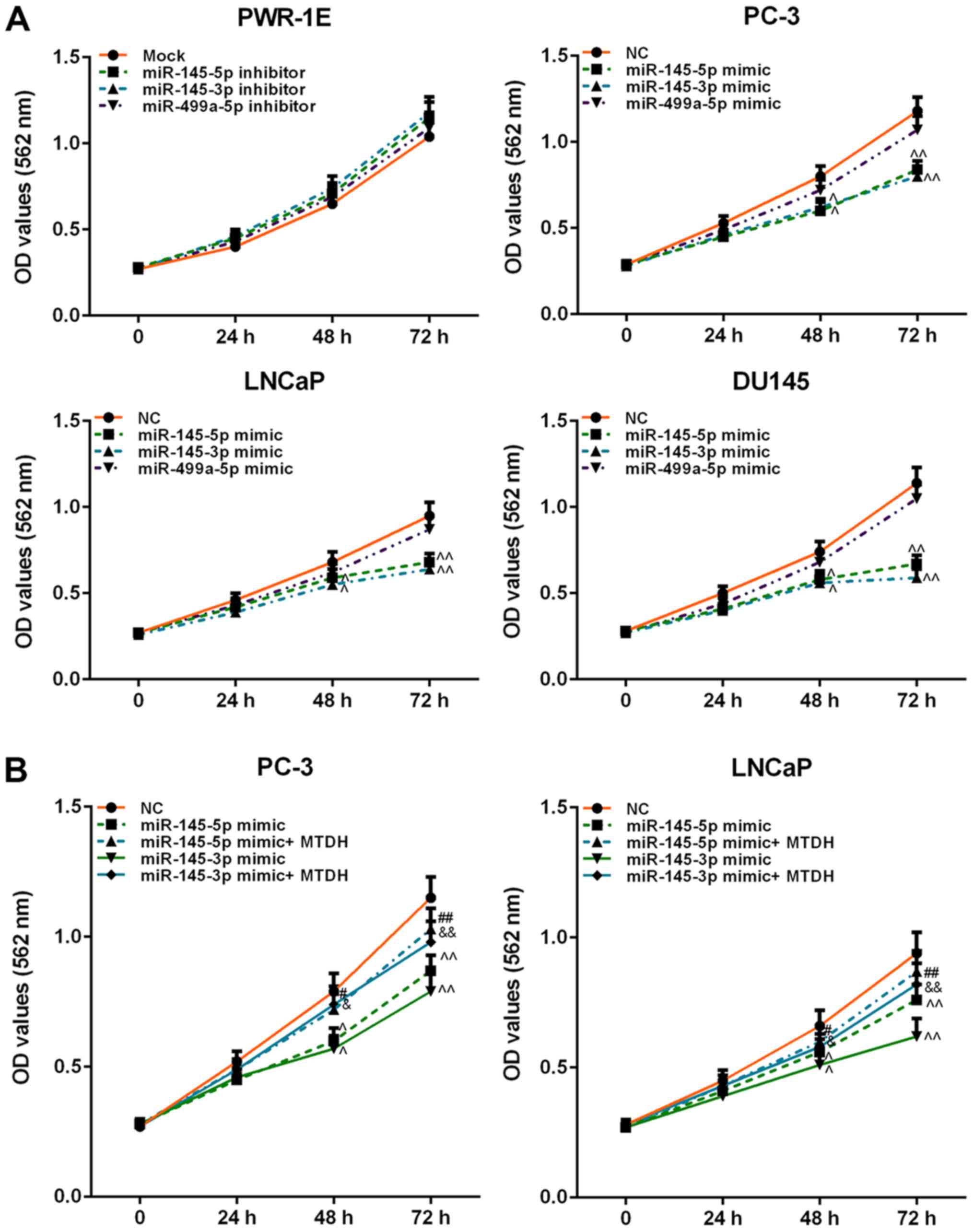 | Figure 6Effects of inhibitors or mimics of
miR-145-5p, miR-145-3p and miR-499a-5p, as well as in combination
with MTDH on cell viability. (A) Effects of inhibitors or mimics of
miR-145-5p, miR-145-3p and miR-499a-5p on PWR-1E or PC-3, LNCaP,
DU145 cell viability, detected by MTT assay. (B) Effects of MTDH on
decreasing cell viability induced by miR-145-5p/ miR-145-3p mimic
in PC-3 and LNCaP cells. Data are shown as the means ± SD from 3
independent experiments [^P<0.05 and
^^P<0.01, compared with mock/negative control (NC);
#P<0.05 and ##P<0.01, compared with
miR-145-5p mimic; &P<0.05 and
&&P<0.01, compared with miR-145-3p mimic].
MTDH, metadherin. |
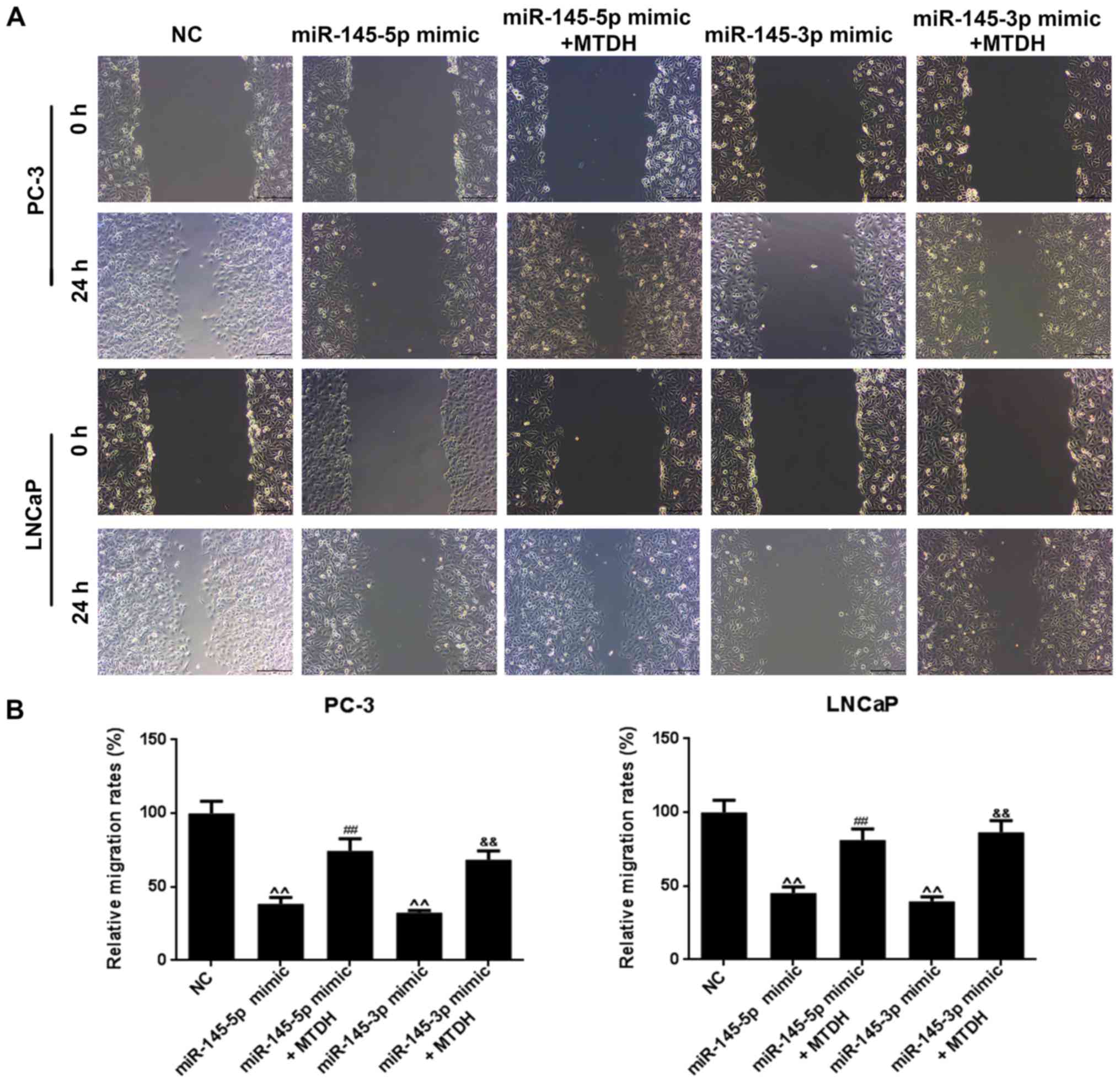
MTDH overexpression reverses the
suppressive effects of miR-145-5p and miR-145-3p mimics on the
viability, migration and invasion of PCa cells
Furthermore, we examined the effects of MTDH in
combination with mimics of miR-145-5p/miR-145-3p on PCa cell
viability, cell migration and invasion. As shown in Fig. 6B, both miR-145-5p and miR-145-3p
mimics significantly decreased PC-3 and LNCaP cell viability at 48
h, compared to the NC group (P<0.05; Fig. 6B). However, when the MTDH
overexpression plasmid and miR-145-5p /miR-145-3p mimics were
co-transfected into the PC-3 or LNCaP cells, cell viability
significantly increased at 72 h in comparison with the group
transfected with the mimics alone (P<0.01). As regards cell
migration (Fig. 7A), the results
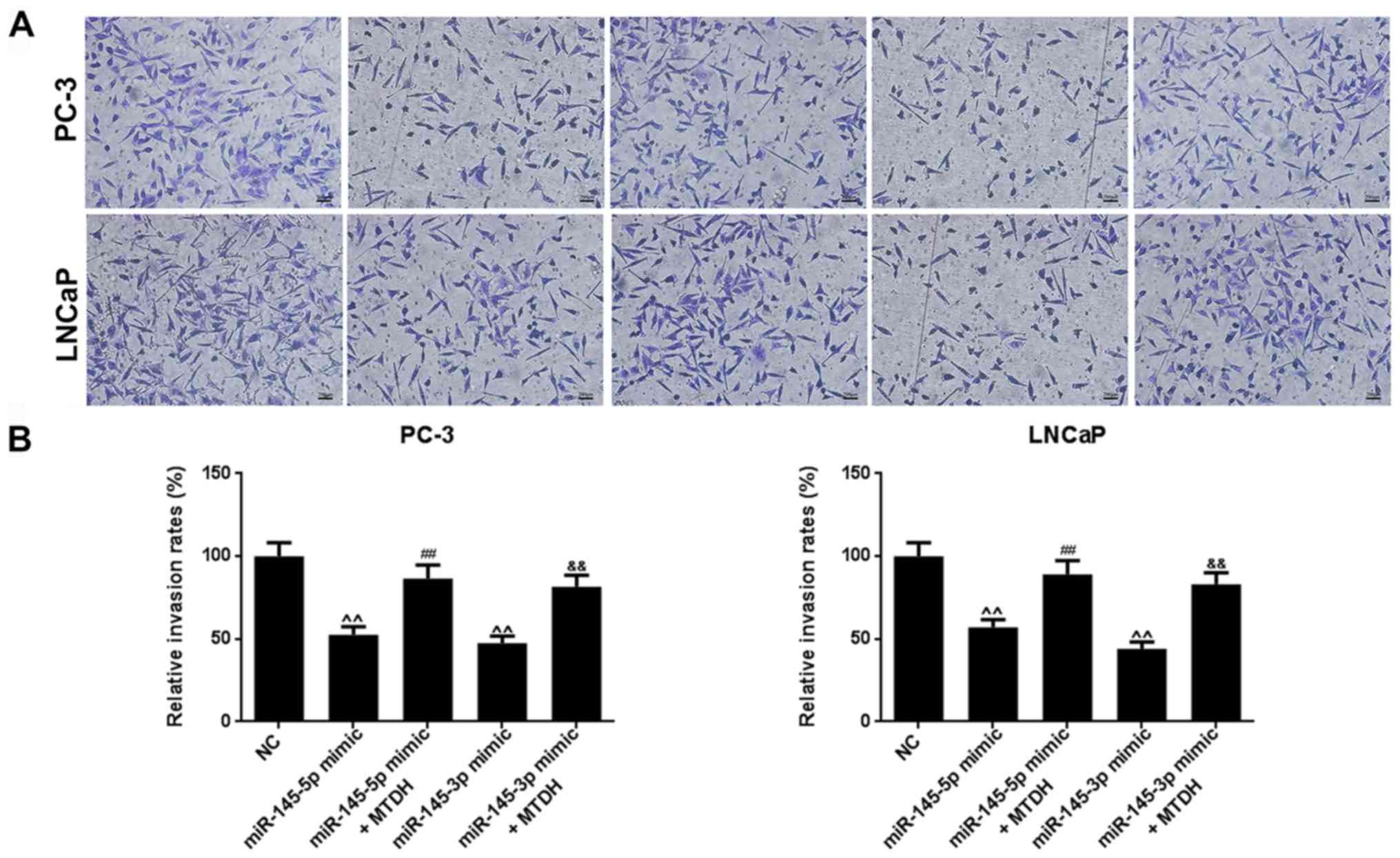
revealed that miR-145-5p and miR-145-3p mimics significantly
inhibited PC-3 and LNCaP cell migration (P<0.01; Fig. 7B). Similarly, as shown in Fig. 8, miR-145-5p and miR-145-3p mimics
significantly inhibited PC-3 and LNCaP cell invasion (P<0.01,
Fig. 8). However, the
overexpression of MTDH, reversed the inhibitory effects of the 2
miRNAs on migration and invasion (P<0.01).
miR-145-5p or miR-145-3p directly target
MTDH to negatively regulate its expression
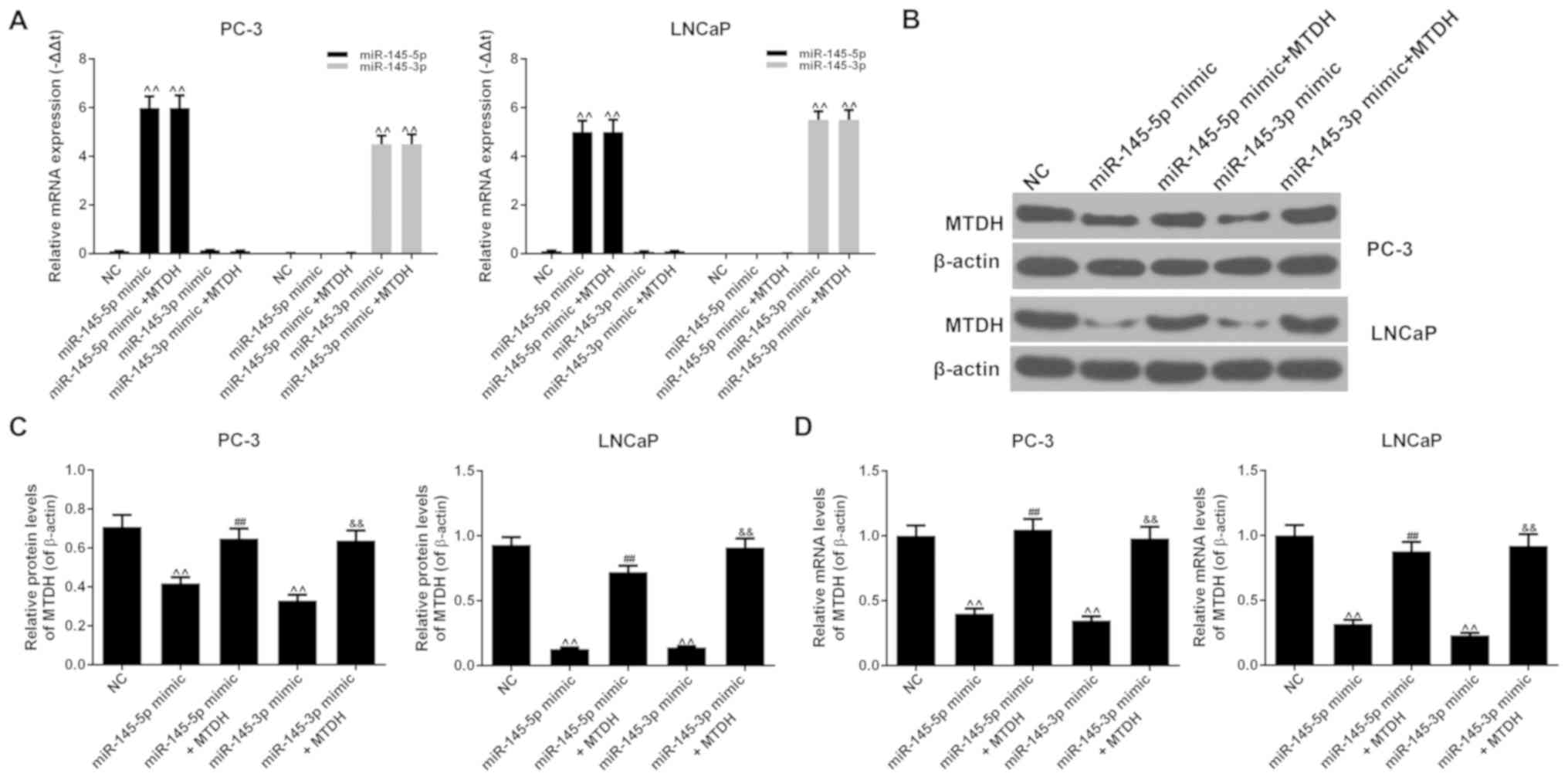
The results revealed that the mRNA expression levels
of miR-145-5p or miR-145-3p increased significantly when miR-145-5p
or miR-145-3p mimic were respectively transfected into the PC-3 or
LNCaP cells (P<0.01; Fig. 9A).
The overexpression of MTDH did not affect the expression of
miR-145-5p or miR-145-3p compared to the NC group (P<0.01). The
results also revealed that miR-145-5p mimic noticeably decreased
the protein and mRNA expression levels of MTDH, compared to the NC
group in both the PC-3 and LNCaP cells (P<0.01; Fig. 9B-D). However, when miR-145-5p mimic
and MTDH were used in combination, the expression of MTDH increased
at both the protein and mRNA level (P<0.01). Similar results
were also observed with miR-145-3p mimic (P<0.01). Therefore,
the suppression of PCa cell growth and metastasis induced by the
upregulation of miR-145-5p/miR-145-3p involve the downregulation of
MTDH expression.
Discussion
MTDH is a highly conserved protein located at 8q22
(30). It is believed that MTDH
may be used not only as an independent molecular marker to
determine tumor prognosis, but also a molecular target for
antitumor therapy (34,35). Li et al found that the mRNA
and protein expression levels of MTDH in breast cancer cells were
significantly higher than those in normal cells; their statistical
data indicated that the expression of MTDH was associated with the
clinical features of breast cancer, including staging, the number
of positive lymph nodes and tumor type; therefore, MTDH was
considered as an independent prognostic factor (36). In this study, we found that a high
expression of MTDH was closely associated with a poor prognosis in
501 PCa cases analyzed from the TCGA database. Thus, we were
interested in further exploring the effects of MTDH on PCa. For
this purpose, 46 cases of PCa tissues and adjacent normal tissues
were collected, and RT-qPCR and IHC revealed that MTDH had a
significantly increased expression in the cancer tissues. Moreover,
Kaplan-Meier analysis also confirmed that the upregulation of MTDH
was related to a poor 5-year survival rate of patients with PCa.
The analysis of the association between MTDH expression and the
clinicopathological characteristics of patients with PCa revealed
that MTDH expression was significantly associated with tumor size
and osseous metastasis. Cellular experiments revealed that MTDH had
a high expression in several PCa cell lines, but a low expression
in 2 normal prostate epithelial cells. Thus, a difference between
the mRNA expression level and protein expression level of MTDH in
22RV1 cells was identified, and such a phenomenon may be explained
by the participation of other factors, such as transcription
factors, small RNA, methylation and acetylation. Studies have also
suggested that MTDH may be expected to be a novel target in
tumor-targeted therapy as it is overexpressed in various types of
tumors, including breast cancer, neuroblastoma, esophageal cancer,
cervical cancer, non-small cell lung cancer and hepatocellular
carcinoma (36-41). The findings of this study
demonstrated that on the one hand, the silencing of MTDH inhibited
the viability, migration and invasion of PC-3, LNCaP and DU145 PCa
cells; on the other hand, the overexpression of MTDH increased
PWR-1E normal cell viability, cell migration and invasion. Some
studies, for instance, have proposed that MTDH overexpression can
enhance the adhesion of tumor cells and promote tumor metastasis
(25,42). Hu et al also found that 8q22
genomic repeated gain significantly accelerated the lung metastasis
of cancer cells (43).
miRNAs have recently become a main focus in tumor
progression. Studies have demonstrated that some miRNAs are
associated with PCa. For example, miR-200c-3p/141-3p, miR-375 and
miR-93-5p/106b-5p/25-3p have been frequently found to be
upregulated in PCa, whereas the expression levels of miR-221-3p and
miR-222-3p in PCa tissues have often been proven to be
downregulated (15-17,44-46).
In a number of tumor cells, miRNAs such as miR-145, miR-497,
miR-1297 and miR-655, have been reported to regulate cell
proliferation and invasion by targeting MTDH; however, only a few
studies to date have investigated the upstream regulatory
mechanisms of MTDH in PCa, at least to the best of our knowledge
(27,28,47).
In this study, by using the DianaTools, TargetScan, miRWalk and
miRTarBase databases, 16 miRNAs were screened to determine whether
they were associated with MTDH. As miR-145-5p, miR-145-3p,
miR-499a-5p and miR-22-3p were overexpressed in PWR-1E cells and
downregulated in the PC-3, LNCap and DU145 cells, these 4 miRNAs
were selected to help detect their association with MTDH. Western
blot analysis, RT-qPCR and dual-luciferase reporter assay confirmed
that miR-145-5p, miR-145-3p and miR-499a-5p bound to MTDH and
decreased its expression in PCa cells. MTT assay further revealed
that only miR-145-5p and miR-145-3p significantly inhibited tumor
cell viability. Previous studies have suggested that miR-145-5p and
miR-145-3p, as the dual strands of the miR-145 duplex, play roles
in a variety of tumors (48-50).
In addition, the two miRNAs can inhibit the proliferation of PCa
cells (51,52).
In addition, in this study, we found that MTDH
reversed the suppressive effects of miR-145-3p or miR-145-5p mimics
on the viability, invasion and migration of PC-3 and LNCaP cells.
The ‘rescue’ experiments confirmed that miR-145-3p and miR-145-5p
directly targeted MTDH to negatively regulate its expression,
thereby regulating cell viability and cell invasion and migration.
However, this study still had some limitations, as for example, the
‘rescue’ experiments were not validated in DU145 cells, and the
sample size of the patients with PCa was not large enough. Thus, we
aim to carry out a more comprehensive study on
miR-145-5p/miR-145-3p and MTDH in the future.
Collectively, this study, for the first time, at
least to the best of our knowledge, demonstrated the effects of the
miR-145-5p/MTDH and miR-145-3p/MTDH pathways on the growth and
metastasis of PCa cell lines in vitro, and suggests that
these pathways may become novel treatment targets for PCa.
Funding
No funding was received.
Availability of data and materials
The analyzed data sets generated during the study
are available from the corresponding author on reasonable
request.
Authors’ contributions
DP made substantial contributions to the conception
and design of the study. ZJ, WL and ZD was involved in data
acquisition, data analysis and interpretation. DP, WL and ZD
drafted the article or critically revised it for important
intellectual content. All authors agree to be accountable for all
aspects of the study in ensuring that questions related to the
accuracy or integrity of the study are appropriately investigated
and resolved. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
The Ethics Committees of the First Affiliated
Hospital and College of Clinical Medicine of Henan University of
Science and Technology approved the study. A total of 46 cases of
patients signed informed consent forms prior to participation.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have not competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
Statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cotignola J, Leonardi DB, Shahabi A, Acuña
AD, Stern MC, Navone N, Scorticati C, De Siervi A, Mazza O and
Vazquez E: Glutathione-S-transferase (GST) polymorphisms are
associated with relapse after radical prostatectomy. Prostate
Cancer Prostatic Dis. 16:28–34. 2013. View Article : Google Scholar :
|
|
3
|
Isaacs JT, Schulze H and Coffey DS:
Development of androgen resistance in prostatic cancer. Prog Clin
Biol Res. 243A:21–31. 1987.PubMed/NCBI
|
|
4
|
Han M, Partin AW, Zahurak M, Piantadosi S,
Epstein JI and Walsh PC: Biochemical (prostate specific antigen)
recurrence probability following radical prostatectomy for
clinically localized prostate cancer. J Urol. 169:517–523. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Djebali S, Davis CA, Merkel A, Dobin A,
Lassmann T, Mortazavi A, Tanzer A, Lagarde J, Lin W, Schlesinger F,
et al: Landscape of transcription in human cells. Nature.
489:101–108. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Esquela-Kerscher A and Slack FJ: Oncomirs
- microRNAs with a role in cancer. Nat Rev Cancer. 6:259–269. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Filipowicz W, Bhattacharyya SN and
Sonenberg N: Mechanisms of post-transcriptional regulation by
microRNAs: Are the answers in sight. Nat Rev Genet. 9:102–114.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Friedman RC, Farh KK, Burge CB and Bartel
DP: Most mammalian mRNAs are conserved targets of microRNAs. Genome
Res. 19:92–105. 2009. View Article : Google Scholar :
|
|
10
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Adams BD, Kasinski AL and Slack FJ:
Aberrant regulation and function of microRNAs in cancer. Curr Biol.
24:R762–R776. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Calin GA, Dumitru CD, Shimizu M, Bichi R,
Zupo S, Noch E, Aldler H, Rattan S, Keating M, Rai K, et al:
Frequent deletions and down-regulation of micro- RNA genes miR15
and miR16 at 13q14 in chronic lymphocytic leukemia. Proc Natl Acad
Sci USA. 99:15524–15529. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Garzon R, Calin GA and Croce CM: MicroRNAs
in Cancer. Annu Rev Med. 60:167–179. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ma J, Dong C and Ji C: MicroRNA and drug
resistance. Cancer Gene Ther. 17:523–531. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wach S, Nolte E, Szczyrba J, Stöhr R,
Hartmann A, Ørntoft T, Dyrskjøt L, Eltze E, Wieland W, Keck B, et
al: MicroRNA profiles of prostate carcinoma detected by
multiplatform microRNA screening. Int J Cancer. 130:611–621. 2012.
View Article : Google Scholar
|
|
16
|
Szczyrba J, Löprich E, Wach S, Jung V,
Unteregger G, Barth S, Grobholz R, Wieland W, Stöhr R, Hartmann A,
et al: The microRNA profile of prostate carcinoma obtained by deep
sequencing. Mol Cancer Res. 8:529–538. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Schaefer A, Jung M, Mollenkopf HJ, Wagner
I, Stephan C, Jentzmik F, Miller K, Lein M, Kristiansen G and Jung
K: Diagnostic and prognostic implications of microRNA profiling in
prostate carcinoma. Int J Cancer. 126:1166–1176. 2010.
|
|
18
|
Ambs S, Prueitt RL, Yi M, Hudson RS, Howe
TM, Petrocca F, Wallace TA, Liu CG, Volinia S, Calin GA, et al:
Genomic profiling of microRNA and messenger RNA reveals deregulated
microRNA expression in prostate cancer. Cancer Res. 68:6162–6170.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Szczyrba J, Nolte E, Wach S, Kremmer E,
Stöhr R, Hartmann A, Wieland W, Wullich B and Grässer FA:
Downregulation of Sec23A protein by miRNA-375 in prostate
carcinoma. Mol Cancer Res. 9:791–800. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Brown DM and Ruoslahti E: Metadherin, a
cell surface protein in breast tumors that mediates lung
metastasis. Cancer Cell. 5:365–374. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Su ZZ, Kang DC, Chen Y, Pekarskaya O, Chao
W, Volsky DJ and Fisher PB: Identification and cloning of human
astrocyte genes displaying elevated expression after infection with
HIV-1 or exposure to HIV-1 envelope glycoprotein by rapid
subtraction hybridization, RaSH. Oncogene. 21:3592–3602. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li JW, Huang CZ, Li JH, Yuan JH, Chen QH,
Zhang WF, Xu ZS, Liu YP, Li Y, Zhan MX, et al: Knockdown of
metadherin inhibits cell proliferation and migration in colorectal
cancer. Oncol Rep. 40:2215–2223. 2018.PubMed/NCBI
|
|
23
|
Chen X, Li XY, Long M, Wang X, Gao ZW, Cui
Y, Ren J, Zhang Z, Liu C, Dong K, et al: The FBXW7 tumor suppressor
inhibits breast cancer proliferation and promotes apoptosis by
targeting MTDH for degradation. Neoplasma. 65:201–209. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen Y, Wu H, Wang X, Wang C, Gan L, Zhu
J, Tong J and Li Z: Huaier Granule extract inhibit the
proliferation and metastasis of lung cancer cells through
down-regulation of MTDH, JAK2/STAT3 and MAPK signaling pathways.
Biomed Pharmacother. 101:311–321. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ma Z, Chen Y, Dong S, Xu X, Liu J, Song P,
Yu C and Dai L: AEG-1 mRNA expression in non-small cell lung cancer
is associated with increased tumor angiogenesis. Pathol Res Pract.
213:1257–1263. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Huang LL, Wang Z, Cao CJ, Ke ZF, Wang F,
Wang R, Luo CQ, Lu X and Wang LT: AEG-1 associates with metastasis
in papillary thyroid cancer through upregulation of MMP2/9. Int J
Oncol. 51:812–822. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yin Q, Han Y, Zhu D, Li Z, Shan S, Jin W,
Lu Q and Ren T: miR-145 and miR-497 suppress TGF-β-induced
epithelial-mesenchymal transition of non-small cell lung cancer by
targeting MTDH. Cancer Cell Int. 18:1052018. View Article : Google Scholar
|
|
28
|
Chen Z, Ma Y, Pan Y, Zhu H, Yu C and Sun
C: MiR-1297 suppresses pancreatic cancer cell proliferation and
metastasis by targeting MTDH. Mol Cell Probes. 40:19–26. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fan N, Zhang J, Cheng C, Zhang X, Feng J
and Kong R: MicroRNA-384 represses the growth and invasion of
non-small-cell lung cancer by targeting astrocyte elevated
gene-1/Wnt signaling. Biomed Pharmacother. 95:1331–1337. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Qiao W, Cao N and Yang L: MicroRNA-154
inhibits the growth and metastasis of gastric cancer cells by
directly targeting MTDH. Oncol Lett. 14:3268–3274. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Abrahante JE, Daul AL, Li M, Volk ML,
Tennessen JM, Miller EA and Rougvie AE: The Caenorhabditis elegans
hunchback-like gene lin-57/hbl-1 controls developmental time and is
regulated by microRNAs. Dev Cell. 4:625–637. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
34
|
Gu C, Feng L, Peng H, Yang H, Feng Z and
Yang Y: MTDH is an oncogene in multiple myeloma, which is
suppressed by Bortezomib treatment. Oncotarget. 7:4559–4569. 2016.
View Article : Google Scholar :
|
|
35
|
Noch EK and Khalili K: The role of
AEG-1/MTDH/LYRIC in the pathogenesis of central nervous system
disease. Adv Cancer Res. 120:159–192. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li J, Zhang N, Song LB, Liao WT, Jiang LL,
Gong LY, Wu J, Yuan J, Zhang HZ, Zeng MS and Li M: Astrocyte
elevated gene-1 is a novel prognostic marker for breast cancer
progression and overall patient survival. Clin Cancer Res.
14:3319–3326. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liu L, Wu J, Ying Z, Chen B, Han A, Liang
Y, Song L, Yuan J, Li J and Li M: Astrocyte elevated gene-1
upregulates matrix metalloproteinase-9 and induces human glioma
invasion. Cancer Res. 70:3750–3759. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yu C, Chen K, Zheng H, Guo X, Jia W, Li M,
Zeng M, Li J and Song L: Overexpression of astrocyte elevated
gene-1 (AEG-1) is associated with esophageal squamous cell
carcinoma (ESCC) progression and pathogenesis. Carcinogenesis.
30:894–901. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Song L, Li W, Zhang H, Liao W, Dai T, Yu
C, Ding X, Zhang L and Li J: Over-expression of AEG-1 significantly
associates with tumour aggressiveness and poor prognosis in human
non-small cell lung cancer. J Pathol. 219:317–326. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Emdad L, Sarkar D, Su ZZ, Randolph A,
Boukerche H, Valerie K and Fisher PB: Activation of the nuclear
factor kappaB pathway by astrocyte elevated gene-1: Implications
for tumor progression and metastasis. Cancer Res. 66:1509–1516.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Li C, Wu X, Zhang W, Li J, Liu H, Hao M,
Wang J, Zhang H, Yang G, Hao M, et al: AEG-1 Promotes Metastasis
Through Downstream AKR1C2 and NF1 in Liver Cancer. Oncol Res.
22:203–211. 2014. View Article : Google Scholar
|
|
42
|
Liu DC and Yang ZL: MTDH and EphA7 are
markers for metastasis and poor prognosis of gallbladder
adenocarcinoma. Diagn Cytopathol. 41:199–205. 2013. View Article : Google Scholar
|
|
43
|
Hu G, Chong RA, Yang Q, Wei Y, Blanco MA,
Li F, Reiss M, Au JL, Haffty BG and Kang Y: MTDH activation by 8q22
genomic gain promotes chemoresistance and metastasis of
poor-prognosis breast cancer. Cancer Cell. 15:9–20. 2009.
View Article : Google Scholar :
|
|
44
|
Fuse M, Kojima S, Enokida H, Chiyomaru T,
Yoshino H, Nohata N, Kinoshita T, Sakamoto S, Naya Y, Nakagawa M,
et al: Tumor suppressive microRNAs (miR-222 and miR-31) regulate
molecular pathways based on microRNA expression signature in
prostate cancer. J Hum Genet. 57:691–699. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Porkka KP, Pfeiffer MJ, Waltering KK,
Vessella RL, Tammela TL and Visakorpi T: MicroRNA expression
profiling in prostate cancer. Cancer Res. 67:6130–6135. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Tong AW, Fulgham P, Jay C, Chen P, Khalil
I, Liu S, Senzer N, Eklund AC, Han J and Nemunaitis J: MicroRNA
profile analysis of human prostate cancers. Cancer Gene Ther.
16:206–216. 2009. View Article : Google Scholar
|
|
47
|
Wang Q, Lv L, Li Y and Ji H: MicroRNA 655
suppresses cell proliferation and invasion in oral squamous cell
carcinoma by directly targeting metadherin and regulating the
PTEN/AKT pathway. Mol Med Rep. 18:3106–3114. 2018.PubMed/NCBI
|
|
48
|
Matsushita R, Yoshino H, Enokida H, Goto
Y, Miyamoto K, Yonemori M, Inoguchi S, Nakagawa M and Seki N:
Regulation of UHRF1 by dual-strand tumor-suppressor microRNA-145
(miR-145-5p and miR-145-3p): Inhibition of bladder cancer cell
aggressiveness. Oncotarget. 7:28460–28487. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Misono S, Seki N, Mizuno K, Yamada Y,
Uchida A, Arai T, Kumamoto T, Sanada H, Suetsugu T and Inoue H:
Dual strands of the miR-145 duplex (miR-145-5p and miR-145-3p)
regulate oncogenes in lung adenocarcinoma pathogenesis. J Hum
Genet. 63:1015–1028. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Mataki H, Seki N, Mizuno K, Nohata N,
Kamikawaji K, Kumamoto T, Koshizuka K, Goto Y and Inoue H:
Dual-strand tumor-suppressor microRNA-145 (miR-145-5p and
miR-145-3p) coordinately targeted MTDH in lung squamous cell
carcinoma. Oncotarget. 7:72084–72098. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Ozen M, Karatas OF, Gulluoglu S, Bayrak
OF, Sevli S, Guzel E, Ekici ID, Caskurlu T, Solak M, Creighton CJ,
et al: Overexpression of miR-145-5p inhibits proliferation of
prostate cancer cells and reduces SOX2 expression. Cancer Invest.
33:251–258. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Goto Y, Kurozumi A, Arai T, Nohata N,
Kojima S, Okato A, Kato M, Yamazaki K, Ishida Y, Naya Y, et al:
Impact of novel miR-145-3p regulatory networks on survival in
patients with castration-resistant prostate cancer. Br J Cancer.
117:409–420. 2017. View Article : Google Scholar : PubMed/NCBI
|