Introduction
Hypopharyngeal squamous cell carcinoma (HSCC) is a
common type of head and neck cancer, which has a poor prognosis and
a 5-year survival rate of 25-40% (1-3). The
majority of patients with HSCC are usually asymptomatic at the
early stages and are diagnosed at the advanced stage (4,5).
Metastasis remains a leading cause of HSCC-associated mortality,
despite marked improvements in the clinical comprehensive treatment
of this malignancy (6,7). Therefore, the underlying mechanisms
that regulate tumor invasion and metastasis should be explored to
develop novel strategies to treat HSCC.
Fascin-1 is a 55-kDa globular actin-bundling
protein, which specifically interacts with F-actin to form parallel
actin bundles, and participates in the regulation of cell adhesion,
interactions and migration (8,9).
Fascin-1 expression in normal tissues is highly restricted to
certain filopodium-rich cell types, including neurons and mature
dendritic cells (10,11), and fascin-1 expression is absent or
low in normal epithelia (12).
Conversely, fascin-1 is highly expressed in various epithelial
neoplasms, including laryngeal squamous cell carcinoma (13), oral squamous cell carcinoma
(14), and head and neck squamous
cell carcinoma (15). In addition,
fascin-1 overexpression is associated with cancer invasion and
metastasis (14,15).
Hypoxia is one of the most common types of
microenviron-mental stress in solid tumors, and is the result of
overwhelming tumor growth and inadequate blood supply (16). Hypoxia has been reported to serve a
key role in tumor metastasis (17,18).
Hypoxia-inducible factor-1 (HIF-1) is an important heterodi-meric
transcription factor composed of highly regulated HIF-1α and
constitutively expressed HIF-1β subunits. HIF-1α is a key regulator
of the cellular response to hypoxia, which has a vital role in
HIF-1 transcriptional activity (19), and controls the expression of
target genes associated with tumor invasion and metastasis
(20). HIF-1α promotes tumor
invasion and metastasis in head and neck squamous cell carcinoma
(21). A previous study also
reported that HIF-1α may promote invasion and metastasis by
upregulating fascin expression in pancreatic ductal adenocarcinoma,
and fascin is a direct target gene of HIF-1α (22).
Fascin-1 expression in HSCC, and the potential roles
of fascin-1 in the invasion and metastasis of HSCC remain unclear.
Furthermore, the molecular mechanism underlying regulation of
fascin-1 by HIF-1α under hypoxia in HSCC is still poorly
understood. The present study aimed to investigate the effects of
fascin-1 on the invasion and metastasis of HSCC, and to determine
the molecular mechanism underlying regulation of fascin-1 by HIF-1α
under hypoxia in HSCC. The results demonstrated that fascin-1 was
overexpressed in HSCC, and promoted the invasion and migration of
HSCC. Furthermore, HIF-1α promoted the invasion and migration of
HSCC by upregulating fascin-1 expression.
Materials and methods
Reagents
Cobalt chloride hexahydrate
(CoCl2·6H2O; Sigma-Aldrich; Merck KGaA) was
dissolved in deionized water to obtain a 200 mmol/l
CoCl2 stock solution. The solution was further diluted
with DMEM/F12 (Gibco; Thermo Fisher Scientific, Inc.) to achieve
the desired concentration prior to use. Rhodamine phalloidin
(Invitrogen; Thermo Fisher Scientific, Inc.) was dissolved in
methanol to yield a stock concentration of 200 U/ml. The solution
was diluted with PBS to achieve the staining solution required for
subsequent experiments. DAPI (cat. no. D1306) was purchased from
Invitrogen; Thermo Fisher Scientific, Inc. Mouse anti-human
fascin-1 (cat. no. TA807305), rabbit anti-human HIF-1α (cat. no.
ZA0552) and mouse anti-human β-actin (cat. no. TA-09) antibodies
were purchased from Origene Technologies, Inc. Mouse anti-human
HIF-1α antibody (cat. no. 79233) was purchased from Cell Signaling
Technology, Inc. Matrix metalloproteinase (MMP)-2 (cat. no.
10373-2-AP) and MMP-9 (cat. no. 10375-2-AP) antibodies were
purchased from ProteinTech Group, Inc. E-cadherin antibody (cat.
no. 610181) was purchased from BD Biosciences. Vimentin antibody
(cat. no. ab92547) was purchased from Abcam.
Immunohistochemistry
HSCC tissues and adjacent normal tissues were
obtained from 96 patients with HSCC at the Shandong Provincial ENT
Hospital Affiliated to Shandong University between January 2010 and
November 2014. Tissues were collected in accordance with the
ethical approval and institutional guidelines of the Shandong
Provincial ENT Hospital Affiliated to Shandong University. The
tissue samples were fixed in 10% neutral buffered formalin at room
temperature for 24 h, embedded in paraffin, and cut into
5-µm serial sections. Sections of each tissue sample were
stained with hematoxylin and eosin for histological diagnosis. All
diagnoses of primary HSCC were confirmed by at least two
pathologists. No patient had received chemotherapy or radiotherapy
prior to surgery. Written informed consent for tissue donation for
research purposes was obtained from each patient prior to tissue
collection, and the present study was approved by the Ethics
Committee of Shandong University. Immunohistochemical staining was
performed according to the manufacturer's protocol. The following
primary antibodies were used: Mouse anti-human fascin-1 antibody
(1:200) and rabbit anti-human HIF-1α antibody (1:100). The sections
were incubated with the primary antibodies overnight at 4°C.
Sections were then incubated with secondary biotinyl-ated goat
anti-rabbit immunoglobulin G antibody (IgG) (1:500; cat. no.
SP-9001; OriGene Technologies, Inc.) or goat anti-mouse IgG
antibody (1:500; cat. no. SP-9002; OriGene Technologies, Inc.) at
37°C for 15 min. All histopathological images were captured under
an Olympus BX53 microscope (Olympus Corporation). The results of
immunohistochemical analysis were scored by two examiners who were
blinded to the clinicopathological data. Cytoplasmic staining of
cancer or paracarcinoma cells was regarded as positive fascin-1 or
HIF-1α immunostaining. Five random fields were observed under a
light microscope at x200 magnification. For each visual field, 200
cells were counted (i.e., a total of 1,000 cells). Staining
intensity was scored as follows: 0, no staining; 1, pale yellow
staining; 2, yellowish-brown staining; and 3, brown staining. The
extent of staining was scored as follows: 0, 0% stained; 1, 1-25%
stained; 2, 26-50% stained; and 3, 51-100% stained. The final score
was determined by multiplying the scores of intensity with the
extent of staining. Therefore, the final score ranged between 0 and
9, and final scores ≥3 were considered positive expression.
Cell culture
The human FaDu cell line was purchased from the
American Type Culture Collection. FaDu cells were cultured in
DMEM/F12 (Gibco; Thermo Fisher Scientific, Inc.) containing 10%
fetal bovine serum (FBS; Biological Industries) and incubated at
37°C in a humidified atmosphere consisting of 5% CO2 and
95% air.
Silencing gene expression using small
interfering RNA (siRNA)
FaDu cells were plated at a density of
3×105 cells/well in 6-well plates. The siRNA sequences
were as follows: Control siRNA duplexes,
5′-UUCUCCGAACGUGUCACGUTT-3'; fascin-1 siRNA duplexes,
5'-GCAGCCTGAAGAAGAAGCA-3'; HIF-1α siRNA duplexes,
5'-CUAACUGGACACAGUGUGUTT-3'. The siRNAs were synthesized by
Shanghai GenePharma Co., Ltd. Lipofectamine-RNA MAX Transfection
Reagent (Invitrogen; Thermo Fisher Scientific, Inc.) was used to
transfect cells with 1.25 µl fascin-1 siRNA (20
µmol/l), 1.25 µl HIF-1α siRNA (20 µmol/l) or
1.25 µl control siRNA (20 µmol/l) according to the
manufacturer's protocol; cells were incubated with the siRNAs at
37°C for 48 h. Gene silencing effects were evaluated by western
blot analysis.
Plasmid transient transfection
FaDu cells were seeded at a density of
3×105 cells/well in 6-well plates and were transfected
with 2.5 µg GV144-fascin-1 plasmids (263 ng/µl;
Shanghai GeneChem Co., Ltd.) or 2.5 µg empty GV144 (control;
701 ng/µl) using Lipofectamine-RNA LTX Transfection Reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol; cells were incubated with the plasmids at
37°C for 48 h. The gene overexpressing effects of the plasmids were
evaluated by western blot analysis.
Fluorescently labeled phalloidin
staining
FaDu cells were seeded at a density of
4.2x104 cells/well in 48-well plates containing
coverslips. Cells cultured on a cover slip were fixed with 4%
paraformaldehyde at room temperature for 10 min and stained with 5
µl rhodamine phalloidin (6.6 µmol/l), which
specifically binds to F-actin, at room temperature for 20 min
according to the manufacturer's protocol. DAPI was used as a
counterstain for 10 min at room temperature, for detection of the
nuclei. Images were captured under a confocal microscope (TCS SPE;
Leica Microsystems GmbH).
Establishment of hypoxia model
The hypoxia model was established by
CoCl2 treatment, which is a well-known hypoxia mimetic.
The hypoxia model was established as follows: FaDu cells were
seeded at a density of 3×105 cells/well in 6-well
plates. Subsequently, the cells were treated with 0, 25, 50, 100,
200 or 400 µmol/l of CoCl2 for 6, 12 or 24 h, and
with 200 µmol/l CoCl2 for 0, 3, 6, 12 or 24 h at
37°C. Alterations in HIF-1α expression were evaluated by western
blot analysis.
Cell Counting kit-8 (CCK-8) assay
Cell viability was detected using the CCK-8 assay
(Beyotime Institute of Biotechnology). FaDu cells were seeded at a
density of 3×103 cells/well in 96-well plates and
incubated for 24 h, after which the cells were treated with 0, 25,
50, 100, 200 or 400 µmol/l CoCl2. CCK-8 assays
were performed after treatment with CoCl2 for 6, 12 or
24 h at 37°C. Subsequently, 10 µl CCK-8 solution was added
to each well, and the cells were incubated for 2 h at 37°C.
Absorbance was measured at 450 nm using a microplate reader (BioTek
Instruments, Inc.). The results were analyzed by SPSS 17.0 (SPSS,
Inc.) and the graph was generated using GraphPad Prism 5 (GraphPad
Software, Inc.).
Western blot analysis
The preparation of whole-cell protein lysates and
western blot analysis were performed as previously described
(23). Total proteins were
extracted in lysis buffer and protein concentrations were measured
using the protein bicinchoninic acid assay kit (Beyotime Institute
of Biotechnology). Protein lysates (40 µg) were separated by
10% SDS-PAGE and transferred to polyvinylidene difluoride membranes
(EMD Millipore). Subsequently, the membranes were blocked with 5%
non-fat milk in Tris-buffered saline containing 0.05% Tween-20 for
1 h at room temperature, and were incubated at 4°C overnight with
primary antibodies against fascin-1 (1:1,000), HIF-1α (1:500),
MMP-2 (1:1,000), MMP-9 (1:2,000), E-cadherin (1:6,0000), Vimentin
(1:1,000) and β-actin (1:20,000). After washing with Tris-buffered
saline containing 0.05% Tween-20, the membranes were incubated at
room temperature for 1 h with horseradish peroxidase-linked
secondary antibodies (1:10,000; goat anti-mouse IgG, cat. no.
ZB-5305; goat anti-rabbit IgG, cat. no. ZB-5301; ZSGB-BIO). The
protein bands were visualized using enhanced chemiluminescence
reagent (cat. no. WBKLS0500; EMD Millipore) and analyzed by ImageJ
software (version 1.37; National Institutes of Health).
Wound-healing assay
Cells were cultured in 6-well plates; once the cells
reached 100% confluence, the medium containing 10% FBS was replaced
with medium containing 1% FBS. A sterile 200-µl pipette tip
was used to generate a wound in the cell layer. Wound healing was
observed using an inverted light microscope after 0 and 24 h, and
images were captured using a Leica microscope image system (Leica
Microsystems, Inc.). The fold change of wound healing was
calculated using the following formula: Wound size at 24 h/wound
size at 0 h. Wound size was measured using ImageJ software
(version1.37; National Institutes of Health).
Transwell assays
Cell migration assays were performed using Transwell
chambers (Costar; Corning, Inc.). The upper chamber of the
Transwell system was filled with 250 µl serum-free DMEM/F-12
containing 1x105 cells. Medium containing 20% FBS was
added to the lower chamber as a chemoattractant, and the cells were
incubated for 24 h at 37°C. Migrating cells were fixed with 4%
formaldehyde at room temperature for 15 min and stained with 0.1%
crystal violet at room temperature for 25 min. The number of cells
that migrated onto the lower side of the membrane was determined
using a light microscope (magnification, x100). Cell invasion
assays were performed in the same manner as the cell migration
assays; however, the Transwell chambers were pre-coated with 21
µl Matrigel (BD Biosciences; 1/3 diluted in DMEM/F12) for 30
min at 37°C. Each experiment was performed in triplicate, and five
random microscopic fields per well were analyzed.
Statistical analysis
Analyses were performed using SPSS 17.0 statistical
analysis software (SPSS, Inc.). The difference in fascin-1
expression between the paracarcinoma and HSCC tissues was assessed
using Wilcoxon rank sum test. The correlation between fascin-1 and
clinicopathological parameters or HIF-1α was assessed using the
Spearman rank correlation coefficient test. The relationship
between fascin-1 and overall survival rate was assessed using the
Kaplan-Meier method and compared with the log-rank test. All
experiments were performed in triplicate. Data are presented as the
mean ± SD. The statistical significance of differences between two
groups was analyzed with Student's t-test. The statistical
significance of differences between multiple groups was analyzed
with one-way ANOVA followed by the least-significant difference
post-hoc test. All P-values were two sided, and P<0.05 was
considered to indicate a statistically significant difference.
Results
Expression of fascin-1 in HSCC
tissues
Immunohistochemistry was performed to validate
fascin-1 expression using a cohort of 96 patients with HSCC. The
results demonstrated that fascin-1 was mainly localized in the
cytoplasm and cell membrane of tumor cells (Fig. 1). Paracarcinoma tissues revealed
weak or partly moderate fascin-1 staining restricted to the basal
layers (Fig. 1D), whereas HSCC
tissues revealed variable distribution and intensity of fascin-1
staining, and fascin-1 expression at the invasive front of the
tumor was considerably stronger than in other regions (Fig. 1H). Fascin-1 positive expression in
HSCC tissues was detected in 88.54% of cases, which was
significantly higher than in paracarcinoma tissues (Table I).
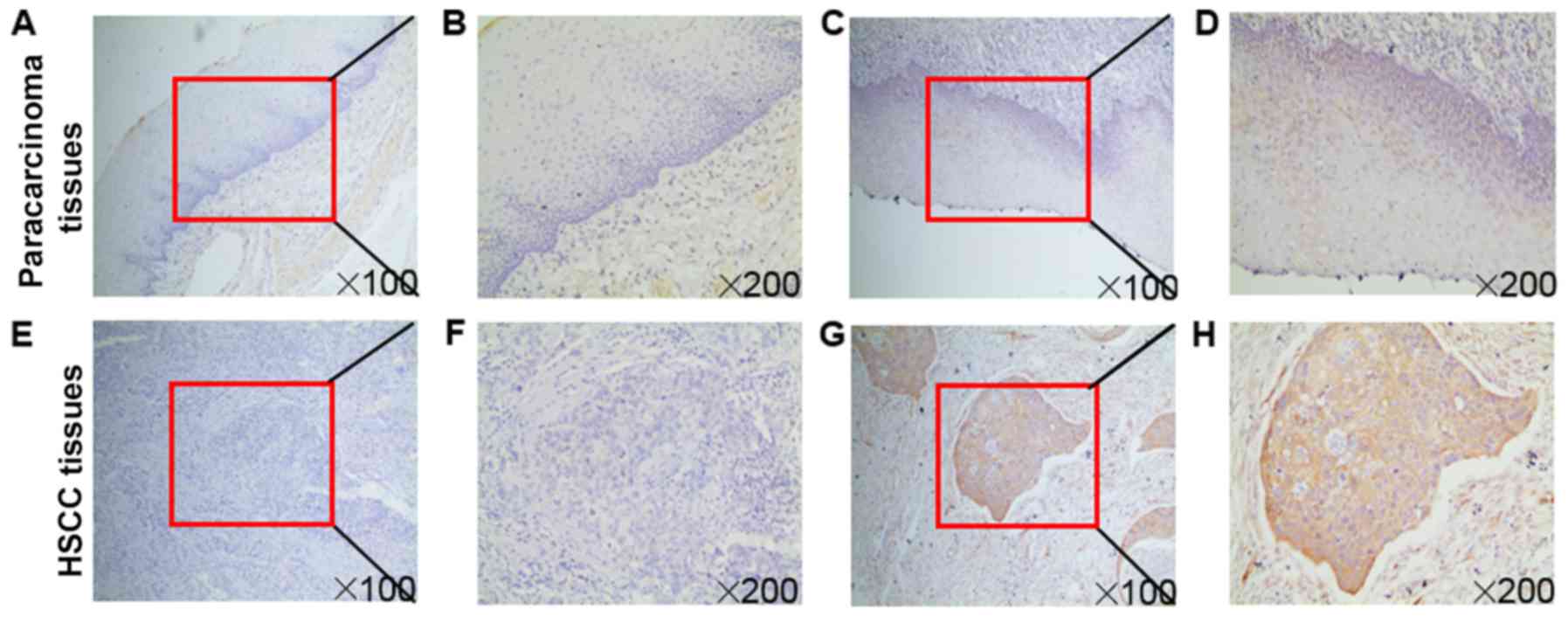 | Figure 1Immunochemical analysis of fascin-1
expression in HSCC. Fascin-1 protein expression in (A-D)
paracarcinoma and (E-H) HSCC tissues. Higher magnification images
of the boxed areas in (A), (C), (E) and (G) are shown in (B), (D),
(F) and (H), respectively. (A, B, E and F) Negative expression of
fascin-1 in paracarcinoma and HSCC tissues. (C, D, G and H)
Positive expression of fascin-1 in paracarcinoma and HSCC tissues.
(D) In the paracarcinoma tissues, fascin-1 revealed weak or partly
moderate staining restricted to the basal layers. (H) In HSCC
tissues, fascin-1 expression at the invasive front of the tumor was
markedly stronger than in the other regions. Magnifications, x100
and x200. HSCC, hypopharyngeal squamous cell carcinoma. |
 | Table IFascin-1 expression in paracarcinoma
tissues and HSCC tissues. |
Table I
Fascin-1 expression in paracarcinoma
tissues and HSCC tissues.
| Tissue | Number | Fascin-1 expression
| P-value |
|---|
| Negative (%) | Positive (%) |
|---|
| Paracarcinoma | 96 | 72 (75) | 24 (25) | |
| HSCC | 96 | 11 (11.46) | 85 (88.54) | <0.001 |
The Spearman rank correlation coefficient test was
used to determine the relationships between fascin-1 expression and
the clinicopathological characteristics of patients with HSCC. As
shown in Table II, fascin-1
expression was significantly correlated with lymph node metastasis
(P<0.001) and the pathological tumor-node-metastasis (TNM) stage
(P<0.001) (24). These data
suggested that fascin-1 may serve a vital role in the invasion and
metastasis of HSCC. Furthermore, the relationship between fascin-1
and survival rate was analyzed; the results revealed that fascin-1
expression was not associated with 3-year survival (P=0.365;
Fig. S1). Therefore, fascin-1 may
not serve an important role in HSCC prognosis.
| Table IICorrelation between fascin-1
expression and clinicopathological parameters of patients with
hypopharyngeal squamous cell carcinoma. |
Table II
Correlation between fascin-1
expression and clinicopathological parameters of patients with
hypopharyngeal squamous cell carcinoma.
| Parameter | Number | Fascin-1 expression
| rs | P-value |
|---|
| Negative (%) | Positive (%) |
|---|
| Sex | | | | | |
| Male | 91 | 10 | 81 | | |
| Female | 5 | 1 | 4 | −0.063 | 0.543 |
| Age (years) | | | | | |
| <60 | 57 | 5 | 52 | | |
| ≥60 | 39 | 6 | 33 | −0.102 | 0.323 |
|
Differentiation | | | | | |
| Well and
moderate | 63 | 6 | 57 | | |
| P oor | 33 | 5 | 28 | −0.084 | 0.416 |
| T
classification | | | | | |
| T1-T2 | 20 | 4 | 16 | | |
| T3-T4 | 76 | 7 | 69 | 0.138 | 0.181 |
| TNM stage | | | | | |
| I-II | 5 | 3 | 2 | | |
| III-IV | 91 | 8 | 83 | 0.357 | <0.001 |
| Lymph node
metastasis | | | | | |
| Negative | 16 | 6 | 10 | | |
| Positive | 80 | 5 | 75 | 0.366 | <0.001 |
| Lung
metastasis | | | | | |
| Negative | 70 | 9 | 61 | | |
| Positive | 26 | 2 | 24 | 0.072 | 0.485 |
Fascin-1 promotes the invasion and
migration of FaDu cells
To determine the effects of fascin-1 on FaDu cells,
fascin-1 knockdown and overexpression were induced to reveal
whether fascin-1 could modulate the efficiency of cell migration
and invasion. Western blot analysis was used to determine the
transfection efficiency of fascin-1 siRNA and expression plasmids
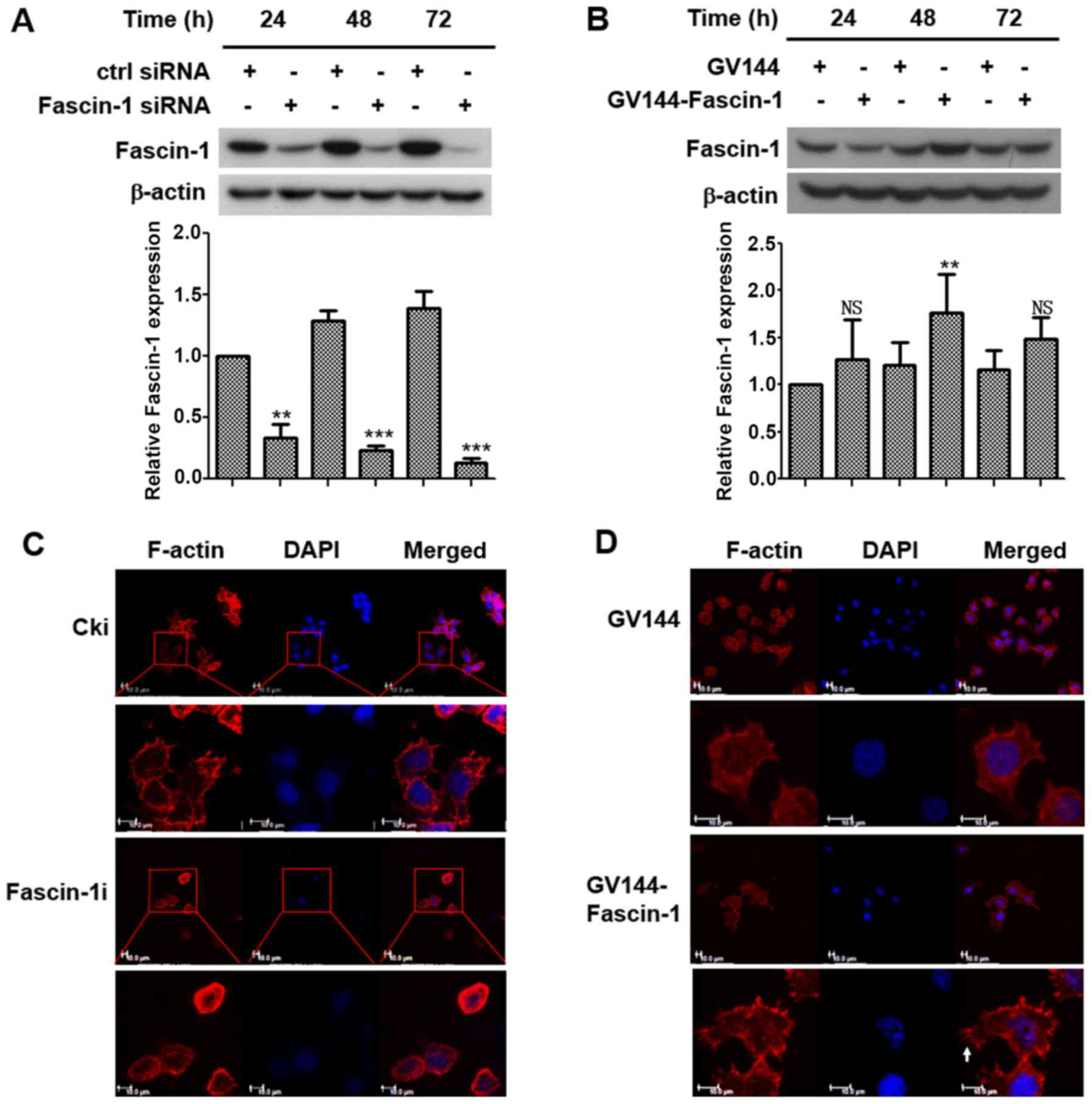
in FaDu cells. The results demonstrated that fascin-1 expression
was decreased with prolonged time post-transfection with fascin-1
siRNA (Fig. 2A), and the increase
in fascin-1 expression peaked at 48 h post-transfection with
fascin-1 expression plasmids in FaDu cells (Fig. 2B). Therefore, cells were analysed
48 h post-transfection in subsequent experiments.
Fluorescently labeled phalloidin staining of F-actin
was performed to observe alterations in cell morphology. The
results demonstrated that the control cells exhibited a polarized
shape with membrane protrusions; however, the fascin-1-silenced
cells exhibited a more rounded morphology with fewer membrane
protrusions (Fig. 2C). Cells in
which fascin-1 was overexpressed exhibited a more polarized shape
with more membrane protrusions (Fig.
2D). Filopodia were some of the membrane protrusions observed.
These results suggested that fascin-1 may lead to changes in cell
morphology, which could increase the motility and migratory ability
of FaDu cells.
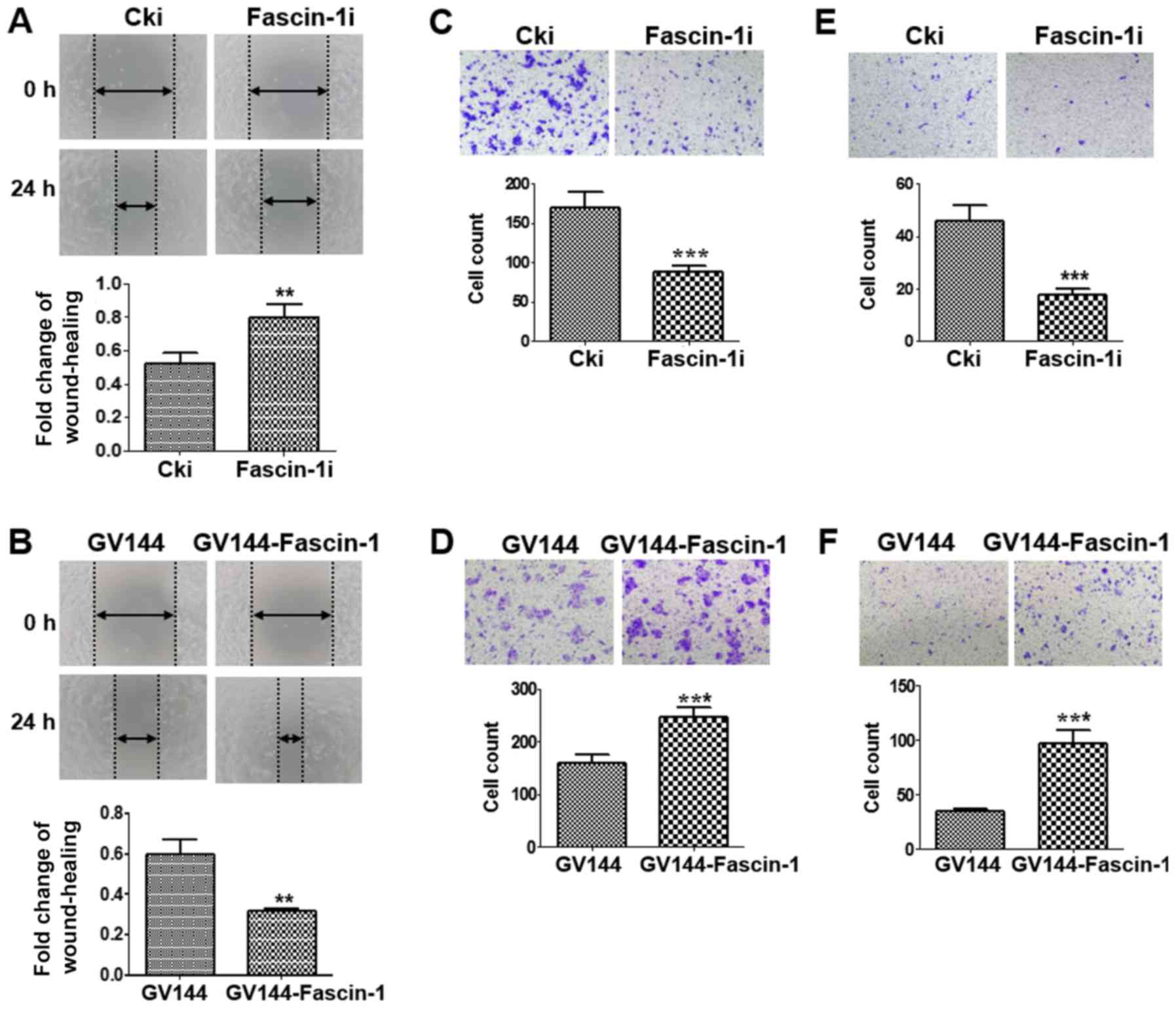
Wound-healing and Transwell assays were performed to
further determine the effects of fascin-1 on cell migration and
invasion. A wound-healing assay revealed that fascin-1-silenced
cells migrated more slowly than the control cells (Fig. 3A), whereas fascin-1-overexpressed
cells migrated more quickly than the control cells (Fig. 3B). A similar trend was observed in
the migration assay. The results confirmed that the number of
migrated cells was reduced in the fascin-1-silenced group compared
with in the control group (Fig.
3C), whereas the number of migrated cells was higher in the
fascin-1-overexpressed group than in the control group (Fig. 3D). These results indicated that
fascin-1 promoted the migration of FaDu cells. The results from the
invasion assay revealed a reduction in the number of invasive cells
in the fascin-1-silenced cells compared with in the control group
(Fig. 3E); however, the number of
invasive cells was increased in the fascin-1-overexpressed group
compared with in the control group (Fig. 3F). Therefore, fascin-1 may promote
the invasion of FaDu cells.
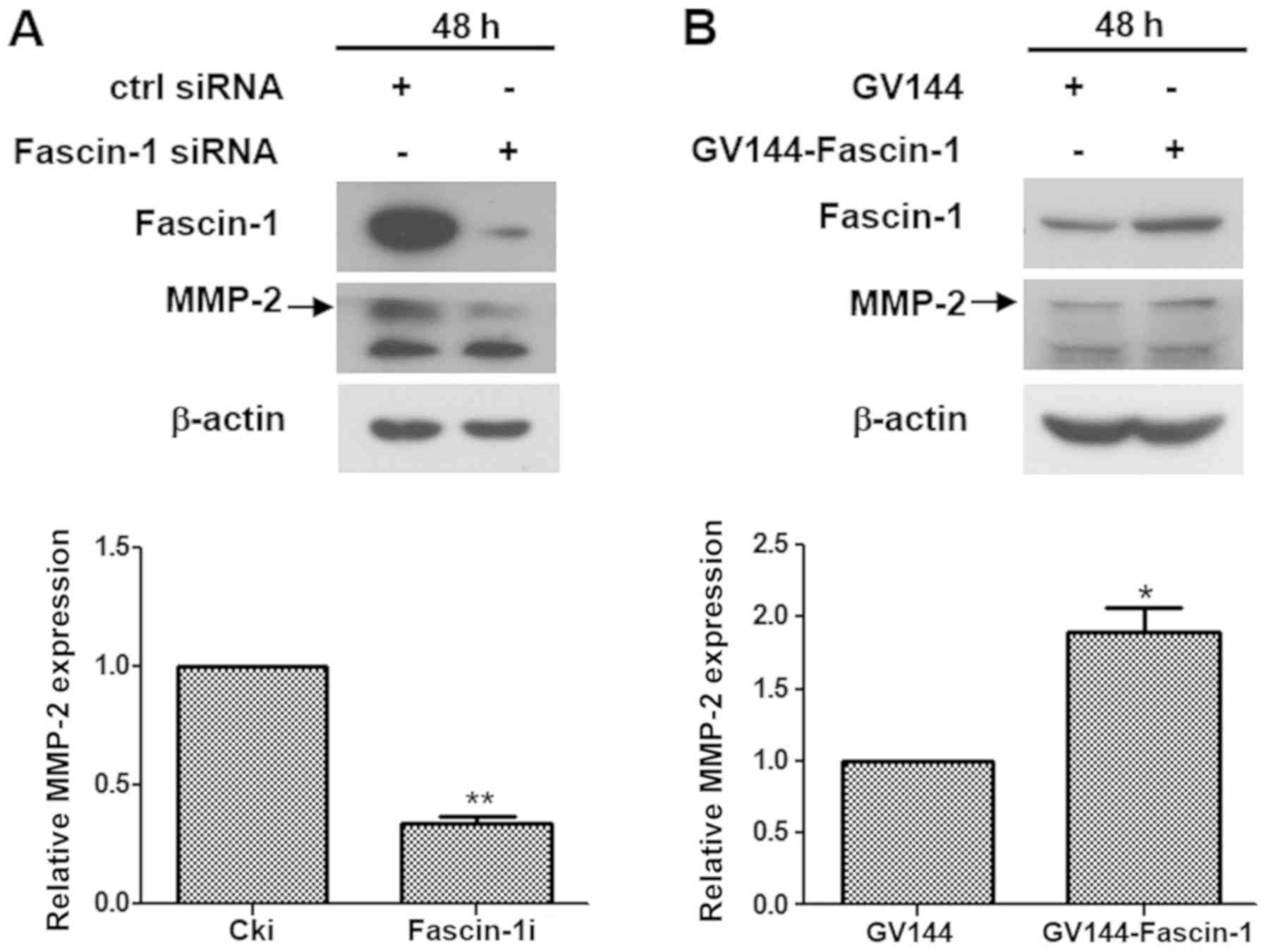
The expression of proteins associated with invasion,
metastasis and epithelial-mesenchymal transition, including MMP-2,
MMP-9, E-cadherin and vimentin were examined by western blotting to
determine whether fascin-1 influenced cell invasion and migration
at the molecular level. The results revealed that fascin-1
knockdown downregulated MMP-2 expression in FaDu cells (Fig. 4A), whereas fascin-1 overexpression
upregulated MMP-2 expression in FaDu cells (Fig. 4B). However, no significant effect
was observed on the other proteins(Fig. S2). These findings suggested that
fascin-1 promoted invasion and migration of FaDu cells by
upregulating MMP-2.
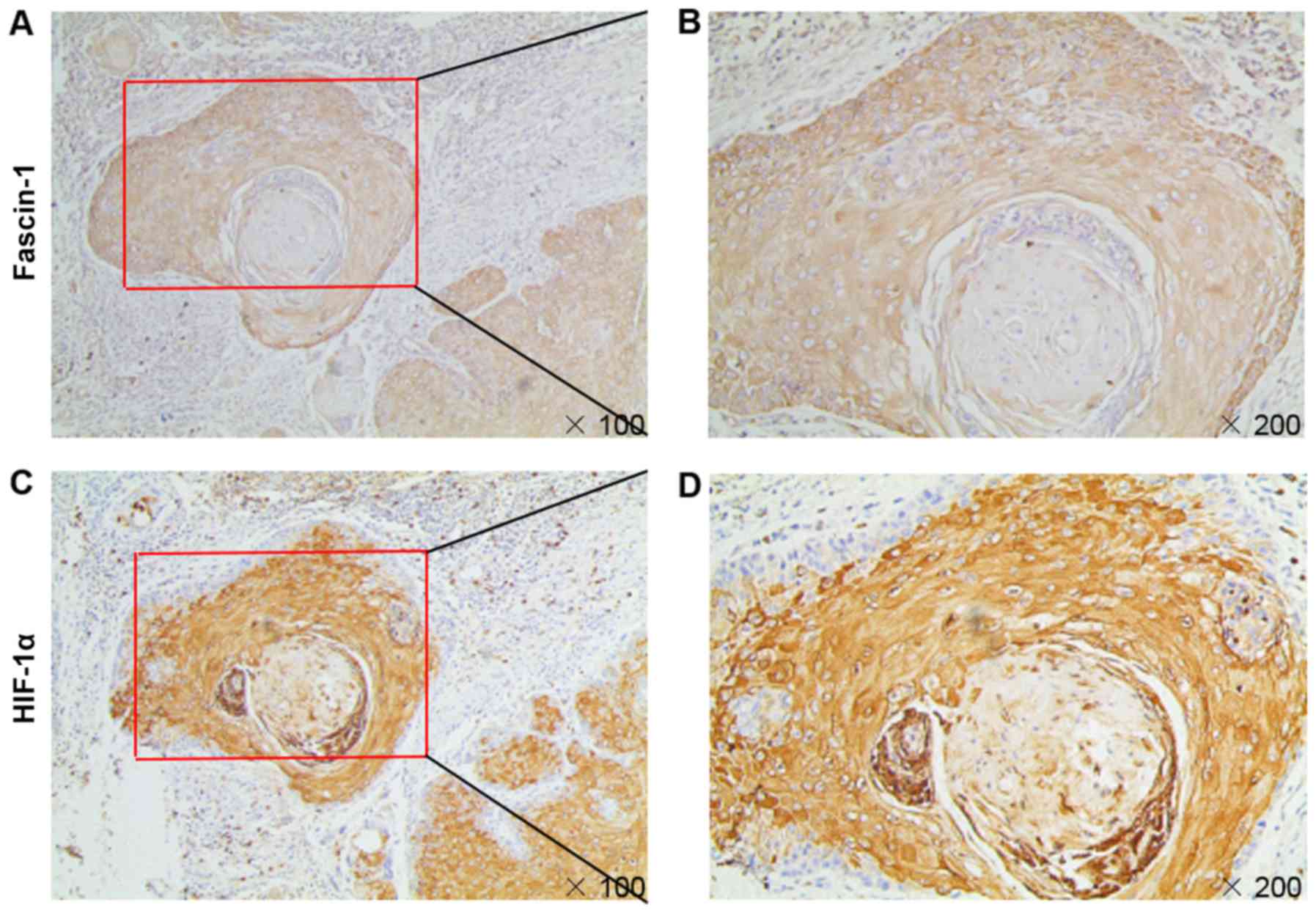
Correlation between HIF-1α and fascin-1
expression in HSCC tissues
The results of immunohistochemical analysis
confirmed that fascin-1 expression colocalized with HIF-1α
expression in consecutive sections of HSCC tissues (Fig. 5). Fascin-1 expression was
significantly correlated with HIF-1α expression in HSCC tissues
(Table III). These results
suggested that HIF-1α may be involved in fascin-1 overexpression in
HSCC tissues.
| Table IIICorrelation between HIF-1α and
fascin-1 expression in hypopharyngeal squamous cell carcinoma. |
Table III
Correlation between HIF-1α and
fascin-1 expression in hypopharyngeal squamous cell carcinoma.
| HIF-1α
expression | Fascin-1 expression
| rs | P-value |
|---|
| Negative (%) | Positive (%) |
|---|
| Negative | 7 | 20 | | |
| Positive | 4 | 65 | 0.284 | 0.005 |
Upregulation of fascin-1 by HIF-1α under
hypoxia in FaDu cells
Previous results indicated that HIF-1α may be
involved in fascin-1 overexpression in HSCC tissues. Therefore, a
hypoxia model was further established by CoCl2
treatment, which can upregulate HIF-1α expression. The results
demonstrated that the expression levels of HIF-1α were increased in
concentration-dependent and time-dependent manners after FaDu cells
were treated with CoCl2 (Fig. 6A and B). CCK-8 assay revealed that
cell viability was significantly decreased following treatment with
400 µmol/l CoCl2 (Fig. 6C). Therefore, 200 µmol/l
CoCl2 was selected as the experimental concentration in
subsequent studies.
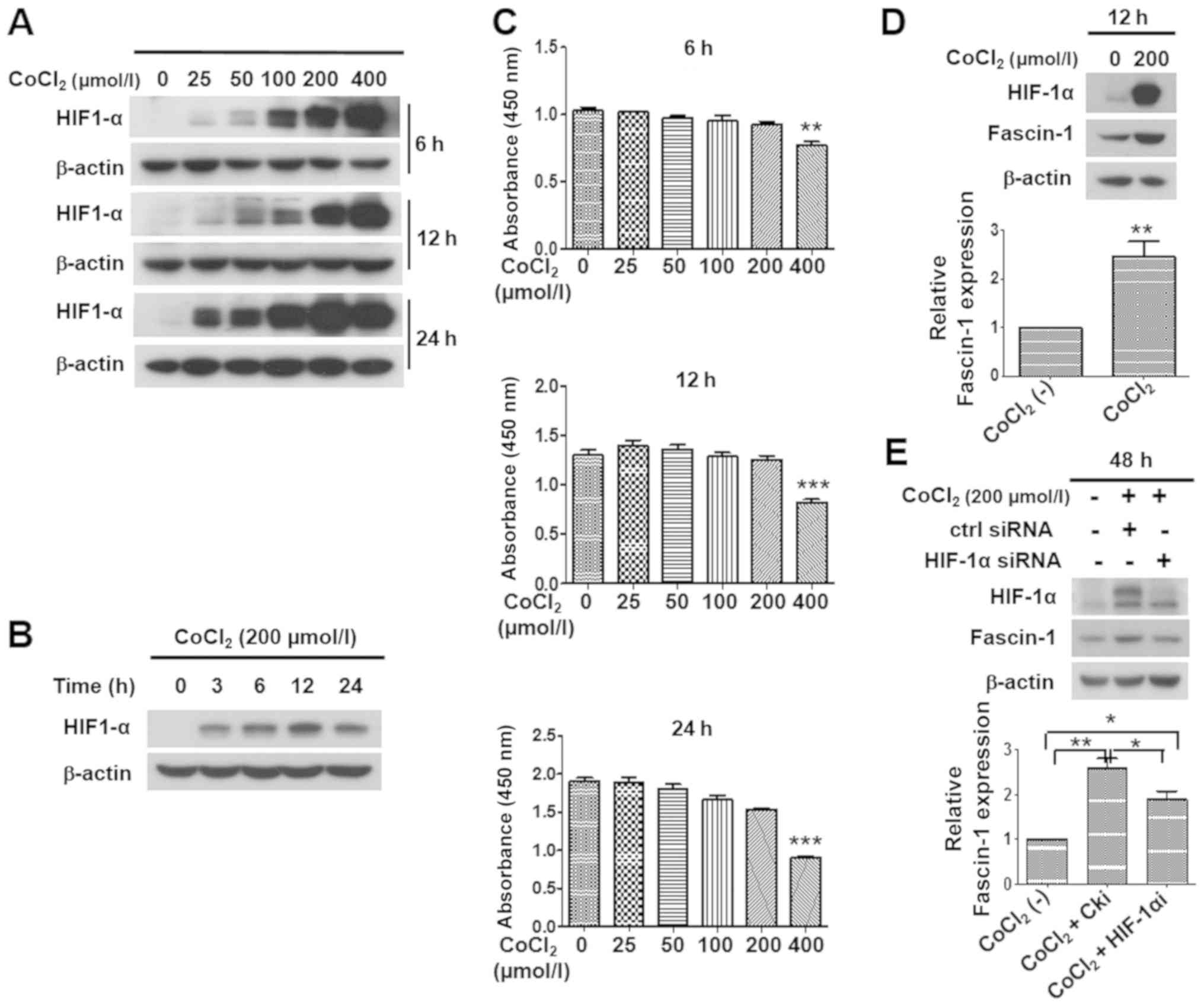 | Figure 6Effects of hypoxia induced by
CoCl2 on HIF-1α expression, and the upregulation of
fascin-1 expression by HIF-1α under hypoxia in FaDu cells. (A)
Expression of HIF-1α was observed following treatment with 0, 25,
50, 100, 200 or 400 µmol/l of CoCl2 for 6, 12 or
24 h in FaDu cells. Western blotting revealed that HIF-1α
expression was upregulated in a concentration-dependent manner.
β-actin was used as an internal reference. (B) Expression of HIF-1α
in FaDu cells was observed following treatment with 200
µmol/l CoCl2 for 0, 3, 6, 12 or 24 h. Western
blotting revealed that CoCl2 induced HIF-1α expression
in a time-dependent manner, and HIF-1α expression peaked at 12 h in
FaDu cells. β-actin was used as an internal reference. (C)
Viability of FaDu cells treated with 0, 25, 50, 100, 200 or 400
µmol/l CoCl2 for 6, 12 or 24 h, as determined by
Cell Counting kit-8 assay. No significant difference was detected
in cell viability at concentrations ranging between 0 and 200
µmol/l. However, following treatment with 400 µmol/l
CoCl2, cell viability was significantly decreased.
**P<0.01, ***P<0.001 vs. 0
µmol/l. Western blotting and semi-quantitative analysis
revealed that (D) compared with under normoxia, hypoxia induced by
treatment with 200 µmol/l CoCl2 for 12 h
upregulated HIF-1α and fascin-1 expression in FaDu cells. Student's
t-test was used for analysis. (E) HIF-1α knockdown reduced the
expression of fascin-1 under hypoxia post-transfection with HIF-1α
siRNA for 48 h in FaDu cells. One-way ANOVA was used for analysis.
β-actin was used as an internal reference. Protein expression was
semi-quantified using ImageJ software, and the CoCl2 (-)
group was set at 1. Data are presented as the mean ± SD.
*P<0.05, **P<0.01. CoCl2,
cobalt chloride; HIF-1α, hypoxia inducible factor-1α; siRNA, small
interfering RNA. |
To identify whether hypoxia promoted the expression
of fascin-1 in FaDu cells, the cells were exposed to 200
µmol/l CoCl2 for 12 h. Subsequently, western blot
analysis was performed. The results demonstrated that, with the
upregulation of HIF-1α, fascin-1 expression was considerably
increased in FaDu cells under CoCl2-induced hypoxia
compared with under normoxia (Fig.
6D). These data suggested that hypoxia could upregulate
fascin-1 expression in FaDu cells.
HIF-1α knockdown was performed to examine the role
of HIF-1α in regulating the expression of fascin-1 under hypoxia in
FaDu cells. Since HIF-1α protein undergoes rapid degradation under
normoxic conditions, the expression levels of HIF-1α were hardly
detected in FaDu cells under normoxia. Therefore, the transfection
efficiency of HIF-1α siRNA could not be analyzed under normoxic
conditions. Conversely, hypoxia promotes the stability of HIF-1α
via preventing its proteasomal degradation. Therefore, cells that
underwent CoCl2-induced hypoxia for 48 h were
transfected with HIF-1α siRNA. As shown in Fig. 6E, the expression of HIF-1α under
CoCl2-induced hypoxia was markedly reduced 48 h
post-transfection with HIF-1α siRNA. Further results demonstrated
that HIF-1α knockdown significantly downregulated the expression of
fascin-1 under CoCl2-induced hypoxia (Fig. 6E). These data suggested that
hypoxia upregulated the expression of fascin-1 through HIF-1α, and
HIF-1α may serve a critical role in fascin-1 overexpression under
hypoxia in FaDu cells.
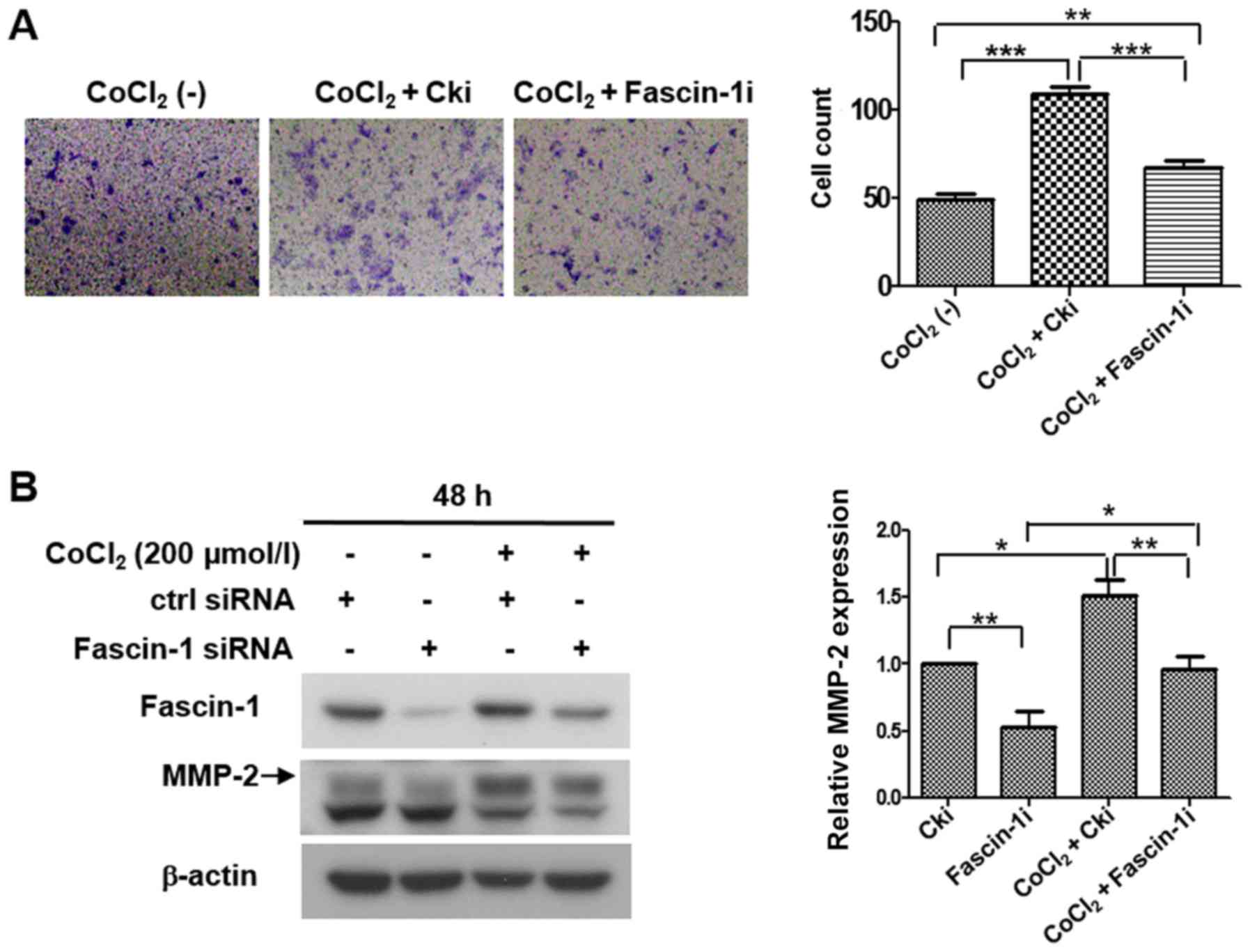
Hypoxia promotes invasion and migration
of FaDu cells by upregulating fascin-1 expression
An invasion assay and western blot analysis were
performed to further determine whether hypoxia promoted cell
invasion and migration through fascin-1. The invasion assay
revealed that the number of invasive cells was significantly
increased under hypoxia compared with under normoxia, whereas the
number of invasive cells under hypoxia was reduced when fascin-1
was silenced (Fig. 7A). Western
blot analysis revealed that hypoxia markedly increased the
expression levels of MMP-2. However, fascin-1 knockdown partially
reversed MMP-2 expression promoted by hypoxia in FaDu cells
(Fig. 7B). These data suggested
that hypoxia may promote the invasion and migration of FaDu cells
by increasing fascin-1 expression.
Discussion
Tumor invasion and metastasis are complex processes.
The acquisition of spindle-like morphology by reorganization of the
cytoskeleton to enhance motility is regarded as an important step
in tumor invasion and metastasis (25,26).
Another key step in tumor invasion and metastasis is degradation of
the extracellular matrix; this degradation is associated with MMPs,
such as MMP-2 and MMP-9 (27,28).
Fascin-1 is a cytoskeletal protein, which is
involved in regulating the cytoskeleton structure and formation of
plasma membrane protrusions by crosslinking actin microfilaments
into tight, parallel bundles (29). Alam et al (14) reported that fascin overexpression
is significantly correlated with tumor stage, lymph node metastasis
and reduced differentiation in oral squamous cell carcinoma.
Papaspyrou et al (15)
reported the fascin is overexpressed in tumor tissues and is
associated with lymph node metastases; furthermore, it was
demonstrated that this overexpression is significantly correlated
with lymph node metastasis in head and neck squamous cell
carcinoma. Zhao et al (30)
reported that fascin-1 overexpression is significantly correlated
with age groups, clinical stages and lymph node metastases in lung
cancer. Consistent with these previous studies, the present data
indicated that fascin-1 was overexpressed in HSCC tissues and was
significantly correlated with lymph node metastasis and
pathological TNM stage. Therefore, fascin-1 overexpression may
promote invasion and metastasis in HSCC. In addition, according to
epidemiological investigations, sex is significantly associated
with the incidence of hypopharyngeal carcinoma worldwide (31,32).
Consistently, the 96 patients with HSCC enrolled in this study
exhibited a skew with regards to sex (91 male and 5 female).
However, there was no significant correlation between fascin-1
expression and sex. To obtain fairly accurate statistical data,
larger samples containing more female patients are required for
further study.
Previous studies have indicated that fascin-1
promotes the invasion and migration of carcinoma in vitro.
Alam et al (14) reported
that fascin overexpression alters cell morphology, and increases
cell migration, invasion and MMP-2 activity in oral squamous cell
carcinoma cells. Zhao et al (33) demonstrated that fascin-1 knockdown
suppresses cell invasion and migration in non-small cell lung
cancer. In the present study, a HSCC cell line was used to
determine the specific function of fascin-1 in invasion and
migration of HSCC. At present, the FaDu cell line is the only
available hypopharyngeal carcinoma cell line; therefore, only this
cell line was used to conduct subsequent experiments, which may be
a limitation to this study. To compensate for this, we are
currently working on the construction of novel hypopharyngeal
cancer cell lines using clinical specimen resources; however, this
is yet to be successful. In future studies, we will consider using
primary cells to conduct cell experiments. In the present study,
consistent with previous studies (14,33),
the in vitro analyses revealed that fascin-1 led to cells
with a more polarized shape and more membrane protrusions, such as
filopodia, which increased the motility of FaDu cells. Further
functional studies revealed that fascin-1 significantly promoted
the migration and invasion of FaDu cells. Molecular mechanism
studies indicated that fascin-1 upregulated the expression of
MMP-2, which may be associated with the invasion and migration of
FaDu cells. In summary, these data suggested that fascin-1 promoted
cell invasion and migration in HSCC.
The present results also identified colocalization
of HIF-1α and fascin-1 expression, and the correlation between
HIF-1α and fascin-1 expression was significant in HSCC specimens.
Therefore, an underlying regulatory mechanism may exist between
fascin-1 and HIF-1α. Zhao et al (22) also reported that fascin expression
is colocalized with HIF-1α and is significantly correlated with
HIF-1α expression in pancreatic ductal adenocarcinoma tissues.
Therefore, the underlying regulatory mechanism between HIF-1α and
fascin-1 in HSCC should be established.
Hypoxia and CoCl2 increase HIF-1α
expression; CoCl2 can be used to stabilize HIF-1α and
mimic true hypoxic conditions, which activate HIF-1α (34). In the present study, the
CoCl2-induced chemical hypoxia model was established.
Further studies revealed that CoCl2-induced hypoxia
increased the expression of fascin-1. This result suggested that
hypoxia increased fascin-1 expression in FaDu cells. A previous
study demonstrated that HIF-1α knockdown markedly downregulates the
expression of fascin under true hypoxia (1% O2), and
fascin is a direct target gene of HIF-1α (22). In line with this previous report,
our further studies revealed that HIF-1α knockdown downregulated
the expression of fascin-1 under CoCl2-induced hypoxia
in FaDu cells. These results suggested that hypoxia elevated the
expression of fascin-1 though HIF-1α in FaDu cells, and HIF-1α, as
a transcription factor, may upregulate fascin-1 expression.
Zhao et al (22) reported that hypoxia promotes the
invasion and migration of pancreatic ductal adenocarcinoma by
inducing fascin overexpression. The present invasion assay data
revealed that CoCl2-induced hypoxia significantly
promoted the invasion of FaDu cells, whereas fascin-1 knockdown
partially decreased the invasion of FaDu cells under hypoxia.
Furthermore, mechanistic studies indicated that fascin-1 knockdown
partially reversed the MMP-2 overexpression promoted by HIF-1α
under CoCl2-induced hypoxia in FaDu cells. These results
suggested that fascin-1 was involved in the HIF-1α-dependent
invasion and migration of HSCC, and fascin-1 may serve a vital role
in promoting cell invasion and migration under a hypoxic
microenvironment in HSCC.
In conclusion, these results indicated that fascin-1
was overexpressed in HSCC, and was significantly associated with
the invasion and metastasis of HSCC. Furthermore, the present data
suggested that HIF-1α may promote the invasion and migration of
HSCC cells by upregulating fascin-1 expression. Several
small-molecule inhibitors targeting fascin-1 have been reported to
prevent tumor metastasis (35).
Therefore, Fascin-1 may provide a potential target for the
treatment of invasion and metastasis in HSCC.
Supplementary Data
Acknowledgments
Not applicable.
Funding
The present study was supported by grants from the
National Natural Science Foundation of China (grant no. 81702679),
the Natural Science Foundation of Shandong province (grant no.
ZR2017BH060) and the Key Project of Shandong Provincial Programs
for Research and Development (grant no. 2018GSF118225).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
MB and XL contributed equally to this article. WX
designed the experiments. MB wrote the paper and performed the
experiments. XL performed the experiments and revised the paper. MB
and XL were responsible for collecting all data and analyzed the
data in the study. WX checked the manuscript. All authors have read
and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the ethics
committee of Shandong University and written informed consent was
obtained from patients prior to sample collection.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Gooi Z, Fakhry C, Goldenberg D, Richmon J
and Kiess AP; Education Committee of the American Head and Neck
Society (AHNS): AHNS Series: Do you know your guidelines?
Principles of radiation therapy for head and neck cancer: A review
of the National Comprehensive Cancer Network guidelines. Head Neck.
38:987–992. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lewis CM, Hessel AC, Roberts DB, Guo YZ,
Holsinger FC, Ginsberg LE, El-Naggar AK and Weber RS: Prereferral
head and neck cancer treatment: Compliance with national
comprehensive cancer network treatment guidelines. Arch Otolaryngol
Head Neck Surg. 136:1205–1211. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wycliffe ND, Grover RS, Kim PD and
Simental A Jr: Hypopharyngeal cancer. Top Magn Reson Imaging.
18:243–258. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ge N, Lin HX, Xiao XS, Guo L, Xu HM, Wang
X, Jin T, Cai XY, Liang Y, Hu WH, et al: Prognostic significance of
Oct4 and Sox2 expression in hypopharyngeal squamous cell carcinoma.
J Transl Med. 8:942010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kotwall C, Sako K, Razack MS, Rao U,
Bakamjian V and Shedd DP: Metastatic patterns in squamous cell
cancer of the head and neck. Am J Surg. 154:439–442. 1987.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hui AB, Bruce JP, Alajez NM, Shi W, Yue S,
Perez-Ordonez B, Xu W, O'Sullivan B, Waldron J, Cummings B, et al:
Significance of dysregulated metadherin and microRNA-375 in head
and neck cancer. Clin Cancer Res. 17:7539–7550. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Loberg RD, Bradley DA, Tomlins SA,
Chinnaiyan AM and Pienta KJ: The lethal phenotype of cancer: The
molecular basis of death due to malignancy. CA Cancer J Clin.
57:225–241. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Adams JC: Formation of stable microspikes
containing actin and the 55 kDa actin bundling protein, fascin, is
a consequence of cell adhesion to thrombospondin-1: Implications
for the anti-adhesive activities of thrombospondin-1. J Cell Sci.
108:1977–1990. 1995.PubMed/NCBI
|
|
9
|
Adams JC: Fascin protrusions in cell
interactions. Trends Cardiovasc Med. 14:221–226. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Duh FM, Latif F, Weng Y, Geil L, Modi W,
Stackhouse T, Matsumura F, Duan DR, Linehan WM, Lerman MI, et al:
cDNA cloning and expression of the human homolog of the sea urchin
fascin and Drosophila singed genes which encodes an actin-bundling
protein. DNA Cell Biol. 13:821–827. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
De Arcangelis A, Georges-Labouesse E and
Adams JC: Expression of fascin-1, the gene encoding the
actin-bundling protein fascin-1, during mouse embryogenesis. Gene
Expr Patterns. 4:637–643. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang FR, Tao LH, Shen ZY, Lv Z, Xu LY and
Li EM: Fascin expression in human embryonic, fetal, and normal
adult tissue. J Histochem Cytochem. 56:193–199. 2008. View Article : Google Scholar
|
|
13
|
Zou J, Yang H, Chen F, Zhao H, Lin P,
Zhang J, Ye H, Wang L and Liu S: Prognostic significance of
fascin-1 and E-cadherin expression in laryngeal squamous cell
carcinoma. Eur J Cancer Prev. 19:11–17. 2010. View Article : Google Scholar
|
|
14
|
Alam H, Bhate AV, Gangadaran P, Sawant SS,
Salot S, Sehgal L, Dange PP, Chaukar DA, D'cruz AK, Kannanl S, et
al: Fascin overexpression promotes neoplastic progression in oral
squamous cell carcinoma. BMC Cancer. 12:322012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Papaspyrou K, Brochhausen C, Schmidtmann
I, Fruth K, Gouveris H, Kirckpatrick J, Mann W and Brieger J:
Fascin upregulation in primary head and neck squamous cell
carcinoma is associated with lymphatic metastasis. Oncol Lett.
7:2041–2046. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shay JE and Celeste Simon M:
Hypoxia-inducible factors: Crosstalk between inflammation and
metabolism. Semin Cell Dev Biol. 23:389–394. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Subarsky P and Hill RP: The hypoxic tumour
microenvironment and metastatic progression. Clin Exp Metastasis.
20:237–250. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gilkes DM, Semenza GL and Wirtz D: Hypoxia
and the extracellular matrix: Drivers of tumour metastasis. Nat Rev
Cancer. 14:430–439. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Semenza GL: Hydroxylation of HIF-1: Oxygen
sensing at the molecular level. Physiology (Bethesda). 19:176–182.
2004.
|
|
20
|
Brahimi-Horn MC, Bellot G and Pouysségur
J: Hypoxia and energetic tumour metabolism. Curr Opin Genet Dev.
21:67–72. 2011. View Article : Google Scholar
|
|
21
|
Zhu G, Peng F, Gong W, She L, Wei M, Tan
H, Chen C, Zhang D, Li G, Huang D, et al: Hypoxia promotes
migration/invasion and glycolysis in head and neck squamous cell
carcinoma via an HIF-1α-MTDH loop. Oncol Rep. 38:2893–2900. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhao X, Gao S, Ren H, Sun W, Zhang H, Sun
J, Yang S and Hao J: Hypoxia-inducible factor-1 promotes pancreatic
ductal adenocar-cinoma invasion and metastasis by activating
transcription of the actin-bundling protein fascin. Cancer Res.
74:2455–2464. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu X, Yue P, Zhou Z, Khuri FR and Sun SY:
Death receptor regulation and celecoxib-induced apoptosis in human
lung cancer cells. J Natl Cancer Inst. 96:1769–1780. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Edge SB, Byrd DR, Compton CC, Fritz AG,
Greene FL and Trotti A: AJCC Cancer Staging Manual. 7th edition.
Springer; New York, NY: 2010
|
|
25
|
Haynes J, Srivastava J, Madson N, Wittmann
T and Barber DL: Dynamic actin remodeling during
epithelial-mesenchymal transition depends on increased moesin
expression. Mol Biol Cell. 22:4750–4764. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu SC, Jen YM, Jiang SS, Chang JL, Hsiung
CA, Wang CH and Juang JL: G(alpha)12-mediated pathway promotes
invasiveness of nasopharyngeal carcinoma by modulating actin
cytoskeleton reorganization. Cancer Res. 69:6122–6130. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sato H, Kida Y, Mai M, Endo Y, Sasaki T,
Tanaka J and Seiki M: Expression of genes encoding type IV
collagen-degrading metalloproteinases and tissue inhibitors of
metalloproteinases in various human tumor cells. Oncogene. 7:77–83.
1992.PubMed/NCBI
|
|
28
|
Okazaki I and Inagaki Y: Novel strategies
for hepatocellular carcinoma based on MMPs science. Anticancer
Agents Med Chem. 12:753–763. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Edwards RA and Bryan J: Fascins, a family
of actin bundling proteins. Cell Motil Cytoskeleton. 32:1–9. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhao W, Gao J, Wu J, Liu QH, Wang ZG, Li
HL and Xing LH: Expression of Fascin-1 on human lung cancer and
paracarcinoma tissue and its relation to clinicopathological
characteristics in patients with lung cancer. Onco Targets Ther.
8:2571–2576. 2015.PubMed/NCBI
|
|
31
|
Zhang ZM, Tang PZ, Xu ZG, Li QH, Hu YH, Xu
GZ, Gao L and Tu GY: Long-term results of different treatment
modalities in 464 hypopharyngeal squamous-cell carcinoma patients.
Zhonghua Zhong Liu Za Zhi. 27:48–51. 2005.In Chinese. PubMed/NCBI
|
|
32
|
Hoffman HT, Karnell LH, Shah JP, Ariyan S,
Brown GS, Fee WE, Glass AG, Goepfert H, Ossoff RH and Fremgen AM:
Hypopharyngeal cancer patient care evaluation. Laryngoscope.
107:1005–1017. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhao D, Zhang T, Hou XM and Ling XL:
Knockdown of fascin-1 expression suppresses cell migration and
invasion of non-small cell lung cancer by regulating the MAPK
pathway. Biochem Biophys Res Commun. 497:694–699. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Huang BW, Miyazawa M and Tsuji Y: Distinct
regulatory mechanisms of the human ferritin gene by hypoxia and
hypoxia mimetic cobalt chloride at the transcriptional and
post-transcriptional levels. Cell Signal. 26:2702–2709. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chen L, Yang S, Jakoncic J, Zhang JJ and
Huang XY: Migrastatin analogues target fascin to block tumour
metastasis. Nature. 464:1062–1066. 2010. View Article : Google Scholar : PubMed/NCBI
|