Introduction
Lung cancer is a leading cause of cancer morbidity
(11.6% in 2018) and cancer-related mortality (18.4% in 2018)
worldwide (1); it induces a number
of symptoms, such as cough and dyspnea, and occlusion and
cavitation by tumors sometimes cause severe lung infections
(2-5). Non-small cell lung carcinoma (NSCLC)
accounts for ~85% of all cases of lung cancer, of which 20-30% are
squamous cell lung carcinoma (SQCLC) (6). Although molecular targeted therapies
have markedly prolonged the survival of patients with lung
adenocarcinoma, there are currently no developed effective targeted
therapies for SQCLC. Amplification of the fibroblast growth factor
receptor 1 (FGFR1) gene is one of the most common oncogenic
alternations in SQCLC. Although clinical trials on FGFR inhibitors
for patients with FGFR-amplified SQCLC have been conducted,
the overall response rates were only 8-11% (7,8).
Therefore, the prognosis of patients with SQCLC remains poor and
treatment options are limited, and further research on the
development of novel effective therapies for SQCLC is needed.
The third-generation anthracycline amrubicin is a
potent topoisomerase II inhibitor that is approved in Japan for the
treatment of NSCLC and small cell lung carcinoma (SCLC). A
randomized phase III study comparing amrubicin and docetaxel (DOC)
treatment in patients with previously treated NSCLC was conducted,
and the median progression-free survival was 3.6 months in the
amrubicin group and 3.0 months in the DOC group (P=0.54) (9). Although this study was unable to
demonstrate the superiority of amrubicin over DOC, amrubicin is
still regarded as one of the treatment options for patients with
previously treated NSCLC in Japan.
Histone deacetylases (HDACs) are enzymes that serve
important roles in changing epigenetic conditions and regulating
gene expression (10). The strong
expression of HDACs has been reported in various cancers, including
lung cancer, and HDAC inhibitors block the proliferation of various
lung cancer cell lines (11-13).
Although HDAC inhibition is expected to become a new prospective
treatment for lung cancer, HDAC inhibitor monotherapies for
patients with NSCLC have failed to exhibit clinical efficacy in
clinical trials (14,15). Therefore, several clinical trials
on combination therapies with HDAC inhibitors and cytotoxic drugs
have been conducted, but failed to demonstrate the efficacy and
safety of these therapies (16,17).
OBP-801, also known as YM753, is one of the most potent HDAC
inhibitors that was discovered in our previous study by screening
for cyclin-dependent kinase inhibitor
p21WAF1/Cip1-inducing agents (18). OBP-801 is currently in a clinical
trial in the USA and is a promising HDAC inhibitor.
To develop a novel combination therapy with the HDAC
inhibitor OBP-801 against SQCLC, cytotoxic drugs approved for the
treatment of SQCLC were screened and it was revealed that
co-treatment with OBP-801 and amrubicin synergistically inhibited
the viability of human SQCLC cells by inducing apoptosis. Moreover,
the combined treatment was effective in H520 SQCLC xenograft model
mice. The combined treatment with OBP-801 and amrubicin may have
potential as a treatment option for patients with SQCLC.
Materials and methods
Reagents
Amrubicin was purchased from APExBIO Technology LLC,
and OBP-801 was from Oncolys BioPharma Inc. Doxorubicin
hydrochloride was purchased from FUJIFILM Wako Pure Chemical
Corporation. Selonsertib, SB203580 and SP600125 were purchased from
Selleck Chemicals. The pan-caspase inhibitor Z-VAD-FMK was
purchased from R&D Systems, Inc. These agents were dissolved in
DMSO for in vitro experiments. N-acetyl-L-cysteine (NAC) was
purchased from Nacalai Tesque, Inc.
Lung cancer cell lines and cell
cultures
The human SQCLC cell lines Calu-1 and H520, and the
lung adenocarcinoma cell line A549 were obtained from the American
Type Culture Collection. Calu-1 and A549 cells were cultured in
DMEM (Nissui Pharmaceutical Co., Ltd.) supplemented with 10% FBS
(Sigma-Aldrich; Merck KGaA), 4 mM glutamine, 50 U/ml penicillin and
100 µg/ml streptomycin at 37°C in 5% CO2. H520
cells were cultured in RPMI-1640 medium with 10% FBS, 2 mM
glutamine, 50 U/ml penicillin and 100 µg/ml streptomycin at
37°C in 5% CO2.
Treatments with agents
For cell viability assay, Calu-1 (1×103
cells/well), A549 (1×103 cells/well) and H520 cells
(2×103 cells/well) in a 96-well plate were treated with
various concentrations of OBP-801 or amrubicin, or OBP-801 (Calu-1,
2.75 nM; A549, 2.5 nM; H520, 4.5 nM) with or without amrubicin
(Calu-1, 2 µM; A549, 400 nM; H520, 4 µM) at 37°C for
72 h. For quantification of apoptosis, Calu-1 (5×104
cells/well), A549 (2×104 cells/well) and H520 cells
(1.5×105 cells/well) in a 6-well plate were treated with
OBP-801 (Calu-1, 2.75 nM; A549, 2.5 nM; H520, 4.5 nM) with or
without amrubicin (Calu-1, 2 µM; A549, 400 nM; H520, 4
µM) in the presence or absence of 5 mM NAC or 20 µM
Z-VAD-FMK at 37°C for 72 h. In addition, Calu-1 cells
(5×104 cells/well in a 6-well plate) were also treated
with 2.75 nM OBP-801 with or without 2 µM amrubicin in the
presence or absence of 50 µM selonsertib, 50 µM
SB203580 or 50 µM SP600125 at 37°C for 72 h, after which
apoptosis was analyzed. For western blot analysis, Calu-1 cells
(3×105 cells/10-cm dish) were treated with 2.75 nM
OBP-801 with or without 2 µM amrubicin at 37°C for 72 h. For
measuring intracellular ROS, Calu-1 cells (5×104
cells/well in a 6-well plate) were treated with 2.75 nM OBP-801
with or without 2 µM amrubicin at 37°C for 48 h.
Cell viability assay
Following the various treatments of Calu-1, A549 and
H520 cells, viability was evaluated using Cell Counting Kit-8
(CCK-8; Dojindo Molecular Technologies, Inc.), as previously
described (19).
Quantification of apoptosis
Treated Calu-1, A549 and H520 cells were stained for
1 min at room temperature with 50 µg/ml propidium iodide.
The cells were analyzed using a BD FACSCalibur (Becton, Dickinson
and Company), and the degree of apoptosis was determined by
quantifying the sub-G1 population (the left side of the G1 peak)
using BD CellQuest Pro software (version 6.0; Becton, Dickinson and
Company), as previously described (19).
Western blot analysis
Treated Calu-1 cells were lysed with RIPA buffer (50
mM Tris-HCl, pH 8.0; 150 mM NaCl; 1% NP-40; 0.5% deoxycholic acid;
0.1% SDS; 1 mM dithiothreitol; and 0.5 mM phenylmethylsulfonyl
fluoride) for 30 min at 4°C, and western blotting was performed as
previously described (19). Total
protein concentration was measured using Bio-Rad Protein Assay Dye
Reagent Concentrate (Bio-Rad Laboratories, Inc.). A total of 5-20
µg of protein was separated by 12% SDS-PAGE. The proteins
were subsequently transferred to Immobilon-P membranes (Millipore;
Merck KGaA), which were then blocked in Tris-buffered saline
containing 5% skim milk for 1 h at room temperature. The membranes
were incubated with the following primary antibodies for 1 h at
room temperature: Anti-thioredoxin-interacting protein (TXNIP;
1:500; cat. no. ab188865; Abcam), anti-thioredoxin 2 (Trx2; 1:500;
cat. no. sc-133201; Santa Cruz Biotechnology, Inc.), anti-cleaved
poly (ADP-ribose) polymerase (Asp214) (PARP; 1:1,000; cat. no.
5625; Cell Signaling Technology, Inc.) and anti-β-actin (1:2,000;
cat. no. A5441; Sigma-Aldrich; Merck KGaA). Membranes were then
incubated with the following secondary antibodies for 1 h at room
temperature: Horseradish peroxidase (HRP)-conjugated sheep
anti-mouse IgG (1:2,000; cat. no. NA931; GE Healthcare) or
HRP-conjugated donkey anti-rabbit IgG (1:2,000; cat. no. NA934; GE
Healthcare). Proteins were visualized with Chemi-Lumi One L
(Nacalai Tesque) or Immobilon Western Chemiluminescent HRP
Substrate (Millipore; Merck KGaA) and detected on BioMax XAR film
(Carestream Health, Inc.).
Measurement of intercellular ROS
Treated Calu-1 cells were incubated with 5 µM
CellROX Deep Red Reagent (Thermo Fisher Scientific, Inc.) for 30
min at 37°C. Fluorescence signals were measured in the FL-4 channel
by FACSCalibur and BD CellQuest Pro software (version 6.0).
siRNA transfection
The following siRNAs targeting Trx2 were purchased
from Thermo Fisher Scientific, Inc.; only sense strands are shown:
siTrx2#1, 5′-CCC GGA CAA UAU ACA CCA CGA GGA U-3′; siTrx2#2, 5′-CCA
CAC AGA CCU CGC CAU UGA GUA U-3′; and siTrx2#3, 5′-GCC UUC CUGAAG
AAG CUG AUU GGC U-3′; and a negative control siRNA (cat. no.
12935-113). Calu-1 cells (3×104 cells/well in a 6-well
plate) were transfected with each siRNA (30 pmol) at 37°C using
Lipofectamine® RNAiMAX (Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol. After 5 h, the medium was
replaced. A total of 48 h post-transfection, the cells were lysed
with RIPA buffer, and the knockdown efficiency of each siRNA was
confirmed by western blotting. At 120 h post-transfection, the
proportion of apoptotic cells (the sub-G1 population) was analyzed
using FACSCalibur and BD CellQuest Pro software (version 6.0)
aforementioned.
Mouse xenograft model
Female BALB/c nu/nu mice (n=20; age, 5 weeks;
weight, 16.85-21.78 g) were purchased from Charles River
Laboratories, Inc. The mice were housed in cages at 24±2°C and
45±5% humidity under pathogen-free conditions and fed CLEA rodent
diet CE-2 commercial pellets (CLEA Japan, Inc.) and tap water ad
libitum. All experiments were performed in accordance with the
institutional animal care and use committee guidelines, and the
present study was approved by the Committee for Animal Research of
Kyoto Prefectural University of Medicine (permission no. M29-576).
A total of 4×106 untreated H520 cells mixed with
Matrigel (BD Biosciences) were injected subcutaneously into the
right flank of mice. Tumor volumes were calculated using the
following formula: ½ x length x width2. When tumor
volumes reached 40 mm3 on average on day 13, mice were
randomized into four groups (n=5 mice per group) and treatments
were initiated. OBP-801 was dissolved in saline containing 20%
hydroxypropyl-β-cyclodextrin, and mice were injected through the
tail vein once a week (on days 15, 22, 29 and 36) with diluent or
OBP-801 (10 mg/kg). Amrubicin hydrochloride (Nippon Kayaku Co.,
Ltd.) was dissolved in saline, and mice were injected through the
tail vein only once (on day 14) with diluent or amrubicin (25
mg/kg). The concentrations of OBP-801 and amrubicin used were based
on the results of pre-experiments (data not shown). Tumor sizes
were measured twice per week, and the experiment was finished on
day 72. The humane endpoints established for this study were mice
exhibiting abnormalities such as a sudden weight loss of ≥20%, or
tumor size in a single mouse exceeds 2,000 mm3. The mice
were euthanized by intraperitoneal injection of 200 mg/kg
pentobarbital.
Statistical analysis
Data are expressed as the mean ± SD of three
measurements. Statistical analyses were performed using ANOVA
followed by Bonferroni's post hoc test. Combination index (CI)
values were calculated using CalcuSyn software (version 2.0;
Biosoft, Cambridge, UK); a combination was judged to be synergistic
when CI <1.0. P<0.05 was considered to indicate a
statistically significant difference.
Results
Combined OBP-801 and amrubicin treatment
synergistically inhibits human lung cancer cell line viability
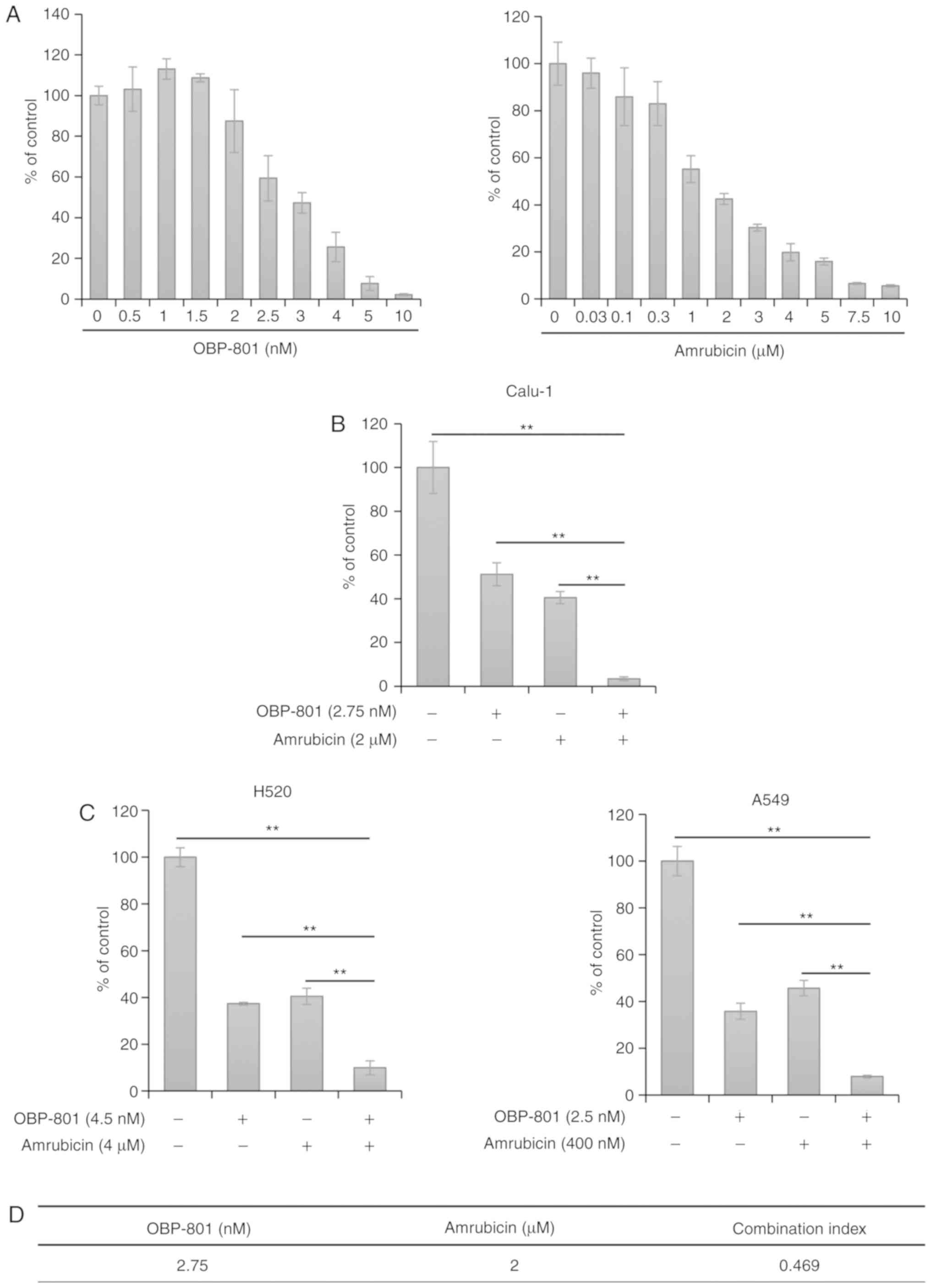
To evaluate the effects of combined treatments with
the potent HDAC inhibitor OBP-801, the effects of OBP-801 or
amrubicin alone on the viability of human SQCLC Calu-1 cells were
examined. Each agent inhibited the viability of Calu-1 cells in a
dose-dependent manner (Fig. 1A).
The effects of several combinations were then analyzed by combining
OBP-801 and amrubicin at various concentrations near the
IC50; the combined treatment with OBP-801 and amrubicin
more strongly inhibited the viability of Calu-1 cells than the
treatment with each agent alone (Fig.
1B). Based on the viability inhibition curves concerning human
SQCLC H520 cells (Fig. S1A) and
human lung adenocarcinoma A549 cells (Fig. S1B), the effects of several
combinations were also examined by combining OBP-801 and amrubicin
at various concentrations near the IC50, and similar
results were obtained using H520 and A549 cells (Fig. 1C). Moreover, OBP-801 and another
anthracycline, doxorubicin, also coordinately inhibited the
viability of Calu-1 cells (Fig.
S2). The CI value for the combination of OBP-801 and amrubicin
against Calu-1 cells was markedly <1.0 (Fig. 1D), which indicated synergistic
inhibition against the viability of Calu-1 cells.
Co-treatment with OBP-801 and amrubicin
induces caspase-dependent and ROS-dependent apoptosis in lung
cancer cells
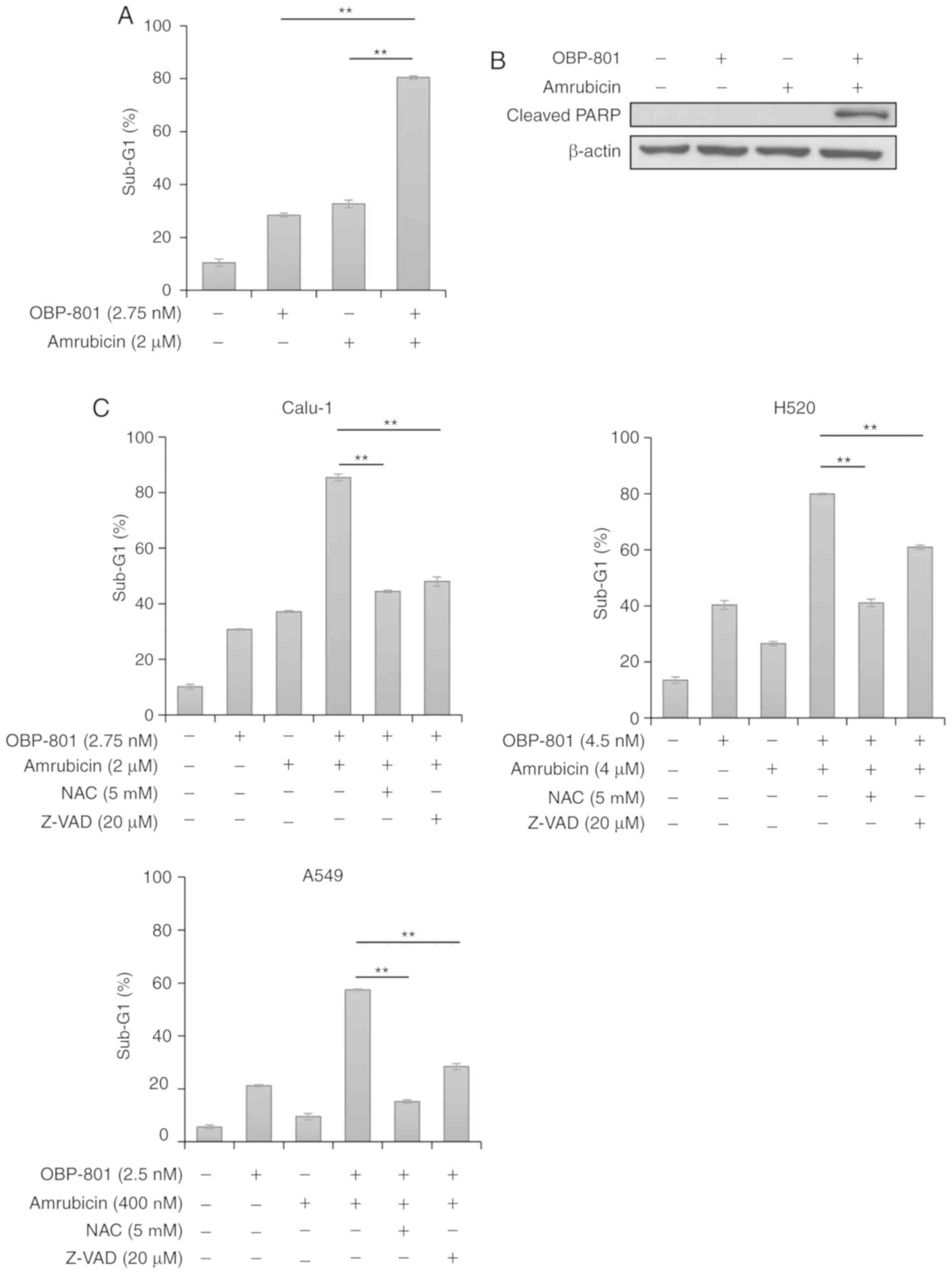
To clarify the mechanisms underlying synergistic
viability inhibition by the combination of OBP-801 and amrubicin,
the effects of this combination on Calu-1 cells was examined using
flow cytometric analysis of apoptotic cell proportions at Sub-G1.
OBP-801 or amrubicin alone slightly induced apoptosis compared with
untreated cells, but the differences were not significant, whereas
the co-treatment with OBP-801 and amrubicin significantly increased
apoptosis in Calu-1 cells compared with either treatment alone
(Figs. 2A and S3). The combination notably induced the
cleavage of the PARP protein (Fig.
2B). Since amrubicin and HDAC inhibitors are both known to
increase the production of ROS (20-24),
whether apoptosis was inhibited by the free radical scavenger NAC
was examined. NAC was used at 5 mM according to our previous
reports (25,26). NAC treatment inhibited apoptosis
induced by the co-treatment with OBP-801 and amrubicin in Calu-1,
H520 and A549 cells (Figs. 2C and
S4-6). In addition, the
pan-caspase inhibitor Z-VAD-FMK was used at 20 µM according
to our previous reports (26,27);
Z-VAD-FMK also significantly inhibited apoptosis in OBP-801 and
amrubicin co-treated Calu-1, H520 and A549 cells (Figs. 2C and S4-6). These results suggested that the
combined treatment with OBP-801 and amrubicin may induce
ROS-mediated and caspase-dependent apoptosis in human lung cancer
cell lines.
OBP-801 and amrubicin co-treatment
induces apoptosis signal-regulating kinase 1 (ASK1)-dependent and
JNK- and p38 mitogen-activated protein kinase (MAPK)-independent
apoptosis
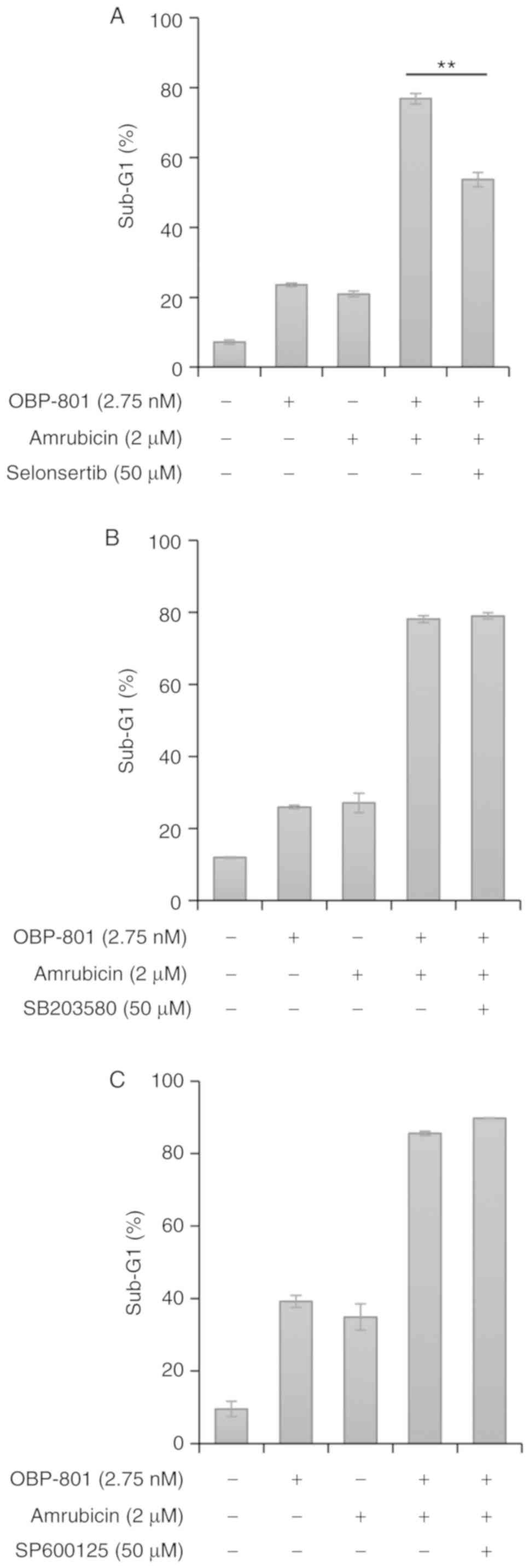
Since ROS is known to induce apoptosis by activating
ASK1 and its downstream targets JNK and p38 MAPK (28), whether apoptosis induced by the
co-treatment with OBP-801 and amrubicin depended on ASK1, JNK or
p38 MAPK was investigated using their respective inhibitors. Based
on previous reports (29,30), these inhibitors were used at the
uniform concentration of 50 µM. The ASK1 inhibitor
selonsertib significantly suppressed apoptosis induced by the
combined treatment in Calu-1 cells (Figs. 3A and S7). However, neither the p38 MAPK
inhibitor SB203580 nor JNK inhibitor SP600125 suppressed apoptosis
induced by the combined treatment (Fig. 3B and C, respectively, and Figs. S8 and S9). These results indicated
that apoptosis induced by the co-treatment with OBP-801 and
amrubicin may be ASK1-dependent and JNK- and p38
MAPK-independent.
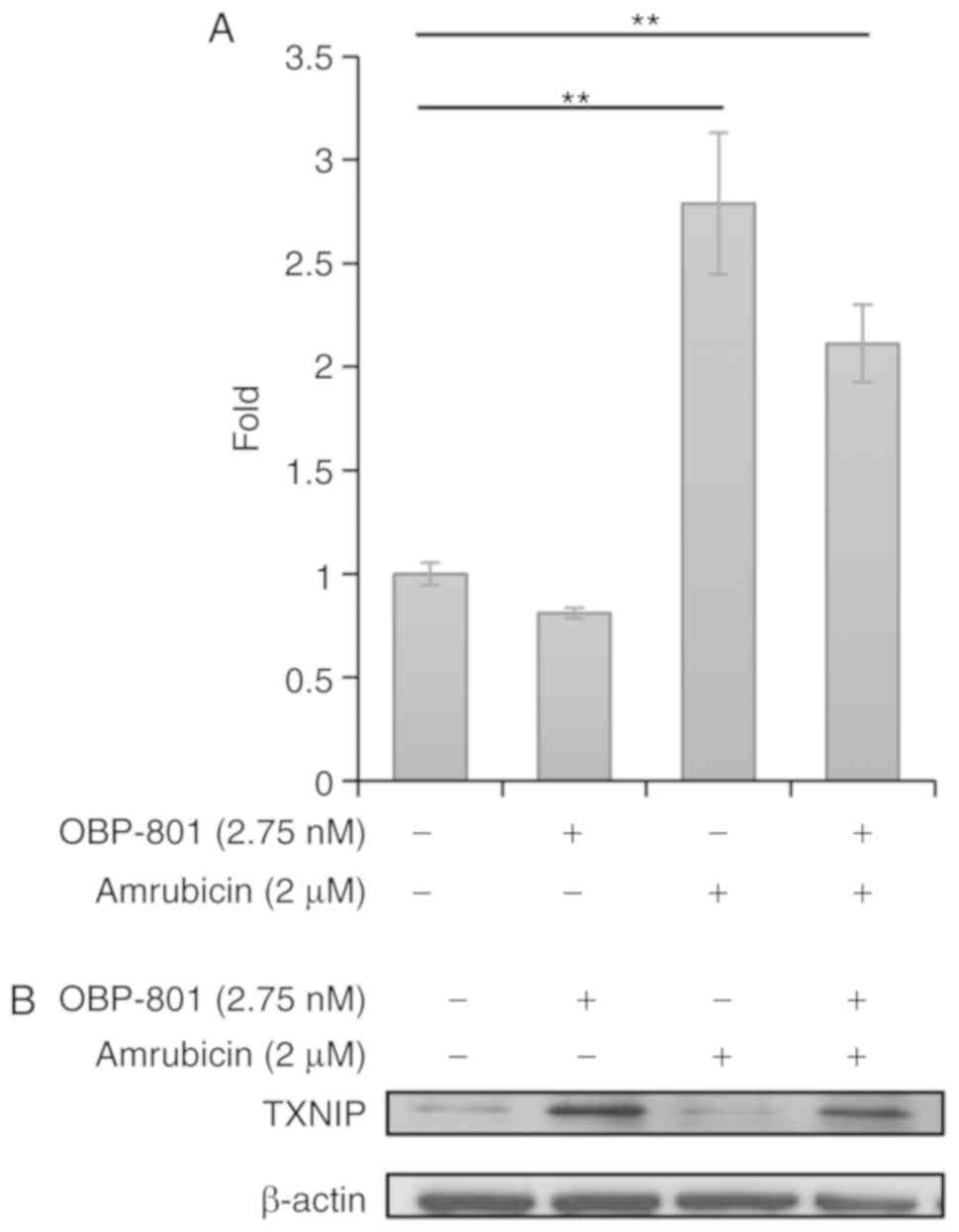
OBP-801 upregulates TXNIP protein
expression and amrubicin increases intracellular ROS
ASK1 is inactivated by the binding of Trx, and ROS
oxidize Trx, resulting in the dissociation of Trx from ASK1 and
activation of ASK1 (31). A
previous study reported that ASK1-dependent and JNK- and p38
MAPK-independent apoptosis was induced by mitochondrial ASK1, which
was inactivated by Trx2 (32).
Therefore, the relationship between the combined treatment with
OBP-801 and amrubicin and the regulation of mitochondrial ASK1 was
examined. Although OBP-801 alone slightly reduced intracellular
ROS, which oxidizes Trx2, amrubicin alone or combination with
OBP-801 increased it (Figs. 4A and
S10). The TXNIP protein is also known to oxidize Trx2 in
mitochondria and to dissociate Trx2 from mitochondrial ASK1
(32). We found that OBP-801 and
its combination potently induced TXNIP protein expression (Fig. 4B). These results suggested that ROS
and TXNIP induced by amrubicin and OBP-801 might contribute to
ASK1-dependent and JNK- and p38 MAPK-independent apoptosis.
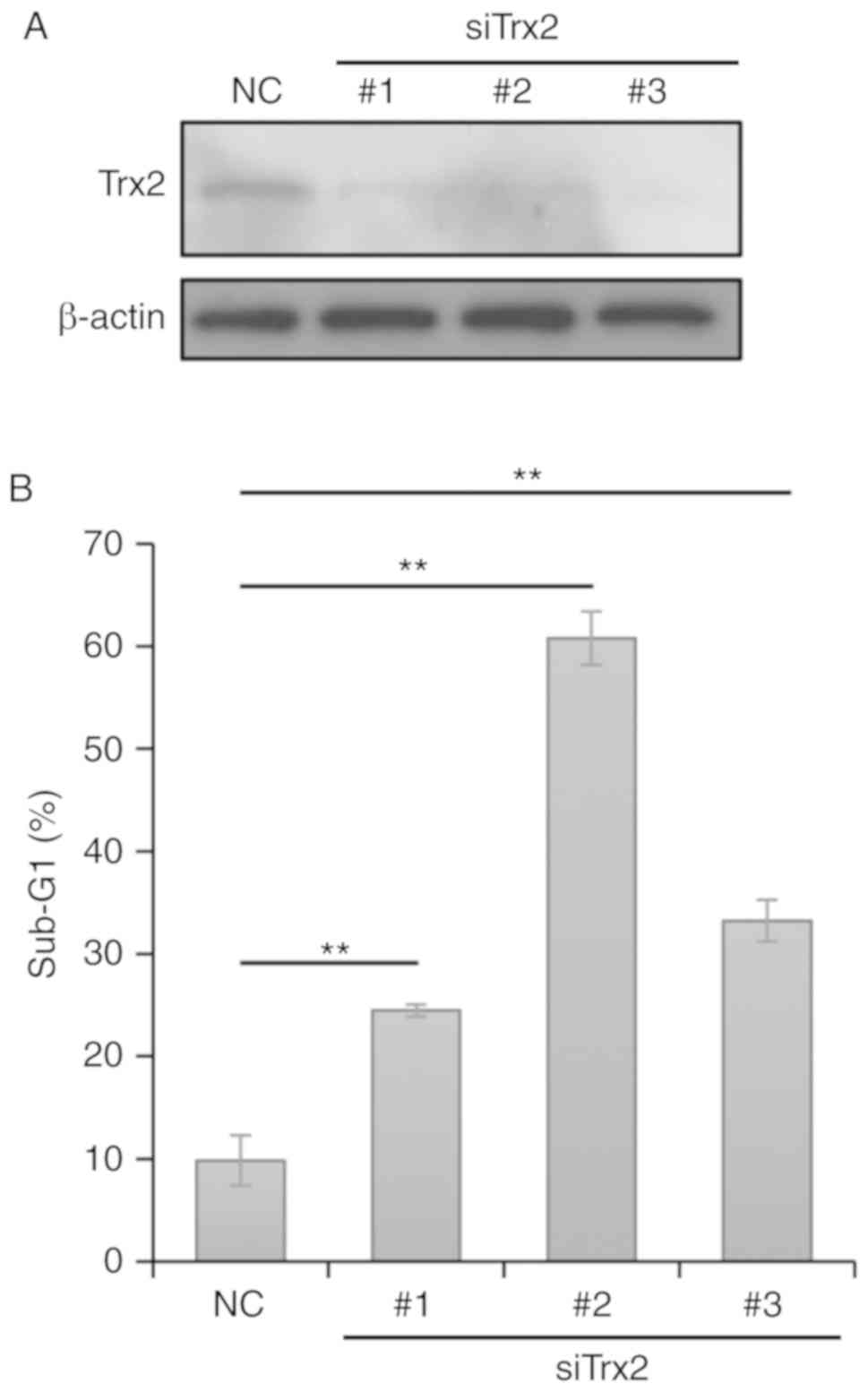
Trx2 knockdown induces apoptosis in
Calu-1 cells
To examine the significance of the disassociation of
Trx2 from ASK1, Trx2 expression was knocked down in Calu-1 cells
using siRNAs. siRNAs targeting Trx2 notably reduced Trx2 protein
expression levels in transfected Calu-1 cells (Fig. 5A). The depletion of Trx2
significantly induced apoptosis in Calu-1 cells compared with
control cells (Figs. 5B and
S11), which suggested that
inactivation of Trx2 induces apoptosis in SQCLC cells.
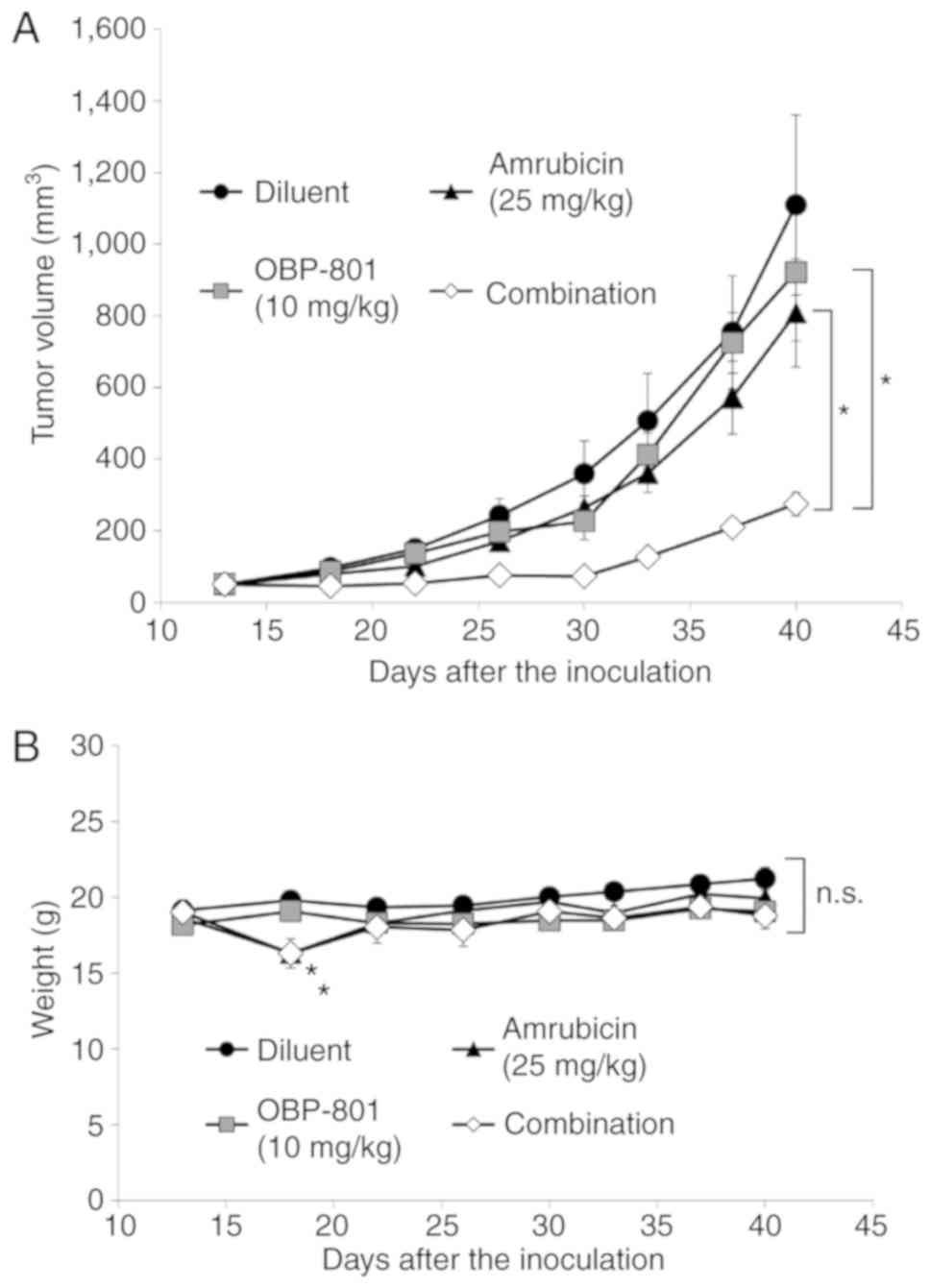
Combined treatment with OBP-801 and
amrubicin inhibits tumor growth in vivo
Furthermore, the antitumor effects of the combined
treatment with OBP-801 and amrubicin were examined in a mouse
xenograft model. Since SQCLC H520 cells, but not Calu-1 cells,
could be engrafted in BALB/c nu/nu mice, the SQCLC H520
xenograft model was used. The combined treatment significantly
suppressed tumor growth compared with either treatment alone on day
40 (Fig. 6A). Although the
treatment with amrubicin reduced the body weights of mice after the
injection on day 18 as previously reported (33), body weights recovered within a few
days, and no significant differences were observed between groups
at the end of the study period (Fig.
6B). These results indicated that the combined treatment with
OBP-801 and amrubicin effectively prevented the tumor growth of
SQCLC in vivo.
Discussion
The treatment of advanced NSCLC has been markedly
improved by the arrival of novel molecular targeted agents, such as
gefitinib (34) and crizotinib
(35), and immune checkpoint
inhibitors, such as nivolumab (36) and pembrolizumab (37). However, no molecular targeted
agents are currently approved for the treatment of SQCLC; thus, the
development of novel effective treatment strategies for SQCLC is
needed. In the present study, a combined treatment with the potent
HDAC inhibitor OBP-801 and amrubicin was demonstrated to
synergistically inhibit the viability of Calu-1 SQCLC cells by
inducing mitochondrial ASK1-dependent apoptosis. Moreover, this
combination strongly inhibited tumor growth in a SQCLC xenograft
model. Limited information is currently available on combined
treatments with HDAC inhibitors for SQCLC. Although the HDAC
inhibitor ITF2357 was previously reported to enhance the
cytotoxicity of pemetrexed against SQCLC cells (38), pemetrexed is not approved for the
treatment of SQCLC. Since amrubicin is approved for the treatment
of SQCLC in Japan, a combined treatment with OBP-801 and amrubicin
may be a realistic combination therapy for SQCLC. Moreover, since
this co-treatment was also effective against human lung
adenocarcinoma A549 cells, it may have potential in the treatment
of NSCLC.
Apoptosis induced by the combined treatment with
OBP-801 and amrubicin was suppressed by the ROS scavenger NAC.
Since HDAC inhibitors and amrubicin are known to increase
intracellular ROS (20-24), it was speculated that OBP-801 and
amrubicin both increased ROS, resulting in the strong induction of
apoptosis. However, OBP-801 treatment alone did not induce ROS, and
ROS levels in co-treatment cells did not exceed those by amrubicin
alone. The mechanisms underlying the induced increase in apoptosis
were examined, and it was demonstrated that OBP-801 strongly
induced the TXNIP protein. Since the overexpression of TXNIP is
known to enhance oxidative stress (39), OBP-801 might augment the anticancer
effects of amrubicin, which has an ability to increase ROS, by
inducing the TXNIP protein. The low dose of OBP-801 (2.75 nM),
which did not increase ROS, enhanced the growth-inhibitory effect
of amrubicin by strongly inducing TXNIP protein. In a mouse
xenograft model, the low dose of OBP-801 (10 mg/kg), which could
not suppress tumor growth alone, exhibited strong tumor growth
inhibition in combination with amrubicin. Since enhanced expression
of TXNIP protein is reported to augment oxidative damage (39), low doses of OBP-801 may potentiate
the anticancer effects of the agents that exhibit cytotoxicity by
inducing ROS.
Although some clinical trials on combination
therapies with HDAC inhibitors and cytotoxic drugs against NSCLC
have been conducted, safety was an important issue in these trials.
A phase I trial of a combination with the HDAC inhibitor vorinostat
and docetaxel was stopped due to excessive toxicity (16). In a randomized phase II trial
evaluating the efficacy of vorinostat in combination with
carboplatin and paclitaxel for advanced NSCLC, adverse events were
more frequent than with a treatment involving carboplatin and
paclitaxel (17). However, since
the feasibility of the combination with the HDAC inhibitor
valproate and doxorubicin was reported in a phase II trial on
patients with malignant mesothelioma (40), combinations with HDAC inhibitors
and anthracyclines may be relatively tolerable. Amrubicin is also
reported to be more tolerable than doxorubicin concerning
cardiotoxicity, which is a noteworthy adverse event associated with
the use of anthracyclines (41,42).
Therefore, the combined treatment with OBP-801 and amrubicin may be
tolerable.
In conclusion, the present study demonstrated that a
combined treatment with the potent HDAC inhibitor OBP-801 and
amrubicin synergistically inhibited the viability of SQCLC cells by
strongly inducing apoptosis. Furthermore, this combined treatment
inhibited tumor growth in an in vivo xenograft model. To the
best of our knowledge, the present study is the first to show the
synergistic efficacy of a combined treatment with a HDAC inhibitor
and anthracycline against Calu-1 SQCLC cells. Since combined
treatments with HDAC inhibitors and anthracyclines may be
tolerable, the combination of OBP-801 and amrubicin has potential
in the treatment of SQCLC.
Supplementary Data
Funding
This study was supported by a commercial research
grant from Oncolys BioPharma, Inc. to TS.
Availability of data and materials
Not applicable.
Authors' contributions
YC, YI, YS, KT and TS conceived the study. YC, YI,
MH and TY designed the experiments. YC, MW, WG, MM and EN performed
the experiments. YC, YI, YS, TY, KT and TS wrote the manuscript and
revised it critically. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
The mouse xenograft study was performed in
accordance with the institutional animal care and use committee
guidelines and approved by the Committee for Animal Research of
Kyoto Prefectural University of Medicine (Kyoto, Japan; permission
no. M29-576).
Patient consent for publication
Not applicable.
Competing interests
The presented study was supported by a commercial
research grant from Oncolys BioPharma, Inc. (Tokyo, Japan) to Dr
Sakai. The remaining authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ungureanu A, Zlatian O, Mitroi G, Drocas
A, Ţîrcă T, Calina D, Dehelean C, Docea AO, Izotov BN, Rakitskii
VN, et al: Staphylococcus aureus colonisation in patients from a
primary regional hospital. Mol Med Rep. 16:8771–8780. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zlatian O, Balasoiu AT, Balasoiu M,
Cristea O, Docea AO, Mitrut R, Spandidos DA, Tsatsakis AM, Bancescu
G and Calina D: Antimicrobial resistance in bacterial pathogens
among hospitalised patients with severe invasive infections. Exp
Ther Med. 16:4499–4510. 2018.PubMed/NCBI
|
|
4
|
Tanase A, Colita A, Ianosi G, Neagoe D,
Branisteanu DE, Calina D, Docea AO, Tsatsakis A and Ianosi SL: Rare
case of disseminated fusariosis in a young patient with graft vs.
host disease following an allogeneic transplant. Exp Ther Med.
12:2078–2082. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Călina D, Roşu L, Roşu AF, Ianoşi G,
Ianoşi S, Zlatian O, Mitruţ R, Docea AO, Rogoveanu O, Mitruţ P, et
al: Etiological diagnosis and pharmacotherapeutic management of
parapneu-monic pleurisy. Farmacia. 64:946–952. 2016.
|
|
6
|
Travis WD: Pathology of lung cancer. Clin
Chest Med. 32:669–692. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Paik PK, Shen R, Berger MF, Ferry D, Soria
JC, Mathewson A, Rooney C, Smith NR, Cullberg M, Kilgour E, et al:
A phase Ib open-label multicenter study of AZD4547 in patients with
advanced squamous cell lung cancers. Clin Cancer Res. 23:5366–5373.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nogova L, Sequist LV, Perez Garcia JM,
Andre F, Delord JP, Hidalgo M, Schellens JH, Cassier PA, Camidge
DR, Schuler M, et al: Evaluation of BGJ398, a fibroblast growth
factor receptor 1-3 kinase inhibitor, in patients with advanced
solid tumors harboring genetic alterations in fibroblast growth
factor receptors: Results of a global phase I, dose-escalation and
dose-expansion study. J Clin Oncol. 35:157–165. 2017. View Article : Google Scholar
|
|
9
|
Yoshioka H, Katakami N, Okamoto H, Iwamoto
Y, Seto T, Takahashi T, Sunaga N, Kudoh S, Chikamori K, Harada M,
et al: A randomized, open-label, phase III trial comparing
amrubicin versus docetaxel in patients with previously treated
non-small-cell lung cancer. Ann Oncol. 28:285–291. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Petta V, Gkiozos I, Strimpakos A and
Syrigos K: Histones and lung cancer: Are the histone deacetylases a
promising therapeutic target? Cancer Chemother Pharmacol.
72:935–952. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Miyanaga A, Gemma A, Noro R, Kataoka K,
Matsuda K, Nara M, Okano T, Seike M, Yoshimura A, Kawakami A, et
al: Antitumor activity of histone deacetylase inhibitors in
non-small cell lung cancer cells: Development of a molecular
predictive model. Mol Cancer Ther. 7:1923–1930. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sun L, He Q, Tsai C, Lei J, Chen J, Vienna
Makcey L and Coy DH: HDAC inhibitors suppressed small cell lung
cancer cell growth and enhanced the suppressive effects of
receptor-targeting cyto-toxins via upregulating somatostatin
receptor II. Am J Transl Res. 10:545–553. 2018.
|
|
13
|
You BR and Park WH: Down-regulation of
thioredoxin1 is involved in death of calu-6 lung cancer cells
treated with suberoyl bishydroxamic acid. J Cell Biochem.
117:1250–1261. 2016. View Article : Google Scholar
|
|
14
|
Reid T, Valone F, Lipera W, Irwin D,
Paroly W, Natale R, Sreedharan S, Keer H, Lum B, Scappaticci F and
Bhatnagar A: Phase II trial of the histone deacetylase inhibitor
pivaloyloxy-methyl butyrate (Pivanex, AN-9) in advanced non-small
cell lung cancer. Lung Cancer. 45:381–386. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Traynor AM, Dubey S, Eickhoff JC, Kolesar
JM, Schell K, Huie MS, Groteluschen DL, Marcotte SM, Hallahan CM,
Weeks HR, et al: Vorinostat (NSC# 701852) in patients with relapsed
non-small cell lung cancer: A wisconsin oncology network phase II
study. J Thorac Oncol. 4:522–526. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Schneider BJ, Kalemkerian GP, Bradley D,
Smith DC, Egorin MJ, Daignault S, Dunn R and Hussain M: Phase I
study of vorinostat (suberoylanilide hydroxamic acid, NSC 701852)
in combination with docetaxel in patients with advanced and
relapsed solid malignancies. Invest New Drugs. 30:249–257. 2012.
View Article : Google Scholar
|
|
17
|
Ramalingam SS, Maitland ML, Frankel P,
Argiris AE, Koczywas M, Gitlitz B, Thomas S, Espinoza-Delgado I,
Vokes EE, Gandara DR and Belani CP: Carboplatin and paclitaxel in
combination with either vorinostat or placebo for first-line
therapy of advanced non-small-cell lung cancer. J Clin Oncol.
28:56–62. 2010. View Article : Google Scholar
|
|
18
|
Shindoh N, Mori M, Terada Y, Oda K, Amino
N, Kita A, Taniguchi M, Sohda KY, Nagai K, Sowa Y, et al: YM753, a
novel histone deacetylase inhibitor, exhibits antitumor activity
with selective, sustained accumulation of acetylated histones in
tumors in the WiDr xenograft model. Int J Oncol. 32:545–555.
2008.PubMed/NCBI
|
|
19
|
Ono H, Iizumi Y, Goi W, Sowa Y, Taguchi T
and Sakai T: Ribosomal protein S3 regulates XIAP expression
independently of the NF-κB pathway in breast cancer cells. Oncol
Rep. 38:3205–3210. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Salvatorelli E, Menna P, Gonzalez Paz O,
Surapaneni S, Aukerman SL, Chello M, Covino E, Sung V and Minotti
G: Pharmacokinetic characterization of amrubicin cardiac safety in
an ex vivo human myocardial strip model. II. Amrubicin shows
metabolic advantages over doxorubicin and epirubicin. J Pharmacol
Exp Ther. 341:474–483. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Petruccelli LA, Dupéré-Richer D,
Pettersson F, Retrouvey H, Skoulikas S and Miller WH Jr: Vorinostat
induces reactive oxygen species and DNA damage in acute myeloid
leukemia cells. PLoS One. 6:e209872011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
You BR and Park WH: Trichostatin A induces
apoptotic cell death of HeLa cells in a Bcl-2 and oxidative
stress-dependent manner. Int J Oncol. 42:359–366. 2013. View Article : Google Scholar
|
|
23
|
You BR, Kim SH and Park WH: Reactive
oxygen species, glutathione, and thioredoxin influence suberoyl
bishydroxamic acid-induced apoptosis in A549 lung cancer cells.
Tumour Biol. 36:3429–3439. 2015. View Article : Google Scholar
|
|
24
|
Han BR, You BR and Park WH: Valproic acid
inhibits the growth of HeLa cervical cancer cells via
caspase-dependent apoptosis. Oncol Rep. 30:2999–3005. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yoshioka T, Yogosawa S, Yamada T, Kitawaki
J and Sakai T: Combination of a novel HDAC inhibitor OBP-801/YM753
and a PI3K inhibitor LY294002 synergistically induces apoptosis in
human endometrial carcinoma cells due to increase of Bim with
accumulation of ROS. Gynecol Oncol. 129:425–432. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yamada T, Horinaka M, Shinnoh M, Yoshioka
T, Miki T and Sakai T: A novel HDAC inhibitor OBP-801 and a PI3K
inhibitor LY294002 synergistically induce apoptosis via the
suppression of survivin and XIAP in renal cell carcinoma. Int J
Oncol. 43:1080–1086. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ono H, Sowa Y, Horinaka M, Iizumi Y,
Watanabe M, Morita M, Nishimoto E, Taguchi T and Sakai T: The
histone deacetylase inhibitor OBP-801 and eribulin synergistically
inhibit the growth of triple-negative breast cancer cells with the
suppression of survivin, Bcl-xL, and the MAPK pathway. Breast
Cancer Res Treat. 171:43–52. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ichijo H, Nishida E, Irie K, ten Dijke P,
Saitoh M, Moriguchi T, Takagi M, Matsumoto K, Miyazono K and Gotoh
Y: Induction of apoptosis by ASK1, a mammalian MAPKKK that
activates SAPK/JNK and p38 signaling pathways. Science. 275:90–94.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tai LM, Holloway KA, Male DK, Loughlin AJ
and Romero IA: Amyloid-beta-induced occludin down-regulation and
increased permeability in human brain endothelial cells is mediated
by MAPK activation. J Cell Mol Med. 14:1101–1112. 2010.
|
|
30
|
Chen YY, Liu FC, Chou PY, Chien YC, Chang
WS, Huang GJ, Wu CH and Sheu MJ: Ethanol extracts of fruiting
bodies of antrodia cinnamomea suppress CL1-5 human lung
adenocarcinoma cells migration by inhibiting matrix
metalloproteinase-2/9 through ERK, JNK, p38, and PI3K/Akt signaling
pathways. Evid Based Complement Alternat Med.
2012:3784152012.PubMed/NCBI
|
|
31
|
Noguchi T, Takeda K, Matsuzawa A, Saegusa
K, Nakano H, Gohda J, Inoue J and Ichijo H: Recruitment of tumor
necrosis factor receptor-associated factor family proteins to
apoptosis signal-regulating kinase 1 signalosome is essential for
oxidative stress-induced cell death. J Biol Chem. 280:37033–37040.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Saxena G, Chen J and Shalev A:
Intracellular shuttling and mitochondrial function of
thioredoxin-interacting protein. J Biol Chem. 285:3997–4005. 2010.
View Article : Google Scholar :
|
|
33
|
Hatakeyama Y, Kobayashi K, Nagano T,
Tamura D, Yamamoto M, Tachihara M, Kotani Y and Nishimura Y:
Synergistic effects of pemetrexed and amrubicin in non-small cell
lung cancer cell lines: Potential for combination therapy. Cancer
Lett. 343:74–79. 2014. View Article : Google Scholar
|
|
34
|
Mok TS, Wu YL, Thongprasert S, Yang CH,
Chu DT, Saijo N, Sunpaweravong P, Han B, Margono B, Ichinose Y, et
al: Gefitinib or carboplatin-paclitaxel in pulmonary
adenocarcinoma. N Engl J Med. 361:947–957. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Solomon BJ, Mok T, Kim DW, Wu YL, Nakagawa
K, Mekhail T, Felip E, Cappuzzo F, Paolini J, Usari T, et al:
First-line crizotinib versus chemotherapy in ALK-positive lung
cancer. N Engl J Med. 371:2167–2177. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Brahmer J, Reckamp KL, Baas P, Crinò L,
Eberhardt WE, Poddubskaya E, Antonia S, Pluzanski A, Vokes EE,
Holgado E, et al: Nivolumab versus docetaxel in advanced
squamous-cell non-small-cell lung cancer. N Engl J Med.
373:123–135. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Reck M, Rodríguez-Abreu D, Robinson AG,
Hui R, Csőszi T, Fülöp A, Gottfried M, Peled N, Tafreshi A, Cuffe
S, et al: Pembrolizumab versus chemotherapy for PD-L1-positive
non-small-cell lung cancer. N Engl J Med. 375:1823–1833. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Del Bufalo D, Desideri M, De Luca T, Di
Martile M, Gabellini C, Monica V, Busso S, Eramo A, De Maria R,
Milella M and Trisciuoglio D: Histone deacetylase inhibition
synergistically enhances pemetrexed cytotoxicity through induction
of apoptosis and autophagy in non-small cell lung cancer. Mol
Cancer. 13:2302014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yu Y, Xing K, Badamas R, Kuszynski CA, Wu
H and Lou MF: Overexpression of thioredoxin-binding protein 2
increases oxidation sensitivity and apoptosis in human lens
epithelial cells. Free Radic Biol Med. 57:92–104. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Scherpereel A, Berghmans T, Lafitte JJ,
Colinet B, Richez M, Bonduelle Y, Meert AP, Dhalluin X, Leclercq N,
Paesmans M, et al: Valproate-doxorubicin: Promising therapy for
progressing mesothelioma. A phase II study Eur Respir J.
37:129–135. 2011.
|
|
41
|
Suzuki T, Minamide S, Iwasaki T, Yamamoto
H and Kanda H: Cardiotoxicity of a new anthracycline derivative
(SM-5887) following intravenous administration to rabbits:
Comparative study with doxorubicin. Invest New Drugs. 15:219–225.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Noda T, Watanabe T, Kohda A, Hosokawa S
and Suzuki T: Chronic effects of a novel synthetic anthracycline
derivative (SM-5887) on normal heart and doxorubicin-induced
cardiomyopathy in beagle dogs. Invest New Drugs. 16:121–128. 1998.
View Article : Google Scholar : PubMed/NCBI
|