1. Introduction
The gastric epithelium is continually renewed over a
lifetime, and is maintained through the proliferation and
differentiation of pluripotent stem cells from the isthmus of the
gastric gland (1). Stem cells
generate precursors that migrate to the gastric lumen and, in turn,
generate parietal, gastric zymogenic and foveolar cells. Parietal
cells produce hydrochloric acid, while gastric zymogenic cells have
a half-life of ~6 months and synthesize trefoil factor 2 and mucin
6 (1-3). Foveolar cells or surface mucous cells
(SMCs), whose half-life is 2-3 days, produce mucous granules, mucin
5AC, gastrokine 1 (GKN1) and trefoil factor 1 (1-3) and
play an important role in the restitution of the gastric mucosa in
the event of Helicobacter pylori infection (4). The integrity and continuity of the
gastric epithelium are rapidly restored after damage, prior to cell
proliferation (5). Epithelial
restitution is achieved through the migration of epithelial cells
from the adjacent area or the cell stratum below the surface cells
in the injured area. Epithelial cell restitution in the stomach of
mammals takes place in minutes (5).
GKN1 is a protein secreted by SMCs of the gastric
antrum and fundus (6), which
contributes to maintaining gastric homeostasis, inhibits
inflammation and acts as a tumor suppressor (7-16).
The expression of GKN1 decreases due to H. pylori infection,
inflammation or atrophy, and is absent in gastric cancer (17-25).
Although methylation of the GKN1 promoter (20), modification of histones (26) and proteasomal degradation of GKN1
in some cases (27) have been
reported, the mechanisms that cause a decrease in GKN1 levels or
its absence entirely have not been completely described. A total of
~10% of gastric tumors contain the Epstein-Barr virus (EBV), while
the Epstein-Barr nuclear antigen 1 (EBNA1) binds to the promoter
region of the GKN1 gene and induces the reduction of its
transcription (28,29).
Epigenetic modifications are as important to the
regulation of gene expression and the initial stages of disease as
genetic modifications. Differential changes have been documented in
the expression profile of microRNAs (miRNAs) in gastritis or cancer
patients infected with H. pylori (17,30-33),
gastric cancer cell lines (34-36),
CD4 T lymphocytes, macrophages, monocytes and dendritic cells
(37-39). It has been proposed that some
components of H. pylori induce the activation of signals
that modify the expression of miRNAs in the host cells, and that
changes in the global expression profile of miRNAs are related to
the genotype of the bacteria (31).
The reduction of GKN1 cannot be explained by
mutations in its gene, the methylation of its promoter or the
protea-somal degradation of the protein (20,26,27).
It is probable that some miRNAs modulate the decreased translation
of GKN1 mRNA and, consequently, the reduction of the level of
protein in the gastric mucosa. The role of miRNAs in the regulation
of GKN1 expression in the normal gastric mucosa, or mucosa infected
with H. pylori, affected by preneoplasic lesions or with
gastric cancer, has not been explored. The present review comprises
an analysis of the information published on the regulation of GKN1
expression, proposing a model that integrates the probable
regulatory mechanisms at the transcriptional, post-transcriptional
and translational levels.
2. GKN1
GKN1 (also known as CA11, AMP-18, foveolin or TFIZ2)
is a small protein of 181-184 amino acids, specifically expressed
in the stomach. The GKN1 gene is located on chromosome
2p13.3 and is composed of six exons separated by relatively short
introns (6). GKN1 is composed of:
i) A hydrophobic signal peptide in the extreme
NH2-terminal, whose processing generates a protein of
160 amino acids with a molecular mass of 18 kDa; ii) a BRICHOS
domain with three conserved amino acid residues, one aspartic acid
residue and two cysteine acid residues; and iii) a COOH-terminal
domain (6,19,40-43).
GKN1 is a member of the BRICHOS superfamily of proteins, which
includes proteins associated with the development of cancer. It is
a protein with both an autocrine and paracrine function, which
promotes the healing of the mucosa and facilitates cellular
restitution and proliferation (43). GKN1 modulates the progression of
the cell cycle, cellular proliferation and viability, and
apoptosis.
Additionally, GKN1 regulates the production of
reactive oxygen species (ROS) and the PI3K/Akt signaling pathway,
thus influencing epithelial mesenchymal transition (EMT) and the
migration of cancerous cells. GKN1 significantly inhibits the
expression of the mRNA of DNA (cytosine-5)-methyltransferase 1
(DNMT1) and histone-lysine N-methyltransferase EZH2 (EZH2) and the
activity of DNMT1, functions that link this protein to the
inhibition and progression of cancer (43,44).
In the normal gastric mucosa, GKN1 is expressed by
epithelial cells on the surface, but not at the depth of the glands
of the gastric mucosa (21,45,46).
GKN1 reduces the expression of the gastrin receptor,
gastrin/cholecystokinin type B receptor, thus inhibiting the cell
proliferation induced by this hormone (13). GKN1 activates the p16/Rb and p21
signaling pathways, inhibits cell growth and drives cells to
senescence (46). GKN1 modulates
the expression of cytokines and other inflammatory mediators
associated with gastric carcinogenesis, inducing the increased
expression of interleukin (IL)-8 and IL-17 and the decreased
expression of nuclear factor (NF)-κB, IL-6 and IL-10. Thus, it
regulates the immune response and inhibits the progression of
epithelial gastric cells to cancerous cells. GKN1 suppresses the
activation of NF-κB, and thus inhibits the onco-genic signaling
regulated by this transcription factor (9).
3. GKN1, H. pylori infection and
gastric cancer
GKN1 and H. pylori infection.
Infection with H. pylori cagA+ strains increases
the risk of gastric cancer and is related to the reduced expression
of GKN1 in the mucosa (47). In
mice infected with H. pylori-cagA+, the increased
expression of the antiapoptotic proteins Bcl-2, Bcl-XL and induced
myeloid leukemia cell differentiation protein Mcl-1, as well as
NF-κB and proteins related to EMT, is found, while the expression
of p53, p21, p16 and stress response genes decreases (48). The ectopic expression of GKN1
suppresses the effects of H. pylori-cagA+ in the
human gastric cancer cell lines AGS, MKN1 and MKN28. Based on these
findings, it has been suggested that GKN1 suppresses the malignant
transformation of gastric epithelial cells and the progression to
gastric cancer (48).
The expression of GKN1 decreases at the mRNA and
protein levels in dyspeptic patients and is not detected in the
mucosa of subjects with intestinal-type gastric cancer, both with
and without H. pylori infection (19,20,21,22,30,49-52),
or with a diffuse-type cancer (19,23,25).
GKN1 and gastric cancer
GKN1 is absent in human gastric tumors and acts as a
tumor suppressor, regulating cell proliferation, apoptosis,
migration and invasion in gastric cancer cell lines (10). Stimulating the expression of Fas
receptor and the activation of caspase-3, this protein modulates
apoptotic signals, playing an important role in the repair of
tissues during the early stages of neoplastic transformation
(7).
In AGS, MKN-1 and MKN-28 gastric cancer cell lines
transfected with GKN1, the re-expression of p16 and a reduction in
CDK4, cyclin D1 and E2F levels was observed (8) In gastric cancer SGC7901 cells, GKN1
reduces the expression of MMP2, through the deactivation of NF-κB
(15), and induces the expression
of miRNA (miR)-185. The extreme 3′ untranslated region (UTR) of
RhoA mRNA has sequences with affinity to miR-185 and, when this
hybrid miRNA with RhoA mRNA reduces its translation, the silencing
of RhoA is indirectly mediated by GKN1. c-Myc is a transcription
factor that activates RhoA expression and is a target of miR-34a, a
miRNA whose expression is promoted by GKN1. Thus, GKN1 also
deactivates RhoA via miR-34a. These data suggest that GKN1 inhibits
cell motility and invasion by means of the deactivation of RhoA
(16).
4. GKN1 as a potential biomarker of gastric
carcinogenesis
In ~80% of gastric cancer cases, symptoms are scarce
and non-specific at the early stages of the disease, with the
majority of patients diagnosed at an advanced stage with metastasis
already occurring (44,53). Thus, the treatment of this
malignancy is ineffective and the prognosis for patients is
unfavorable. Due to the late diagnosis and consequent limited
therapy options for most patients, the 5-year survival rate is
<20% (54). The lack of
criteria and useful markers for early diagnosis has led to studies
being conducted on the expression of genes associated with gastric
carcinogenesis, with the objective of identifying biomarkers
characteristic of premature stages of the disease. GKN1 is one of
the proteins considered to be potential biomarker of
carcinogenesis.
There are few reports in the available literature on
the identification of GKN1 in samples taken from patients. Nardone
et al (17) identified the
presence of GKN1 in human gastric tissue, finding that its
expression decreases in the event of H. pylori infection,
deteriorates progressively from chronic gastritis to atrophic
gastritis, and is not detected in areas in which intestinal
metaplasia or H. pylori-positive tumors are found (17). GKN1 is absent in cases of gastric
cancer without H. pylori infection (17-25).
Villano et al (55)
analyzed the level of GKN1 mRNA in serum taken from patients
with gastric cancer and apparently healthy volunteers, finding no
statistically significant differences between patients with cancer
and healthy volunteers. The aforementioned results indicate that
GKN1 mRNA is not a useful biomarker for the diagnosis of
gastric cancer (55). Yoon et
al (56) found that the serum
levels of GKN1 are significantly lower in gastric cancer patients
than in either apparently healthy subjects or patients with
hepatocel-lular and colorectal carcinoma (P<0.0001). These data
suggest that the serum levels of GKN1 may be used for the
differentiation of patients with gastric cancer from those with
other malignancies of the digestive system and clinically healthy
subjects. The authors concluded that the serum concentration of
GKN1 may be an informative diagnostic biomarker for gastric cancer
(56). Dokhaee et al
(44) reported that GKN1
mRNA is significantly reduced in the gastric tissue of patients
with gastric cancer, compared to normal tissue. The results led to
the hypothesis that GKN1 may be a reliable biomarker for the
detection of gastric cancer in its early stages.
The aforementioned data indicate that the
measurement of circulating GKN1 concentration, the protein or the
mRNA in gastric tissue may be of utility for the early diagnosis of
cancer. However, it is necessary to strengthen these findings with
more research in patients with preneoplasic lesions (atrophic
gastritis, intestinal metaplasia and dysplasia) and cancer in
distinct stages of evolution.
5. Regulation of GKN1 expression in mucosa
infected with H. pylori or with gastric cancer
At the chromosomal level, cytogenetic aberrations,
such as duplications, translocations, deletions or the loss of
heterozygosity in the 2p13 chromosome (in which the GKN1
gene is found) have not been detected (57,58).
The sequence of the GKN1 gene was analyzed in
81 gastric tumors and 40 adenomas, confirming a lack of mutations
(20) These data suggested that
the reduction of GKN1 cannot be attributed to cytogenetic
aberrations or mutations, and that other mechanisms are involved in
the deregulation of this protein.
Gene expression is regulated at different levels,
from transcription to translation (59). At the transcriptional level,
regulation occurs via epigenetic modifications, such as the
modification of histones and the methylation of DNA (60,61).
At the post-transcriptional level, the role of small RNAs (miRNAs)
in the modulation of translation should be taken into account
(62), while at post-translational
level, ubiquitination, followed by the proteasomal degradation of
the marked protein, is the best-known mechanism involved in the
reduction in cytoplasmic levels of proteins (61).
Transcription factors
Transcription factors are able to activate or
repress the expression of a gene (63,64).
Little is known about the transcriptional regulation of
GKN1. Yoon et al (65), using luciferase and chromatin
immunoprecipitation assays, confirmed that NKX6.3 is a
transcription factor for GKN1, and located the recognition
sequence corresponding to NKX6.3 in the promoter region of the
GKN1 gene (Fig. 1A). NKX6.3
positively modulates the transcription of GKN1, which is
reflected in the increased level of both mRNA and protein (65).
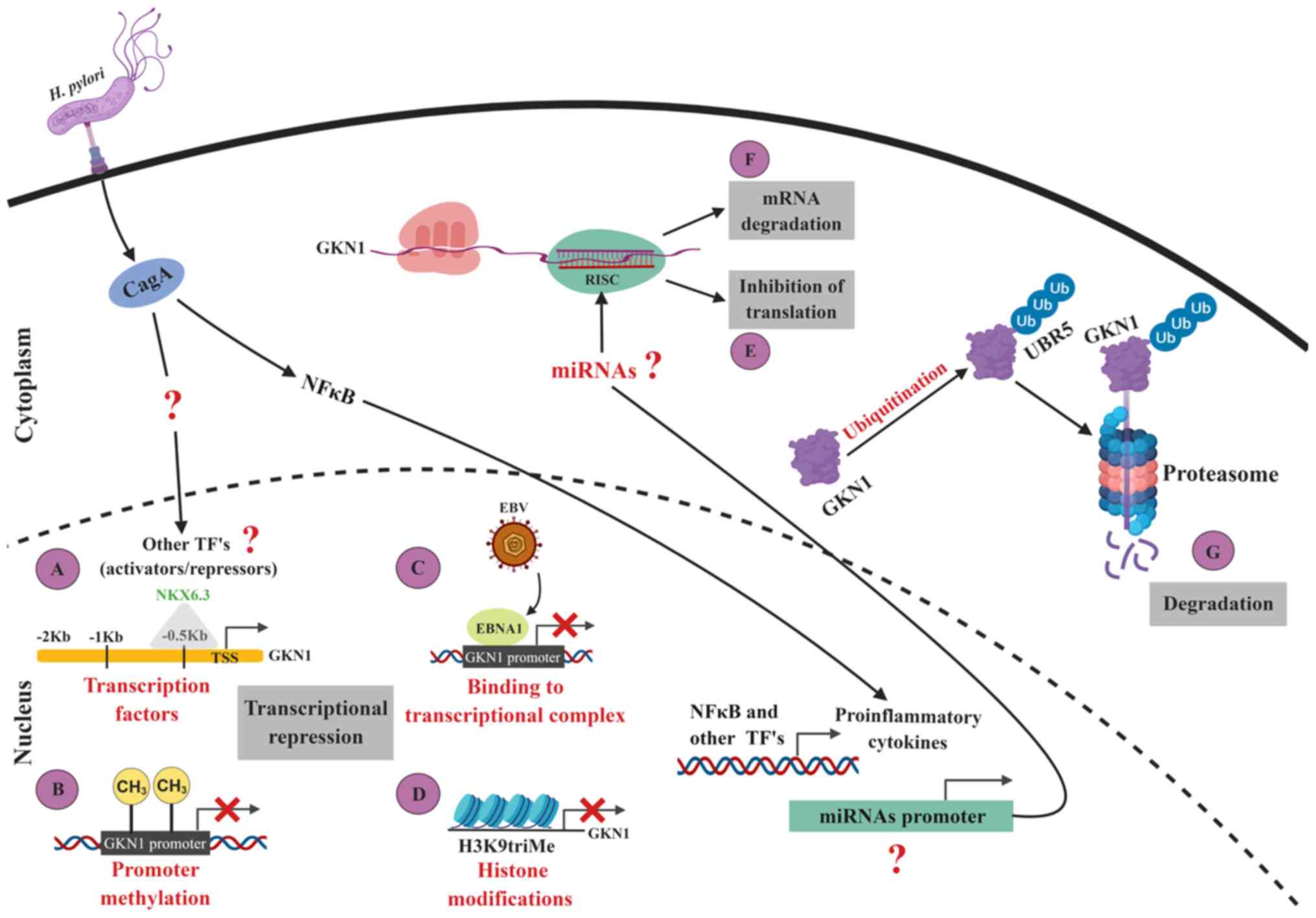 | Figure 1Regulation of GKN1 expression. (A)
NKX6.3 is the only transcription factor validated as a positive
regulator of GKN1 transcription. It is likely that another
transcription factor or factors act as activators or repressors of
GKN1 transcription during the infection of the gastric epithelium
by H. pylori. Evidence indicates that H. pylori
activates different signaling pathways that induce the expression
of various transcription factors. The decrease in or loss of GKN1
expression in gastric cancer may be a consequence of: (B) GKN1
promoter methylation; (C) EBNA1 binding to the transcriptional
complex; or (D) histone modification, such as trimethylation of
lysine 9 in histone 3. Additionally, it is possible that the GKN1
mRNA is targeted by some miRNAs. By in silico analysis,
miRNAs were found with sequences complementary to sites located in
the 3' untranslated region of GKN1 mRNA, and are likely to
contribute to its negative post-transcriptional regulation through:
(E) Inhibition of translation; or (F) mRNA degradation. The
expression of the proposed miRNAs may or not be induced by H.
pylori. GKN1 is degraded in the cytoplasm of epithelial cells
when (G) the ubiquitin ligase UBR5 marks GKN1 for its degradation
in the proteasome. These and other mechanisms can act
synergistically to promote the diminution or silencing of GKN1
expression, manifesting as a decreased or absent mRNA and protein
expression in gastric tissue or in the circulation. Figure created
using BioRender. com. TFs, transcription factors; GKN1, gastrokine
1; UBR5, E3 ubiquitin-protein ligase UBR5; EBNA1, Epstein Barr
nuclear antigen 1; EBV, Epstein Barr virus; RISC, RNA-induced
silencing complex; H. pylori, Helicobacter pylori;
miRNA, microRNA; Ub, ubiquitin; NF-κB, nuclear factor-κB; TSS,
transcriptional start site. |
By means of in silico analysis, conducted
using the MatInspector (66)
(http://www.genomatix.de/matinspector.html), AliBaba2.1
(67) (http://gene-regulation.com/pub/programs/alibaba2/index.html)
and TfsiteScan 68) (http://www.ifti.org) programs, transcription factors
were identified with affinity to recognition sequences in the
GKN1 promoter region (Fig.
2A and Table I). It is likely
that one or more of these transcription factors, predicted
bioinformatically, are involved in the transcriptional regulation
of GKN1. Experimental confirmation of the effect exerted by
the proposed transcription factors on the modulation of GKN1
expression will improve understanding of the mechanisms involved in
the regulation of the expression of this protein.
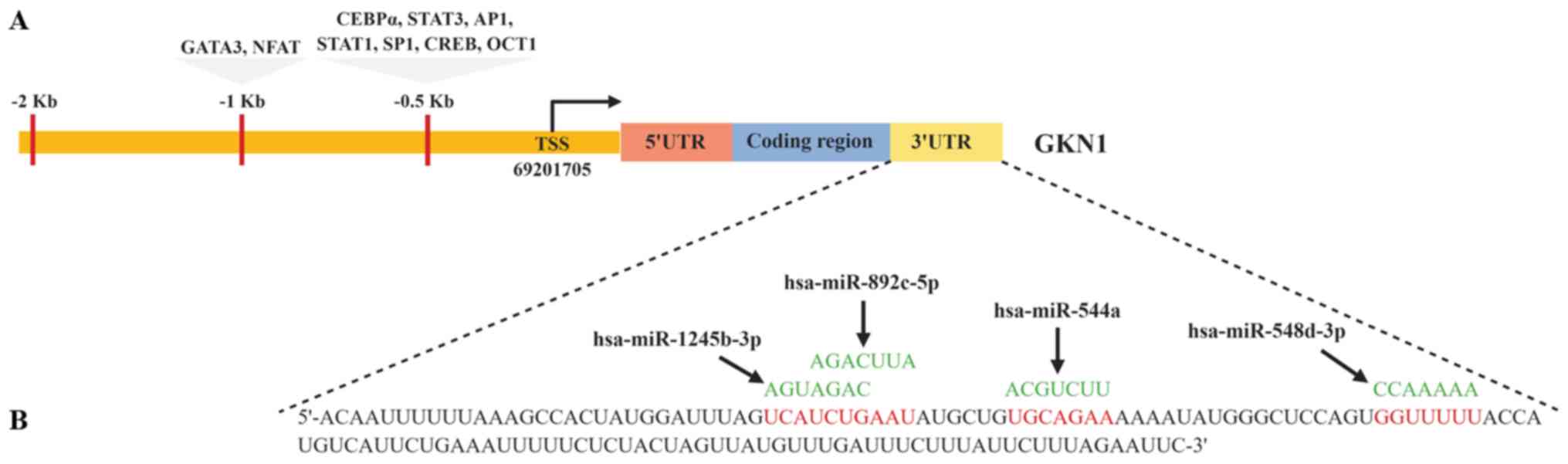 | Figure 2Transcription factors and miRNAs
predicted in silico as regulators of GKN1 expression. By
in silico analysis (A) transcription factors were identified
with affinity to recognition sequences in the GKN1 promotor region
and (B) miRNAs were found with sequences complementary to sites
located in the 3'UTR of GKN1 mRNA. It is likely that the
transcription factors act as activators or repressors of the
transcriptional regulation of GKN1, and the miRNAs contribute to
post-transcriptional regulation, inhibiting translation or inducing
mRNA degradation. Figure created using BioRender.com TSS, transcriptional start site; UTR,
untranslated region; miRNA/miR, microRNA; GKN1, gastrokine 1;
GATA3, T-cell-specific transcription factor GATA-3; NFAT, nuclear
factor of activated T cells; CEBPα, CCAAT enhancer binding
protein-α; AP-1, AP-1 transcription factor; Sp1, transcription
factor Sp1; CREB, cyclic AMP-responsive element-binding protein
3-like protein 4. |
 | Table ITranscription factors with affinity
to binding sequences in the gastrokine 1 gene promoter region. |
Table I
Transcription factors with affinity
to binding sequences in the gastrokine 1 gene promoter region.
| Transcription
factor | Number of binding
sites | Binding
sequence | Genomic position of
binding sequencea |
|---|
| GATA-3 | 7 | CAGAGATAAAATG |
68974044-68974056 |
| CEBPα | 7 |
GAAATTGAGGAAGGT |
68974539-68974553 |
| Oct-1 | 6 |
GTCATGCAATTGATC |
68973972-68973986 |
| AP-1 | 4 | TGATGAGTCAGGT |
68974444-68974456 |
| STAT-3 | 3 |
AGGTTTCCTGGTACACTGG |
68974502-68974520 |
| Sp1 | 2 |
GCTGTGGGCGTGAGTAT |
68974361-68974377 |
| STAT-1 | 1 |
AGTGTACCAGGAAACCTTT |
68974500-68974518 |
| CREB | 1 |
AGGGTCCTATGTAATAAGATT |
68973801-68973821 |
| NFAT | 1 |
CTTTGGAAATCTTATTACA |
68973794-68973812 |
In patients with gastric pathology, in murine models
or in gastric epithelial cell lines, the expression of trans-acting
T-cell-specific transcription factor GATA-3 (GATA-3), STAT-1,
STAT-3, transcription factor Sp1 (Sp1), cyclic AMP-responsive
element-binding protein 3-like protein 4 (CREB), AP-1 transcription
factor (AP-1) and Oct-1 increases, while the levels of CCAAT
enhancer binding protein-α (CEBPα) and NKX6.3 decrease (65,69-82).
The expression of GATA-3 was found to have increased at different
stages of the carcinogenesis associated with H. pylori in
patient biopsies, murine models and human gastric epithelial cells
(73,74).
CagA and OipA of H. pylori induce the
activation of transcription factors such as AP-1, NF-κB, STAT-3,
CREB and nuclear factor of activated T cells (NFAT), which favor
the expression of IL-6, and cytokines, which promote inflammation
(83-89). IL-6 stimulates the activation of
the signaling pathway gp130/STAT3 in gastric cancer cell lines
(90), while CagA stimulates the
expression of the NFAT transcription factor in AGS cells (88).
The protein Tipα, produced by H. pylori,
activates the IL-6/STAT3 pathway (89). H. pylori
cagA+ strains induce signaling through the MAPK
pathway, thus increasing proliferation and activating transcription
factors such as AP-1 (70).
Through toll like receptor (TLR)2 and TLR9, H. pylori
activates the MAPK pathway and, downstream, the factors AP-1 and
CREB, which positively regulate the transcription of cyclooxygenase
2 (COX-2) (91). CREB and STAT-3
are activated by H. pylori and positively regulate the
transcription of COX-2 in gastric epithelial cells (72,83).
Increased STAT-3 expression has been found in biopsies of the
gastric mucosa infected with H. pylori cagA+
(80), as well as in cell lines
and murine models (86). CagA
promotes the phosphorylation of STAT-3 in gastric epithelial cells
(92).
In vitro and in vivo experiments have
shown that the protein OipA of H. pylori stimulates the
phosphorylation of STAT-1 (93)
and that H. pylori alters the STAT-1 signaling induced by
IFN-γ in gastric epithelial cells. This event may represent an
adaptation of the bacteria in order to modulate the immune response
of the host mucosa, allowing the bacteria to survive in the stomach
(94).
The expression level of the Sp1 transcription factor
increases in gastric adenocarcinoma and is related to the cancer
stage, the depth of infiltration and an unfavorable prognosis for
patients (82). The expression of
Sp1 differs between intestinal-type and diffuse-type cancer, while
low-level expression of Sp1 is related to the progression and
metastasis of intestinal-type cancer, in contrast to diffuse-type
cancer (95). Sp1 is essential in
the regulation of genes that determine the characteristics of
cancer (96). In AGS cells, the
ERK1/2 signaling pathway, activated in response to H. pylori
infection, in turn activates Sp1, which modulates the transcription
of vascular endothelial growth factor-A (69). It is probable that the factors
AP-1, Oct-1, STAT-1, STAT-3, GATA-3, Sp1, CREB and NFAT, with
recognition sequences in the GKN1 promotor and being
activated by H. pylori, repress the transcription of
GKN1 in infected mucosa or mucosa with gastric cancer.
In normal gastric mucosa, CEBPα is expressed in the
foveolar epithelium and is reduced in the tumor tissue of patients
with gastric cancer (79), and in
the cell lines MKN45 and MKN74 (97). The ectopic expression of CEBPα in
gastric cancer cell lines reduces cell viability (97). The level of CEBPα expression
gradually decreases in line with the advancing carcinogenesis
associated with H. pylori infection (73). Given the function of CEBPα in the
regulation of the viability of cancerous cells and the fact that
GKN1 and CEBPα levels gradually decrease in line with the progress
of the lesion, it is probable that this factor is an activator of
GKN1 transcription. The reduced expression of CEBPα and
NKX6.3 in gastric cancer may be due to the negative regulation
mediated by microRNAs.
DNA methylation
Changes in the methylation of DNA lead to changes in
gene expression. The hypermethylation of CpG islands located in the
promoter region of a gene results in the decrease or silencing of
the expression of the gene.
The methylation of the GKN1 promoter was
previously studied, finding hypermethylation in the CpG islands of
the promoter region in only two of 25 gastric tumors. This evidence
indicates that the low or null GKN1 expression in the inflamed
tissue, either tumoral or infected by H. pylori, is not due
to the methylation of its promoter in all cases (Fig. 1B) (20).
The protein EBNA1 of EBV is able to bind to the
promoter region of various genes of the host (28). It has been found to have an
affinity with the sequences contained in the GKN1 promoter
and, binding at these sites, contributes to the deregulation of
GKN1 in gastric cancer associated with Epstein-Barr infection
(Fig. 1C) (29). Only a small proportion of gastric
tumors contain EBV.
Histone modification
The modification of histones is an epigenetic
mechanism influencing gene expression. Altieri et al
(26) analyzed six gastric tumors
in order to determine whether histone modification contributes to
GKN1 regulation. Chromatin immunoprecipitation assays were
conducted on a fragment of 600 pb of the GKN1 gene promoter,
including the 5′UTR, finding trimethylation in lysine 9 of histone
3 (H3K9triMe), among bases -148 and -310 of the GKN1 gene
promoter in the six gastric tumors. H3K9triMe is a gene repression
marker that generates binding sites for histone deacetylase I
(HDAC1) (98) (Fig. 1D). The inhibition of HDAC1 activity
with trichostatin A, a hypomethylating agent, is related to
increased GKN1 mRNA levels but not to the protein itself.
These findings suggest that the regulation of GKN1 may occur at the
post-transcriptional level via miRNAs (26).
miRNAs
In the gastric mucosa, miRNAs can be expressed by
epithelial cells, infiltrating inflammatory cells, transformed
cells or cancerous cells (31). In
the regulation of gene expression, miRNAs inhibit the translation
or induce the degradation of target transcripts. It is likely that
some miRNAs impede the translation of GKN1 mRNA and,
consequently, are responsible for the reduction in the protein
level, although there are no reports indicating whether a miRNA is
involved in the regulation of GKN1 expression to the best of
our knowledge.
In order to explore whether GKN1 mRNA has
binding sites for one or more miRNAs, an in silico analysis
was conducted using programs for predicting miRNA targets:
TargetScan (99) (2015, http://www.targetscan.org), miRanda (100) (http://www.microrna.org), miRDB (101) (http://mirdb.org),
miRSystem (102) (http://mirsystem.cgm.ntu.edu.tw/index.php) and
DianaTools (103) (http://www.microrna.gr/microT-CDS), based on
thermodynamic and base complementarity analysis. miRNAs were found
with sequences complementary to sites located in the 3′UTR region
of GKN1 mRNA (Fig. 2B),
four of which are able to hybridize with canonical sites of
GKN1 mRNA and possess two or three guanine or cytosine
residues in the seed region of the miRNA, conferring them greater
binding stability. An adenine in position 1 of the 3′UTR region of
GKN1 mRNA ensures the recognition of the transcript by the
RNA-induced silencing complex. These characteristics increase the
probability that a miRNA will interact with the 3′UTR of
GKN1 mRNA (Table II).
| Table IImiRNAs proposed as a candidate to
regulate GKN1 expression. |
Table II
miRNAs proposed as a candidate to
regulate GKN1 expression.
| miRNAa | 3'UTR GKN1b | miRNA recognition
site | Site type |
|---|
| hsa | 46-53 | GKN1 5′ | u | c | u | g | a | a | u | a | u | g | c | u | g | U | G | C | A | G | A | A | A | 8mer |
| miR-544a | | miRNA 3′ | u | u | g | a | a | c | g | a | u | u | u | u | u | A | C | G | U | C | U | U | A | |
| hsa | 70-77 | GKN1 5′ | u | a | u | g | g | g | c | u | c | c | a | g | u | G | G | U | U | U | U | U | A | 8mer |
| miR-548d-3p | | miRNA 3′ | g | u | u | u | u | c | a | u | u | g | a | u | a | C | C | A | A | A | A | A | C | |
| hsa | 30-37 | GKN1 5′ | a | c | u | a | u | g | g | a | u | u | u | a | g | U | C | A | U | C | U | G | A | 8mer |
| miR-1245b-3p | | miRNA 3′ | u | a | u | c | c | g | g | a | a | a | u | c | u | A | G | U | A | G | A | C | U | |
| hsa | 33-40 | GKN1 5′ | a | u | g | g | a | u | u | u | a | g | u | c | a | U | C | U | G | A | A | U | A | 8mer |
| miR-892c-5p | | miRNA 3′ | a | c | u | g | a | c | c | g | u | g | g | a | a | A | G | A | C | U | U | A | U | |
Multiple prediction programs may be used to locate
binding sites for miRNAs in gene transcripts. The results of the
analysis facilitated the selection of miRNAs with a higher
probability of binding to their target, miR-544a was predicted by
five programs, according to the aforementioned criteria. It is
highly probable that miR-544a is a regulator of GKN1
(104,105). This proposal is strengthened by
experimental data on the expression of miR-544a, which is found at
increased levels in gastric cancer cell lines (106). To the best of our knowledge,
research has not been conducted on the expression of
hsa-miR-1245b-3p, hsa-miR-892c-5p and hsa-miR-548d-3p in the
gastric mucosa, be that infected, inflamed, atrophic or with
gastric cancer. However, the results of the in silico
analysis suggested a high probability that these miRNAs regulate
the expression of GKN1, either inhibiting the translation of
the transcript or promoting its degradation (Fig. 1E and F).
Currently, >5,000 miRNAs are registered on
miRBase, while in silico predictions estimate that more than
one-third of the human transcriptome can be regulated by miRNAs. In
gastric cancer, inflammatory processes and H. pylori
infection, miRNAs fulfil an important function in the deregulation
of gene expression (31,33). GKN1 is absent in cancer, both with
and without H. pylori, and is reduced in patients with
gastritis, in gastric mucosa infected by H. pylori and in
atrophic gastritis (30). The
reduction in or absence of GKN1 transcripts or protein is not
completely explained by the studied mechanisms of transcriptional
and post-translational regulation. It is likely that some miRNAs
regulate GKN1 expression directly or indirectly at the
post-transcriptional level.
It has been reported that ROS deregulate the
expression of miRNAs in tissue infected with H. pylori or
gastric cancer tissue (107,108). It is also known that ROS induce a
decrease in the number of copies of GKN1 mRNA in tissue
infected by H. pylori (48). These data support the hypothesis
that, in H. pylori infection, ROS alter the expression of
miRNAs, among which are those with the GKN1 transcript as a
target. The cytotoxins VacA and CagA, lipopolysaccharide and
peptidoglycan, among other components of H. pylori, are able
to induce the increased expression of miRNAs that inhibit
translation or induce the degradation of the GKN1
transcript, thus modulating the decrease in the levels of this
protein.
From the first stages of H. pylori infection,
the inflammation associated with it causes changes in the
expression of proteins and miRNAs, alterations in cell signaling,
and unbalanced cell proliferation and apoptosis in gastric
epithelial cells, promoting the progression of gastritis to
pre-neoplastic and neoplastic lesions (109). The abnormal expression of miRNAs
is common in different types of cancer (110), with the evidence indicating
changes in the expression profiles of miRNAs in gastric cancer and
in mucosa infected by H. pylori.
Alterations in the expression of miRNAs can
manifest either as increases or decreases (111). In the gastric mucosa, both with
and without H. pylori infection, it has been found that
miRNAs with changes in their expression levels in response to H.
pylori can be similar to or different from those observed in
gastric cancer with a negative result for bacteria (31) (Tables III and IV). Chang et al (33) found that hsa-miR-99b-3p,
hsa-miR-564 and hsa-miR-658 expression increased in cancerous
tissue infected with H. pylori, while hsa-miR-204-5p,
hsa-miR-338-5p, hsa-miR-375 and hsa-miR-548c-3p were found to be
overexpressed in cancer tissue without H. pylori (33).
| Table IIImiRNA expression in patients and
in vitro models with H. pylori infection. |
Table III
miRNA expression in patients and
in vitro models with H. pylori infection.
| Author, year | Model | H. pylori
strain | Method used to
determine miRNA expression | Upregulated
miRNAs | Downregulated
miRNAs | Refs. |
|---|
| Cho et al,
2016 | Murine | C57BL/6 mice | SS1 | Mouse whole genome
miRNA array (Agilent Technologies, Inc.) release 15 | - | mmu-miR-1
mmu-miR-133a
mmu-miR-133b
mmu-miR-203
mmu-miR-205
mmu-miR-490-3p | (116) |
| Belair et al,
2011 | Intestinal type
gastric cancer cell lines | AGS | 26695 | Reverse
transcription-quantitative PCR | hsa-miR-21 |
hsa-miR-371-3p
hsa-miR-372
hsa-miR-373
hsa-miR-19b
hsa-miR-160b
hsa-miR-320 | (117) |
| Santos et
al, 2017 | | AGS | 26695 y P12 | Cancer Pathway
Finder miRNA PCR Array (MIHS-102Z; Qiagen, Inc.) |
hsa-miR-150-5p
hsa-miR-155-5p
hsa-miR-3163 | | (35) |
| Chang et
al, 2015 | Patients diagnosed
with gastric cancer | Eight intestinal
type gastric cancer patients with H. pylori infection | No data | Human miRNA
microarray (Agilent Technologies, Inc.), release 16.0 |
hsa-miR-32-3p
hsa-miR-514a-3p
hsa-miR-181c-3p
hsa-miR-146a-5p
hsa-miR-4319
hsa-miR-99b-3p
hsa-miR-543
hsa-miR-564
hsa-miR-645
hsa-miR-431-3p
hsa-miR-650
hsa-miR-638
hsa-miR-139-3p
hsa-miR-508-3p
hsa-miR-2355-3p
hsa-miR-934
hsa-miR-548f
hsa-miR-1304-5p | - | (33) |
| Table IVmiRNAs expression in cell lines and
patients with gastric Cancer, grouped based on Lauren's
Classification (118). |
Table IV
miRNAs expression in cell lines and
patients with gastric Cancer, grouped based on Lauren's
Classification (118).
| Author, year | Model | Mefhod used to
determine the miRNAs expression | Upregulated
miRNAs | Downregulated
miRNAs | Refs. |
|---|
| Yu e tal,
2012 | Intestinal type
gastric Cancer cell lines | NCI-N87
AGS
MKN28
BGC-823
SGC-7901 | Human microRNA
Microarray v.2 (Agilent Technologies), containing probes for 723
human microRNAs |
hsa-miR-196a
hsa-miR-615
hsa-miR-196b
hsa-miR-92b
hsa-miR-149 |
hsa-miR-376a
hsa-miR-145
hsa-miR-143
hsa-miR-451
hsa-miR-142-5p | (119) |
| Diffuse type
gastric Cancer cell lines | SNU-1
SNU-16
KATO III
MKN45 | |
hsa-miR-550
hsa-miR-183
hsa-miR-301
hsa-miR-18a
hsa-miR-106a
hsa-miR-17-5p
hsa-miR-18b
hsa-miR-19a
hsa-miR-221
hsa-miR-93
hsa-miR-20a | hsa-miR-1
hsa-miR-377
hsa-miR-495
hsa-miR-409-3p
hsa-miR-368
hsa-miR-142-3p
hsa-miR-150
hsa-miR-497
hsa-miR-214
hsa-miR-199a
hsa-miR-146b
hsa-miR-133b
hsa-miR-127
hsa-miR-381
hsa-miR-195
hsa-miR-648
hsa-miR-223
hsa-miR-135a
hsa-miR146a
hsa-miR-136
hsa-miR 126
hsa-miR-29c
hsa-miR-572 |
| Guo et al,
2009 | Patients diagnosed
with gastric Cancer | Intestinal
type | μParaflo™
microfiuidic chip (LC Sciences) |
hsa-miR-31
hsa-miR-133b
hsa-miR-139-5p
hsa-miR-195
hsa-miR-378
hsa-miR-497
hsa-miR-768-3p |
hsa-miR-17
hsa-miR-18a
hsa-miR-18b
hsa-miR-19a
hsa-miR-20a
hsa-miR-20b
hsa-miR-21
hsa-miR-106a
hsa-miR-106b
hsa-miR-340
hsa-miR-421
hsa-miR-658 | (120) |
| Udea et al,
2010 | | | ArrayExpress, v3.0
(European Bioinformatics Institute), contains 1,100 microRNA
probes, 326 human and 249 mouse |
hsa-miR-373
hsa-miR-498
hsa-miR-202
hsa-miR-494 | - | (121) |
| Tsukamoto et
al, 2010 | | Diffuse type | G4470A Human MiRNA
Microarray (Agilent Technologies, Inc.), of 470 human and 64 virus
mature miRNAs based on Sanger miRBase release 9.1 |
hsa-miR-18a
hsa-miR-106a
hsa-miR-17-5p
hsa-miR-146a
hsa-miR-93
hsa-miR-19a
hsa-miR-20a
hsa-miR-20b
hsa-miR-25
hsa-miR-15b
hsa-miR-425-5p
hsa-miR-92
hsa-miR-194
hsa-miR-10a
hsa-miR-222
hsa-miR-7
hsa-miR-106b
hsa-miR-320
hsa-miR-21
hsa-miR-34a
hsa-miR-19b
hsa-miR-103
hsa-miR-215
hsa-miR-192
hsa-miR-429
hsa-miR-279
hsa-miR-223
hsa-miR-23a
hsa-miR-107
hsa-miR-200b
hsa-miR-24
hsa-miR-15a
hsa-miR-16 |
hsa-miR-375
hsa-miR-29c
hsa-miR-148a
hsa-miR-30a-5p
hsa-miR-30e-5p
hsa-miR-638 | (122) |
| Su et al,
2012 | | | AFFX miRNA
expression chips (Affymetrix; Thermo Fisher Scientific, Inc.) |
hsa-miR-455-3p
hsa-miR-34a |
hsa-let-7g
hsa-miR-200b
hsa-miR-768-3p
hsa-let-7d
hsa-miR-104-3p
hsa-miR-27b
hsa-miR-1207-5p
hsa-miR-663
hsa-miR-486-5p
hsa-miR-222
hsa-miR-574-3p
hsa-miR-768-5p
hsa-miR-378
hsa-miR-31 | (123) |
| Juzenas et
cd, 2015 | | | TaqMan Array Human
MiRNA Card A v2.1 (Applied Biosystems; Thermo Fisher Scientific,
Inc.), include 377 human miRNAs of miRBase v20 |
hsa-miR-146b-5p
hsa-miR-155-5p
hsa-miR-214-3p
hsa-miR-223-3p
hsa-miR-224-5p
hsa-miR-331-3p
hsa-miR-484 |
hsa-let-7a-5p
hsa-let-7g-5p
hsa-miR- 135a-5p
hsa-miR-148a-3p
hsa-miR-204-5p
hsa-miR-26b-5p
hsa-miR-30b-5p
hsa-miR-375 | (124) |
| Katada et
cd, 2009 | | | TaqMan miRNA
assays |
hsa-miR-34b
hsa-miR-34c
hsa-miR 128a |
hsa-miR128b
hsa-miR-129
hsa-miR-148a | (125) |
| Ueda et cd,
2010 | | | ArrayExpress,
Version 3.0 (European Bioinformatics Institute), A-MEXP-620,
contains 1,100 microRNA probes, 326 human and 249 mouse |
hsa-miR-105
hsa-miR-100
hsa-miR-125b
hsa-miR-199a
hsa-miR-99a
hsa-miR-143
hsa-miR-145
hsa-miR133a | | (121) |
In infected mucosa, hsa-miR-223 expression was
found to be increased, while in mucosa without H. pylori,
hsa-miR-203, hsa-miR-204, hsa-miR-455, hsa-miR-141 and hsa-let-7f
were found to be overexpressed (31). The levels of let-7, miR-125a and
miR-500 were found to be significantly reduced in cells infected
with cagA+ strains, although not in those
infected with cagA− strains. These results
indicate that miRNAs participate in gastric pathogenesis, whether
associated and not associated with H. pylori, and suggest
that the CagA oncoprotein of H. pylori regulates the
differential expression of miRNAs in epithelial gastric cells
(31). Increased hsa-miR-127-5p,
hsa-miR195, hsa-miR-196a, hsa-miR-206, hsa-miR-216 and miR-488
expression has been found, while decreased hsa-miR-103,
hsa-miR-141, hsa-miR-17-3p, hsa-miR34a and let-7i expression has
been found in gastric epithelial cells infected with different
H. pylori-cagA+ strains (36).
H. pylori is able to modify the expression
of miRNAs by means of inflammatory effectors (112). In gastric epithelial cells, the
pro-inflammatory cytokines IL-8, tumor necrosis factor-α and IL-1β
induce the expression of miR-146a (113), while the oncoprotein CagA
positively regulates c-myc, which is related to the
decreased expression of miR-26a and miR-101. The decrease in the
expression of these miRNAs contributes to increased levels of the
histone methyltransferase EZH2 and methyltransferase DNMT3B, which
promote the methylation of the let-7 promoter (114).
Ubiquitination
E3 ubiquitin-protein ligase UBR5 (UBR5) is an E3
ubiquitin ligase that participates in the ubiquitin-proteasome
system, regulating protein concentration via ubiquitination and
degradation, and is deregulated in different types of cancer
(115). UBR5 increases in the
cancerous tissues of gastric cancer patients, while an interaction
between UBR5 and GKN1 has been observed through immunoprecipitation
assays. These results suggest that UBR5 participates in the
ubiquitination of GKN1, and that at least part of this protein is
sent to be degraded by the proteasome (27) (Fig.
1G). Thus, UBR5 contributes to the regulation of gastric
carcinogenesis, inducing the degradation of tumor suppressing
proteins, such as GKN1 (27).
Therefore, promoter methylation, trimethylation of
histones and ubiquitination are mechanisms that contribute to the
regulation of the GKN1 expression in gastric cancer; however, they
do not explain the absence of the protein in cell lines and
cancerous human tissue.
6. Conclusion
GKN1 plays an important role in the maintenance of
gastric homeostasis. In inflamed mucosa, both with and without
H. pylori infection, GKN1 levels decrease, while this
protein is absent in gastric cancer. The measurement of circulating
GKN1 concentration, the protein itself or its mRNA in gastric
tissue could be useful for the early diagnosis of cancer. However,
little is known about the mechanisms that explain the reduction or
silencing of GKN1 expression in gastric carcinogenesis. No
mutations or polymorphisms have been found in the GKN1
promoter region, which explains the reduction in the levels of this
protein. While the modification of histones seems to be involved in
the transcriptional regulation of GKN1, further research is
required to confirm its level of participation in the regulation of
GKN1 in the population. The information available suggests that the
methylation of the GKN1 promotor is an epigenetic mechanism
that reduces the transcription rate of the gene. However, this
mechanism only occurs in some cases of gastric cancer and,
moreover, it is probable that it is determined by the genetic
characteristics of the individual or the presence of EBV in the
tumor. While only factor NKX6.3 has been confirmed as a positive
regulator of GKN1 transcription, in silico analysis
suggests the existence of other transcription factors with affinity
for sequences in the GKN1 promotor region, among which are
GATA-3, CEBP-α, Oct-1, AP-1, STAT-3, SP1, STAT-1, CREB and NFAT. It
is unknown whether miRNAs regulate GKN1 expression at the
post-transcriptional level. In silico analysis revealed that
hsa-miR-544a, hsa-miR1245b-3p, hsa-miR-892c-5p and hsa-miR-548d-3p
have sequences complementary to sites located in the 3′UTR of
GKN1 mRNA. It is likely that, together, they regulate the
expression of GKN1 in vivo, in mucosa infected by H.
pylori or in gastric cancer (Fig.
1). Functional studies are required to show whether miRNAs play
a role in the regulation of GKN1 expression. At the
post-translational level, UBR5 mediates the ubiquitination of GKN1,
marking it for degradation in the proteasome; however, this
mechanism does not explain the absence or minimal level of GKN1
expression in gastric cancer. Clarifying the mechanisms that
regulate GKN1 expression will contribute useful information for
evaluating the possible clinical applications for the detection of
this protein in mucosa, or in the circulation of patients with
gastric diseases both associated and not associated with H.
pylori.
Funding
This work was supported by a doctoral fellowship
from CONACYT to JAM (grant no. 296364).
Availability of data and materials
Not applicable.
Authors' contributions
JAM, DNMC, OPZ and GFT contributed the idea, wrote
the text, and generated the tables and the figure.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declares that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Khurana S and Mills JC: The gastric mucosa
development and differentiation. Prog Mol Biol Transl Sci.
96:93–115. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dimaline R and Varro A: Attack and defence
in the gastric epithelium - a delicate balance. Exp Physiol.
92:591–601. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hoffmann W: Self-renewal of the gastric
ephitelium from stem and progenitor cells. Front Biosci.
S5:720–731. 2013. View
Article : Google Scholar
|
|
4
|
Mueller A, Merrell DS, Grimm J and Falkow
S: Profiling of microdissected gastric epithelial cells reveals a
cell type-specific response to Helicobacter pylori infection.
Gastroenterology. 127:1446–1462. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Silen W and Ito S: Mechanisms for rapid
re-epithelialization of the gastric mucosal surface. Annu Rev
Physiol. 47:217–229. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Menheniott TR, Kurklu B and Giraud AS:
Gastrokines: Stomach-specific proteins with putative homeostatic
and tumor suppressor roles. Am J Physiol Gastrointest Liver
Physiol. 304:G109–G121. 2013. View Article : Google Scholar
|
|
7
|
Rippa E, La Monica G, Allocca R, Romano
MF, De Palma M and Arcari P: Overexpression of gastrokine 1 in
gastric cancer cells induces Fas-mediated apoptosis. J Cell
Physiol. 226:2571–2578. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yoon JH, Choi YJ, Choi WS, Ashktorab H,
Smoot DT, Nam SW, Lee JY and Park WS: GKN1-miR-185-DNMT1 axis
suppresses gastric carcinogenesis through regulation of epigenetic
alteration and cell cycle. Clin Cancer Res. 19:4599–4610. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yoon JH, Cho ML, Choi YJ, Back JY, Park
MK, Lee SW, Choi BJ, Ashktorab H, Smoot DT, Nam SW, et al:
Gastrokine 1 regulates NF-κB signaling pathway and cytokine
expression in gastric cancers. J Cell Biochem. 114:1800–1809. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kim O, Yoon JH, Choi WS, Ashktorab H,
Smoot DT, Nam SW, Lee JY and Park WS: GKN2 contributes to the
homeostasis of gastric mucosa by inhibiting GKN1 activity. J Cell
Physiol. 229:762–771. 2014. View Article : Google Scholar
|
|
11
|
Yoon JH, Seo HS, Choi WS, Kim O, Nam SW,
Lee JY and Park WS: Gastrokine 1 induces senescence and apoptosis
through regulating telomere length in gastric cancer. Oncotarget.
5:11695–11708. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen P, Li YC and Toback FG: AMP-18
targets p21 to maintain epithelial homeostasis. PLoS One.
10:e01254902015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kim O, Yoon JH, Choi WS, Ashktorab H,
Smoot DT, Nam SW, Lee JY and Park WS: Gastrokine 1 inhibits
gastrin-induced cell proliferation. Gastric Cancer. 19:381–391.
2016. View Article : Google Scholar
|
|
14
|
Rippa E, Altieri F, Di Stadio CS, Miselli
G, Lamberti A, Federico A, Quagliariello V, Papale F, Guerra G and
Arcari P: Ectopic expression of gastrokine 1 in gastric cancer
cells up-regulates tight and adherens junction proteins network.
Pathol Res Pract. 211:577–583. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Xing R, Cui JT, Xia N and Lu YY: GKN1
inhibits cell invasion in gastric cancer by inactivating the
NF-kappaB pathway. Discov Med. 19:65–71. 2015.PubMed/NCBI
|
|
16
|
Yoon JH, Choi WS, Kim O, Choi BJ, Nam SW,
Lee JY and Park WS: Gastrokine 1 inhibits gastric cancer cell
migration and invasion by downregulating RhoA expression. Gastric
Cancer. 20:274–285. 2017. View Article : Google Scholar
|
|
17
|
Nardone G, Martin G, Rocco A, Rippa E, La
Monica G, Caruso F and Arcari P: Molecular expression of Gastrokine
1 in normal mucosa and in Helicobacter pylori-related preneoplastic
and neoplastic gastric lesions. Cancer Biol Ther. 7:1890–1895.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
He QY, Cheung YH, Leung SY, Yuen ST, Chu
KM and Chiu JF: Diverse proteomic alterations in gastric
adenocarcinoma. Proteomics. 4:3276–3287. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Moss SF, Lee JW, Sabo E, Rubin AK, Rommel
J, Westley BR, May FE, Gao J, Meitner PA, Tavares R, et al:
Decreased expression of gastrokine 1 and the trefoil factor
interacting protein TFIZ1/GKN2 in gastric cancer: Influence of
tumor histology and relationship to prognosis. Clin Cancer Res.
14:4161–4167. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yoon JH, Song JH, Zhang C, Jin M, Kang YH,
Nam SW, Lee JY and Park WS: Inactivation of the Gastrokine 1 gene
in gastric adenomas and carcinomas. J Pathol. 223:618–625. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mao W, Chen J, Peng TL, Yin XF, Chen LZ
and Chen MH: Downregulation of gastrokine-1 in gastric cancer
tissues and restoration of its expression induced gastric cancer
cells to apoptosis. J Exp Clin Cancer Res. 31:49–58. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xiao JW, Chen JH, Ren MY, Tian XB and Wang
CS: Relationship between expression of gastrokine 1 and
clinicopathological characteristics in gastric cancer patients.
Asian Pac J Cancer Prev. 13:5897–5901. 2012. View Article : Google Scholar
|
|
23
|
Choi WS, Seo HS, Song KY, Yoon JH, Kim O,
Nam SW, Lee JY and Park WS: Gastrokine 1 expression in the human
gastric mucosa is closely associated with the degree of gastritis
and DNA methylation. J Gastric Cancer. 13:232–241. 2013. View Article : Google Scholar
|
|
24
|
Guo XY, Dong L, Qin B, Jiang J and Shi AM:
Decreased expression of gastrokine 1 in gastric mucosa of gastric
cancer patients. World J Gastroenterol. 20:16702–16706. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hasan AA, Igci M, Borazan E, Khailany RA,
Bayraktar E and Arslan A: Down-regulated gene expression of GKN1
and GKN2 as diagnostic markers for gastric cancer. WASET9. 532–535.
2015.
|
|
26
|
Altieri F, Di Stadio CS, Federico A,
Miselli G, De Palma M, Rippa E and Arcari P: Epigenetic alterations
of gastrokine 1 gene expression in gastric cancer. Oncotarget.
8:16899–16911. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yang M, Jiang N, Cao QW, Ma MQ and Sun Q:
The E3 ligase UBR5 regulates gastric cancer cell growth by
destabilizing the tumor suppressor GKN1. Biochem Biophys Res
Commun. 478:1624–1629. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lu F, Wikramasinghe P, Norseen J, Tsai K,
Wang P, Showe L, Davuluri RV and Lieberman PM: Genome-wide analysis
of host-chromosome binding sites for Epstein-Barr virus nuclear
antigen 1 (EBNA1). Virol J. 7:2622010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lu F, Tempera I, Lee HT, Dewispelaere K
and Lieberman PM: EBNA1 binding and epigenetic regulation of
gastrokine tumor suppressor genes in gastric carcinoma cells. Virol
J. 11:122014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Nardone G, Rippa E, Martin G, Rocco A,
Siciliano RA, Fiengo A, Cacace G, Malorni A, Budillon G and Arcari
P: Gastrokine 1 expression in patients with and without
Helicobacter pylori infection. Dig Liver Dis. 39:122–129. 2007.
View Article : Google Scholar
|
|
31
|
Matsushima K, Isomoto H, Inoue N, Nakayama
T, Hayashi T, Nakayama M, Nakao K, Hirayama T and Kohno S: MicroRNA
signatures in Helicobacter pylori-infected gastric mucosa. Int J
Cancer. 128:361–370. 2011. View Article : Google Scholar
|
|
32
|
Lario S, Ramírez-Lázaro MJ, Aransay AM,
Lozano JJ, Montserrat A, Casalots Á, Junquera F, Álvarez J, Segura
F, Campo R, et al: microRNA profiling in duodenal ulcer disease
caused by Helicobacter pylori infection in a Western population.
Clin Microbiol Infect. 18:E273–E282. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chang H, Kim N, Park JH, Nam RH, Choi YJ,
Lee HS, Yoon H, Shin CM, Park YS, Kim JM, et al: Different microRNA
expression levels in gastric cancer depending on Helicobacter
pylori infection. Gut Liver. 9:188–196. 2015. View Article : Google Scholar :
|
|
34
|
Zhu Y, Jiang Q, Lou X, Ji X, Wen Z, Wu J,
Tao H, Jiang T, He W, Wang C, et al: MicroRNAs up-regulated by CagA
of Helicobacter pylori induce intestinal metaplasia of gastric
epithelial cells. PLoS One. 7:e351472012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Santos JC, Brianti MT, Almeida VR, Ortega
MM, Fischer W, Haas R, Matheu A and Ribeiro ML: Helicobacter pylori
infection modulates the expression of miRNAs associated with DNA
mismatch repair pathway. Mol Carcinog. 56:1372–1379. 2017.
View Article : Google Scholar
|
|
36
|
Chung JW, Jeong SH, Lee SM, Pak JH, Lee
GH, Jeong JY and Kim JH: Expression of microRNA in host cells
infected with Helicobacter pylori. Gut Liver. 11:392–400. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sugihara H, Ishimoto T, Watanabe M,
Sawayama H, Iwatsuki M, Baba Y, Komohara Y, Takeya M and Baba H:
Identification of miR-30e* regulation of Bmi1 expression
mediated by tumor-associated macrophages in gastrointestinal
cancer. PLoS One. 8:e818392013. View Article : Google Scholar
|
|
38
|
Stumpfova Z, Hezova R, Meli AC, Slaby O
and Michalek J: MicroRNA profiling of activated and tolerogenic
human dendritic cells. Mediators Inflamm. 2014:259689–259699. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Teteloshvili N, Smigielska-Czepiel K,
Kroesen BJ, Brouwer E, Kluiver J, Boots AM and van den Berg A:
T-cell activation induces dynamic changes in miRNA expression
patterns in CD4 and CD8 T-cell subsets. MicroRNA. 4:117–122. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Sánchez-Pulido L, Devos D and Valencia A:
BRICHOS: A conserved domain in proteins associated with dementia,
respiratory distress and cancer. Trends Biochem Sci. 27:329–332.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hedlund J, Johansson J and Persson B:
BRICHOS - a super-family of multidomain proteins with diverse
functions. BMC Res Notes. 2:180–189. 2009. View Article : Google Scholar
|
|
42
|
Pavone LM, Del Vecchio P, Mallardo P,
Altieri F, De Pasquale V, Rea S, Martucci NM, Di Stadio CS, Pucci
P, Flagiello A, et al: Structural characterization and biological
properties of human gastrokine 1. Mol Biosyst. 9:412–421. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Yoon JH, Choi YJ, Choi WS, Nam SW, Lee JY
and Park WS: Functional analysis of the NH2-terminal hydrophobic
region and BRICHOS domain of GKN1. Biochem Biophys Res Commun.
440:689–695. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Dokhaee F, Mazhari S, Galehdari M,
Bahadori Monfared A and Baghaei K: Evaluation of GKN1 and GKN2 gene
expression as a biomarker of gastric cancer. Gastroenterol Hepatol
Bed Bench. 11(Suppl 1): S140–S145. 2018.
|
|
45
|
Toback FG, Walsh-Reitz MM, Musch MW, Chang
EB, Del Valle J, Ren H, Huang E and Martin TE: Peptide fragments of
AMP-18, a novel secreted gastric antrum mucosal protein, are
mitogenic and motogenic. Am J Physiol Gastrointest Liver Physiol.
285:G344–G353. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Xing R, Li W, Cui J, Zhang J, Kang B, Wang
Y, Wang Z, Liu S and Lu Y: Gastrokine 1 induces senescence through
p16/Rb pathway activation in gastric cancer cells. Gut. 61:43–52.
2012. View Article : Google Scholar
|
|
47
|
Conteduca V, Sansonno D, Lauletta G, Russi
S, Ingravallo G and Dammacco F: H. pylori infection and gastric
cancer: State of the art (review). Int J Oncol. 42:5–18. 2013.
View Article : Google Scholar
|
|
48
|
Yoon JH, Seo HS, Choi SS, Chae HS, Choi
WS, Kim O, Ashktorab H, Smoot DT, Nam SW, Lee JY, et al: Gastrokine
1 inhibits the carcinogenic potentials of Helicobacter pylori CagA.
Carcinogenesis. 35:2619–2629. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Yoshikawa Y, Mukai H, Hino F, Asada K and
Kato I: Isolation of two novel genes, down-regulated in gastric
cancer. Jpn J Cancer Res. 91:459–463. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Shiozaki K, Nakamori S, Tsujie M, Okami J,
Yamamoto H, Nagano H, Dono K, Umeshita K, Sakon M, Furukawa H, et
al: Human stomach-specific gene, CA11, is down-regulated in gastric
cancer. Int J Oncol. 19:701–707. 2001.PubMed/NCBI
|
|
51
|
Oien KA, Vass JK, Downie I, Fullarton G
and Keith WN: Profiling, comparison and validation of gene
expression in gastric carcinoma and normal stomach. Oncogene.
22:4287–4300. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Oien KA, McGregor F, Butler S, Ferrier RK,
Downie I, Bryce S, Burns S and Keith WN: Gastrokine 1 is abundantly
and specifically expressed in superficial gastric epithelium,
down-regulated in gastric carcinoma, and shows high evolutionary
conservation. J Pathol. 203:789–797. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Koper-Lenkiewicz OM, Kamińska J, Gawrońska
B and Matowicka-Karna J: The role and diagnostic potential of
gastrokine 1 in gastric cancer. Cancer Manag Res. 11:1921–1931.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Zamanian-Azodi M, Rezaei-Tavirani M,
Hasanzadeh H, Rahmati Rad S and Dalilan S: Introducing biomarker
panel in esophageal, gastric, and colon cancers; a proteomic
approach. Gastroenterol Hepatol Bed Bench. 8:6–18. 2015.PubMed/NCBI
|
|
55
|
Villano V, Di Stadio CS, Federico A,
Altieri F, Miselli G, De Palma M, Rippa E and Arcari P: Gastrokine
1 mRNA in human sera is not informative biomarker for gastric
cancer. J Negat Results Biomed. 15:142016. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Yoon JH, Ham IH, Kim O, Ashktorab H, Smoot
DT, Nam SW, Lee JY, Hur H and Park WS: Gastrokine 1 protein is a
potential theragnostic target for gastric cancer. Gastric Cancer.
21:956–967. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Noguchi T, Wirtz HC, Michaelis S, Gabbert
HE and Mueller W: Chromosomal imbalances in gastric cancer.
Correlation with histologic subtypes and tumor progression. Am J
Clin Pathol. 115:828–834. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Panani AD: Cytogenetic and molecular
aspects of gastric cancer: Clinical implications. Cancer Lett.
266:99–115. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Orphanides G and Reinberg D: A unified
theory of gene expression. Cell. 108:439–451. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Jaenisch R and Bird A: Epigenetic
regulation of gene expression: How the genome integrates intrinsic
and environmental signals. Nat Genet. 33(Suppl): 245–254. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Shilatifard A: Chromatin modifications by
methylation and ubiquitination: Implications in the regulation of
gene expression. Annu Rev Biochem. 75:243–269. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Catalanotto C, Cogoni C and Zardo G:
MicroRNA in control of gene expression: An overview of nuclear
functions. Int J Mol Sci. 17:17122016. View Article : Google Scholar :
|
|
63
|
Levine M and Tjian R: Transcription
regulation and animal diversity. Nature. 424:147–151. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Lambert SA, Jolma A, Campitelli LF, Das
PK, Yin Y, Albu M, Chen X, Taipale J, Hughes TR and Weirauch MT:
The human transcription factors. Cell. 175:598–599. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Yoon JH, Choi WS, Kim O, Choi SS, Lee EK,
Nam SW, Lee JY and Park WS: NKX6.3 controls gastric differentiation
and tumorigenesis. Oncotarget. 6:28425–28439. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Cartharius K, Frech K, Grote K, Klocke B,
Haltmeier M, Klingenhoff A, Frisch M, Bayerlein M and Werner T:
MatInspector and beyond: Promoter analysis based on transcription
factor binding sites. Bioinformatics. 21:2933–2942. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Grabe N: AliBaba2: Context specific
identification of transcription factor binding sites. In Silico
Biol. 2:S1–S15. 2002.PubMed/NCBI
|
|
68
|
Ghosh D: Object-oriented transcription
factors database (ooTFD). Nucleic Acids Res. 28:308–310. 2000.
View Article : Google Scholar
|
|
69
|
Strowski MZ, Cramer T, Schäfer G, Jüttner
S, Walduck A, Schipani E, Kemmner W, Wessler S, Wunder C, Weber M,
et al: Helicobacter pylori stimulates host vascular endothelial
growth factor-A (vegf-A) gene expression via MEK/ERK-dependent
activation of Sp1 and Sp3. FASEB J. 18:218–220. 2004. View Article : Google Scholar
|
|
70
|
Mitsuno Y, Yoshida H, Maeda S, Ogura K,
Hirata Y, Kawabe T, Shiratori Y and Omata M: Helicobacter pylori
induced transactivation of SRE and AP-1 through the ERK signalling
pathway in gastric cancer cells. Gut. 49:18–22. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Han JC, Zhang KL, Chen XY, Jiang HF, Kong
QY, Sun Y, Wu ML, Huang L, Li H and Liu J: Expression of seven
gastric cancer-associated genes and its relevance for Wnt,
NF-kappaB and Stat3 signaling. APMIS. 115:1331–1343. 2007.
View Article : Google Scholar
|
|
72
|
Xiong H, Du W, Sun TT, Lin YW, Wang JL,
Hong J and Fang JY: A positive feedback loop between STAT3 and
cyclooxygenase-2 gene may contribute to Helicobacter
pylori-associated human gastric tumorigenesis. Int J Cancer.
134:2030–2040. 2014. View Article : Google Scholar
|
|
73
|
Hu TZ, Huang LH, Xu CX, Liu XM, Wang Y,
Xiao J, Zhou L, Luo L and Jiang XX: Expressional profiles of
transcription factors in the progression of Helicobacter
pylori-associated gastric carcinoma based on protein/DNA array
analysis. Med Oncol. 32:2652015. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Liu X, Cao K, Xu C, Hu T, Zhou L, Cao D,
Xiao J, Luo L, Guo Y and Qi Y: GATA-3 augmentation down-regulates
Connexin43 in Helicobacter pylori associated gastric
carcinogenesis. Cancer Biol Ther. 16:987–996. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Qian J, Kong X, Deng N, Tan P, Chen H,
Wang J, Li Z, Hu Y, Zou W, Xu J, et al: OCT1 is a determinant of
synbindin-related ERK signalling with independent prognostic
significance in gastric cancer. Gut. 64:37–48. 2015. View Article : Google Scholar :
|
|
76
|
Xu G, Li K, Zhang N, Zhu B and Feng G:
Screening driving transcription factors in the processing of
gastric cancer. Gastroenterol Res Pract. 2016:84314802016.
View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Shakya A, Cooksey R, Cox JE, Wang V,
McClain DA and Tantin D: Oct1 loss of function induces a coordinate
metabolic shift that opposes tumorigenicity. Nat Cell Biol.
11:320–327. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Kong Y, Ma LQ, Bai PS, Da R, Sun H, Qi XG,
Ma JQ, Zhao RM, Chen NZ and Nan KJ: Helicobacter pylori promotes
invasion and metastasis of gastric cancer cells through activation
of AP-1 and up-regulation of CACUL1. Int J Biochem Cell Biol.
45:2666–2678. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Regalo G, Resende C, Wen X, Gomes B,
Durães C, Seruca R, Carneiro F and Machado JC: C/EBP α expression
is associated with homeostasis of the gastric epithelium and with
gastric carcinogenesis. Lab Invest. 90:1132–1139. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Jackson CB, Judd LM, Menheniott TR,
Kronborg I, Dow C, Yeomans ND, Boussioutas A, Robb L and Giraud AS:
Augmented gp130-mediated cytokine signalling accompanies human
gastric cancer progression. J Pathol. 213:140–151. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
O'Reilly LA, Putoczki TL, Mielke LA, Low
JT, Lin A, Preaudet A, Herold MJ, Yaprianto K, Tai L, Kueh A, et
al: Loss of NF-κB1 causes gastric cancer with aberrant inflammation
and expression of immune checkpoint regulators in a STAT-1
dependent manner. Immunity. 48:570–583.e8. 2018. View Article : Google Scholar
|
|
82
|
Zhang J, Zhu ZG, Ji J, Yuan F, Yu YY, Liu
BY and Lin YZ: Transcription factor Sp1 expression in gastric
cancer and its relationship to long-term prognosis. World J
Gastroenterol. 11:2213–2217. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Jüttner S, Cramer T, Wessler S, Walduck A,
Gao F, Schmitz F, Wunder C, Weber M, Fischer SM, Schmidt WE, et al:
Helicobacter pylori stimulates host cyclooxygenase-2 gene
transcription: Critical importance of MEK/ERK-dependent activation
of USF1/-2 and CREB transcription factors. Cell Microbiol.
5:821–834. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Lu H, Wu JY, Kudo T, Ohno T, Graham DY and
Yamaoka Y: Regulation of interleukin-6 promoter activation in
gastric epithelial cells infected with Helicobacter pylori. Mol
Biol Cell. 16:4954–4966. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Bronte-Tinkew DM, Terebiznik M, Franco A,
Ang M, Ahn D, Mimuro H, Sasakawa C, Ropeleski MJ, Peek RM Jr and
Jones NL: Helicobacter pylori cytotoxin-associated gene A activates
the signal transducer and activator of transcription 3 pathway in
vitro and in vivo. Cancer Res. 69:632–639. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Zhao J, Dong Y, Kang W, Go MY, Tong JH, Ng
EK, Chiu PW, Cheng AS, To KF, Sung JJ, et al: Helicobacter
pylori-induced STAT3 activation and signalling network in gastric
cancer. Oncoscience. 1:468–475. 2014. View Article : Google Scholar
|
|
87
|
Piao JY, Lee HG, Kim SJ, Kim DH, Han HJ,
Ngo HK, Park SA, Woo JH, Lee JS, Na HK, et al: Helicobacter pylori
activates IL-6-STAT3 signaling in human gastric cancer cells:
Potential roles for reactive oxygen species. Helicobacter.
21:405–416. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Yokoyama K, Higashi H, Ishikawa S, Fujii
Y, Kondo S, Kato H, Azuma T, Wada A, Hirayama T, Aburatani H, et
al: Functional antagonism between Helicobacter pylori CagA and
vacuolating toxin VacA in control of the NFAT signaling pathway in
gastric epithelial cells. Proc Natl Acad Sci USA. 102:9661–9666.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Chen G, Tang N, Wang C, Xiao L, Yu M, Zhao
L, Cai H, Han L, Xie C and Zhang Y: TNF-α-inducing protein of
Helicobacter pylori induces epithelial-mesenchymal transition (EMT)
in gastric cancer cells through activation of IL-6/STAT3 signaling
pathway. Biochem Biophys Res Commun. 484:311–317. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Mejías-Luque R, Peiró S, Vincent A, Van
Seuningen I and de Bolós C: IL-6 induces MUC4 expression through
gp130/STAT3 p athway in gastric cancer cell lines. Biochim Biophys
Acta. 1783:1728–1736. 2008. View Article : Google Scholar
|
|
91
|
Chang YJ, Wu MS, Lin JT and Chen CC:
Helicobacter pylori-Induced invasion and angiogenesis of gastric
cells is mediated by cyclooxy-genase-2 induction through TLR2/TLR9
and promoter regulation. J Immunol. 175:8242–8252. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Lee KS, Kalantzis A, Jackson CB, O'Connor
L, Murata-Kamiya N, Hatakeyama M, Judd LM, Giraud AS and Menheniott
TR: Helicobacter pylori CagA triggers expression of the
bactericidal lectin REG3γ via gastric STAT3 activation. PLoS One.
7:e307862012. View Article : Google Scholar
|
|
93
|
Yamaoka Y, Kudo T, Lu H, Casola A, Brasier
AR and Graham DY: Role of interferon-stimulated responsive
element-like element in interleukin-8p romoter in Helicobacter
pylori infection. Gastroenterology. 126:1030–1043. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Mitchell DJ, Huynh HQ, Ceponis PJM, Jones
NL and Sherman PM: Helicobacter pylori disrupts STAT1-mediated
gamma interferon-induced signal transduction in epithelial cells.
Infect Immun. 72:537–545. 2004. View Article : Google Scholar :
|
|
95
|
Lee HS, Park CK, Oh E, Erkin ÖC, Jung HS,
Cho MH, Kwon MJ, Chae SW, Kim SH, Wang LH, et al: Low SP1
expression differentially affects intestinal-type compared with
diffuse-type gastric adenocarcinoma. PLoS One. 8:e555222013.
View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Beishline K and Azizkhan-Clifford J: Sp1
and the 'hallmarks of cancer'. FEBS J. 282:224–258. 2015.
View Article : Google Scholar
|
|
97
|
Tomizawa M, Shinozaki F, Motoyoshi Y,
Sugiyama T, Yamamoto S and Ishige N: CCAAT/enhancer-binding protein
α decreases the viability of gastric cancer cells. Oncol Lett.
13:4322–4326. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Peterson CL and Laniel MA: Histones and
histone modifications. Curr Biol. 14:R546–R551. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Agarwal V, Bell GW, Nam JW and Bartel DP:
Predicting effective microRNA target sites in mammalian mRNAs.
eLife. 4:e050052015. View Article : Google Scholar :
|
|
100
|
John B, Enright AJ, Aravin A, Tuschl T,
Sander C and Marks DS: Human microRNA targets. PLoS Biol.
3:e2642005. View Article : Google Scholar
|
|
101
|
Wong N and Wang X: miRDB: An online
resource for microRNA target prediction and functional annotations.
Nucleic Acids Res. 43D:D146–D152. 2015. View Article : Google Scholar
|
|
102
|
Lu TP, Lee CY, Tsai MH, Chiu YC, Hsiao CK,
Lai LC and Chuang EY: miRSystem: An integrated system for
characterizing enriched functions and pathways of microRNA targets.
PLoS One. 7:e423902012. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Maragkakis M, Reczko M, Simossis VA,
Alexiou P, Papadopoulos GL, Dalamagas T, Giannopoulos G, Goumas G,
Koukis E, Kourtis K, et al: DIANA-microT web server: Elucidating
microRNA functions through target prediction. Nucleic Acids Res.
37(Web Server): W273–6. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Sethupathy P, Megraw M and Hatzigeorgiou
AG: A guide through present computational approaches for the
identification of mammalian microRNA targets. Nat Methods.
3:881–886. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Leitão AL, Costa MC and Enguita FJ: A
guide for miRNA target prediction and analysis using web-based
applications. Methods Mol Biol. 1182:265–277. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Zhi Q, Guo X, Guo L, Zhang R, Jiang J, Ji
J, Zhang J, Zhang J, Chen X, Cai Q, et al: Oncogenic miR-544 is an
important molecular target in gastric cancer. Anticancer Agents Med
Chem. 13:270–275. 2013. View Article : Google Scholar
|
|
107
|
Chaturvedi R, de Sablet T, Asim M,
Piazuelo MB, Barry DP, Verriere TG, Sierra JC, Hardbower DM,
Delgado AG, Schneider BG, et al: Increased Helicobacter
pylori-associated gastric cancer risk in the Andean region of
Colombia is mediated by spermine oxidase. Oncogene. 34:3429–3440.
2015. View Article : Google Scholar :
|
|
108
|
Ishimoto T, Sugihara H, Watanabe M,
Sawayama H, Iwatsuki M, Baba Y, Okabe H, Hidaka K, Yokoyama N,
Miyake K, et al: Macrophage-derived reactive oxygen species
suppress miR-328 targeting CD44 in cancer cells and promote redox
adaptation. Carcinogenesis. 35:1003–1011. 2014. View Article : Google Scholar
|
|
109
|
Libânio D, Dinis-Ribeiro M and
Pimentel-Nunes P: Helicobacter pylori and microRNAs: Relation with
innate immunity and progression of preneoplastic conditions. World
J Clin Oncol. 6:111–132. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Zhang X, Peng Y, Jin Z, Huang W, Cheng Y,
Liu Y, Feng X, Yang M, Huang Y, Zhao Z, et al: Integrated miRNA
profiling and bioinformatics analyses reveal potential causative
miRNAs in gastric adenocarcinoma. Oncotarget. 6:32878–32889. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Zhang W, Dahlberg JE and Tam W: MicroRNAs
in tumori-genesis: A primer. Am J Pathol. 171:728–738. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Noto JM and Peek RM: The role of microRNAs
in Helicobacter pylori pathogenesis and gastric carcinogenesis.
Front Cell Infect Microbiol. 1:212012. View Article : Google Scholar :
|
|
113
|
Li N, Xu X, Xiao B, Zhu ED, Li BS, Liu Z,
Tang B, Zou QM, Liang HP and Mao XH: H. pylori related
proinflammatory cytokines contribute to the induction of miR-146a
in human gastric epithelial cells. Mol Biol Rep. 39:4655–4661.
2012. View Article : Google Scholar
|
|
114
|
Hayashi Y, Tsujii M, Wang J, Kondo J,
Akasaka T, Jin Y, Li W, Nakamura T, Nishida T, Iijima H, et al:
CagA mediates epigenetic regulation to attenuate let-7 expression
in Helicobacter pylori-related carcinogenesis. Gut. 62:1536–1546.
2013. View Article : Google Scholar
|
|
115
|
Qi J and Ronai ZA: Dysregulation of
ubiquitin ligases in cancer. Drug Resist Updat. 23:1–11. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Cho CH, Yu J and Wu WKK: Identification of
pathogenic microRNAs in Helicobacter pylori-associated gastric
cancer using a combined approach of animal study and clinical
sample analysis. Hong Kong Med J. 22(Suppl 6): 13–18.
2016.PubMed/NCBI
|
|
117
|
Belair C, Baud J, Chabas S, Sharma CM,
Vogel J, Staedel C and Darfeuille F: Helicobacter pylori interferes
with an embryonic stem cell micro RNA cluster to block cell cycle
progression. Silence. 2:72011. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Lauren P: The two histological main types
of gastric carcinoma: Diffuse and so-called intestinal-type
carcinoma. An attempt at a histo-clinical classification. Acta
Pathol Microbiol Scand. 64:31–49. 1965. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Yu BQ, Su LP, Li JF, Cai Q, Yan M, Chen
XH, Yu YY, Gu QL, Zhu ZG and Liu BY: microrna expression signature
of gastric cancer cells relative to normal gastric mucosa. Mol Med
Rep. 6:821–826. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Guo J, Miao Y, Xiao B, Huan R, Jiang Z,
Meng D and Wang Y: Differential expression of microRNA species in
human gastric cancer versus non-tumorous tissues. J Gastroenterol
Hepatol. 24:652–657. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Ueda T, Volinia S, Okumura H, Shimizu M,
Taccioli C, Rossi S, Alder H, Liu CG, Oue N, Yasui W, et al:
Relation between microRNA expression and progression and prognosis
of gastric cancer: A microRNA expression analysis. Lancet Oncol.
11:136–146. 2010. View Article : Google Scholar
|
|
122
|
Tsukamoto Y, Nakada C, Noguchi T, Tanigawa
M, Nguyen LT, Uchida T, Hijiya N, Matsuura K, Fujioka T, Seto M, et
al: MicroRNA-375 is downregulated in gastric carcinomas and
regulates cell survival by targeting PDK1 and 14-3-3zeta. Cancer
Res. 70:2339–2349. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Su Y, Ni Z, Wang G, Cui J, Wei C, Wang J,
Yang Q, Xu Y and Li F: Aberrant expression of microRNAs in gastric
cancer and biological significance of miR-574-3p. Int
Immunopharmacol. 13:468–475. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Juzėnas S, Saltenienė V, Kupcinskas J,
Link A, Kiudelis G, Jonaitis L, Jarmalaite S, Kupcinskas L,
Malfertheiner P and Skieceviciene J: Correction: Analysis of
deregulated microRNAs and their target genes in gastric cancer.
PLoS One. 10:e01357622015. View Article : Google Scholar
|
|
125
|
Katada T, Ishiguro H, Kuwabara Y, Kimura
M, Mitui A, Mori Y, Ogawa R, Harata K and Fujii Y: MicroRNA
expression profile in undifferentiated gastric cancer. Int J Oncol.
34:537–542. 2009.PubMed/NCBI
|














